Изобретение направлено на способ получения тилозина, предусматривающий трансформацию стрептомицетов последовательностью ДНК, содержащей гены блосинтеза антибиотика.
Изобретение является одним из первых и очень важных промышленных разработок технологии рекомбинантной ДНК для практического использования в продуцирующих антибиотик организмах, таких как стрептомицеты.
Большинство технологий рекомбинантной ДНК, применяемой для стрептомицетов и других продуцирующих антибиотики организмов, ограничивались разработкой векторов клонирования. Первыми примерами таких технологий являются описание патента США N 4 332 898 Reusser U.S. и патентов США N N 4273875; 4332900; 4338400; и 4340674 Manis и др.
Трансформация стрептомицетов не была описана ни в одной из ранних публикаций. Усовершенствованные векторы, проявляющие значительно большие возможности для использования в продуцирующих антибиотики организмах, были описаны, например, Feyerman и др. патент США N 4513086, и Nakatsukasa и др. патенты США N 4513085 и 4416994. Эти усовершенствованные векторы включают сигнальные гены, которые являются отбираемыми в стрептомицетах и могут использоваться для трансформации многих важных штаммов Streptomyces, и составляют средства, необходимые для проведения более сложных экспериментов генного клонирования.
Об одном таком эксперименте недавно сообщалось Hopwood и др. 1985 г, в журнале Nature 314, 642. Хотя Hopwood и др. сообщали о продуцировании новых гибридных антибиотических пигментов, в данной публикации ничего не говорится об увеличении способности к продуцированию антибиотика или об эффективности биосинтеза данной клетки хозяина, но говорится о переносе генов биосинтеза актинородинового пигмента от одного штамма Streptomyces к другому.
Известен также способ получения тилозина путем культивирования штамма Streptomyces fradiae в питательной среде, выделение и очистку целевого продукта (А.С. СССР N 755837, кл. С 12 N 15/00, 15.08.80 прототип).
Согласно данному изобретению используются гены биосинтеза тилозина, которые увеличивают способность продуцировать антибиотик. Один такой ген кодирует фермент, макроцин 0 метилтрансфераза, катализирует конечный этап биосинтеза тилозина. Трансформация продуцирующих тилозин микроорганизмов геном, кодирующим макролин-0-метилтрансферазу, обозначаемым как tyl F. приводит в результате к улучшенному пути биосинтеза тилозина ввиду повышенных содержаний генного продукта tyl F в трансформированных клетках. Было клонировано небольшое количество генов биосинтеза антибиотиков. Способы выделения генов биосинтеза антибиотиков уже были разработаны, но один особенно предпочтительный способ выделения генов был описан Baltz и др. патентная заявка США, номер серии 742 349, 07.06.85, (Эквивалента Европейской патентной заявке N 86304239, 6), которая рассматривается здесь как ссылочный материал. Отвечающие данному изобретению гены биосинтеза антибиотика тилозина, используемые в описываемых примерах первоначально выделялись из библиотеки λ, построение которой в основном соответствует методике, описанной Fishman и др. 1985, J. Bacteriology 166, 199-206.
Способ поясняется фиг. 1-7.
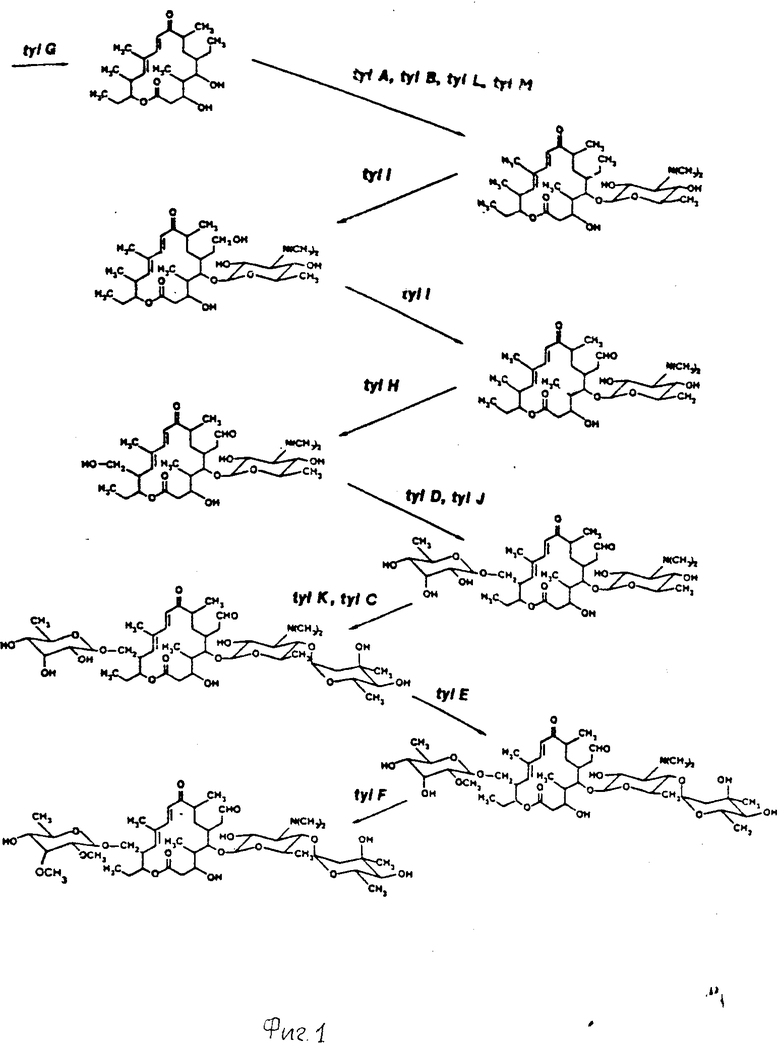
На фиг. 1 показано схематическое воспроизведение пути биосинтеза тилозина, каждая стрелка показывает этап, который катализируется одним или несколькими генными продуктами биосинтеза тилозина. Ген (гены), ответственный за каждую конверсию, показан над каждой стрелкой. Каждое обозначение генотипа может относиться к классу генов, который способствует образованию одного и того же фенотипа. Для осуществления данного изобретения используется целый ряд векторов экспрессии. Эти векторы включают один или несколько генов биосинтеза тилозина и могут быть получены из библиотеки Северной Региональной Научно-Исследовательской Лаборатории (NRRL). Пеория, Иллинойс, 61604.
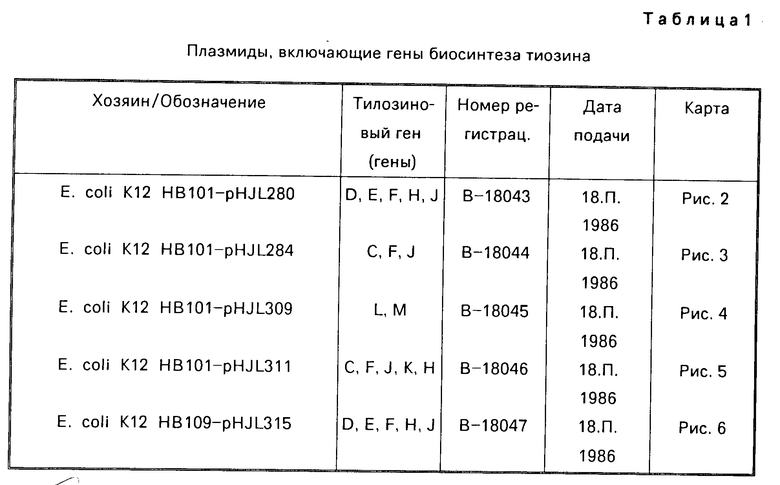
В тал. 1 представлено краткое описание каждой из плазмид, используемых для осуществления способа данного изобретения.
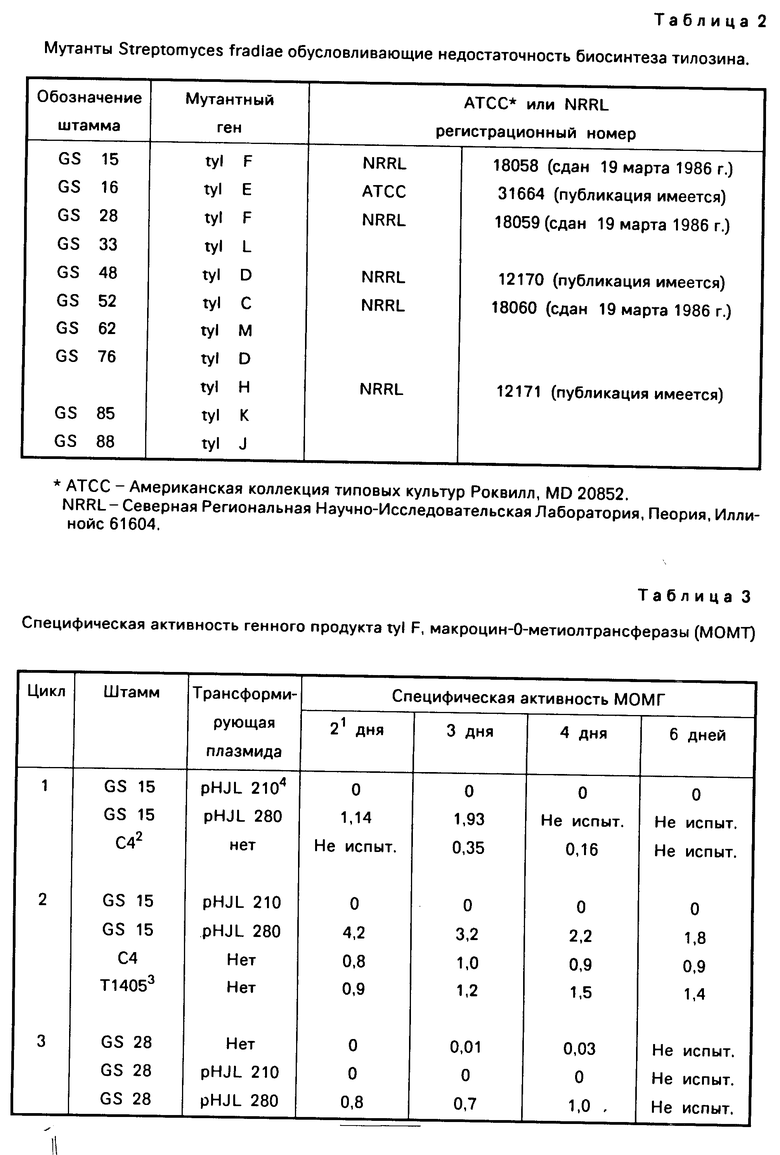
Описывается ряд штаммов Streptomyces fradial, которые имеют гены биосинтеза мутанта тилозина, и, следовательно, образуют значительно меньше тилозина, чем штамм, от которого они образованы. В табл. 2 дается краткое описание этих мутантных штаммов.
