Изобретение относится к новому производному тканевого активатора плазминогена t-PA, ДНК-последовательности, которая кодирует новое -t-PA-производное, плазмидам экспрессии, содержащим кодирующую t-PA-производное ДНК-последовательность, способу получения t-PA-производного и композиции с фибринолитической активностью, содержащей t-PA-производное.
Свернувшаяся кровь содержит в качестве основной компоненты полимерного белкового матрикса фибрин. При физиологических условиях фибрин растворяется благодаря последовательности реакции с участием фибринолитической системы. Центральной реакцией при этом является активирование плазминогена в плазмин, которое вызывается, например, тканевым активатором плазминогена (t-PA). Плазмин, в свою очередь, растворяет фибрин.
Ферментативная активность природного или полученного генноинженерным методом в клетках эукариот t-PA, а именно каталитическое активирование плазминогена в плазмин, в отсутствие фибрина или продуктов расщепления фибриногена очень незначительна, однако в присутствии этих продуктов может повышаться примерно на порядок.
В крови t-PA под действием протеаз расщепляется на A- и B-цепь. Обе образовавшиеся цепи остаются связанными через цистеиновый мостик. Возможность простимулировать активность t-PA представляет собой решающее преимущество по сравнению с другими известными активаторами плазминогенов, например урокиназой или стрептокиназой (M.Hoylaerts и и др., J."Biol. Chem". 257(1982), 2912 - 2919; W.Nieuwenhuizen и др., Biochem Biophys.Acta, 755, (1983) 531 - 533).
Механизм действия t-PA in vivo описан, например, в Korniger u Collen, Thromb. Hamostasis, 46 (1981), 561-565. Присущая t-PA активность позволяет использовать его как средство для борьбы с патологическими состояниями, связанными с образованием тромбов (EP-A 0302456, кл. C 12 N 9/54, 1989, прототип), что непосредственно подтверждено и клиническими испытаниями [Cokken и др., Circulation 70 (1984), 112; Circulation 73 (1986/511)].
Однако в качестве недостатка t-PA нужно рассматривать быстрое уменьшение его концентрации в плазме (клиренс). Следствием этого является то, что необходимо относительно большое количество t-PA, чтобы получить in vivo эффективное лизирование тромбов. Высокие же лечебные дозы оказывают часто побочные действия, в частности вызывают кровотечение.
В европейском патенте N 019620 описан природный продукт распада t-PA, который содержит только домены протеазы и крингель-11 и N-конец которого начинается с аланина 160 (в указанной Pennica и др. в Nature 301 (1983), 214 - 221, последовательности аминокислот).
Скорость клиренса этого продукта распада t-PA, однако, несущественно отличается от таковой природного t-PA. Лишь благодаря химической модификации каталитической области за счет введения блокирующей группы этот показатель можно улучшить.
Задача изобретения - изменение t-PA таким образом, чтобы образовавшееся производное обладало сильно сниженной скоростью клиренса (более длительным временем полураспада в плазме крови), при этом должны сохраняться лизирующее тромбы действие, а также стимулируемость фибрином.
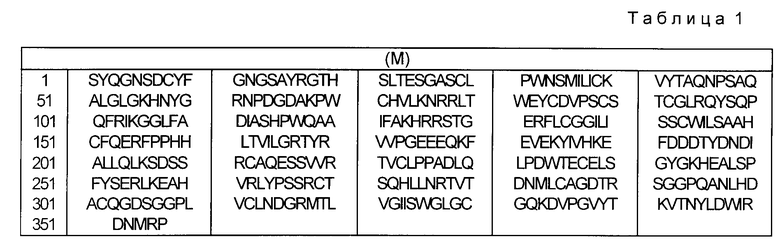
Предметом изобретения поэтому является измененный тканевой активатор плазминогена t-PA-производное), который отличается тем, что он не гликозилирован и представлен последовательностью аминокислот (см. табл. 1).
которая на амино-конце может также иметь дополнительный Met.
Как выяснилось, делеция части имеющихся в нативном t-PA областей не оказывает никакого влияния на тромболитическую эффективность протеина и зависящую от фибрина стимуляцию мутеина, которая сравнима с таковой нативного t-PA. Правда, установлено, что в предлагаемом согласно изобретению t-PA-производном отсутствует свойство связывать фибрин, но несмотря на это, фибринолитическая активность in vivo не только не снижена, но даже существенно улучшена по сравнению с таковой нативного t-PA. Неожиданным также является тот факт, что при введении тромболитически эффективной дозы производного почти не наблюдается влияния на системный фибронолиз.
Таким образом, оказалось, что предлагаемое согласно изобретению t-PA-производное обладает типичной для нативного t-PA специфичностью к фибрину при физиологических условиях. Эти результаты получены при фармакологических исследованиях предлагаемого согласно изобретению t-PA-производного (примеры 6 и 7). Кроме того, предлагаемый протеин обладает очень высокой удельной (специфической) активностью. При применении описанного далее способа ренатурирования получены формы в активностью 500-800 и KU/мг.
Следующим предметом изобретения является ДНК-последовательность, которая кодирует предлагаемое согласно изобретению t-PA-производстное и имеет нуклеотидную последовательность (см. табл. 2).
Предлагаемая в изобретении ДНК-последовательность используется в составе рекомбинантного вектора для экспрессии нового t-PA производного. Рекомбинантная плазмида для t-PA экспрессии составляет следующий предмет изобретения. Такая плазмида экспрессии может также включать ДНК-последовательность, которая колируя предлагаемое в изобретении t-PA-производное, может отклоняться от указанной ДНК-последовательности в рамках вырожденности кода.
Плазмида экспрессии содержит, наряду с кодирующей t-PA-производное последовательностью регулируемый промотор (например, tac), эффективный терминатор (например, td), селективный маркер (например, ген- β -лактамазы) и область начала репликации.
Конкретным примером такой плазмиды является pA 27.3, получение которой описано в примере 1.
Понятно, что выбор плазмиды, в которую внедряется кодирующая предполагаемое в изобретении t-PA-производное ДНК-последовательность, зависит от используемых позднее для экспрессии клеток-хозяев. Пригодные плазмиды, а также минимальные требования, которые предъявляются к такого рода плазмиде (например, источник репликации, необходимый сайт рестрикции) известны любому специалисту. Вместо плазмиды также можно применять космиду, репликативные двунитевые формы фагов ( λ , М13) и другие известные специалисту векторы. Методы направленного мутагенеза (сайт-специфического мутагенеза) описаны Morinaga и др., Biotechnology 21 (1984), 634.
Предпочтительно для получения предлагаемого в изобретении t-PA-производного в качестве клеток-хозяев применяют прокариотические клетки. При этом образующиеся так называемые "внутриклеточные тельца" (нерастворимые протеиновые агрегаты) сначала отделяют от растворимых частиц клеток, солюбилизируют при восстанавливающих условиях путем обработки гуанидин-гидрохлоридом, затем обрабатывают GSSG и, наконец, ренатурируют t-PA-производное путем добавки L-аргинина и GSH. Точные методики активирования t-PA из "внутриклеточных телец" описаны, например, в европейских патентах А-О 219 874 и А-О 241 022. Однако можно использовать также любые другие способы получения активного протеина из "внутриклеточных телец".
В случае предлагаемого в изобретении способа для очистки производного t-PA K2P предпочтительно работают в присутствии L-аргинина, обычно при концентрации 10 - 1000 ММоль/л.
