Изобретение относится к рекомбинантному химерному полипептиду-иммуногену, направленному на стимуляцию протективного иммунного ответа против ВИЧ-инфекции, и может быть использовано в качестве компонента вакцины в области биотехнологии, генной инженерии и медицины.
Количество ВИЧ-инфицированных в России ежегодно возрастает, и к концу 2016 превысило 1 миллион человек. При этом к настоящему времени отсутствуют схемы лечения ВИЧ-инфекции, которые позволили бы элиминировать вирус из организма. Современные способы терапии лишь замедляют прогрессирование ВИЧ-инфекции и её переход в стадию СПИДа. Кроме того, через несколько лет вирус мутирует, приобретая резистентность к применяемым препаратам. ВИЧ-инфекция поражает преимущественно молодое и наиболее трудоспособное население, что, безусловно, отражается как на экономике государства, так и на здоровье будущего поколения. Поэтому разработка эффективной вакцины против вируса иммунодефицита человека также является крайне актуальной задачей.
Для создания анти-ВИЧ вакцин было испробовано множество подходов, включая использование инактивированных, аттенуированных форм вируса, субъединичные вакцины, состоящие из отдельных вирусных гликопротеинов, создание генно-инженерных конструкций, ДНК-вакцины а также препараты на основе рекомбинантных вирусных и бактериальных векторов [1]. Однако ни одна из кандидатных анти-ВИЧ вакцин не продемонстрировала достаточный уровень эффективности. Наиболее успешной разработкой, прошедшей III фазу клинических испытаний, является созданная совместно компаниями Merck и Sanofi Pasteur прайм-буст вакцина ALVAC-HIV vCP1521/AIDSVAX B/E. Она включала в свой состав иммуногены, предназначенные для одновременной стимуляции В- и Т-клеточного иммунного ответа. В качестве праймирующего антигена использовался рекомбинантный вирус оспы канареек, экспрессирующий гены env, gag и протеазы ВИЧ-1 (ALVAC-HIV vCP1521), а бустерную вакцинацию осуществляли с помощью препарата AIDSVAX B/E (Vax003) на основе гликопротеина gp120 ВИЧ-1. Данное сочетание обеспечило снижение риска инфицирования на 31,2% [2]. Более детальный анализ результатов показал, что в первый год испытаний вакцина обеспечивала защиту на уровне 60%, после чего в крови вакцинированных начинала падать концентрация ВИЧ-специфических антител, и эффективность препарата снижалась. Дальнейшие исследования позволили установить причину невысоких показателей защиты от ВИЧ, связанных, по-видимому, с бустирующим компонентом AIDSVAX, ответственным за индукцию гуморального ответа. Выяснилось, что антитела, образующиеся в ответ на введение вакцины, либо не способны нейтрализовать вирус, либо обладают слабой нейтрализующей активностью [3, 4].
Приблизительно в это же время началось активное исследование гуморального ответа при ВИЧ-инфекции, что позволило добиться значительного прогресса в данной области. В частности, были обнаружены антитела, обладающие нейтрализующей активностью в отношении очень большого числа изолятов ВИЧ различных субтипов. Такие антитела, называемые нейтрализующими антителами широкого спектра действия (bnAbs - broadly neutralizing antibodies), связываются с консервативными областями поверхностных белков ВИЧ-1, практически не подверженных мутагенезу, за счет чего способны нейтрализовать до 98% всех известных первичных изолятов ВИЧ-1 [5]. Они обнаруживаются в крови многих ВИЧ-инфицированных, но их титр, как правило, очень низок. Кроме того, длительное аффинное созревание (которое может продолжаться более года) приводит к тому, что bnAbs появляются в организме с сильным опозданием, и к этому моменту уже неспособны существенно снизить вирусную нагрузку. В то же время при достаточно высоких концентрациях они весьма эффективно препятствуют развитию ВИЧ-инфекции. В качестве примера, подтверждающего эффективность bnAbs, можно привести работу Флориана Кляйна с соавторами, которые показали, что пассивная иммунизация гуманизированных мышей смесью из пяти антител широкого спектра действия PG16, PGT121, PGT128, 3BC176 и 45-46G54W подавляла репликацию ВИЧ-1, и еще в течение 60 дней после окончания курса позволяла поддерживать контроль над вирусом. Также на приматах было продемонстрировано протективное действие комбинации широконейтрализующих антител в отношении рекомбинантного вируса SHIV89.6P (вирус иммунодефицита обезьян, содержащий часть структурных компонентов ВИЧ) [6].
Однако использование моноклональных широконейтрализующих антител возможно только в качестве терапевтического средства. Кроме того, из-за высокой стоимости МКА данная терапия доступна лишь очень небольшому проценту населения. Тем не менее, учитывая уникальные свойства нейтрализующих антител широкого спектра действия, очень перспективной выглядит идея создания иммуногена, способного индуцировать их наработку, поскольку это потенциально решает проблему сильной антигенной вариабельности ВИЧ-1.
Один из распространенных подходов, использующихся при создании таких иммуногенов, основан на концепции рационального дизайна и заключается в получении растворимых фрагментов поверхностного гликопротеина gp120 ВИЧ-1, модифицированных таким образом, чтобы они содержали только те эпитопы, которые узнаются нейтрализующими антителами широкого спектра действия. Для этого вносят изменения, позволяющие стабилизировать структуру молекулы (точечные мутации, введение модификаций, приводящих к образованию дисульфидных связей, удаление петлевых фрагментов гликопротеина).
