Изобретение относится к микробиологической и медицинской промышленности.
В аэробных организмах целый ряд нормальных или ненормальных событий приводит к образованию перекисного радикала и образующихся из него более токсичных радикалов. Перекисный радикал является важным медиатором как при воспалительной реакции фагоцитов, так и при реакции отторжения трансплантированных органов. В эукариотических клетках синтезируется специализированный цитоплазматический фермент, иногда обнаруживаемый также в сыворотке, супероксиддисмутаза, который нейтрализует перекисный радикал. Показано, что супероксиддисмутаза обладает противовоспалительным действием, а также существенно препятствует отторжению пересаженных органов сердца, почек и др. Для терапевтических целей, когда есть показания к регулярному применению препаратов супероксиддисмутазы, необходимо использовать белок, идентичный естественному человеческому, так как чужеродная иммунологически отличная супероксиддис- мутаза (например, бычья) будет провоцировать нежелательную иммунную реакцию. Показано, что дрожжи являются подходящим, если не наилучшим, реципиентом для экспрессии гена человеческой супероксиддисмутазы, так как дрожжевая клетка обеспечивает аминоконцевое ацетилирование целевого белкового продукта, что делает последний неотличным от естественной человеческой эритроцитарной супероксид- дисмутазы.
Описанный в настоящее время дрожжевой штамм продуцент супероксиддисмутазы человека, обладает достаточно высокой продуктивностью.
Недостатком этого штамма является невозможность его селективного культивирования на средах сложного неопределенного состава, например на мелассных средах, что существенно удорожает и усложняет процесс наработки биомассы дрожжей, содержащей супероксиддисмутазу человека.
Целью изобретения является создание штамма дрожжей продуцента супероксиддисмутазы человека, способного к поддержанию и продукции целевого белка при росте на среде с мелассой.
Штамм Saccharomyces cerevisiae YBS13Cpgk/pBr-SOD-PGKdel получен следующим образом.
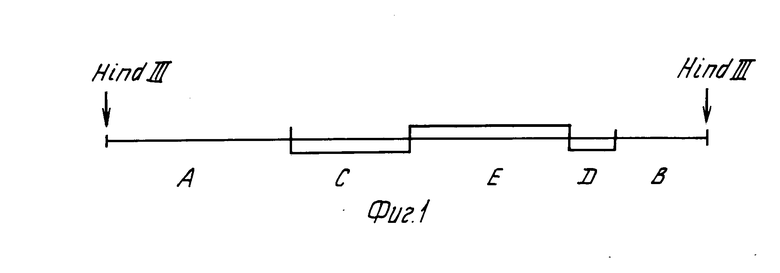
Штамм дрожжей YBS13C генотипа ura3 трансформировали фрагментом ДНК PGK1: URA3. Схема этого фрагмента представлена на фиг.1 где А промоторная область гена PGK1, 0,7 т.п.н. соответствует HindIII/ClaI фрагменту;
В область терминации транскрипции гена PGK1, 0,4 т.п.н. соответствует BglII/HindIII фрагменту;
С фрагмент 0,15, т.п.н. соответствующий BglII/HpaI фрагменту гена LEU2;
D фрагмент 0,52 т.п.н. соответствующий BglII/BglII фрагменту гена LEU2;
Е фрагмент 1,17 т.п.н. соответствующий HindIII/HindIII фрагменту гена UPA3.
Трансформанты отбирали по способности расти на минимальной дрожжевой среде, содержавшей 2% этанола, 3% глицерина и 1% казаминовых кислот. Среди трансформантов идентифицировали клоны с разрушенным хромосомным локусом PGK1 по неспособности к росту на среде с 2%-ной глюкозой. Один из таких клонов обозначен YBS13Cpgk. Культура YBS13Cpgk была использована в дальнейшем в качестве реципиента.
