Настоящее изобретение относится к протеину: структурам фосфолипида, которые используют для стабилизации вторичной и третичной структуры протеинов, способных к переходу в распрямленное состояние глобулы. В особенности это изобретение относится к G-CSF:фософолипидным композициям с повышенной стабильностью и способностью к использованию в G-CSF рецептурах и новых переносчиках G-CSF.
Было обнаружено несколько видов протеинов, способных к переходу глобулы в распрямленное состояние (MGS) (Van der Goot, F.G., Nfture 354, 408-410 (1991)). Протеины в распрямленном состоянии глобулы проявляют вторичную структуру, сравнимую со структурой природного протеина, однако у них отсутствует жесткая третичная структура (Pitsyn et al., FEBS Letters 262:1, 20-24 (1990)). В некоторых случаях переход в это состояние сопровождается раскрытием ранее скрытых гидрофобных звеньев протеина. При раскрытии критических гидрофобных остатков, MGS может находиться в промежуточном состоянии между агрегированием и осаждением протеинов. MGS конформация может быть обнаружена при сравнении кругового дихроизма в дальнем УФ диапазоне со спектром ароматических боковых цепей (ближний УФ круговой дихроизм и флуоресценция). Распрямленное состояние глобулы показывает спектральные изменения ароматических групп в отсутствие изменений дальнего УФ кругового дихроизма (Bychkova et al.FEBS Letters 238: 231-234 (1988)) и может быть вовлеченным в мембранную проницаемость некоторыми видами протеинов (Bychkova et al., FEBS Letters 238: 231-234 (1988); Van der Goot, F. G., Nature 354, 408-410 (1991)).
Для перехода в MGS до агрегирования известны два вида протеинов: гранулоцит колониестимулирующий фактор (G-CSF) и гранулоцит макрофаг колониестимулирующий фактор (GM-CSF). В то время, как два вида протеинов могут быть стабилизированы при некоторых определенных условиях, существует необходимость продления жизни этих материалов стабилизацией вторичной и третичной структуры протеинов.
Человеческий рекомбинантный G-CSF селективно стимулирует нейтрофилы, вид белых кровяных клеток, используемых для борьбы с инфекцией. Общеизвестно, Filgrastim, что рекомбинантный G-CSF является пригодным для использования в терапии. Структура G-CSF в различных условиях подверглась интенсивному изучению (Lu et al. , J.Biol. Chem. Vol. 267, 8770-8777 (1992)). Ввиду гидрофобного характера трудно получить G-CSF с продолжительной жизнеспособностью. Рецептуры некоторых гидрофобных протеинов теряют свою активность в результате образования димера или агрегатов более высокого порядка (макроагрегатов) в процессе продолжительного хранения. Другие химические превращения, такие, как деамидация и окисление, также могут возникнуть в процессе хранения. Кроме того, G-CSF должен защищать против денатурации и, в особенности, быть склонным к стабилизации вторичной и третичной структуры протеина.
Человеческий GM-CSF является 22-kDa гликопротеином, непрерывно требуемым для быстрого размножения in vitro клеток-предшественников макрофагов и гранулоцитов. Он также контролирует необратимое превращение этих клеток-предшественников при образовании гранулоцитов и макрофагов. Другие виды биологической активности могут включать регулирование функциональной активности зрелых клеточных видов (Gough et al., Nature, 309, 763-767 (1984)) и увеличение химотаксиса по отношению к опознанным химоаттрактантам (Williаms et al., Hematology, 4th ed (1990)). GM-CSF также стимулирует образование моноцитов и поэтому может быть использован при лечении моноцитных расстройств, таких как моноцитопения.
Человеческий GM-CSF может быть получен и очищен из нескольких источников. Способы получения рекомбинантного человеческого GM-CSF были описаны ранее Burgess et al., Biood, 69:1, 43-51 (1987). Согласно цитируемому здесь патенту US 5047504 (Boone) предлагается получение в промышленном масштабе GM-CSF в негликосилированной форме как продукт прокариотной клетки-хозяина.
Одним из путей, предпринятым ранее при работе с такими протеинами, как G-CSF и GM-CSF, явилось использование липосом. Липосомы представляют собой полностью закрытые липидные двухслойные мембраны, образованные водонерастворимыми полярными липидами, особенно фосфолипидами.
Пузырьки липосом могут иметь один единственный мембранный двойной слой (однослойные) или множество мембранных двойных слоев (многослойные). Двойной слой состоит из двух липидных монослоев, имеющих гидрофильную (полярную) "головную" зону и гидрофобную (неполярную) "хвостовую" зону, в которой гидрофобные хвосты ориентированы по направлению к центру двойного слоя, в то время как гидрофильные головы ориентированы по направлению к водной фазе. Стабильность, жесткость и проницаемость липосом может изменяться путем изменений фосфолипидной композиции или температуры, включением стирола или введением заряженных амфифил. Основная структура липосом может быть получена различными известными методами.
В процессе своего образования липосомы могут захватывать растворенные в воде вещества в водных каналах и освобождать их с различной скоростью. После обнаружения способности липосом к введению энзимов в клетки и к изменению их метаболизма (Gregoriadis, New Enfl. J. Med. 295, 704-710, 765-770 (1976)) липосомы были объявлены средством, отвечающим требованию целенаправленной доставки медикаментов. В результате наблюдается усиленное развитие исследований в фармацевтической индустрии, включающих использование липосом в качестве медленных поставщиков лекарств, витаминов и протеинов, секвитированных в гидрофобном ядре липосом.
Успешное использование липосом в качестве переносчиков медикаментов было ограничено, т.к. исследователи, предпринимающие попытки в направлении такого применения липосом, встретились с рядом проблем. Например, известна способность липосом действовать в качестве сильных иммунологических помощников по отношению к захваченным антигенам, и должны быть приняты меры предосторожности в том случае, когда липосомами захвачены энзимы или протеины ксеногенного происхождения. Кроме того, трудно контролировать скорость диффузии медикамента. Это является следствием природной нестабильности липосом и присутствия некоторых компонентов крови, которые ускоряют диффузию некоторых медикаментов. Помимо этого, по своей природе некоторые вещества слабо захватываются липосомами и поэтому быстро рассасываются при кровообращении. Наконец, существует проблема нацеливания на любые клетки или органы, кроме печени или селезенки. Замечательным обзором по липосомам, веществам, которые были введены в липосомы, и проблемам, связанным с использованием липосом в качестве переносчиков медикаментов, является Gregory Gregoriadis, "Liposomes", Drug Carriers in Biology and Medicine, Chapter 14, 287-341 (Academic Press, N.Y., 1979).
В то время как в области попыток использования липосом в качестве переносчиков медикаментов опубликовано много работ, мало публикаций появилось в области использования липосом с целью увеличения периода жизни терапевтических пептидов или протеинов путем стабилизации структуры пептидов и/или протеинов. В PCT/US90/05163 "Терапевтические пептиды и протеины", Hostetler et al. описано использование пустых липосом в качестве фармацевтически пригодных разбавителей для растворения полипептидов и/или протеинов с целью предотвращения аккумуляции полипептидов и/или протеинов на поверхности раздела воздух/вода и для предотвращения адсорбции полипептидов и/или протеинов на поверхности контейнера. Hostetler et al. описывает, что отрицательно заряженные фосфолипиды могут быть добавлены в количестве до около 50 мол. процентов и что предпочтительными липосомами являются фосфатидилхолин, нейтральный фософолипид. Hostetler et al. не описывает разбавителя, способного стабилизировать структуру полипептида и/или протеина.
В PCT/US91/07694 "Получение и характеристика липосомных рецептур фактора некрозного новообразования (TNF)", Hung et al., описана липофильная молекула фактора некрозного новообразования, связанная с поверхностью или инкапсулированная внутри липосомы. Липосомные липофильные TNF молекулы описаны как имеющие повышенную стабильность in vivo. Стабильность рассматривается как уменьшение или как тенденция к уменьшению TNF-липосомы выделять TNF в систему in vivo. Предпочтительными липосомами были нейтральные липиды. Heing et al. не описывают композиции TNF, в которой эксипиенты оказывают стабилизирующее действие на структуру протеина.
В литературных источниках отсутствуют какие-либо сведения, касающиеся контактирования протеина, например G-CSF, с отрицательно заряженными пузырьками (полостями) липид, в результате которого непосредственно стабилизируется протеин против термической агрегации, денатурации, потери активности и развертывается вторичная структура. Существует потребность в таких композициях, которые могут быть использованы в технологических процессах, требующих высоких температур, а также в качестве новых проводников (например, оральное применение лекарств, закрепленных в G-CSF). Настоящее изобретение касается таких композиций.
Сущность изобретения.
Настоящее изобретение направлено на добавление гидрофобных эксипиентов, например лизофосфолипидов или других липосом, к протеину в условиях распрямленного состояния глобулы для стабилизации вторичной и третичной структуры протеина, защищая таким образом протеин против термической агрегации, денатурации и потери активности. В особенности целью изобретения являются стабильные G-CSF: фосфолипидные композиции. Неожиданно оказалось, что предпочтительные G-CSF композиции могут быть несколько раз подвергнуты циклическому изменению температуры в диапазоне 10 - 95oC с последующим полным восстановлением вторичной структуры протеина после охлаждения. Эти композиции имеют двойное преимущество: быть полезными при технологических процессах, требующих высоких температур, а также при использовании в новых проводниках G-CSF.
В предпочтительном варианте, комплекс протеин:фосфолипид включает отрицательно заряженную липосому, выбранную из группы:диолеилфосфатицилглицерол (DOPG), димиристоилфосфатицилглицерол (DMPG), дипальмитоилфосфатидилглицерол (DPPG), ячеистый фосфатидилдиглицерол, диолеоилфосфатидилэтаноламин (DOPE), ячеистый фосфатидилэтаноламин, диолеоилфосфатидовая кислота (DOPA), димиристоилфосфатидовая кислота (DMPA), дипальмитоилфосфатидовая кислота (DPPA), диолеилфосфатидилсерин (DOPS), димиристоилфосфатидилсерин (DMPS), дипальмитоилфосфатидилсерин (DPPS), яичный фосфатидилсерин, лизофосфатидилглицерол, лизофосфатидилэтаноламин, лизофосфатидилсерин.
DOPG, отрицательно заряженный ненасыщенный фосфолипид, является особенно предпочтительным. Согласно изобретению pH поддерживают в пределах 3,0 - 7,5, а отношение липид:протеин не менее 10:1.
Дополнительным элементом согласно предпочтительному варианту изобретения является использование химически модифицированных протеинов в составе комплекса протеин:фосфолипид, а также использование одного или нескольких из следующих агентов: агент, устанавливающий изотонию, буферный агент и агент, устанавливающий pH. Специалисту понятно, что изобретение включает стабильные композиции протеин:фосфалипид с различным сочетанием дополнительных элементов.
На фиг. 1 изображен спектр флуоресцентной эмиссии rhG-CSF в присутствии (кривая 1) и в отсутствии (кривая 2) пузырьков DOPG. Концентрация DOPG: rhG-CSF равна 0,2 мг/мл. Молярное отношение DOPG:rhG-CSF (кривая 1) равно 100:1.
