Изобретение относится к области биотехнологии, генетической и белковой инженерии, т.е. к технологии получения белков с видоизмененными природными или новыми свойствами.
Более конкретно, настоящее изобретение относится к новому модифицированному активатору плазминогена урокиназного типа (мАПУТ), который обладает улучшенными тромболитическими свойствами.
Изобретение относится также к последовательности ДНК, которая кодирует новый мАПУТ, рекомбинантной плазмиде, содержащей кодирующую мАПУТ последовательность ДНК, штамму Escherichia coli - продуценту нового мАПУТ, способу получения нового мАПУТ и к фармацевтической композиции с тромболитическим действием, содержащей новый мАПУТ.
Природные и модифицированные активаторы плазминогена (АП) применяются в качестве тромболитических средств для лечения заболеваний, обусловленных закупоркой сосудов фибрином, таких как инфаркт миокарда, легочная тромбоэмболия, тромбозы вен, тромболитические поражения глаз, в частности гемофтальм, гифема и др. Активаторы плазминогена катализируют превращение плазминогена в плазмин, который, в свою очередь, лизирует фибрин. Одним из активаторов плазминогена является урокиназа.
Известно, что урокиназа, выделяемая из мочи, представлена тремя формами: низкомолекулярной (33 кДа), высокомолекулярной (50-55 кДа) двухцепочной и высокомолекулярной одноцепочной (проурокиназа), со значительным преобладанием низкомолекулярной формы. Наибольшую ценность для медицины представляют тромболитические препараты на основе высокомолекулярного предшественника (зимогена) урокиназы - проурокиназы, вследствие ее низкой активности до момента ее активации на тромбе. Проурокиназа синтезируется в клетке рибосомами, ассоциированными с эндоплазматическим ретикулом. Первоначально происходит синтез полипептида-предшественника (так называемой препроурокиназы), от которого в процессе ко-трансляционной секреции отщепляется сигнальный пептид. Секретируемая из клетки форма с отщепленным сигнальным пептидом и представляет собой проурокиназу. Проурокиназа ферментативно практически не активна и активируется, как и большинство сериновых протеаз, путем ферментативного гиролиза. Активаторами проурокиназы в организме обычно являются плазмин, калликреин и катепсин В.
Проурокиназа преимущественно активирует фибрин-связанный плазминоген, имеющий другую конформацию по сравнению с циркулирующим плазминогеном. Использование проурокиназы в качестве тромболитического препарата особенно эффективно по причине ее крайне низкой активности до момента активации непосредственно вблизи тромба.
Проурокиназу принято также называть активатором плазминогена урокиназного типа (АПУТ).
Каталитическая активность проурокиназы и время ее жизни в организме контролируются взаимодействием проурокиназы с ингибитором активатора плазминогена I типа PAI-1.
В организме, кроме участия в процессах тромболизиса, АПУТ выполняет ряд других функций: участвует в воспалительных процессах и активирует пролиферацию клеток, стимулируя тем самым процессы заживления поврежденных тканей и метастазирования опухолей. Участие в этих процессах опосредовано взаимодействием с рядом других белков. Так, процессы пролиферации контролируются взаимодействием N-концевого домена АПУТ с клеточным рецептором.
Для использования АПУТ в качестве тромболитического препарата желательно при сохранении уровня биологической активности снизить ингибирование PAI-1, что может быть достигнуто внесением изменений в один из участков связывания ингибитора, и снизить побочные эффекты, связанные с взаимодействием АПУТ с рецептором, что достигается видоизменением амино-терминального участка АПУТ.
Известны полная аминокислотная и нуклеотидная последовательности препроурокиназы (Holmes W.E. et al., Biotechnology 0: 923-929, 1985; SWISS-PROT: Р00749).
В настоящее время взаимодействие PAI с активатором плазминогена урокиназного типа считается хорошо изученным процессом. Известны трехмерные структуры этих белков: АПУТ (Spraggont G.S et al., Structure 3: 681-688, 1995) и PAI-1 (Mottonen J. et al., Nature 355: 270-273, 1992). Сформулированы модели взаимодействия АПУТ с ингибитором PAI-1 (Xue Y. et al., Structure 6:627-636, 1998).
Из приведенных выше работ следует, что взаимодействие между АПУТ и PAI осуществляется по крайней мере в двух участках: активном центре АПУТ и так называемой ингибиторной петле, представляющей из себя последовательность с 198 по 204 аминокислоту природного АПУТ: ArgArgHisArgGlyGlySer и выступающей из глобулы АПУТ близко к активному центру. По данным Spraggont G.S et al. (Structure 3: 681-688, 1995) эта петля может образовывать лишь 2 водородные связи с окружающими поверхностными аминокислотами, что говорит о малой связи ингибиторной петли с остальной частью АПУТ.
Ввиду изложенного выше представлялось естественным, что делеция ингибиторной петли приведет к снижению чувствительности к PAI, не затронув укладки и ферментативной активности АПУТ. Однако попытка получения модифицированного АПУТ с делетированной петлей (Adams D.S., J Biol Chem 266:13 8476-8482,1991) привела к резкому снижению выхода активного АПУТ.
В патенте США №5728654 по аналогии с данными, полученными на другой сериновой протеазе - тканевом активаторе плазминогена (tPA), теоретически предсказаны модификации в, по крайней мере, одном из положений ингибиторной петли АПУТ, которые по мнению авторов патента способны привести к снижению чувствительности и даже резистентности к PAI-1 за счет уменьшения положительного заряда в области ингибиторной петли протеазы. Однако конкретные модифицированные АПУТ в патенте не получены и не описаны. Прямой перенос результатов, полученных для tPA, на проурокиназу весьма сомнителен, поскольку не учитывает влияние мутационных изменений на всю пространственную структуру фермента. tPA имеет совершенно уникальное строение активного центра, что выделяет его из всех сериновых протеаз. А имеющиеся литературные данные позволяют утверждать, что главным местом связывания PAI-1 является не ингибиторная петля, а активный центр ферментов. Как следствие, tPA по-другому взаимодействует с ингибиторами ферментативной активности.
Таким образом, существенные различия в строении активных центров tPA и АПУТ и, как следствие, различия в степени связи и характере взаимодействия ингибиторной петли с активным центром фермента (т.е. различия во влиянии области ингибиторной петли на поддержание конформации активного центра) не позволяют теоретически предсказать влияние мутационных изменений в ингибиторной петле АПУТ на его связь с PAI-1 и ферментативную (фибринолитическую и амидолитическую) активность.