Другой подход заключается в создании не отдельных фрагментов gp120, а цельных тримеров со стабилизированной структурой, способных имитировать нативную конформацию поверхностного белка Env ВИЧ-1 [7, 8, 9]. В качестве альтернативы было предложено использовать мономерную стабилизированную форму оболочечного гликопротеина вируса для индукции антител к CD4bs - ключевому участку gp120, ответственному за связывание с CD4. Для получения иммуногена в соответствии с таким подходом сердцевина белка Env модифицируется с помощью направленного мутагенеза таким образом, чтобы полученная конструкция имитировала CD4bs сайт [10, 11, 12].
Перечисленные подходы, несмотря на большие перспективы, технически сложны и ресурсозатратны. Вследствие дороговизны и сложности данного метода пока не было создано ни одного вакцинного препарата, основанного на данной концепции.
Более технологически простой подход, применяемый для создания иммуногенов, направленных на индукцию вируснейтрализующих антител, заключается в использовании каркасных молекул, которые экспонируют эпитопы ВИЧ-1, узнаваемые нейтрализующими антителами широкого спектра действия. Данная технология, в частности, была использована для получения иммуногена, вызывающего наработку широконейтрализующих антител 2F5. Каркасные белки в данном случае подбирались таким образом, чтобы обеспечить расположение эпитопа в конформации, оптимальной для связывания с антителом [13]. Однако недостаток данного подхода заключался в том, что при создании описываемой конструкции не учитывался тот факт, что аминокислотное окружение эпитопа, при нахождении его в составе гликопротеина gp41, оказывает влияние на его конформацию. Вследствие этого иммуноген не продемонстрировал на модели животных образование вируснейтрализующих антител при иммунизации [13].
Помимо перечисленных конструкций, известен вакцинный препарат против ВИЧ-1 «ВИЧРЕПОЛ» (ГНЦ Институт иммунологии) [14]. Основной компонент вакцины - химерный белок rec (24-41), состоящий из фрагментов иммуногенных белков р24, gp41 и p17 ВИЧ-1 [15], конъюгированный с иммуностимулятором - носителем полиоксидонием (фармакологически активное соединение с молекулярной массой 100 кД с выраженной иммуномодулирующей активностью). Первая фаза клинических испытаний показала безопасность, иммуногенность и антигенную активность представленной вакцины. Кроме того, при иммунизации лабораторных животных данная вакцина вызывает наработку специфических антител к белкам р24 и gp41 ВИЧ-1, входящих в состав вакцины. Однако эти антитела обладают низкой вирус-нейтрализующей активностью: индекс нейтрализации составляет 54,9% при разведении иммунной сыворотки 1:50. Клеточный ответ был также относительно невысоким, как показали данные в реакции бласт-трансформации.
Известна также вакцинная композиция против вируса иммунодефицита человека [16]. Данный патент защищает рекомбинантные белки AFP (artificial fusion proteins), представляющих собой слитые друг с другом в различных комбинациях домены гликопротеинов ВИЧ-1. AFP содержат CTL-эпитопы, ассоциированные с контролем виремии и длительным отсутствием прогресса у индивидуумов с хронической ВИЧ-1 инфекцией. Кроме того, допускается включение в состав белков дополнительных доменов. Недостатком данного иммуногена является то, что AFP направлен на индукцию только CD8+ ответа, в то время как современные исследования показывают, что для обеспечения надежной защиты от ВИЧ-инфекции необходимо также стимулировать CD4+ T-h ответ и наработку нейтрализующих и не нейтрализующих ВИЧ-1-специфических антител.
Наиболее близким аналогом (прототипом) к настоящему изобретению является белок TBI, компонент вакцины против ВИЧ-инфекции КомбиВичВак [17]. TBI представляет собой рекомбинантный белок, включающий пять B-клеточных эпитопов, чередующихся с четырьмя T-клеточными эпитопами и формирующих конформацию четырех α-спиралей и петлевых участков [18]. На модели лабораторных животных было показано, что вакцина КомбиВичВак обеспечивает формирование ВИЧ-специфических антител, обладающих вируснейтрализующей активностью, а также стимулирует вирус специфический цитотоксический иммунный ответ у вакцинированных добровольцев [19].
Основным недостатком белка TBI является то, что TBI не содержит эпитопов, узнаваемых нейтрализующими ВИЧ-1 антителами широкого спектра действия. Это не позволяет использовать данный иммуноген в качестве универсального средства (вакцины) от ВИЧ-инфекции.
Техническим результатом заявляемого изобретения является создание рекомбинантного химерного полипептида-иммуногена nTBI, обеспечивающего повышение ВИЧ-специфического иммунного ответа за счет включения в состав указанного полипептида-иммуногена nTBI уникальных линейных имитаторов конформационных эпитопов, узнаваемых широконейтрализующими антителами, способного индуцировать образование антител, нейтрализующих ВИЧ-1.
Указанный технический результат достигается тем, что иммуноген nTBI содержит в качестве вакцинных антигенов эпитопы, узнаваемые широконейтрализующими ВИЧ-1 антителами. Ген, кодирующий nTBI, клонирован в плазмидный вектор pET21a по сайтам рестрикции NdeI и XhoI. Для экспрессии полипептида используется штамм Е.coli BL-21. При этом полипептид nTBI представляет собой стерильный очищенный белок и имеет аминокислотную последовательность SEQ ID NO:1, представленную на фиг. 1 и в приложении.
Заявляемый полипептид-иммуноген, направленный на индукцию нейтрализующих ВИЧ-1 антител широкого спектра действия, получают следующим образом.