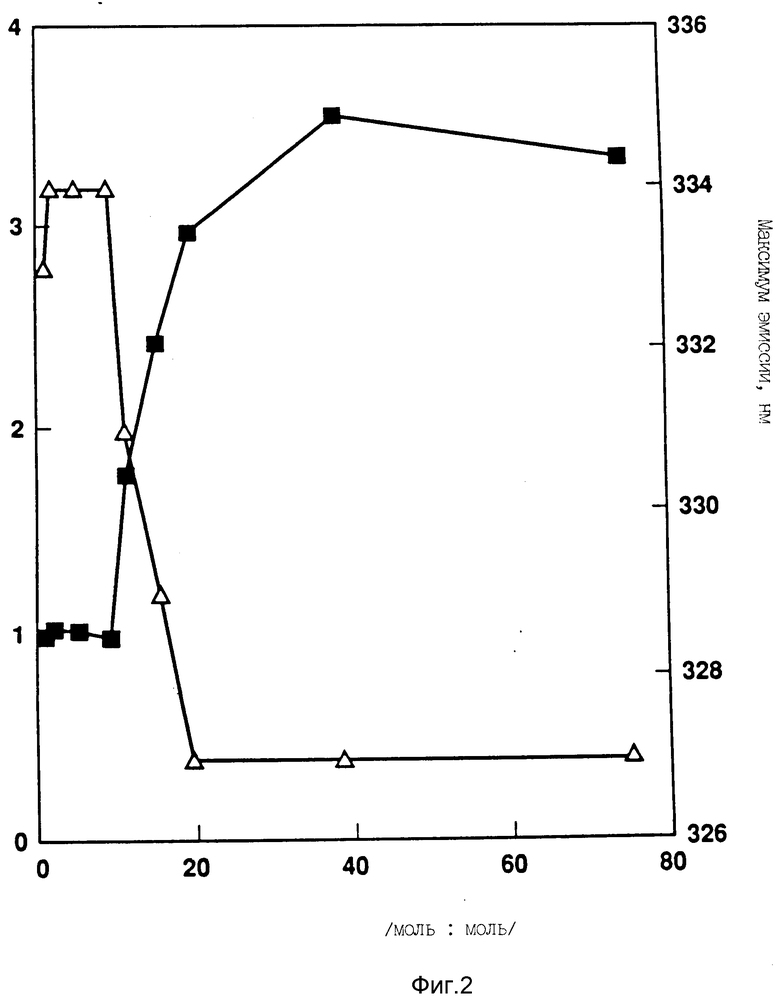
На фиг. 2 показано влияние увеличения отношения липид:протеин на rhG-CSF флуоресценцию. F0 означает исходную флуоресценцию (без липида) и f означает флуоресценцию после добавления липида для достижения указанного молярного отношения F/F0(n) и максимум длины волны эмиссии (Δ) для смесей DOPG:rhG-CSF.
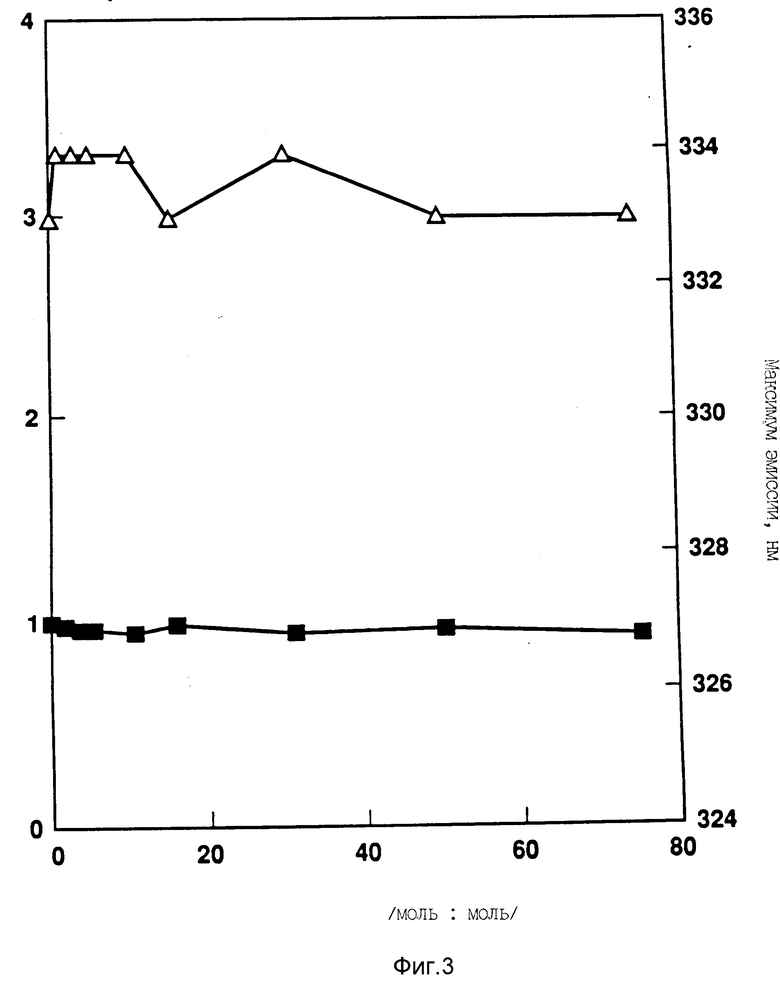
На фиг. 3 показано отношение F/F0(n) и максимум длины волны эмиссии (Δ) для смесей DOPG:rhG-CSF.
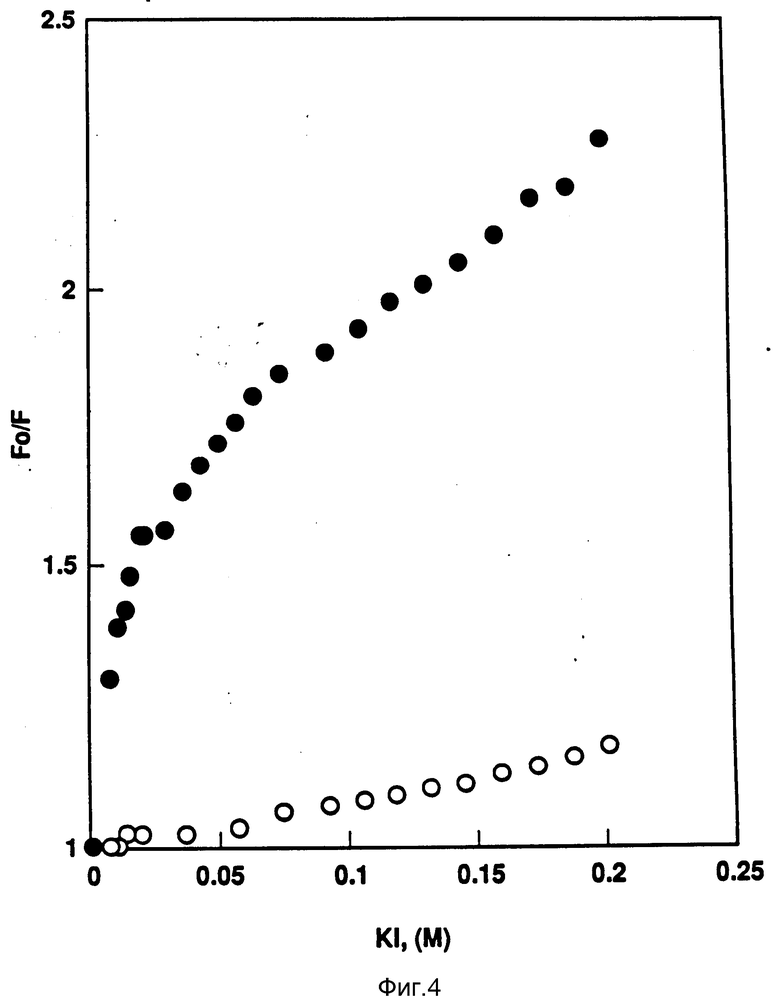
На фиг. 4 показана кривая Стерн-Волмера (Stern-Volmer) затухания rhG-CSF флуоресценции посредством KJ в отсутствии (•) и в присутствии (°) пузырьков DOPG. Эксперимент по затуханию проводился добавлением равных порций KJ к rhG-CSF (0,2 мг/мл) и DOPG:rhG-CSF 100:1.
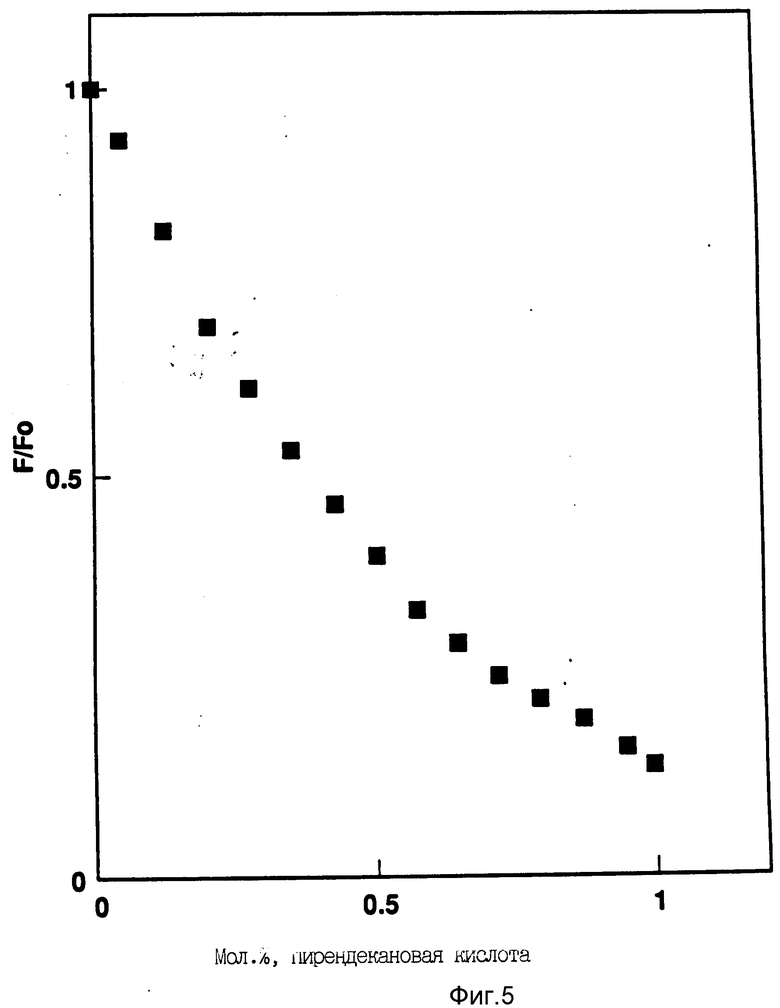
На фиг. 5 показано затухание rhG-CSF флуоресценции триптофана при добавлении пирендекановой кислоты (pyrene decanoic acid). Длина волны эмиссии равна 327 нм. Молярное отношение DOPG:rhG-CSF равно 100:1.
