Изобретение относится к области биотехнологии, в частности к медицине, иммунологии. Кандидатная рекомбинантная вакцина против ВИЧ-1 представляет собой комбинацию иммуногенов, включающую рекомбинантную плазмидную ДНК pcDNA-TCI, обеспечивающую экспрессию искусственного гена TCI в клетках эукариот, покрытую конъюгатом спермидин-полиглюкин с рекомбинантным полиэпитопным белком TBI. На модели лабораторных животных вакцина обеспечивает высокий титр ВИЧ-специфических антител, обладающих вируснейтрализующей активностью, а также стимулирует вирус специфический цитотоксический иммунный ответ.
Учитывая стремительное распространение эпидемии СПИДа как во всем мире, так и в России, задача создания вакцины против ВИЧ-инфекции имеет чрезвычайно актуальное значение. Вместе с этим по трудности создания вакцина против ВИЧ - наиболее сложная из всех, созданных до сих пор или создаваемых в настоящее время вакцин.
Известные стратегии создания вакцины против ВИЧ-1 основаны на использовании различных форм вирусного антигена, включая инактивированный вирус, модифицированный вирус, аттенуированный живой вирус, нативные белки, генноинженерные пептиды, а также рекомбинантные бактерии, вирусы и плазмидные ДНК [1]. Каждая из этих стратегий была опробована, однако проблемы создания анти-ВИЧ-1-вакцины до сих пор не решены.
При создании вакцин против таких сложных вирусов, как ВИЧ, необходимы нетрадиционные подходы [2]. Один из наиболее перспективных подходов к созданию эффективных и безопасных вакцин связан с идентификацией Т- и В-клеточных эпитопов в вирусных белках и созданием на их основе синтетических полиэпитопных анти-ВИЧ вакцин. Такие вакцины, как ожидается, будут свободны от многих дефектов, свойственных вакцинам, создаваемым на основе нативных субъединиц или живых аттенуированных и цельновирионных патогенов.
Однако, как правило, генно-инженерные вакцины, представляющие собой рекомбинантные полипептиды, обладают сравнительно небольшой молекулярной массой, от тысячи до нескольких десятков тысяч дальтон. Поэтому при вакцинации используют различные адъюванты, что не всегда желательно и вызывает побочные эффекты. Наиболее сильным из адъювантов является адъювант Фрейнда, который для людей использовать запрещено.
Поэтому для усиления иммуногенности вакцин, разрабатываемых для людей, желательно использовать другие подходы, которые исключают применение адьювантов. Один из таких подходов может быть на создании вакцинных конструкций, обладающих большой молекулярной массой, сравнимой с размерами вирусов.
Известны примеры повышения иммуногенности генно-инженерных рекомбинантных поливалентных антиВИЧ вакцинных конструкций путем введения в состав слитых белков аминокислотных последовательностей либо нескольких различных иммуногенных эпитопов ВИЧ-1 [3, 4, 5], либо эпитопов, кодирующих антигены различных патогенов - HbsAg, env HIV, HbcAg, B-субъединица холерного токсина [6].
Другим подходом к созданию вакцинных конструкций с молекулярной массой, сравнимой с молекулярной массой вируса, является разработка препаратов в виде искусственных частиц, несущих различные иммуногенные эпитопы ВИЧ-1 (gag, pol, env) в составе генома другого вирусного агента-бакуловируса [7, 8], ленти - или альфавируса [9], или слитого белка, выбранного из TNF, интерлейкина, GM CSF, онкоантигена [10].
Однако рассмотренные выше искусственные полиэпитопной вакцинные конструкции содержат ограниченное количество CTL-эпитопов ВИЧ-1 (не более 20) из ограниченного числа белков ВИЧ-1 (gag, pol, env), что не может гарантировать высокую эффективность этой вакцины. К тому же использование другого вируса в качестве вектора для доставки ДНК-вакцины не всегда индуцирует адекватный иммунный ответ в силу того, что в популяции существует коллективный иммунитет к вирусу-носителю, как это имеет место в случае использования аденовирусных векторов.
Известна кандидатная вакцина против ВИЧ-1 «ВИЧРЕПОЛ» (ГНЦ Институт иммунологии) [11, прототип]. Основной компонент вакцины - химерный белок rec (24-41), состоящий из фрагментов иммуногенных белков р24 и gp41 ВИЧ-1 [12], конъюгированный с иммуностимулятором - носителем полиоксидонием (фармакологически активное соединение с молекулярной массой 100 кД с выраженной иммуномодулирующей активностью). При иммунизации лабораторных животных данная вакцина вызывает наработку специфических антител к белкам р24 и gp41 ВИЧ-1, входящих в состав вакцины. Однако эти антитела обладают низкой вируснейтрализующей активностью: индекс нейтрализации составляет 54,9% при разведении иммунной сыворотки 1:50. Клеточный ответ был также относительно невысоким, как показали данные в реакции бласт-трансформации.
Технической задачей изобретения является создание безопасной и эффективной кандидатной рекомбинантной вакцины против ВИЧ-1, способной индуцировать как ВИЧ-1 специфический цитотоксический ответ, так и высокий уровень антител, обладающих вируснейтрализующей активностью. Чтобы исключить применение адьювантов, вакцинная конструкция должна по молекулярному весу приближаться к размерам вириона.
