Изобретение относится к биотехнологии и молекулярной биологии, а именно к генной и клеточной инженерии, представляет интерес для крупномасштабного получения рекомбинантного природного эритропоэтина (ЭПО) человека, который может быть использован в медицинских и исследовательских целях.
В литературе описано несколько работ по экспрессии гена ЭПО в животных клетках и по созданию штаммов-продуцентов рекомбинантного ЭПО на основе клеток грызунов и приматов, например на основе линии клеток СНО (клетки яичника китайского хомячка).
Известна плазмида pDSVL-gHuEPO, содержащая полноразмерный ген ЭПО человека под контролем промотора поздних генов вируса SV40, а также миниген дигидрофолат редуктазы (DHFR) мыши [1, 2]. После трансфекции экспрессирующим вектором pDSVL-gHuEPO клеток линии CHOdhfr и селекции в среде без нуклеотидов получен клон клеток, секретирующий рекомбинантный ЭПО в культуральную жидкость с выходом 18,2 единицы активности (ед.акт.) на мл [3]. Путем проведения нескольких раундов амплификации в присутствии возрастающих концентраций метотрексата был получен штамм-продуцент, стабильно секретирующий ЭПО с выходом до 100 ед.акт./мл.
Известна плазмида pRKFL13, содержащая кДНК гена ЭПО под контролем основного позднего промотора аденовируса, сайт полиаденилирования, энхансер и участок инициации репликации вируса SV40, вирус-ассоциированный ген аденовируса, а также ген DHFR под контролем раннего промотора SC40 [4]. Путем трансфекции плазмидой pRKFL13 клеток CHOdhfr с последующей селекцией трансфектантов в среде без нуклеотидов, а также проведением трех раундов амплификации в среде с метотрексатом получен штамм-продуцент ЭПО, секретирующий гормон с выходом до 50 ед/мл [3].
К недостаткам описанных аналогов следует отнести невысокие уровни секреции рекомбинантного ЭПО клетками-продуцентами, что значительно усложняет процедуры выделения и очистки гормона.
Известна плазмидная ДНК pSV-sEp-poly-Neo и штамм культивируемых клеток китайского хомячка CHO-PE ВСКК(П) N 626D - продуцент ЭПО человека, описанные в работе [5, прототип]. Плазмида pSV-sEp-poly-Neo получена путем клонирования DstEII - BglII ДНК фрагмента длиной 2386 нуклеотидных пар (н.п.), содержащего полный ген ЭПО человека в составе вектора pSV2 gpt [6, 7]. Встройка проводится по сайтам рестрикции HindIII - BglII, с последующей делецией района HindIII - BstEII и слиянием с плазмидой pSV2 neo по сайту рестрикции Eco- RI. Путем трансфекции рекомбинантной плазмидой pSV-sEp-poly-Neo и плазмидой pTK-1 клеток яичника китайского хомячка CHOtk- и последующей селекции трансфектантов в среде, содержащей гипоксантин, аминоптерин и тимидин (ГАТ), отобран клон клеток CHO-PE, содержащий в своем геноме интегрированные копии гена ЭПО и стабильно секретирующий рекомбинантный гормон в культуральную среду.
Недостатком данного продуцента является то, что интегрированный ген эритропоэтина находится в клетке в единичной копии и это не позволяет достичь повышенной продукции данного белка за счет увеличенной дозы целевого гена.
Технической задачей изобретения является упрощение и удешевление процессов получения рекомбинантного ЭПО человека за счет увеличения уровня биологического синтеза ЭПО клетками-продуцентами.
Поставленная задача решается использованием в качестве вектора плазмиды pNut [8], у которой использован активируемый металлтионеионовый промотор мыши МТ-1 и ген дигидрофолат редуктазы DHFR и конструированием на этой основе рекомбинантной плазмиды pKEP-9. С целью повышения уровня биологического синтеза промотор приближен к структурной части полноразмерного гена ЭПО человека, сразу за геном находится сигнал сплайсинга и сайт полиаденилирования ранней области вируса SV40. Такое сочетание регуляторных элементов определяет высокий уровень экспрессии гена ЭПО и эффективность синтеза рекомбинантного гормона в животных клетках. Наличие в плазмиде pKEP9 активного гена дигидрофолат редуктазы DHFR под контролем раннего промотора SV-40 позволяет проводить селекцию и амплификацию чужеродных последовательностей, интегрированных в геном клетки DURX-B11 (dhfr- - вариант линии клеток CHO-K1), в среде, содержащей метотрексат. Сочетание процессов трансфекции, селекции и амплификации позволяет получить линию клеток продуцентов CHOpE-9, содержащую в своем геноме интегрированные копии гена ЭПО (природная форма ЭПО) и стабильно секретирующую рекомбинантный гормон в культуральную жидкость.
