Изобретение относится к генно-инженерным лекарственным препаратам, получаемым биотехнологическим путем, а именно к способам промышленного получения рекомбинантного лейкоцитарного интерферона альфа-2b человека медицинского назначения (далее интерферон), а также к рекомбинантньм штаммам продуцентам Escherichia coli (E.coli) и плазмидным ДНК, кодирующим синтез интерферона.
Интерфероны представляют собой белковые молекулы с молекулярной массой от 15000 до 21000 дальтон, продуцируемые и секретируемые клетками в ответ на вирусную инфекцию или другие возбудители. Выделяют три основные группы интерферонов: альфа, бета и гамма. Сами по себе эти группы не являются однородными и могут содержать несколько различных молекулярных разновидностей интерферона. Так, выделено более 14 генетических разновидностей интерферона альфа, которые представляют интерес и находят широкое применение в медицине в качестве противовирусных, антипролиферативных и иммуномодулирующих средств.
Известны способы получения лейкоцитарного интерферона человека из лейкоцитов донорской крови человека, индуцированных вирусами и другими индукторами (SU1713591, RU 2066188, RU 2080873).
Основным недостатком этих способов получения интерферонов являются вероятность контаминации конечного продукта вирусами человека, такими как вирус гепатитов В и С, вируса иммунодефицита и др.
В настоящее время более перспективным признан способ получения интерферона микробиологическим синтезом, который обеспечивает возможность получения целевого продукта со значительно более высоким выходом из сравнительно недорогого исходного сырья. Используемые при этом подходы позволяют создать оптимальные для бактериальной экспрессии варианты структурного гена, а также регуляторных элементов, контролирующих его экспрессию.
В качестве исходных микроорганизмов используют различные конструкции штаммов Pichia pastoris, Pseudomonas putida и Escherichia coli.
Недостатком использования P. pastoris в качестве продуцента интерферона (J.N. Garcia, J.A. Aguiar et. al. //High level expression of human IFN-2b in Pichia pastoris.//Biotecnologia Aplicada, 12(3),152-155, 1995), является крайне сложные условия ферментации этого типа дрожжей, необходимость строго поддерживать концентрацию индуктора, в частности метанола, в процессе биосинтеза. Недостатком использования штаммов Ps. putida (SU1364343, SU1640996, SU1591484, RU1616143, RU2142508) является сложность процесса ферментации при низком уровне экспрессии (10 мг интерферона на 1 л культуральной среды). Более продуктивным является использование штаммов Escherichia coli (Semin. Oncol.,1997, Iun; 24 (3 Suppl. 9):S9-41-S9-51).
Известно большое количество плазмид и созданных на их основе штаммов Е. coli, экспрессирующих интерферон: штаммы Е. coli ATCC 31633 и 31644 с плазмидами Z-pBR322 (Psti) HclF-11-206 или Z-pBR 322(Pstl)/HclN SN 35-AHL6 (SU 1764515), штамм Е. coli pINF-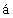 AP2 (SU 1312961), штамм Е. coli pINF-
AP2 (SU 1312961), штамм Е. coli pINF-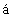 F-Pa (AU 1312962), штамм E.Coli SG 20050 с плазмидой p280/21FN (Кравченко В.В. и др. Биоорганическая химия, 1987, т.13, №9, с.1186-1193), штамм E.Coli SG 20050 с плазмидой pINF14 (SU 1703691), штамм E.coli SG 20050 с плазмидой pINF16 (RU 2054041) и др. Недостатком технологий, основанных на использовании этих штаммов, является их нестабильность, а также недостаточный уровень экспрессии интерферона.
F-Pa (AU 1312962), штамм E.Coli SG 20050 с плазмидой p280/21FN (Кравченко В.В. и др. Биоорганическая химия, 1987, т.13, №9, с.1186-1193), штамм E.Coli SG 20050 с плазмидой pINF14 (SU 1703691), штамм E.coli SG 20050 с плазмидой pINF16 (RU 2054041) и др. Недостатком технологий, основанных на использовании этих штаммов, является их нестабильность, а также недостаточный уровень экспрессии интерферона.
Наряду с особенностями используемых штаммов эффективность процесса во многом зависит от используемой технологии выделения и очистки интерферона.
Известен способ получения интерферона, включающий в себя культивирование клеток Ps. putida, разрушение биомассы, обработку полиэтиленимином, фракционирование сернокислым аммонием, гидрофобную хроматографию на фенилсилохроме С-80, рН-фракционирование лизата, его концентрирование и диафильтрацию, ионообменную хроматографию на целлюлозе DE-52, элюирование в градиенте рН, ионообменную хроматографию полученного элюента на целлюлозе СМ-52, концентрирование пропусканием через кассету фильтров и гель-фильтрацию на Сефадексе G-100 (SU 1640996). Недостатком этого способа кроме сложной многостадийной ферментации является многостадийность при получении конечного продукта.
Известен также способ получения интерферона, включающий в себя культивирование штамма E.coli SG 20050/pIF16, в LB-бульоне в колбах в термостатированном шейкере, центрифугирование биомассы, ее промывку буферным раствором и обработку ультразвуком для разрушения клеток. Полученный лизат центрифугируют, промывают 3М раствором мочевины в буфере, растворяют в растворе гуанидин хлорида в буфере, обрабатывают ультразвуком, центрифугируют, проводят окислительный сульфитолиз, диализ против 8 М мочевины, ренатурацию и окончательную двухстадийную хроматографию на СМ-52 целлюлозе и сефадексе G-50 (RU 2054041). Недостатками этого способа является его относительно невысокая производительность основных этапов процесса выделения и очистки. В особенности это относится к ультразвуковой обработке продукта, диализу и окислительному сульфитолизу, что приводит к нестабильности выхода интерферона, а также к невозможности использования этого метода для промышленного производства интерферона.
В качестве наиболее близкого аналога (прототипа) может быть указан способ получения лейкоцитарного интерферона человека, заключающийся в культивировании рекомбинантного штамма E.coli, замораживании полученной биомассы при температуре не выше -70°С, размораживании, разрушении клеток микроорганизма лизоцимом, удалении ДНК и РНК введением в лизат ДНК-азы и очисткой выделенной нерастворимой формы интерферона отмывкой буферным раствором с детергентами, растворении осадка интерферона в растворе гуанидин гидрохлорида, ренатурации и одностадийной очистке ионообменной хроматографией. В качестве продуцента используют штамм E.coli SS5, полученный с помощью рекомбинантной плазмиды pSS5, содержащей три промотора: Plac, Pt7 и Ptrp, и ген альфа -интерферона с введенными нуклеотидными заменами.
Экспрессия интерферона штаммом E.coli SS5, содержащим эту плазмиду, контролируется тремя промоторами: Plac, Pt7 и Ptrp. Уровень экспрессии интерферона составляет около 800 мг на 1 л клеточной суспензии (RU 2165455).