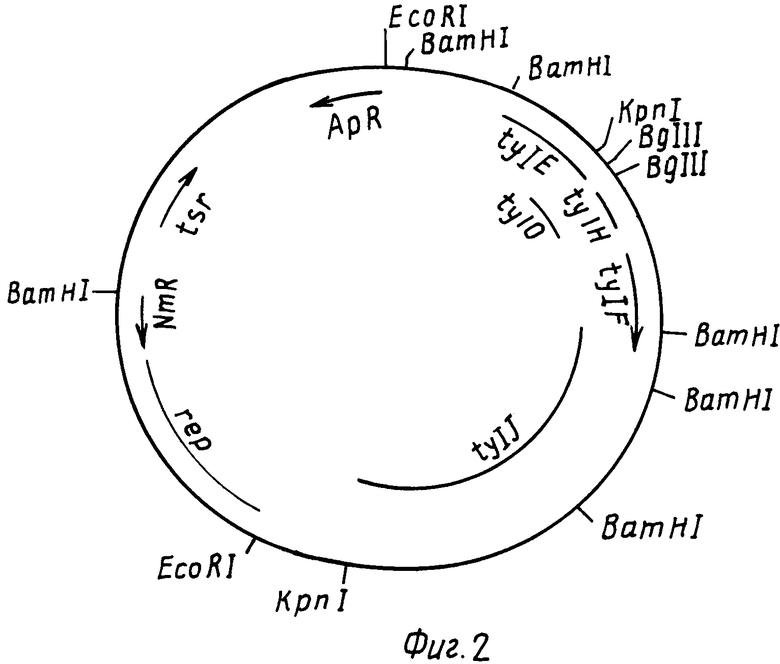
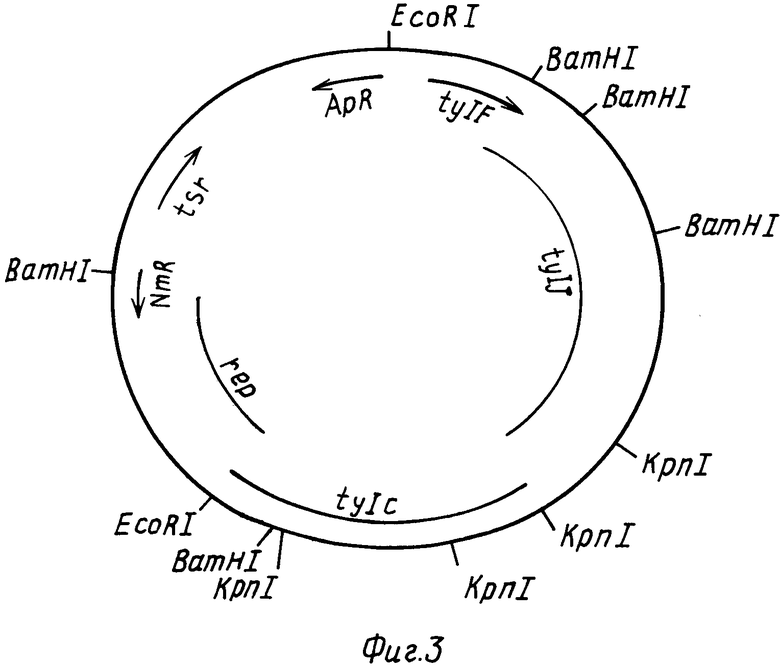
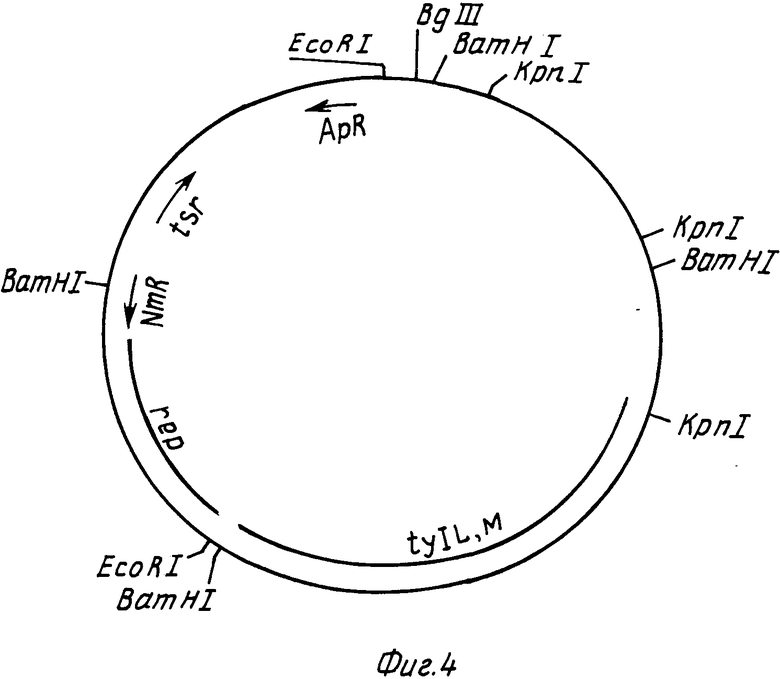
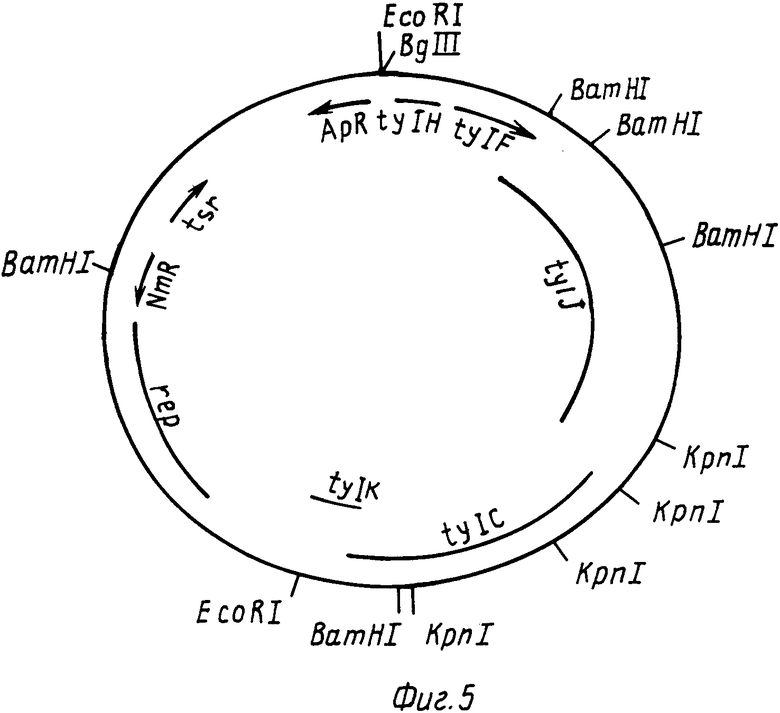
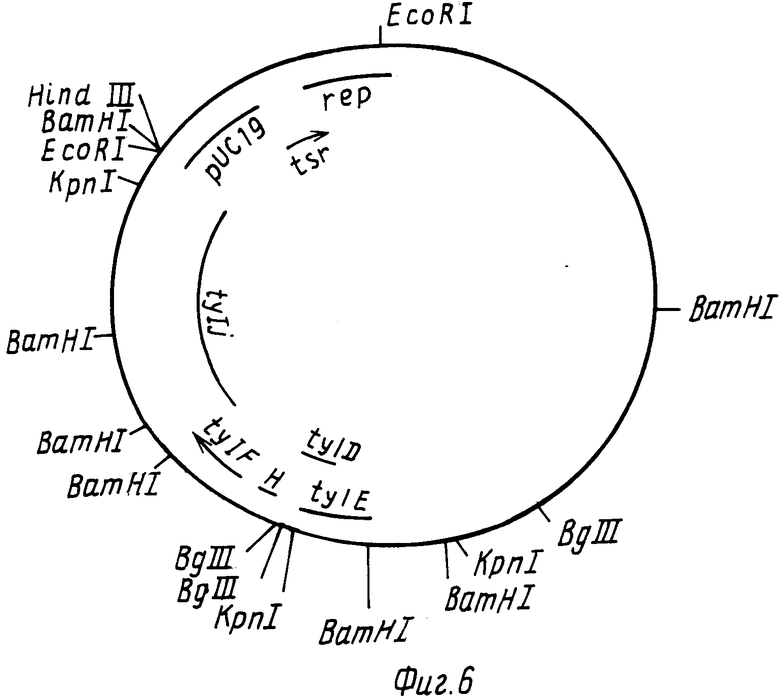
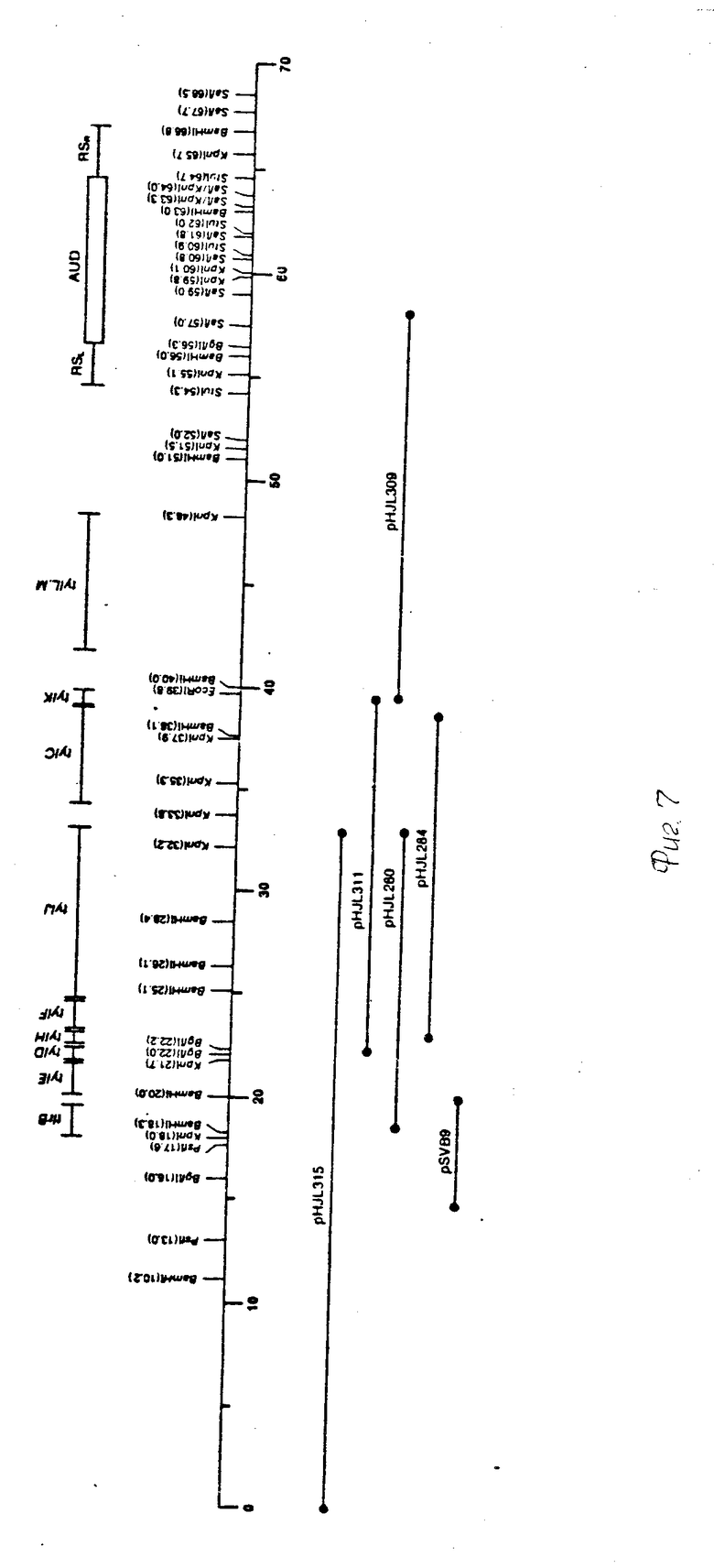
На фиг. 2 РНJL 280 показана рестрикционная и функциональная карта плазмиды; на фиг. 3 рНJL 284 рестрикционная и функциональная карта плазмиды; на фиг. 4 рHJL 309 рестрикционная и функциональная карта плазмиды; на фиг. 5 рНJL311 рестрикционная и функциональная карта плазмиды; на фиг. 6 рНJL 315 рестрикционная и функциональная карта плазмиды; на фиг. 7 хромосомная организация генов биосинтеза тилозина.