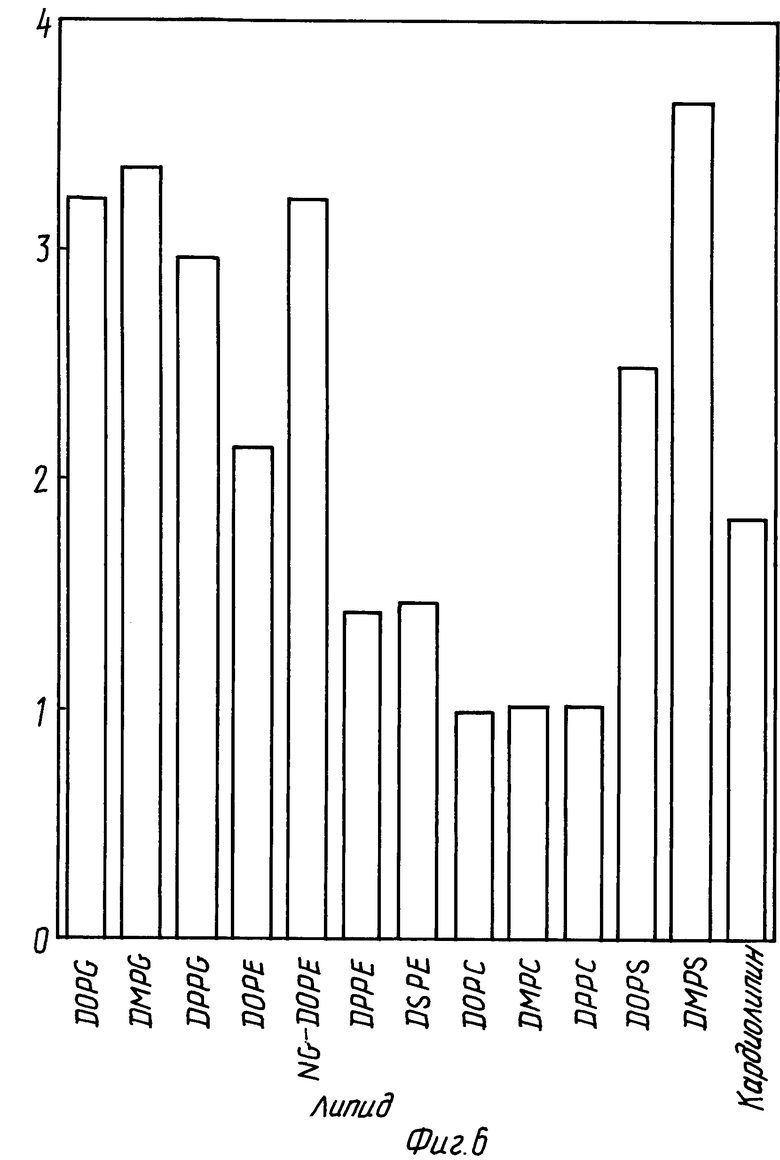
Фиг. 6 представляет собой диаграмму, показывающую в сравнении изменения F интенсивности для rhG-CSF в отсутствии и в присутствии различных липидов. В каждом случае молярное отношение липид:протеин равно 100:1.
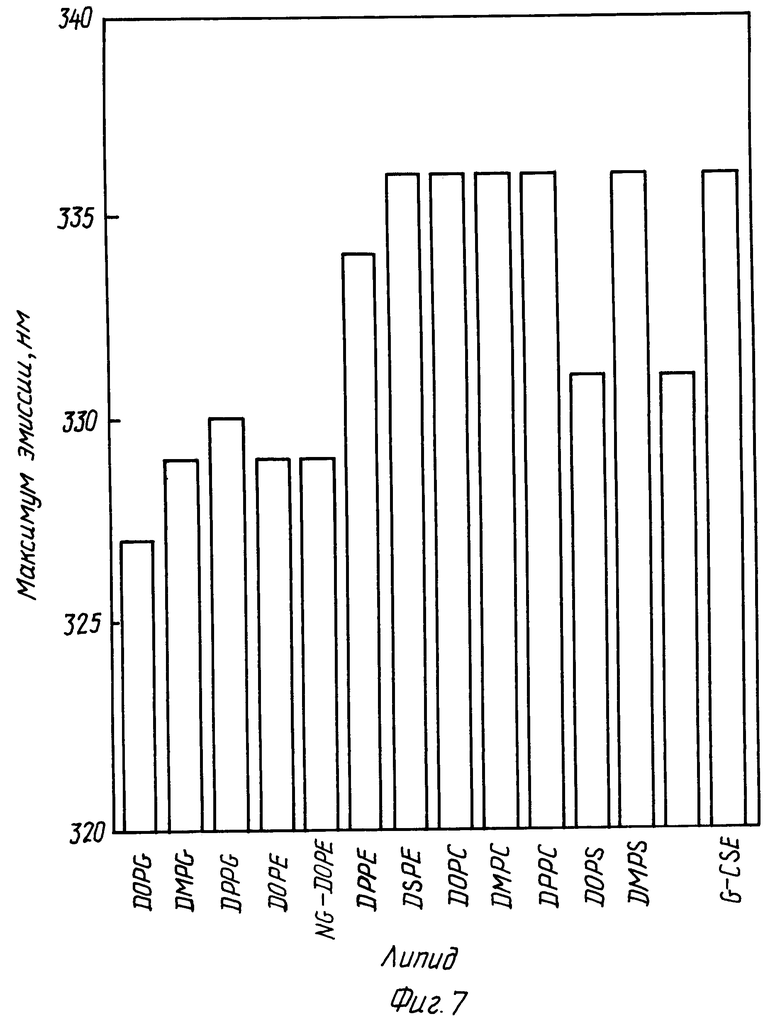
Фиг. 7 представляет собой диаграмму, показывающую в сравнении сдвиги максимумов эмиссии для rhG-CSF в отсутствии и в присутствии различных липидов. В каждом случае молярное отношение липид:протеин равно 100:1.
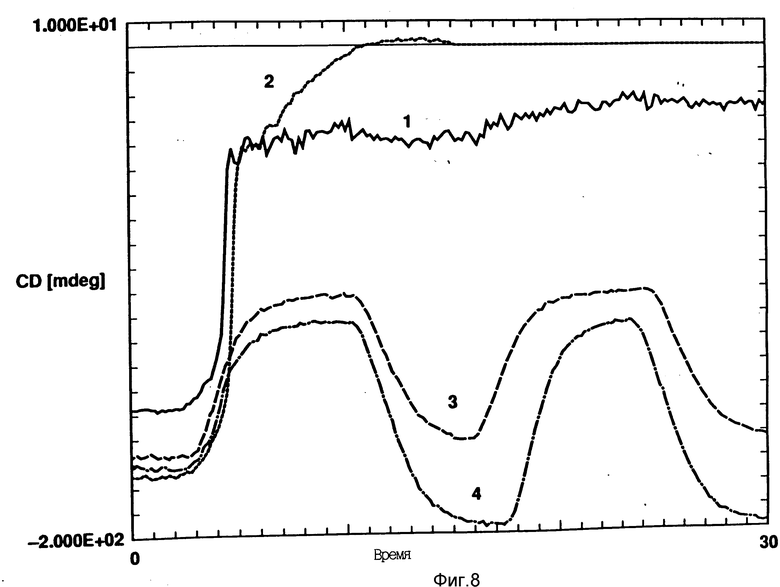
На фиг. 8 показано влияние DMPC (кривая 2), DMPG (кривая 3) и DMPA (кривая 4) на CD rhG-CSF (кривая 1). В каждом случае молярное отношение липид:протеин в воде равно 50:1, pH 6,0.
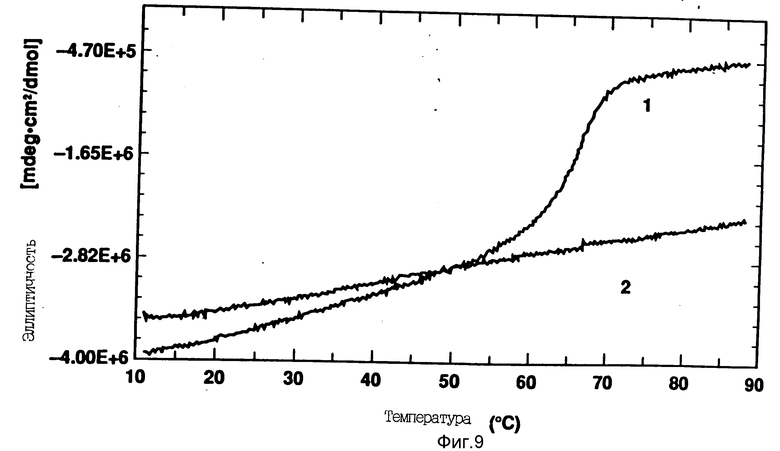
На фиг. 9 показано влияние повышения температуры на CD rhG-CSF (кривая 1) и DOPG:rhG-CSF (140:1 молярное) (кривая 2). Концентрация rhG-CSF в воде равна 80 мг/мл, pH 6,0. Температура изменялась в пределах 10 - 90oC со скоростью 100oC/ч.
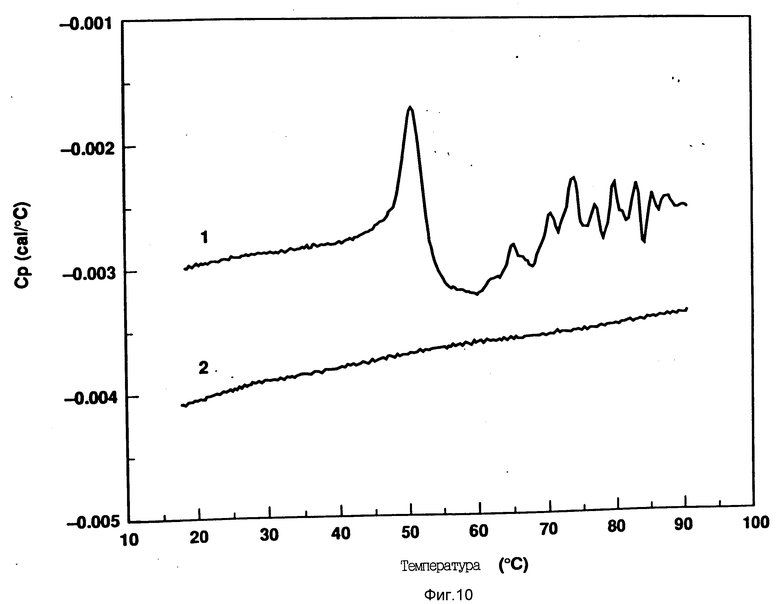
На фиг. 10 показана дифференциальная сканирующая калориметрическая термограмма для rhG-CSF (кривая 1) и DOPG:rhG-CSF (45:1 молярное) (кривая 2). Концентрация rhG-CSF в образцах равна 1 мг/мл, pH 7,0 в воде. Скорость сканирования 90oC/ч.
