Изобретение относится к биотехнологии и может быть использовано в молекулярной энзимологии, а также для получения гомогенных препаратов Мп2 зависимой ДНКазы, хроматина печени крыс методом аффинной хроматографии.
Штамм гибридных культивируемых клеток Mus musculus L. получают следующим образом.
В качестве доноров иммунных селезенок используют 8-12-недельных мышей линии Balb/c. Мышей им.мунизируют внутрибрюшинно и подкожно нейтральной Мп2+-зависимой ДНКазой хроматина печени крыс с удельной активностью 600 ед/мг. За три дня до гибридизации животным вводят высокоочищенный препарат фермента с удельной активностью 2358 ед/мг. Культуру клеток миеломы мыши NS/1-Ag.4.1., у.которой потеряна способность к синтезу собственных иммуноглобулинов, получают из
коллекции института Цитологии А СССР. Клетки культивируют на полноценной среде следующего состава: 1 литр исходной среды RPNI - 1640, 2 г МаНСОз, 110 мг пирувата натрия, 15% нативной эмбриональной телячьей сыворотки, 2 мМ L-глуга- мина, 10 мМ HEPES, 50 мг гентамицина, 10 мМуЗ-меркаптоэтанола, 5 103 ед/мл инсулина. Клетки в течение трех недель выращивают на полноценной среде в присутствии 20 мкг/мл 8-азагуанина для освобождения от предшествующих ревертантов. За три дня до гибридизации клетки переводят на полноценную среду без 8-азагуанина. Для гибридизации монослой миеломных клеток, находящихся в логарифмической фазе роста, смывают полноценной средой без сыворотки. Подсчет клеток ведут в камере Горяева. Количество жизнеспособных клеток определяют при окрашивании 0,5%-ным раствором трипанового синего. Слияние
00 О 00 00 VJ Ю
клеток миеломы с иммунными спленоцита- ми проводят по методу Келера и Милстейна с помощью полиэтиленгликоля, Селезенку иммунизированного животного промывают 2-3 раза полноценной средой без сыворотки и переносят в стеклянный гомогенизатор, Тремя осторожными давительными движениями получают суспензию клеток. После оседания крупных частиц собирают полученную суспензию клеток и смешивают с миеломными в соотношении 2 10 спле- ноцитов и 1 107 миеломных. Клетки осаждают центрифугированием в течение 10 минут при 800 д. Затем по каплям добавляют 2 мл 50%-ного раствора полиэтиленгликоля (молекулярная масса 1540). приготовленного на полноценной среде, но без сыворотки и HEPES. После мягкого перемешивания в течение минуты, приливают полноценную среду (50 мл) и центрифу- . гируют 10 минут при 800 д. Полученный осадок заливают полноценной средой. Суспензию клеток оставляют на 12 часов в термостате. Затем полноценную среду заменяют на селективную среду HAT - полноценную среду, содержащую10 мол/л гипоксантина, 1,6-10 мол/л ти- мидина, 4- .10 мол/л аминоптерина - и раскапывают в 96-луночные панели с питающим слоем, из перитонеальных макрофагов мыши. Клетки выращивают в 5%-ной атмосфере С02 при 37°С. На 14-ый день появляются первые колонии гибридом, су- пернатант этих лунок исследуют на присутствие моноклональных антител путем иммуноферментного анализа.
Далее путем клонирования осуществляют выделение стабильных клонов гибридных кле ток. Клетки из тех лунок, супернатант которых дает положительный результат в ИФА, используют для клонирования и реклонирования. Для этого клетки, находящиеся в середине логарифмической фазы роста, разводят свежей полноценной средой и рассевают в новую панель из расчета 1 клетка на лунку. Предварительно в эти панели рассевают питающие клетки. Клонирование проводят трехкратно, так, чо 60% клонов дают положительный ответ при исследовании методом ИФА. Клоны, дающие положительный результат в ИФА и ингибирующие активность ДНКазы в системе In vitro, выращивают в сосудах Карреля.
Полученный штамм обозначают 4Е, он хранится в Специализированной коллекции перевиваемых соматических клеток позвоночных Института цитологии АН СССР под
0
5
0
5
0
5
0
5
0
5
номером 530Д и характеризуется следующими признаками.
Культуральные признаки.
Среда RPMI - 1640с 10% телячьей эмбриональной сыворотки, 2 мМ L-глутамина, 50 г/л гентамицина, 10 мМ 2-меркаптоэта- нола, 5 ед/мл инсулина. Клетки выращивают при 37°С в 5%-иой атмосфере на стеклянной или пластиковой культураль- ной посуде. Посевная доза 5- 104 кл/мл. Пассажи проводят каждые 2-3 дня с кратностью рассева 1:5. Для получения асцитной жидкости 5-7 -10 клеток вводят перитони- ально мышам Balb/c, сенсибилизированным пристанем. Опухоль развивается через 7-10 дней.
Криоконсервирование.
Клетки штамма ресуспендируют вереде RPNI с 45% эмбриональной телячьей сыворотки в присутствий 10% ДМСО, разливают в пластиковые ампулы по 4-6 -10 клеток в 1 мл.