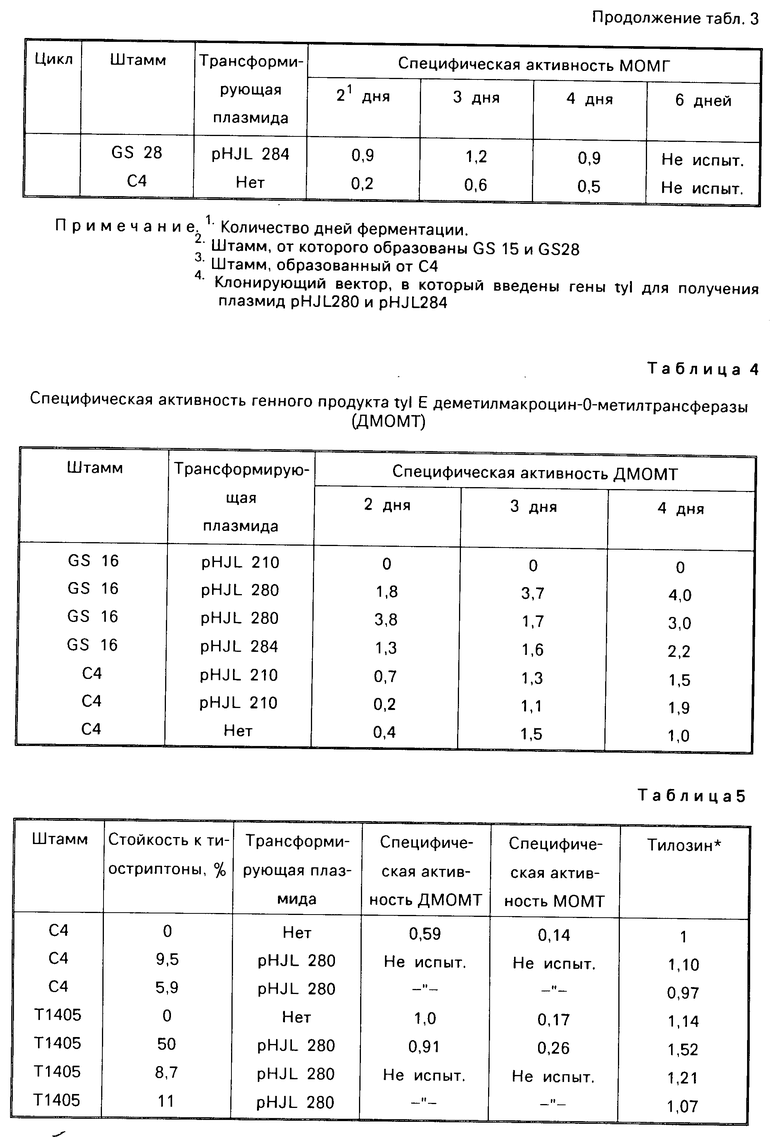
Плазмиды рНJL280, рНJL284 и рНJL315 были использованы для трансформации Streptomyces fradial GS15 и Streptomyces fradiae GS28. Штаммы GS15 и GS28 были получены от S.fradiae С4 путем мутагенеза нитрозогуанидином. S.fradiae C4 был получен от S.fradiae T59235 (АТСС 19609) путем мутагенеза. Штамм GS 15 почти не образует тилозина и штамм GS 28 дает очень маленькие концентрации тилозина по сравнению с штаммом С4. Пониженная способность к продуцированию тилозина у штаммов GS15 и GS28 или отсутствие этой способности вообще является результатом мутаций, влияющих на ген tyl F, который кодирует макроцин-0-метилтрансферазу (МОМТ). Фермент МОМТ, который требуется для превращения макроцина в тилозин путем биосинтеза тилозина, очень часто присутствует в количествах, ограничивающих скорость реакции, в продуцирующих тилозин штаммах. Плазмиды рНJL280, рНJL284 и рНJL315 устраняют это ограничение реакции посредством увеличения как числа копий гена биосинтеза так и концентрации макроцин-0-метилтрансферазы, доступной для синтеза тилозина. Согласно этому ферментация S. fradiae GS 15/рНJL280, S.fradiae GS 6515/рНJL 284, S. fradiae GS 15/рНJL 315, S.fradiae GS 28/рНJL 284, S.fradiae GS28/рНJL280 и S.fradiae GS 28/рНJL 315 в течение 72 ч приводит в результате примерно к 2-6 кратному увеличению продуцирования микроцин-0-метилтрансферазы по сравнению с той, которая образуется в случае штамма С4 и к 120 кратному увеличению по сравнению с той, которая образуется в случае штамма GS28.
Плазмида рНJL280 использовалась также для трансформации: (1) Streptomyces fradiae GS 16; S.fradiae GS 48, (3) S.fradiae GS 76 и (4). S.fradiae GS 88, которые продуцируют тилозин ниже точки обнаружения и получены путем мутагенеза штамма С4. Нетрансформированные штаммы GS16, GS48, GS76 и GS88 продуцируют соответственно дефектный фермент или ограничивающее скорость реакций количество (1) tyl E, деметилмакроцин-0-метилтрансферазы, фермента; (2) фермента tyl D, который необходим для добавления или биосинтеза-6-деокси-Д-аллозы; (3) фермента tyl H,
который необходим для окисления С-23 метилового положения тилактона; и (4) фермента tyl J. Нетрансформированные штаммы GS 16, GS48, GS76 и GS88 соответственно имеют тенденцию к накоплению деметилмакроцина, демицинозилтилозина, 23 деоксидемицинозилтилозина и демицинозилтилозина, а не желаемых тилозиновых антибиотических соединений.
Плазмиды рНJL280 обеспечивают также средство увеличения эффективности биосинтеза тилозина не только за счет недефектного гена, но также за счет увеличения числа воспроизведения генов биосинтеза tylD, tylE, tylH и tylJ, а также за счет увеличения внутриклеточного количества продукта, определяемых этими генами. Концентрация имеющегося генного продукта tyl E таким образом увеличивается, приводя в результате к повышенному количеству фермента, способного вызывать конверсию деметилмакролина в макроцин и в тилозин в процессе биосинтеза тилозина.
Аналогично этому концентрация имеющихся генных продуктов tylD, tylH и tylJ также увеличивается, приводя в результате к продуцированию повышенных количеств ферментов, способных обеспечивать ввод -6-деокси-D-аллозы и С-23 окисления предшественников тилозина. Ферментация Streptomyces fradiae GS 16/рНJL280, S. fradiae GS 48/рНJL280, S.fradiae GS76 рНJL280 и S.fradiae GS 88/рНJL280 в течение 144-168 ч приводит в результате к выходам тилозина, которые значительно выше выходов в случае нетрансформированных мутантных штаммов, продуцирующих небольшое количество тилозина. Такие трансформированные штаммы имеют более высокие концентрации специфических ферментов, кодируемых плазмидой рНJL280, чем родственный штамм С4, и таким образом служит дополнительным примером осуществления настоящего изобретения. Плазмида рНJL280 может использоваться для повышения способности к продуцированию тилозина любого организма, в котором генные продукты tylD, tylE, tylF, tyl Н или tylJ)(или любая их комбинация) присутствуют в количествах, ограничивающих скорость реакции биосинтеза тилозина.
Плазмида рНJL284 была использована для трансформации Streptomyces fradiae GS 52, мутантного штамма, продуцирующего небольшое количество тилозина, образованного от штамма С4, который продуцирует ограничивающие скорость реакции количества фермента, необходимого для биосинтеза или добавления микарозы к де-0-метил-лактеноцину. Таким образом, реакция биосинтеза тилозина Streptomyces fradiae GS 52 приводит к образованию десмикозина, а не желаемого антибиотика тилозина. Плазмида рНJL284 обеспечивает средство повышения эффективности синтеза за счет недефектного гена биосинтеза и за счет увеличения числа воспроизведения гена биосинтеза tylС. Концентрация имеющегося генного продукта tylC в трансформированном штамме таким образом увеличивается, приводя в результате к повышенному продуцированию фермента, способного обеспечивать желаемую реакцию присоединения. Таким образом, ферментация Streptomyces fradiae GS 52/рНJL284 в течение 144-168 ч приводит в результате к продуцированию такого количества тилозина, которое значительно превышает количество нетрасформированного мутантного штамма и которое приводит в результате к более высоким концентрациям фермента tylС, чем концентрации в родственном штамме С4. Плазмида рНJL284 используется также в способе данного изобретения с целью увеличения способности Streptomyces fradiae GS88, мутанта tylJ, продуцировать тилозин, и таким образом она может быть использована согласно данному способу для повышения способности продуцирования тилозина любого организма, в котором генные продукты tyl C, tylF или tyl J (или любая их комбинация) присутствуют в количествах, ограничивающих скорость реакции биосинтеза тилозина.
Плазмида рНJL309 содержит гены биосинтеза tylL и tylM, была использована в способе данного изобретения для повышения способности к продуцированию тилозина организма Streptomuces fradiae GS 33, Mутанта tyl L и GS 62, мутанта tyl М. Плазмида рНJL309 может использоваться также в способе данного изобретения для повышения способности продуцирования тилозина любого организма, в котором генные продукты tyl L или tyl M (или оба) присутствуют в количествах, ограничивающих скорость реакции биосинтеза тилозина.
Плазмида рНJL311 содержит гены биосинтеза tyl C, tyl F, tyl H, tyl J и tyl K, и таким образом она использована в способе данного изобретения с целью повышения способности к продуцированию тилозина у организма Streptomyces fradiae GS 52, мутанта tyl GS; 88, мутанта tyl J; GS 15 и GS 28, которые оба являются мутантами tyl F и GS 85 мутанта tyl K. Плазмида рНJL 311 может использоваться также в способе данного изобретения для повышения способности к продуцированию тилозина у любого организма, в котором генные продукты tyl C, tyl F, tyl H, tyl J или tyl K (или любая их комбинация) присутствуют в количествах ограничивающих скорость реакции биосинтеза тилозина.
Плазмида рНJL315 содержит гены биосинтеза tyl D, tyl E, tyl F, tyl Н и tyl J и таким образом она использовалась в способе данного изобретения для повышения способности к продуцированию тилозина у организма Streptomuces fradiae GS 48, мутанта tyl D; GS 88, мутанта tyl J; GS 16. мутанта tyl E. GS 76, двойного мутанта tyl D, tyl H; и GS 15 и GS 28, которые оба представляют собой мутанты tyl F. Плазмида рНJL315 может быть также использована в способе данного изобретения для повышения способности к продуцированию тилозина у любого организма, в котором генные продукты tyl D, tyl E, tyl F, tyl H или tyl J (или любая их комбинация) присутствуют в количествах, ограничивающих скорость реакции биосинтеза тилозина.
Данные результаты показывают, что векторы, отвечающие настоящему изобретению, могут увеличивать способность к продуцированию антибиотика у образующего антибиотик организма за счет более высокой концентрации фермента или другого генного продукта, который является ограничителем скорости реакции в процессе биосинтеза антибиотика, по сравнению с нетрансформированным организмом.