Штамм-продуцент CHO-pE-9 отличается от штамма DURX-B11 по признакам, передаваемым последовательностями ДНК, использованными для трансфекции, т.е. является устойчивыми к высоким дозам метотрексата (500 нМ), и синтезирует рекомбинантный эритропоэтин человека с выходом до 400 ед.акт./мл.
Полученный штамм характеризуется следующими признаками:
1. Морфологические признаки.
Культура представлена эпителиоподобными и веретеновидными клетками с крупными ядрами неправильной формы, содержащими от одного до трех ядрышек. Цитоплазма зернистая, вакуолизированная.
2. Культуральные признаки.
Штамм CHOpE-9 поддерживается на смеси отечественных сред Игла МЕМ с добавлением 10% эмбриональной телячьей сыворотки (ЭТС) и 1% неосновных аминокислот. Клетки субстрат зависимы, образуют монослой. Отделение клеток от стекла или пластика проводят смесью растворов 0,02% версена (2/3) и 0,25% трипсина (1/3), кратность рассева 1:3 - 1:4, посевная доза - 100-120 тыс. клеток в 1 мл. Плотного монослоя культура достигает через 3 суток после посева.
3. Устойчивость к антиметаболитам.
При культивировании клеток штамм проявляет устойчивость к метотрексату в концентрации до 500 нМ, обусловленную наличием интегрированных в геном последовательностей ДНК.
4. Криоконсервация.
Криоконсервирование проводят на среде Игла МЕМ (80%) с добавлением 10% эмбриональной телячьей сыворотки и 10% DMSO в качестве криопротектора. Концентрация клеток 3-5 млн. в 1 мл. Жизнеспособными после восстановления остаются 95% клеток.
5. Кариологическая характеристика.
Кариологический анализ штамма CHOpE-9 проводят на препаратах, приготовленных без выжигания фиксатора в пламени спиртовки и окрашенных азуром-зозином.
Частоту полиплоидии в популяции делящихся клеток определяют на основании подсчета 1000 клеток. Доля полиплоидных клеток составляет 8%.
Анализ распределения клеток по числу хромосом на препаратах, окрашенных азуром-зозином. Просмотру подвергают 50 клеток. Модальный класс - 20 хромосом.
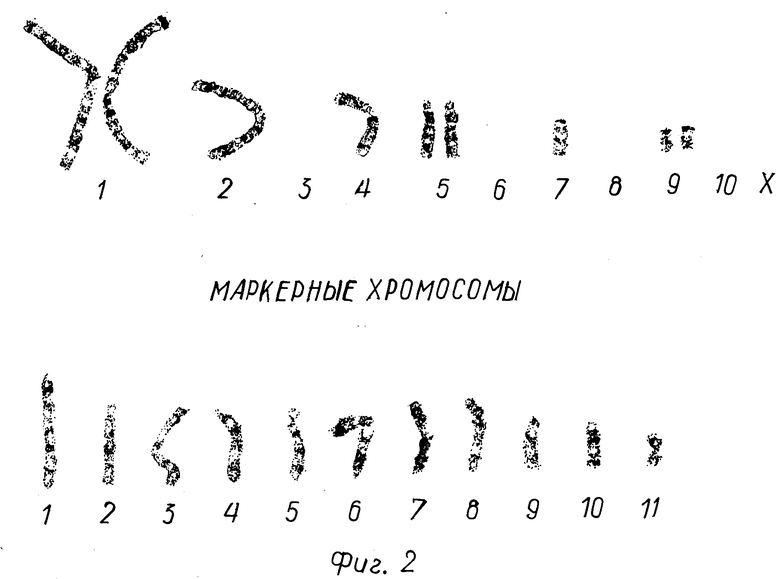
Проведена фотореконструкция 15-ти метафазных пластинок. Установлено, что кариотип клеток CHOpE-9 стабилен. Основной набор состоит их хромосом 1-, 2-, 4-, 5-, 7-, 9-, 10-й пары. X-хромосома перестроена и образует маркерную хромосому М3. Выявлено 11 постоянно встречающихся маркерных хромосом (фиг. 2).
