Изобретение относится к микробиологической промышленности, в частности касается выделения нового штамма Pseudomonas species Ш-22. Данные клетки после культивирования на среде, содержащей в качестве единственного источника углерода и азота L-пролин, специфически окисляют L-пролин и практически не окисляют другие аминокислоты и некоторые распространенные сахара.
Штамм Pseudomonas species Ш-22 представляет интерес для разработки метода количественного определения L-пролина в различных средах и может найти применение в микробиологической промышленности, клинических и лабораторных исследованиях.
Известно, что способностью окислять L-пролин обладают представители таких родов микроорганизмов, как, например, Escherichia, Salmonella и Pseudomonas. Причем показано, что это свойство наиболее ярко выражено у представителей родов Escherichia и Pseudomonas [1, 2]
Описаны дикий и мутантный штаммы Salmonella typhimurium 15 59, обладающие пролинокисляющий активностью [3] Глубинное культивирование клеток обоих штаммов в среде, содержащей L-пролин, приводит к активации их пролинокисляющей активности в 25 и 10 раз соответственно. Однако активированные клетки и дикого, и мутантных штаммов сохраняют способность окислять некоторые сахара (например, глюкозу и мальтозу) и аминокислоты (например, гистидин) с практически той же скоростью, что и L-пролин. Таким образом, основным недостатком данных штаммов является низкая субстратная специфичность.
Наиболее близкими к предлагаемому штамму являются дикий штамм E. coli-B и ауксотрофный E. coli-B4, описанные в работе [4] Глубинное культивирование клеток обоих штаммов в среде, содержащей L-пролин, приводит к активации их пролинокисляющей активности в 14 и 10 раз соответственно. Однако активированные клетки и дикого, и ауксотрофного штаммов сохраняли способность окислять многие аминокислоты со скоростью, составляющей от 20 до 50% от скорости окисления L-пролина. Недостатком данного штамма является низкая субстратная специфичность.
Настоящее изобретение относится к выделению нового дикого штамма Pseudomonas species Ш-22, обладающего специфичной пролинокисляющей активностью.
Предлагаемый штамм был получен в результате селекционного отбора колоний бактерий, выделенных из почвы, и их последующего культивирования на средах, содержащих 0,05 0,45% L-пролина.
Штамм Pseudomonas species Ш-22 хранится во Всесоюзной коллекции промышленных микроорганизмов Института "ВНИИГенетика" (Москва), N B-4877.
Штамм имеет следующие характеристики.
Морфологические особенности штамма. Клетки прямые, палочковидные, 0,1 - 0,2 х 0,9 1,0 мкм, встречаются в длинных и коротких цепочках, подвижные, грамотрицательные, спор не образуют.
Культуральные признаки. При росте на агаризованных питательных средах (мясопептонный агар, LB-среда) через 24 ч при 25oC образуются колонии, светло-желтые, выпуклые, блестящие, гладкие, край слегка волнистый, достигающие в диаметре 1,5 мм. Рост по уколу в МПА обильный, в основном на поверхности среды. В жидких питательных средах также наблюдается обильный рост.
Хорошо растет на минеральных средах простого состава, дополненных L-пролином. В добавлении факторов роста и других аминокислот не нуждается. При культивировании на пролинсодержащей среде наблюдается синтез внеклеточного желто-зеленого пигмента.
Физиолого-биохимические признаки. По отношению к кислороду облигатный аэроб. Максимальная температура роста 37oC, минимальная 15oC, оптимальная 25 30oC, рН среды в пределах 6,8 7,5.
Азот утилизирует в виде солей аммония, мочевины, нитратов.
Пигменты. Образует желто-зеленый водорастворимый флюоресцирующий пигмент на пролинсодержащей синтетической среде. Внутриклеточных пигментов не образует. Реакция на оксидазу и каталазу положительная. Реакция Фогес-Проскауэра отрицательная. Индол не образует. Тест на анаэробную аргинин-дегидрогеназу положительный. Реакция на анаэробную денитрификацию отрицательная. Лецитиназная активность отсутствует.
Не гидролизует крахмал, желатин, твин-80.
В качестве источников углерода использует многие сахара, аминокислоты и органические кислоты.
Штамм Pseudomonas species Ш-22 непатогенен для человека и животных.
В соответствии с отмеченными морфологическими, физиологическими и культурально-биохимическими признаками штамм был идентифицирован как Pseudomonas species.
При определении систематического положения использовали определитель Bergey:
Bergey's Manual of Determination Bacteriology. 1974. 8-th ed. Baltimore. The William Wilkins Co.
Пример 1.