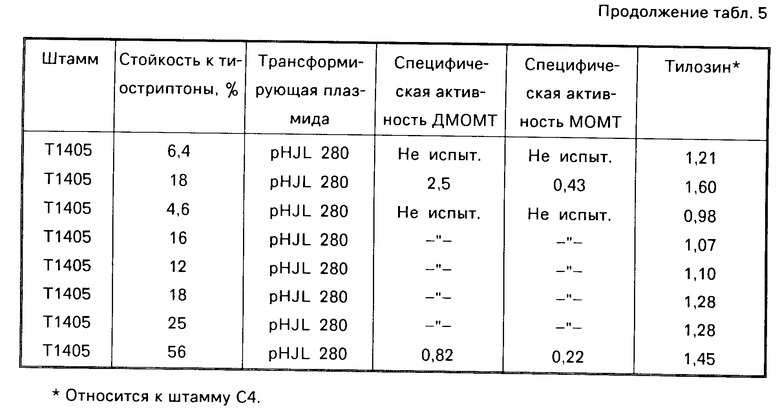
Для устранения возможности отрицательного влияния рекомбинантной плазмиды (либо автономно реплицируемой, либо интегрированной) на продуцирование тилозина можно использовать способность Streptomuces fradiae поглощать предшественники тилозина из питательной среды культивации и превращать их в тилозин. В одной ферментации продуцирующего тилозин штамма, который был трансформирован плазмидой рНJL280 и культивирован для получения интегрантов, лишь субпупуляция ( ≈18%) клеток была стойкой к тиострептону, что указывает на присутствие последовательности плазмида рНJL280. Однако эта субпупуляция содержит множество копий генов для двух ограничивающих скорость реакций ферментов, деметил-макроцин-0-метилтрансферазы (ДМОМТ) и макроцин-0-метилтрансферазы (МОМТ), и следовательно имеет повышенную концентрацию (примерно 9-кратную) этих двух ферментов, и в состоянии превращать все номинальное накопление диметилмакроцина и макроцина в тилозин (см. табл. 5).
Таким образом, можно получить специфические штаммы S.fradiae, содержащие множество копий, ограничивающих скорость реакции генов и высокое содержание ферментов для действия их как трансформаторов аккумулированных предшественников в тилозин. Эти штаммы трансформаторов могут использоваться различными путями:
(1) штамм трансформатор может быть введен в ферментер совместно с нормальным продуцирующим штаммом при низком отношении штамма трансформатора к продуцирующему штамму;
(2) штамм трансформатор может быть введен в среду ферментации в более позднем цикле для превращения в промежуточные продукты (3) штамм трансформатор может находиться в отдельном "реакторе", в который будет добавляться питательный бульон ферментации от продуцирующего штамма или (4) штамм трансформатора может быть иммобилизован на колонке, через которую может быть пропущен бульон ферментации от продуцирующего штамма. Для специалистов в данной области должно быть ясно, что популяции отдельного продуцирования и трансформации устраняют отрицательные влияния, которые иногда рекомбинантные плазмиды оказывают на продуцирование антибиотиков в случае высокопродуцирующих штаммов.
Тилозин состоит из 16 членного разветвленного лактона (тилонолид), к которому подсоединены три молекулы сахара (микароза, микаминоза и мициноза). Данный лактон образован от двух ацетатов, пяти пропионатов и бутирата путем конденсации молекулы пропионил-S-коэнзима А с двумя молекулами малонил-S-коэнзина А, четырьмя молекулами метилмалонил-S-коэнзима А по схеме, аналогичной схеме биосинтеза жирной кислоты.
Образование лактона, биосинтеза присоединение сахара и конверсия полученных в результате промежуточных соединений в тилозин катализируются рядом ферментов. Клонирующие гены, которые кодируют такие ферменты, обеспечивают модифицикацию и увеличение эффективности процесса биосинтеза тилозина и которые могут быть использованы при осуществлении способа данного изобретения, включают, например, гены tyl C, tyl D, tyl E, tyl F, tyl J, tyl K, tyl L и tyl М.
Из этой группы ген tyl F является предпочтительным ввиду того, что кодированный таким путем фермент макроцин-0-метилтрансфераза является ограничителем скорости реакции в процессе биосинтеза тилозина для большинства продуцирующих тилозин штаммов. Макроцин аккумулируется до неприемлемых концентраций в условиях оптимальной ферментации Streptomyces fradiae ввиду ограничивающих скорость реакции этапов, катализированных генным продуктом tyl F, фермент tyl F катализирует конверсию макроцина в тилозин, как представлено на фиг. 1. Чрезмерное продуцирование генного продукта tyl F макроцин-0-метилтрансферазы приводит в результате к более эффектному процессу биосинтеза тилозина, что определяется по повышенному выходу антибиотика и пониженной стоимости ферментаций.
Для специалистов в данной области должно быть ясно, что настоящее изобретение не ограничивается использованием плазмид рНJL280, рНJL284, рНJL309, рНJL311 или рНJL315. Гены биосинтеза антибиотиков, содержащиеся в этих векторах, могут быть экспрессированы всей группой или по отдельности и клонированы в различные подходящие векторы. Так, например, расщепление плазмиды рНJL280 ферментами BamHI и Bgl 11 дает пять фрагментов BamHI-BamHI размерами ≈10,3 кб ≈6,54 кб 2,3 кб ≈1,7 кб ≈1,0 кб; два фрагмента BamHI-BglII размерами ≈2,9 кб и 2,0 кб; и один фрагмент BglII-BglII размером ≈0,2 кб. Фрагмент ≈2,9 кб BamHI-BglII плазмиды рНJL280 содержит ген tylF. Расщепление плазмиды рНJL280 ферментами BglII и EcoRI генерирует четыре фрагмента фрагмент ≈11,24 кб ЕсоRI-EcoRI; фрагмент ≈11,5 кб BglII-EcoRI; фрагмент ≈4,0 кб ЕсоRI-BglII и фрагмент ≈2,0 кб BglII-BglII; фрагмент ≈4,0 кб EcoRI-BglII плазмиды рHJL280 содержит ген типа tyl E.
Расщепление плазмиды рНJL284 ферментами BamHI и ЕсоRI генерирует три фрагмента BamHI-BamHI размерами ≈9,7 кб ≈2,3 кб и ≈1,0 кб; и четыре фрагмента ЕсоRI-BamHI размерами ≈6,24 кб, ≈4,3 кб, ≈2,3 кб и ≈1,1 кб. Фрагмент ≈2,3 кб BamHI-EcoRI плазмиды рНJL284 содержит ген tyl F. Расщепление плазмиды рНJL284 ферментом EcoRI генерирует два фрагмента размерами ≈16,4 кб и 10,54 кб фрагмент ≈16,4 кб содержит гены tyl F, tyl C, tyl J.
Фрагмент ≈1,7 кб EcoRI-BamHI плазмиды рНJL311 включает ген tylK фрагмент ≈18,5 кб ЕсоRI, а также фрагмент ≈8,3 кб BamHI-КрnI плазмиды pHJL309 содержит гены tyl L и tyl M.
Любой из указанных выше содержащих ген tyl фрагментов может быть лигирован с другими векторами, образуя векторы, используемые согласно настоящему изобретению. Эти другие векторы включают, например, те векторы, которые описаны в патентах США NN 4 468 462; 4 513 086; 4 416 994; 4 503 155; и 4 513 185, а также плазмиды pIJL101, pIJL350, pIJL702 (АТСС 39155), SСР2Х (NRRL 15041), рНJL192, рНJL197, рН198, рНJL210, рНJL211, рНJL400, рНJL401, рНJL302, pIJ922, pIJ903, рIJ941, pIJ940 и pIJ916. Эти векторы воспроизводятся в Streptomyces fradiae и других продуцирующих тилозин штаммах и таким образом используются для клонирования, отвечающих данному изобретению генов биосинтеза антибиотиков.
В качестве примеров штаммов Streptomyces, которые могут использоваться для осуществления целей данного изобретения, являются S.fradiae, S.fradiae GS 52, S.fradiae GS 48, S.fradiae GS 16, S.fradiae GS 28, S.fradiae GS 15, S.fradiae GS 76.
Из числа Streptomyces штаммы, S.fradiae GS 16, S.fradiae GS 15 и S.fradiae GS 28 являются предпочтительными, особенно для трансформации плазмидой pHJL280. Streptomyces fradiae является хорошо известным микроорганизмом, и некоторые штаммы уже могут поставляться без особых ограничений из Северной Региональной Научно-Исследовательской лаборатории (NRRL). Пеория, Иллюнойс 61604 и из АТСС под соответствующими регистрационными номерами NRRL 2702, NRRL 2703 и АТСС 19609.
Рекомбинантные плазмиды, описанные в данной патентной заявке, включают каждая один из несколько генов биосинтеза антибиотиков. Если не считать часть полицистрона, то каждый ген биосинтеза антибиотика обычно включает (1) стимулятор, который направляет транскрипцию гена (2) последовательность, которая при транскрипции в информационную РНК направляет трансляцию транскрипта; (3) кодирующую белок последовательность, ии (4) завершитель транскрипции. Каждый из этих элементов используется независимо и посредством технологии рекомбинантной ДНК может служить для образования огромной разновидности рекомбинантных генов. Так, например, кодирующая белок последовательность гена tyl F может быть связана со стимулятором, активирующей трансляцию последовательностью и с завершающей транскрипцию последовательностью от не Streptomyces fradiae гена, образуя рекомбинантный ген, который функционирует в хозяине, из которого выделены последовательности не S.fradiae. Такой новый ген может быть использован для продуцирования гибридного антибиотика при вводе его в организм, который продуцирует антибиотик или антибиотический промежуточный продукт, который не обнаруживается в синтезе тилозина, но который служит в качестве субстрата для нового генного продукта. Аналогично этому стимулятор и другие регуляторные элементы гена tyl F могут быть связаны с кодирующей последовательностью гена биосинтеза нетилозинового антибиотика, образуя гибридный ген, который будет функционировать S.fradiae. Так, индивидуальные элементы каждого из генов биосинтеза антибиотиков описанных плазмид включают очень важный компонент данного изобретения.
Так, например, данные ряда на нуклеотидной последовательности tylF позволили идентифицировать стимулятор tyl F, который является очень важным аспектом данного изобретения. Данная последовательность (представлена для удобства лишь одна цепь этой последовательности) показана ниже. Стимулятор и активирующая трансляцию последовательность гена tyl F сохраняются в данной последовательности между остаточными группами 1 и 207. Данная последовательность завершается началом кодирующей зоны гена tyl F.
