Изобретение касается последовательности ДНК, кодирующей гликопротеин человеческого происхождения.
Гликопротеин проявляет цитотоксическую активность по отношению к различным линиям опухолевых клеток, но не по отношению к нормальным клеткам, выступает как новый противоопухолевый цитотоксический фактор, фактор, способствующий дифференциации лейкемических клеток, фактор, усиливающий клеточную иммунологию, фактор, обеспечивающий рост клеток эндотелия сосудов, фактор, обеспечивающий рост гепацитов.
В выложенной патентной заявке Японии N 64-10998 описаны вся последовательность аминокислот, а также нуклеотидная последовательность в комплементарной ДНК (кДКН) кодирующая противоопухолевый цитотоксический фактор, получаемый из культуры фибробластов человеческого происхождения и имеющего характеристики: молекулярная масса 36000±1000, изоэлектрическая точка свыше 10,5.
Согласно изобретению праймерная последовательность TCF-II была выведена из его кДНК, которая была клонирована сортировкой библиотеки кДНК, полученной с использованием мРНК, выделенной в чистом виде из общего количества всей РНК, которая в свою очередь была экстрагирована из клеток IMR-90 фибробластов зародыша человека согласно следующей методике.
(1) Экстрагирование поли(A) РНК из клеток UMR-90.
Вся РНК была приготовлена методом, предусматривающим использование тиоцианата гуанидина и хлорида цезия. Из клеток IMR-90, взятых в количестве 200 млн, которые культивируют в среде, предложенной Иглом (Eagle) и модифицированной Дулбэкко (Dulbecco), содержащей 5% сыворотки новорожденного теленка (NBCS). Клетки IMR-90, подвергнутые усреднению, представляют собой суспензию в 28 мл 6М тиоцианата гуанидина, содержащую также цитрат натрия (5 мМ), 0,5% препарата Саркосил и бета-меркаптоэтанол (0,1 М). В пробирки центрифуги, выполненные из полиалломера, помещают по 4 мл 5,7 М раствора хлорида цезия, содержащего 0,1 М этилендиаминтетрауксусной кислоты. Затем к раствору хлорида цезия добавляют по 7 мл гомогенизированной клеточной суспензии и осуществляют центрифугирование с частотой вращения 35000 1/мин при 20oC в течение 16 ч на бекмановской (Beckman) центрифуге с ротором 40Т1. После центрифугирования частицы дважды промывают 95%-ным этиловым спиртом и растворяют в 200 мкл буферного раствора Tris HCl (10 мм, pH 75,5), содержащего 1 мМ этилендиаминтетрауксусной кислоты, посредством нагревания при 65oC в течение 5 мин. Полученный раствор далее обозначен как раствор всей РНК. Поли (A) РНК выделяют в чистом виде из общей (всей) РНК методом хроматографии в колонке с олиго (dT) целлюлозой. Раствор всей РНК заливают в колонку с олиго (dT) целлюлозой, приведенную в равновесие с использованием 10 мМ буферного раствора Tris HCl (10 мМ, pH 7,4), содержащего 1 мМ этилендиаминтетрауксусной кислоты и 0,05% SDS. Полученный раствор ниже обозначен как раствор поли(A) РНК.
(2) Синтез библиотеки кДНК.
Двойную спираль кДНК синтезируют с использованием (поли)A РНК из параграфа (1) в качестве шаблона, а также с использованием фирменного средства синтеза кДНК (производство фирмы Pharmacia Co. Ltd), причем дополнительно присоединяют адаптор EcoR 1. Методику синтеза осуществляют согласно рекомендациям указанной фирмы за исключением добавления (на стадии синтеза одинарной спирали ДНК) обратной транскриптазы (40 ед. на реакционную смесь, производство фирмы Life Science Co. Ltd), полученной из культуры несферического болезнетворного вируса, поражающего костный мозг птиц.
(3) Подготовка библиотеки кДНК.