В литературе описан искусственный рекомбинантный полиэпитопный белок, получивший название TBI (Т and В cell epitopes containing immunogen) [13, 14]. TBI-четырех-α-спиральный искусственный белок, который содержит четыре Т-клеточных эпитопа и пять В-клеточных нейтрализующих эпитопа из белков ВИЧ-1 Env и Gag. У мышей и обезьян, иммунизированных белком TBI, регистрировали появление вирусспецифических антител, обладающих вируснейтрализующей активностью [13].
При разработке новых перспективных подходов, нацеленных на индукцию нейтрализующих антител, в ряде исследований были получены убедительные данные, свидетельствующие о том, что ответы цитотоксических Т-лимфоцитов (CD8+ CTL), ассоциированные с ВИЧ-инфекцией, могут быть важным компонентом эффективной вакцины против ВИЧ-1 [15, 16, 17, 18].
Для стимуляции ВИЧ-специфического цитотоксического ответа авторами из ГНЦ ВБ «Вектор» был проведен дизайн искусственного поли-CTL-эпитопного Т-клеточного иммуногена TCI (Т cell immunogen) - кандидата для использования в качестве ДНК-вакцины. Белок TCI состоит из 392 аминокислотных остатков и содержит более 80 оптимально отобранных эпитопов (как CD8+ CTL, так CD4+ Th) из основных вирусных белков Env, Gag, Pol и Nef [19, 20]. Проведена сборка гена, кодирующего белок TCI, и его клонирование в векторной плазмиде pcDNA3.1 [21]. При иммунизации лабораторных животных полученная ДНК-вакцинная конструкция pcDNA-TCI вызывает индукцию цитотоксических Т-лимфоцитов, при этом уровень вирус-специфических антител был незначительным [20].
Поставленная задача решается путем создания кандидатной вакцины «КомбиВИЧвак», которая представляет собой комбинацию иммуногенов, состоящую из рекомбинантной плазмидной ДНК pcDNA-TCI с геном, кодирующим искусственный белок-иммуноген TCI, содержащий набор большого числа цитотоксических эпитопов ВИЧ-1 и обеспечивающая экспрессию искусственного гена в клетках млекопитающих, покрытую конъюгатом спермидин-полиглюкин с рекомбинантным полиэпитопным белком TBI, содержащим консервативные вируснейтрализующие В- и Т-клеточные эпитопы ВИЧ-1.
Кандидатная рекомбинантная вакцина против ВИЧ-1 "КомбиВИЧвак" представляет собой мицеллоподобные частицы (фиг.2), содержащие в центре рекомбинантную плазмидную ДНК, кодирующую искусственный белок-иммуноген TCI, содержащий набор большого числа цитотоксических эпитопов ВИЧ-1, а оболочка состоит из конъюгата спермидин-полиглюкин с рекомбинантным белком TBI, содержащим консервативные Т- и В-клеточные вируснейтрализующие эпитопы ВИЧ-1.
Одна доза вакцины объемом 0,2 мл содержит:
- 75±5 мкг рекомбинантной плазмидной ДНК pcDNA-TCI;
- 50±5 мкг рекомбинантного белка TBI;
- 1,0±0,1 мкг конъюгата полиглюкина со спермидином;
- 50±2 мг/мл реополиглюкина;
- 1,35±0,05 мг/мл калия фосфорнокислого двузамещенного 3 водного;
- 5,84±0,20 мг/мл натрия хлористого.
При иммунизации животных созданной конструкцией индуцируется выработка специфических антител, обладающих вирус-нейтрализующей активностью, а также специфический цитотоксический (ЦТЛ) иммунный ответ. При этом в отношении гуморального ответа наблюдается эффект синергизма при объединении в одной конструкции двух В- и Т-клеточных иммуногенов (TBI и TCI). Уровень антител при иммунизации отдельно взятыми конструкциями только с TBI или только с pcDNA-TCI был достоверно ниже в эксперименте, чем при использовании "КомбиВИЧвак".
Изобретение иллюстрируется чертежами.
Фиг.1. Электрофоретический анализ чистоты белка TBI в 12% ПААГ.
Дорожка 1 - маркеры молекулярной массы.
Дорожка 2 - клетки рекомбинантного штамма Е.coli JМ103, продуцирующего белок TBI.
Дорожка 3 - очищенный белок TBI.
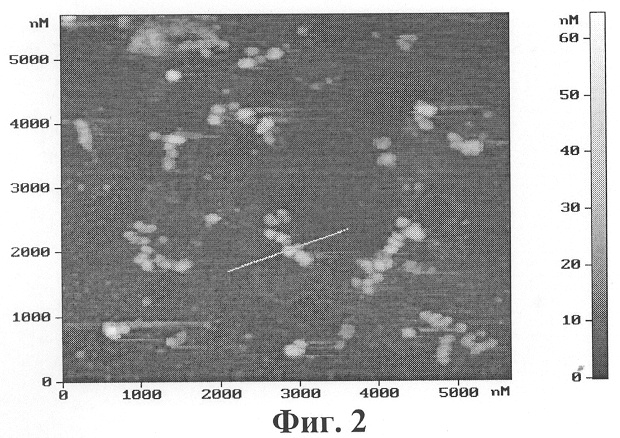
Фиг.2. Атомно-силовая микроскопия адсорбированных на пленку частиц "КомбиВИЧвак".