Дополнительная генная последовательность для гена tyl F. В частности ген tyl F в целом, включающий стимулятор и активирующую трансляцию последовательность, о которых упоминалось выше, имеет указанную ниже последовательность:
5'-TTC GCG GGA  G ATG CTG A
G ATG CTG A CGG GGG TC
CGG GGG TC GCC AGC AGC
GCC AGC AGC  C CGG ACG T
C CGG ACG T TCT GGC GG
TCT GGC GG AGA TCA GCC
AGA TCA GCC  A CCG GCG C
A CCG GCG C CGT CCC AC
CGT CCC AC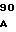 GCT CGG CCC
GCT CGG CCC 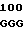 CGA TCG G
CGA TCG G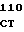 CCT CCG C
CCT CCG C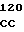 GGA GGG
GGA GGG 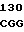 CGT ACT GCT C
CGT ACT GCT C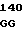 GAG GGC TG
GAG GGC TG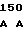 GG GAC AGG
GG GAC AGG 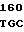 GGG CGA C
GGG CGA C GCC AGG CG
GCC AGG CG GC TGC GCC
GC TGC GCC  CTG CGG C
CTG CGG C CGT CGG TG
CGT CGG TG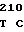 GT TGG CGC
GT TGG CGC 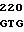 CTG CGG G
CTG CGG G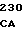 ACA GAA TC
ACA GAA TC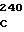 CCT TTT GTG
CCT TTT GTG 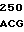 GGC GGG C
GGC GGG C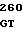 CCC CGG AC
CCC CGG AC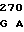 GG ACA CGA
GG ACA CGA 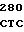 GCT GCG G
GCT GCG G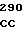 TCA ACG AA
TCA ACG AA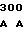 CA CCG TGT
CA CCG TGT 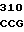 GTG CCC A
GTG CCC A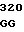 CCA CGA AC
CCA CGA AC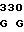 TG ACC GGT
TG ACC GGT 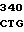 TGT CAG G
TGT CAG G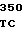 GCC CGT GG
GCC CGT GG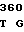 AC GGG CTC
AC GGG CTC 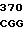 GGC GGC G
GGC GGC G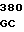 GCG GGC GG
GCG GGC GG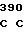 GA CCT TGA
GA CCT TGA 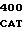 ACC CGC G
ACC CGC G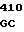 CGG GCT CC
CGG GCT CC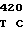 GT TCC GGC
GT TCC GGC 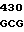 GCC CGC G
GCC CGC G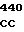 GAT AGC GT
GAT AGC GT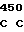 GT CCT CAC
GT CCT CAC 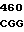 CTC CGG C
CTC CGG C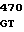 CCG CGT CC
CCG CGT CC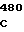 CGC CGG GAC
CGC CGG GAC  CCA CCT C
CCA CCT C CCG ACC CC
CCG ACC CC GA GCC GAT
GA GCC GAT  CCC GCT A
CCC GCT A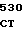 GGA GGA CC
GGA GGA CC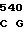 TG GCA CCT
TG GCA CCT 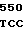 CCG GAC C
CCG GAC C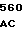 GCC CGC GA
GCC CGC GA TC TAC
TC TAC
VAL ALA PRO SER  RO ASP HIS ALA ARG
RO ASP HIS ALA ARG  P LEU TYR ATC
P LEU TYR ATC  CTG CTG AAG A
CTG CTG AAG A GTC GT
GTC GT CG AAC GTC
CG AAC GTC  TAC GAG G
TAC GAG G CCC ILE GLU LEU
CCC ILE GLU LEU 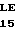 U LYS LYS VAL
U LYS LYS VAL 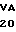 L SER ASN VAL ILE
L SER ASN VAL ILE 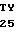 R GLU ASP PRO ACC CA
R GLU ASP PRO ACC CA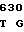 TG GCG GGG
TG GCG GGG 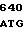 ATC ACC G
ATC ACC G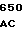 GCG TCG TT
GCG TCG TT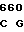 AC CGG ACG
AC CGG ACG 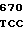 THR
THR 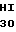 S VAL ALA GLY MET
S VAL ALA GLY MET 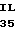 E THR ASP ALA SER
E THR ASP ALA SER 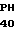 E ASP ARS THR SER CGT GAG A
E ASP ARS THR SER CGT GAG A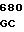 GGC GAG GA
GGC GAG GA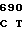 AC CCC ACG
AC CCC ACG 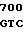 GCC CAC A
GCC CAC A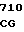 ATG ATC GG
ATG ATC GG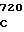
 G GLU SER GLY GLU
G GLU SER GLY GLU 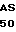 P TYR PRO THR VAL
P TYR PRO THR VAL 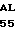 A HIS THR MET ILE
A HIS THR MET ILE 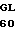 Y CTC AAG CGT
Y CTC AAG CGT 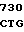 GAC AAT C
GAC AAT C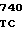 CAC CGG TG
CAC CGG TG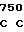 TC GCG GAC
TC GCG GAC 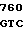 GTG GAG LEU LYS ARG LEU
GTG GAG LEU LYS ARG LEU 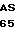 P ASN LEU HIS ARG
P ASN LEU HIS ARG  S LEU ALA ASP
S LEU ALA ASP  L VAL GLU G
L VAL GLU G CGC GTC CC
CGC GTC CC GT GAC TTC
GT GAC TTC 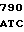 GAG ACC G
GAG ACC G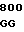 GTG TGC CG
GTG TGC CG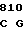 CG CCG ASP GLY VAL
CG CCG ASP GLY VAL 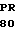 O GLY ASP PHE ILE
O GLY ASP PHE ILE  U THR GLY VAL CYS
U THR GLY VAL CYS  G ALA PRO TGC
G ALA PRO TGC 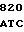 TTC GCC C
TTC GCC C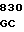 GGA CTG CT
GGA CTG CT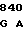 AC GCG TAC
AC GCG TAC 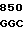 CAG GCC G
CAG GCC G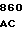 CGC CYS ILE
CGC CYS ILE 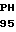 E ALA ARG GLY
E ALA ARG GLY 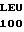 LEU ASN ALA TYR GLY
LEU ASN ALA TYR GLY 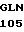 ALA ASP ARG ACC GT
ALA ASP ARG ACC GT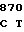 GG GTC GCC
GG GTC GCC 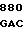 TCC TTC C
TCC TTC C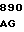 GGC TTT CC
GGC TTT CC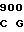 AG CTG ACC
AG CTG ACC 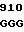 THR
THR 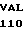 TRP VAL ALA ASP
TRP VAL ALA ASP 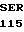 PHE GLN GLY PHE
PHE GLN GLY PHE 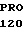 GLU LEU THR GLY TCC GAC C
GLU LEU THR GLY TCC GAC C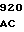 CCG CTG G
CCG CTG G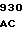 GTC GAG ATC
GTC GAG ATC 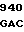 CTC CAC C
CTC CAC C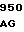 TAC AAC GA
TAC AAC GA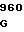
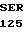 ASP HIS PRO LEU
ASP HIS PRO LEU 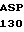 VAL GLU ILE ASP
VAL GLU ILE ASP  HIS GLN TYR ASN
HIS GLN TYR ASN 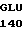 GCC GTG GAC
GCC GTG GAC 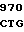 CCC ACC A
CCC ACC A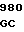 GAG GAG AC
GAG GAG AC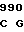 TG CGG GAG
TG CGG GAG 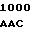 TTC GCC ALA VAL ASP LEU
TTC GCC ALA VAL ASP LEU  THR SER GLU GLU
THR SER GLU GLU  VAL ARG GLU ASN
VAL ARG GLU ASN 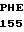 ALA C
ALA C AC GGG CT
AC GGG CT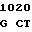 C GAC GAC
C GAC GAC 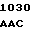 GTC CGT TT
GTC CGT TT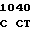 G GCG GG
G GCG GG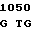 G TTC ARG TYR GLY
G TTC ARG TYR GLY  LEU ASP ASP ASN
LEU ASP ASP ASN 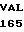 ARG PHE LEU ALA
ARG PHE LEU ALA 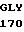 TRP PHE AAG
TRP PHE AAG 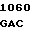 ACC ATG C
ACC ATG C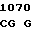 CT GCG CC
CT GCG CC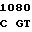 G AAG CAG
G AAG CAG  GCG GTG A
GCG GTG A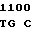 GC LYS ASP
GC LYS ASP  MET PRO ALA ALA
MET PRO ALA ALA  VAL LYS GLN LEU
VAL LYS GLN LEU  VAL MET ARG CTG GA
VAL MET ARG CTG GA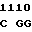 C GAC TCC
C GAC TCC  GGC GCC A
GGC GCC A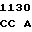 TG GAT GT
TG GAT GT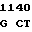 C GAC AGC
C GAC AGC  LEU
LEU 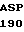 GLY ASP SER TYR
GLY ASP SER TYR 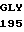 ALA THR MET ASP
ALA THR MET ASP 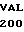 LEU ASP SER LEU TAC GAG C
LEU ASP SER LEU TAC GAG C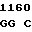 TG TCG CC
TG TCG CC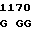 C GGT TAC
C GGT TAC  ATC GTC G
ATC GTC G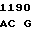 AC TAC TG
AC TAC TG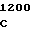
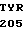 GLU ARG LEU SER
GLU ARG LEU SER 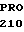 GLY GLY TYR VAL
GLY GLY TYR VAL 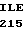 VAL ASP ASP TYR
VAL ASP ASP TYR 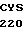 ATC CCG GCC
ATC CCG GCC  CGC GAG C
CGC GAG C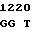 GC ACG AC
GC ACG AC C GCG ACC
C GCG ACC 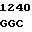 TCG GCA ILE PRO ALA CYS
TCG GCA ILE PRO ALA CYS 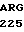 GLU ARG CYS THR
GLU ARG CYS THR 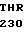 SER ALA THR GLY
SER ALA THR GLY 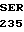 ALA T
ALA T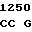 CG ACA CG
CG ACA CG C ACC GGA
C ACC GGA  ACC GCC A
ACC GCC A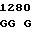 CG CTA TT
CG CTA TT G GCA SER ALA THR
G GCA SER ALA THR 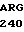 SER THR GLY SER
SER THR GLY SER 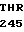 ALA ARG ALA LEU
ALA ARG ALA LEU 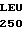 ALA ALA CAG
ALA ALA CAG 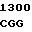 CTG AGT C
CTG AGT C CC GCC CG
CC GCC CG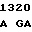 G CCC GAC
G CCC GAC 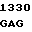 AGC AGG A
AGC AGG A AT GLN ARG
AT GLN ARG  SER ARG SER ALA
SER ARG SER ALA 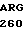 GLU PRO ASP GLU
GLU PRO ASP GLU 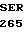 ARG ARG TYR GCG AG
ARG ARG TYR GCG AG C GAC GCG
C GAC GCG  GCT CGG C
GCT CGG C GA GGG AG
GA GGG AG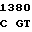 G GGT GAT
G GGT GAT  ALA
ALA 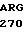 HIS ASP ALA PRO
HIS ASP ALA PRO 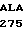 ARG HIS GCC GGA G
ARG HIS GCC GGA G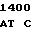 CA TCC GG
CA TCC GG G GCG CGG
G GCG CGG 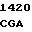 GTT CCA C
GTT CCA C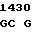 TG GTT CC
TG GTT CC
GAG CCA GCC 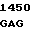 TTC CGG C
TTC CGG C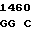 TG ACC GG
TG ACC GG C TCC TTC
C TCC TTC  GTG CCG
GTG CCG
C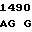 TC GTC AA
TC GTC AA C GCG TGT
C GCG TGT  GGA AAG G
GGA AAG G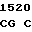 CG CTG CG
CG CTG CG T CCA
T CCA
CTT  CCG AGG T
CCG AGG T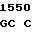 AC CGG GC
AC CGG GC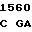 G GCC AAG
G GCC AAG 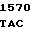 AGC GGC G
AGC GGC G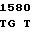 GT
GT
GCA GG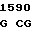 C CGG TGT
C CGG TGT 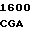 GGT CGT C
GGT CGT C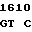 GA CGC GC
GA CGC GC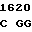 T GTC GAG
T GTC GAG 
GTC GTC G '
'
где А представляет собой группу дезоксиаденила. G группу деоксигуанила, С группу деоксицитидила и Т тимидиловую группу. Данный структурный ген, как указывалось выше, начинается с группы 541 и продолжается до группы 1371, заканчиваясь стоп кодоном, расположенным у группы 1372. Аминокислотная последовательность данного структурного гена tyl F это последовательность, которая указана под соответствующей нуклеотидной последовательностью. Ввиду вырождения данного генетического кода могут быть получены последовательности, эквивалентные тем, которые специально указаны выше, которые кодируют тот же самый генный продукт tyl F. Этот способ получения таких эквивалентных последовательностей должен быть знаком для специалистов в данной области. Однако данная специфическая последовательность никак не должна рассматриваться как ограничение настоящего изобретения.