Предлагаемое в изобретении отделение от чужеродного протеина путем аффинной хроматографии осуществляется в предпочтительном варианте осуществления изобретения через ET1-адсорбирующую колонку (ET1 = эритрин-трипсин-ингибитор). При этом ET1 фиксируется на материале носителя, например, на сефарозе. Очистка через ET1-адсорбирующую колонку имеет то преимущество, что материал ET1-адсорбирующей колонки может загружаться непосредственно из концентрированной смеси ренатурирования даже в присутствии таких высоких концентраций аргинина, как 0,8 моль/л. Благодаря этому избегают агрегации K2P, которая имеет место при низких концентрациях аргинина (ниже 10 ммоль/л). Желательно осуществлять очистку K2P через ET1-адсорбирующую колонку в присутствии 0,6 - 0,8 моль/л аргинина. Предпочтительно, чтобы содержащий K2P раствор имел pH выше 7 и особенно предпочтительно 7,5 - 8,6.
Элюирование продукта с ET1-колонки осуществляется путем снижения pH, как в присутствии, так и в отсутствии аргинина, при условиях, которые обеспечивают хорошую растворимость K2P. Предпочтительное значение pH при элюировании лежит в кислой области, особенно предпочтительно в области 3 - 5,5.
Полученный таким образом K2P обладает удельной t-PA-активностью 550.000±200.000 IU/мг при чистоте свыше 95%, предпочтительно выше 99%.
Согласно изобретению получается t-PA-производное, которое обладает отчетливо более продолжительным временем сохранения в плазме за счет пониженной скорости клиренса. Названное производное, однако при этом не теряет никаких свойств, делающих его пригодным для использования в качестве лекарственного средства. Например, необходимые для тромболитической терапии с помощью K2P дозы снижается по крайней мере, на четверть в сравнении с дозой нативного t-PA. В случае применения одинаковых доз K2P и нативного t-PA в первом случае система свертывания оказывается менее затронутой, и время кровотечения в отличие от наблюдаемого с нативным t-PA, удлиняется незначительно. Таким образом, вероятность осложнения за счет кровотечения при терапии с помощью K2P уменьшается.
Указанные свойства предлагаемого t-PA-производного делают его особенно пригодным для использования в фармкомпозиции, что составляет еще один предмет изобретения. При применении предлагаемого в изобретении t-PA-производного в виде лекарственного препарата для достижения одинаковой с нативным t-PA эффективности требуется значительно более низкая доза введения.
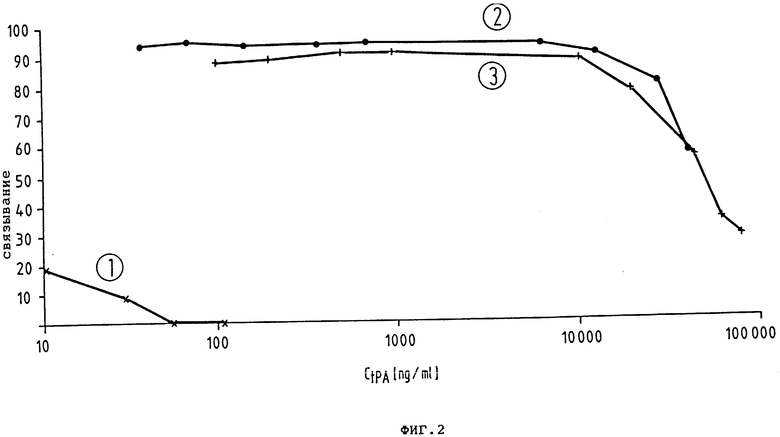
На чертежах показано следующее: фиг. 1 - схематически получение плазмиды pA27.3; фиг. 2 - сравнение связывания фибрина предлагаемым в изобретении t-PA-производным (кривая 1) и полученным в CHO-клетках нативным t-PA-производным (двунитевым t-PA, расщепленным в физиологическом сайте расщепления Arg 275-1le276 [кривая 2) и однонитевым t-PA (кривая 3)];
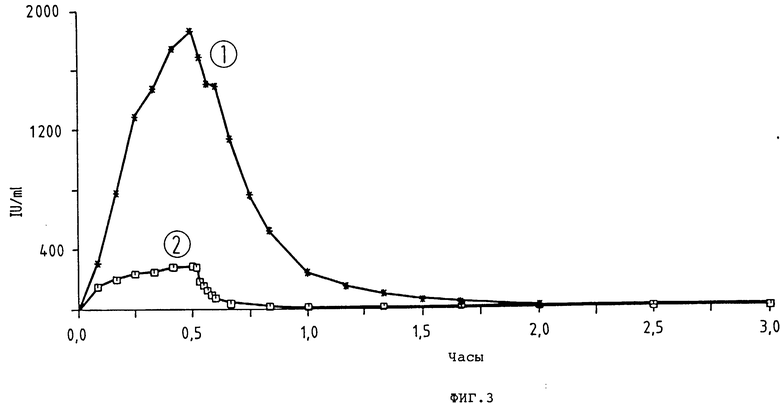
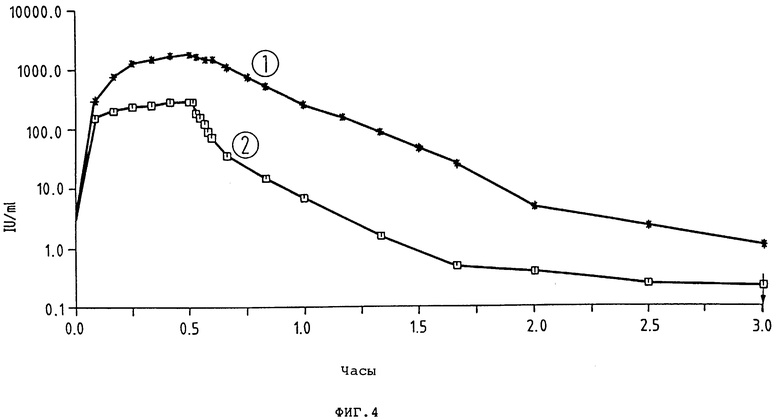
фиг. 3 и 4 - диаграммы фармакокинетики t-PA-активности предлагаемого согласно изобретению t-PA-производного и выпускаемого в продажу t-PA-препарата (ActilyseR); кривая 1: K2P, доза 200 000 ед/кг = 0,25 мг/кг; внутривенное вливание через 30 мин; число исследованных животных (кролики) = 4; кривая 2: ActilyseR доза 200 000 ед/кг; внутривенное вливание через 30 мин; число исследованных животных (кролики) = 6;
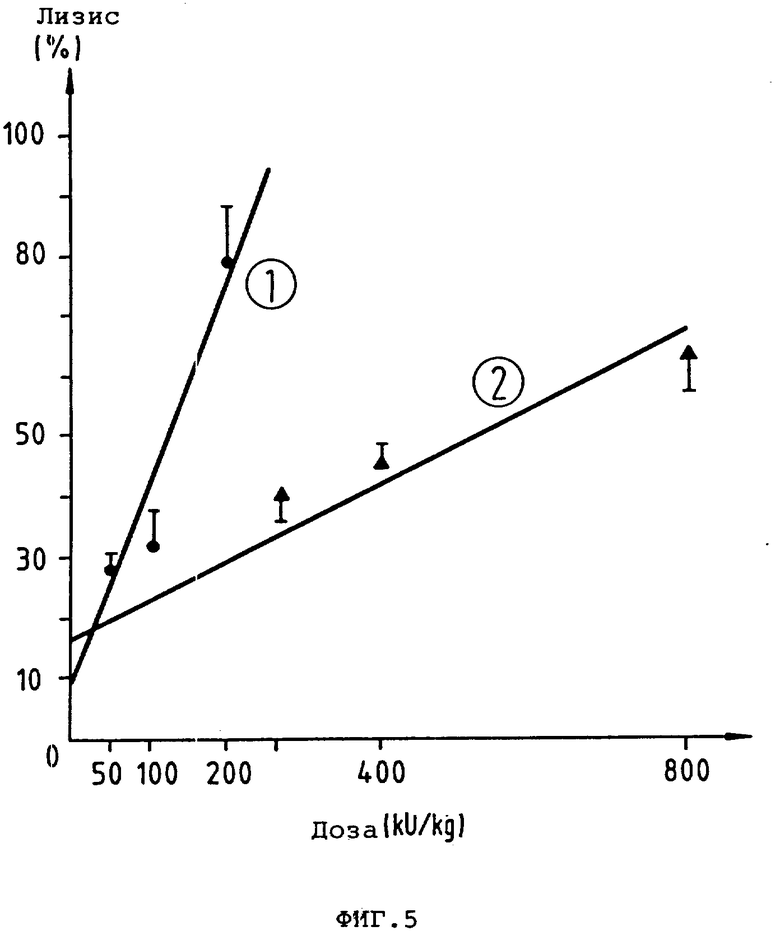
фиг. 5 - сравнительные кривые действия доз (для кроликов) на тромболиз t-PA-производных (показано среднее значение +SEM: 1kU = 1000 1uU; кривая 1: K2P; кривая 2: ActilyseR);
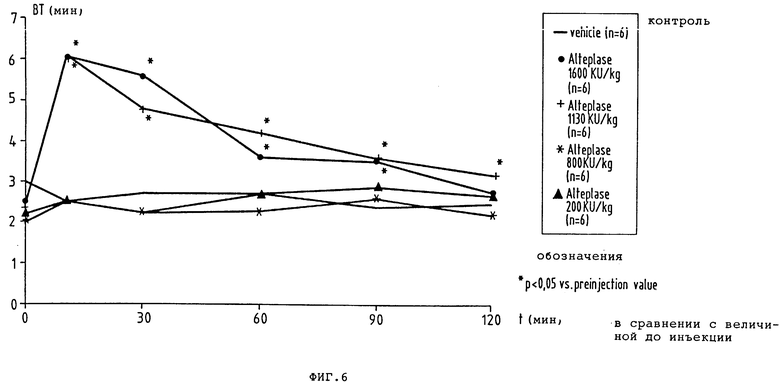