Недостатком способа является низкая технологичность использования ферментативного разрушения клеток, ДНК и РНК микроорганизма и одностадийная хроматографическая очистка интерферона. Это обуславливает нестабильность процесса выделения интерферона, приводит к снижению его качества и ограничивает возможность использования приведенной схемы для промышленного производства интерферона. Недостатками данной плазмиды и штамма на ее основе являются использование в плазмиде сильного нерегулируемого промотора фага Т7 в штамме Е. coli BL21 (DE3), в котором ген Т7 РНК полимеразы находится под промотором lac оперона и который всегда "течет". Следовательно, в клетке непрерывно происходит синтез интерферона, что приводит к диссоциации плазмиды и снижению жизнеспособности клеток штамма, и в результате - снижение выхода интерферона.
Задачей данного изобретения является конструирование рекомбинантого промышленного штамма продуцента Е. coli с помощью новой рекомбинантной плазмидной ДНК, обладающего высоким уровнем биосинтеза интерферона, и разработка эффективной промышленной технологии получения субстанции интерферона медицинского назначения, соответствующей по качеству "European Pharmacopoeia " для субстанции интерферона альфа-2b.
Указанная задача решалась созданием рекомбинантной плазмидной ДНК pSX50 и штамма Escherichia coli SX50, депонированный во Всероссийской коллекции промышленных штаммов ФГУП ГосНИИ генетики, номер ВКПМ В-8550,
а также способом получения рекомбинантного альфа-2b интерферона, основанным на использовании рекомбинантного штамма Е. coli SX50 и предусматривающим его глубинное культивирование на питательной среде с пониженным содержанием триптофана при непрерывном добавлении питательных субстратов в процессе биосинтеза, механическое разрушение клеток микроорганизма при высоком давлении, растворение агрегированного белка в концентрированном растворе гуанидин гидрохлорида с последующей ренатурацией интерферона в физиологических буферных растворах в присутствии хаотропных агентов и трехстадийной хроматографической очисткой интерферона на смолах типа Chelating Sepharose Fast Flow, иммобилизованной ионами Сu+2, ионообменной хроматографии на ионообменных смолах типа CM Sepharose Fast Flow и гель фильтрационной хроматографии на смолах типа Superdex 75.
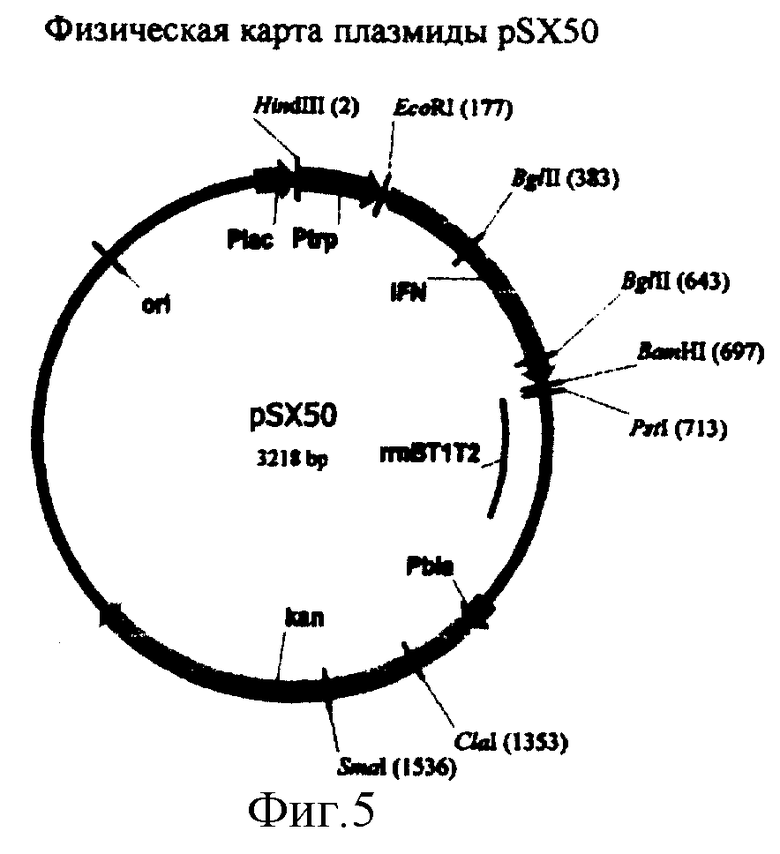
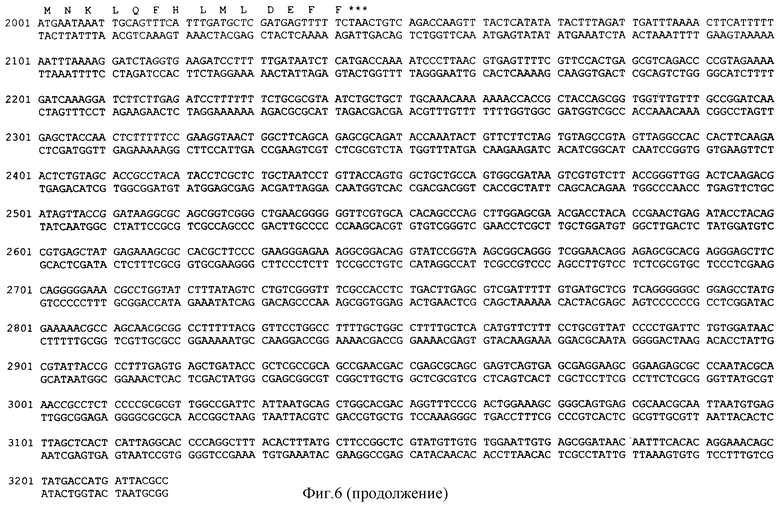
Согласно изобретению предлагается новая рекомбинантная мультикопийная плазмидная ДНК pSX50, кодирующая синтез лейкоцитарного альфа-2b интерферона человека, экспрессия которого находится под контролем лактозного и триптофанового промоторов и терминатора транскрипции. Плазмида pSX50 имеет 3218 пар оснований (п.о.) и характеризуется наличием следующих фрагментов:
- Последовательность с 1 нуклеотида по 176 нуклеотид (н.) включает фрагмент ДНК размером 176 п.о., содержащий триптофановый промотор (Ptrp);
- Последовательность с 177 н. по 194 н. включает синтетический фрагмент ДНК размером 18 п.о., содержащий последовательность Шайн Дельгарно, ответственный за инициацию трансляции;
- Последовательность с 195 н. по 695 н. включает фрагмент ДНК размером 501 п.о., содержащий последовательность гена интерферона со следующими нуклеотидными заменами: в положении 37 замена А на С, в положении 39 замена G на Т, в положении 40 замена А на С, в положении 42 замена G на Т, в положении 67 замена А на С, в положении 69 замена G на Т, в положении 70 замена А на С, в положении 72 замена А на Т, в положении 96 замена G на А, в положении 100 замена А на С, в положении 102 замена А на Т, в положении 114 замена А на С, в положении 120 замена С на G, в положении 126 замена G на А, в положении 129 замена G на А, в положении 330 замена С на G, в положении 339 замена G на А, в положении 342 замена G на А, в положении 487 замена А на С, в положении 489 замена А на Т, в положении 495 замена G на А;
- Последовательность с 696 н. по 713 н. включает синтетический фрагмент ДНК размером 18 п.о., содержащий синтетический полилинкер;
- Последовательность с 714 н. по 1138 н. включает фрагмент ДНК плазмиды рКК223-3 с 4129 н. по 4553 н. размером 425 п.о., содержащий последовательность строгого терминатора транскрипции rrnBT1T2;
- Последовательность с 1139 н. по 1229 н. включает фрагмент ДНК плазмиды pUC19 с 2487 н. по 2577 н. размером 91 п.о., содержащий промотор гена 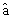 -лактомазы (ген устойчивости к ампициллину - АmрR);
-лактомазы (ген устойчивости к ампициллину - АmрR);
- Последовательность с 1230 н. по 2045 н. включает фрагмент ДНК плазмиды pUC4K с 720 н. по 1535 н. размером 816 п.о., содержащий структурную область гена kan;
- Последовательность с 2046 н. по 3218 н. включает фрагмент ДНК плазмиды pUC19 с 1625 по 453 н. размером 1173 п.н., содержащий последовательность, ответственную за репликацию плазмиды (ori) и lac промотора (Plac).
