Изобретение относится к области биотехнологии и органической химии, к способам получения изотопомодифицированных природных соединений, а именно: L-α-аминокислот, и может найти применение в экспериментальной биологии, медицине, ветеринарии, сельском хозяйстве.
Известен способ получения равномерно изотопомодифицированных природных L-a-аминокислот путем инкубации клеток микроорганизмов на среде, содержащей единственный источник углерода в виде изотопомодифицированных солей уксусной кислоты с последующим выделением целевого продукта. В качестве изотопомодифицированной соли в данном известном способе используют 14C ацетат натрия. Клетки микроорганизма Bacillus pumilus при росте в присутствии данного источника углерода включают 14C в вещество своего тела. После завершения инкубации белок клеток гидролизуют и из гидролизата выделяют целевые продукты равномерные изотопомодифицированные природные L-a-аминокислоты.
Недостатком известного способа является сложная процедура разделения гидролизата белка клеток микроорганизма на индивидуальные изотопомодифицированные L-a-аминокислоты. Кроме того, данным известным способом получают L-a-аминокислоты, модифицированные только изотопом 14C. Последнее время возрос интерес к природным соединениям, в том числе, к аминокислотам, меченым стабильным изотопом 13C. Но для работы с ними такие соединения требуются в больших количествах, недоступных для получения их данным известным способом. Большая часть природных L-a-аминокислот, меченых стабильным изотопом 13C, в настоящее время еще не получена и не описана. Не описаны и природные L-a-аминокислоты, которые равномерно модифицированы двумя изотопами: 14C и 15N или 13C и 15N.
Техническим результатом, достигаемым при реализации настоящего изобретения, является получение новых соединений, равномерно изотопомодифицированных изотопом 13C или изотопами 13C и 15N, а также упрощение технологии получения природных L-a-аминокислот, равномерно модифицированных изотопом 14C или изотопами 14C и 15N.