При создании иммуногена nTBI за основу был взят ранее спроектированный белок TBI. Модификации исходного варианта белка TBI были направлены, в первую очередь, на улучшение его антигенных характеристик. С этой целью была произведена замена нескольких наименее иммуногенных пептидов, входящих в состав оригинального TBI, на аминокислотные последовательности, представляющие собой эпитопы (либо их линейные имитаторы), с которыми связываются нейтрализующие ВИЧ-1 антитела широкого спектра действия. Из молекулы TBI были удалены 2 B-клеточных эпитопа из белка Gag (99-109 и 351-361), имеющих низкую иммуногенность, а также эпитоп env (255-266), который распознается антителами, не обладающими способностью нейтрализовать вирусные частицы. Удаленные пептиды были заменены на эпитопы из MPER региона гликопротеина gp41, узнаваемые нейтрализующими ВИЧ-1 антителами широкого спектра действия 10e8 и 2F5. Также в состав нового белка был включен линейный пептид-имитатор эпитопа, расположенного в области CD4bs gp120 и узнаваемого bnAb VRC01, выявленный ранее с помощью фагового дисплея [20].
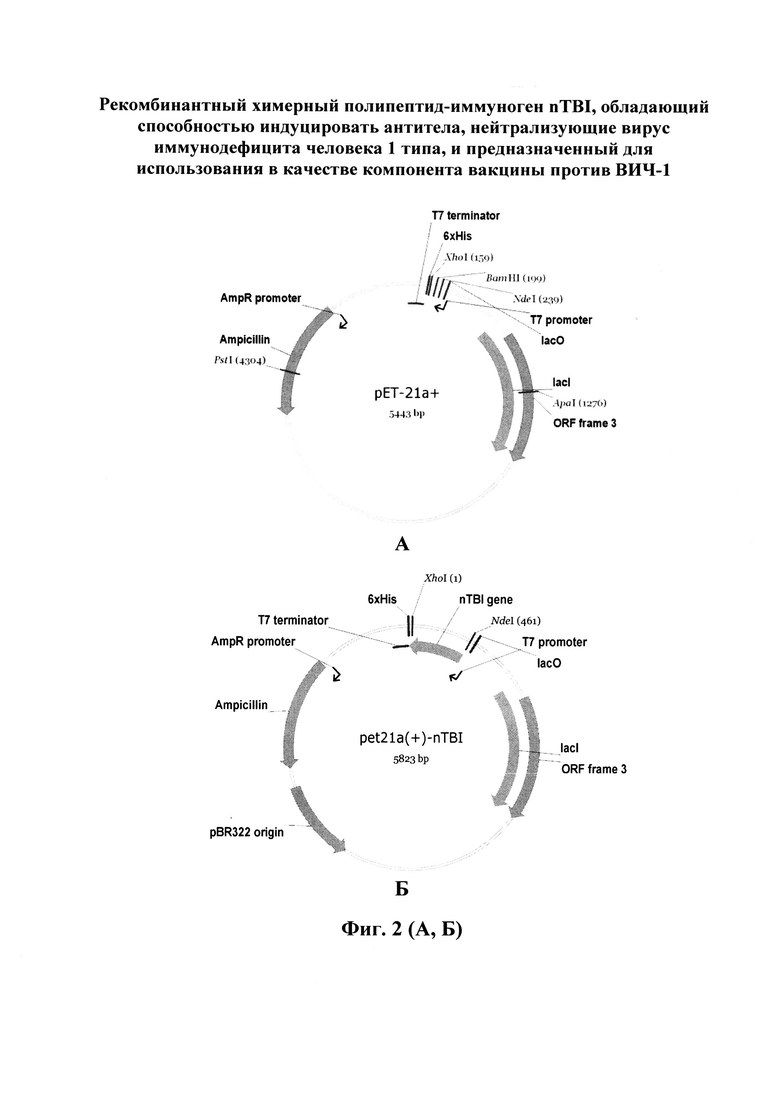
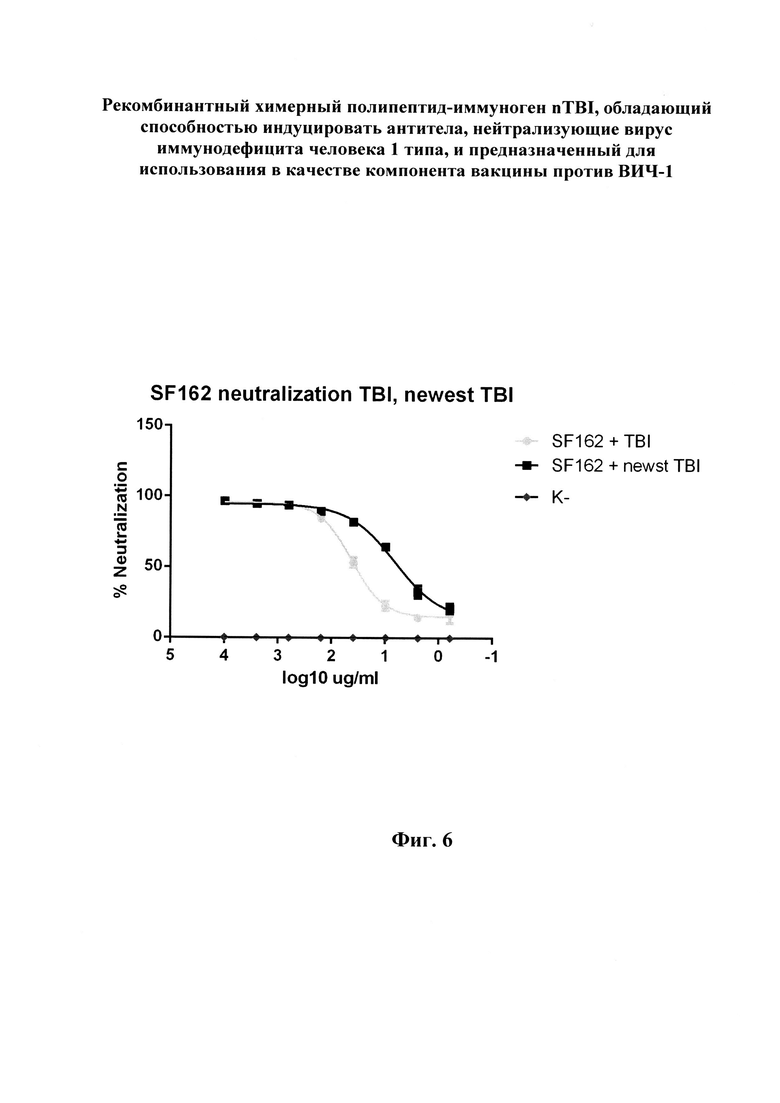
Помимо замены пептидов, были проведены модификации, направленные на повышение экспрессии гена целевого белка. В структуру nTBI с N-конца был добавлен фрагмент транскрипционного фактора InfB E. coli, позволяющий увеличить уровень экспрессии гена в прокариотической системе экспрессии [21]. Также была проведена процедура оптимизации кодонного состава гена nTBI с использованием онлайн-сервиса Codon Optimization OnLine. Спроектированный ген клонировали в составе вектора pET21a+. Использовали сайты, узнаваемые эндонуклеазами рестрикции XhoI и NdeI. Полученной конструкцией трансформировали клетки E. coli BL21, биомассу трансформированных клеток наращивали в течение ночи на среде LB. Полученную биомассу обрабатывали с помощью ультразвукового дезинтегратора, центрифугировали и ресуспендировали в лизирующем буфере, содержащем 8 М мочевину. Очистку целевого белка проводили с помощью металл-хелатной аффинной хроматографии на колонке с Ni-NTA-агарозой. Для перевода белка в нативное состояние использовали ступенчатый диализ против буферов с постепенным снижением концентрации мочевины. Наличие фракции целевого белка и степень очистки оценивали с помощью электрофореза в ПААГ в денатурирующих условиях. На фиг. 1. представлена нуклеотидная последовательность гена, кодирующего полипептид-иммуноген nTBI, и аминокислотная последовательность указанного полипептида. На фиг. 2 (А, Б) изображена схема клонирования данного гена в вектор pET21a(+). На фиг. 3 приведена первичная структура белка TBI и полученного на его основе nTBI. Замененные B-клеточные эпитопы белка nTBI выделены жирным шрифтом, более бледным шрифтом - Т-h эпитопы, а также