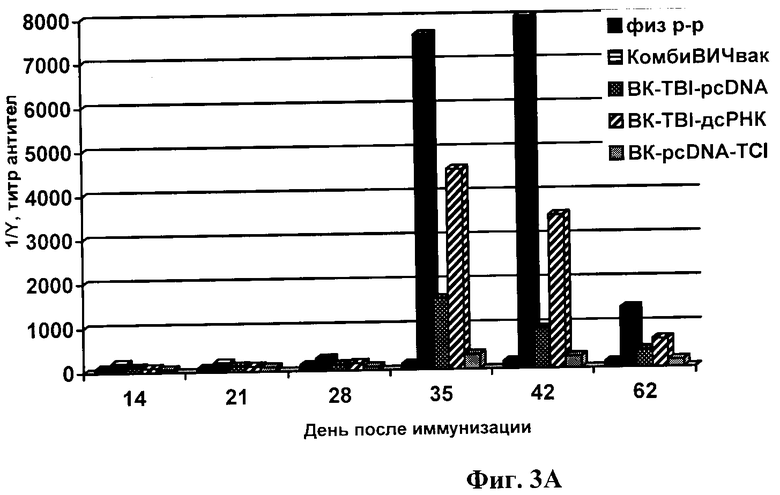
Фиг.3А. Титры антител в сыворотке иммунизированных животных. В качестве антигена в ИФА был использован рекомбинантный белок TBI.
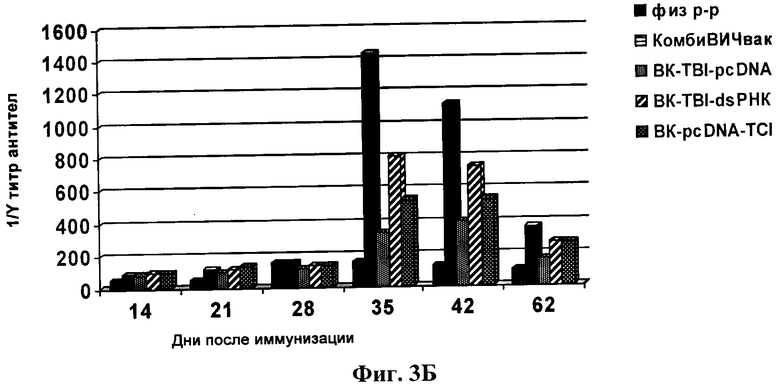
Фиг.3Б. Титры антител в сыворотке иммунизированных животных. В качестве антигена в ИФА был использован лизат ВИЧ-1.
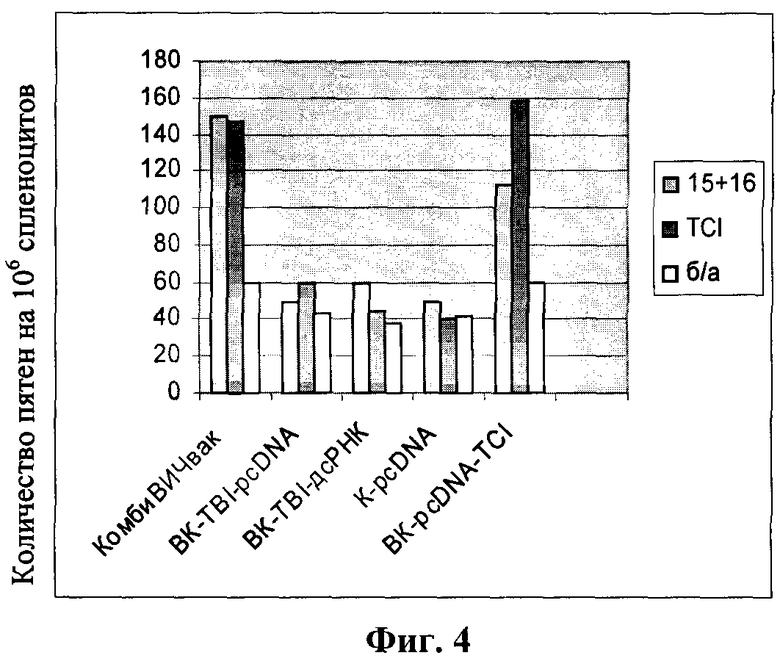
Фиг.4. Результаты оценки специфических цитотоксических Т-лимфоцитов у иммунизированных животных на 35 сутки после начала иммунизации.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Очистка рекомбинантной плазмидной ДНК pcDNA-TCI.