Комплементарную ДНК (кДНК), полученную, как указано в параграфе (2), встраивают EcoR 1 часть (производство фирмы Promega Co Ltd) вектора λgt 10 фага. Эту кДНК в количестве 3,3 мкг, синтезированную из (поли)A РНК, растворяют в 150 мкл буферного раствора 66 мМ Tris-HCl (pH 7,6) для хроматографической колонки, содержащего спермидин (1 мМ), хлорид магния (10 мМ), дитиотрейтол (15 мМ), а также альбумин бычьей сыворотки (0,2 мг/мл). Данный раствор в количестве 6,2 мкл смешивают с 1 мкг EcoR 1 части фага (вектор λgt 10), после чего осуществляют осаждение этиловым спиртом. Рекомбинантный фаг ДНК, включающий оба компонента, λgt 10 и кДНК, готовят следующим образом. Упомянутый выше осадок переводят в суспензию с использованием 9 кмл буферного раствора, предназначенного для хроматографической колонки, и оставляют на инкубирование на ночь при 16oC при условии добавления 1 мкл 10 мМ трифосфата аденозина, а также 1 мкл лигазы Т4 ДНК в количестве 350 ед. на 1 мкл.
(4) Скрининг библиотеки ДНК.
(i) Приготовление олигонуклеотидной пробы.
Для приготовления пробы составляют смесь (маркировка 5' terminus) комплементарного олигонуклеотида с характеристикой 17 мер (mer) (сочетание 384 разновидностей) соответствующего последовательности аминокислот от Va11 до Pro6 в N-терминальной последовательности аминокислот бета-цепи фактора TCF-II, с полинуклеотидной киназой (производство фирмы TAKARA SHUZO) и фактором [γ-32P]ATP (производство фирмы Amersham Co., Ltd). Указанная проба ниже показана следующим образом: дополнительная спираль, использованная в качестве эталона (смесь 384 разновидностей):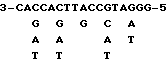
(ii) Скренинг рекомбинантного фага.
500 тыс. дисков фага получают лабораторной расфасовкой раствора рекомбинантного фага ДНК, приготовленного согласно параграфу (3), с использованием препарата Gigapack Gold (Stratagene) и с последующим заражением кишечной палочкой E.coli С600hf1. После адсорбирования дисков фага на фильтре марки Hybond-N (фирма Amersham) эти диски денатурируют щелочью, нейтрализуют и подвергают термообработке при 80oC в течение 2 ч. Гибридизацию по методике, которую разработали Белл и сотр., проводят с использованием смешанной пробы, полученной согласно параграфу (1). Один из клонов, который должен был содержать фрагмент TCF-II, был найден среди позитивных клонов, обнаруженных при первой сортировке.
(5) Клонирование полной длины кДНК фактора TCF-II.
Внутренние последовательности аминокислот (однобуквенные коды): (α) NYMGNLSQTRSGL и (β) TSXSVYGWGYTGLINYDGLL (аминокислота X не идентифицирована) определяют из альфа- и бета- цепей TCF-II посредством обработки лизилэндопептидазой при условии последующего картирования (mapping) фрагментов. N-терминальная последовательность аминокислот бета-цепи TCF-II совпадала с аналогичной последовательностью бета-цепи одного из факторов (hHGF) роста гепатоцитов человека. Более того, упомянутые выше внутренние последовательности (α) и (β) фактора TCF-II имелись соответственно и в альфа- и бета-цепях hHGF. Следовательно, было сделано заключение о том, что фактор TCF-II есть следствие экспрессии одного из генов семейства hHGF. Были сообщены данные о библиотеках кДНК генов hHGF из плаценты и печени соответственно (3-u). Сравнение полных праймерных последовательностей, выведенных из обеих кДНК, выявило различия аминокислот на 14 позициях в их последовательности. На основании этих данных было выдвинуто предположение о присутствии генов семейства hHGF. Области, идентичные для кДНК (сDNA) генов hHGF как из плаценты, так и из печени, были выбраны в качестве праймерных (primer) последовательностей для цепной реакции полимеразы (Polymerase Chain Reaction-PCR). Химически синтезируют идентичные олигонуклеотиды обеих кДНК генов hHGF на некодирующих позициях 5' и 3'. Сортировку кДНК фактора TCF-II проводят методом PCR c использованием этих олигонуклеотидов в качестве праймеров (primer). Праймер Sa1-77, имеющий участок расщепления ограничивающего энзима Sa1 I, и праймер Sph 2203, имеющий участок расщепления ограничивающего энзима Sph1, синтезируют с использованием реагента, способствующего синтезу ДНК (производство фирмы Applied Co, Ltd). Ниже эти праймеры показаны следующим образом:
праймер Sa1 - 77: 5'-GGTCGACTAGGGACTGACTCCGAACAGGATTC-3' Sa1 I
праймер Sph 2203: 5'-GGCATGCACAGTTGTATTGGTGGGTGCTTCAG-3' Sph I
Клонирование методом PCR проводят по следующей методике.