Нельзя не отметить еще одно важное обстоятельство: предлагаемая авторами патента США №5728564 резкая смена зарядов в ингибиторной петле (а эта петля действительно заметно выступает из глобулы АПУТ) может привести к появлению на поверхности белка нового антигенного детерминанта и, как следствие, - к развитию иммунных реакций, особенно в случае повторного применения. Кроме того, снижать чувствительность к PAI-1 следует очень осторожно, чтобы введение препарата не вызывало геморрагических осложнений.
Итак, при получении фармакологически приемлемых модификаций природного АПУТ необходимо соблюдение тонкого баланса между снижением чувствительности к ингибитору PAI-1 и его способностью вызывать геморрагические и иммунологические осложнения и сохранением функциональной активности фермента, что может быть достигнуто преимущественно экспериментальным путем с последующей коррекцией представлений о структуре и функции такой сложной белковой молекулы, какой является АПУТ.
Ранее авторами данного изобретения была получена рекомбинантная плазмидная ДНК pUABC22, кодирующая модифицированный активатор плазминогена урокиназного типа (мАПУТ), и штамм Escherichia coil - продуцент мАПУТ (Патент РФ №2140453). В патенте не приводятся аминокислотная последовательность мАПУТ и нуклеотидная последовательность ДНК, его кодирующей. Известно, что мАПУТ, кодируемый данной плазмидой pUABC22, представляет собой рекомбинантную проурокиназу, которая отличается от природной проурокиназы заменой первых 24 аминокислот N-концевого домена, гомологичного эпидермальному ростовому фактору, на 15 чужеродных аминокислот. Однако мАПУТ (далее мАПУТ22), кодируемый плазмидой pUABC22, не содержит видоизменений в участке связывания с ингибитором PAI-1.
Основная задача настоящего изобретения состояла в получении такого модифицированного АПУТ, который обладал бы пониженной чувствительностью к PAI-1, не имел побочных эффектов и при этом полностью сохранял биологическую активность, присущую природному АПУТ, т.е. в целом характеризовался лучшими, чем природный АПУТ, тромболитическими свойствами.
Новый модифицированный активатор плазминогена урокиназного типа, заявляемый в соответствии с настоящим изобретением, обладает улучшенными тромболитическими свойствамии и имеет выведенную аминокислотную последовательность SEQ ID NO 1. В последовательности выделены замененные по сравнению с природным АПУТ аминокислоты: 2 аминокислоты ингибиторной петли и 16 чужеродных аминокислот N-концевого домена. При этом данный белок обладает фибринолитической и амидолитической активностями, присущими природному АПУТ.
Новый модифицированный АПУТ имеет молекулярную массу 43 кДа, определенную методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия.
Снижение чувствительности мАПУТ к ингибитору PAI-1 достигается за счет произведенных аминокислотных замен в участке связывания ингибитора активатора плазминогена I типа PAI-1 с АПУТ.
Более конкретно, видоизменение, приводящее к уменьшению чувствительности к PAI, состоит в замене естественной последовательности аминокислот ингибиторной петли RRHRGGS (ArgArgHisArgGlyGlySer) на последовательность RHHAGGS (ArgHisHisAlaGlyGlySer).
Данная последовательность аминокислот соответствует аминокислотам №178-184 в нумерации аминокислот природной человеческой проурокиназы, №198-204 в случае ее предшественника - препроурокиназы (Holmes W.E. et al., Biotechnology 0: 923-929, 1985; SWISS-PROT: P00749) и №170-176 в случае заявляемой последовательности (SEQ ID NO 1). Такая замена позволяет избежать нарушений укладки белка при одновременном уменьшении чувствительности к PAI-1.
За счет указанных видоизменений достигается большая ферментативная активность в присутствии PAI-1, чем в случае природного АПУТ. Это позволяет достичь аналогичного тромболитического эффекта при меньших дозах препарата. Кроме того, новый модифицированный АПУТ может оказаться особенно эффективным для пациентов с повышенным содержанием PAI-1 в крови.
Новый модифицированный АПУТ содержит дополнительно видоизмененную по сравнению с природным АПУТ последовательность амино-терминального домена, гомологичного эпидермальному ростовому фактору, а именно: 24 аминокислоты N-концевого домена, вызывающие пролиферацию клеток, заменены на 15 чужеродных аминокислот.
Следующим объектом заявляемой группы изобретений является последовательность ДНК, которая кодирует новый модифицированный АПУТ и имеет нуклеотидную последовательность SEQ ID NO 2.
В приведенной последовательности SEQ ID NO 2 выделены нуклеотиды, кодирующие замененные аминокислоты. Выделенные кодоны могут быть заменены любыми другими аналогичными кодонами, определяемыми соответствующими замененными аминокислотами.
Последовательность ДНК, кодирующую новый модифицированный АПУТ, получают с использованием метода сайт-направленного мутагенеза.
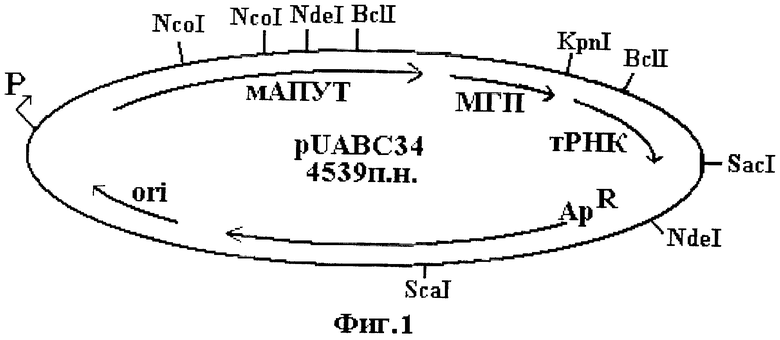
Еще одним объектом изобретения является рекомбинантная плазмидная ДНК pUABC34 для экспрессии нового модифицированного АПУТ, имеющего аминокислотную последовательность SEQ ID NO 1. Плазмида pUABC34 имеет размер 4539 п.н. и содержит следующие последовательно соединенные конструктивные элементы:
- фрагмент ДНК, кодирующий мАПУТ и имеющий нуклеотидную последовательность SEQ ID NO 2, содержащую 2 участка узнавания рестриктазой NcoI, участок узнавания рестриктазой NdeI и участок узнавания рестриктазой BelI;
- искусственную межгенную последовательность МГП14 и ген тРНК Arg;
- ген устойчивости к ампициллину;
- участок начала репликации;
- lac-промотор.