пептидная последовательность, обеспечивающая усиление экспрессии (expressivity tag). Цилиндрами обозначены участки, имеющие структуру α-спиралей, серыми стрелками - β-складок. На фиг. 4, А приведены результаты электрофоретического разделения белков TBI и nTBI в 14% SDS-ПААГ, а на фиг. 4, Б - а вестерн-блоттинг белков TBI и nTBI, перенесенных на нитроцеллюлозную мембрану, где: 1 - образец белка nTBI; 2 - образец белка TBI; M - маркер молекулярных масс PAGERuler (TermoFisher). На фиг. 5 представлены данные анализа сывороток кроликов, иммунизированных белком nTBI, с использованием тест-системы NEW LAV-BLOT1, где: 3 - сыворотка ВИЧ-положительного пациента, взятая из набора New LAv Blot (положительный контроль); 4 - сыворотка кролика, иммунизированного nTBI; К- - сыворотка из образца крови кролика, взятой до иммунизации nTBI (отрицательный контроль). На фиг. 6. приведена кривая нейтрализации env-псевдотипированных частиц ВИЧ-1 сыворотками кроликов, иммунизированных белками TBI и nTBI. Черным и серым цветами обозначены кривые, построенные для сывороток кроликов, иммунизированных белками TBI и nTBI соответственно. Линия (с ромбами) представляет нейтрализацию сывороток животных из контрольной группы. Ниже приведены примеры осуществления изобретения.
Пример 1. Получение плазмиды pET21a-nTBI
Конструирование плазмиды проводят с помощью стандартных методов молекулярного клонирования [22]. Ген, кодирующий белок nTBI, был химически синтезирован компанией Евроген, нуклеотидная последовательность гена представлена на фиг. 1. Ген клонируют в составе плазмиды pET21a(+) (фиг. 2, Б).
Плазмидный вектор pET21a(+) (Novagen) гидролизуют эндонуклеазами рестрикции NdeI и XhoI, полученную ДНК выделяют с помощью набора Qiagen MiniPrep и лигируют с геном, кодирующим белок nTBI. Полученной лигазной смесью трансформируют клетки Е. coli XL1-Blue (Novagen) методом электропорации согласно методике, описанной в [23]. Трансформированные клетки высевают на чашки с LB агаром, содержащим 100 мкг/мл апмициллина. Для проверки наличия в трансформантах плазмидной ДНК с целевой вставкой отдельные колонии пересевают в пробирки с 2 мл среды LB, содержащей 100 мкг/мл ампициллина, и подращивают в течение ночи при 37°С. Плазмидную ДНК из полученных клеток выделяют методом щелочного лизиса [24]. Выделенную ДНК анализируют путем секвенирования по методу Сэнгера с использованием праймеров T7 promoter primer #69348-3 и T7 terminator primer #69337-3 (Novagen). Используют набор CEQ2000Dye Terminator Cycle Sequencing Kit и 16-капилярный автоматический секвенатор ABI 3130xl. Полученные секвенограммы должны подтверждать наличие гена, кодирующего белок nTBI, в составе рекомбинантной плазмиды pET21a-nTBI. Секвенограмма должна быть идентична нуклеотидной последовательности целевого гена, представленного на фиг. 1.