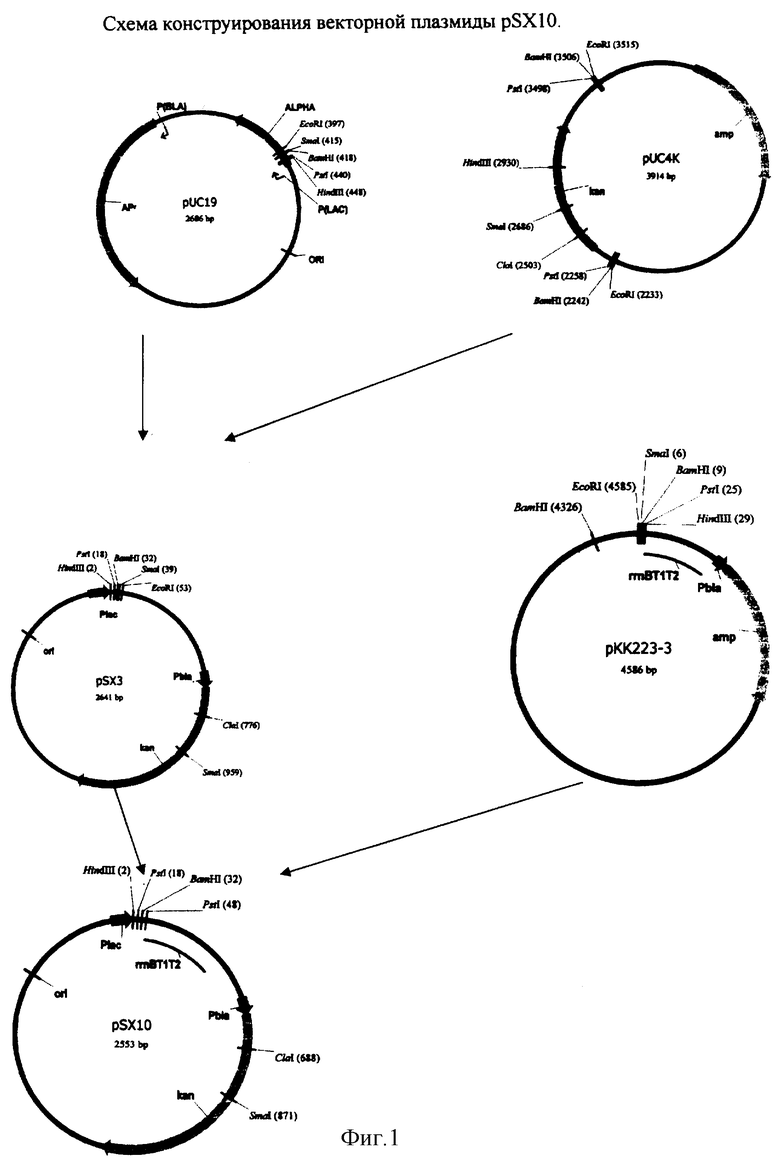
На фиг.1-5 изображены схемы конструирования и физическая карта плазмиды рSХ50.
На фиг.6 представлена установленная для плазмиды pSX50 полная последовательность нуклеотидов.
Штамм Escherichia coli SX50 получен трансформацией клеток Escherichia coli BL21 плазмидой pSX50 с использованием традиционной генно-инженерной технологии. Штамм E.Coli SX50 характеризуется следующими признаками.
Культурально-морфологические признаки
Клетки мелкие, прямые, утолщенной палочковидной формы, грамотрицательные, неспороносные. Клетки хорошо растут на простых питательных средах. При росте на агаре "Дифко" образуются круглые, гладкие, выпуклые, мутные, блестящие, серые колонии, с ровными краями. При росте в жидких средах (в минимальной среде с глюкозой или в LB-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки
Аэроб. Температурный диапазон роста 4-42°С при оптимуме рН 6,5-7,5.
В качестве источника азота используют как минеральные соли в аммонийной и нитратной формах, так и органические соединения в виде аминокислот, пептона, триптона, дрожжевого экстракта и т.д.
В качестве источника углерода используют аминокислоты, глицерин, углеводы. Устойчивость к антибиотикам. Клетки проявляют устойчивость к канамицину (до 100 мкг/мл).
Штамм Escherichia coli 8Х50 - продуцент интерферона.
Способ, условия и состав среды для хранения штамма
В L-arape с добавлением канамицина до концентрации 20 мкг/мл под маслом, в L-бульоне, содержащем 15% глицерина и соответствующими антибиотиками в ампулах при температуре минус 70°С, в лиофилизированном состоянии в ампулах при температуре плюс 4°С.
Штамм Escherichia coli SX50 идентифицирован по Определителю Берги (1974) как штамм вида Escherichia coli.
Способ промышленного получения альфа-2b интерферона
Особенностью заявляемого способа является разработка технологии, позволяющая выделять интерферон из нерастворимой формы, накапливающейся в течение ферментации, что позволяет существенно упростить технологическую схему процесса выделения и повысить выход целевого продукта.
Способ заключается в культивировании штамма Escherichia coli SХ50 в питательной среде, с постоянным добавлением питательных субстратов, предпочтительно глюкозы и дрожжевого экстракта, в процессе биосинтеза, предпочтительно с пониженным содержанием триптофана, механическом разрушении клеток микроорганизма при высоком давлении 700-900 bar, растворении интерферона в буферном растворе гуанидин гидрохлорида, ренатурации интерферона в физиологических буферных растворах в присутствии хаотропных агентов, с последующей трехстадийной хроматографической очистке интерферона на смолах типа Chelating Sepharose Fast Flow, иммобилизованных ионами Сu+2, ионообменную хроматографию на ионообменных смолах типа CM Sepharose Fast Flow и гель фильтрационную хроматографию на смолах типа Superdex 75.