Карта плазмиды представлена на фиг.1. Карта не дает исчерпывающей информации о деталях последовательности, например о сайтах рестрикции, присутствующих в данной ДНК, однако указанные на этой фигуре сайты являются достаточными для однозначного распознавания функциональных элементов плазмиды.
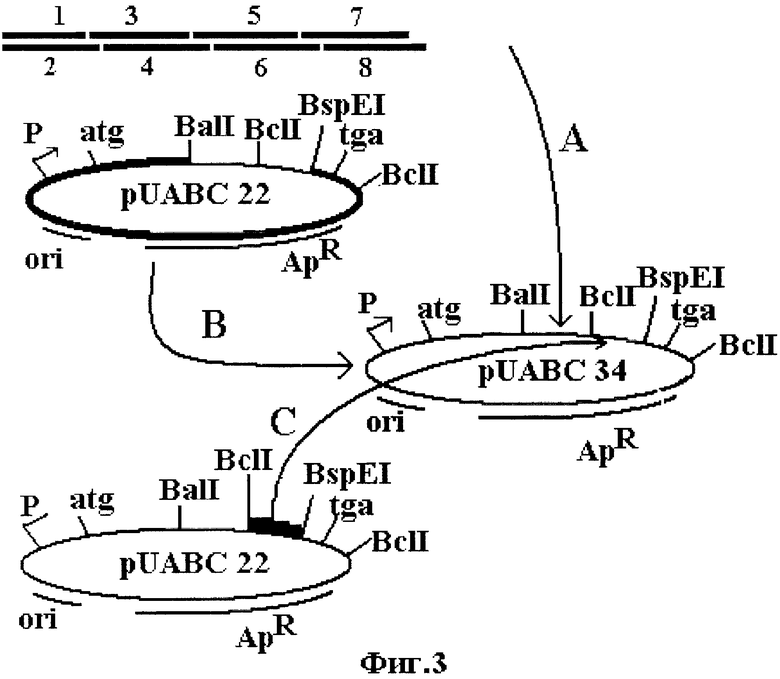
В основе получения плазмиды pUABC34 лежит технология рекомбинантных ДНК, все методы которой хорошо известны специалистам в данной области. Для этого BalI - BspEI - фрагмент плазмиды pUABC22 размером 4 т.п.н., содержащий ген устойчивости к антибиотику, участок начала репликации, ген аргининовой тРНК, искусственную межгенную последовательность МГП 14, а также N- и С-концевые фрагменты кодирующей области мАПУТ, и BspEI - BslI - фрагмент плазмиды pUABC22 размером 0.33 т.п.н, представляющий собой фрагмент кодирующей области мАПУТ, не содержащий видоизменений последовательности, лигируют с синтетическим фрагментом "А", который представляет собой последовательность, кодирующую измененный участок связывания с PAI-1 (см. фиг.3). В свою очередь, для получения синтетического фрагмента "А" синтезируют 8 попарно комплементарных олигонуклеотидов (см. фиг.2), фосфорилируют их при помощи полинуклеотидкиназы фага Т4, попарно отжигают и лигируют с образованием синтетического фрагмента рамером 150 п.н.
Следующим объектом заявляемой группы изобретений является штамм Escherichia coli - продуцент мАПУТ. Штамм получают путем трансформации клеток E.coli K-12 JM109 рекомбинантной плазмидной ДНК pUABC34.
Штамм депонирован во Всероссийской Коллекции Промышленных Микроорганизмов и имеет коллекционный номер ВКПМ В-8145.
Еще одним объектом заявляемой группы изобретений является способ получения нового модифицированного АПУТ. Способ заключается в том, что культивируют клетки штамма-продуцента Escherichia coli ВКПМ В-8145 в условиях, обеспечивающих экспрессию мАПУТ, и выделяют из них целевой продукт.
Для этого клетки штамма E.coli ВКПМ В-8145, содержащие плазмиду pUABC34, выращивают до плотности OD600=2.5 в LB-среде с ампициллином при 37°С и интенсивной аэрации, собирают с помощью центрифугирования, обрабатывают лизоцимом и разрушают с помощью ультразвука. Осадочную фракцию клеточного лизата растворяют в денатурирующих условиях, а затем проводят реконституцию ферментативной активности мАПУТ.
Еще одним объектом заявляемой группы изобретений являются фармацевтические композиции, обладающие тромболитическим действием. Фармацевтические композиции включает в себя новый мАПУТ в качестве действующего вещества и фармацевтически приемлемый носитель.
Фармацевтические композиции могут быть получены известными в данной области техники способами.
Фармацевтические композиции на основе нового мАПУТ могут быть использованы в качестве лекарственного средства в любой лекарственной форме, преимущественно в виде парентеральных препаратов, таких, например, как растворы для инъекций.
При приготовлении фармацевтических композиций в форме раствора для инъекций в качестве носителей могут быть использованы: растворители, такие как вода, этиловый спирт, макрогель (полигликоль), пропиленгликоль, этоксилированный изостеариловый спирт, полиоксилированный изостеариловый спирт, полиоксиэтиленсорбитановый эфир жирной кислоты, полисахара, такие как манетол, декстран и т.п.; регуляторы рН и буферы, такие как цитрат натрия, ацетат натрия или фосфат натрия, стабилизаторы, такие как пиросульфит натрия, глютатион, этилендиаминтетрауксуасная кислота, тиогликолевая кислота или тиомолочная кислота и др. В этом случае лекарственный препарат может содержать хлорид натрия, глюкозу или глицерин в количестве, достаточном для получения изотонического раствора. Указанные носители могут быть добавлены для получения растворов для подкожных, внутримышечных и внутривенных инъекций способом, известным в данной области техники.
Количество мАПУТ по настоящему изобретению, которое должно содержать вышеописанные фармацевтические композиции, изменяется в соответствии с типом композиции, способом введения, схемой дозирования и поэтому не может быть точно определено, а выбирается в соответствии с потребностью из определенного диапазона. Композиция может предпочтительно содержать примерно 0,1-10 мас.% мАПУТ от массы композиции.
Для лучшего понимания настоящего изобретения описание сопровождается следующими иллюстрациями:
На фиг.1 представлена карта рекомбинантной плазмидной ДНК pUABC34, содержащая основные функциональные элементы плазмиды и характерные участки узнавания рестриктазами. Промотор lac-оперона обозначен как "Р"; мАПУТ означает кодирующую область мАПУТ; АрR - ген устойчивости к ампициллину, ori- участок начала репликации; ген аргининовой тРНК (Е.соli) обозначен как тРНК, искусственная межгенная последовательность МГП14 обозначена как МГП. Для привязки расположения функциональных элементов к физической карте (последовательности) приведены участки узнавания рестриктазами: NcoI, NdeI, KpnI, SacI и ScaI.