Пример 2. Наработка и очистка белка nTBI
Для получения белка nTBI плазмиду pET21-nTBI трансформируют в штамм-продуцент E. coli BL-21 методом электропорации. Клоны отбирают на чашках с агаризованной средой LB по устойчивости к ампициллину. Наращивание биомассы клеток E. coli, трансформированных полученной плазмидой, осуществляют на среде LB с ампициллином (40 мкл/мл) на термостатированных качалках при 37°С до оптической плотности 0,8-0,9. Индукцию синтеза целевого белка проводят путем добавления в среду 1 мМ ИПТГ и последующей инкубации в течение 4-6 ч. Полученную биомассу осаждают центрифугированием при 4°С и суспендируют в лизирующем буфере, содержащем 50 мМ NaH2PO4, 300 мМ NaCl, 0,1% Tween 20, 10 мМ Tris, 30 мМ имидазол и PMSF (Sigma Aldrich) в соотношении 7 мл буфера на 1 г биомассы. Далее суспензию помещают в ледяную баню и производят обработку ультразвуком 18 раз по 2 мин с перерывом по 1 мин для охлаждения смеси. Далее проводят центрифугирование при 34000 g в течение 15 мин, полученный дебрис ресуспендируют в лизирующем буфере, повторно дезинтегрируют и центрифугируют. Очистку рекомбинантного белка осуществляют с помощью аффинной хроматографии на Ni-NTA-агарозе в присутствии 8 М мочевины. Для этого осадок ресуспендируют в 2 М растворе мочевины в 20 мМ Tris-буфере (pH 8.0), центрифугируют. Полученный дебрис растворяют в лизирующем буфере, содержащем 8 М мочевину, и наносят на колонку с Ni-NTA агарозой. Материал элюируют буфером, содержащим 8 М мочевину, 100 мМ NaH2PO4, 10 мМ Tris, pH 8.0, 250 мМ имидазол. Для перевода белка в нативное состояние используют ступенчатый диализ против буферов, содержащих 50 мМ NaH2PO4 и 6, 3,1, 0,1 М мочевину. На конечном этапе диализ проводят против физиологического раствора без мочевины. Наличие фракции целевого белка и степень очистки оценивают с помощью электрофоретического разделения белков по Лэммли в 14% SDS-ПААГ [26]. После электрофореза гель фиксируют в 40%-ном спиртовом растворе, содержащем 10% уксусной кислоты, отмывают в дистиллированной воде и окрашивают кумасси G-250.
Электрофореграмма (фиг. 4, А) должна подтверждать наличие в образце гомогенного препарата белка размером около 18 кДа, что соответствует расчетному значению.
Пример 3. Подтверждение антигенных свойств nTBI
Для подтверждения способности nTBI связываться с МКА, эпитопы которых были включены в его состав, а также оценки его антигенных свойств в сравнении с оригинальным белком TBI, проводят вестерн-блот анализ. Разделенные электрофоретически препараты белков TBI и nTBI переносят на нитроцеллюлозную мембрану методом полусухого переноса [23]. Мембрану блокируют 1% раствором бычьего сывороточного альбумина в PBS (0,002 М KH2PO4, 0,01 M Na2HPO4x7H2O, 0,137 M NaCl, 0,0027 M KCl, pH 7,4) для предотвращения неспецифического связывания. Иммунодетекцию проводят согласно протоколу для системы «SNAP i.d.» (Merck Millipore, США) с первичными моноклональными человеческими антителами 2F5, 10E8, а также с мышиными МКА 29F2, связывающимися с белком p24 ВИЧ-1. Связывание специфического антитела визуализируют с использованием вторичных антител rabbit anti-human IgG alkaline phosphatase (AP) и rabbit anti-mouse IgG-AP (Sigma-Aldrich, США) в разведениях, рекомендованных производителем, и добавлением хромогенного субстрата BCIP/NBT. Результаты Вестерн-блот анализа, представленные на фиг. 4, Б, демонстрируют способность пептидов, включенных в состав nTBI, специфически взаимодействовать с соответствующими МКА. В данном тесте не используют антитело VRC01, поскольку узнаваемый им эпитоп является конформационным, а белок nTBI после переноса на нитроцеллюлозную мембрану в Вестерн-блоте находится в денатурированном виде, лишенном третичной структуры. В отсутствие соответствующей пространственной организации эпитоп не узнается МКА VRC01.
Пример 4. Проверка иммуногенных свойств белка nTBI. Иммунизация лабораторных животных
Для проверки иммуногенных свойств nTBI очищенным препаратом белка иммунизируют лабораторных животных и проводят иммунологические тесты с использованием антисывороток. Иммунизацию проводят по следующей схеме. Первую и вторую группу кроликов иммунизируют очищенными препаратами белков TBI и nTBI соответственно в количестве 0,5 мг на кролика при первой иммунизации и 1 мг при последующих. Первую инъекцию проводят подкожно, в четыре точки вдоль спинного хребта, в смеси с полным адъювантом Фрейнда, последующие две - с интервалом в две недели в смеси неполным адъювантом. Забор крови проводят из краевой ушной вены перед иммунизацией у каждого животного (в дальнейшем сыворотки используют в качестве отрицательного контроля), а также через неделю после последней иммунизации.