фиг. 6 - временная зависимость Simplate времени кровотечения (ВТ) до и после разовой внутривенной инъекции плацебо или повышающихся доз ActilyseR находящимся под наркозом собакам;
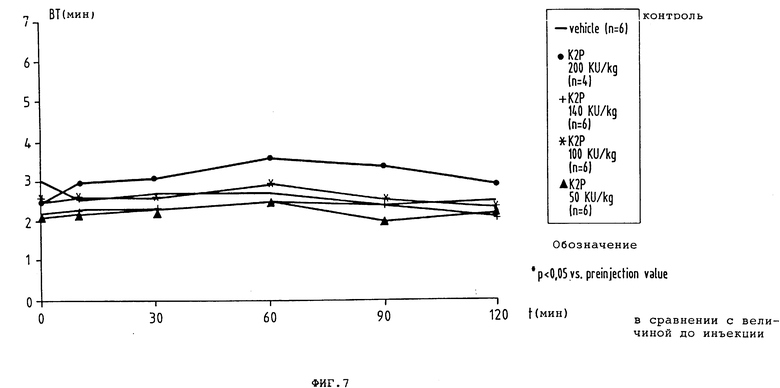
фиг. 7 - временная зависимость Simplate времени кровотечения (ВТ) до и после разовой внутривенной инъекции плацебо или повышающихся доз K2P находящимся под наркозом собакам.
Пример 1. Конструкция плазмиды pA27.3. Исходная плазмида pREM7685, описанная в европейском патенте А-О 242 836, содержит следующие элементы: tac-промотор, область lac-оператора с ATG-стартовым кодоном, участок кодирования t-PA-производного FK2P, терминатор транскрипции из pKK223-3, ген Actilyse® -лактамазы, ген регистентности к канамицину и сайт начала репликации из плазмиды рАСУС177. Последовательность t-PA-производного FK2P образована из нуклеотидов 180-336 (F-домен), 715-1809 (K2-домен, протеаза, меньшая доля 3'-UT) и ATG-стартового кодона. Положения нуклеотидов даны согласно последовательности, описанной в Nature, 301 (1983) 214 - 221.
Для делеции F-доменов из FK2P-конструкции плазмиды pREM7685 используют метод Morinaga и сотр. Biotechnology, 21 (1984) 634. Для образования гетеродуплекса из pREM7685 изолируют два фрагмента. Фрагмент A: pREM7685 расщепляют с помощью рестрикционного фермента EcoR1, продукты расщепления разделяют с помощью гель-электрофореза и больший фрагмент EcoR1 элюируют из геля. Фрагмент B: плазмиду pREM7685 линеаризуют с помощью рестрикционного фермента Xho I, линеаризованную плазмиду также получают препаративно гель-электрофорезом. Для мутагенеза синтезируют следующий олигонуклеотид:
5 TG TCT TAC CAA GGA AAC AGT GA 3'
Для образования гетеродуплекса фрагмента A, фрагмент B (по 450 f моль) и олигонуклеотид (75 p моль) смешивают и в присутствии 50 ммоль/л NaCl 10 ммоль/л Трис/HCl, pH 7,5 и 10 ммоль/л MgSO4 инкубируют 3 мин при 100oC и переносят на лед. Ренатурация ДНК происходит за 30 мин при 60oC. Для репаративного синтеза к гетеродуплексу добавляют дезоксинуклеотидтрифосфаты (0,25 ммоль/л, ATP (1 ммоль/л), NaCl (100 ммоль/л), Трис-HCl, pH 7,5 (6,5 ммоль/л), MgCl2 (8 ммоль/л) Actilyse® -меркапотоэтанол (1 ммоль/л), фрагмент Кленова ДНК-полимеразы из E. coli (0,125 ед/мкл смеси) и Т4-лигазу (0,1 ед/мкл смеси). Репаративный синтез осуществляют в течение 4 ч при 16oC. Затем смесью трансформируют клетки E.coli (R М82 DSM 3689) с помощью 1ac 1q-плазмиды и трансформанты путем добавки к питательной среде 25 мкг/мл нанамицина.
С помощью способа гибридизации колоний при применении вышеописанного олигонуклеотида в качестве зонда можно отбирать клоны, несущие плазмиду pA27.3, которая кодирует предлагаемое в изобретении t-PA-производное К2Р. Эта плазмида отличается от исходной плазмиды pREM7685, также отсутствием сайта PstI и сайта SspI. Эти оба места разреза содержатся в области исходной плазмиды, которая кодирует F-домен. Конструкция плазмиды pA27.3 схематически представлена на фиг. 1.
Пример 2. Получение активного t-PA-производного К2Р из E.coli:
а) Лизис клеток и получение внутриклеточных телец IB'S
1,6 кг влажной клеточной массы E.coli DSM 3689, трансформированная с помощью плазмиды pA27.3 суспендируют в 10 л 0,1 моль/л Трис-HCl, 20 ммоль/л ЭДТА, pH 6,5, при 4oC. К этой смеси добавляют 2,5 г лизоцима и инкубируют 30 мин при 4oC; затем клетки переводят в удобную для переработки форму путем диспергирования при высоком давлении. К полученному раствору примешивают 5 л 0,1 ммоль/л Трис-HCl, 20 ммоль/л ЭДТА, 6% Тритона Х100 и 1,5 ммоль/л NaCl, pH= 6,5, и инкубируют 30 мин при 4oC. Затем осуществляют отделение нерастворимых составных частей (IB's) путем центрифугирования с помощью центрифуги Padberg.