Клетки Pseudomonas species Ш-22 выращивают на синтетической среде, содержащей 0,05% L-пролина, 0,2% K2HPO4 • 3H2O, 0,05% KH2PO4, 0,02% MgSO4 • 7H2O, pH 7,2 7,5, при 30oC в 750 мл конических колбах (объем ферментационной среды 200 мл) на круговой качалке (180 об/мин) в течение 20 ч. Биомассу отделяют центрифугированием (10000 об/мин, 15 мин), суспендируют в 50 мМ калий-фосфатном буфере рН 7,2. Пролинокисляющую активность определяют амперометрически. Для этого в измерительную ячейку (объем 5 мл), снабженную перемешивающим устройством, помещают 50 мМ калий-фосфатный буфер рН 7,2. В ячейку вводят 0,05 мл клеточной суспензии, исходная концентрация которой составляет 0,5 0,1 г/мл и 0,05 мл 5 мМ раствора L-пролина. Изменение содержания кислорода в измерительной ячейке регистрируют с помощью мембранного кислородного электрода Кларка. Пролинокисляющую активность клеток вычисляют по их скорости потребления кислорода в присутствии L-пролина и выражают как количество кислорода (мкмоль), потребленное 1 мг клеток (влажный вес) в 1 мин [5]
Для данной концентрации L-пролина скорость потребления кислорода составляла 3,00 мкмоль/мин•мг клеток.
Пример 2.
Культивирование клеток и измерения проводили также, как в примере 1, только в среду роста вместо 0,05% добавляли 0,45% L-пролина. Пролинокисляющая активность клеток составляла 4,50 мкмоль O2/мин•мг клеток.
Пример 3.
Культивирование клеток и измерения проводили также, как описано в примере 2, только в ячейку последовательно вводили 0,05 мл 5 мМ растворов различных аминокислот (табл. 1). Клетки Pseudomonas species Ш-22, выращенные на среде, содержащей L-пролин, практически не окисляли другие аминокислоты.
Пример 4.
Выращивание клеток и измерения проводили также, как описано в примере 2, только вместо L-пролина в измерительную ячейку последовательно вводили 0,05 мл 5 мМ растворов глюкозы, фруктозы, галактозы, лактозы и сахарозы. Клетки не окисляют данные сахара.
Пример 5.
Клетки Pseudomonas species Ш-22 выращивали на среде, содержащей 1% пептона, 0,5% дрожжевого экстракта, 1% хлорида натрия (рН 7,2). Условия культивирования и определения активности такие же, как в примере 2. Пролинокисляющая активность клеток составляла 0,5% от активности клеток, выращенных на среде с пролином. Кроме того, клетки проявляли высокое сродство к другим аминокислотам и сахарам (табл. 2).
Пример 6.
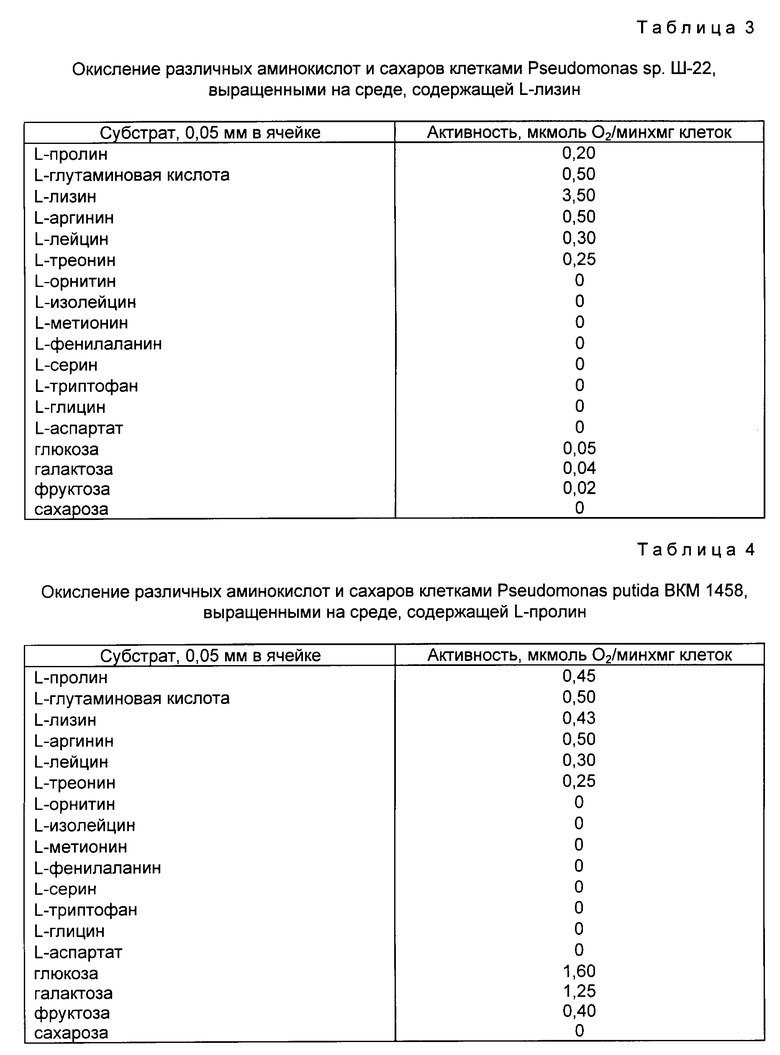
Культивирования клеток и измерения проводили также, как описано в примере 2, только в качестве единственного источника углерода и азота в среду культивирования добавляли вместо L-пролина L-лизин. Пролинокисляющая активность клеток, выращенных на L-лизине, составляла 0,5% от активности клеток, выращенных на среде с L-пролином. Кроме того, клетки проявляли высокое сродство к другим аминокислотам и сахарам (табл. 3).