Штаммы Streptomyces fradiae могут быть культивированы рядом способов с использованием различных сред. Предпочтительными углеводными источниками в среде культивации являются, например, меласса, глюкоза, декстран и глицерил, предпочтительными азотными источниками являются, например, соевая мука, смеси аминокислот и пептоны. Питательная среда содержит также неорганические соли, которые включают выпускаемые промышленностью соли, способные образовывать йоны фосфатов, хлоридов сульфатов и т.д. натрия, калия, аммония, кальция. При необходимости для роста и образования других микроорганизмов вводятся основные микрокомпоненты. Такие микрокомпоненты вводятся как случайные примеси в добавление к другим составляющим компонентам питательной среды. Штаммы S.fradiae выращиваются в аэробных условиях с широким пределом величины рН примерно от 5,5 до 8, при температуре в пределах примерно от 25оС до 37оС. В частности, тилозин может быть получен путем культивации продуцирующих тилозин штаммов, например S.fradiae таких как штаммы, содержащие векторы, отвечающие настоящему изобретению. Используемая питательная среда может быть любой из числа различных сред, поскольку данный организм способен потреблять многие источники энергии. Однако, из соображений экономичности продуцирования максимальных выходов антибиотика и легкости извлечения антибиотика предпочтительны определенные среды культивации. Среда, которая особенно полезна для продуцирования тилозина, включает ассимилируемый источник углерода, такой как глюкоза, сахароза, фруктоза, крахмал, глицерин, меласса, декстрин, бурый сахар, кукурузная дробина, и т.д. Предпочтительными источниками углерода являются глюкоза и крахмал. Кроме того, питательная среда культивации включает источник ассимилируемого азота, такой как льняная мука, отстой брожения, рыбная мука, мука семян хлопка, овсяная мука, пшеничная мука, мука соевых бобов, мясной экстракт, пептоны (мяса или сои), казеин, смеси аминокислот, т.д. Предпочтительными источниками азота являются мука соевых бобов, казеин и кукурузная дробина.
Положительный эффект достигается при вводе в питательную среду минеральных солей, например, солей, обеспечивающих ионы сульфата, хлорида, фосфата, карбоната, ацетата и нитрата натрия, калия, аммония, кальция, магния, кобальта, а также источников факторов роста, таких как барда и дрожжевой экстракт.
В питательную среду для роста микроорганизмов, используемых согласно настоящему изобретению, должны быть введены также основные микрокомпоненты, обеспечивающие рост и развитие других микроорганизмов. Такие микрокомпоненты обычно вводятся как случайные примеси в добавление к другим составляющим компонентам среды.
Начальная величина рН среды культивации может изменяться в широких пределах. Однако установлено что желательная величина рН питательной среды находится в интервале примерно от 5,5 до 8,0 и предпочтительная величина рН составляет примерно от 6,5 до 7,0. Как установлено для случая других организмов, величина рН среды постепенно увеличивается в ходе периода роста организма, и в течение этого времени образуется тилозин, и может достигаться величина рН примерно от 7,2 до 8,0 или более и конечная величина рН зависит по крайней мере частично от начальной величины рН среды, буферных систем, присутствующих в питательной среде, и от периода времени, в течение которого происходит рост организма.
Аэробные условия культивации с погружением являются условиями выбора продуцирования больших количеств тилозина. Для получения относительно небольших количеств могут использоваться взбалтываемые колбы и поверхностная культура в склянках, но для получения больших количеств предпочтительны аэробные условия погружения культуры в стерильные емкости. Среда в этой стерильной емкости может быть инокулирована спорулированной суспензией. Однако ввиду задержки роста в случае использования спорулированной суспензии в инокулуме растительная форма культуры является предпочтительной для исключения явной задержки роста позволяющей более эффективно использовать оборудование для брожения. Таким образом, желательно прежде всего продуцировать растительный инокулум организмов путем инокулирования относительно небольшого количества питательной среды культивации споровой формой организма, и в случае получения молодого активного растительного инокулума асептически переносить растительный инокулум в большую емкость. Среда, в которой продуцируется растительный инокулум, может быть той же самой или иной средой, чем та, которая используется для продуцирования тилозина в больших масштабах.
Данные организмы наилучшим образом выращиваются при температурах в интервале примерно от 25 до 37оС. Оптимальное продуцирование тилозина происходит при температуре примерно 26-30оС.
При осуществлении обычной обработки погруженной культуры через данную среду культивации продувается стерильный воздух. Для эффективного роста организма и продуцирования тилозина объем воздуха, используемого в емкости продуцирования тилозина составляет более 0,1 объема в минуту на объем среды культивации. Эффективный рост и оптимальные выходы тилозина получаются в тех случаях, когда объем используемого воздуха составляет не менее одного объема в минуту на объем среды культивации.
Концентрацию тилозина в среде культивации можно легко проследить в период брожения путем испытания образцов в среде культивации на их ингибирующее действие против роста известного организма, который ингибируется в присутствии тилозина.
Обычно после инокулирования максимальное продуцирование тилозина происходит в течение примерно 2-7 дней при использовании погруженной аэробной культуры или культуры во встряхиваемой колбе, и в течение примерно 5-10 дней при использовании поверхностной культуры.
При желании мицелий и нерастворенные твердые вещества удаляются из питательного бульона ферментации обычными способами, такими как фильтрация или центрифугирование. При желании тилозин удаляется из отфильтрованного или центрифугированного питательного бульона способами адсорбции или экстракции, которые известны для специалистов в данной области.
Для экстракции тилозина из отфильтрованного питательного бульона используются предпочтительно несмешиваемые с водой полярные органические растворители, например включающие сложные эфиры жирных кислот, такие как этилацетат и амилацетат, хлорированные углеводороды, например хлороформэтилендихлорид и трихлорэтилен, смешиваемые с водой спирты, например бутиловый и амиловый спирты, несмешиваемые с водой кетоны, например метилизобутилкетон и метиламилкетон, и другие растворители, например, простой диэтиловый эфир и простой метилпропиловый эфир. Могут использоваться также другие растворители аналогичного типа. Предпочтительными растворителями экстракции являются хлороформ и амилацетат.
Для извлечения тилозина методом адсорбции могут использоваться различные абсорбенты и ионообменные смолы, например уголь, силикагель, окись алюминия и ионообменные смолы кислого характера, такие как "ХЕ" и 64 и "IRC" 50 (слабо кислые катионообменные смолы, выпускаемые фирмой "Rohm and Haas Company) карбоксиметилцеллюлозная смола и "Dewex" 50 (сильно кислая катионообменная смола, выпускаемая фирмой "Dow Chemical Company). Тилозин может адсорбироваться на одном из указанных выше или на аналогичных адсорбентах из раствора в хлороформе, ацетоне, бензоле или других подходящих растворителей. Адсорбированный тилозин может быть растворен в соответствующих растворителях.
Затем адсорбированный тилозин элюируется из адсорбента соответствующими способами элюирования, такими как промывка адсорбента, на котором адсорбирован тилозин, низшим спиртом, таким как метанол или этанол, или низшим спиртом, содержащим до примерно 50% низшего кетона, например ацетона.
Экстракт органического растворителя, полученный предпочтительным способом экстракции, может непосредственно выпариваться досуха до получения сырого тилозина. Кроме того, экстракт органического растворителя может использоваться для получения очищенного тилозина путем выпаривания в вакууме экстракта тилозина в органическом растворителе, путем обесцвечивания концентрата углем, и путем осаждения тилозина с добавлением неполярного растворителя, например петролейного эфира. Полученный таким путем осадок представляет собой твердый продукт, очищенный тилозин, который обычно является аморфным веществом. Этот аморфный осадок может кристаллизоваться с использованием одного из кристаллизующих растворителей, указанных выше. Кроме того, тилозин может быть извлечен из содержащего тилозин органического экстракта, путем абсорбционной хроматографии, путем вывода абсорбированного тилозина из абсорбента элюированием.
Другие способы получения желаемого продукта из среды культивации должны быть уже известны для специалистов в данной области. Кислые аддитивные соли тилозина могут быть образованы неорганическими кислотами, например серной, соляной и азотной кислотой, и органическими кислотами, например винной, глюконовой, щавелевой и уксусной кислотой. Кислые аддитивные соли могут быть получены путем растворения свободного основания тилозина в растворителе, в котором этот тилозин растворим, таком как ацетон или простой эфир, и ввода в раствор равномолярного количеств подходящей кислоты. Соль, которая при этом образуется, обычно выпадает в осадок из раствора. В случае, когда соль не выпадает в осадок, она может быть извлечена путем испарения раствора до меньшего объема, обеспечивающего осаждение, или путем ввода смешивающегося растворителя, в котором эта соль нерастворима.
П р и м е р 1. Выделение плазмиды рНJL280
А.Культура Е.coli К12 НВ 101/рНJL280
Лиофилы Е coli К12 НВ 101/рНJL280 могли быть получены из коллекции NRRL под порядковым номером NRRL В-18043. Лиофилизированные клетки наносились на чашки с L-агаром (L-агар содержал 10 г. Триптона Бакто, 5 г дрожжевого экстракта Бакто, 10 г NaCl, 2 г. глюкозы и 15 г агара на литр), содержащим 50 мкг/мл ампициллина для получения одиночной колонии изолята Е.coli К 12 НВ 101/рНJL280. Одна такая колония использовалась для инокулирования 100 мл питательного бульона L (L питательный бульон представляет собой L агар без агара), который затем инкубировался в аэробных условиях при температуре 37оС в течение ночи (примерно 16 ч). На следующее утро клетки извлекали путем центрифугирования со скоростью 10 000 об/мин в течение 10 мин. Примерно 1,0 г клеток, полученных в результате такой процедуры, использовались для получения плазмиды рНJL280 ДНК согласно описанной ниже процедуре.
В. Выделение плазмиды. Клеточную гранулу, полученную в примере 1А. повторно суспензировали в 10 мл раствора, состоящего из 25% сахарозы и 50 ммол Трис-НСl при величине рН 8,0. В эту клеточную суспензию вводили примерно 1 мл раствора лизозима концентрацией 10 мг/мл в 50 ммол Трис-НСl, и полученную смесь инкубировали на льду в течение 5 мин. Затем в эту клеточную суспензию вводили примерно 4 мл 0,215 мол этилендиаминтетрауксусной кислоты (ЕДТА), рН 8,0 и инкубирование на льду продолжалось еще в течение 5 мин. В обработанные лизозимом клетки вводили примерно 16 мл лизисного раствора (лизисный раствор содержал 0,4% деоксихолят; 1% Brij 58 Sigma Chemical Co.P. 0. Вох 14508, шт. Миссури, Санта Луис, 63178); 0,05 Мол. Трис-НСl, рН 8,0; и 0,0625 Мол. ЕДТА), и полученную смесь инкубировали при температуре 37оС в течение 15 минут.
Клеточный лизат осветляли путем центрифугирования со скоростью 48000 об/мин в течение 25 мин. Всплывший слой декантировали в отдельную трубку, в которую добавляли 0,1 объема 3,0 мол ацетата натрия (NaOAc) при величине рН 8,0 и 0,64 объема изопропилового спирта. Осажденную ДНК извлекали путем центрифугирования со скоростью вращения центрифуги 20000 об/мин в течение 10 мин. Затем снова растворяли в 01, объеме буферного раствора ТЕ (10 ммол. Трис-НСl, рН 7,8, и 1 ммол ЕДТА). Раствор ДНК инкубировали при 65оС в течение 30 мин, затем очищали путем центрифугирования с равновесным перепадом плотности в CsCl и в дийодиде пропидия. Полученную таким образом плазмиду рНJL280 ДНК растворяли в буфере ТЕ до концентрации примерно 1 мкг/мкл. Карта ограничительной точки плазмида рНJL280 представлена на фиг. 2, приложенном к данному описанию.
П р и м е р 2. Выделение плазмид рНJL284, рНJL309, рНJL311 и рНJL315.