Осадок после центрифугирования суспендируют в 10 л 0,1 ммоль/л Трис-HCl, 20 ммоль/л ЭДТА, pH 6,5, инкубируют 30 мин при 4oC и IB-пррепарат отделяют путем последующего центрифугирования.
б) Солюбилизация 1B
100 г 1B (мокрый вес) суспендируют в 450 мл 0,1 ммоль/л Трис-HCl, 6 ммоль/л гуанидин-гидрохлорида, 0,2 ммоль/л ДТЕ, 1 ммоль/л ЭДТА, pH 8,6 и перемешивают при 25oC 2,5 ч.
После установления pH 3 с помощью HCl (25%) осуществляют диализ против 10 ммоль/л HCl (3 • 50 л, 24 ч, 4oC).
в) Дериватизация
Вводят гуанидин-гидрохлорид (твердый) так, чтобы в последнем разбавлении вышеуказанного диализа 10 ммоль/л HCl концентрация гуанидин-гидрохлорида составляла 6 ммоль/л.
Реакционную смесь инкубируют при 25oC в течение 1,5 ч, затем вводят окисленный глютатион GSSG до 0,1 моль/л и Трис-HCl до 0,05 ммоль/л, а pH-значение доводят по титру до pH 9,3 с помощью 5 ммоль/л NaOH. Смесь перемешивают в течение 3,5 ч при 25oC.
После установления pH 3 с помощью HCl (25%) осуществляют диализ против 10 ммоль/л HCl (3 • 100 л и 48 ч, 4oC). После диализа реакционную смесь центрифугируют и прозрачную надосадочную жидкость обрабатывают далее.
г) Ренатурация
Реакционный сосуд емкостью 10 л заполняют смесью 0,1 ммоль/л Трис-HCl, 0:8 ммоль/л L-аргинина, 2 ммоль/л GSH (восстановленная форма), 1 ммоль/л ЭДТА, pH 8,5. В зависимости от условий ренатурирование проводят при 20oC путем трехкратной добавки 100 мл указанного выше смешанного дисульфида в отрезок времени 24 ч.
После ренатурирования получают материал с удельной активностью 1500 - 10000 1U (мг) определение (пример 46). Единица 1U - единица активности по определению WHO, Национального Института по биологическим стандартам и контролю.
д) Концентрирование ренатурированной реакционной смеси
Ренатурированную смесь при необходимости можно концентрировать через гемодиализатор.
Пример 3. Очистка К2Р из E.coli.
1. Очистка К2Р из E.cjli путем афинной хроматографии на ET1 - сефарозе после предварительного концентрирования
а) Элюирование с помощью лимонной кислоты
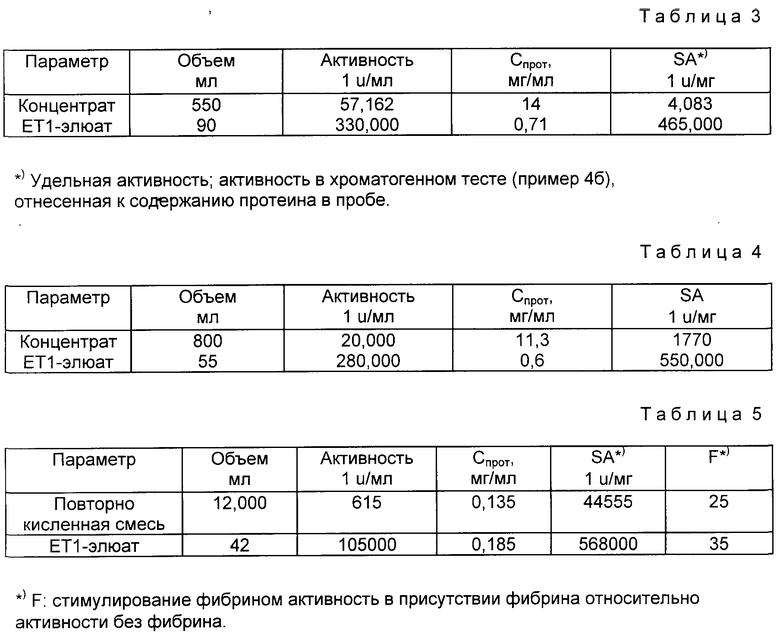
Смесь после ренатурирования концентрируют через Memo, диализатор (Asahi AM 300) 1: 23 и дополняют с помощью 0,5 ммоль/л. Уравновешенную смесью 0,1 ммоль/л Трис-HCl, pH 7,5, 0,8 ммоль/л аргинина, 0,5 ммоль/л NaCl колонку с ET1 (эритрин-трипсин-ингибитор)-сефарозой (объем = 50 мл) загружают 550 мл концентрата подвергнутой повторному окислению смеси (10 колоночных объектов/ч, 10 SV/ч) и промывают уравновешивающим буфером до тех пор, пока абсорбция при 280 нм не достигнет значения буфера. Элюирование связанного материала осуществляют с помощью 20 ммоль/л лимонной кислоты, pH 3,2 (см. табл. 3).
б) Элюирование с помощью 0,3 ммоль/л аргинина, pH 4,5
Ренатурированную реакционную смесь концентрируют, как описано в примере 3.1. а). Уравновешенную с помощью смеси 0,1 ммоль/л Трис-HCl, pH 7,5, 0,8 ммоль/л аргинина, 0,5 ммоль/л NaCl колонку с ET1-сефарозой (25 мл) нагружают 800 мл концентрат (12 BV/ч) и промывают уравновешивающим буфером до тех пор, пока экстинкция элюата при 280 нм не достигает значения экстинкции буфера. Связанный материал элюируют с помощью 0,3 ммоль/л аргинина, pH 4,5, (см. табл. 4).
2. Очистка К2Р из E.coli путем афинной хроматографии на ET1-сефарозе без предварительного концентрирования.
Уравновешенную с помощью смеси 0,1 ммоль/л Трис-HCl, pH 7,5, 0,8 ммоль/л аргинина, 0,5 ммоль/л NaCl колонку с ET1-сефарозой (объем = 10 мл) загружают 12 л повторно окисленной смеси и промывают уравновешивающим буфером до тех пор, пока экстинкция элюата при 280 нм не достигнет значения экстинкции буфера. Элюирование связанного материала осуществляют с помощью 0,8 ммоль/л аргинина, pH 5 (см табл. 5).
Пример 4. Экспрессия К2Р-гена при использовании других векторов. Последовательность, колирующую К2Р, клонируют в космиде pHC79. Космиду, как описано Коллинзом, Дж., и др. в PNAS USA 75 (1978) 424, упаковывают в Actilyse® - фаги, трансдуцируют в E.coli ED 8654 и экспрессируют.
Протеин очищают, как описано в примере 3.
Аналогично клонируют ген K2P в векторах M13+, MP18, M13-, MP19, pUC18, pUC19, pBR322, pBR328 и pSP64.
Молекулярный вес, аминокислотную последовательность и активность определяли, как описано ниже.
Пример 5. Характеристика очищенного K2P из E.coli
а) Протеин-химическая характеристика - SDS - PAGE и жидкостная хроматография высокого давления с обращенной фазой.