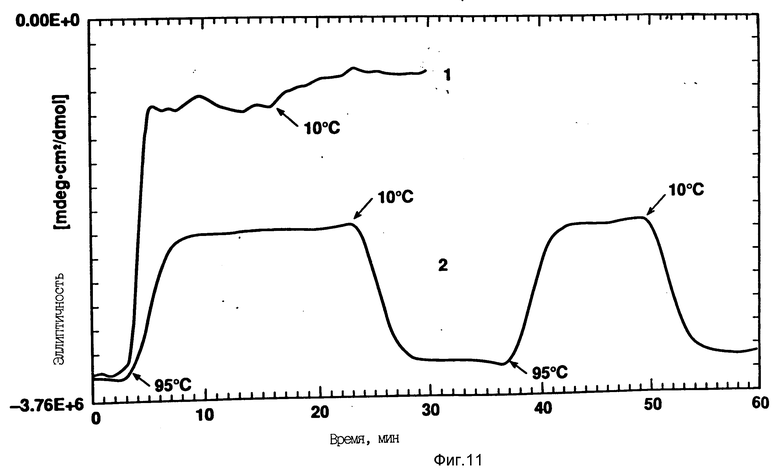
На фиг. 11 показано влияние циклического изменения температуры на CD rhG-CSF (кривая 1) и DOPG: rhG-CSF (140:1 молярное) (кривая 2). Образцы быстро нагревали до 95oC и охлаждали до 10oC, как показано стрелками. Концентрация rhG-CSF в образцах равна 80 мг/мл, pH 6,0.
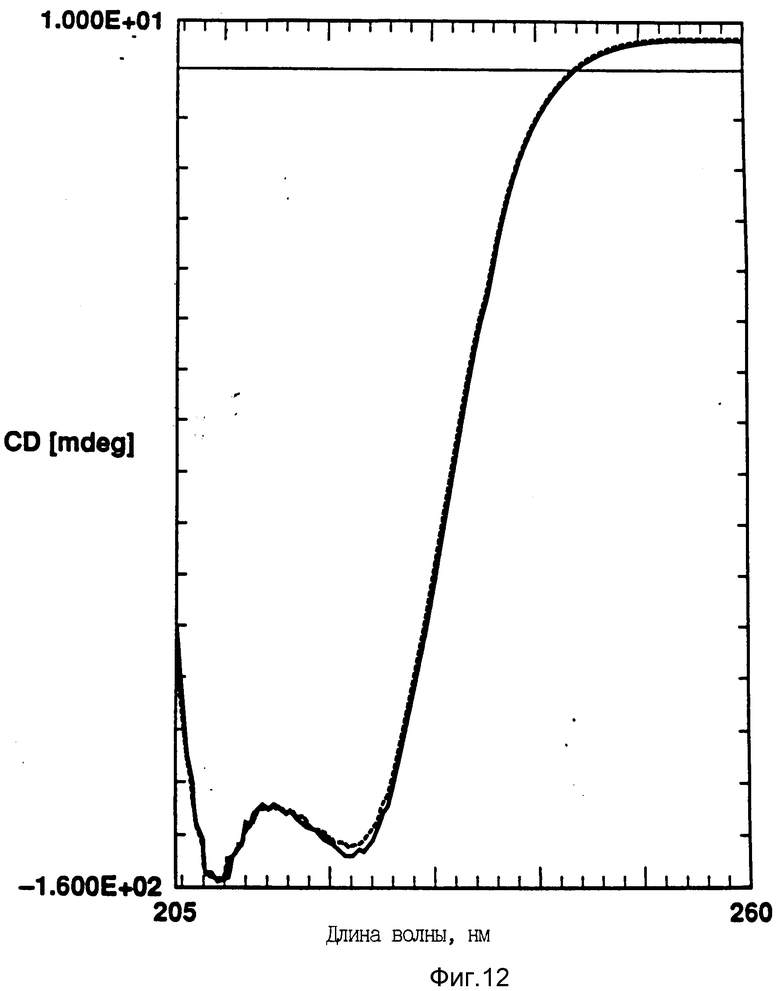
На фиг. 12 показано влияние циклического изменения температуры на CD rhG-CSF (кривая 1) и DMPG:rhG-CSF (150:1 молярное) (кривая 2). Образцы нагревают до 95oC и охлаждают до 10oC. Концентрация rhG-CSF в образцах равна 80 мг/мл, pH 6,0.
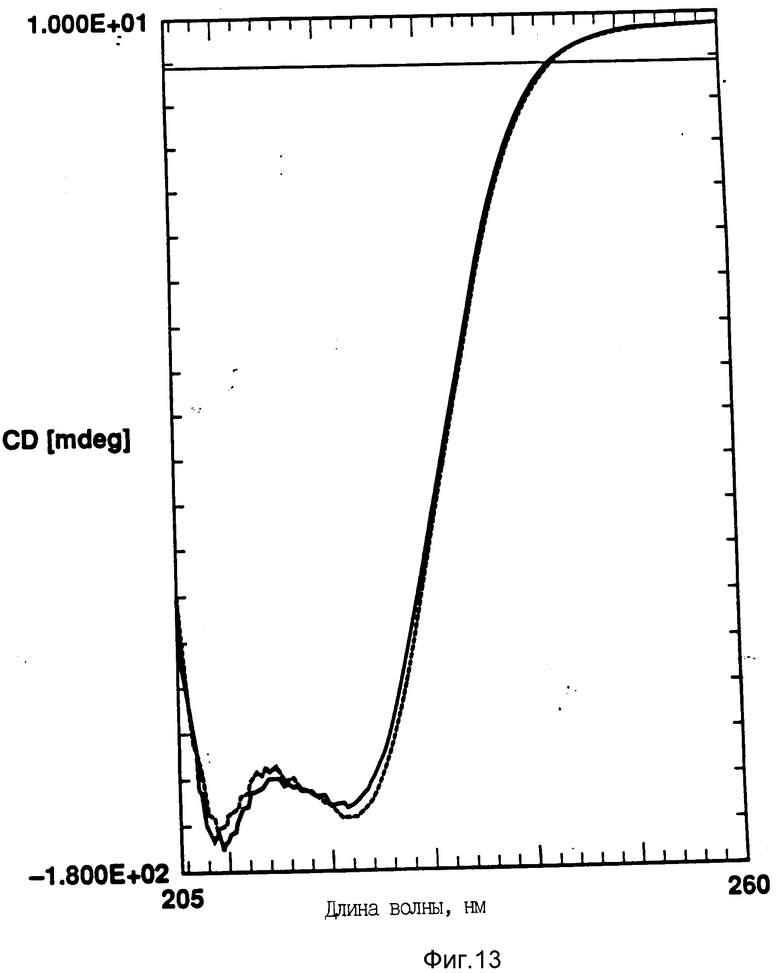
На фиг. 13 показано влияние температурных циклов на CD rhG-CSF (кривая 1) и DPPG:rhG-CSF (150:1) (кривая 2). Образцы нагревают до 95oC и охлаждают до 10oC. Концентрация rhG-CSF в образцах равна 80 мг/мл, pH 6,0.
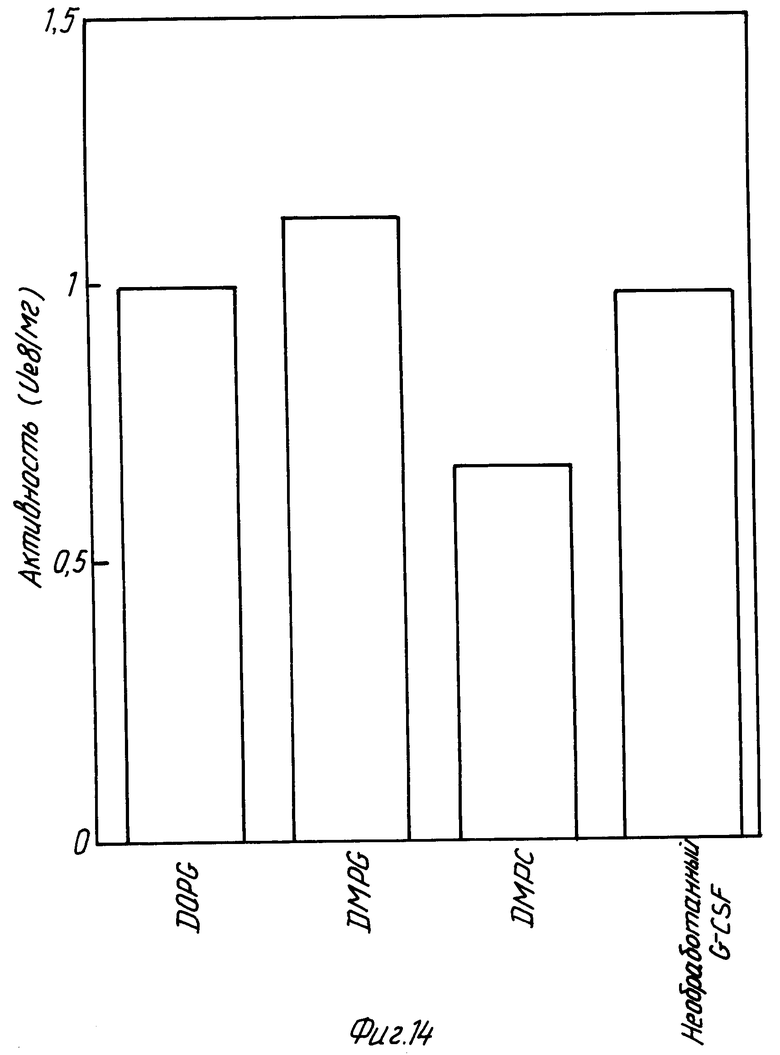
На фиг. 14 изображает диаграмму, показывающую способность различных липидов к стабилизации rhG-CSF в процессе замораживание-сушка. Отношение липид: протеин в каждом случае равно 100:1. Стабильность определялась по сохранению активности in vitro в образце костного мозга. Один rhG-CSF не выдерживает процесса замораживание-сушка, поэтому контрольным образцом является необработанный rhG-CSF в отсутствии липида.
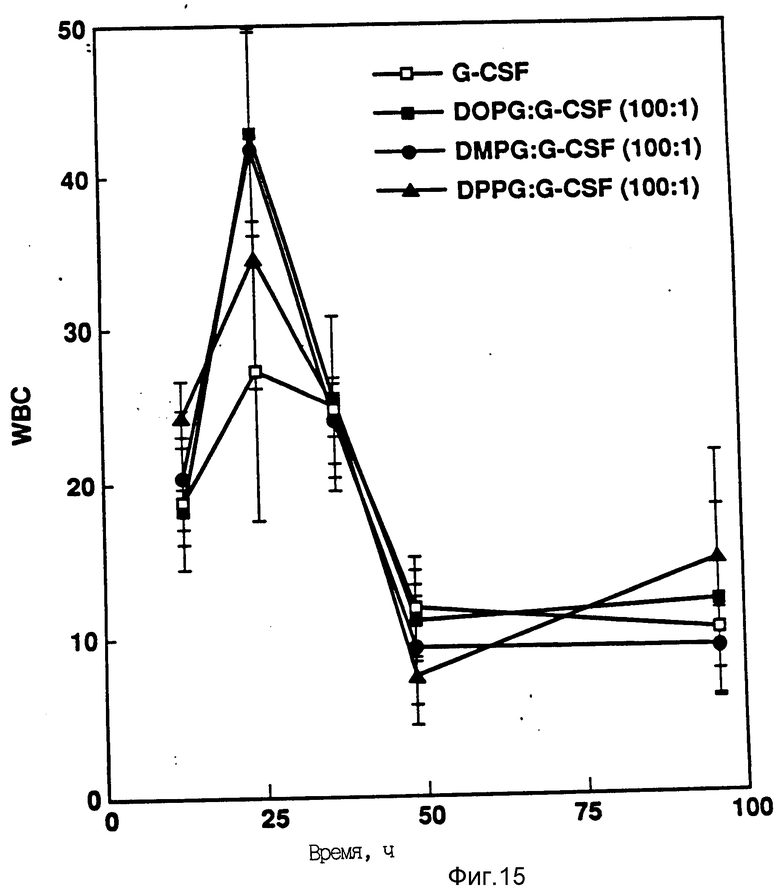
На фиг. 15 показано влияние различных липидов на активность in vitro rhG-CSF. Активность (в расчете на WBC) измеряли после подкожной инъекции хомячков. Доза rhG-CSF равна 100 мг/кг при отношении липид:протеин 100:1.