6. Контроль видовой идентичности.
Соответствие виду подтверждается с помощью анализа электрофоретической подвижности изоферментов - глюкозо-6-фосфат дегидрогеназы и лактатдегидрогеназы в полиакриламидном геле. Клетки имеют изоферментный набор, характерный для клеток китайского хомячка.
7. Контаминация.
При обследовании клеток штамма на стерильность (наличие бактерий, грибов и микоплазмы) получают отрицательные результаты на уровнях 10 и 20 пассажей.
8. Продуктивность штамма.
Штамм-продуцент CHOpE-9 синтезирует рекомбинантный эритропоэтин человека с выходом в культуральную среду до 400 ед.акт./мл.
Предложенные рекомбинантная плазмида pKEP-9 и штамм культивируемых клеток китайского хомячка CHOpE-9 впервые получены авторами данного изобретения, в литературе не описаны, следовательно, можно сделать вывод о соответствии технического решения критериям изобретения "новизна" и "изобретательский уровень".
Перечень фигур графического изображения
Фиг. 1. Схема конструирования и генетическая карта рекомбинантной плазмиды pKEP9. Светлыми блоками обозначены ДНК-последовательности вируса SV40, темными блоками - районы гена дигидрофолат редуктазы (DHFR), тонкой линией - ДНК-фрагменты плазмиды pUC18, точками отмечен промотор MT-1, штриховкой - остаток гена ксантин-гуанин фосфорибозил трансферазы E.coli (gpt), серыми блоками - ген ЭПО. Стрелками указаны направления транскрипции генов. Плазмида pKEP9 имеет в своем составе: - BstEII-BglII-фрагмент плазмидной ДНК pSV-Ep-gpt размером 2383 н.п., содержащий ген эритропоэтина;
-EcoRI-SalGI - фрагмент плазмидной ДНК pNut размером 2659 н.п., содержащий участок плазмиды pUC18 и имеющий ген β- -лактамазы (Amp);
- BglII/SmaI-EcoRI - фрагмент плазмидной ДНК pNut размером 600 н.п., содержащий сигнал сплайсинга и сайт полиаденилирования;
- KpnI-HindIII - фрагмент плазмидной ДНК pNut размером 324 н.п., содержащий сайт инициализации репликации (ori) и промотор ранних генов вируса SV40;
- KpnI-SmaI/BstEII - фрагмент плазмидной ДНК pNut размером 650 н.п., содержащий промотор металлотионеинового гена мыши (MT-1);
- HindIII-SalGI - фрагмент плазмидной ДНК pNut размером 1500 н.п., содержащий кДНК DHFR и сайт полиаденилирования.
Фиг. 2. Кариотип культуры. Дифференцированная G-окраска хромосом клеток CHOpE-9.
Ниже следуют примеры осуществления предлагаемого изобретения.
Пример 1. Конструирование рекомбинантной плазмиды pKEP-9.
В качестве источника генетических последовательностей, кодирующих ЭПО человека, используют плазмиду pSV-Ep-gpt (фиг. 1), которая содержит BstEII-BglII-фрагмент ДНК ЭПО человека длиной 2386 н.п. [5]. Клетки бактерий E. coli HB101, трансформированные плазмидой pSV-Ep-gpt, и клетки бактерий E. coli HB101, трансформированные плазмидой pNut [8], выращивают в одном литре среды LB [9] с ампициллином в концентрации 50 мкг/мл в течение 20 ч. Клетки осаждают центрифугированием (5000 g, 5 мин, 4oC) и выделяют из них плазмидные ДНК, как описано в работе [10].