Гомогенность очищенного с помощью афинной хроматографии с ETI-сефарозой материала показана при помощи SDS - PAGE и жидкостной хроматографии высокого давления с обращенной фазой (R P-HPLC). Из относительной длины пробега для K2P из прокариотов рассчитан молекулярный вес 38,550 + 2,000 Да. Денситометрическая оценка дает чистоту препарата выше 95%.
RP-HPLC основана на различном взаимодействии протеинов с гидрофобной матрицей. Это семейство используется в качестве аналитического метода для квалификации степени чистоты.
Анализ очищенного K2P из E.cjli осуществляют через разделительную колонку с Нуклеозилом 300 (Kаduer) с помощью градиента трифторуксусная кислота - ацетонитрил (буфер А: 1,2 мл трифторуксусной кислоты в 1000 мл воды; буфер B: 300 мл воды, 700 мл ацетонитрила, 1 мл трифторуксусной кислоты; 0-100%). Интегрирование хроматографического анализа дает чистоту выше 95%.
-N-терминальная последовательность аминокислот
N-терминальная последовательность аминокислот определяется с помощью AB1 470 - секвенатора со стандартной программой и линейным PTH-детектированием. Найденная последовательность ST-Y2-Q3-G4-N5-S6-D7-C8-Y9 совпадает с ожидаемой, происходящей из ДНК-последовательности.
б) определение активности
"Ин витро" активность K2P из E.coli определяется в соответствии с тестом, описанном в "Zeitschrift f Actilyse® r die gesamte innere Medizin" (ZGIMAL), 42, (17) 478 - 486 (1987). Удельная активность составляет 550000/мг±200,001/мг. Стимулируемость K2P из E.coli BrCN-фрагментами фибриногена (активность в присутствии фибриногеновых фрагментов, отнесенная к активности без фибриногеновых фрагментов) в этой тест-системе составляет не более 25.
в) Связывание с фибрином in vitro
"Ин витро"-связывание K2P из E.coli с фибрином определяют как описано (Higgins, D.L. und Vehar, G.A. (1987), Biochem, 26, 7786-7791).
На фиг. 2 показано, что K2P из E.coli в противоположность полному t-PA на CHO или β t-PA из E.coli не обладает сколько-нибудь заметной способностью связывать фибрин.
Пример 6. Повышение уровня экспрессии.
Для увеличения выхода целевого продукта кодирующую K2P-ген последовательность переклонируют в плазмиду с более высоким числом копий. Для этой цели используют плазмиду pePa 126.1, описанную в патенте ФРГ 3838378.0. Эта плазмида состоит из вектора pKK223-3 и кодирующей последовательности для t-PA, как описано в европейском патенте A-O 242 835.
В эту плазмиду прежде всего интегрируют последовательность fd-терминатора. Для этой цели плазмиду pePA 126.1 линеаризируют с помощью рестрикционного фермента Hind III. Расщепленную таким образом плазмиду разделяют с помощью гель-электрофореза и получают препаративно. Плазмиду pLBU1 (Beck и др., (1978), Nucl. Acids Res. 5, 4495-4503; Gentz и др. (1981), PNAS 78(8): 4963) подвергают рестрикции с помощью Hind III и препаративно получают фрагмент Hind III величиной примерно 360 п.о., который содержит fd-терминатор (путем гель-электрофореза и гель-элюирования). Линеаризированную плазмиду pePa 126.1 и Hind III-фрагмент из pLBU1 длиной 360 п.о. лигируют. Смесь после лигирования контрасформируют с описанной в заявке на патент ФРГ 3838378.0 плазмидой pUBS 500 в E.coli DSM 2102. Из клонов выбирают те, которые содержат желательную плазмиду pePA 126 fd, которая отличается от исходной плазмиды pePA 126.1 тем, что она содержит второй сайт разрезания Hind III.
Из плазмиды pePA 126 fd получают два фрагмента:
BamHI/PvuI фрагмент величиной 3,4 т.п.о. и PvuI/XmaI-фрагмент, размером примерно 1,3 т.п.о. Оба указанных фрагмента лигируют с 1,3 т.п.о. BamHI/XmaI фрагментом из плазмиды pA27.3 и трансформируют E.coli вместе с плазмидой pVBS 500. Результирующая плазмида название pA 27 fd.
Пример 7. Приготовление фармацевтической композиции.
Получаемое из E.coli t-PA- производное K2P после описанной выше очистки может быть использовано для тромболитической терапии, предпочтительно в виде фармацевтической композиции.
Соответствующие фармацевтические препараты в частности могут иметь следующий состав:
1. 0,2-0,6 моль/л аргинин/HCl
0,01-0,06 моль/л фосфата натрия, pH 7,2 и
0,05-0,2 мас.% твин 80
2. 0,2-0,6 моль/л аргинин/H3P04, pH 7,2
0,05-02 моль/л твин 80
3. 0,1-0,2 моль/л фосфата натрия или калия, pH 6,0
10 моль/л транексамовой кислоты,
50 мг/мл сахарозы или трехалозы и
0,01-0,2 мас.% твин 80 или твин 20.
Для K2P из примера 3 концентрация протеина во всех указанных выше составах 4 - 10 мг/мл (амидолитическая активность 2,4-4,9 М/мл).
Пример 8. Фармакологические данные получаемого в прокариотах K2P.
1. Фармакокинетика K2P на кроликах
K2P на белых новозеландских кроликах сравнивают с Actilyseβ по их фармакокинетическим свойствам. Оба фибринолитика вводят в дозе 200 000 IU/ кг KG в течение 30 мин. Пробы плазмы получают в определенные моменты времени до, во время и после вливания. t-PA-активность измеряется с помощью спектрофотометрического теста по J.H.Verheijen и др. (Thromb. Haemostas. 48, 266) (1982)) и модифицированному по H. Lill (Z.ges Inn. Med. 42, 478 (187)).
Для расчета фармакокинетических параметров используют расчетную программу нелинейной регрессии, модифицированную по H.J. Huang (Aero-Astronautica-Report 64. Rice University, 1 - 30, 1969). Параметры рассчитывают индивидуально с помощью биэкспоненциальной фармакокинетической модели.