(i) Цепная реакция полимеразы (PCR), мкл:
Дополнительная ДНК (cDNA), синтезированная как описано в параграфе (2) и растворенная в 150 мкл буферного раствора хроматографической колонки - 1
20 мкМ праймер Sa1-77 - 2,5
20 мкМ праймер Sph 2203 - 2,5
10 порций реакционного раствора для проведения реакции PCR (буферный раствор на основе 100 мМ Tris HCl с величиной pH 8,3, содержащий 500 мМ KCl, 15 мМ MgCl и 0,1% (мас.ч./об.ч) желатина) - 10
Смесь 1,25 мМ dGTP, dATP, dTTP, dCTP - 16
Amp1i Tag (5 ед. на 1 мкл, производство фирмы TAKARA SHUZO) - 0,5
Дистиллированная вода - 67,5
Затем указанные выше растворы смешивают в пробирке микрофуги объемом 0,5 мл, причем поверхность жидкости закрывают минеральным маслом в количестве 100 мкл (производство фирмы Sigma Co., Ltd). Реакцию PCR проводят с использованием системы Quick Thermo (производство фирмы Japan Genetics Co., Ltd). После предварительной обработки при 94oC в течение 7 мин 35 раз повторяют трехстадийную реакцию, включающую: 1) термообработку при 55oC в течение 3 мин; 2) реакцию с участием полимеразы при 72oC в течение 2 мин, и 3) реакцию денатурирования при 94oC в течение 2 мин. Далее реакционную смесь нагревают при 55oC в течение 3 мин, а затем при 72oC в течение 11 мин, после чего охлаждают ее до комнатной температуры (в указанное время включают также время перехода с одного режима на другой). При анализе части реакционной смеси методом электрофореза с использованием геля агарозы получают фрагмент ДНК, состоящий из приблизительно 2,3 Kb оснований (Kirobases) и признанный в качестве искомой кДНК фактора TCF-II. Далее ДНК, полученную из четырех пробирок, содержащих вышеупомянутую реакционную смесь, осаждают этиловым спиртом и гидролизуют энзимами Sa1 I и Sph I. В результате электрофореза на геле агарозы с использованием бумажного фильтра DE81 (производство фирмы Watmen Co., Ltd) был получен фрагмент ДНК с приблизительно 2,3 Kb.
(ii) Субклонирование.
Фрагмент ДНК (2,3 Kb), содержащий Sa1 I и Sph I, сайты, полученный согласно параграфу (1), встраивают с использованием (ligation) буфера для лигирования (производство фирмы TАKARA SHUZO) в плазмидный вектор pUC18 (производство фирмы Japan gene Сo., Ltd), обработанный энзимами Sa1 I и Sph I. Рекомбинантной ДНК трансфецируют кишечные палочки Esherichia Co1i DH5 по методике фирмы BRL Co, Ltd. Удавалось получить более 20 субклонов.
(iii) Определение последовательности оснований.
Последовательности оснований в полученных субклонах определяют дидеокси-методом с использованием секвеназы (версия 2,0 производства фирмы TOYOBO). Ошибочно включенные нуклеотиды на препарате Ampi Tag (производство фирмы TAKARA SHUZO) были исправлены путем анализа нуклеотидной последовательности оснований в нескольких субклонах.