Пример 7.
Среда культивирования и условия определения пролинокисляющей активности, как в примере 2, только вместо клеток Pseudomonas species Ш-22 использовали штамм Pseudomonas putida ВКМ В-1458. Пролинокисляющая активность данного штамма составляла 10% от активности Pseudomonas species Ш-22. Клетки Pseudomonas putida ВКМ 1458 кроме L-пролина окисляли ряд аминокислот и сахаров (табл. 4).
Предлагаемый штамм Pseudomonas species Ш-22 отличается от Е. coli B и E. coli B4 тем, что после культивирования на среде, содержащей в качестве единственного источника углерода и азота L-пролин, специфически окисляет L-пролин и практически не окисляет другие аминокислоты и некоторые распространенные сахара.
На основе данного штамма может быть разработан метод количественного определения пролина, что представляет интерес для микробиологической промышленности, клинических и лабораторных исследований.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ L-ПРОЛИНА | 1993 |
|
RU2086652C1 |
| Штамм @ @ ВКМ В-1458-продуцент лизин-2-монооксигеназы | 1984 |
|
SU1310427A1 |
| ШТАММ BACILLUS SUBTILIS ИБ-22-ПРОДУЦЕНТ ЦИТОКИНИНОВ | 2000 |
|
RU2178970C2 |
| Бактерия вида Escherichia coli - продуцент L-треонина, способ микробиологического синтеза L-треонина с ее использованием. | 2018 |
|
RU2697499C1 |
| ШТАММ БАКТЕРИЙ PSEUDOMONAS SPECIES ДЛЯ ПОЛУЧЕНИЯ ПРЕПАРАТА ПРОТИВ ВОЗБУДИТЕЛЕЙ ЗАБОЛЕВАНИЙ РАСТЕНИЙ | 1997 |
|
RU2130262C1 |
| СПОСОБ ПРОМЫШЛЕННОГО ПОЛУЧЕНИЯ ЧЕЛОВЕЧЕСКОГО ЛЕЙКОЦИТАРНОГО ИНТЕРФЕРОНА-АЛЬФА-2 | 1996 |
|
RU2118366C1 |
| ФРАГМЕНТ ДНК ИЗ ESCHERICHIA COLI, ОПРЕДЕЛЯЮЩИЙ ПОВЫШЕННУЮ ПРОДУКЦИЮ L-АМИНОКИСЛОТ (ВАРИАНТЫ), И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 1999 |
|
RU2175351C2 |
| ШТАММ KLUYVEROMYCES MARXIANUS VAR. BULGARICUS T - ПРОДУЦЕНТ СУПЕРОКСИД ДИСМУТАЗЫ И СОПУТСТВУЮЩИХ ФЕРМЕНТОВ | 1992 |
|
RU2113477C1 |
| ШТАММ БАКТЕРИЙ Pseudomonas aureofaciens ВКМ В-2501 Д ДЛЯ БИОДЕГРАДАЦИИ ПОЛИЦИКЛИЧЕСКИХ АРОМАТИЧЕСКИХ УГЛЕВОДОРОДОВ В УСЛОВИЯХ ЗАГРЯЗНЕНИЯ ПОЧВ СОЛЯМИ НИКЕЛЯ | 2008 |
|
RU2396339C2 |
| ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ L-ГИСТИДИНА | 1997 |
|
RU2119536C1 |

Использование: биотехнология, микробиологическая промышленность. Сущность изобретения: новый штамм, Pseudomonas species ВКПМ В-4787. Данные клетки после культивирования на среде, содержащей в качестве единственного источника углерода и азота L-пролин, специфически окисляют L-пролин и практически не окисляют другие аминокислоты и некоторые распространенные сахара. Предлагаемый штамм был получен в результате селекционного отбора колоний бактерий, выделенных из почвы, и культивирования на средах, содержащих 0,05 - 0,45% L-пролина. Штамм Pseudomonas species представляет интерес для разработки метода количественного определения L-пролина в различных средах и может найти применение в микробиологической промышленности, клинических и лабораторных исследованиях. 4 табл.
Штамм бактерий Pseudomonas species ВКПМ В-4877, способный специфически окислять L-пролин.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Kameda et al., Nature, 1963, p | |||
| Способ утилизации отработанного щелока из бучильных котлов отбельных фабрик | 1923 |
|
SU197A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Смирнов В., Киприянов Е | |||
| Бактерии рода Pseudomonas, 1991 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| S | |||
| Newell et al., 1972, Bacteriol., III, p | |||
| Ручной дровокольный станок | 1921 |
|
SU375A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| L | |||
| Frank et al., 1961, Arch | |||
| Biochem | |||
| and Biophys, 95, p | |||
| Кинематографический аппарат | 1918 |
|
SU441A1 |