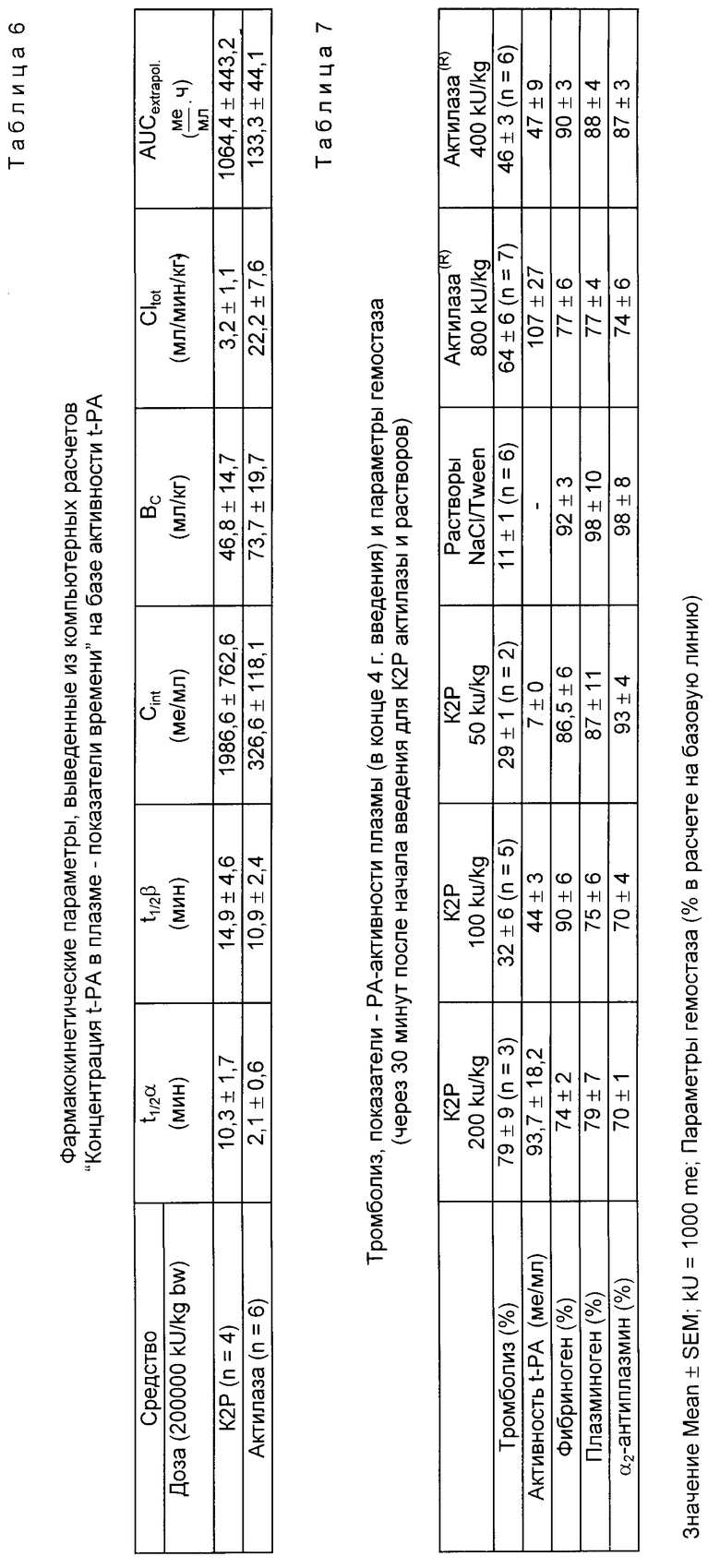
K2P показывает в пять раз более длительное время полураспада (t1/2 λ = 10,3 мин, уменьшение концентрации в плазме), чем Actilyse® (t-PA-препарат фирмы Thomal) (табл. 6, рис. 5 и 6). По окончании вливания (спустя 30 мин) в случае K2P-Pro концентрация в плазме активности t-PA (Cинф./равна 1966 1 U/мл и, таким образом, в шесть раз выше, чем в случае Actilyse α . Распределение объема (Vc) составляет для K2P 46,8 мл/кг по сравнению с 73,7 мл/кг для Actilyse. Клиренс всей плазмы (C1общ) по сравнению с Actilyse Actilyse® (C1общ= 22,2 мл/мин/кг) в случае K2P-Pro снижен до 3,2 мл/мин/кг. При применении фибронолитика особенно интересен показатель AUC, так как он отражает преимущественную концентрацию в плазме. K2P показывает AUC в 8 раз выше (1064 1/мл•ч), чем Actilyse Actilyse® (133,3 1U/мл•ч).
В целом K2P по сравнению с Actilyse® (это в настоящее время единственный продажный рекомбинантный t-PA-протеин) при равной дозе показывает улучшенный в 5 - 8 раз фармакокинетический профиль.
2. Фармакодинамика K2P на кроликах
Для испытания тромболитической эффективности используют разработанную D. Collen и др. модель на кроликах по тромбозу яремной вены (J.Clin Invtst. 71,368, 1983). K2P и Actilyse Actilyse® исследуют в трех, смотря по обстоятельствам, дозах. Фибринолитики вливают в течение 4 ч и затем определяют скорость тромболиза (табл. 7, рис. 5).
Рассчитанная с помощью линейной степени регрессии доза для скорости 50%-ного тромболиза (ЭД50) составляет для K2P 124000 1U/ кг KG и для Actilyse 520,000 1U/кг KG. K2Pпоказывает, таким образом, в 4 раза более высокое тромболитическое действие, чем Actilyse®. K2P обеспечивает в зависимости от дозы также концентрирование активности t-Pa в плазме которое при четырехкратно сниженной дозе сравнимо с Actilyse®. Сравнимая по тромболитическому действию с 800 kU Actilyse® KG доза в 200 kU K2P/ кг KG приводит к незначительным изменениям параметров получения фибриногена, плазминогена и α2 -антиплазмина, которые при этом не отличаются от воздействий дозы 800 к U Actilyse /кг KG.
K2P представляет собой t-PA-Мутеин, который на модели тромбоза яремной вены кролика при четырехчасовом вливании в дозе, составляющей 1/4 дозы Actilyse®, проявляет такое же тромболитическое действие, как и Actilyse® K2P не отличается при пониженной дозе от Actilyse® по воздействиям на систему свертывания и по концентрации в плазме t-PA -активности.
Пример 9. Фармакологические свойства K2P из E.coli на используемой в качестве модели собаке с тромбозом коронарной артерии
Для того, чтобы изучить тромболитические действия K2P из E.coli на артериальных тромбах экспериментальной моделью было выбрано животное с острым инфарктом миокарда. В качестве одного из таких животных исследовали собаку. Метод образования тромба коронарной артерии представляет собой модификацию способа Romson и др. (Thromb. Res. 17, 841 (1980). При открытой грудной клетке у находящихся под наркозом и при искусственном дыхании собак раздражают поверхность интимы окружающей ветви A.coronaria sinistra (=leftcirtum- flex coronary artery = LCX) электрическим импульсом (150 μ ) благодаря чему возникает тромбоз.
При этом проксимально коронарный тромбоз получали при помощи электромагнитной плавающей измерительной головки, обеспечивающей возможность измерения реперфузии (восстановление кровотока).
Для установления необходимых доз находящимся под действием гепарина собакам в течение 1 мин вводили путем однократно внутривенной инъекции ВМ 06.022 с эукаристическим t-Pa(Alteplase, Actilyse®, Dr. Karl Thomae Гмбх. Biberoch, ФРГ) в виде четырех различных доз (по 6 животных на каждую дозу). До и в определенные моменты времени после инъекции отбирали пробы плазмы, чтобы определить концентрацию в плазме t-PA-активности, фибриногена, плазминогена и α2 -антиплазмина, а также, чтобы определить число тромбоцитов во всей крови. Фибриноген измеряют коагулометрически по Клаусу (Acta haemat. 17, 237, 1957), плазмиген и α2 -антиплазмин измеряют, как описано у Collen и др. (J. Clin Invest 71,368, (1983) спектрометрически.
Далее Simplate время кровотечения измеряли на задней ноге собаки с помощью защелки (Simplate®I, Organon I, Organon Teknika, Eppelheim, ФРГ) при 40 мм Hg (J. Surg. Res. 27, 244, 1979). Статическое сравнение измеренных величин после инъекции с контрольным значением до инъекции осуществляют с помощью теста Wilcoxon для каждой пары.