На фиг.2 представлены структуры олигонуклеотидов In1-In8, которые имеют нуклеотидные последовательности SEQ ID NO 3-10 и применяются при синтезе искусственного фрагмента "А" кодирущей области мАПУТ. Направление последовательности задается указанием 3'- и 5'-концов; для наглядности олигонуклеотиды расположены таким образом, как они входят в состав искусственного фрагмента кодирущей области мАПУТ; указано, какие из олигонуклеотидов фосфорилированы; при этом иллюстриуется сборка обеих цепей. Непосредственно под последовательностями олигонуклеотидов указаны кодируемые ими последовательности аминокислот. Аминокислоты, отличные от природных, подчеркнуты.
Фиг.3 иллюстрирует конструирование рекомбинантной плазмиды pUABC34, кодирующей мАПУТ. Буквами А, В и С обозначены фрагменты ДНК, из которых составляется плазмида pUABC34. Фрагмент "А" - искусственный фрагмент кодирующей области мАПУТ размером 0.15 т.п.н. Фрагмент "В" - это BalI - BspEI-фрагмент плазмиды pUABC22 размером 4 тпн. Он содержит ген усточивости к антибиотику, участок начала репликации, ген аргининовой тРНК, искусственную межгенную последовательность МГП14, а также N- и С-концевые фрагменты кодирующей области мАПУТ. Фрагмент С представляет собой BspEI - BelI - фрагмент плазмиды pUABC22 размером 0.33 т.п.н. Он содержит фрагмент кодирующей области мАПУТ, не содержащий видоизменений.
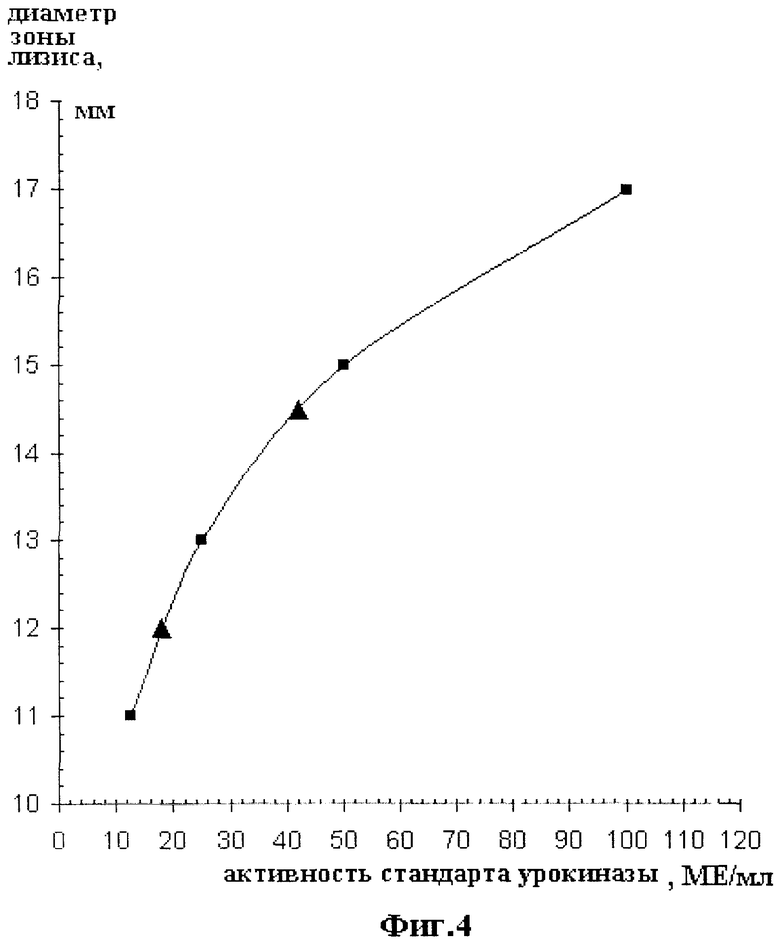
Фиг.4 иллюстрирует построение калибровочной кривой фибринолитического теста и определение активности мАПУТ. По горизонтальной оси откладывается активность стандарта урокиназы в МЕ/мл. При построении использовались точки 12.5, 25, 50 и 100 МЕ/мл. По вертикальной оси откладывается диаметр зоны лизиса в мм. Калибровочные точки обозначены квадратными значками. Точки, по которым производилось измерение экспериментальных образцов мАПУТ, обозначены треугольниками.
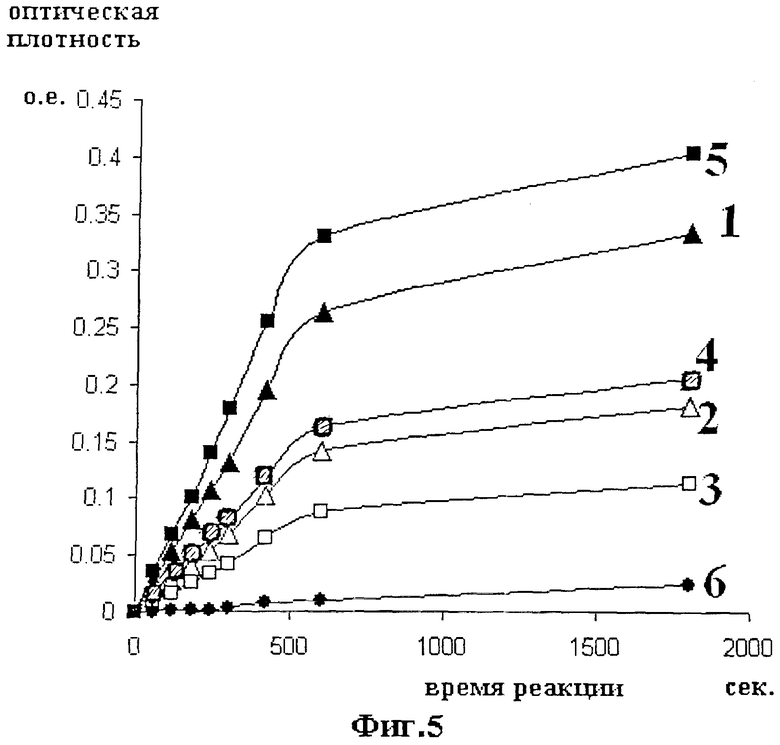
Фиг.5 иллюстрирует результаты амидолитического теста. По горизонтальной оси откладывается время реакции в секундах. По вертикальной оси откладывается оптическая плотность реакционной смеси после остановки реакции. Оптическая плотность измеряется при 405 нм. Значению OD=0.1 соответствует 0.28 мМ концентрация гидролизованного хромогенного субстрата S2444.