Полученные препараты ДНК плазмид pSV-Ep-gpt и pNut используют для конструирования плазмиды pKEP (схема на фиг. 1), которое проводят следующим образом. Осуществляют гидролиз 60 мкг ДНК pSV-Ep-gpt эндонуклеазами рестрикции BstEII и BglII (этап 1 на фиг. 1) в буфере, содержащем 0,01M Трис-HСl (pH 8,0); 0,01M MgCl2, 0,05M NaCl, 1 mM β-меркаптоэтанол. Аналогичным образом проводят гидролиз 20 мкг ДНК pNut рестриктазой SmaI (этап 2 на фиг. 1). Гидролизаты плазмид подвергают электрофорезу в 4%-ном ПААГ, гель прокрашивают бромистым этидием и под длинноволновым УФО вырезают полосу, соответствующую линейной ДНК, для SmaI гидролизата pNut и полосу, соответствующую фрагменту 2383 н.п., для BstEII-BglII гидролизата pSV-Ep-gpt. Фрагменты ДНК выделяют из кусочков геля электроэлюцией на бумажные диски DE-81 (Whatman, США) в 0,01M трис-боратном буфере, pH 8,3, при 350 V в течение 2-3 ч. ДНК с дисков смывают 1,5M NaCl; 0,01M Трис-HCl (pH 7,5); 1мМ ЭДТА (420 мкл) в течение 40 сек при 56oC, осаждают этанолом. Осадки промывают спиртом, высушивают, растворяют в 10 мкл воды. Выступающие 5' концы выделенного фрагмента BstEII-BglII достраивают до тупых в ходе полимеразной реакции с помощью фрагмента Кленова ДНК-полимеразы I E.coli [11]. После достройки 1 пмоль фрагмента смешивают с 0,2 пмолями SmaI-линеаризованной и дефосфорилированной векторной ДНК pNut в 30 мкл раствора, содержащего 66 мМ Трис-Hcl (pH 7,5); 5 мМ MgCl2; 5мМ дитиотреит (ДТТ); 1мМ АТФ; 14 ед.акт. ДНК-лигазы фага Т4. Реакцию проводят при 80oC в течение 20 ч. Аликвоты лигазной смеси используют для трансформации компетентных клеток E.coli HB101, как описано в работе [12] . Трансформированные клетки высевают на чашки с твердым агаром, содержащим ампициллин (50 мкг/мл), и растят 18 ч при 37oC. Выросшие колонии скалывают в 5 мл среды LB, содержащей ампициллин (50 мкг/мл), инкубируют при 37oC и интенсивном встряхивании в течение 18 ч. Клетки отделяют центрифугированием, из них выделяют плазмидную ДНК для последующего рестрикционного анализа. Полученную плазмиду называют pKEP-9 (фиг. 1).
Пример 2. Получение штамма-продуцента CHOpE-9.
Для получения стабильного продуцента рекомбинантного ЭПО человека проводят трансформацию клеток яичника китайского хомячка CHO-K1 dhfr - плазмидой pKEP-9. Клетки культивируют в среде F-12 (90%), с добавлением 10% эмбриональной телячьей сыворотки (ЭТС). В культуральные флаконы площадью 25 см2 засевают 5 мл клеточной суспензии (1,5 млн клеток) и через 16-20 ч стандартным методом кальций-фосфатной преципитации проводят трансформацию [13]. Плазмидную ДНК добавляют к клеткам в виде кальций-фосфатного преципитата. Преципитат готовят следующим образом: при комнатной температуре во флакон A вносят 30 мкг ДНК плазмиды в 300 мкл буфера TE (1мМ Трис-HCl; 0,1 мМ ЭДТА; pH 9,0), добавляют 188 мкл 2М CaCl2 и воды до 1500 мкл. Во флакон B вносят 1500 мкл буфера HBS (0,3М NaCl; 50 mM Hepes; 2,8 mM Na2HPO4, pH 7,1). Содержимое флакона A по каплям добавляют к содержимому флакона B и оставляют на 30-40 мин при комнатной температуре до образования полупрозрачного преципитата. Из флаконов с клеточной культурой CHO-K1 dhfr - сливают среду, на монослой наносят 3 мл преципитата и инкубируют 45-60 мин при 37oC, затем добавляют 10 мл среды F-12, содержащей 2% ЭТС. Через 16 ч после трансформации клетки отмывают и помещают в ростовую среду на 24 ч. Далее клетки снимают с флакона смесью трипсин-версена и рассевают в концентрации 100 клеток в мл на две чашки Петри диаметром 90 мм в селективной среде a-МЕМ (без тимидина и гипоксантина), содержащей 10% ЭТС и 500 нМ метотрексата. Клетки инкубируют в селективной среде в течение двух недель, смену среды проводят каждые трое суток. Через 14 дней после помещения в селективные условия формируются колонии, устойчивые к метотрексату. Колонии выделяют, рассеивают в лунки 48-луночных пластиковых планшетов в среде a-МЕМ, содержащей 10% ЭТС и 500 нМ метотрексата и инкубируют при температуре 37oC, 98% влажности, в атмосфере 5% CO2 до формирования монослоя. Методом гибридизации определяют, клетки каких клонов содержат в своем геноме ДНК плазмиды pKEP-9 [14].