Размораживание.
Быстро при 37°С клетки разводят в 10 мл полноценной среды и осаждают, ресуспендируют в 5 мл той же среды и переносят в культуральный флакон. Выживамость после размораживания более составляет более 30%.
Контаминация.
Бактерии и грибы в культуре не обнаружены при длительном наблюдении и посевах на питательные среды.
Характеристика полезного продукта.
Моноклональные антитела связывают
О 1
Мп -зависимую ДНКазу, при этом они способны ингибировать нуклеазную активность данного фермента. Титр. МКА в культуральной жидкости, определенный с помощью ИФА, составляет 1:8, в асцитной жидкости - 1:10 .
Пример 1, Для определения специфического взаимодействия моноклональных антител с Мп -зависимой ДНКазой проводят определение нуклеазной активности Мп +-зависимой ДНКазы в присутствии антител. Нуклеазную активность определяют спектрофотометрически. В работе используют реакционную смесь следующего состава (общий обьем 200 мкл):100 мкг денатурированной нагреванием ДНК из тиму- са теленка, 20 мМ трис HCI буфера рН 7,2, 25 мМ Мп2+. Во все пробы добавляют по 20 мкл препарата ДНКазы, инкубированной в течение 30 минут при 37°С с моноклональ- ными антителами. Контролем служат пробы, где фермент инкубируют без антител. Затем реакционную смесь с ферментом инкубируют при 37°С в течение часа. Реакцию останавливают добавлением 3,8 мл 3% HCI04 на холоду. Осадок кислотнонерастворимых продуктов удаляют через 15 мин центрифугированием при 7000 g в течение 15 мин. В надосадочной жидкости прирост кислотора- створимых продуктов определяют на спектрофотометре по поглощению при 260 нм. За единицу активности . фермента принимают изменение оптической плотности /Е260/ реакционной смеси за 1 ч инкубации в пересчете на 1 мл ферментного препарата (ед/мл). Удельную активность определяют как отношение числа единиц активности ферментного препарата на количество мг белка (ед/мг). Активность фермента, инкубированного без антител, принимают за 100%, а активность фермента, инкубированного в присутствии антител, пересчитывают в пропорции:
100% - активность фермента без антител (Е260)
Х% - активность фермента с антителами (Е260)
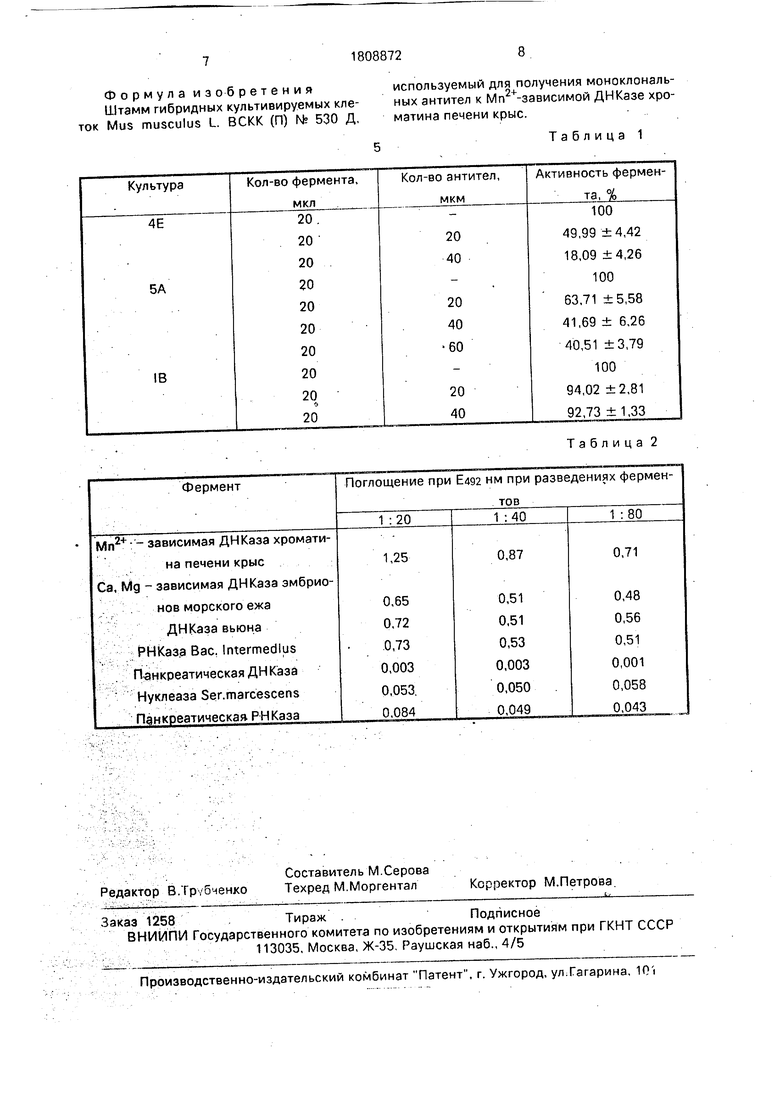
Полученные результаты представлены в табл.1.