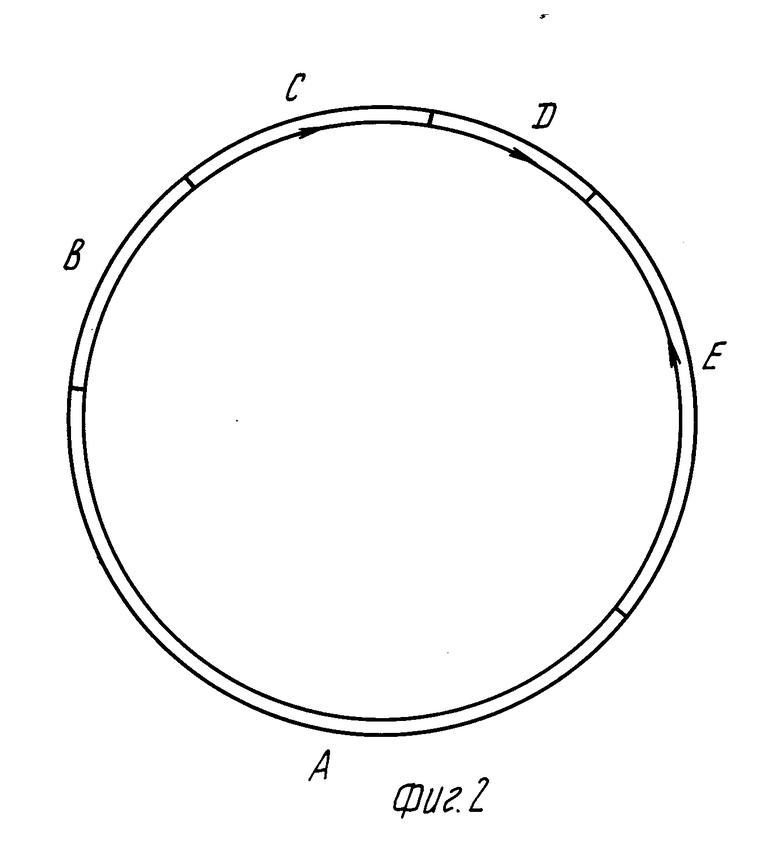
Сконструирована экспрессионная плазмида, содержащая ген супероксиддисмутазы (SOD) человека. Плазмида обозначена pBR-SOD-PGKdel. Схема этой плазмиды представлена на фиг. 2, где А соответствует EcoRI/BamHI фрагменту плазмиды pBR322;
В соответствует EcoRI/HindIII фрагменту дрожжевой 2 мкм плазмиды В-типа;
С соответствует HindIII/HindIII фрагменту дрожжевого гена URA3;
D соответствует фрагменту ДНК, кодирующему PGK1: начало фрагмента -33 н. п. относительно стартового кодона, конец фрагмента 2355 н.п. фрагмента ДНК, кодирующего ген PGK1;
Е экспрессионная кассета с геном SOD, которая имеет следующий вид
1 11 21 31 41 51 61
aagcttggcgtccattctattagcgcaactacagagaacagggcacaaacaggcaaaaaa
ttcgaaccgcaggtaagataatcgcgttgatgtctcttgtcccgtgtttgtccgtttttt
61 71 81 91 101 111 121
cgggcacacctcaatggagtgatgcaacctgcctggagtaaatgatgacacaaggcaatt
gcccgtgtggagttacctcactacgttggacggacctcatttactactgtgttccgttaa
121 131 141 151 161 171 181
gacccacgcatgtatctatctcattttcttacaccttctattaccttctgctctctctga
ctgggtgcgtacatagatagagtaaaagaatgtggaagataatggaagacgagagagact
181 191 201 211 221 231 241
tttggaaaaagctgaaaaaaaaggtttaaaccagttccctgaaattattcccctacttga
aaacctttttcgacttttttttccaaatttggtcaagggactttaataaggggatgaact
241 251 261 271 281 291 301
ctaataagtatataaagacggtaggtattgattgtaattctgtaaatctatttcttaaac
gattattcatatatttctgccatccataactaacattaagacatttagataaagaatttg
301 311 321 331 341 351 361
ttcttaaattctacttttatagttagtcttttttttagttttaaaacaccaagaacttag
aagaatttaagatgaaaatatcaatcagaaaaaaaatcaaaattttgtggttcttgaatc
361 371 381 391 401 411 421
MetAlaThrLysAlaValCysValLeuLysGly 11
tttcgactaaacacacataaataaacaccatggctacgaaggccgtgtgcgtgctgaagg aaagctgatttgtgtgtatttatttgtggtaccgatgcttccggcacacgcacgacttcc
421 431 441 451 461 471 481
AspGlyProValGlnGlyIleIleAsnPheGluGlnLysGluSerAsnGlyProValLys 31 gcgacggcccagtgcagggcatcatcaatttcgagcagaaggaaagtaatggaccagtga cgctgccgggtcacgtcccgtagtagttaaagctcgtcttcctttcattacctggtcact
481 491 501 511 521 531 541
ValTrpGlySerIleLysGlyLeuThrGluGlyLeuHisGlyPheHisValHisGluPhe 51 aggtgtggggaagcattaaaggactgactgaaggcctgcatggattccatgttcatgagt tccacaccccttcgtaatttcctgactgacttccggacgtacctaaggtacaagtactca
541 551 561 571 581 591 601
GlyAspAsnThrAlaGlyCysThrSerAlaGlyProHisPheAsnProLeuSerArgLys 71 ttggagataatacagcaggctgtaccagtgcaggtcctcactttaatcctctatccagaa aacctctattatgtcgtccgacatggtcacgtccaggagtgaaattaggagataggtctt