Наработка плазмиды
Для наработки плазмиды pcDNA-TCI в препаративных количествах использовали следующую схему:
1. Бактериальную культуру E.coli DH 5 альфа - pcDNA-TCI, содержащую плазмиду, засевали в 5 мл среды LB с ампициллином в концентрации 100 мг/мл;
2. Клетки растили ночь при 37°С при интенсивном перемешивании;
3. Ночную культуру клеток пересевали с разведением 1:1000 в 10 мл среды LB (концентрация ампициллина 100 мг/мл);
4. Клетки растили 6-8 часов при 37°С при интенсивном перемешивании;
5. Полученную культуру клеток пересевали в четыре колбы, содержащие по 150 мл среды LB (концентрация ампициллина 100 мг/мл) с разведением 1:100;
6. Клетки растили ночь (16 часов) при 37°С при интенсивном перемешивании;
7. Клетки собирали центрифугированием при 4000 об/мин в течение 20 минут;
8. Удаляли надосадочную жидкость и повторяли центрифугирование при 4000 об/мин в течение 2 минут, удаляли остатки жидкости;
9. Полученную биомассу замораживали в центрифужных стаканах при -60°С и затем переносили в индивидуальные пакеты для хранения.
Очистка плазмиды
Применяли метод очистки плазмидной ДНК, основанный на использовании Wizard® PureFection Plasmid DNA Purification System (Promega). Эта система включает запатентованную смолу для очистки от эндотоксинов, которая позволяет очистить плазмидную ДНК до низкого уровня загрязнения эндотоксинами.
Методы контроля качества препаратов плазмидной ДНК
При контроле качества плазмидной ДНК использовали методы, которые предусматривают оценку соответствия препарата исходной форме и его чистоты. Были использованы следующие методы:
1. Рестрикционный анализ с последующим электрофорезом в 1% агарозном геле. Рестрикцию проводили с помощью эндонуклеаз рестрикции (Fermentas, Литва и Сибэнзим, Россия) в стандартных условиях, рекомендованных фирмой-производителем.
2. Для оценки чистоты препарата исследовали спектр его оптического поглощения в диапазоне длин волн 230-280 нм. Форма спектра и отношения оптических плотностей А260нм/А230нм и А260нм/А280нм характеризуют чистоту препаратов ДНК от примесей белков и полисахаридов. В норме эти параметры должны быть не ниже 1,9 и 1,75, соответственно.
Пример 2. Очистка рекомбинантного белка TBI
В работе использовали штаммы Escherichia coli SG 200-50/p TBI [22].
Приготовление сред, ферментативные реакции, электрофорез в геле агарозы и ПААГ, трансформацию клеток E.coli проводят, как описано в руководстве Маниатиса и соавт. [23].
Для наработки биомассы рекомбинантных клеток выросшие на селективной (Ар) среде клоны вносят в 100 мл L-бульона; подращивают до оптической плотности Д550=1;0 о.е., добавляют ампициллин до конечной концентрации 500 мкг/мл и инкубируют в течение 2 ч для лизиса безплазмидных клеток. После этого засевают 1,6 л L-бульона (8 колб по 200 мл). Индукцию биосинтеза целевого белка проводят добавлением в среду налидиксовой кислоты до конечной концентрации 0,1 мМ. Рекомбинантый белок recA-TBI накапливается в клетках в нерастворимой форме, в тельцах включения.
Для очистки белка recA-TBI 3 г клеток суспендируют в 15 мл буфера А следующего состава: 10 мМ Трис-HCl (рН 8,3), 0,1 мМ CuCl2, 0,1 ZnCl2. Суспензию подвергают обработке ультразвуком до снижения оптической плотности суспензии при длине волны 550 нм до 50% от исходной. Тельца включения осаждают центрифугированием и дважды промывают буфером А. Гибридный белок экстрагировали из осадка 6 М раствором мочевины в буфере А в течение 2 ч, удаляют осадок центрифугированием, и затем проводят ренатурацию целевого белка, последовательно инкубируя при концентрациях мочевины 3 М (5 ч) и 1,5 М (16 ч). После этого проводят диализ против физиологического раствора. Оценку чистоты выделенного белка проводят с помощью электрофореза по Лэммли. Результаты анализа представлены на фиг.1.
Пример 3. Конструирование вакцины КомбиВИЧвак
Центральным ядром кандидатной вакцины "КомбиВИЧвак" является плазмида pcDNA-TCI, а оболочкой является конъюгат спермидина-полиглиглюкина-TBI.
Для получения оболочки 15 мг (0,25 мкМ) полиглюкина (60000 Д) обрабатывают 0,05 М раствором периодата натрия в течение 20-30 мин и затем отделяют гель-фильтрацией на колонке с сефадексом G-50 в 20 мМ фосфатном буфере (рН 7,6). После этого в раствор активированного полиглюкина вносят 5,5 мг белка TBI (0,25 мкМ) и 0,4 мг спермидина (2,6 мкМ). Смесь инкубируют в течение 5 ч, затем добавляют боргидрид натрия, перемешивают еще 2 ч и образовавшийся конъюгат отделяют гель-фильтрацией на сефадексе G-50, используя 100 мМ Трис-HCl (рН 8,3). Так как исходные компоненты (TBI и спермидин) вносились в соотношении на 0,25 мкМ полиглюкина 0,25 мкМ TBI и 2,6 мкМ спермидина, то можно предположить, что и полученный конъюгат содержит их в соотношении на одну молекулу полиглюкина - одна молекула TBI и десять молекул спермидина.