Достигается это тем, что в способе получения равномерно изотопомодифицированных природных L-a-аминокислот путем инкубации клеток микроорганизмов на среде, содержащей единственный источник углерода в виде изотопомодифицированных солей уксусной кислоты, с последующим выделением целевого продукта отличительной особенностью является то, что используют клетки штаммов микроорганизмов продуцентов целевого продукта, которые предварительно инкубируют на среде, содержащей источники углерода в виде солей уксусной кислоты в сочетании с углеводами с последующим отделением и отмыванием полученных клеток, а по завершении инкубации на среде с равномерно изотопомодифицированными солями уксусной кислоты клетки отделяют и отбрасывают, а целевой продукт выделяют из среды.
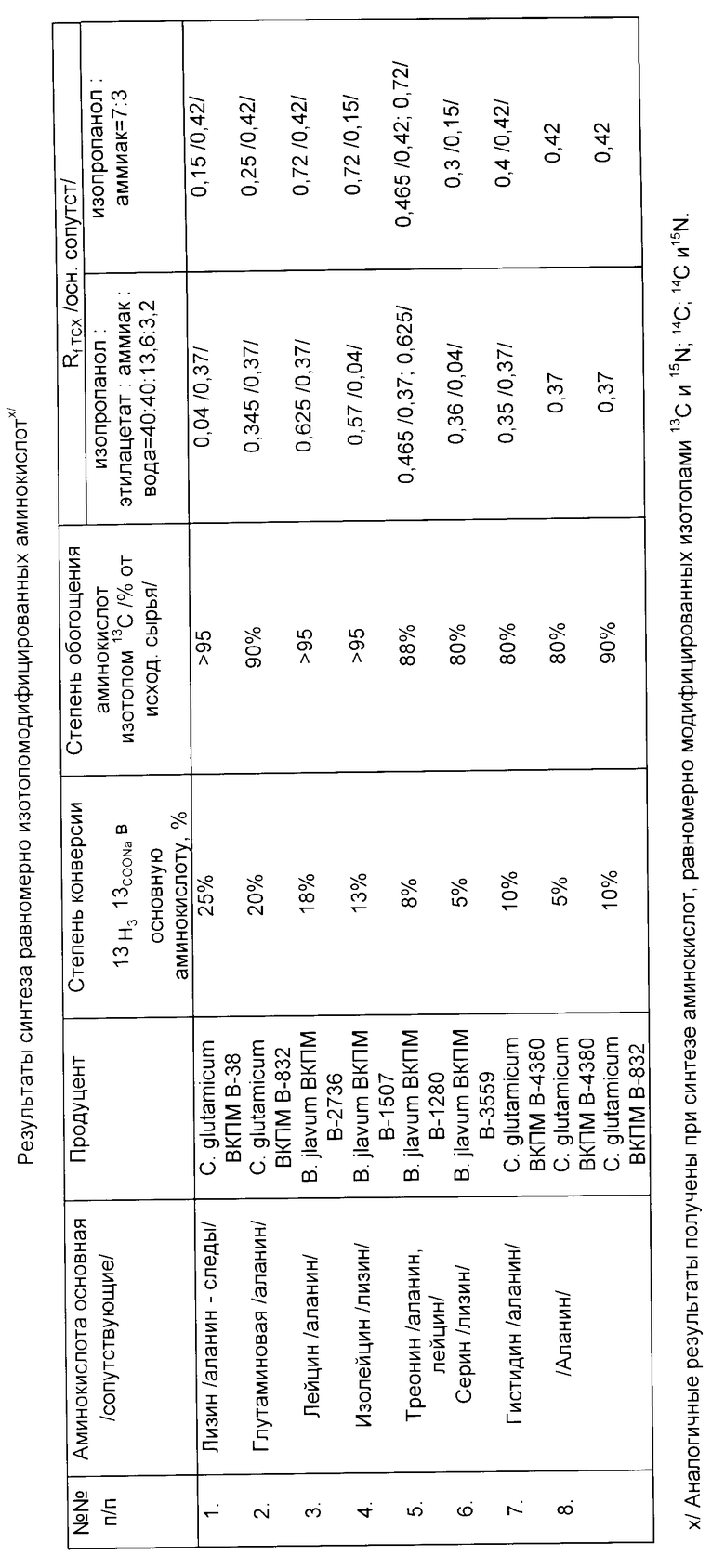
При этом в качестве штаммов продуцентов L-a-аминокислот используют продуцент лизина Corynebacterium glutamicum ВКПМ В-38, продуцент глутаминовой кислоты Corynebacterium glutamicum ВКПМ В-832, продуцент лейцина Brevibacterium flavum ВКПМ В-2736, продуцент изолейцина Brevibacterium flavum ВКПМ В-1570, продуцент треонина Brevibacterium flavum ВКПМ В-1280, продуцент серина Brevibacterium flavum ВКПМ В-3559, продуцент гистидина Corynebacterium glutamicum ВКПМ В-4380, причем в качестве продуцента аланина используют продуцент глутаминовой кислоты Corynebacterium glutamicum ВКПМ В-832 или продуцент гистидина Corynebacterium glutamicum ВКПМ В-4380.
В способе в качестве солей уксусной кислоты используют ацетат натрия и/или ацетат аммония, немодифицированные или равномерно модифицированные изотопами 13C или 14C, причем ацетат аммония может быть дополнительно модифицирован изотопом 15N.
В способе выделения наряду с целевой изотопомодифицированной аминокислотой сопутствующих ей аминокислот повышается степень конверсии изотопов углерода из изотопомодифицированных солей уксусной кислоты в получаемые изотопомодифицированные аминокислоты. Для этого после пропускания надосадочной жидкости инкубационной смеси через Дауэкс и элюции раствором аммиака элюат дополнительно пропускают через Амберлит GC-50 II в медной форме, фракции аминокислот элюируют раствором аммиака и пропускают через карбоксилметилцеллюлозу в Н+ форме.
Получены новые соединения природные L-a-аминокислоты: аланин, лизин, глутаминовая кислота, лейцин, изолейцин, треонин, серин и гистидин, равномерно модифицированные изотопом 13C или изотопом 13C и 15N.