Лиофилы штаммов E. coli, заключающие в себе плазмиды рНJL284, рНJL309. рНJL311 и рНJL315, могут быть получены из коллекции NRRL порядковых номеров, указанных в табл. 10. Каждую из желаемых плазмид получали и очищали от лиофилизированных клеток согласно описанию примера 1. Карты ограничительных точек этих плазмид представлены на рисунках 2-6, приложенных в настоящей патентной заявке.
П р и м е р 3. Построение Streptomyces fradiae GS 28/рНJL280.
Культура Streptomyces fradiae GS28 инокулировалась в 20 мл триптиказного питательного бульона (TSB) и инкубировалась в водяной бане при 29оС при вращении мешалки 260 об/мин в течение ночи (примерно 16 ч). Эта культура гомогенизировалась с использованием гомогенизирующего сосуда (Томас Сайнтифик, Сведсборо, Нью Джерси) и лабораторной мешалки Т-Line, а затем фрагметировалась посредством клеточного разрывателя (Хит Системс Ультразоникс Инк) в течение 7 с при мощности 76 Вт. Четыре миллилитра гомогенизированной фрагментированной культуры инокулировались в 20 мл TSB (BBh), содержащем 0,3 мас. /об. глицина, и эту культуру снова инкубировали в течение ночи при 29оC. На следующее утро культуру гомогенизировали и снова культивировали как описано выше. После третьей ночи инкубирования культуру гомогенизировали, собирали, а затем дважды промывали Р средой. Р среда приготавливалась путем ввода 103 г сахарозы в 0,25 г K2SO4 и 2,03 г MgCl2-6H2O и последующего ввода деионизированной воды до конечного объема 700 мл. Затем смесь стерилизовали, и в каждые 70 мл раствора вводили примерно по 10 мл каждого из следующих реагентов 0,05 г КН2РО4/100 мл дейонизированной воды; 2,78 г СаСl2/100 мл деионизированной воды, и 0,25 Мол ТЕS (2-([трис-(оксиметил)метил]-амино)этансульфокисло- ты) при величине рН 7,2.
Данную клеточную гранулу снова суспензировали в 15 мл среды Р, содержащей 1 мг/мл лизозима (Cobbiochem, La Jolla, Калифорния 92037) и затем инкубировали при комнатной температуре в течение примерно полутора часов до образования протопластов. Эти протопласты осторожно извлекали путем центрифугирования, промывали двукратно Р средой, снова суспензировали в 2 мл Р среды, и инкубировали на льду до использования.
Примерно 1 мкг плазмиды рНJL280 ДНК вводили примерно в 50 мкл гепаринсульфата концентрацией 1 мг/мл (Сигма) и инкубировали на льду в течение примерно 10 мин. Для трансформации Streoptomyces fradiae, если она получена от хозяина fradiae, можно использовать значительно меньше плазмидной ДНК, примерно 5-100 нг. Процедура выделения ДНК плазмиды Streptomyces описана в работе Hopwood и др. 1985 г. "Генетическая манипуляция Streptomyces": Лабораторное руководство (John Jnnes Foundation, Norwich, Англия). Сначала раствор ДНК (гепарин вводили примерно в 200 мкл протопластов и затем примерно 0,9 мл раствора, состоящего из 55% РЕG 1000 (Сигма) в Р среде вводили в смесь ДНК (протопласт, и полученную смесь осторожно перемешивали при комнатной температуре. Эту смесь высевали в различных аликвотах на чашках R2, используя 4 мл поверхностных слоев мягкого -R2 агара. R2-чашки содеpжали 30 мл среды R2 и были высушены при 37оС в течение примерно четырех дней. Среда R2 приготавливалась путем ввода 103 г сахарозы, 0,25 г K2SO4, 2 мл раствора микрокомпонентов, 10,12 г MgCl2- -6H2O, 10,0 г глюкозы, 2,0 г L аспарагина, 0,1 г Казаминовых кислоты и 22 г агара в 700 мл воды; стерилизации полученного в результате раствора, и в конечном счете ввода по 100 мл каждого из нижеследующих растворов: 0,05 г КН2РО4 (100 мл деионизированной воды; 2,22 г CaCl2/100 мл деионизированной воды и 0,25 мол ТЕS, рН 7,2. Величину рН конечного раствора доводили до 7,2. Раствор микрокомпонентов содержал 40 мг ZnCl2, 200 мг FeCl-6H2O, 10 мг CuCl2-2Н2О, 10 мг MnCl2 4H2O, 10 мг Na2B4O7 10H2O и 10 мг (NH4)6Mo7O24˙4H2O на 1 л. Поверхностные слои мягкого -R2- агара приготавливали путем ввода 51,5 г сахарозы, 5,06 г MgCl2-6H2O, 1,11 г СаСl2, 50 мл 0,25 мол. TES с величиной рН 7,2 и 2,05 г агара в достаточно йонизированную воду до достижения конечного объема 500 мл. Смесь обрабатывалась водяным паром для расплавления агара, декантировалась в 4 мл аликвоты и до использования обрабатывалась в автоклаве. После высевания трансформированных протопластов чашки инкубировались при 29оС в течение 24 ч и затем на протопласты распылялось 4 мл мягкого R2 агара, содержащего 25 мкл 50 мг/мл тиострептона (E.R. Sguibb, Princeton. Hью Джерси 08540). Инкубирование чашек при температуре 29оС продолжалось до тех пор, пока не прекращалась полностью регенерация, обычно в течение примерно 7-14 дней, для отбора желаемых трансформантов S.fradiae GS 28/рНJL280.
Штамм Streptomyces fradiae GS28 культивировали и продуцировали макроцин-0-метилтрансферазу и тилозин концентрациями выше тех, которые получались в случае нетрансформированного штамма S.fradiae GS 28. Активность макроцин-0-метилтрансферазы определяли в основном согласно описанию Jch и др. 1984 г. Journal of Chromatography, 288, 157-165. Сравнение активностей макроцин-0-метилтрансферазы в трансформированных, GS 28/рНJL280, и родственных, GS 28, штаммах показало 60-100-кратное увеличение продуцирования фермента и 14-18-кратное увеличение продуцирования тилозина в трансформированном штамме. Продуцирование тилозина определяли в основном согласно описанию Baltz и Seno 1981. Антимикробные агенты и Хемотерапия, 20, 214-225; Kennedy J.H. 1983 г. Journal of Chromatography, 281, 288-292.
П р и м е р 4. Построение Streptomyces fradiae SG15pHJL280.
Осуществляли построение желаемого штамма в основном согласно описанию примера 3 с той разницей, что использовался Streptomyces fradiae GS 15, а не S. fradiae GS 28. Желаемый штамм культивировался в течение 72 ч и продуцировались макроцин-0-метилтрансфераза и тилозин в концентрациях выше тех, которые получаются у нетрансформированного штамма, который продуцирует нелегко обнаруживаемый тилозин.
П р и м е р 5. Построение Streptomyces fradiae GS 15/pHJL284.
Осуществляли построение желаемого штамма в основном согласно описанию примера 4 с той разницей, что использовалась плазмида рНJL284, а не плазмида рНJL280. Желаемый штамм культивировался и продуцировались макроцин-0-метилтрансфераза и тилозин концентрациями выше тех, которые получались в случае нетрансформированного штамма S.fradiae GS 15.
П р и м е р 6. Построение Streptomyces fradiae GS 16/рНJL280.
Осуществляли построение желаемого штамма в основном согласно описанию примера 3 с той разницей, что использовался Streptomyces fradiae GS 16, а не S. fradiae GS 28. Желаемый штамм культивировался, и продуцировались генный продукт tyl E, деметилмакроцин-0-метилтрансфераза, и тилозин концентрациями выше тех, которые получались в случае нетрансформированного штамма. Активность деметилмакроцин-0-метилтрансферазы и продуцирование тилозина соответственно определяли в основном согласно описанным выше процедурам с той разницей, что макроцин был заменен деметилмакроцином в качестве субстрата.
П р и м е р 7. Построение Streptomyces fradiae GS 76/рНJL280.
Осуществляли построение желаемого штамма в основном согласно описанию примера 3 с той разницей, что использовался Streptomyces fradiae GS 76, а не S. fradiae GS 28. Желаемый штамм культивировался, и продуцировались генные продукты tyl D и tyl H концентрациями выше тех, которые получались в случае нетрансформированного штамма.
П р и м е р 8. Построение Streptomyces fradiae GS 48/pHJL280.
Осуществляли построение желаемого штамма в основном согласно описанию примера 3 с той разницей, что использовался Streptomyces fradiae GS 48, а не S. fradiae GS 28. Желаемый штамм культивировался, и продуцировались генный продукт tyl D и тилозин концентрациями выше тех, которые получались в случае нетрансформированного штамма.
П р и м е р 9. Построение Streptomyces fradiae GS52/рНJL284.
Осуществляли построение желаемого штамма в основном согласно описанию примера 3 с той разницей, что использовались Streptomyces fradiae GS 52 и плазмида рНJL284, а не S.fradiae GS 28 и плазмида рНJL280. Желаемый штамм культивировался и продуцировались генный продукт tyl C и тилозин концентрациями выше тех, которые получались в случае нетрансформированного штамма.
П р и м е р 10. Специфическая активность ограничивающих скорость процесса ферментов и повышенное продуцирование тилозина с использованием способа данного изобретения.
В нижеследующих таблицах представлены эффективности, достигаемые благодаря способу данного изобретения. Все трансформанты, приведенные в таблицах, получены в основном согласно процедуре, описанной в примере 3. Результаты, приведенные в табл. 12 и 13, получены при использовании штаммов, культивированных в ферментационной среде (Baltz иSeno 1981 г. Антимикробные агенты и Хемотерапия, 20, 214-225), которая содержала также 20 мкг/мл тиострептона, если культивированный штамм содержал плазмиду. Следует отметить, что трансформированные штаммы, приведенные в табл. 12 и 13, продуцировали небольшое количество тилозина или количество, которое не могло легко обнаруживаться, и эти штаммы культивировались при избирательном давлении (тиострептона) для сохранения плазмиды как автономно воспроизводящего вектора.
Результаты, представленные в табл. 14, получены от трансформантов, штаммов, высоко продуцирующих тилозин, которые культивировались после трансформации с получением интегрантов, трансформантов, в которых вся плазмидная ДНК или ее часть интегрировалась в геном клетки хозяина. Для получения интегрантов были использованы два способа. В первом способе трансформанты направляются на избирательные (содержащие тиострептон) и неизбирательные чашки и инкубируются в течение примерно 16 ч при температуре 29оС для получения одиночных колоний. Эти одиночные колонии на неизбирательных чашках, которые стойки к тиострептону на избирательных чашках, осуществляют несколько повторных проходов таким же образом до тех пор, пока одиночная колония не будет относительно стойкой без отбора. Во втором способе получения интегрантов трансформанты неизбирательно проходят несколько раз путем переноса спор от поверхности чашки с помощью хлопковой щетки. После нескольких таких проходов колонии выращиваются в неизбирательной жидкой среде (TSB), гомогенизируются, фрагментируются путем пропускания звуковых волн, разбавляются и высевают в чашках в избирательной и неизбирательной среде с тем, чтобы идентифицировать относительно стойкие интегранты. Другие способы получения интегрантов уже известны для специалистов в данной области и настоящее изобретение не ограничивается лишь каким-то одним способом получения интегрантов.