Для постановки иммуноферментного анализа белок nTBI разводят в буфере PBS до концентрации 1 мкг/мл и вносят по 100 мкл в лунки 96-луночного планшета. Планшет блокируют добавлением в лунки 3%-ного раствора казеина в буфере PBS, инкубируют 2 часа при 37°С, после чего 3 раза промывают буфером PBS-T. Для титрования используют разведения сывороток в буфере PBS с 3%-ным казеином в диапазоне от 1:10 до 1:10240. В лунки заблокированного и отмытого планшета вносят по 100 мкл последовательных разведений сывороток, инкубируют при покачивании 1-1,5 часа при 37°С, после чего трижды промывают буфером PBS-Т. Специфически связавшиеся антитела выявляют добавлением 40000-кратного разведения вторичных антител mouse anti-rabbit IgG-AP. Инкубируют 1 час при 37°С при покачивании, после чего планшет 3 раза промывают PBS-Т и добавляют хромогенный субстрат на основе п-нитрофенилфосфата (pNPP). Инкубацию с субстратом проводят в течение 16 часов при комнатной температуре в термостате при 37°С. Результат регистрируют с помощью ИФА-ридера Bio Rad 680 Microplate reader при длине волны 405 нм. Результаты ИФА показывают, что титры сывороток кроликов, иммунизированных как TBI, так и nTBI, составляют 1:600.000.
Наличие в антисыворотках антител, специфичных к ВИЧ-1, оценивают с помощью тест-системы NEW LAV-BLOT 1 (Bio Rad), представляющей собой набор нитроцеллюлозных полосок (стрипов), на которые сорбирован лизат белков ВИЧ-1. Сыворотки иммунных животных разводят в 50 раз в буфере ТБС-Т (0,05 М трис HCl рН 7,5; 0,15 М NaCl, Твин-20 v/v 200:1) и инкубируют со стрипами в течение двух часов при покачивании. В качестве положительного контроля использовалась сыворотка, полученная из крови ВИЧ-положительного человека. Инкубация проводится по аналогичной схеме, описанной для сывороток кроликов. Далее полоски отмывают ТБС-Т 3 раза по 5 мин и добавляют 5000-кратные разведения вторичных антител rabbit anti-human IgG-AP (goat anti-rabbit IgG-AP в случае положительного контроля), инкубируют в течение 1 ч при покачивании. После отмывки вносят хромогенный субстрат BCIP/NBT. После проявления нитроцеллюлозных стрипов в течение 20 мин наблюдают окрашенные полосы, по электрофоретической подвижности соответствующие определенным вирусным белкам. На фиг. 5 представлен результат проявки стрипов из набора NEW LAV BLOT, подтверждающий наличие вирус-специфических антител в сыворотках иммунизированных кроликов, при этом выявляемые белки соответствуют по подвижности расчетным (эпитопы, включенные в nTBI, входят в состав гликопротеинов gp120, gp41 и p24).
Пример 5. Постановка реакции вируснейтрализации для оценки иммуногенных свойств белка nTBI
Вируснейтрализующую активность антисывороток определяют путем постановки реакции вируснейтрализации с использованием панели env-псевдотипированных вирусных частиц, полученных на основе штаммов ВИЧ-1 субтипа B. Псевдовирусы получают котрансфекцией клеток 293Т/17 плазмидами pcDNA3.1/V5, несущими различные полноразмерные гены gp160, и плазмидой pSG3Δenv, несущей все остальные гены ВИЧ-1, за исключением gp160. Процедуру проводят с использованием липофектамина 2000 (Invitrogen): на 5 мкл липофектамина 2000 берется по 300 нг плазмид pEnv и pSG3Δenv. Смесь перемешивают, инкубируют 15 мин при комнатной температуре и переносят в 24-луночный планшет к монослою клеток 293Т/17. Через 48 ч инкубации в СО2-термостате при 37°С отбирают культуральную среду. От клеток псевдовирусы отделяют путем низкоскоростного центрифугирования с последующей фильтрацией через нитроцеллюлозные фильтры с диаметром пор 0,22 мкм. Полученную псевдовирус-содержащую надосадочную жидкость хранят при -80°С в среде DMEM, содержащую 20 % фетальной бычьей сыворотки.
Постановку реакции вируснейтрализации проводят согласно протоколу Монтефиори [25]. Для проведения эксперимента из антисывороток предварительно выделяют фракцию IgG осаждением сульфатом аммония и последующей аффинной очисткой на сефарозе с белком A. В лунки планшета, содержащие 150 мкл культуральной среды, вносят последовательные пятикратные разведения фракции IgG из антисывороток объемом 11 мкл. В качестве отрицательного контроля (фоновый люминесцентный сигнал) используют один ряд лунок, содержащих только среду DMEM без добавления антисывороток. Затем в каждую лунку за исключением контрольного ряда добавляют по 50 мкл суспензий псевдовирусов с концентрацией, соответствующей значению 200 TCID50/100 мкл. Далее во все лунки вносят 100 мкл суспензии клеток TZM-bl с концентрацией 100000 клеток/мл и инкубируют 48 ч при 37°С. После инкубации из лунок отбирают по 150 мкл их содержимого и добавляют LAR реактив (Promega, США), использующийся в качестве субстрата для люциферазы. После лизирования клеток содержимое лунок переносят в лунки планшета для измерения люминесцентного сигнала. Уровень люминесценции измеряют с помощью люминометра Wallac 1420 Multilabel counter (Perkin Elmer, Waltman, MA) при длине волны 482 нм. На фиг. 6 представлены кривые нейтрализации на примере env-псевдотипированного вируса, полученного на основе ВИЧ-1 SF162. Можно заметить, что сыворотки кроликов, иммунизированных TBI и nTBI, обладают дозозависимой нейтрализующей активностью, в отличие от сыворотки неиммунизированных животных, которая не продемонстрировала нейтрализующий эффект. Кроме того, концентрация полумаксимального ингибирования (IC50) антисывороток к nTBI составляет 5,58 мкг/мл, превышая аналогичное значение концентрации антисывороток к TBI (34,61 мкг/мл) примерно в 6 раз. При этом значение IC50, измеренное для МКА 2F5, составило 1,17 мкг/мл. Данный факт говорит о существенном улучшении иммуногенных свойств белка nTBI по сравнению с исходным вариантом белка TBI, что подтверждает заявленный технический результат.