Наличие эритропоэтина в культуральной жидкости определяют по включению in vitro тритий-меченного тимидина в растущие спленоциты мышей с фенилгидразиновой анемией [15] . Анализ проводят следующим образом. Мышам линии C3H/He весом 20-30 г вводят раствор фенилгидразина из расчета 60 мг на кг веса, двукратно через сутки. На третий день после последней инъекции стерильно извлекают селезенки и готовят суспензию спленоцитов с концентрацией 4 млн клеток/мл в среде RPMI 1640, содержащей 20% ЭТС. В лунки плоскодонного 96-луночного планшета вносят по 50 мкл клеточной суспензии, добавляют равный объем разведений тестируемого материала в культуральной среде. В качестве положительного контроля используют разведения стандартного образца ЭПО с активностью 250 ед. акт./мл производства Boehringer Mannheim (Германия). В качестве отрицательного стандарта служит среда культивирования. Планшет инкубируют в течение 22 ч при 37oC во влажной атмосфере, содержащей 5% CO2. Далее в каждую лунку вносят по 1 мкКи [3H]-тимидина в 20 мкл среды RPMI 1640. Инкубацию продолжают в тех же условиях в течение 2 ч. Содержимое лунок переносят на стекловолоконные фильтры GF/C (Whatman, США) с помощью харвестера Titertek (Flow, США), промывают несколько раз водой. Фильтры высушивают на воздухе и просчитывают в толуольном сцинтилляторе на счетчике радиоактивности MarkIII (Beckman, США). Биологическую активность тестируемых проб оценивают путем сравнения с калибровочной кривой ЭПО-стандарта.
Таким образом, отбирают клон CHOpE-9, клетки которого стабильно продуцируют эритропоэтин в культуральную жидкость с выходом до 400 ед.акт. (4 мкг) на мл среды при стационарном культивировании.
Пример 3. Выделение и характеризация рекомбинантного эритропоэтина.
Культуральную жидкость после выращивания клеток-продуцентов CHOpE-9 в объеме 3000 мл фильтруют через бумажный фильтр, проводят последовательное концентрирование до объема 50-60 мл и диафильтрацию против 500-600 мл раствора 15 мМ трис-HCl, pH 7,0, содержащего 0,2 мМ сульфата меди и 0,02% твин-20 (буфер A) с использованием разделительного ультрафильтрационного аппарата на полых волокнах АР-0.2н производства ОКБ ТБМ г. Кириши. Полученный концентрат культуральной жидкости наносят на колонку с 50 мл ДЕАЕ-сефарозы, уравновешенной буфером Б (15 мМ трис-HCl, pH 7,0, 0,02% твин-20). Колонку промывают последовательно 50 мл буфера А; 150 мл раствора 6М мочевины, подкисленной уксусной кислотой до pH 4,5; 100 мл раствора 25 мМ NaCl в буфере В; 100 мл раствора 80 мМ NaCl в буфере В; 100 мл раствора 160 мМ NaCl в буфере В со скоростью 4 мл/мин. Фракции элюата собирают по оптической плотности, определяемой при длине волны 254 нм с помощью проточного УФ-денситометра и определяют их активность методом иммуноферментного анализа (ИФА) с использованием ЭПО-специфической поликлональной кроличьей антисыворотки и конъюгата антивидовых антител (коза-антикролик) с пероксидазой хрена. Фракции, обладающие активностью в ИФА, объединяют и наносят на колонку 8 x 150 мм с обращеннофазовым широкопористым (диаметр пор 30 нм) сорбентом, содержащим бутильные группы, уравновешенную 5%-ным раствором ацетонитрила в 0,1%-ной трифторуксусной кислоте. Элюцию