601 611 621 631 641 651 661
HisGlyGlyProLysAspGluGluArgHisValGlyAspLeuGlyAsnValThrAlaAsp 91 aacacggtgggccaaaggatgaagagaggcatgttggagacttgggcaatgtgactgctg ttgtgccacccggtttcctacttctctccgtacaacctctgaacccgttacactgacgac
661 671 681 691 701 711 721
LysAspGlyValAlaAspValSerIleGluAspSerValIleSerLeuSerGlyAspHis 111 acaaagatggtgtggccgatgtgtctattgaagattctgtgatctcactctcaggagacc tgtttctaccacaccggctacacagataacttctaagacactagagtgagagtcctctgg
721 731 741 751 761 771 781
CysIleIleGlyArgThrLeuValValHisGluLysAlaAspAspLeuGlyLysGlyGly 131 attgcatcattggccgcacactggtcgtccatgaaaaagcagatgacttgggcaaaggtg taacgtagtaaccggcgtgtgaccagcaggtactttttcgtctactgaacccgtttccac
781 791 801 811 821 831 841
AsnGluGluSerThrLysThrGlyAsnAlaGlySerArgLeuAlaCysGlyValIleGly 151 gaaatgaagaaagtacaaagacaggaaacgctggaagtcgtttggcttgtggtgtaattg ctttacttctttcatgtttctgtcctttgcgaccttcagcaaaccgaacaccacattaac
841 851 861 871 881 891 901
IleAlaGln
ggatcgcccaataaacattcccttggatgtagtctgaggccccttaactcatctgttatc 154 cctagcgggttatttgtaagggaacctacatcagactccggggaattgagtagacaatag
901 911 921 931 941 951 961
ctggtagctgtagaaatgtatcctgataaacattaaacactgtaatcttaaaaaaaaaaa
gaccatcgacatctttacataggactatttgtaatttgtgacattagaattttttttttt
961 971 981 991 1001 1011 1021
aggaattccggatccaagatggcctttggtgggttgaagaaggaaaaagacagaaacgac
tccttaaggcctaggttctaccggaaaccacccaacttcttcctttttctgtctttgctg
1021 1031 1041 1051 1061 1071 1081
ttaattacctacttgaaaaaagcctgtgagtaaacaggccccttttcctttgtcgatatc aattaatggatgaacttttttcggacactcatttgtccggggaaaaggaaacagctatag
1081 1091 1101 1111 1121 1131 1141
atgtaattagttatgtcacgcttacattcacgccctccccccacatccgctctaaccgaa tacattaatcaatacagtgcgaatgtaagtgcgggaggggggtgtaggcgagattggctt
1141 1151 1161 1171 1181 1191 1201
aaggaaggagttagacaacctgaagtctaggtccctatttatttttttatagttatgtta ttccttcctcaatctgttggacttcagatccagggataaataaaaaaatatcaatacaat
1201 1211 1221 1231 1241 1251 1261
gtattaagaacgttatttatatttcaaatttttcttttttttctgtacagacgcgtgtac cataattcttgcaataaatataaagtttaaaaagaaaaaaaagacatgtctgcgcacatg
1261 1271 1281 1291 1301 1311 1321
gcatgtaacattatactgaaaaccttgcttgagaaggttttgggacgctcgaaggcttta cgtacattgtaatatgacttttggaacgaactcttccaaaaccctgcgagcttccgaaat
1321 1331
atttgcaagctgagatct
taaacgttcgactctaga
При трансформации реципиента YBS13Cpgk плазмидой pBR-SOD-PGKdel отобраны искомые трансформанты, способные к росту на среде с глюкозой.