Источники патентной и научно-технической информации
1. Gorry P.R., McPhee D.A., Verity E., Dyer W.B., Wesselingh S.L., Learmont J., Sullivan J.S., Roche M., Zaunders J.J., Gabuzda D., Crowe S.M., Mills J., Lewin S.R., Brew B.J., Cunningham A.L., Churchill M.J. Pathogenicity and immunogenicity of attenuated, nef-deleted HIV-1 strains in vivo // Retrovirology. - 2007. - V. 4. - N 3. - P.1.
2. Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E, Benenson M, Gurunathan S, Tartaglia J, McNeil JG, Francis DP, Stablein D, Birx DL, Chunsuttiwat S, Khamboonruang C, Thongcharoen P, Robb ML, Michael NL, Kunasol P, Kim JH; MOPH-TAVEG Investigators. Vaccination with ALVAC and AIDSVAX to Prevent HIV-1 Infection in Thailand // New England Journal of Medicine. - 2009. - V. 361. - N. 23. - P. 2209-2220.
3. Kim J.H., Excler J.L., Michael N.L. Lessons from the RV144 Thai Phase III HIV-1 Vaccine Trial and the Search for Correlates of Protection // Annual Review of Medicine. - 2015. - V. 66. - P. 423-427.
4. Montefiori D.C., Karnasuta C., Huang Y., Ahmed H., Gilbert P., De Souza M.S., McLinden R., Tovanabutra S., Laurence-Chenine A., Sanders-Buell E., Moody M.A., Bonsignori M., Ochsenbauer C., Kappes J., Tang H., Greene K., Gao H., Labranche C.C., Andrews C., Polonis V.R., Rerks-Ngarm S., Pitisuttithum P., Nitayaphan S., Kaewkungwal J., Self S.G., Berman P.W., Francis D., Sinangil F., Lee C., Tartaglia J., Robb M.L., Haynes B.F., Michael N.L., Kim J.H. Magnitude and breadth of the neutralizing antibody response in the RV144 and Vax003 HIV-1 vaccine efficacy trials // Journal of Infectious Diseases. - 2012. - V. 206. - N 3. - P. 431-441.
5. Sok D., Burton D. R. HIV Broadly Neutralizing Antibodies: Taking Good Care Of The 98%. // Immunity. - 2016. - V. 45. - N. 5. - P. 958-960.
6. Ferrantelli F., Rasmussen R.A., Hofmann-Lehmann R., Xu W., McClure H.M., Ruprecht R.M. Do not underestimate the power of antibodies - Lessons from adoptive transfer of antibodies against HIV // Vaccine. - 2002. - V. 20. - N SUPPL. 4. - P. A61-A65.
7. Kovacs JM, Nkolola JP, Peng H, Cheung A, Perry J, Miller CA, Seaman MS, Barouch DH, Chen B. HIV-1 envelope trimer elicits more potent neutralizing antibody responses than monomeric gp120 // Proceedings of the National Academy of Sciences. - 2012. - V. 109. - N. 30. - P. 12111-12116.
8. Yang X., Lee J., Mahony E.M., Kwong P.D., Wyatt R., Sodroski J. Highly stable trimers formed by human immunodeficiency virus type 1 envelope glycoproteins fused with the trimeric motif of T4 bacteriophage fibritin // Journal of Virology. - 2002. - V. 76. - N 9. - P. 4634-4642.
9. Yasmeen A., Ringe R., Derking R., Cupo A., Julien J.P., Burton D.R., Ward A.B., Wilson I.A., Sanders R.W., Moore J.P., Klasse P.J. Differential binding of neutralizing and non-neutralizing antibodies to native-like soluble HIV-1 Env trimers, uncleaved Env proteins, and monomeric subunits // Retrovirology. - 2014. - V. 11. - N 1. DOI: 10.1186/1742-4690-11-41.
10. Wu X.L., Yang Z.Y., Li Y.X., Hogerkorp C.M., Schief W.R., Seaman M.S., Zhou T.Q., Schmidt S.D., Wu L., Xu L., Longo N.S., McKee K., O'Dell S., Louder M.K., Wycuff D.L., Feng Y., Nason M., Doria-Rose N., Connors M., Kwong P.D., Roederer M., Wyatt R.T., Nabel G.J., Mascola J.R. Rational Design of Envelope Identifies Broadly Neutralizing Human Monoclonal Antibodies to HIV-1 // Science. - 2010. - V. 329. - N 5993. - P. 856-861.
11. Zhou T.Q., Xu L., Dey B., Hessell A.J., Van Ryk D., Xiang S.H., Yang X.Z., Zhang M.Y., Zwick M.B., Arthos J., Burton D.R., Dimitrov D.S., Sodroski J., Wyatt R., Nabel G.J., Kwong P.D. Structural definition of a conserved neutralization epitope on HIV-1 gp120 // Nature. - 2007. - V. 445. - N 7129. - P. 732-737.