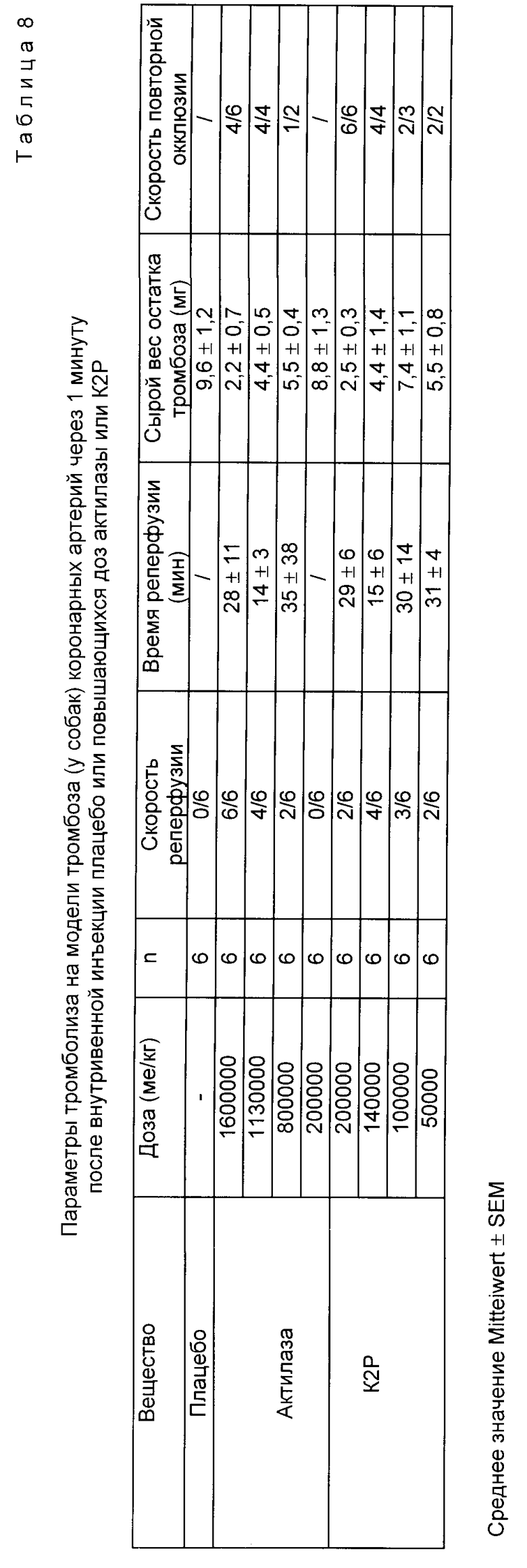
Для того, чтобы описать тромболитический эффект в каждой "доза-группе" указывается число животных с восстановленным кровотоком (скорость реперфузии), а также время до начала восстановления (время реперфузии). Далее определяют сырой вес еще имеющегося через 2 ч после начала инъекции остаточного тромба и определяют количество животных с новой закупоркой после реперфузии (= скорость повторной окклюзии). С помощью полулогарифмического регрессионного анализа действия доз (отношение скоростей реперфузии) для каждого вещества рассчитывается эффективная доза для 50%-ной скорости восстановления кровотока (=ЭД50). Статистическое сравнение веса остаточного тромба осуществляют с помощью теста Wilcoxon Mann-Whithey.
Концентрация в плазе t-PA-активности измеряется с помощью спектрофотометрического теста по Verhejen и др. (Thromb Haemost. 48, 266, 1982), модифицированного по Lill (Z. gesamte Inn Med. 42, 478 (1987). Для расчета фармакокинетических параметров используется расчетная программа нелинейной регрессии, модифицированная по HY. Huang (Aero-Astronauties Report 64 Rice Univercity, США, 1 - 30, 1969). Параметры рассчитываются индивидуально после снижения свойственного организму базального уровня t-PA-активности нижеследующих измеренных значений с помощью биэкспоненциальной фармакокинетической модели.
При этом получаются следующие результаты:
1. Фармаколинамика на собаке
K2P после внутривенной инъекции приводит к зависящей от дозы скорости реперфузии. Максимальный эффект (скорость = 100%) достигается после инъекции 200 кU/кг KG. Доза для 100%-ной реперфузии в случае Actilyse® составляет 1600 кU/кг KG. Сравнение значение ЭД50 для K2P Actilyse® показывает, что первое (83 кU/кг KG) примерно в 11,5 раза ниже, чем второе (961 кU/кг KG). Введение плацебо не дает восстановления кровотока. Вес остаточного тромба с введенным плацебо составляет 9,6 ± 1,6 кг (среднее значение ± SEM; как K2P, так и также Actilyse® приводят с увеличением доз к статистически значительному снижению веса остаточного тромба по сравнению с плацебо-контролем. Реперфузия происходит в случае обоих фибринолитиков в среднем у всех животных спустя 25,9 ± 3,5 мин (K2P) и спустя 24,2 ± 6,2 мин (Actilyse Actilyse®).
Большинство обработанных K2P или Actilyse® собак повторно окклюдируют после восстановления (табл.8).
2. Фармакокинетика на собаке
После внутривенной инъекции 200 кU/кг K2P или Actilyse® на наркотизированной собаке показано, что быстрая фаза уменьшения концентрации в плазме, выраженная в виде t 1/2 α , в случае K2P (7,2 ± 1,1 мин) в 4:5 раза длиннее, чем в случае Actilyse® (1,6 ± 0,2 мин) (табл.9). Найденная непосредственно после окончания инъекции концентрация в плазме K2P примерно вдвое выше, чем в случае Actilyse®. Удаление K2P из плазмы (клиренс плазмы = C1общ) осуществляется в девять раз медленнее, чем в случае Actilyse®. Соответственно площадь под кривой концентрации в плазме K2P примерно в 9,5 раз больше, чем таковая в Actilyse®.
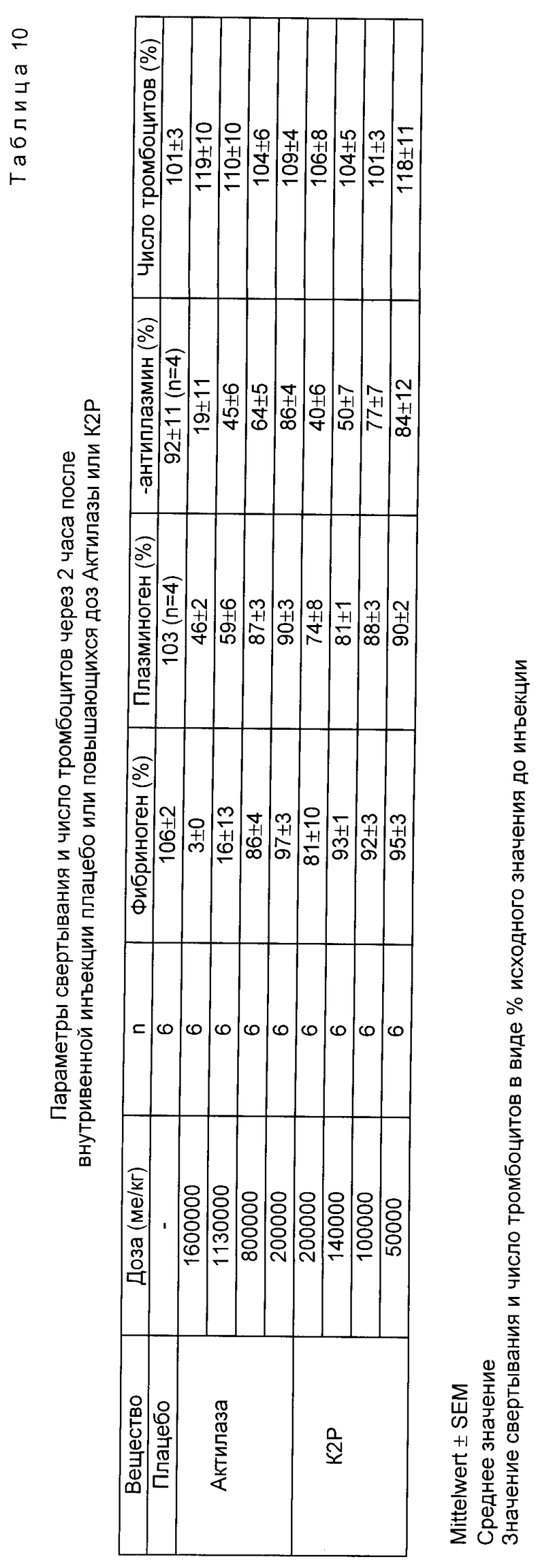
3. Специфичность к фибрину на собаке
Спустя 2 ч после инъекции K2P находят зависящее от дозы небольшое снижение остаточной концентрации фибриногена (до 81 ± 10% в случае самой высокой дозы 200 кU/кг KG). В противоположность этому концентрация фибриногена после введения самой высокой дозы  (1600 кU/кг KG почти полностью снижена, до 3 ± 0% (табл. 10). Если осуществляют полулогарифмический регрессионный анализ зависимости "доза - побочное действие" (соотношение снижения фибрина) и определяют ЭД50 тромболитического действия в отношении остаточной концентрации фибриногена, то для эквипотентных доз в случае K2P остаточное содержание фибриногена получается 92,5% (по сравнению с 38,6% в случае Actilyse®). Для плазминогена и α2 -антиплазмина находят также зависящее от дозы снижение остаточного содержания спустя 2 ч после инъекции, которое в случае Actilyse® более ярко выражено, чем в случае K2P. Лишь концентрация тромбоцитов в случае обоих веществ одинаково не затрагивается.