Для сборки вакцины к раствору, содержащему 7,5 мг pcDNA-TCI (около 1,8 нМ) в физиологическом растворе добавляют полисахаридный конъюгат, смесь инкубируют 2 ч при 4°С. Полученную конструкцию отделяют от несвязавшихся компонентов гель-фильтрацией на сефарозе CL-6B в физиологическом растворе.
Исследование физических свойств полученной молекулярной конструкции
По данным атомно-силовой микроскопии препарат молекулярной конструкции содержал шарообразные (мицеллоподобные) частицы, имеющие в основном диаметр от 10 до 25 нм (фиг.2). Встречались также более крупные частицы, диаметр которых достигал 50-100 нм. Возможно, это обусловлено тем, что полисахаридная вставка содержит на поверхности не одну молекулу спермидина, а десять, и происходит объединение ряда частиц. Размер полученной конструкции сравним с размером полиовируса и вируса полиомы и приближается к размеру ВИЧ.
Пример 4. Биологические испытания молекулярной конструкции
Мыши. Биологические испытания вакцинной молекулярной конструкции "КомбиВИЧвак" проводили на мышах линии BALB/c весом 18-20 грамм. Животные содержались на стандартном рационе. Все манипуляции с животными в эксперименте проводили с применением седативных средств в соответствии с ветеринарным законодательством.
Иммунизация. Для изучения иммуногенности вакцины "КомбиВИЧвак" животных (по 5 мышей в каждой группе) иммунизировали внутримышечно двукратно с интервалом 28 дней по одной дозе вакцины.
В качестве контроля использовали следующие вакцинные конструкции (ВК):
а) искусственные мицеллоподобные частицы, содержащие в центре векторную плазмидную ДНК pcDNA3.1, покрытую конъюгатом спермидин-полиглюкин с рекомбинантным полиэпитопным белком TBI (45 мкг по белку TBI) - BK-TBI-pcDNA;
б) искусственные частицы, содержащие в центре двуцепочечную дрожжевую РНК (Ридостин), покрытую конъюгатом спермидин-полиглюкин с рекомбинантным полиэпитопным белком TBI (45 мкг по белку TBI) - BK-TBI-дсРНК;
в) искусственные частицы, содержащие в центре рекомбинантную плазмидную ДНК pcDNA-TCI, обеспечивающую экспрессию искусственного гена TCI в клетках эукариот, и покрытую конъюгатом спермидин-полиглюкин (75 мкг по плазмидной ДНК) - ВК-TCI-pcDNA;
г) искусственные частицы, содержащие в центре только векторную плазмидную ДНК pcDNA 3.1 - K-pcDNA (контроль);
д) физиологический раствор.
На 14, 21, 28, 35, 42 и 62 сутки после начала иммунизации у мышей всех групп из ретроорбитального синуса забиралась кровь. После удаления центрифугированием сгустка отбиралась сыворотка и закладывалась на хранение в холодильник (-20°С). В конце эксперимента специфические анти-ВИЧ-антитела одновременно были определены во всех полученных сыворотках в ИФА.
Определение титра антител в сыворотках крови иммунизированных мышей с рекомбинантным белком TBI
Сыворотки иммунизированных и интактных мышей (отрицательный контроль) в двукратных разведениях, начиная с 1:10, вносили в лунки иммунологических планшетов Costar с орбированными на них рекомбинантным белком TBI (1 мкг на лунку). ИФА проводили по стандарной методике [24]. В работе использовали конъюгат анти-IgG мыши и ПХ.
Определение титра антител в сыворотках крови иммунизированных мышей на иммуносорбенте с антигеном лизата ВИЧ-1
Антиген лизата вируса ВИЧ-1 (ФГУН ГНЦ ВБ «Вектор») сорбируют в 0,05М карбонат-бикарбонатном буфере рН 9,6±0,1 на иммунологические планшеты (Costar или Nunc Maxi Sorp) в конечной концентрации 1 мкг/лунку и в объеме 100 мкл при 4°С в течение 15 ч. Затем в лунки подготовленного иммуносорбента вносят разведения сывороток крови иммунизированных и интактных мышей. Для проявления комплекса антиген-антитело используют конъюгат анти-IgG мыши с ПХ. Результаты экспериментов приведены на фиг.3.
Из данных, представленных на фиг.3, видно, что 2-кратная иммунизация мышей вакцинными конструкциями, содержащими специфические иммуногены TCI и TBI, сопровождалась образованием антиВИЧ-специфических IgG, однако, величина гуморального ответа у мышей из разных групп различалась (фиг.3). Максимальное значение индукции синтеза IgG регистрировалось при иммунизации кандидатной вакциной «КомбиВИЧвак», которое значительно превышало соответствующий показатель при введении конструкций, содержащих только один из иммуногенов (либо BK-TBI-pcDNA, либо BK-TCI-pcDNA). Это было показано в ИФА при использовании в качестве антигена как рекомбинантного белка TBI, так и лизата ВИЧ-1 (фиг.3А и фиг.3Б). Вместе с тем, иммунный ответ в случае конструкции BK-TBI-pcDNA (TBI без TCI) был выше, чем при иммунизации ВК-TCI-pcDNA (TCI без TBI). Тот факт, что конструкция BK-TCI-pcDNA обеспечивала наименьший титр антител можно объяснить тем, что она спроектирована для индукции Т-клеточного ответа и поэтому, содержит в основном CTL-эпитопы [19].