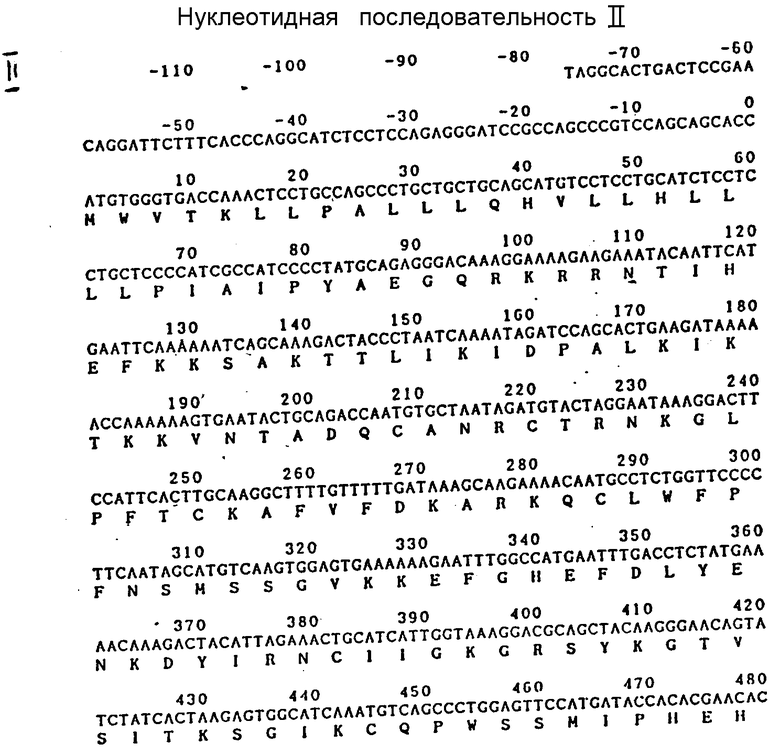
На фиг. 1 и 2 представлены нуклеотидная последовательность кДНК фактора TCF-II, полученная по упомянутой выше методике, а также последовательность аминокислот, выведенная из этой нуклеотидной последовательности.
Нуклеотидная последовательность содержит 2172 пары оснований (п.о.) от АТС запускающего кодона до кодона ТАС терминации транскрипции. После аминокислотного транслирования фактор TCF-II содержит 723 аминокислоты. Было сделано допущение о том, что последовательность аминокислот от первого, метионинового (Met) до 29-го аланинового (A1a) остатков представляет собой сигнальную последовательность. Как показано на фиг.1, фактор TCF-II - это полипептид, содержащий цепи α и β, связанные дисульфидной связью, которые сначала синтезируются как единая цепь. Поскольку N-замыкание цепи фактора TCF-II было блокировано, его не удалось идентифицировать. Однако N-замыкающая последовательность аминокислот цепи β, а также несколько внутренних последовательностей аминокислот фактора TCF-II были определены, как упомянуто выше.
Полученная нуклеотидная последовательность кДНК TCF-II очень похожа на соответствующую последовательность в hHGF, которую установил Миядзава и сотр. Однако в кДНК фактора TCF-II отсутствуют кодоны пяти аминокислотных остатков (F-L-P-SS) от Phe162 до Ser166, присутствующие в последовательности аминокислот, характерной для hHGF. Таким образом, факты свидетельствуют о том, что кДНК фактора TCF-II - одна из новых разновидностей семейства генов hHGF.