Относительно стойкие интегранты были использованы для инокуляции растительной среды (комплексная растительная среда содержит на один литр 10 г кукурузного экстракта, 5 граммов дрожжевого экстракта, 5 г зерен соевых бобов, 3 грамма карбоната кальция и 4,5 г сырого соевого масла, и величина рН была доведена до 7,8 посредством NaOH. Вполне пригодной растительной средой была также TSB) без тиостриптона (отсутствие избирательного давления), и растительная культура использовалась для инокуляции (10% инокулум) среды ферментации, в которой также отсутствовал тиострептон. Ферментации осуществлялись при скорости вращения 260 об/мин при температуре 29оС в течение семи дней. Общее содержание макролида в бульоне ферментации определяли путем экстракции смесью метанол СНСl3, измерения спектральной поглощательной способности при 290 нм и сопоставления со стандартной кривой. Тилозиновые факторы идентифицировались по нанесению точек питательного бульона ферментации на чашки (тонкослойная хроматография на силикагеле) и по проявлению этих чашек системой растворителя этилацетат диэтиламин в соотношении 95:5. Концентрация отдельных макролидных компонентов составляет А290 кратный процент каждого компонента, как определено методом жидкостной хроматографии высокого разрешения (НРLC).
П р и м е р 11. Получение тилозина.
Спорулированная культура Streptomy-ces fradiae содержащая плазмиды, отвечающие данному изобретению, может быть продуцирована путем роста организма на питательном скошенном агаре, имеющем следующий состав: Дрожжевой экстракт 1,0 г Мясной экстракт 1,0 г
Гидролизованный
казеин ("N-Z-Амин-
Тип А", выпускается
фирмой "Шеффилд Кемикэл Ко") 2,0 г Декстрин 10,0 г
Шестигидрат хлорида
кобальта (двухвалент- ного) 20,0 мг Агар 20 г Вода 1 л
Величину рН среды доводили до 7,3 путем добавления гидрата окиси натрия.
Скошеный агар инокулировали в течение пяти дней при примерно 30оС. Спорулированную культуру, выращенную на скошенном агаре, покрывали водой, и этот скошенный агар осторожно соскабливали для удаления спор, получая водную споровую суспензию.
1 мл этой споровой суспензии использовали для инокуляции в асептических условиях 100 мл порции стерильной растительной среды культивации, имеющей следующий состав, г: Глюкоза 15 Соевая мука 15 Вымоченная кукуруза 15 Хлористый натрий 5 Карбонат кальция 2
Водопроводная вода,
вводимая до общего
объема 1 л.
Инокулированную растительную среду инкубировали примерно при 30оС в течение 48 ч и при этом инкубат встряхивался со скоростью 114 циклов в минуту с помощью встряхивателя обратно-поступательного действия, с длиной хода 2 дюйма (50,8 мм).
Пять миллилитров этого растительного инокулума использовали для инокуляции асептически 100 мл порций указанной ниже стерилизованной продуцирующей среды, помещенных в 500-миллилитровые колбы Эрленмейера, г: Мука сои 15 Казеин 1 Сироп сырой глюкозы 20 мл Карбонат кальция 2,5 Нитрат натрия 3
Водопроводная вода,
вводимая до общего объема 1 л
Эту инокулированную культуру затем инкубировали в течение 100 ч при 26-28оС. В ходу инкубирования инкубат встряхивали со скоростью 114 циклов в минуту с помощью встряхивателя возвратно-поступательного действия с длиной хода 2 дюйма (50,8 мм). Величина рН исходной среды составляла примерно 6,5 и в конце периода инкубации величина рН среды обычно увеличивалась примерно до 7,5.
Подвергнутый ферментации питательный бульон фильтровали с целью удаления мицелия и других нерастворимых твердых веществ. Отфильтрованный питательный бульон содержал тилозин.
П р и м е р 12. Альтернативный способ получения тилозина.
Спорулированную культуру желаемого трансформированного микроорганизма получали путем выращивания организма на питательном скошенном агаре, имеющем следующий состав,
Агар Томатная паста овсяная мука г Томатная паста 20
Предварительно
термообработанная овсяная мука 20 Агар 15
Вода вводимая до
общего объема 1 литр
Этот скошенный агар инокулировали спорами данного организма и инокулированный скошенный агар инкубировали в течение 9 дней при температуре примерно 30оС. После инкубации спорулированную культуру скошенного агара покрывали водой и поверхность скошенного агара соскребали очень осторожно для удаления спор, получая при этом водную споровую суспензию.
Используя асептические способы, половину инокулума, полученного от одного скошенного агара, использовали для инокулирования 500 миллилитровой порции стерилизованной растительной среды культивации, имеющей указанный ниже состав, помещенной в 2 литровую колбу Эрленмейера: Кукурузные дрожжи 1 Глюкоза 15 Вымоченная кукуруза 5 Дрожжи 5 Карбонат кальция 5
Вода, вводимая до
общего объема 1 л
Инкубирование осуществлялось при 28оС в течение 48 ч со встряхиванием 110 циклов в минуту с использованием встряхивателя возвратно-поступательного действия с длиной хода 2 дюйма (50,8 мм).
0,25 галлона (0,96 л) растительного инокулума из данной колбы вводили асептически как инокулум в 250 галлонов (937,5 л.) стериальной питательной среды кукурузных дрожжей, описанной выше, содержащейся в стальной емкости ферментации на 350 галлонов (1312,5 л). В питательную среду вводили 0,025 галлона (0,096 л. ) Антипены А (противопенный продукт, выпускаемый фирмой "Доу Конинг Кампани") для предотвращения чрезмерного пенообразования, и по мере необходимости вводили дополнительные количества в ходе ферментации. Осуществлялась ферментация инокулированной среды в течение 24 ч при 24оС. В ходе ферментации среда подвергалась аэрации стерильным воздухом со скоростью 27 фут3/м в минуту (0,765 м3/мин) и перемешивалась двумя 16-дюймовыми мешалками (406 мм) со скоростью 160 об/мин.
В стальную емкость ферментации объемом 1700 галлонов (6375 л) вводили 1200 галлонов (4500 л) питательной среды культивации имеющей следующий состав:
Соя из вымоченной кукурузы XII кг Глюкоза 30 Соевая мука 15
Вымоченная кукуруза (мука) 5
Сырое соевое масло (соевых бобов) 10 Карбонат кальция 2 Хлористый натрий 5
Вода, вводимая до общего объема 1000 л
Данная среда инокулировалась 96 галлонами (360 л) инокулума, выращенного в емкости ферментации. Ферментация осуществлялась при 28оС в течение четырех дней и любое пенообразование предотвращалось путем ввода по мере необходимости "Larex" N 1 (противопенный продукт, выпускаемый фирмой "Свифт энд Кампани"). Эта среда ферментации подвергалась аэрации путем ввода стерильного воздуха со скоростью 128 фут3/мин (3,56 м3) мин) и перемешивалась двумя 24-дюймовыми мешалками (600 мм) работающими со скоростью 130 оборотов в 1 мин. В питательный бульон вводили 600 фунтов "Silfo" (272 кг) (диатомовая земля, выпускаемая фирмой "Сильфо Кампани"), и смесь фильтровалась. Величину рН фильтрата доводили до 8,5 путем добавления 20% гидрата окиси натрия, вводили 500 галлонов (1875 л) хлороформа и смесь перемешивалась в течение 30 мин, и хлороформный слой, который имел форму эмульсии, декантировался. Экстракцию хлороформом повторяли дважды с использованием 500-галлоновые порции хлороформа (1875 г.) Хлороформные эмульсии, содержащие тилозин, соединялись и пропускались через сепаратор Дэ Лаваля для разбивки эмульсии, и хлороформный раствор затем концентрировался в вакууме до объема 25 л. Примеси большей частью были удалены из раствора путем пропускания его колонку диаметром 6 дюймов (152,4 мм), содержащую 10 кг активированного угля, выпускаемого фирмой "Питсбург Коук энд Кемикэл Ко". Колонку с активированным углем промывали 16 л хлороформа и соединенные хлороформные потоки, содержащие тилозин, выпаривали в вакууме до объема примерно 2 л. Хлороформный концентрат медленно вводили с одновременным перемешиванием в 20 л петролейного эфира, и смесь перемешивали в течение 15 мин, затем ее фильтровали, удаляя белый аморфный осадок тилозина.
Этот аморфный тилозин кристаллизовался путем растворения его в 355 мл ацетона, фильтрации ацетоновой смеси с целью удаления небольшой мути, и медленного ввода отфильтрованной ацетоновой смеси со слабым перемешиванием в 20 мл воды при температуре 5оС. Водно-ацетоновый раствор тилозина выстаивался при комнатной температуре при слабом перемешивании с тем, чтобы ацетон медленно испарялся, после чего выкристаллизовывался тилозин. Кристаллы тилозина удаляли путем фильтрации и высушивали в вакууме при комнатной температуре. Тилозин имел температуру плавления примерно 127-132оС.
П р и м е р 13. Получение тартрата тилозина.
Пять граммов кристаллического тилозина растворяли в 100 мл ацетона и 1,5 г D-винной кислоты, растворенной в 20 мл ацетона, вводили в этот раствор с одновременным перемешиванием. Раствор выстаивался при комнатной температуре, после чего виннокислая соль тилозина выкристаллизовывалась из раствора.
Кристаллы виннокислой соли тилозина удалялись путем фильтрации, промывались ацетоном и высушивались воздухом. Кристаллическая виннокислая соль тилозина имела температуру плавления примерно 140-146оС.
П р и м е р 14. Получение глюконата тилозина.
1,03 грамма глюконо-дельта лактона растворяли в 10 мл воды, и водный раствор нагревали до температуры 85оС в течение двух часов с тем, чтобы вызвать гидролиз лактона с образованием глюконовой кислоты. В этот водный раствор вводили 15 мл теплого метанола. Пять граммов тилозина, растворенного в 10 мл метанола, вводили в данную метаноловую смесь с одновременным перемешиванием. Тилозиновая смесь выстаивалась в течение ночи при комнатной температуре. Метанол удалялся из смеси путем выпаривания в вакууме при комнатной температуре. После удаления метанола в водно-тилозиновую смесь вводили 40 мл воды. Разбавленную смесь фильтровали, и фильтрат, содержащий тилозин, подвергали сушке с замораживанием, получая белый твердый продукт, состоящий из глюконатной соли тилозина. Эта глюконатная соль тилозина имела температуру плавления примерно 114-117оС.
П р и м е р 15. Получение хлоргидрата тилозина 890 мг тилозина растворяли в 200 мл простого эфира. Эфирную смесь подкисляли путем добавления 0,082 мл 12 нормальной соляной кислоты. Осадок хлоргидратной соли тилозина, который был образован, отфильтровывали, промывали простым эфиром, и высушивали в вакууме. Хлоргидратная соль тилозина выкристаллизовывалась из смеси этано-простой эфир. Эта хлоргидратная соль тилозина имела температуру плавления примерно 141-145оС.
Использование: генетическая инженерия, в частности, к способу получения тилозина в результате культивирования штаммов трансформированной рекомбинантной плазмидной ДНК. Сущность изобретения: способ получения тилозина предусматривает культивирование штамма Streptomyces fradiae, трансформированного рекомбинантной плазмидной ДНК pHJ 2280 или pH I2 311 или pH I2 315 содержащей гены биосинтеза тилозина tyl C, tyl D, tyl E, tyl F, или tyl H, или tyl K, tyl L или tyl M. 7 ил. 5 табл.
СПОСОБ ПОЛУЧЕНИЯ ТИЛОЗИНА, предусматривающий культивирование штамма Streptomyces fradiae в питательной среде, выделение и очистку целевого продукта, отличающийся тем, что культивируют штамм Str. fradiae, предварительно трансформированный рекомбинатной плазмидной ДНК pHJ 2280, или pH J 284, или pH J2 311, или pH J2 315, содержащей гены биосинтеза тилозина tylC, tylD, или tylE, или tylF, или tulH, или tylK, или tylL, или tylM.
| Авторское свидетельство СССР N 755837, C 12N 15/00, 1980. |