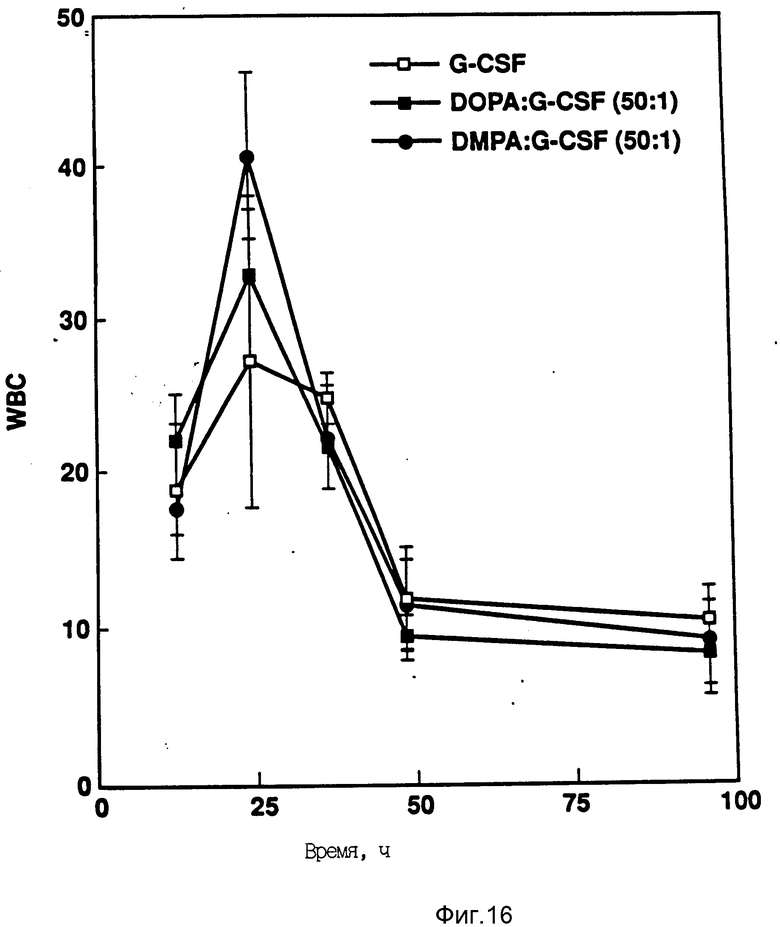
На фиг. 16 показано влияние различных липидов на активность in vitro rhG-CSF. Активность (в расчете на WBC) измеряли после подкожной инъекции хомячков. Доза rhG-CSF равна 100 мг/кг при отношении липид:протеин 50:1.
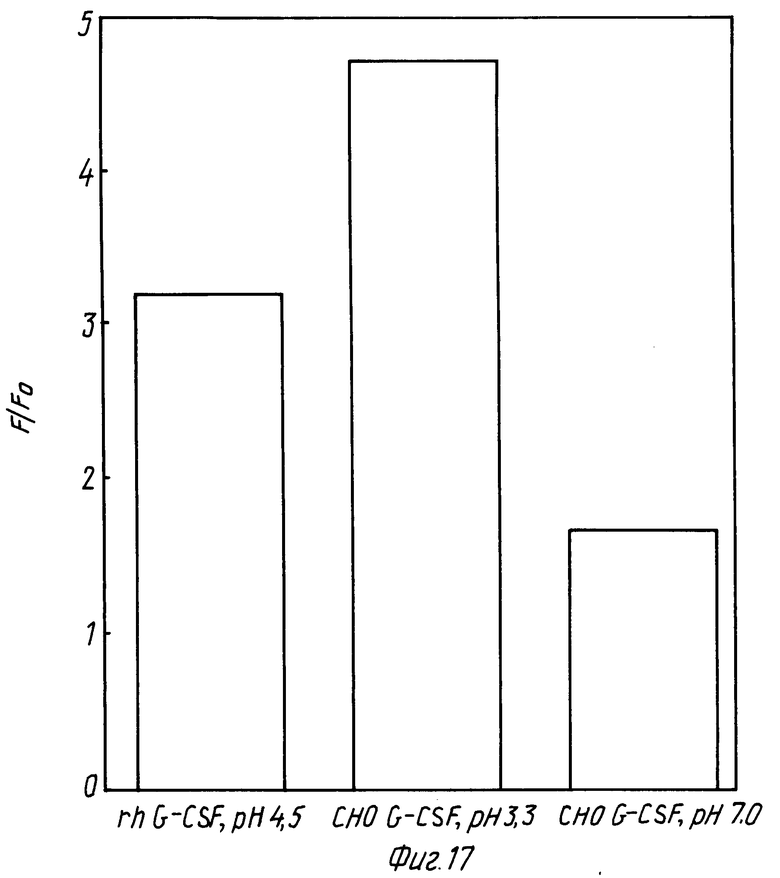
На фиг. 17 изображена диаграмма, показывающая сравнительное изменение CHO-G-CSF в отсутствии и в присутствии DOPG при различном pH. В каждом случае молярное отношение липид:протеин равно 100:1.
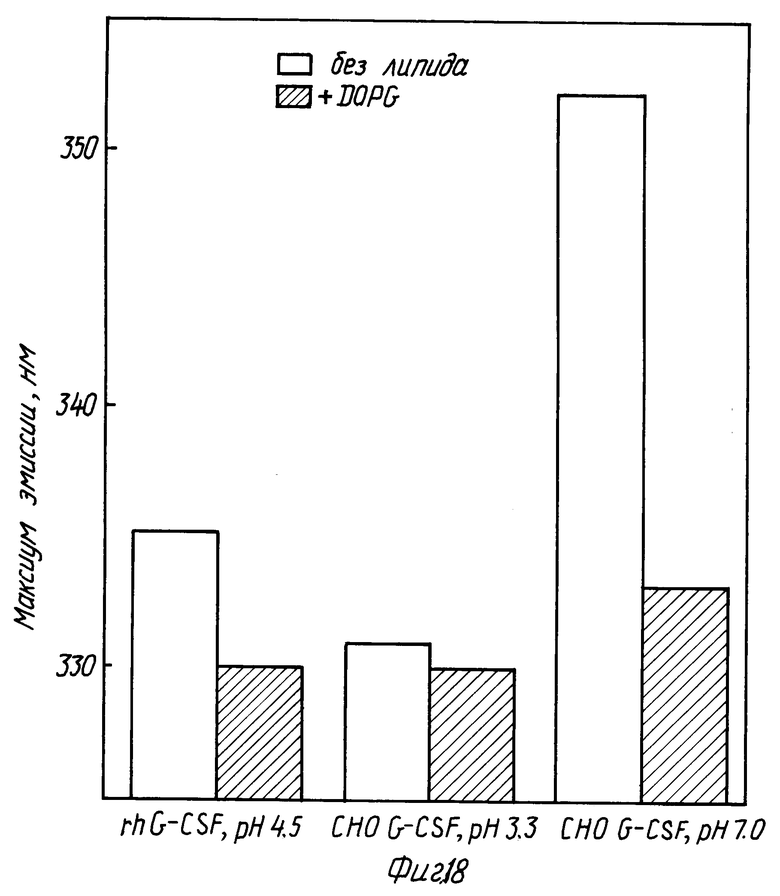
На фиг. 18 изображена диаграмма, показывающая сравнение сдвигов максимумов эмиссии для CHO-G-CSF в отсутствии и в присутствии DOPG при различном pH. В каждом случае молярное отношение липид:протеин равно 100:1.
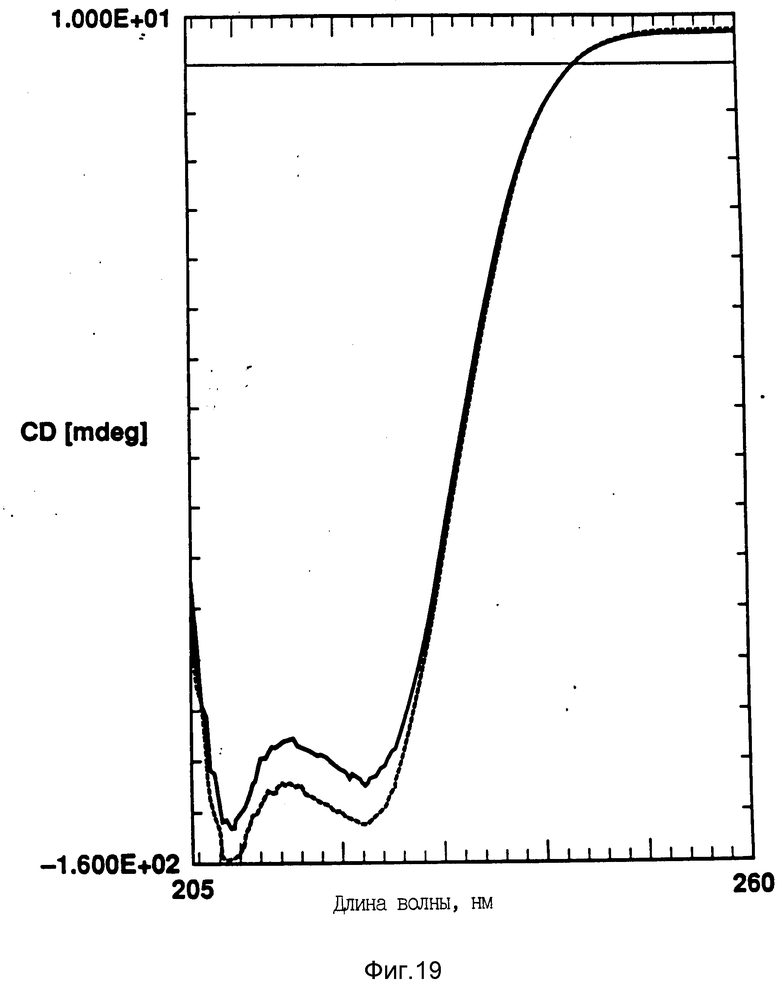
На фиг. 19 показано влияние температурных циклов на CD PEG-G-CSF (-) и DMPG:PE-G-CSF (17:1 молярное) (--). Образцы нагревают до 90oC и охлаждают до 10oC.