Кинетические кривые 3, 4 и 5 получены для концентраций стандарта 625, 1250 и 2500 МЕ/мл соответственно. Треугольными значками обозначены точки, по которым производилось измерение экспериментальных образцов мАПУТ. Кинетические кривые 1 и 2 получены для концентраций мАПУТ 20 и 10 мкг/мл соответственно. Круглые значки соответствуют значениям фона, создаваемого плазмином (кривая 6).
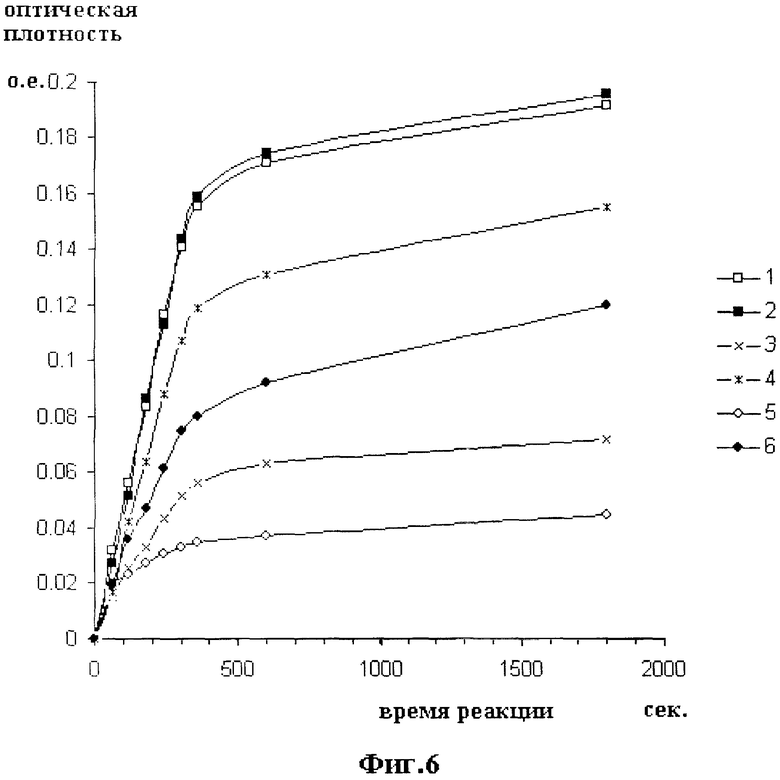
На фиг.6 отражены результаты амидолитического теста в присутствии ингибитора PAI-1. Кривые иллюстрируют влияние PAI-1 на заявляемый модифицированный АПУТ (мАПУТ34), содержащий модификацию аминокислотной последовательности в участке связывания ингибитора, и известный модифицированный АПУТ (мАПУТ22, Патент РФ №2140453), у которого такая модификация отсутствует. По горизонтальной оси откладывается время реакции в секундах. По вертикальной оси откладываются значения оптической плотности реакционной смеси после остановки реакции при 405 нм. Значению OD=0.1 соответствует 0.28 мМ концентрация гидролизованного хромогенного субстрата S2444.
Кривая 1 - мАПУТ22 без PAI; кривая 2 - мАПУТ34 без PAI, кривая 3 - молярное соотношение мАПУТ22/РАI=1:1; кривая 4 - молярное соотношение мАПУТ34/РАI=1:1; кривая 5 - молярное соотношение мАПУТ22/РАI=1:2, кривая 6 - молярное соотношение мАПУТ34/РАI=1:2.
Перечень последовательностей нуклеотидов и аминокислот, характеризующих настоящее изобретение, включает следующие последовательности:
SEQ ID NO 1 представляет собой полную выведенную аминокислотную последовательность нового модифицированного АПУТ. В последовательности выделены замененные по сравнению с природным АПУТ аминокислоты.
SEQ ID NO 2 представляет собой фрагмент ДНК, кодирующий новый мАПУТ. Выделенные кодоны могут быть заменены любыми аналогичными, соответствующими замененным аминокислотам.
SEQ ID NO 3 - SEQ ID NO 10 представляют собой последовательности олигонуклеотидов In1-In10 соответственно, используемых при получении синтетического фрагмента "А" кодирующей области мАПУТ.
Ниже приводятся примеры конкретного осуществления настоящего изобретения нижеследующими примерами. Однако следует иметь в виду, что эти примеры иллюстрирует, но не ограничивают настоящее изобретение.
Пример 1. Получение синтетического фрагмента "А".
На фиг.3 представлена сборка искусственного фрагмента кодирующей области из олигонуклеотидов, который обозначен как синтетический фрагмент "А". Он заменяет собой природный BalI-BclI фрагмент. Синтетические олигонуклеотиды In1-In8 синтезируют фосфорамидитным методом. Используют последовательности нуклеотидов, обозначенные как SEQ ID NO 3 - SEQ ID NO 10 в Перечне последовательностей и представленные на фиг.2.
5×10-10 моль каждого олигонуклеотида растворяют в 200 мкг буферного раствора состава: трис-НСl 25 мМ, рН 7.5; MgCl2 10 мМ; АТР 0.5 мМ; EDTA 0.1 мМ. К олигонуклеотидам In2-In7 добавляют полинуклеотидкиназу фага Т4 в концентрации 0.1 ед./мкл. Фосфорилирование продолжается 1 час при 37°С. Далее полинуклеотидкиназу инактивируют при 90°С в течение 20 сек.
Фосфорилированные олигонуклеотиды объединяют попарно: In3 с In4, In5 с In6. К фосфорилированному олигонуклеотиду In2 добавляют нефосфорилированный олигонуклеотид In1 (5·10-10 моль), к фосфорилированному олигонуклеотиду In7 добавляют нефосфорилированный олигонуклеотид In8 (5·10-10 моль). Объединенные попарно олигонуклеотиды прогревают при 90°С в течение 10 сек, а затем медленно (2 часа или более) охлаждают до +20°С в металлическом контейнере. Четыре полученных сшитых попарно олигонуклеотида смешивают, добавляют ДНК-лигазу фага Т4 (до 10 м/мл), инкубируют в течение 20 часов при +15°С, а затем инактивируют ДНК-лигазу 60°С за 20 сек.