(1600 кU/кг KG почти полностью снижена, до 3 ± 0% (табл. 10). Если осуществляют полулогарифмический регрессионный анализ зависимости "доза - побочное действие" (соотношение снижения фибрина) и определяют ЭД50 тромболитического действия в отношении остаточной концентрации фибриногена, то для эквипотентных доз в случае K2P остаточное содержание фибриногена получается 92,5% (по сравнению с 38,6% в случае Actilyse®). Для плазминогена и α2 -антиплазмина находят также зависящее от дозы снижение остаточного содержания спустя 2 ч после инъекции, которое в случае Actilyse® более ярко выражено, чем в случае K2P. Лишь концентрация тромбоцитов в случае обоих веществ одинаково не затрагивается.
4. Влияние на время кровотечения у собаки
Внутривенная инъекция K2P не дает никакого статически значительного удлинения времени кровотечения по сравнению с контрольным значением перед инъекцией во всех четырех различных дозах. В противоположность этому Actilyse® в дозах 1130 и 1600 кU/кг KG статистически значительно удлиняет время кровотечения (фиг.6 и 7).
5. Общая оценка
На описанной модели тромбоза коронарных артерий K2P проявляет себя как тромболитик, который после внутривенной инъекции порции может достигать 100-ной скорости восстановления кровотока не влияя сильно на концентрацию фибриногена и без заметного удлинения времени кровотечения. По сравнению с Actilyse® K2P в тромболитической потенции после внутривенной инъекции оказался отчетливо превосходящим (фактор 11,5). Исследование фармакокинетического профиля K2P, также показывает, что по сравнению с Actilyse® клиренс K2P, как выражение для более продолжительного удаления из плазмы, в девять раз длиннее.


Использование: биотехнология, генетическая инженерия. Сущность изобретения: получен полипептид К2Р со свойствами активатора плазминогена путем трансформации бактериального штамма рекомбинантной плазмидной ДНК рА27.3, включающей фрагмент ДНК, кодирующий указанный полипептид и имеющий следующие свойства: фибринолитическая активность 550000 ± 200000 Ед/мг; молекулярный вес 38500 ± 2000 Да; увеличенный период полураспада по сравнению с природным аналогом; фибринзависимая стимуляция; тромболитическая активность; растворимость в растворе аргинина в концентрации 10 - 1000 ммоль/л при чистоте продукта 95%; получена также фармакологическая композиция, содержащая указанный полипептид в эффективном количестве и фармакологически приемлемой носитель. 4 с.п. ф-лы, 7 ил., 10 табл.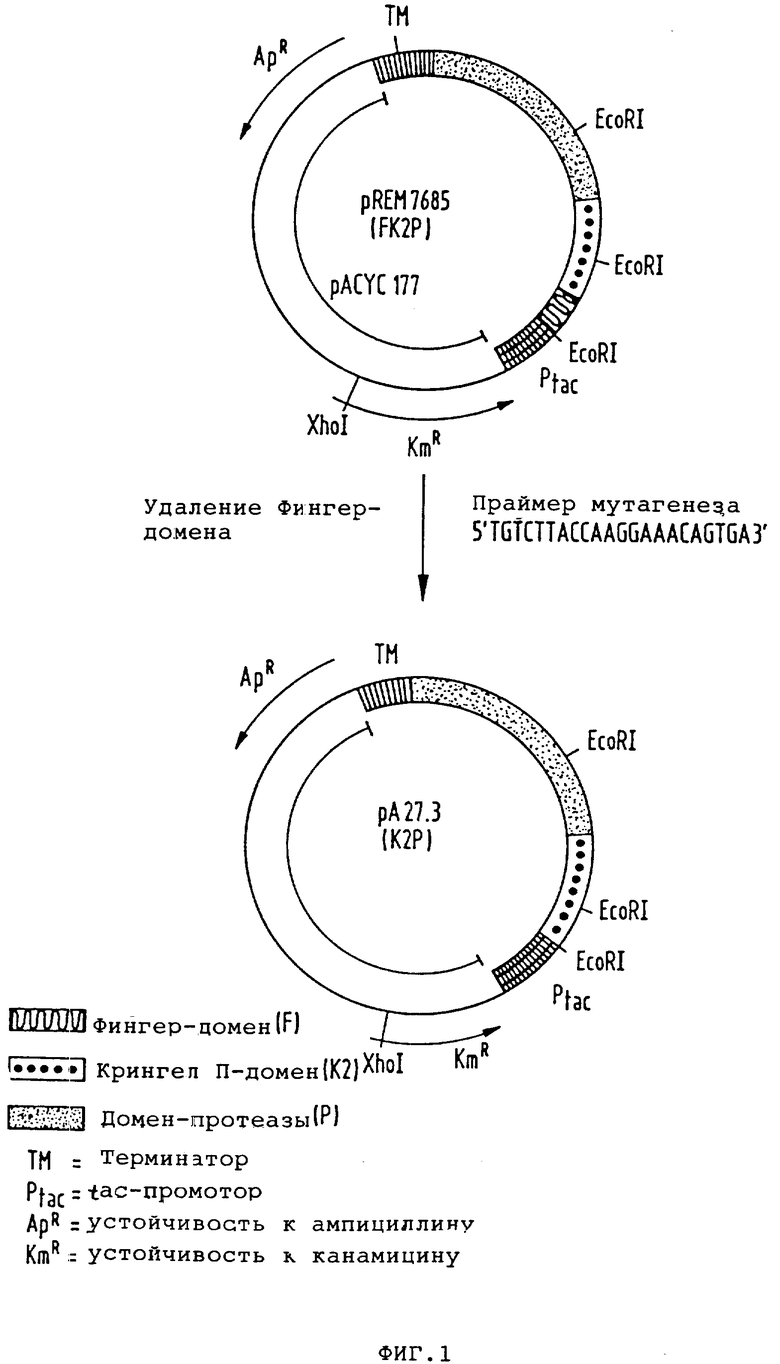
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| EP, А0302456, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