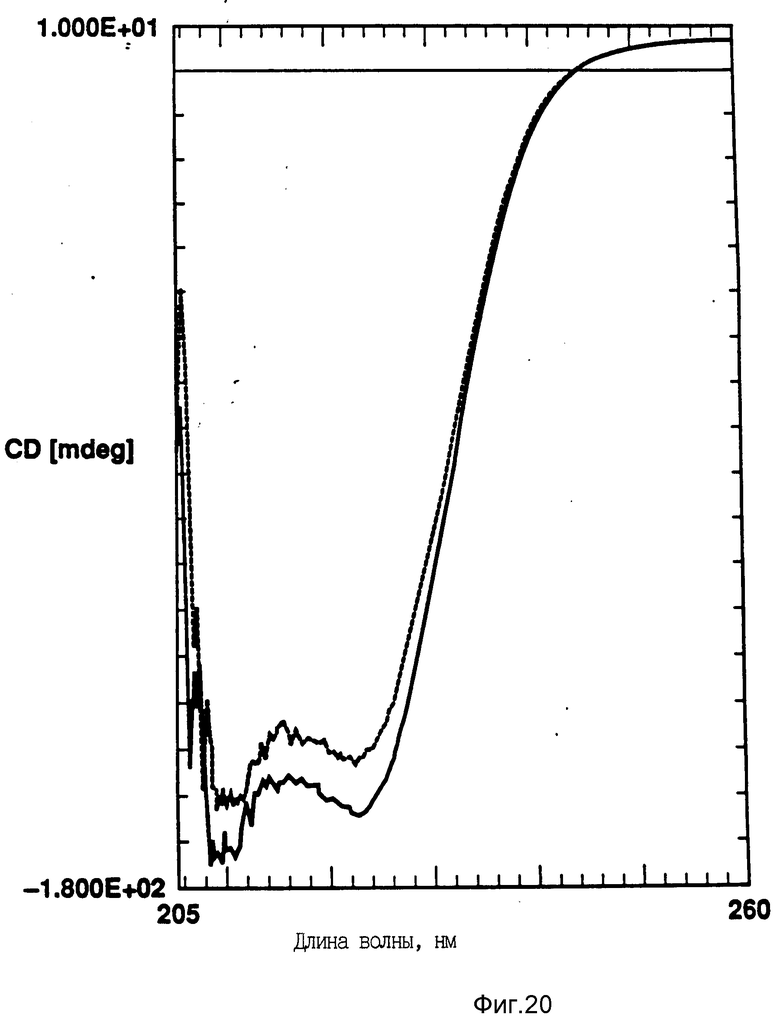
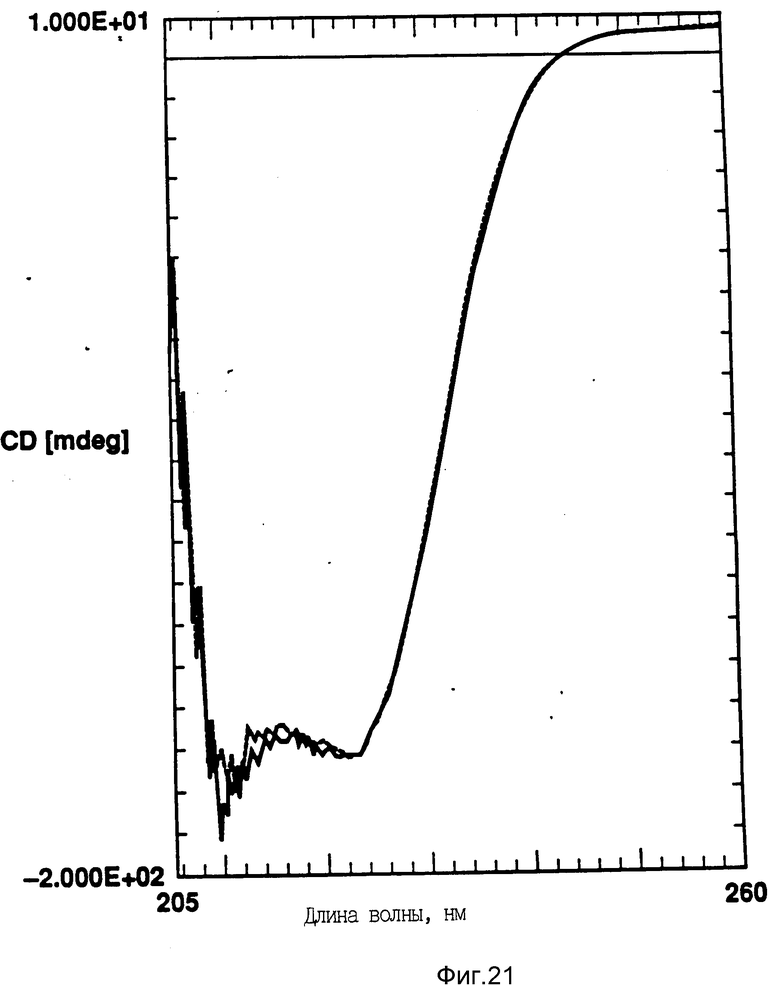
На фиг. 20 показано влияние температурных циклов на CD GM-CSF в PBC, pH 7,0. GM-CSF при 10oC (-) сравнительно с GM-CSF, который нагревают до 90oC и охлаждают до 10oC (--); на фиг. 21 - влияние температурных циклов на CD DPPG: PEG-G-CSF (17:1 молярное). DPPG:GM-CSF при 10oC (-) сравнивают с DPPG: -GM-CSF, который нагревают до 90oC и затем охлаждают до 10oC (--).
Полное описание изобретения.
Композиция по изобретению более подробно описывается ниже и иллюстрируется нижеследующими примерами. Примеры показывают различные аспекты изобретения и включают данные испытаний по стабильности и биологической активности различных протеин:фосфалипидных композиций. Неожиданно взаимодействие протеинов с липидными пузырьками непосредственно стабилизирует протеиновую структуру протеина, оказывая, таким образом, стабилизирующее воздействие на протеин даже в условиях, приводящих к денатурации протеина в отсутствии липида. В настоящем изобретении предлагается использовать различные протеины, способные переходить в распрямленное состояние глобулы. Примерами таких протеинов являются цитокины, включая различные гемопоэтические факторы, такие как упомянутые выше G-CSF, GM-CSF, M-CSF, интерфероны (альфа, бета и гамма), интерлейкины (1-11), эритропоэтин (EPO), фактор роста фибробластов, клеточный фактор (stem all factor), фактор роста нерва, BDNF, NT3, полученный из тромбоцита фактор роста, и фактор роста опухолевых новообразований (альфа, бета). Другие протеины могут быть оценены по способности к переходу в MGS. Если такой протеин способен к превращению в MGS, то такой протеин может затем быть приведен в контакт с пузырьками отрицательно заряженной липосомы и определен эффект стабилизации.
В общем случае G-CSF, используемый в этом изобретении, может быть в нативной форме, выделенной в чистом виде из организма млекопитающих, или альтернативно может быть продуктом химического синтеза или продуктом эукариотной или прокариотной экспрессии клетки хозяина экзогенной последовательности ДНК, полученной геномным или сДНК клонированием или генным синтезом. Подходящие прокариотные хозяева включают различные бактериальные клетки (например, E. coli). Подходящие эукариотные хозяева включают дрожжи (например, S.cerevisiae) и клетки млекопитающих (например, яичника китайского хомячка, обезьяны). В зависимости от используемого хозяина продукт G-CSF может быть гликосилирован млекопитающими или другими эукариотными карбогидратами, или может быть негликосилирован. Настоящее изобретение предполагает использование любых или всех таких форм G-CSF, хотя предпочтительным, с точки зрения наибольшей коммерческой практичности, является рекомбинантный G-CSF, особенно полученный E. coli.
G-CSF, который подвергают химической модификации с целью использования в настоящем изобретении, может быть также либо природным человеческим G-CSF (nhG-CSF), либо продуктом рекомбинантного процесса нуклеиновой кислоты в прокариотной или эукариотной клетке хозяина.
В общем случае предполагаемой химической модификацией является присоединение химической половины к самой молекуле G-CSF.
Обзорной статьей, описывающей модификацию протеина и слияние протеинов, является Francis, Focus on Growth Factors 3; 4-10 (май 1992) (опубликовано Mediscript, Mountview Court, Frun Barnet Lane, London N 20 OLD, UK). B EP 0401384 описаны материалы и методы G-CSF, к которым присоединены молекулы полиэтиленгликоля. Присоединение может быть непосредственным связыванием с протеином или к половине, действующей в качестве мостика для активного агента. Наиболее предпочтительной, как наиболее стабильной, является ковалентная связь. Химическая модификация может способствовать контролируемому, длительному или расширенному эффекту G-CSF. Это может приводить, например, к контролированию времени, необходимого для достижения химически модифицированным G-CSF свернутого в клубок положения. Примером химического модификатора являются полиэтиленгликолевые композиции, включая их производные.
По изобретению предполагается использовать любые химически модифицированные G-CSF препараты, эффективные при применении. Эффективность может быть определена различными методами, известными специалисту. Предпочтительными являются пегилированный G-CSF, в особенности полученный E.coli G-CSF и еще более предпочтительно три-тетра пегилированный полученный E.coli G-CSF. G-CSF описан как наиболее стабильный в кислой среде, несмотря на то, что при pH 2,5-5,0 происходит конформационное изменение, сопровождающееся потерей третичной структуры и повышением содержания альфа спиральной формы (Narhi et al., J.Protein Chem. 10, 359-367 (1991). Это конформационное изменение характерно для распрямленного состояния глобулы (MGS). Так, как это происходит в случае работы препаратора с другими протеинами, способными к превращению в MGS, препаратор, имеющий дело с G-CSF, должен принимать защитные меры против термического распрямления вторичной и третичной структуры для предотвращения агрегации и денатурации. GM-CSF, пригодный для использования по изобретению, может быть нативной формой, выделенной в чистом виде из организма млекопитающих, или продуктом прокариотной или эукариотной экспрессии клетки хозяина экзогенной последовательности ДНК, полученной геномом или сДНК клонированием или генным синтезом. Подходящие прокариотные хозяева включают различные бактериальные клетки (например, E.coli). Подходящие эукариотные хозяева включают дрожжи (например, S.cerevisiae) и клетки млекопитающих (например, яичника китайского хомячка, обезьяны). В зависимости от используемого хозяина продукт GM-CSF выражения может быть гликосилирован млекопитающим или другими эукариотными карбогидритами или может быть негликосилирован. Настоящее изобретение предполагает использование любых и всех таких форм GM-CSF, хотя предпочтительным является рекомбинантный GM-CSF, в особенности полученный E. coli, по соображениям коммерческой практичности.
Липидные пузырьки, используемые в композициях по изобретению, являются теми отрицательно заряженными липосомами, которые способны к взаимодействию с протеином. Особенно предлагается использовать следующие липосомы:
диолеилфосфатидилглицерол (DOPG), димиристоилфосфатидилглицерол (DMPG), дипальмитоилфосфатидилглицерол (DPPG), яичный фисфатидилглицерол, диолеилфосфатидилэтаноламин (DOPE), яичный фосфатидилэтаноламин, диолеилфосфатидовая кислота (DOPA), димиристоилфосфатидовая кислота (DMPA), дипальмитоилфосфатидовая кислота (DPPA), диолеилфосфатидилсерин (DOPS), димиристоилфосфатидилсерин (DMPS), дипальмитоилфосфатидилсерин (DPPS), яичный фосфатидилсерин, лизофосфатидилглицерол, лизофосфатидилэтаноламин, лизофосфатидилсерин.