Продукты реакции разделяют в 5% полиакриламидном геле. После окрашивания бромистым этидием из геля вырезают продукт длиной 150 п.н. и экстрагируют его в буферный раствор (трис-НСl 20 мМ, рН 7.5, NaCl 20 мм; EDTA 0.1 мМ) путем пассивной диффузии в течение 60 часов при 50°С. Продукт (синтетический искусственный фрагмент "А", кодирующий измененный участок связывания с PAI-1) концентрируют бутанолом-1 до объема от 100 до 400 мкл, а затем высаживают добавлением 3 объемов этанола. Продукт растворяют в буферном растворе состава: трис-НСl 20 мМ, рН 7.5; EDTA 0.1 мМ и его концентрацию определяют спектрофотометрически.
Пример 2. Получение рекомбинантной плазмидной ДНК pUABC34.
1. 10 мкг плазмиды pUABC22 (Патент РФ №2140453) гидролизуют 20 ед. рестриктаз BalI и BspEI в буфере следующего состава: трис-НСl 25 мМ, рН 7.5; MgCl2 10 мМ; NaCl 50 мМ; EDTA 0.1 мМ. Продукты гидролиза разделяют в 0.8% легкоплавкой агарозе (Clontech Lab. Inc.). После окрашивания бромистым этидием идентифицируют фрагмент размером 4 т.п.н., кусочек геля вырезают, подвергают инкубации при 70°С в течение 15 мин., удаляют агарозу из образца экстракцией водным раствором фенола, а примеси фенола удаляют экстракцией бутанолом-1.
Полученный продукт высаживают добавлением 3 объемов этанола и растворяют в буферном растворе: трис-НСl 20 мМ, рН 7.5; EDTA 0.1 мМ. На фиг.3 фрагмент размером 4 т.п.н. обозначен как "В".
2. 10 мкг плазмиды pUABC22 гидролизуют 20 ед. рестриктаз BelI и BspEI в буфере следующего состава: трис-НСl 25 мм, рН 7.5; MgCl2 10 мМ; NaCl 50 мМ; EDTA 0.1 мМ. Продукты гидролиза разделяют в 1.2% легкоплавкой агарозе (Clontech Lab. Inc.). После окрашивания бромистым этидием идентифицируют продукт длиной 0.33 т.п.н., кусочек геля вырезают; удаляют агарозу из образца экстракцией водным фенолом, а примеси фенола удаляют экстракцией бутанолом-1. Полученный продукт высаживают добавлением 3 объемов этанола. Полученный фрагмент растворяют в буфере: трис-НСl 20 мМ, рН 7.5; EDTA 0.1 мМ. На фиг.1 фрагмент размером 0.33 т.п.н. обозначен как "С".
3. Оба выделенных фрагмента смешивают друг с другом и синтетическим фрагментом "А", добавляют MgCl2 до концентрации 10 мМ, АТР - до 0.5 мМ и ДНК-лигазу фага Т4 - до 10 ед./мл. После 20-часовой инкубации при +15°С полученную смесь трансформируют в клетки E.coli K-12 JM109. Для этого клетки E.coli K-12 JM109 выращивают при 37°С в среде LB до достижения оптической плотности 0.3 (при 600 нм). Клетки собирают центрифугированием и ресуспендируют в буфер (КСН3СОО 20 мМ, КСl 100 мМ, СаСl2 10 мМ, МnСl2 10 мМ, glycerol 10%, рН 5.5) при 0°С в объеме, составляющем 1/2 объема исходной культуры. Клетки вторично собирают центрифугированием и ресуспендируют при 0°С в том же буфере в объеме, составляющем 1/20 объема исходной культуры.
К 0.2 мл суспензии клеток добавляют лигазную смесь. После инкубации в течение 40 мин при 0°С и последующей инкубации при 42°С в течение 2 мин к клеткам добавляют 1 мл среды LB. Клетки инкубируют при 37°С в течение 40 мин и высевают на чашки с ампициллином (0.1 мг/мл). Из полученных клонов с помощью рестриктного анализа отбирают клоны, содержащие плазмиды с 2 участками узнавания рестриктазой NcoI и 2 участками узнавания рестриктазой NdeI. Нуклеотидную последовательность одного из отобранных клонов подтверждают методом Сэнгера и плазмиду с подтвержденной нуклеотидной последовательностью обозначают как pUABC34.
Пример 3. Получение штамма-продуцента мАПУТ.
Клетки штамма Escherichia coli JM109 трансформируют плазмидой pUABC34 по методике примера 2 и получают штамм-продуцент модифицированного активатора плазминогена урокиназного типа. Штамм депонирован во Всероссийской Коллекции Промышленных Микроорганизмов и имеет коллекционный номер ВКПМ В-8145.
Пример 4. Получение рекомбинантного модифицированного активатора плазминогена урокиназного типа.
Бактерии штамма E.coli ВКПМ В-8145, содержащие плазмиду pUABC34, выращивают до плотности OD600=2.5 в течение 16 ч в 50 мл LB-среды с ампициллином (100 мкг/мл) при 37°С и интенсивной аэрации. По окончании выращивания клетки собирают центрифугированием в течение 30 мин при 4000 об/мин, суспендируют в 2.5 мл буферного раствора (трис-НСl 20 мМ, рН 6.7, NaCl 30 мМ), добавляют 2.5 мг лизоцима и инкубируют 10 мин при 4°С. К полученной смеси добавляют Трис-HCl, рН 8.3, до 50 мМ и ЭДТА до 20 мМ, инкубируют 30 мин при 4°С, после чего обрабатывают ультразвуком 1 мин. Осадочную фракцию клеточного экстракта отделяют центрифугированием в течение 90 мин при 5000 об/мин, суспендируют в 1 мл 2%-ного Тритона Х-100 и центрифугируют 5 мин при 14000 об/мин. Осадок суспендируют в 100 мкл раствора, содержащего 200 мМ ЭДТА и 500 мМ Трис-HCl, рН 8.0, добавляют 800 мкл денатурирующего раствора (6 М гуанидинхлорид, 100 мМ 2-меркаптоэтанол), инкубируют 20 мин при 65°С, нерастворимый осадок убирают центрифугированием.
Процедуру реконституции ферментативной активности проводят следующим образом. Раствор разбавляют в 20 раз буфером: 100 мМ Трис-HlCl, рН 9.0, 200 мМ аргинин-хлорид, 5 мМ восстановленный глютатион, 1 мМ окисленный глютатион, денатурирующий агент (1 М гуанидинхлорид) и инкубируют в течение 40 ч при 4°С. Полученную смесь подвергают диализу против 1 л раствора, содержащего 100 мМ NaCl, 20 мМ Трис-HCl, рН 8.0, в течение 6 ч. Выпавший осадок удаляют центрифугированием и измеряют активность в тесте на фибринолизис.