Путем инкубации этих культур в присутствии этидийбромида получены клоны дрожжей, являющиеся дыхательными мутантами rho-, т.е. не способными к аэробной ассимиляции всех глюконеогенетических субстратов. Один из таких клонов отобран в качестве продуцента и обозначен YBS13Cpgk/pBR-SOD-PGKdel. Исходя из метода конструирования, данный продуцент способен расти на среде со сбраживаемыми сахарами анаэробно лишь при наличии экспрессионной плазмиды. В случае утери плазмиды образуется реципиентная клетка генотипа rho-pgk-. Такие клетки не способны усваивать ни один из субстратов и являются нежизнеспособными. Таким образом, созданный штамм продуцент обладает способностью к селективному поддержанию экспрессионной плазмиды на средах любого состава, например на мелассных средах.
Штамм Saccharomyces cerevisiae YBS13Cpgk/pBR-SOD-PGKdel депонирован во Всесоюзной коллекции промышленных микроорганизмов (ВКПМ) под номером Y2134.
Штамм характеризуется следующими культурально-морфологическими и физиолого-биохимическими признаками.
Вегетативные клетки двухсуточной культуры, выращенной на твердой питательной среде с 2% глюкозы в качестве единственного источника углерода, имеют овальную форму, размер клеток 3,5 х 7,0 мкм, протоплазма гомогенная, размножение почкованием.
При выращивании на твердой среде, содержащей дрожжевой экстракт и пептон (YEP) при 30оС через 72 ч роста колонии имеют следующий вид:
1) на среде YEP с глюкозой колонии белого цвета с ровным краем, блестящей поверхностью, конусообразным профилем, сметанообразной консистенцией;
2) на среде YEP с крахмалом колонии белого цвета с узорчатым краем, матовой поверхностью, линзовидным профилем и крупинчатой консистенцией;
3) на среде YEP с мелассой колонии белого цвета с матовой морщинистой поверхностью, узорчатым краем, выпуклым профилем и сметанообразной консистенцией.
Рост в жидкой среде на среде YEP с крахмалом при 32оС в течение первых 24 ч культивирования жидкость мутная, осадок белый, не комкуется, пристеночных пленок не образует.
Факультативный анаэроб.
Температура роста 23-33оС (оптимум 31оC, рН культивирования 3,8-6,7 (оптимум 5,0).
Ассимиляция источников углерода: сбраживает глюкозу, галактозу, фруктозу, мальтозу, сахарозу, декстрины, крахмал.
Ассимиляция источников азота: усваивает аминокислоты, мочевину, сернокислый аммоний, азотнокислый аммоний.
Отличительные особенности: при культивировании на богатой среде с крахмалом (2% ) вокруг колоний образуются зоны просветления крахмала, окруженные темным ободком после инкубации чашек при +4оС в течение 24 ч.
Штамм Saccharomyces cerevisiae Y2134 непатогенен.
Штамм хранится на агаризованной богатой среде с глюкозой в течение 3 мес при +4оС.
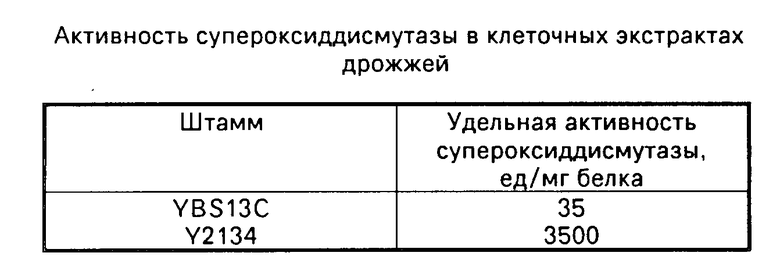
П р и м е р использования по назначению: клетки штамма-продуцента Y2134 и исходного штамма YBS13C выращивали в колбах при 30оС и аэрации на среде следующего состава: глюкоза 5% дрожжевой экстракт 1% пептон 2% вода водопроводная. Исходный титр составлял 5х10о кл./мл. Через 72 ч выращивания культуры переходили в стационарную фазу роста при титре 1х108. Выход биомассы дрожжей 70%-ной влажности составлял 12 г/л. Биомассу, собранную в этот период культивирования, использовали для анализа супероксиддисмутазы.