В группе мышей, иммунизированных K-pcDNA (контроль), гуморальный иммуный ответ был на уровне фона, также как и в группе, иммунизированной физиологическим раствором.
Способностью эффективно индукцировать специфический гуморальный иммунный ответ кроме «КомбиВИЧвак» также обладает конструкция BK-TBI-дсРНК, содержащая дсРНК - сильный неспецифический индуктор интерферона, который является медиатором иммунного ответа. Величина ответа в случае ВК-TBI-дсРНК значительно превышала ответ иммунной системы на BK-TBI-pcDNA (фиг.3), однако была достоверно ниже, чем ответ на «КомбиВИЧвак».
Тот факт, что максимальный иммунный ответ наблюдается в случае иммунизации животных «КомбиВИЧвак», можно объянить эффектом синергизма TCI и TBI. Эффект синергизма может быть связан с наличием в белках TBI и TCI разных Т-хелперных эпитопов [13, 19], которые, как известно, усиливают гуморальный иммунный ответ за счет стимуляции пролиферации и дифференцировки В-клеток.
Определение количества CTL лимфоцитов на основе реакции ELISPOT
При постановке реакции ELISPOT на первом этапе проводят сорбцию анти-INF-γ MAT с концентрацией 5 мкг/мл на лунку 96-луночного планшета ImmunoSpot M200. После инкубации в течение 12 ч при 4°С каждую лунку дважды промывают раствором PBS и блокируют средой RPMI 1640, содержащей 10% фетальную бычью сыворотку в течение 2 ч. В качестве клеток-эффекторов используют спленоциты иммунизированных животных в концентрации 106/мл. Для стимуляции продукции INF-γ суспензией клеток используют рекомбинантный белок TCI (1 мкг/мл) и два Н-2d-специфически пептида: N15 (DRVIEVVQGAYRAIR), N16 (KQIINMWQEVGKAMYA), которые содержатся в последовательности белка TCI. Для оценки неспецифической продукции используют пептид ЕНЕС (отрицательный контроль). Клетки культивируют в присутствии 5% СО2 при 37°С в течение 24 ч. INF-γ-секретирующие клетки визуализируют, используя 0,5 мкг/мл биотинилированнных анти-INF-γ антитела и 0,25 мкг/мл конъюгата Avidin-HRP. Окрашивание производят добавлением субстратов для пероксидазы (4-хлор-1-нафтол, диаминобензидин фосфат). Реакцию останавливают удалением реагентов, лунки промывают 3 раза дистиллированной водой. Подсчет количества INF-γ-продуцирующих клеток осуществляют с помощью микроскопа. Результаты экспериментов приведены на фиг.4.
Формирование ЦТЛ регистрировалось по антиген-зависимому выбросу IFN-γ в тесте ELISpot. В настоящей работе было показано, что иммунизация конструкциями, содержащими ген TCI (КомбиВИЧвак и BK-pcDNA-TCI) сопровождается формированием клонов ЦТЛ к 35 сут (фиг.4). В группах животных, иммунизированных BK-TBI-pcDNA и K-pcDNA, клеточный ответ был на уровне отрицательного контроля (группы животных, иммунизированных физраствором). В случае иммунизации вакцинными конструкциями «КомбиВИЧвак» и BK-TCI-pcDNA ответы ЦТЛ практически не различаются. Это означает, что в отличие от стимуляции гуморального иммунного ответа в случае индукции Т-клеточного ответа компоненты комбинированной вакцины «КомбиВИЧвак» (TCI и TBI) не вызывают эффекта синергизма. Вероятно это связано с тем, что основной вклад в индукцию Т-клеточного ответа вносит белок TCI, поскольку он содержит около 80-ти клеточных эпитопов (как ЦТЛ, так и Т-хелперных) [19], включая и те ЦТЛ эпитопы, которые содержит TBI. Поэтому, в случае индукции Т-клеточного иммунитета КомбиВИЧвак, основной иммуногенный потенциал сосредоточен в белке TCI.
Пример 5. Оценка вируснейтрализующей активности сывороток
Реакция вирусной нейтрализации
Реакцию нейтрализации проводили, используя дозу вируса равную 100 TCID50, с штаммом ВИЧ-1EVK (получен из Института вирусологии им. Д.И.Ивановского, Москва, депонирован в Государственной коллекции вирусов №4005).