В предпочтительном варианте с использованием E.coli полученного rhGM-CSF липосомным пузырьком является DMPG с отношением DMPG:GM-CSF 17:1, при pH 7,0 в растворе фосфатного буфера (PBS).
В предпочтительном варианте с использованием химически модифицированного (пегилированного) E.coloi полученного rhG-CSF является три-тетра связанным, липосомным пузырьком является DMPG с отношением DMPG:PEG-G-CSF 17:1, при pH 4,5.
Пример 1. Производили первоначальные эксперименты для исследования возможности введения рекомбинантного G-CSF (rhG-CSF) в липидный пузырек. rhG-CSF получили при использовании рекомбинантной ДНК технологии, согласно которой E.coli клетки были подвергнуты трансфекции посредством ДНК-последовательности, кодирующей человеческий G-CSF, как описано в патенте US N4810643, rhG-CSF готовили в виде раствора с концентрацией 4 мг/мл в разбавленной HCl, pH 4,0. Все липиды были получены из Avanti Polar Lipids (Albaster, Ala) и выдержаны при -20oC в атмосфере азота при конечной концентрации 100 мг/мл в хлороформе.
Приготовление G-CSF:фосфолипидных комплексов.
С целью приготовления липидных пузырьков для сочетания с G-CSF 30 мкмоль соответствующего липида помещают в стеклянную трубку и сушат в потоке газообразного азота до образования тонкой пленки. Липидную пленку высушивают не менее двух часов под вакуумом для удаления любых следов хлороформа. Липидную пленку гидрируют в 1 мл либо дистиллированной деионизированной воды (ddH2O), фосфатном буферном растворе, pH 7,2 (Cibro/BRL "D-PBS") или в 150 мМ NaCl. Затем образцы были обработаны ультразвуком в ультразвуковой установке типа ванны (лаборатория Supplies, Hicksville, N.Y.). Ультразвуковую обработку продолжали до тех пор, пока образцы не приобрели оптическую прозрачность (обычно в пределах 10-15 мин). Образцы хранили при 4oC под азотом вплоть до использования. Конечная концентрация липида составляла 30 мМ. Альтернативно, липидные пузырьки могут быть приготовлены высушиванием под азотом как описано выше 300 μмол липида. Сухие липидные пленки гидрировали в 10 мл соответствующего водного раствора, как описано выше. Образцы были затем микроэмульгированы в эмульгаторе (Mictofluidics Model 110S, Microfluidics, Inc., Cambridge, MA), работающем при 10000 пси. Образцы подвергали 10 циклам рециркуляции в приборе. Микроэмульгированные образцы затем хранили при 4oC, как описано выше.
G-CSF:фосфолипидные комплексы готовили смешением G-CSF (как описано выше) со специфичным липидом (как описано выше). Смешение осуществляют в вихревом смесителе перемешиванием или бережным встряхиванием. Готовят образцы с различным мольным соотношением липид:G-CSF для оценки мембранной проницаемости и стабилизации протеина. Например, для приготовления 3 мл образца (в воде), содержащего 0,2 мг/мл G-CSF при мольном соотношении липид:G-CSF 40:1, 150 μл исходного раствора G-CSF соединяют с 44 μл липида (30 мМ исходный раствор в воде, приготовленный при воздействии ультразвука) и добавляют воду до конечного объема образца 3 мл. Рекомендуемая пятиминутная инкубация (но не обязательная) была осуществлена перед использованием и испытанием образца.
G-CSF может быть также соединен с гидрированным липидом перед микроэмульгированием. Последующее микроэмульгирование смеси как описано выше приводит к внедрению G-CSF в липидную мембрану.
Анализ G-CSF:фосфолипидных комплексов.
1. Триптофанный эмиссионный спектр. Существуют две группы триптофана в rhG-CSF, являющиеся очень чувствительными к условиям локальной окружающей среды. Поэтому проводили анализ для определения rhG-CSF триптофанной флуоресценции в условиях контактирования rhG-CSF с липосомой. Голубой сдвиг максимума флуоресцентной эмиссии предполагает, что триптофаны находятся в более гидрофобной окружающей среде и следовательно rhG-CSF внедрен в липидную мембрану. Отличным обзором триптофанного флуоресцентного анализа является Principles of Fluorescence Microscopy, J.Lakowicz, Chap. 11 (Plenum Press, New York, 1983).
Триптофанную флуоресценцию G-CSF:липидных комплексов (как описано выше) анализировали возбуждением образцов при длине волны 280 нм и в процессе сканирования эмиссии от 285 нм до 420 нм с приращением в 1 нм при скорости 1 нм/с. Образец имел объем 3 мл при конечной концентрации G-CSF 0,2 м/мл для всех образцов. Отношение липид:G-CSF варьировалось. Все измерения флуоресценции проводили с использованием PTJ Alphascan флуорометра (South Brunswick, NY). Все изменения осуществлялись при 25oC и эту температуру поддерживали использованием держателя кюветы с водной рубашкой, связанного с циркулирующей водной ванной. Собирали эмиссионный спектр и анализировали, используя данные программного обеспечения PTJ.
На фиг. 1 показан спектр флуоресценции rhG-CSF в присутствии и в отсутствии небольших однослойных пузырьков, состоящих DOPG. rhG-CSF имеет максимум эмиссии в области 334 нм в отсутствии пузырьков DOPG. В присутствии DOPG и при соотношении липид:протеин 100:1 rhG-CSF триптофанная флуоресценция показывает сдвиг максимума флуоресценции в голубую область эмиссии в область 327 нм сильное увеличение интенсивности флуоресценции.
Короткая длина волны флуоресцентной эмиссии в присутствии DOPG предполагает большую гидрофобность среды окружающей триптофаны, чем нативный протеин. Как показано на фиг. 2, сдвиг флуоресценции зависит от мольного отношения DOPG:G-CSF и мембранное внедрение обнаруживается как только достигается соотношение DOPG:G-CSF 10:1.
2. Испытания иодидного тушения. Иодид является эффективным тушителем триптофанной флуоресценции, но он не может проникать через липидные мембраны. Поэтому эффективное тушение триптофанной флуоресценции иодидом показывает доступность групп основной массе водного растворителя, в то время как защита от иодидного гашения наблюдается в случае, когда триптофаны протеина изолированы от водного растворителя.
В этих экспериментах использованы G-CSF и DOPG: G-CSF композиции (отношение липид: протеин 100:1). После того, как первоначальное считывание (F0) на образцах было произведено и зарегистрировано, измеряли интенсивность флуоресценции после введения возрастающих количеств иодида калия (KJ) (5 М исходный раствор). Образцы и KJ растворы были приготовлены с содержанием 1 мМ Na2SO3 (конечная концентрация) как описано Lee et al., Biochem. Biophys. Acta, 984: 174 - 182 (1989) и Le Doan et al., Biochem. Biophys. Acta, 858: 1 - 5 (1986). Добавление Na2SO3 препятствует образованию J, который может распределяться между неполярными участками протеинов и мембранами. Данные испытаний были анализированы с помощью уравнения Stern-Volmer (F0/F = 1+Kkj[KJ] ), где F0 и F представляют собой интенсивности флуоресценции образцов соответственно в отсутствии и в присутствии KJ при концентрации [KJ]. Kkj является константой тушения Stern-Volmer для KJ тушения G-CSF триптофанных радикалов (Lehrer, S., Biochemistry 10: 3254 - 3263 (1979)).
На фиг. 4 показаны данные кривой Stern-Volmer. В отсутствии пузырьков DOPG, rhG-CSF флуоресценция эффективно гасится KJ. В присутствии DOPG данные Stern-Volmer расположены на прямой, что указывает на то, что иодид имеет слабую доступность к обоим триптофанам. Данные показывают, что группы триптофана, доступные для иодида в отсутствии DOPG, становятся недоступными для иодида в присутствии DOPG. Поэтому часть rhG-CSF, содержащая этот триптофан, должна быть внедренной в двойной слой DOPG.
3. Измерение превращения энергии. Как уже указывалось раньше, между донорами триптофана и растворимыми в липиде акцепторами флуоресценции, такими как пирендекановая кислота, может произойти передача энергии, так как спектр возбуждения этой пробы значительно перекрывает спектр эмиссии триптофана (Friere et al., Biochemistry, 22: 1675 - 1680 (1983)). В случае внедрения протеина в липидную мембрану, передача энергии от триптофана к пирену приведет к затуханию флуоресценции триптофана. В этом эксперименте непрерывно измеряют интенсивность эмиссии триптофана различных комплексов G-CSF: липид перед (F0) и после (F) добавление различных количеств пирендекановой кислоты (исходный раствор в тетрагидрофуране с концентрацией 30 μг/мл). Образцы непрерывно перемешивают в процессе добавления пирендекановой кислоты для лучшего смешения пирендекановой кислоты с образцом. Отношение F/F0 пропорционально количеству передачи энергии между G-CSF триптофанами и гидрофобным акцептором энергии - пирендекановой кислотой.
На фиг. 5 показан характер затухания rhG-CSF в присутствии DOPG (отношение липид: протеин 100:1) как функция добавленного количества пирендекановой кислоты. Затухание флуоресценции начинается при очень небольших концентрациях пирендекановой кислоты (< 1 мол.%), так что эффект флуоресцентной пробы на мембранную структуру и поведение является минимальным. Поскольку предполагают, что пирендекановая кислота может очень быстро проникать в липидный двойной слой, настоящие данные показывают, что rhG-CSF довольно глубоко внедряется в DOPG мембраны, что вызывает эффективную передачу энергии от триптофана к пиреновому акцептору. Процесс передачи энергии был подтвержден исследованием спектра возбуждения пирендекановая кислота - меченые DOPG пузырьки в присутствии и в отсутствии rhG-CSF.
Вышеуказанный анализ показывает, что rhG-CSF может тесно взаимодействовать с ненасыщенными фосфолипидами подобно DOPG. В присутствии DOPG пузырьки rhG-CSF триптофан являются защищенными от водорастворимого тушителя флуоресценции, но является поддающимся затуханию флуоресценции в результате передачи энергии гидрофобной и флуоресцентной пробе.
Вместе взятые данные показывают, что rhG-CSF может внедряться в мембраны, состоящие из DOPG. Мембранное внедрение можно обнаружить при достижении отношения липид:G-CSF 10:1 и это число может представлять собой число липидов, окружающих внедренную часть протеина.
Пример 2. В этом примере определяют способность rhG-CSF к взаимодействию с другими фосфолипидами сравнением отношения интенсивностей F/F0 и максимумом эмиссии, как описано выше. В каждом случае отношение липид:rhG-CSF равняется 100:1.
На фиг. 6 показаны значения отношения F/F0 для rhG-CSF в отсутствии и в присутствии различных липидов. На фиг 7 показаны значения максимума эмиссии для тех же самых композиций. Данные на фиг. 6 и 7 показывают, что в дополнение к DOPG, rhG-CSF может внедряться в DMPG, DPPG и менее эффективно в фосфатидилэтаноламины (PE's) и в фосфатидилсерины (PS's). Кроме того, установлено, что NG-DOPG (DOPG образец с более отрицательно заряженной PE головной группой) более способствует внедрению rhG-CSF, чем DOPG.