Оптимальными условиями проведения отдельных стадий получения интерферона являются следующие:
- ферментацию проводят при непрерывном добавлении субстратов в течение всего процесса, что обуславливает высокий уровень экспрессии интерферона;
- разрушение клеток осуществляют в дезинтеграторе типа Гаулин при давлении 900 bar;
- удаление растворимых клеточных компонентов (ДНК, РНК, белков, липополисахаридов и т.д.) производят промыванием нерастворимой формы интерферона буферными растворами, содержащими детергенты (Тритон XI00, мочевина и т.д.);
- образовавшийся осадок, содержащий интерферон, растворяют в буферном растворе 6 М гуанидина гидрохлорида;
- ренатурацию интерферона проводят в физиологическом буферном растворе, содержащем хаотропные агенты;
- трехстадийную хроматографическую очистку интерферона проводят на Chelating Sepharose Fast Flow, иммобилизованной ионами Сu+2, на катионообменной смоле СМ Sepharose Fast Flow и гель фильтрационную хроматографию на смоле типа Superdex 75;
- после каждой хроматографической очистки проводят стерилизующую фильтрацию через апирогенные фильтры с размерами пор 0.22 мкм.
Выход интерферона в результате применения описанного способа составляет примерно 400-800 мг интерферона с 1 л культуральной среды. Качество получаемого продукта соответствует нормам и требованиям "European Pharmacopoeia" для субстанции альфа-2b интерферона.
Существенными отличиями заявляемого способа от прототипа являются:
- использование конструкции штамма, с более высокой производительностью, что позволяет получать при биосинтезе большее количество интерферона с 1 л культуральной среды;
- использование эффективного механического разрушения клеточной биомассы, что позволяет получать более чистый экстракт нерастворимой формы интерферона за более короткое время, с меньшими потерями;
- использование физиологических буферных растворов при ренатурации в присутствии хаотропных агентов позволяет повысить выход корректно ренатурированной формы интерферона;
- трехстадийная хроматографическая очистка интерферона позволяет получать субстанцию интерферона более 99% чистоты по данным электрофореза в редуцирующих и нередуцирующих условиях при окрашивании гелей серебром и более 98% по данным RF HPLC и практически не содержащую пирогенов (LAL-тест).
Сущность и преимущества заявляемой группы изобретений иллюстрируется следующими примерами.
Пример 1. Конструирование рекомбинантной плазмиды рSХ50
Способ конструирования плазмиды pSX50 включает следующие этапы:
- конструирование векторной плазмиды pSX10;
1. конструирование плазмиды pSX3 (2641 п.о.)
2. конструирование векторной плазмиды pSX10 (2553 п.о.)
- конструирование рекомбинантной плазмиды pSX41 (3218 п.о.);
- конструирование рекомбинантной плазмиды pSX43 (3218 п.о.);
- конструирование рекомбинантной плазмиды pSX45 (3218 п.о.);
- конструирование рекомбинантной плазмиды pSX50 (3218 п.о.).
Конструирование векторной плазмиды pSX10
Векторная плазмида pSX10 представляет собой вектор pUC19, в котором кодирующая последовательность гена бета лактомазы, обеспечивающая устойчивость к ампициллину, заменяется кодирующей последовательностью гена kаn и содержит терминатор транскрипции из плазмиды рКК223-3.
Конструирование векторной плазмиды pSS10 проводят в две стадии:
- получение плазмиды pSX3 (2641 п.о.), представляющей собой плазмиду pUC19, в которой кодирующая область аmp гена заменяется на кодирующую область гена kan;
- получение веторной плазмиды pSX10 (2553 п.о.), представляющей собой плазмиду pSX3, в которой за BamHI сайтом вставлен фрагмент ДНК, кодирующий терминатор транскрипции ггBT1T2.
Для получения плазмиды pSX3 проводят пять раундов амплификации ДНК методом ПЦР (полимеразная цепная реакция). В ходе первого раунда, используя ДНК плазмиды pUC19 в качестве матрицы, проводят амплификацию фрагмента ДНК размером 1828 п.о. (фрагмент PU1-PU2) с помощью праймеров:

Данную и последующие ПЦР реакции проводят в следующих условиях: 20 mМ Tis-HCl, рН 8.8, 10 mM (NH4)2SO4,10 mМ КСl, 2 тМ MgCl2, 0.1% Triton Х100,0.1 mg/ml BSA, 0.2 mM каждого dNTP, 1.25 ед. Pfu ДНК полимеразы, 100 нг ДНК. Процесс амплификации состоит из следующих стадий: прогревание при 95°С 5 мин, 35 циклов ПЦР (30 сек 95°С, 30 сек 56°С, 2 мин 72°С) и инкубация 10 мин при 72°С. После амплификации (и после последующих амплификаций) фрагмент ДНК очищают электрофоретически в 1% агарозном геле. В ходе второго и третьего раундов, используя ДНК плазмиды pUC4K в качестве матрицы, проводят амплификацию фрагмента ДНК размером 555 п.о. (фрагмент КМ1-КМ2) с помощью праймеров:
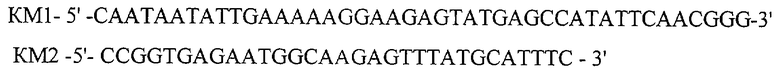
и амплификацию фрагмента ДНК размером 258 п.о. (КМЗ-КМ4) с праймеров
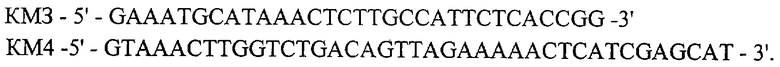
Далее фрагменты (КМ1-КМ2) и (КМ3-КМ4) объединяют в четвертом раунде амплификации, используя праймеры КМ1 и КМ4, получают фрагмент ДНК размером 813 п.о. (КМ1-КМ4), кодирующий структурную часть гена kan.
В пятом раунде ПЦР проводят объединение фрагментов (PU1-PU2) и (КМ1-КМ4) в следующих условиях: прогревание при 95°С 5 мин, 5 циклов ПЦР (30 сек 95°С, 30 сек 56°С, 10 мин 72°С) и инкубация 10 мин при 72°С. ДНК, полученную после последней ПЦР, прямо трансформируют в клетки штамма E.coli DH5 и высевают на среду LA, содержащую 20 мкг/мл канамицина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК и проводят рестрикционный анализ. В результате получают плазмиду рSХ3 размером 2641 п.о.