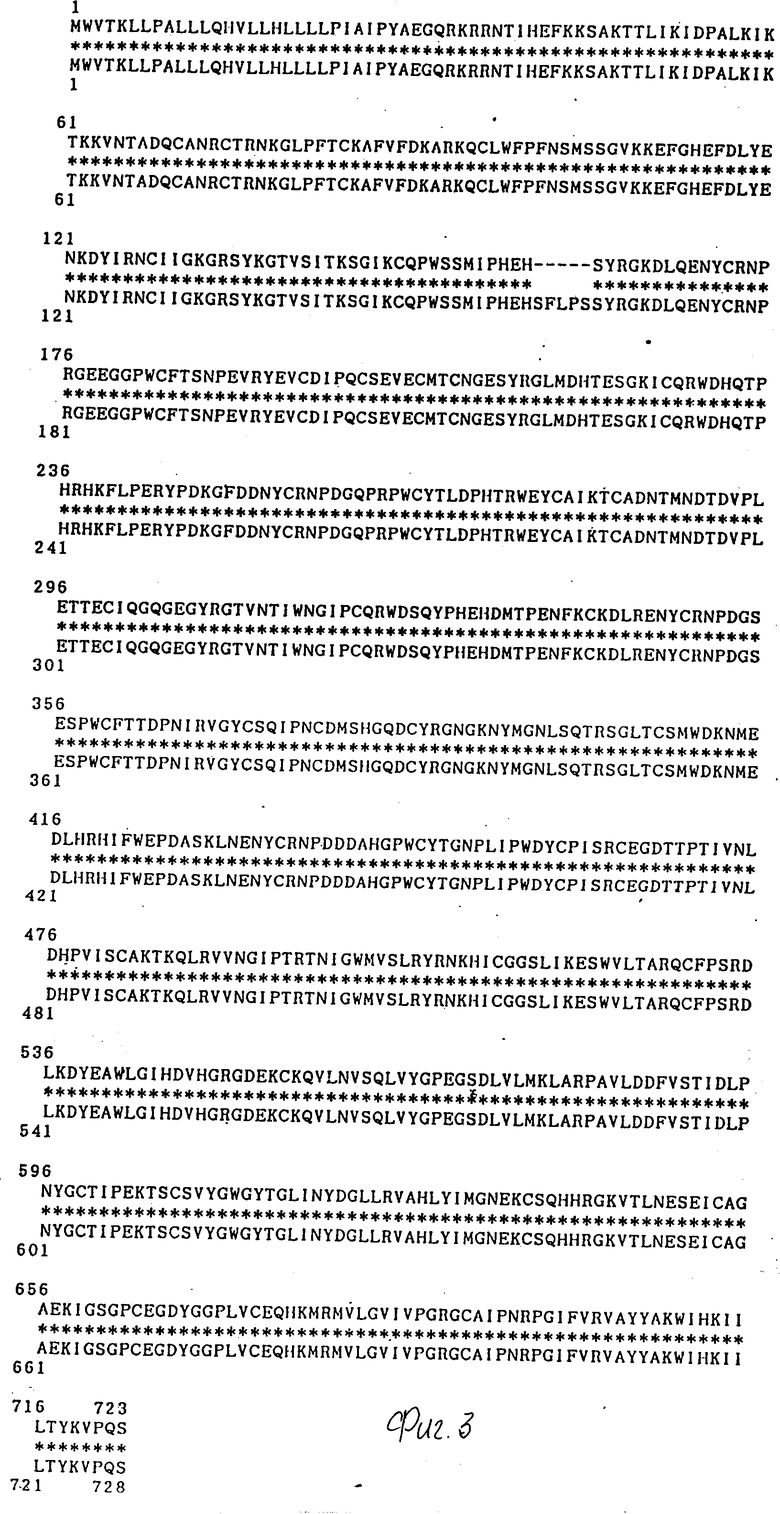
На фиг. 3 представлено сравнение последовательности аминокислот TCF-II, выведенной из упомянутой выше нуклеотидной последовательности, и последовательности аминокислот, соответствующей гену HGF (по данным Миядзавы и сотр. ).
На фиг. 1 и 2 соответственно показаны нуклеотидная последовательность к ДНК, а также последовательность аминокислот фактора TCF-II, выведенная из указанной последовательности оснований.
На фиг.3 дано сравнение последовательности аминокислот TCF-II, выведенной из вышеупомянутой нуклеотидной последовательности и последовательности аминокислот гена hHGF, о которой сообщили Миядзава и сотр.
Полученную согласно изобретению ДНК можно использовать для получения гликопротеина, обладающего противоопухолевой цитотоксической активностью, индуцирующего дифференции линии клеток лейкемии активностью, усиливающего клеточную иммунологию активностью, способствующего росту клеток эндотелии сосудов активностью.







Изобретение относится к биотехнологии. Получение нуклеотидная последовательность фрагмента к ДНК, кодирующего новый гликопротеин ТСF-II человеческого происхождения, обладающий противоопухолевой активностью, активностью по индуцированию дифференциации клеток лейкемии, активностью по усилению клеточной иммунологии, активностью по стимулированию роста клеток эндотелия сосудов, а также активностью по стимулированию роста генатоцитов. 1 з.п. ф-лы, 3 ил.
| JP, заявка, 64-10993, C 12 P 21/02, C 12 N 15/00, 1989. |