Ренатурированный рекомбинантный модифицированный активатор плазминогена представлен, главным образом, проферментной формой с молекулярной массой 43 кДа, что подтверждают данные электрофоретического и зимографических анализов. Выход активного мАПУТ составляет порядка 200000 ед. на 1 г биомассы.
Была исследована специфическая активность нового модифицированного АПУТ в сравнении с природным АПУТ путем определения фибринолитической активности по лизису фибрина и определения амидолитической активности после активации плазмином. Кроме того, было исследовано влияние ингибитора PAI-1 на мАПУТ в тесте на амидолитическую активность. При исследованиях в качестве стандарта использовалась высокомолекулярная урокиназа фирмы Calbiochem с активностью 100000 МЕ/мг.
Пример 5. Определение фибринолитической активности.
Определение фибринолитической активности осуществляют по методу (Gaffney P.J. et al., Thrombos and Heamostas 45: 34-37, 1981). Агарозу (Clontech Lab. Inc.) разводят водой до получения 1.2% суспензии; суспензию кипятят до полного растворения агарозы и охлаждают до температуры 37°С.
Фибриноген (Sigma F 8630) растворяют в фосфатном буфере (20 мМ фосфата натрия, рН 7.5, 300 мМ NaCl, 0.1% Тритон Х-10, 5% глицерин) до концентрации 10 мг/мл при 37°С. Добавляют какодилат натрия (рН 7.4) до концентрации 5 мМ и тромбин (Sigma, T 3399) - до концентрации 0.08 ед/мл. Растворы фибриногена и агарозы смешивают и разливают в чашки Петри. После полимеризации при комнатной температуре чашки могут использоваться или храниться при +4°С.
Приготовляют 0,2% раствор бычьего сывороточного альбумина (БСА) в фосфатном буфере: 10 мМ фосфата натрия, 150 мМ NaCI, pH 7.4. Образец стандарта высокомолекулярной урокиназы разводят в 0,2% растворе БСА таким образом, чтобы получить разведения от 12,5 до 100 МЕ/мл. Затем приготовляют растворы мАПУТ с концентрацией 20 и 40 мкг/мл.
2 мл каждого разведения стандарта высокомолекулярной урокиназы (12.5, 25, 50 и 100 МЕ/мл) и исследуемых образцов наносят на поверхность фибринового геля. После 10-часовой инкубации при 30°С измеряют диаметр зон лизиза для образцов стандарта и мАПУТ. Стандарт используют для построения калибровочной кривой, по которой затем определяют активность мАПУТ. Квадратные значки на фиг.4 иллюстрируют построение калибровочной кривой. Треугольными значками обозначены точки, по которым производилось измерение экспериментальных образцов мАПУТ.
Как следует из результатов фибринолитического теста, активность мАПУТ не отличается от активности природной высокомолекулярной урокиназы и составляет ~105 МЕ/мг.
Пример 6. Определение амидолитической активности.
Образец стандарта высокомолекулярной урокиназы растворяют в 0,2% растворе БСА в фосфатном буфере (10 мМ фосфат натрия, 150 мМ NaCI, pH 7,4) до концентрации 5000 МЕ/мл. Плазмин (Serva, P-32965) растворяют в фосфатном буфере до концентрации 1 ед/мл. 100 мкл раствора стандарта урокиназы инкубируют с 1 мкл раствора плазмина в течение 1,0-1,5 часов при 37°С.
Готовят образцы мАПУТ в 0,2% растворе БСА в фосфатном буфере с концентрацией 10 и 20 мкг/мл. 100 мкл раствора образцов мАПУТ инкубируют с 1 мкл раствора плазмина в течение 1,0-1,5 часов при 37°С.
Готовят последовательные разведения стандарта высокомолекулярной урокиназы, активированной плазмином, с помощью 0.2% раствора БСА в фосфатном буфере. Получают растворы с концентрацией стандарта 625, 1250 и 2500 МЕ/мл.
В качестве контроля используют раствор 1 мкл раствора плазмина в 0.2% раствора БСА в фосфатном буфере.
Амидолитическую активность оценивают по скорости гидролиза хромогенного субстрата S2444 (L-пироглутамил-глицил-L-аргинин-4-нитроанилид гидрохлорид, Serva B-52338) в присутствии добавляемых ферментов. К 100 мкл разведений активированной урокиназы, контрольного раствора и испытуемых образцов мАПУТ добавляют по 10 мкл 10 мМ водного раствора хромогенного субстрата S2444 и инкубируют при 37°С в течение 30 мин. Реакцию останавливают добавлением 500 мкл раствора 10% уксусной кислоты, растворы перемешивают и измеряют оптическую плотность при длине волны 405 нм. Результаты амидолитического теста представлены на фиг.5. Кинетические кривые 3, 4 и 5 получены для концентраций стандарта 625, 1250 и 2500 МЕ/мл соответственно. Треугольными значками обозначены точки, по которым производилось измерение экспериментальных образцов мАПУТ. Кинетические кривые 1 и 2 получены для концентраций мАПУТ 20 и 10 мкг/мл соответственно. Круглые значки соответствуют значениям фона, создаваемого плазмином (кривая 6).
По наклону начального (линейного) участка кривых делают вывод о скорости гиролиза хромогенного субстрата. Скорость гидролиза, отнесенная к концентрации фермента, характеризует удельную активность фермента.
Как следует из результатов амидолитического теста, активность мАПУТ не отличается от активности природной высокомолекулярной урокиназы и составляет ~105 МЕ/мг.
Пример 7. Определение чувствительности мАПУТ к ингибированию PAI-1.
Влияние ингибитора PAI-1 на активность нового мАПУТ (мАПУТ34) определяют в амидолитическом тесте. Для сравнения используют известный мАПУТ22 (Патент РФ №2140453), от которого заявляемый мАПУТ отличается наличием модификации аминокислотной последовательности в участке связывания ингибитора PAI-1.
Готовят образцы мАПУТ22 и мАПУТ34 в 0,2% растворе БСА в фосфатном буфере (10 мМ фосфат натрия, 150 мМ NaCl, pH 7,4) с концентрацией 20 мкг/мл.
Плазмин (Serva, P-32965) растворяют в фосфатном буфере до концентрации 1 ед./мл. По 100 мкл раствора образцов мАПУТ инкубируют с 1 мкл раствора плазмина в течение 1,0-1,5 часов при 37°С.
В качестве контроля используют раствор 1 мкл раствора плазмина в 0.2% раствора БСА в фосфатном буфере.