Для получения векторной плазмиды pSX10 проводят три раунда амплификации ДНК методом ПЦР. В ходе первого раунда, используя ДНК плазмиды pSX3 в качестве матрицы, проводят амплификацию фрагмента ДНК размером 2025 п.о. (фрагмент 10.1-10.2) с помощью праймеров:
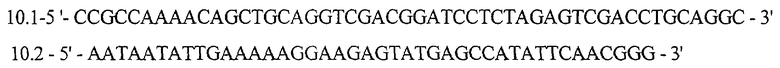
В ходе второго раунда, используя ДНК плазмиды рКК223-3 в качестве матрицы, проводят амплификацию фрагмента ДНК размером 528 п.о. (фрагмент КК1-КК2) с помощью праймеров:

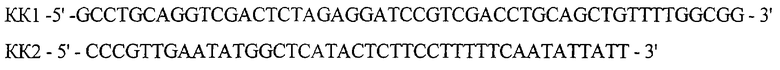
В третьем раунде ПЦР проводят объединение фрагментов (10.1-10.2) и (КК1-КК2) в следующих условиях: прогревание при 95°С 5 мин, 5 циклов ПЦР (30 сек 95°С, 30 сек 56°С, 10 мин 72°С) и инкубация 10 мин при 72°С. ДНК, полученную после последней ПЦР, прямо трансформируют в клетки штамма E.coli DH5 и высевают на среду LA, содержащую 20 мкг/мл канамицина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК и проводят рестрикционный анализ. В результате получают плазмиду pSX10 размером 2553 п.о.
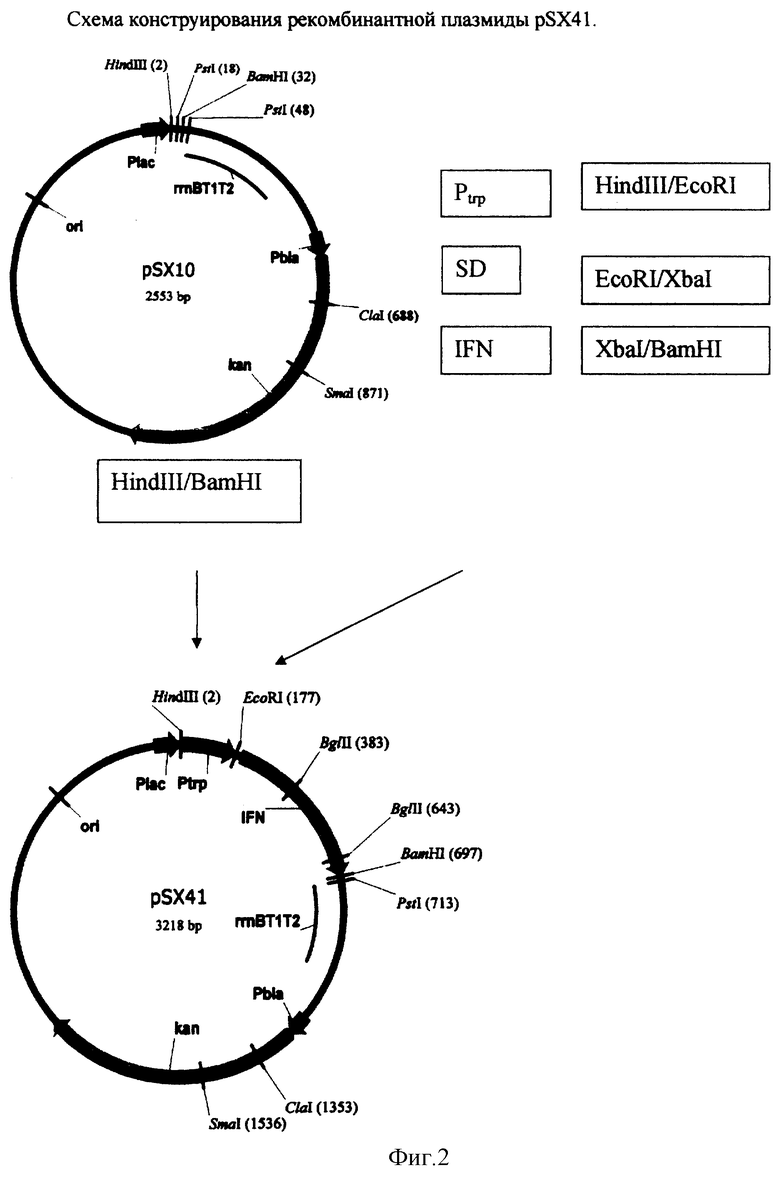
Конструирование рекомбинантной плазмиды pSX41
- Рекомбинантная плазмида pSX41 представляет собой Hind III - BamHI фрагмент ДНК векторной плазмиды pSX3 (2529 п.о.), Hind III - EcoRI фрагмент ДНК размером 168 п.о., кодирующий промотор триптофанового оперона Е. coli (Ptrp), EcoRI-XbaI синтетический фрагмент ДНК размером 20 п.о., кодирующий SD последовательность (Шайн-Дельгарно) и XbaI-BamHI фрагмент ДНК размером 501 п.о., кодирующий ген альфа 2b интеферона человека.
Для получения Hind III - ВаmHI фрагмент ДНК векторной плазмиды pSX3 (2529 п.о.) ДНК плазмиды pSX3 обрабатывают ферментами рестрикции HindIII и BamHI с последующей электрофоретической очисткой в 1% агарозном геле. Hind III EcoRI фрагмент ДНК размером 168 п.о., кодирующий промотор триптофанового оперона (Ptrp) получают методом ПЦР, используя тотальную ДНК Е. coli в качестве матрицы и праймеры TRP1 и PRP2 с последующей обработкой амплифицированного фрагмента рестриктазами Hindlll и EcoRI:
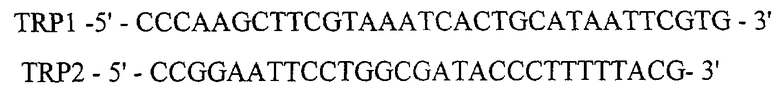
Для получения EcoRI-Xbal синтетический фрагмент ДНК размером 20 п.о., кодирующий SD последовательность (Шайн-Дельгарно), синтезируются следующие комплементарные олигонуклеотиды:
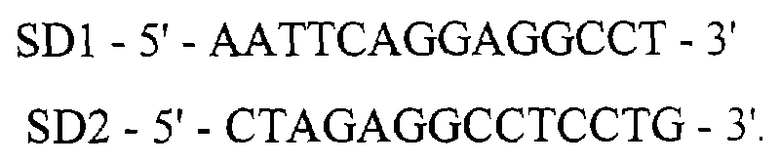
XbaI-ВаmIII фрагмент ДНК размером 501 п.о., кодирующий ген альфа 2b интеферона человека, получают методом ПЦР, используя тотальную ДНК человека в качестве матрицы и праймеры IFN1 и IFN2 с последующей обработкой амплифицированного фрагмента рестриктазами Xbal и ВаmIII:

Далее электрофоретически очищенные фрагменты объединяют, лигируют ферментом лигаза фага Т4, ДНК трансформируют в клетки штамма E.coli DH5 и высевают на среду LA, содержащую 20 мкг/мл канамицина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК, проводят рестрикционный анализ и определяют первичную структуру ДНК. В результате получают плазмиду pSX41 размером 3218 п.о. Далее проводят поэтапный мутагенез гена интерферона для увеличения уровня экспрессии целевого продукта. Мутагенез гена интерферона заключается в замене редко встречающихся в Е. coli триплетов, кодирующих соответствующие аминокислоты на часто встречающиеся в Е. coli триплеты, кодирующие эти же аминокислоты. Мутагенез ДНК гена интерферона проводят методом ПЦР.