Для реакции вирус-нейтрализации были выбраны сыворотки животных, иммунизированных «КомбиВИЧвак», полученные на 35 сутки после начала иммунизации. В качестве контролей использовали сыворотку ВИЧ-инфицированного пациента (получена и охарактеризована в Новосибирском государственном областном центре по профилактике и борьбе со СПИДом и инфекционными заболеваниями) и сыворотку здорового донора, не содержащего антитела к ВИЧ-1. Сыворотки иммунизированных и интактных животных и контрольные сыворотки (ВИЧ-инфицированного пациента и здорового донора) инактивировали на водяной бане при 56°С в течение 30 мин. В лунках 96-луночного планшета Cell-Cult смешивали равные объемы суспензии вируса и предварительно приготовленных рабочих разведении сывороток 1:10, 1:50 и 1:100. После часовой инкубации при 37°С инфицировали этими смесями чувствительные клетки МТ-4 с концентрацией 2 млн кл/мл, используя по 3 лунки 96-луночного культурального планшета на каждый вариант. Адсорбцию смесей проводили в течение 1 часа при 37°С в атмосфере 5% СО2, после чего трижды отмывали инфицированные и контрольные клетки питательной средой RPMI-1640 без добавок путем центрифугирования планшетов при 1600g в течение 10 мин. Затем разводили клеточную суспензию полной ростовой питательной средой RPMI-1640 с добавлением 10% сыворотки плода коров до посевной концентрации 500 тыс кл/мл.
Планшеты инкубировали при 37°С в СО2-инкубаторе в течение 4 суток. По окончании инкубации определяли количество вирусспецифического белка р24 в каждой пробе методом ИФА с использованием коммерческой тест-системы "ВектоВИЧ-1-р24-подтверждающий тест" производства «ЗАО «Вектор-Бест».
Результаты приведены в таблице. В последней колонке указано ингибирование накопления вирусного белка р24. За нулевую точку (т.е. отсутствие ингибирование) принимается цифра по р24 белку 560 нг/мл (см. строку МТ-4 (ВИЧ)-контроль). Эта точка взята на 4-е сутки после заражения вирусом клеток без добавления сывороток животных.
Из приведенных в таблице данных видно, что сыворотки мышей, иммунизированных препаратом "КомбиВИЧвак" активно подавляют размножение вируса в разведении 1:50 и в разведении 1:100. Эффективность подавления вируса мышиных сывороток сравнима с сывороткой человека, больного ВИЧ инфекцией.
Таким образом, кандидатная рекомбинантная вакцина "КомбиВИЧвак" индуцирует высокий гуморальный ответ, причем антитела высоко специфичны, они не только распознают антигены ВИЧ-1, но и способны с высокой эффективностью нейтрализовать вирус в системе in vitro.
Таким образом, использование предлагаемой молекулярной конструкции - кандиданой вакцины «КомбиВИЧвак» по сравнению с прототипом позволяет:
- индуцировать высокий уровень ВИЧ-1 специфических антител, обладающих вируснейтразующей активностью;
- индуцировать вирус-специфический цитотоксический иммунный ответ.
Предложенная кандиданая вакцина «КомбиВИЧвак» может быть использована для создания препаратов для профилактической и терапевтической профилактики ВИЧ-1 инфекции.
Литература
1. Berzofsky J.A., Ahlers J.D., Belyakov I.M. Strategies for designing and optimizing new generation vaccines. Nat Rev Immunol. 2001 Dec; 1 (3): 209-19.
2. Letvin N.L. Strategies for an HIV vaccine. J. Clin Invest. 2002, 110: 15-20.
3. Международная заявка WO №9109625, кл. А61K 39/395, опубл. 11.07.1991.
4. Международная заявка WO №9511701, кл. А61K 39/12, опубл. 04.05.1995 г.
5. Международная заявка WO №9939735, кл. А61K 39/385, опубл. 12.08.1999 г.
6. Патент США №6130082, кл. С12N 1/21, опубл. 10.10.2000 г.
7. Заявка ЕПВ №449116, кл. А61K 39/21, опубл. 02.10.1991 г.
8. Патент США №5443828, кл. А61K 39/21, опубл. 22.08.1995 г.
9. Патент США №6602705, кл. С12N 15/00, опубл. 05.08.2003 г.
10. Патент США №6099847, кл. А61K 39/21, опубл. 08.08.2000 г.
11. Сидорович И.Г., Николева И.А., Шевалье А.Ф., Коробова С.В., Игнатьева Г.А., Карамов Э.В., Корнеева Г.В., Павлова Т.В., Апритян С.А., Некрасов А.В., Пучкова И.Г., Савинова И.В., Иванова А.Г., Губима Г.О., Воробьева М.С. Перспективы контроля эпидемии ВИЧ/СПИД в России: первая отечественная вакцина против ВИЧ/СПИД ВИЧРЕПОЛ готова для проведения I фазы клинических испытаний. // Физиология и патология иммунной системы. 2004. - №2. Стр.74-84.
12. Патент РФ №02214274, кл. А61K 39/21, опубл. 20.10.2003 г.
13. Eroshkin A.M., Karginova E.A., Gileva I.P. et al. // Protein Enginiring. - 1995. - V.8. - N.2. - P.167-173.
14. Loktev V.B., Ilyichev A.A., Eroshkin A.M., Karpenko L.I., Pokrovsky A.G., Pereboev A.V., Svyatchenko V.A., Ignat′ev G.M., Smolina M.I., Melamed N.V., Lebedeva C.D., Sandakhchiev L.S. Design of immunogens as components of a new generation of molecular vaccines. // J Biotechnol. - 1996. - Jan 26; 44 (1-3). - P.129-37.