Для того, чтобы исключить возможность ошибки эксперимента при разведении фермента вследствие внесения в реакционную смесь дополнительного объема жидкости в виде антител, ставят следующие опыты: вместо исследуемых антител в реакционную смесь вносят антитела клонов 5А и 1В, которые дают положительный ответ в ИФА при взаимодействии с ферментатом. Полученные данные тоже представлены в табл.1.
Таким образом, результаты полученных исследований показывают, что только клон 4Е является продуцентом антител, которые способны ингибировать активность фермента на 80%. Клоны 5А и 1В, которые дают высокий положительный ответе ИФА, не способны ингибировать активность изучаемого фермента на 80%, даже при увеличении количества вносимых антител. По литературным данным большего процента ингибирования активности ферментов иммуноглобулинами достигнуть невозможно. Таким образом, из полученных гибридом только клон 4Е ртвеча-. ет поставленным требованиям - максимальной способностью ингибировать активность Мп2+-зависимой ДНКазы. Пример 2. Полученные моноклональ- .ные антитела используют для изучения им- мунохимических свойств нуклеаз (эволюционная биохимия ферментов). В работе используют ферменты различных эволюционных труп: Мп2+-зависимую ДНКазу хроматина печени крыс с удельной активностью 2358 ед/мг; Са, Mg-зависимую ДНКазу ядер эмбрионов морского ежа с удельной активностью 1278 ед/мг; ДНКазу из ядер
эмбрионов вьна с удельной активностью 416 ед/мг; панкреатическую ДНКазу с удельной активностью 4200 ед/мг по Кунит- цу; панкреатическую РНКазу с удельной активностью 40 ед/мг по Кунитцу, а также бактериальные нуклеазы. РНКазу Bac.intermedlus с удельной активностью 12000 ед/мг, и нуклеазу Ser.marcescens с удельной активностью 1,5-2,0 -106 ед/мг.
0 Для изучения взаимодействия монокло- нальных антител с различными ферментами применяют метод твердофазного ИФА, т.к.. он обладает высокой чувствительностью и позволяет в одинаковых условиях провести
5 анализ сотни проб. В качестве антигена, в панель вносят ферменты в различных разве- дениях, а в качестве антител используют фракции моноклональных антител клона 4Е в разведении 1:2000 (концентрация антител
0 1,1 мкг/мл).
Моноклональные антитела разводят на 1%-ном БСА в буфере, содержащем 0,01 М трис-HCI, 0,15 М NaCI. М ЭДТА, поскольку при разведении без БСА активность.
5 антител резко снижается.
Результаты исследований представле- 1 ны в табл.2.
Как видно из таблицы, моноклональные
О I
антитела к Мп -зависимой ДНКазе хрома0 тина печени крыс связываются с этим ферментом; Са, Mg-зависимой ДНКазой ядер эмбрионов морского ежа, нуклеазой эмбрионов вьюна и РНКазой Bacillus intermedius, но не связываются с панкреатической
5 ДНКазой, нуклеазой Ser.marcescens и панкреатической РНКазой, На основании полученных данных можно сделать вывод о наличии сходных антигенных детерминант у трех из семи исследованных ферментов.
0 Способность моноклональных антител распознавать ферменты с высокой степенью специфичности используется для ре- шения ряда проблем молекулярной генетики и эволюционной, биохимии. Инте5 ресно отметить, что в 60% случаев детальное изучение ферментов с помощью моноклональных антител показало, что последние существуют в мультимолекулярной форме. Можно предположить, что изучае0 мый авторами фермент Мп -зависимая ДНКаза хроматина печени крыс существует в такой форме. Но этот вопрос еще подлежит дальнейшему изучению. Используя моноклональные антитела, можно проводить
5 исследования в эволюционном плане, следя за появлением и изменением содержания изучаемого антигена в клетке при развитии организма или клеточном цикле, наблюдая за миграцией антигена в-пределах клетки и всего организма.
Формула из об р е т е н и я Штамм гибридных культивируемых клеток Mus musculus L. ВСКК (П) № 530 Д,
используемый для получения моноклональ- ных антител к М независимой ДНКазе хроматина печени крыс.
Таблица 1


Использование: биотехнология, молекулярная энзимология. Сущность изобретения: получение штамма гибридных культивируемых клеток Mus musculus L., продуцирующего моноклональные антитела (МКА) к Мп -зависимой ДНКазе хроматина печени крысы. Штамм получают гидридиза- цией спленоцитов мышей линии Balb/c и клеток миеломы NS/1 - Ад 4.1. Титр МКА в культуральной жидкости, определенный с помощью иммуноферментного анализа, составляет 1:8, в асцитной - 1:10 .МКА связывают Мп2+-зависимую ДНКазу, при этом они способны ингибировать нуклеазную активность данного фермента. 2 табл.
Таблица 2