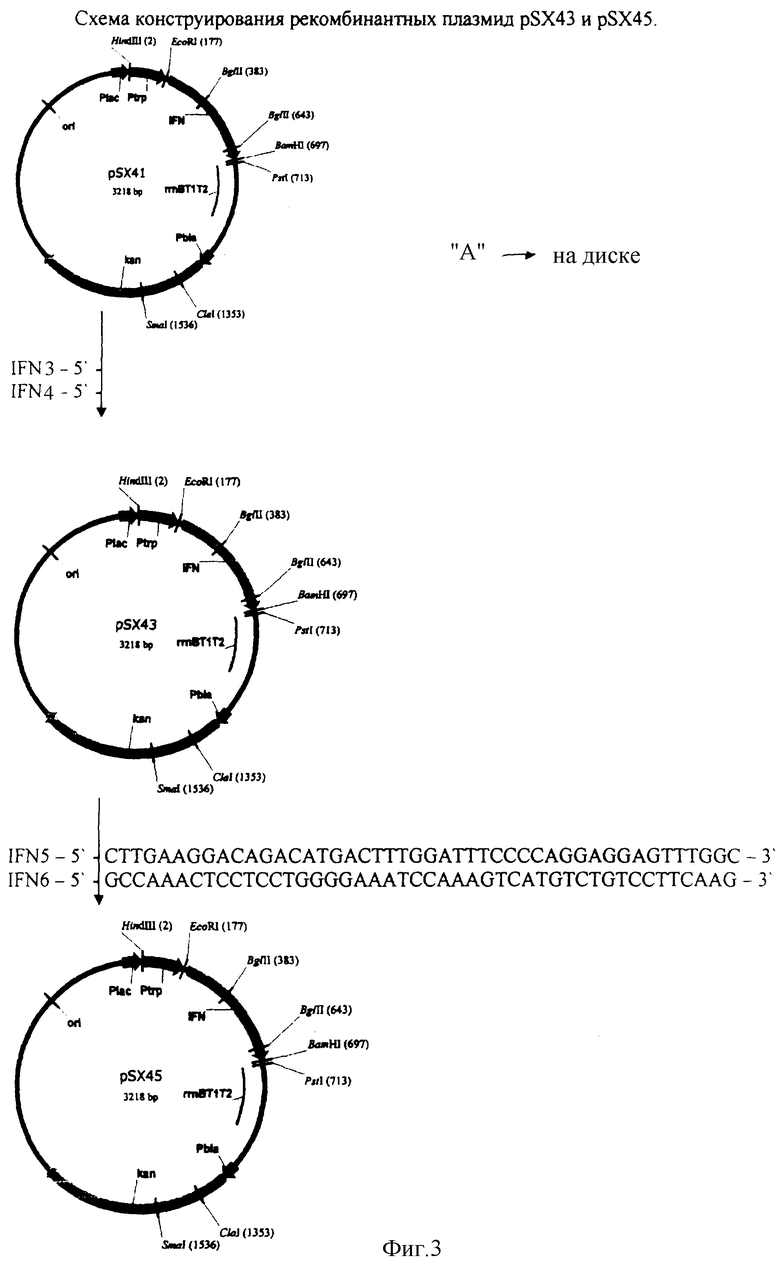
Конструирование рекомбинантной плазмиды pSX43
Для получения рекомбинантной плазмиды pSX43 проводят один раунд амплификации ДНК методом ПЦР, используя ДНК плазмиды pSX41 в качестве матрицы и праймеры IFN3 и IFN4:
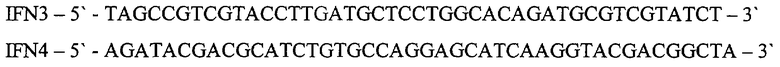
ПЦР проводят в следующих условиях: прогревание при 95°С 5 мин, 20 циклов ПЦР (30 сек 95°С, 30 сек 56°С, 10 мин 72°С) и инкубация 20 мин при 72°С. ДНК, полученную после ПЦР, прямо трансформируют в клетки штамма E.coli DH5 и высевают на среду LA, содержащую 20 мкг/мл канамицина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК, проводят рестрикционный анализ и определяют первичную структуру ДНК. В результате получают плазмиду рSХ43 размером 3218 п.о.
Конструирование рекомбинантной плазмиды pSX45
Для получения рекомбинантной плазмиды pSX45 проводят один раунд амплификации ДНК методом ПЦР, используя ДНК плазмиды pSX43 в качестве матрицы и праймеры IFN5 и IFN6:
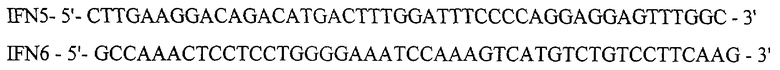
ПЦР проводят в следующих условиях: прогревание при 95°С 5 мин, 20 циклов ПЦР (30 сек 95°С, 30 сек 56°С, 10 мин 72°С) и инкубация 20 мин при 72°С. ДНК, полученную после ПЦР, прямо трансформируют в клетки штамма E.coli DH5 и высевают на среду LA, содержащую 20 мкг/мл канамицина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК, проводят рестрикционный анализ и определяют первичную структуру ДНК. В результате получают плазмиду pSX45 размером 3218 п.о.