Ингибитор PAI-1 (Calbiochem) растворяют в 0,1% растворе БСА в фосфатном буфере (фосфат Na 10 мМ, NaCl 150 мМ, pH 7.2-7.5) до концентрации 500 мкг/мл.
К аликвотам по 10 мкл 10 мМ водного раствора хромогенного субстрата S2444 добавляют 0, 4 и 8 мкл раствора PAI-1, что позволяет затем создать соотношения между мАПУТ и PAI, равные 0, 1:1 и 1:2.
К полученным разведениям PAI-1 в растворе хромогенного субстрата S2444 добавляют по 100 мкл контрольного раствора или растворов испытуемых образцов мАПУТ22 и мАПУТ34 и инкубируют при 37°С в течение 1, 2, 3, 4, 5, 6, 10 и 30 мин. Реакцию останавливают добавлением 100 мкл раствора 10% уксусной кислоты, растворы перемешивают и измеряют оптическую плотность при длине волны 405 нм.
Сравнительную активность нового и известного мАПУТ в отсутствие и в присутствии PAI-1 оценивают по характеру кинетических кривых, представленных на фиг.6. Кривая 1 получена для мАПУТ22 без PAI; кривая 2 - для мАПУТ34 без PAI. Кривая 3 получена при молярном соотношении мАПУТ22 и PAI, равном 1:1; а кривая 5 - при соотношении 1:2. Кривая 4 получена при молярном соотношении мАПУТ34 и PAI, равном 1:1; а кривая 6 - при соотношении 1:2.
Поскольку по ходу реакции происходит не только гидролиз хромогенного субстрата, но и постепенная инактивация ферментов за счет их взаимодействия с PAI, то и скорость реакции меняется не только за счет изменения концентрации субстрата, но и за счет постепенной инактивации фермента. Это обстоятельство делает неадекватным описание системы лишь начальной скоростью реакции, а требует описания изменения скорости во времени.
Как следует из данных фиг.6, в отсутствие PAI интегральный эффект реакции как на начальном участке кривой, так и по выходе реакции на плато идентичны для обоих ферментов мАПУТ22 и мАПУТ34. При проведении реакции в присутствии PAI активность как мАПУТ22, так и мАПУТ34 уменьшается, но в существенно разной степени.
Так при соотношении мАПУТ22/РАI, равном 1:1, активность в течение первых 3-х минут уменьшается в 2.5 раза, а через 30 мин - в 2.6 раза по сравнению с теми же величинами в отсутствие PAI. Уменьшение активности мАПУТ34 в аналогичных условиях происходит соответственно лишь в 1.3 и 1.2 раза.
А при соотношении мАПУТ22/РАI, равном 1:2, активность мАПУТ22 в течение первых 3-х минут реакции уменьшается в 3.8 раза, а за 30 мин - в 4.3 раза по сравнению с теми же величинами в отсутствие PAI. Уменьшение активности мАПУТ34 в аналогичных условиях происходит соответственно лишь в 1.8 и 1.6 раза.
Следует отметить, что кривые для мАПУТ22 в присутствии PAI (кривые 3 и 5 фиг.6) выходят на насыщение уже на 6-ой минуте реакции на низком уровне, что говорит о практически полной инактивации мАПУТ22 к этому времени. При этом кривые реакций в отсутствие PAI (кривые 1 и 2) и кривые реакций мАПУТ34 с PAI (кривые 4 и 6) к этому времени еще не насыщаются, что говорит о наличии в реакционной смеси активного фермента.
Пример 8. Лекарственная форма для инъекций.
Для приготовления лекарственной формы для инъекций используют следующий состав: модифицированный АПУТ - 23,5 мг натрия хлорид - 180 мг.
Модифицированный АПУТ растворяют в воде для инъекций, раствор подвергают стерилизации с использованием стерилизационного фильтра "Миллипор" с величиной пор 0,22 мм. Стерильный раствор разливают во флаконы по 5 мл, замораживают при температуре -46°С в течение 4 часов, а затем осуществляют сублимационную сушку при температуре -40°С с постепенным повышением температуры до 20°С в течение 48 часов.
Перечень последовательностей приведен в конце описания.






Изобретение относится к области генетической и белковой инженерии и может быть использовано в медицине. Предложена модифицированная форма активатора плазминогена урокиназного типа (мАПУТ), аминокислотная последовательность которой отличается от последовательности природного АПУТ заменой последовательности ArgArgHisArgGlyGlySer в составе ингибиторной петли последовательностью ArgHisHisAlaGlyGlySer, а также заменой 24 N-концевых аминокислот чужеродной последовательностью из 16 аминокислотных остатков. Сконструирована рекомбинантная плазмида (pUABC 34), включающая фрагмент ДНК, который кодирует новый мАПУТ. В результате трансформации клеток E.coli К-12 JM109 плазмидой pUАВС 34 получен рекомбинантный штамм E.coli ВКПМ-8145, являющийся продуцентом модифицированной формы АПУТ. При полном сохранении биологической активности, присущей природному активатору, продуцируемый рекомбинантным штаммом полипептид характеризуется пониженной чувствительностью к действию ингибитора PAI-1 и отсутствием ряда побочных эффектов, что обеспечивает эффективное применение нового мАПУТ в составе фармацевтических композиций с тромболитическим действием. 6 с.п. ф-лы, 6 ил.
фрагмент ДНК, кодирующий мАПУТ и имеющий нуклеотидную последовательность SEQ ID № 2, содержащую 2 участка узнавания рестриктазой NcoI, участок узнавания рестриктазой NdeI и участок узнавания рестриктазой BclI;
искусственную межгенную последовательность МГП14 и ген тРНК Arg;
ген устойчивости к ампициллину;
участок начала репликации;
lac-промотор.
| US 5728564, 17.03.1998 | |||
| ФАЗОЗАДАЮЩЕЕ УСТРОЙСТВО | 0 |
|
SU253241A1 |
| US 5550042, 27.04.1996 | |||
| 1990 |
|
RU1692151C | |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК P UA BC22, КОДИРУЮЩАЯ МОДИФИЦИРОВАННЫЙ АКТИВАТОР ПЛАЗМИНОГЕНА УРОКИНАЗНОГО ТИПА, НЕТРАНСЛИРУЕМЫЙ ДНК-ЭЛЕМЕНТ - ИСКУССТВЕННАЯ МЕЖГЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ МГП14 И ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ МОДИФИЦИРОВАННОГО АКТИВАТОРА ПЛАЗМИНОГЕНА УРОКИНАЗНОГО ТИПА | 1999 |
|
RU2140453C1 |