Клетки из 3 мл культуры собирали центрифугированием, промывали водой и разрушали после добавления равного объема стеклянных шариков на встряхивателе 2 раза по 40 с. Для получения фракции растворимых белков дебрис отделяли центрифугированием при 15000 об/мин в течение 20 мин. Осветленный таким образом клеточный экстракт анализировали электрофоретически и определяли в нем активность супероксиддисмутазы. По результатам электрофореза (фиг.3, где дорожка 1 чистый препарат СОД человека; дорожки 2-8 экстракт клеток штамма Y2134; дорожка 9 экстракт клеток штамма YBS13C) видно, что штамм продуцент содержит существенное количество супероксиддисмутазы человека, отсутствующей в экстракте исходного штамма. Из данных денситометрии полос 2-8 этой электрофореграммы следует, что супероксиддисмутаза человека составляет 20% растворимого клеточного белка.
Активность супероксиддисмутазы в клеточных экстрактах измеряли по ингибированию реакции автоокисления кверцетина, которую контролировали спектрофотометрически. За единицу активности супероксиддисмутазы принимали такое количество белка, которое требовалось для ингибирования реакции на 50% Данные по анализу активности приведены в таблице.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pPHO-SOD23, КОДИРУЮЩАЯ СИНТЕЗ СУПЕРОКСИДДИСМУТАЗЫ ЧЕЛОВЕКА, И ШТАММ Saccharomyces cerevisiae № Д-11 ВНИИСХМ-ПРОДУЦЕНТ СУПЕРОКСИДДИСМУТАЗЫ ЧЕЛОВЕКА | 2000 |
|
RU2200196C2 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE - ПРОДУЦЕНТ СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА | 2011 |
|
RU2460795C1 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКА E7-HSP70 И ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2489481C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE И ШТАММ-ПРОДУЦЕНТ ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2011 |
|
RU2446172C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ГИБРИДНОГО БЕЛКА Е7-HSP70 (ВАРИАНТЫ) | 2013 |
|
RU2546917C1 |
| СПОСОБ КОНСТРУИРОВАНИЯ ШТАММОВ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫХ ПРОДУЦЕНТОВ СОМАТОТРОПИНА ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫЙ ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА (ВАРИАНТЫ) | 2009 |
|
RU2420567C1 |
| ГИБРИДНЫЙ БЕЛОК, ОБЛАДАЮЩИЙ ПРОЛОНГИРОВАННЫМ ДЕЙСТВИЕМ, НА ОСНОВЕ РЕКОМБИНАНТНОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ), СПОСОБ ЕГО ПОЛУЧЕНИЯ И ШТАММ Saccharomyces cerevisiae ДЛЯ ОСУЩЕСТВЛЕНИЯ ЭТОГО СПОСОБА (ВАРИАНТЫ) | 2013 |
|
RU2515913C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА, ШТАММ Saccharomyces cerevisiae - ПРОДУЦЕНТ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2010 |
|
RU2427645C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА КЛЕТКАМИ ДРОЖЖЕЙ, СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE-ПРОДУЦЕНТ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА | 1998 |
|
RU2180003C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА ПАУТИНЫ, СЛИТЫЙ БЕЛОК, РЕКОМБИНАНТНАЯ ДНК, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН И ШТАММЫ-ПРОДУЦЕНТЫ | 2010 |
|
RU2451023C1 |

Использование: микробиологическая промышленность, в частности производство ферментных препаратов супероксиддисмутазы. Сущность изобретения: штамм дрожжей Saccharomyces cerevisiae продуцент супероксиддисмутазы человека, получен с применением методов классической генетики и генетической инженерии и депонирован в ВКПМ под номером Y2134. Для получения супероксиддисмутазы штамм культивируется на среде, содержащей глюкозу (5%), дрожжевой экстракт (1% ) и пептон (2%), на качалке при 30°С в течение 72 ч. Активность фермента в клеточном экстракте дрожжей составляет 3500 ед./мг белка. 3 ил. 1 табл.
Штамм дрожжей Saccharomyces cerevisiae ВКПМ У-2134 продуцент супероксиддисмутазы человека.
| "Biotechnology", 1987, V5, 363-366. |