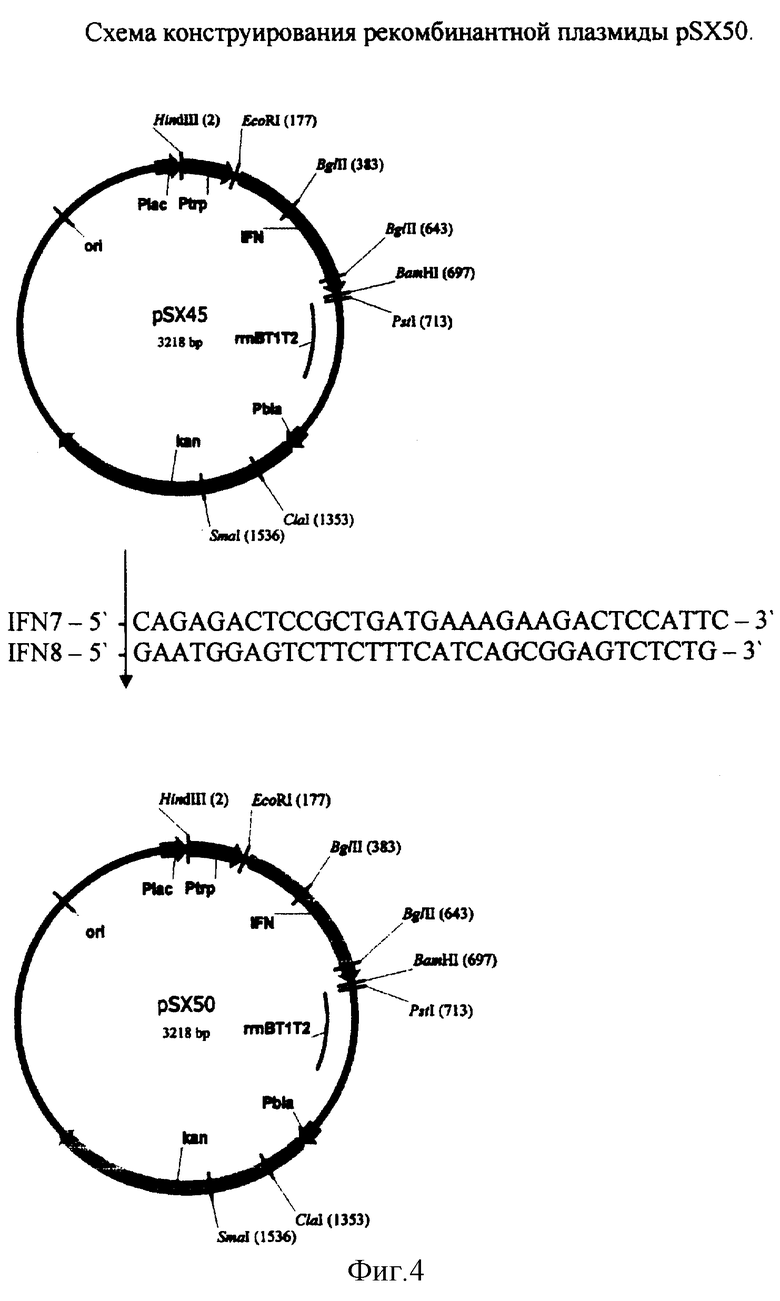
Конструирование рекомбинантной плазмиды pSX50.
Для получения рекомбинантной плазмиды pSX50 проводят один раунд амплификации ДНК методом ПЦР, используя ДНК плазмиды pSX45 в качестве матрицы и праймеры IFN7 и IFN8:

ПЦР проводят в следующих условиях: прогревание при 95°С 5 мин, 20 циклов ПЦР (30 сек 95°С, 30 сек 56°С, 10 мин 72°С) и инкубация 20 мин при 72°С. ДНК, полученную после ПЦР, прямо трансформируют в клетки штамма E.coli DH5 и высевают на среду LA, содержащую 20 мкг/мл канамицина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК, проводят рестрикционный анализ и определяют первичную структуру ДНК. В результате получают плазмиду pSX50 размером 3218 п.о.
Пример 2. Получение штамма Е. coli SX50 - продуцента интерферона
Штамм продуцент интерферона Е. coli SX50 получают путем трансформации клеток штамма Е. coli BL21 рекомбинантной плазмидой рSХ50. Штамм продуцент интерферона выращивают в 30 л ферментере до оптической плотности 25.0-30.0 о.е. в среде М9, содержащей 1% кислотного гидролизата казеина (Difco), 1% глюкозы, 40 мкг/мл канамицина, при температуре 38-39°С. В процессе ферментации проводят непрерывное добавление питательного субстрата, используя гравитометрический контроллер.
Содержание интерферона в биомассе клеток, получаемые с 1 л культуры, составляет в зависимости от серии 0.9-1.0 г интерферона.
Пример 3. Способ выделения интерферона из штамма Е. coli SX50
Получение интерферона проводили в 4 этапа:
1 этап. Культивирование штамма Е. coli SX50.
2 этап. Выделение и очистка нерастворимой формы интерферона.
3 этап. Растворение и ренатурация интерферона.
4 этап. Хроматографическая очистка интерферона.
1 этап. Культивирование штамма Е. coli SX50
Выращенный посевной материал штамма Е. coli SX50 в объеме 3 л богатой среде LB в течение 12 ч при 26°С асептически вносят в ферментер, содержащий 27 л стерильной среды, содержащей М9, 1% кислотного гидролизата казеина, 1% глюкозы, 1 мМ MgCl2, 0.1 mM CaCl2, 40 мг/мл канамицина. Культивирование в ферментере проводят при температуре 38-39°С, поддерживая рН 7±0,15 путем автоматической подтитровки 40%-ным раствором гидроокиси натрия. Концентрацию растворенного кислорода в диапазоне (50±10)% от насыщения поддерживают путем изменения скорости оборотов мешалки от 100 до 800 об/мин и подачи воздуха от 1 до 15 л/мин. Концентрацию субстратов, в частности глюкозы и дрожжевого экстракта, измеряют в течение ферментации и поддерживают их концентрацию путем изменения скорости подачи концентрированных растворов, через перистальтические насосы, используя гравиметрический контроллер.
Накопление интерферона в виде нерастворимой формы контролируют с помощью фазово-контрастной микроскопии, методом электрофореза в 15% полиакриламидном геле (SDS-PAAG) и методом обратнофазной высокоэффективной хроматографии (RF HPLC). Ферментацию останавливают по достижении максимальной оптической плотности (~ 25-30 о.е.) и остановки синтеза интерферона. По окончании ферментации культуральную жидкость сепарируют центрифугированием в проточном роторе при скорости вращения 5000-10000 об/мин. Биомассу фасуют в полиэтиленовые пакеты и замораживают при температуре минус 70°С.
2 этап. Выделение и очистка нерастворимой формы интерферона
300-400 г замороженной биомассы штамма Е. coli SX50 суспедируют в 3000 мл буфера 1 (20 мМ Tris-HCl, pH 8.0, 10 мМ ЭДТА, 0,1% Triton X100). Суспензию пропускают через проточный гомогенизатор типа Гаулин, поддерживают давление 900 бар и центрифугируют в проточном роторе при 15 000 об/мин. Полученный осадок промывают в аналогичных условиях последовательно буферами 2 (20 мМ Tris-HCl, pH 8.0, 1 мМ ЭДТА, 3 М мочевины) и буфером 3 (20 мМ Tris-HCl pH 8.0, 1 мМ ЭДТА) и окончательно осадок интерферона суспедируют в 200 мл буфера 3. При этом время выделения и очистки нерастворимой формы интерферона составляет не более 5 час.