белков с колонки проводят линейным градиентом 5-90% ацетонитрила в течение 60 мин со скоростью 5 мл/мин. Фракции собирают, анализируют на наличие ЭПО-активности, как описано выше, объединяют и концентрируют в три раза упариванием на ротационном испарителе Buchi (Швейцария). Полученный образец рехроматографируют на приведенной выше колонке с обращеннофазовым сорбентом. Фракции, элюирующиеся с колонки в 48-55%-ном ацетонитриле и обладающие ЭПО-активностью, объединяют, упаривают на ротационном испарителе при 30oC, растворяют в 0,02М цитрате натрия, pH 6,9, содержащем 0,1М NaCl. Полученные образцы рекомбинантного эритропоэтина характеризуются следующими показателями:
1. Чистота и гомогенность.
- гомогенность препарата не менее 90-95% (по данным гель-электрофореза в 13%-ном полиакриламидном геле с денситометрией);
- отношение ОП280/ОП260 = 1,05;
- содержание агрегированной формы - до 4,5% (по данным иммуноблоттинга и денситометрии электрофорегамм, окрашенных серебром);
- содержание примесей ДНК клеток-продуцентов не более 10 пг на 10000 ед. активности (по данным дот-блот гибридизации с использованием меченной радиоактивным фосфором суммарной ДНК клеток CHO-9).
2. Идентичность.
- молекулярная масса - 37500 Да (по данным гель-электрофореза с белковыми маркерами молекулярных весов);
- удельная активность - 150000 ед/мг (10000 ед/мл) (определена методом твердофазного ИФА);
- специфическая активность in vitro - 200000 ед/мг (определена по включению [3H] -тимидина в растущие спленоциты мышей с фенилгидразиновой анемией по [15]);
- специфическая активность in vivo - не менее 100000 ед/мг (определена путем подсчета числа ретикулоцитов в образцах крови мышей после введения им образцов ЭПО по [16]);
- аминокислотный состав соответствует расчетным данным (определен после гидролиза 6 н. HCl, 110oC, 24 ч);
- N-концевая последовательность A-P-P-R-L-I-C-D-S-R 10 аминокислот соответствует литературным данным (определена методом Эдмана на автоматическом секвенаторе с идентификацией ФТГ-аминокислот методом ВЭЖХ).
Список литературы
1. Lin F.K., Suggs S., Lin C.H., Browne J.K., Smalling R., Egrie J.C., Chen K. K. , Fox G. M. , Martin F., Stabinsky Z., Badrawi S.M., Lai P.H., Goldwasser E. Proc.Natl.Acad.Sci.USA. 1985. V.82, N 22. P. 7580-7584.
2. Патент США N 4703008. Последовательность ДНК, кодирующая эритропоэтин.
3. Urlaub G., Chasin L.A. Proc.Natl.Acad.Sci.USA. 1980. V. 77, N 7. P. 4216-4220.
4. Международная заявка N 86/03520. Способ получения эритропоэтина.
5. Заявка России N 93029235/13 от 08.06.93. Рекомбинантная плазмидная ДНК pSV-dEp-poly-Neo, кодирующая эритропоэтин человека, штамм культивируемых клеток яичника китайского хомячка CHO-PE - продуцент эритропоэтина человека. Решение о выдаче патента от 28.03.95. (Патент РФ N 2070931, Бюл.Из. 9636, 27.12.96).
6. Mulligan R.C., Berg P. Proc.Natl.Acad.Sci.USA. 1981. V. 48, N 4. P. 2072-2076.
7. Richardson K.K., Fostel J., Skopek T.R. Nucl.Acids Res. 1983. V. 11, N 24. P. 8809-8816.
8. Palmiter R.D., Behringer R.R., Quaife C.J., Maxwell F., Maxwell I.F., Brinster R.L., (1987) Cell 50, P. 435-443.
9. Миллер Дж. Эксперименты в молекулярной генетике. Пер. с англ. - М.: Мир, 1976, с. 395.
10. Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии: Молекулярное клонирование. Пер. с англ. - М.: Мир, 1984, с. 98-103.
11. Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии: Молекулярное клонирование. Пер. с англ. - М.: Мир. 1984. С. 123-124.
12. Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии: Молекулярное клонирование. Пер. с англ. - М.: Мир. 1984. С. 240-241.
13. Горман К. Высокоэффективный перенос генов в клетки млекопитающих. В кн. : Клонирование ДНК. Методы. Под ред. Д. Гловера: Пер. с англ. - М.: Мир. 1988. С. 409-463.
14. Ротстейн Р. Клонирование в дрожжах. В кн.: Клонирование ДНК. Методы. Под ред. Д. Гловера: Пер. с англ. - М.: Мир. 1988. С. 299.
15. Krystal G. Exp.Hematol. 1983. V. 11, N 7. P. 649-660.
16. Goldwasser E. , Gross M. Methods Enzymol. 1975. V. 37, pt.B, р. 109-121.
Изобретение относится к области биотехнологии. Путем встраивания Bst EII - BglII фрагмента (2383 н. п.) плазмиды pSV - Ep - gpt, включающего структурный ген эритропоэтина (ЭПО) человека, в Sma - I сайт векторной плазмиды pNut (5733 н.п.), содержащей активируемый металлотионеиновый промотор мыши МТ - I и ген дигидрофолатредуктазы под контролем раннего промотора SV-40, получают рекомбинантный плазмидный вектор pКЕР-9, имеющий размер 8116 н.п. (мол. м. 5 Мд) и обеспечивающий биосинтез ЭПО человека в культивируемых клетках яичника китайского хомячка. В результате трансформации клеток СНО-КIdhfr сконструированной рекомбинантной плазмидой pКЕР-9, последующего культивирования на ростовой среде и отбора колоний, устойчивых к метотрексату и гибридизующихся с ДНК pКЕР-9, получен рекомбинантный штамм СНОpЕ-9. Клетки нового штамма при стационарном культивировании стабильно продуцируют в культуральную жидкость ЭПО с выходом 400 ед. акт./мл среды. 2 с.п. ф-лы, 2 ил.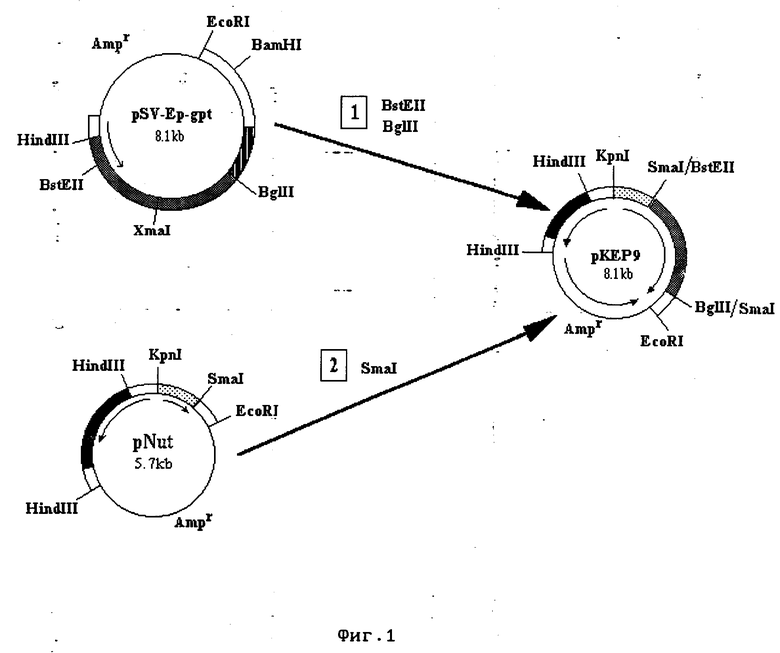
| RU, патент, 2070931, C 12 N 15/12, 1996. |