DOPG, DMPG и DPPG являются нейтральными липидами и эти пузырьки мало влияют или вовсе не влияют на максимум эмиссии или интенсивность флуоресценции rhG-CSF, что указывает на отсутствии взаимодействия с этими фосфалипидами (см. фиг. 6 и 7 и фиг. 8, кривая 2).
Приведенные выше данные свидетельствуют о том, что протеин, способный переходить в распрямленное глобулярное состояние, может внедряться в различные липидные пузырьки. Однако это rhG-CSF:липидное взаимодействие происходит только тогда, когда используют отрицательно заряженные липидные пузырьки. Среди отрицательно заряженных липидных пузырьков пузырьки с небольшим отрицательным зарядом должны вызывать более сильное rhG-CSF взаимодействие.
Пример 3. В этом примере определяют влияние rhG-CSF:DOPG на стабильность протеина. Для этого измеряют круговой дихроизм на приборе Jasco J-720, снабженном штативом термостатированных клеток типа Peltier и магнитной мешалкой. Измеряют круговой дихроизм при 222 нм конечной концентрации rhG-CSF 80 μг/мл, pH 6,0. На колориметре Microcal MC-2 производят измерения дифференциальной сканирующей колориметрии. Сканируют образцы rhG-CSF (1 мг/мл, в воде) или DOPG:rhG-CSF (45:1 моль/моль, в воде) со скоростью 90oC/ч. Данные хранят и анализируют при помощи программного обеспечения Microcol.
Исследуют температурные изменения альфа-спирали G-CSF посредством измерения кругового дихроизма (222 нм) как функции повышения температуры. Развертывание спирали rhG-CSF под действием температуры при pH 6,0 показано на фиг. 9. Кривая показывает, что довольно большое превращение наблюдается при температуре около 60 - 70oC, что приводит к потере альфа-спиральной формы.
После такого превращения rhG-CSF необратимо осаждается из раствора. Температурный интервал процесса развертывания спирали подобен температуре плавления rhG-CSF при pH 7,0, как это было установлено методом дифференциальной сканирующей калориметрии и показано на фиг. 10.
Например, DOPG:rhG-CSF образцы показывают постепенную потерю альфа-спиралевидности с повышением температуры и, в противоположность одному rhG-CSF, развертывание спирали DOPG: rhG-CSF под действием температуры не является совместным (см. фиг. 8). Такой вывод также следует из отсутствия перехода плавления, как это показывает дифференциальная сканирующая калориметра (фиг. 10). Примечательно, что DOPG:rhG-CSF образцы могут восстанавливать альфа-спиралевидность после нагрева до 95oC и могут быть подвергнуты циклическому изменению температуры между 95oC и 10oC с полным восстановлением спиралевидной формы после охлаждения (см. фиг. 11). rhG-CSF сам по себе в таких условиях необратимо развертывается и осаждается из раствора.
Кроме того, исследовали влияние DMPG и DPPG на G-CSF круговой дихроизм. Использовали отношение липид:rhG-CSF 150:1, и, как и в случае с DOPG, DMPG и DPPG также стабилизируют вторичную структуру rhG-CSF (фиг. 12-14).
Эти данные показывают, что взаимодействие rhG-CSF с DOPG, DMPG и DPPG улучшает стабильность протеина в условиях, при которых rhG-CSF сам по себе нестабилен. Это взаимодействие стабилизирует вторичную и третичную структуру rhG-CSF.
Пример 4. В этом примере определяют влияние взаимодействия rhG-CSF:DOPG в соответствии с биологической активностью rhG-CSF. Исследуют активность in vitro rhG-CSF посредством G-CSF зависящего поглощения [3H] тримидина клетками костного мозга мыши, как описано в Zsebo et al., Immunobiology 172: 175-184 (1986). Все испытания проводились трижды. Активность in vivo определяли посредством мышечной инъекции хомячков (доза rhG-CSF 100 μг/кг) и подсчетом количества белых кровяных телец (WBC).
1. Активность in vitro.
A. Определяют специфичную активность rhG-CSF в присутствии и в отсутствии DOPG. Также испытывают термообработанные образцы rhG-CSF и DOPG:rhG-CSF. Результаты испытаний сведены в таблице.
Как видно из данных таблицы, внедрение в DOPG двойной слой не снижает биологической активности rhG-CSF. После нагрева до 85oC в течение 10 мин активность rhG-CSF не поддается определению и протеин осаждается. После такой же обработки DOPG: rhG-CSF сохраняет приблизительно 85% активности rhG-CSF без нагревания и полностью восстанавливает вторичную структуру после охлаждения.
В. Также изучалась способность различных липидов стабилизировать rhG-CSF в процессе сушки вымораживанием. Образцы rhG-CSF в сочетании с различными липидами подвергали сушке вымораживанием и испытывали на активность, как описано выше. При смешении с rhG-CSF DOPG, DMPG и DPPG обеспечивают примерно 100% сохранение биоактивности rhG-CSF после сушки вымораживанием (фиг. 15). Сам по себе rhG-CSF не выдерживает процесса сушки вымораживанием.
2. Активность in vitro.
Определяют активность (подсчетом WBG) rhG-CSF в отсутствии и в присутствии липида. Активность измеряют после мышечной инъекции (доза rhG-CSF 100 μг/кг) на день О. Исследовали пять различных комплексов липид:rhG-CSF в каждом случае комплекс липид:rhG-CSF сохраняет активность in vivo (фиг. 16 и 17).
Вышеописанные исследования показывают, что внедрение в отрицательно заряженный двойной слой липида не оказывает отрицательного воздействия на биологическую активность rhG-CSF. Кроме того, оказалось, что защитное воздействие липида защищает rhG-CSF в течение процесса сушки вымораживанием.
Пример 5. В этом процессе испытывают способность химически модифицированного (пегилированного) G-CSF (PEG-G-CSF) и G-CSF, полученного как продукт эукариотной экспрессии клетки-хозяина (CHO-G-CSF), к взаимодействию с отрицательно заряженными липидными пузырьками. В случае CHO-G-CSF определение проводили на основе сравнения отношения интенсивностей F/Fo и максимума эмиссии (как описано выше в примере 1). В каждом случае мольное отношение липид: протеин равнялось 100:1. В случае же PEG-G-CSF определение проводили на основе анализа кругового дихроизма.
CHO-G-CSF получали по технологии рекомбинатной ДНК, согласно которой клетки яичника китайского хомячка подвергали трансфекции посредством кодирования последовательности ДНК человеческим G-CSF, как это описано в патенте США N 48810643 на имя Souza. CHO-G-CSF приготовляли в виде раствора в PBS с концентрацией 0,6 мг/мл, pH 7,0. Показано, что CHO-G-CSF взаимодействует с DOPG подобно rhG-CSF. Каждый образец показывает увеличение интенсивности флуоресценции в присутствии DOPG и сдвиг максимума эмиссии в голубую область в присутствии DOPG (фиг. 18 и 19). Следовательно, взаимодействие DOPG не является следствием некой специфичности рекомбинантной формы G-CSF.
В этом эксперименте использовали PEG-G-CSF в виде три-тетра пегилированного E-coli полученного G-CSF (используют PEG 6000). Образцы DMPG:PEG-G-CSF (мольное отношение 17: 1) готовили, как описано выше. Найдено, что образцы DMPG: PEG-G-CSF полностью восстанавливают вторичную структуру после нагревания (фиг. 19). Несмотря на присутствие молекул PEG модифицированный протеин был способен к взаимодействию с липидом также, как нативный протеин.
Приведенные выше данные показывают, что эффект стабилизации, связанный с взаимодействием G-CSF с отрицательно заряженными липидными пузырьками, не является присущим только rhG-CSF, полученному как продукт прокариотной экспрессии клетки-хозяина. Химически модифицированный протеин, способный к превращению в MGS, после контактирования с липидным пузырьком также проявляет стабилизирующее воздействие.
Пример 6. В этом примере изучают влияние DMPG и DPPG на GM-CSF. GM-CSF представлял собой рекомбинантный человеческий GM-CSF согласно патенту США N 5047504. Его готовили в виде раствора с концентрацией 1 мг/мл в фосфатном буферном растворе, pH 7,0. Отношение липид: GM CSF равно 17:1. Измеряли термическую стабильность посредством анализа кругового дихроизма, как описано выше. DMPG и DPPG могут приводить к лучшей термической стабильности GM-CSF, то есть к восстановлению вторичной структуры после нагрева (фиг. 20 и 21). Данные этого исследования показывают пример другого образца протеина, способного к превращению в распрямленное состояние глобулы и к взаимодействию с отрицательно заряженным липидным пузырьком, для повышения термической стабильности протеина.

Изобретение относится к области молекулярной биологии и фармакологии, в частности к фосфолипидной композиции и способу ее приготовления. Сущность изобретения заключается в том, что композиция представляет собой липосомный пузырек, содержащий белковый препарат, при этом липосома представляет собой один из фосфолипидов, а протеин представляет собой гемопоэтический фактор, в частности цитокин, G - CSF, или CM - CSF. Описан также способ приготовления композиции, которая имеет рН 3,0 - 7,5 при соотношении липида к протеину не менее 10: 1. Изобретение расширяет арсенал липосомальных форм лекарственных средств. 2 с. и 21 з.п., 21 ил, 1 табл.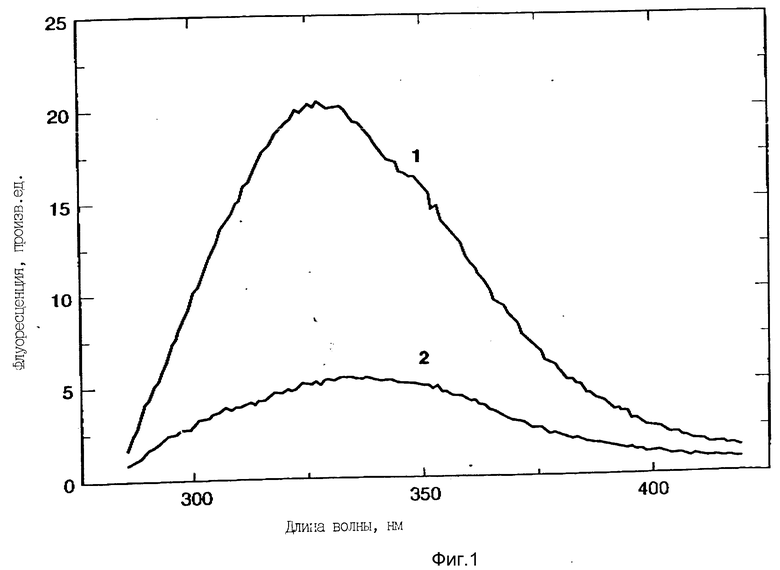
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Сплав на основе меди | 1973 |
|
SU456842A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Мембранное устройство | 1973 |
|
SU457837A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Gregoriadis G | |||
| "Liposomes", Drug Carriers in Biology and Medicine", N.-J | |||
| Academic Press, 1979, т | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| СПОСОБ ИЗГОТОВЛЕНИЯ ЧЕРТЕЖЕЙ ДЛЯ ОДНООБРАЗНОЙ РАСКРОЙКИ ПРЕДМЕТОВ ОДЕЖДЫ | 1919 |
|
SU287A1 |