Пример 1. Ночную культуру клеток штаммов-продуцентов, выросшую на полной питательной среде, выращивают на солевой среде, содержащей 1% ацетата натрия, 0,2% глюкозы и минеральные соли: KH2PO4, NaCl, /NH4/2SO4 и MgSO4; среда имеет нейтральное значение pH. По мере потребления культурой иона ацетата величина pH среды повышается, в связи с чем среду периодически нейтрализуют добавлением уксусной кислоты. По достижении культурой поздней логарифмической фазы роста биомассу осаждают центрифугированием и заливают тем же объемом той же среды, но без глюкозы. При этом свежая среда содержит равномерно модифицированный ацетат натрия и/или ацетат аммония. Концентрация клеток в свежей среде оказывается высокой и поэтому в процессе инкубации полученной суспензии клеток они практически не растут, но катализируют биосинтез равномерно меченых аминокислот из равномерно меченых солей уксусной кислоты, что сопровождается повышением величины pH инкубационной смеси. Смесь периодически нейтрализуют добавлением соляной кислоты. Время инкубации 1-2,5 сут при 30-37oC в зависимости от используемого штамма.
В тех случаях, когда клетки накапливают только одну аминокислоту, инкубационную смесь после завершения инкубации центрифугируют, надосадочную жидкость пропускают через ионообменную смолу Дауэкс и после элюции раствором аммиака выделяют из элюата целевую аминокислоту. Если используемый штамм-продуцент накапливает в инкубационной смеси не только целевую, но и сопутствующие ей иные аминокислоты, то их тоже выделяют.
Разделение основной и сопутствующих аминокислот осуществляют с помощью лигандообменной хроматографии на карбоксильном сорбенте, заполненном ионами меди. Степень зарядки составляет 100, 70, 30% в зависимости от набора аминокислот в инкубационной среде. Таким сорбентом служит Амберлит GC-50 II (частицы размером 200-400 меш/, через него пропускают элюат аминокислот, полученный после пропускания надосадочной жидкости через Дауэкс 50 и удаления аммиака. При элюции раствором аммиака собирают фракции аминокислот в виде их медных комплексов. Удаляют медь пропусканием фракций элюата через карбоксиметилцеллюлозу в H+ форме.
Пример 2. Получение изотопомодифицированного лизина.
Ночную культуру клеток штамма Corynebacterium glutamicum ВКПМ В-38, продуцента лизина, засевают на среду, содержащую 0,2% глюкозы, 0,1% KH2PO4, 1% /NH4/2SO4, 0,04% MgSO4, 1,5% ацетата натрия и 0,05% NaCl и инкубируют при 30oC в течение суток. Полученные клетки отделяют центрифугированием, отмывают, заливают тем же объемом той же солевой среды, но без глюкозы, содержащей равномерно изотопомодифицированный ацетат натрия и/или ацетат аммония. Суспензию клеток инкубируют при 30oC 8 ч. Инкубационную смесь центрифугируют, надосадочную жидкость пропускают через Дауэкс 50•8 в NH
Пример 3. Получение изотопомодифицированной глутаминовой кислоты
В качестве штамма-продуцента использована культура Corynebacterium glutamicum ВКПМ В-832. При инкубации суспензии отмытых клеток в условиях по примеру 2, но в течение 16 ч наряду с глутаминовой кислотой наблюдают образование в небольших количествах аланина. Инкубационную смесь центрифугируют, надосадочную жидкость пропускают через Дауэкс 1•8 в формиатной форме. Элюируют 0,1 н. раствором муравьиной кислоты. Аланин очищают на Дауэкс 50•8 в H+-форме, элюируя 0,5 н. раствором аммиака (см. таблицу).
Пример 4. Получение изтопомодифицировнных лейцина и изолейцина
В качестве штаммов-продуцентов используют культуры Brevibacterium flavum ВКПМ В-2736 и Brevibacterium flavum ВКПМ В-1507 соответственно. Инкубируют суспензии отмытых клеток по примеру 2 на среде с 2% изотопомодифицированного ацетата натрия в течение 1,5 сут. После пропускания надосадочной жидкости инкубационной смеси через Дауэкс 50 для выделения аминокислот используют смолу амберлит GC-50 II в медной форме со степенью зарядки медью 70% Элюент 0,2 н. раствор аммиака. После удаления аммиака из полученных фракций их пропускают через карбоксиметилцеллюлозу в H+-форме. Результаты приведены в таблице.