3 этап. Растворение и ренатурация интерферона
К полученной на предыдущем этапе суспензии нерастворимой формы интерферона добавляют сухой гуанидин гидрохлорид до концентрации 6 М, добавляют дитиотреитол до концентрации 50 мМ, Tris-HCl pH 8.0 до концентрации 50 мМ, NaCl до концентрации 150 мМ и Triton X100 до концентрации 0.1%, инкубируют при комнатной температуре в течение 2 ч. Нерастворившийся материал отделяют при стерилизующей фильтрации через мембраны с диаметрами пор 0.22 микрона.
Ренатурацию интерферона проводят путем медленного разбавления полученного раствора в 100-200 раз буфером 4 (20 мМ Tris-HCl pH 8.0, 100 мМ NaCl, 0.1 мМ ЕДТА). После чего ренатурационную смесь инкубируют при постоянном перемешивании в течение 12-15 час при температуре 4-8°С. Далее добавляют сульфат магния до концентрации 1 мМ и агрегированный материал удаляют стерилизующей фильтрацией через мембранный фильтр с диаметром пор 0.22 микрона.
4 этап. Хроматографическая очистка интерферона
Хроматографическую очистку интерферона осуществляют в три стадии.
1. Полученный ренатурированный интерферон на первом этапе подвергают очистке с помощью аффинной хроматографии на смоле типа Chelating Sepharose Fast Flow (Amersham Biosciences), иммобилизованной ионами Cu+2. Для этого раствор интерферона наносят на колонну с Cu+2 Chelating Sepharose Fast Flow и интерферон элюируют буфером 0.1 М лимонной кислоты pH 2.2.
2. На второй стадии хроматографической очистки раствор интерферона наносят на катионообменную смолу типа CM Sepharose Fast Flow (Amersham Biosciences) и интерферон элюируют градиентом растворов (0.0-0.5 М NaCl) в буфере 50 мМ Nа(СН3СОО), рН 5.5.
3. Очистка мономерной формы интерферона от остатков полимерных форм интерферона проводят на третьей стадии очистки интерферона гель-фильтрацией на смоле типа Superdex 75 (Amersham Biosciences). Хроматографию проводят в буфере 50 мМ Na(CH3COO), рН 5.0, содержащем 0.15М NaCl.
Описанный способ выделения и очистки интерферона позволяет получить 4-8 г высокоочищенного интерферона за один цикл выделения в течение 7-10 дней из биомассы, полученной с 10 л культуральной среды. Качество получаемого интерферона в полной мере соответствует требованиям "European Pharmacopoeia" для субстанции интерферона альфа-2b, а именно:
- Концентрация интерферона не менее 2×108 МЕ/мл;
- Удельная активность интерферона не менее 2.0×108 МЕ/мг;
- Электофоретическая чистота препарата не менее 99% в редуцирующих и не редуцирующих условиях при окрашивании гелей серебром;
- Содержание правильно ренатурированной формы интерферона не менее 98% по данным RF HPLC;
- Изоэлектрическая точка выделенного интерферона находится в районе рН 5.8-6.3;
- Пептидная карта выделенного интерферона принципиально не отличается от пептидной карты для Европейского стандарта интерферона альфа 2b CRS;
- Содержание бактериальных эндотоксинов не более 100 ME на 1 мг интерферона.
Как следует из приведенных примеров, заявляемая группа изобретений позволяет получать интерферон альфа-2b с высоким выходом при относительно простой и надежной технологии.


Изобретение относится к генной инженерии, биотехнологии, медицине, фармакологии. Новая рекомбинантная мультикопийная плазмидная ДНК pSX50, кодирующая синтез лейкоцитарного альфа-2b интерферона человека, экспрессия которого находится под контролем лактозного и триптофанового промоторов и терминатора транскрипции. В результате трансформации клеток реципиентного штамма Е. coli BL21 рекомбинантной плазмидной ДНК pSX50 получен штамм Е. coli SX50 - продуцент рекомбинантного лейкоцитарного альфа-2b интерферона человека с продуктивностью до 0.9-1.0 г альфа-2b интерферона с 1 л культуральной среды. Способ получения рекомбинантного альфа-2b интерферона основан на использовании созданного рекомбинантного штамма Е. coli SX50 и предусматривающий его глубинное культивирование на питательной среде с пониженным содержанием триптофана при непрерывном добавлении питательных субстратов в процессе биосинтеза, механическое разрушение клеток микроорганизма при высоком давлении, растворение агрегированного белка в концентрированном растворе гуанидин гидрохлорида с последующей ренатурацией интерферона в физиологических буферных растворах в присутствии хаотропных агентов и его очисткой с использованием трехстадийной хроматографической очистки интерферона на смолах типа Chelating Sepharose Fast Flow иммобилизованной ионами Cu+2, ионообменной хроматографии на ионообменных смолах типа СМ Sephsrose Fast Flow и гель фильтрационной хроматографии на смолах типа Superdex 75. Способ позволяет получать субстанцию интерферона более 99% чистоты по данным электрофореза в редуцирующих и нередуцирующих условиях при окрашивании гелей серебром и более 98% по данным RF HPLC и не содержащую пирогенов (LAL-тест) в количествах не менее 400-800 мг с 1 л культуральной среды. 3 н. и 3 з.п ф-лы, 6 ил.
 -лактомазы (ген устойчивости к ампициллину -AmpR), последовательность с 1230 по 2045 н. включает фрагмент ДНК плазмиды pUC4K с 720 н. по 1535 н. размером 816 п.о., содержащий структурную область гена kan, последовательность с 2046 н. по 3218 н. включает фрагмент ДНК плазмиды pUC19 с 1625 по 453 н. размером 1173 п.н., содержащий последовательность, ответственную за репликацию плазмиды (ori) и lac промотора (Plac).
-лактомазы (ген устойчивости к ампициллину -AmpR), последовательность с 1230 по 2045 н. включает фрагмент ДНК плазмиды pUC4K с 720 н. по 1535 н. размером 816 п.о., содержащий структурную область гена kan, последовательность с 2046 н. по 3218 н. включает фрагмент ДНК плазмиды pUC19 с 1625 по 453 н. размером 1173 п.н., содержащий последовательность, ответственную за репликацию плазмиды (ori) и lac промотора (Plac).
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pSS5, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА, ШТАММ ESCHERICHIA COLI SS5 - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА И СПОСОБ ПОЛУЧЕНИЯ ИНТЕРФЕРОНА АЛЬФА-2b | 1999 |
|
RU2165455C1 |
| НУКЛЕИНОВО-КИСЛОТНЫЙ КОНСТРУКТ ДЛЯ ЭКСПРЕССИИ АКТИВНЫХ ВЕЩЕСТВ, КОТОРЫЕ МОГУТ БЫТЬ АКТИВИРОВАНЫ ПРОТЕАЗАМИ, ПОЛУЧЕНИЕ И ИСПОЛЬЗОВАНИЕ | 1998 |
|
RU2204414C2 |
| SU 1591484 A1, 20.05.1995 | |||
| Способ получения человеческого лейкоцитарного интерферона альфа-2 | 1984 |
|
SU1364343A1 |