12. Zhou T.Q., Georgiev I., Wu X.L., Yang Z.Y., Dai K.F., Finzi A., Kwon Y.D., Scheid J.F., Shi W., Xu L., Yang Y.P., Zhu J.A., Nussenzweig M.C., Sodroski J., Shapiro L., Nabel G.J., Mascola J.R., Kwong P.D. Structural Basis for Broad and Potent Neutralization of HIV-1 by Antibody VRC01 // Science. - 2010. - V. 329. - N 5993. - P. 811-817.
13. Ofek G., McKee K., Yang Y., Yang Z.Y., Skinner J., Guenaga F.J., Wyatt R., Zwick M.B., Nabel G.J., Mascola J.R., Kwong P.D. Relationship between antibody 2F5 neutralization of HIV-1 and hydrophobicity of its heavy chain third complementarity-determining region // Journal of Virology. - 2010. - V. 84. - N 6. - P. 2955-2962.
14. Патент РФ № 2475264, кл. A61K 39/395, C07K 14/155, опубл.: 20.02.2013.
15. Патент РФ № 2214274, кл. А 61К 39/21, опубл. 20.10.2003.
16. Патент US 2008/0306224 A1, кл. C07K 14/00, опубл. 11.12.2008.
17. Патент РФ № 2317107, кл. C12N 15/48, опубл. 20.07.2007 (прототип).
18. Eroshkin AM, Karginova EA, Gileva IP, Lomakin AS, Lebedev LR, Kamyinina TP, Pereboev AV, Ignat'ev GM. Design of four-helix bundle protein as a candidate for HIV vaccine // Protein engineering. - 1995. - V. 8 - N. 2. - P. 167-173.
19. Карпенко Л.И., Бажан С.И., Богрянцева М.П., Рындюк Н.Н., Гинько З.Т., Кузубов В.И., Лебедев Л.Р., Каплина О.Н., Регузова А.Ю., Рыжиков А.Б., Усова С.В., Орешкова С.Ф., Нечаева Е.А, Даниленко Е.Д., Ильичев А.А. Комбинированная вакцина против ВИЧ-1 на основе искусственных полиэпитопных иммуногенов: результаты I фазы клинических испытаний // Биоорганическая химия. - 2016. - Т. 42. - N 2. - С. 191.
20. Chikaev A.N., BakulinaA.Yu., Burdick R.C., Karpenko L.I., Pathak V.K., Ilyichev A.A. Selection of peptide mimics of HIV-1 epitope recognized by neutralizing antibody VRC01 // PLOS ONE. - 2015. - V. 10. - N 3. - P. e0120847.
21. Hansted JG, Pietikäinen L, Hög F, Sperling-Petersen HU, Mortensen KK. Expressivity tag: a novel tool for increased expression in Escherichia coli // Biotechnology. - 2011. - V. 155. - N. 3. - P. 275-283.
22. Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование. - М.: Мир. - 1984.
23. Sambrook, J., Fritsch, E. F., & Maniatis, T. Molecular cloning. // New York : Cold spring harbor laboratory press. - 1989. - V. 2. - P. 14-23.
24. Holmes D.S., Quigley M. A rapid boiling method for the preparation of bacterial plasmids. // Analytical biochemistry. - 1981. - V. 114. - N. 1. - P. 193-197.
25. Montefiori D.C. Evaluating neutralizing antibodies against HIV, SIV, and SHIV in luciferase reporter gene assays // Current Protocols in Immunology. - 2005. - Chapter 12. - Unit 12.11.
26. Laemmli U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4 // Nature. - 1970. - V. 227. - P. 680-685.



Изобретение относится к области медицины, биотехнологии и генной инженерии. Предложен рекомбинантный химерный полипептид-иммуноген, включающий консервативные Т- и В-клеточные эпитопы вируса ВИЧ-1 и последовательно расположенные пептидные фрагменты белков р24, gp41, gp120, узнаваемые широконейтрализующими антителами 10e8, 2F5, VRC01. Изобретение обеспечивает повышение ВИЧ-специфического иммунного ответа за счет включения в состав полипептида-иммуногена nTBI уникальных линейных имитаторов конформационных эпитопов, узнаваемых широконейтрализующими антителами. 6 ил., 5 пр.
Рекомбинантный химерный полипептид-иммуноген, включающий консервативные Т- и В-клеточные эпитопы вируса ВИЧ-1 и характеризующийся тем, что он содержит в качестве вакцинного антигена последовательно расположенные пептидные фрагменты белков р24, gp41, gp120, узнаваемые широконейтрализующими ВИЧ-1 антителами, причем химерный полипептид-иммуноген экспрессируется в клетках Е.coli, содержащих плазмидный вектор рЕТ21a, в который по сайтам рестрикции NdeI и XhoI вставлен фрагмент ДНК, кодирующий указанный рекомбинантный химерный полипептид, имеющий последовательность SEQ ID NO:1 и представляющий собой очищенный белок, несущий эпитопы и пептидные миметики эпитопов ВИЧ-1, узнаваемых широконейтрализующими антителами 10e8, 2F5, VRC01.
| ЧИКАЕВ А.Н | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Дисс | |||
| канд | |||
| биол | |||
| наук | |||
| Кольцово, 2015, 118 c | |||
| EROSHKIN A.M | |||
| et al | |||
| Design of four-helix bundle protein as a candidate for HIV vaccine | |||
| Protein Eng | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА 1 ТИПА | 2006 |
|
RU2317107C2 |
| УСТРОЙСТВО ДЛЯ АВТОМАТИЧЕСКОГО ПУСКА АСИНХРОННЫХ ДВИГАТЕЛЕЙ | 1929 |
|
SU22788A1 |