Пример 5. Получение изотопомодифицированного треонина
В качестве штамма-продуцента используют культуру Brevibacterium flavum ВКПМ В-1280 (суспензию отмытых клеток инкубируют по примеру 2, но на среде с 1% изотопомодифицированного ацетата натрия и 0,05% глюкозы). Кроме треонина, клетки образуют лейцин и аланин. Разделение смеси аминокислот производят на смоле Амберлит GC-50 II со 100%-ной зарядкой медью. Элюент 0,2 н. раствора аммиака (см. таблицу).
Пример 6. Получение изтопомодифицированного серина
В качестве штамма-продуцента используют культуру Brevibacterium flavum ВКПМ В-3559. Биомассу клеток выращивают по примеру 2, но в течение 2 сут (суспензию отмытых клеток инкубируют по примеру 2, но на среде с 1% изотопомодифицированного ацетата натрия и 0,05% глюкозы). Кроме серина, культура образует аланин и глутаминовую кислоту. Разделение аминокислот в соответствии с примером 5. Результаты приведены в таблице.
Пример 7. Получение изотопомодифицированных гистидина и аланина
Использован штамм-продуцент гистидина Corynebacterium glutamicum ВКПМ В-4380, который дополнительно синтезирует аланин. Биомассу клеток получают на солевой среде с 0,1% глюкозы. Для разделения аминокислот используют Амберлит GC-50 II, заряженный медью на 30% Элюируют 0,5 н раствором аммиака. Остальные операции по примерам 1-2. Результаты приведены в таблице.
| название | год | авторы | номер документа |
|---|---|---|---|
| L-ЛИЗИН-ПРОДУЦИРУЮЩАЯ БАКТЕРИЯ Corynebacterium glutamicum ИЛИ Brevibacterium flavum, УСТОЙЧИВАЯ К ФУЗИДИНОВОЙ КИСЛОТЕ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА L-ЛИЗИНА С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2013 |
|
RU2549690C1 |
| L-Лизин-продуцирующая коринеформная бактерия с мутантным геном fusA и способ получения L-лизина с использованием этой бактерии | 2015 |
|
RU2612159C1 |
| L-Лизин-продуцирующая коринеформная бактерия с инактивированным геном ltbR и способ получения L-лизина с использованием этой бактерии | 2016 |
|
RU2639247C1 |
| ШТАММ БАКТЕРИЙ BREVIBACTERIUM FLAVUM - ПРОДУЦЕНТ L-ГЛУТАМИНА (ВАРИАНТЫ) | 1994 |
|
RU2084520C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-СЕРИНА | 1991 |
|
RU2008356C1 |
| КОСМИДНЫЙ ВЕКТОР PC LF*993 ДЛЯ КЛОНИРОВАНИЯ ФРАГМЕНТОВ ДНК В КОРИНЕБАКТЕРИЯХ | 1992 |
|
RU2063440C1 |
| ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ L-ГИСТИДИНА | 1997 |
|
RU2119536C1 |
| ШТАММ БАКТЕРИЙ BREVIBACTERIUM SP. - ПРОДУЦЕНТ ЛИЗИНА | 1991 |
|
RU2027761C1 |
| ШТАММ БАКТЕРИЙ BREVIBACTERIUM SP. - ПРОДУЦЕНТ L-ЛИЗИНА | 1993 |
|
RU2034921C1 |
| Бактерия Corynebacterium glutamicum с повышенной способностью продуцировать L-валин и способ получения L-валина с использованием этой бактерии | 2020 |
|
RU2753996C1 |
Использование: биотехнология, органическая химия, получение изотопомодифицированных природных аминокислот, используемых в экспериментальной биологии, ветеринарии, медицине, сельском хозяйстве. Сущность изобретения: равномерно изотопомодифицированные природные аминокислоты получают инкубацией штаммов - продуцентов соответствующей аминокислоты. При этом сначала клетки выращивают на среде с глюкозой и с солью уксусной кислоты в качестве источников углерода. Затем клетки отделяют, отмывают и готовят их суспензию на солевой среде с единственным источником углерода в виде изотопомодифицированной соли уксусной кислоты в виде ацетата натрия и/или ацетата аммония, модифицированных изотопами 13C, или 14C и/или 15N. Впервые получены новые соединения, модифицированные 13C и 15N. 4 з.п. ф-лы, 1 табл.
| FR, патент, 2128844, кл | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