15. Yang O.O., Kalams S.A., Trocha A., Cao H., Luster A., Johnson R.P., Walker B.D. Suppression of human immunodeficiency virus type 1 replication by CD8+ cells: evidence for HLA class I-restricted triggering of cytolytic and noncytolyticmechanisms. JVirol 1997, 71: 3120-8.
16. Ogg G.S., Jin X., Bonhoeffer S., Dunbar P.R., Nowak M.A., Monard S., Segal J.P., Cao Y., Rowland-Jones S.L., Cerundolo V., Hurley A., Markowitz M., Ho D.D., Nixon D.F., McMichael A.J. Quantitation of HIV-1-specific cytotoxic Т lymphocytes and plasma load of viral RNA. Science 1998; 279: 2103-6.
17. Wagner E., Yang O.O., Garcia-Zepeda E.A., Ge Y., Kalams S.A., Walker B.D., Pasternack M.S., Luster A.D. Beta-chemokines are release from HIN-1-specific Т cells granules complexed to proteoglycans. Nature 1998; 391: 908-11.
18. Price D.A., Sewell A.K., Dong T., Tan R., Goulder P.J., Rowland-Jones S.L., Phillips R.E. Antigen-specific release of beta-chemokines by anti-HIV-1 cytotoxic T lymphocytes. Cur Biol 1998; 8: 355-8.
19. Патент РФ №2238946, кл. С07К 14/16, опубл. 27.10.2004 г.
20. Bazhan S.I., Belavin P.A., Seregin S.V., Danilyuk N.K., Babkina I.N., Karpenko L.I., Nekrasova N.A., Lebedev L.R., Ignatyev G.M., Agafonov A.P., Poryvaeva V.A., Aborneva I.V., Ilyichev A.A. Designing and engineering of DNA-vaccine construction encoding multiple CTL epitopes of major HIV-1 antigens // Vaccine. - 2004, Apr 16; (13-14): 1672-82.
21. Патент РФ №2248396, кл. С12N 15/48, опубл. 20.03.2005 г.
22. Лебедев Л.Р. Ерошкин A.M. Гшева И.П. «Способ получения синтетического белка - кандидата в вакцины против HIV-1». // Биотехнология. 1997. №7-8. C.38-42.
23. Маниатпис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование. М.: Мир. - 1984.
24. Karpenko L.I., Lebedev L.R., Ignatyev G.M., Agafonov A.P., Poryvaeva V.A., Pronyaeva T.R., Ryabchikova E.I., Pokrovsky A.G., Ilyichev A.A. Construction of artificial virus-like particles exposing HIV epitopes, and the study of their immunogenic properties // Vaccine. - 2003 - Jan 17; 21 (5-6): 386-92.
Оценка вирус-нейтрализующей активности сывороток

Изобретение относится к области биотехнологии, в частности к медицине, иммунологии. Рекомбинантная вакцина против ВИЧ-1 представляет собой комбинацию иммуногенов, в виде мицеллоподобных частиц, включающая рекомбинантную плазмидную ДНК pcDNA-TCI, покрытую конъюгатом спермидина-полиглюкина с рекомбинантным белком TBI, содержащим консервативные Т- и В-клеточные вируснейтрализующие эпитопы, в стабилизирующей смеси. При иммунизации созданной вакцины индуцируется выработка специфических антител, обладающих вируснейтрализующей активностью, а также специфический цитотоксический иммунный ответ. 1 табл., 5 ил.
Рекомбинантная вакцина против вируса иммунодефицита 1 типа, представляющая собой комбинацию иммуногенов в виде мицеллоподобных частиц, включающая рекомбинантную плазмидную ДНК pcDNA-TCI, покрытую конъюгатом спермидина-полиглюкина с рекомбинантным белком TBI, содержащим консервативные Т- и В-клеточные вируснейтрализующие эпитопы, в стабилизирующей смеси.
| СИДОРОВИЧ И.Г | |||
| и др | |||
| Перспективы контроля эпидемии ВИЧ/СПИДа в России | |||
| Первая отечественная вакцина против ВИЧ/СПИД ВИЧРЕПОЛ готова для проведения I фазы клинических испытаний | |||
| Физиология и патология иммунной системы | |||
| - М., 2004, т.8, №2, стр.74-84 | |||
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PCDNA-TCI, ОБЕСПЕЧИВАЮЩАЯ ЭКСПРЕССИЮ ИСКУССТВЕННОГО ГЕНА TCI В КЛЕТКАХ ЭУКАРИОТ, И РЕКОМБИНАНТНЫЙ АТТЕНУИРОВАННЫЙ ШТАММ БАКТЕРИЙ SALMONELLA ENTERITIDIS E-23/PCDNA-TCI КАК КАНДИДАТ ДЛЯ КОНСТРУИРОВАНИЯ ЖИВОЙ ДНК-ВАКЦИНЫ ПРОТИВ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА | 2003 |
|
RU2248396C2 |
| ИСКУССТВЕННЫЙ БЕЛОК-ИММУНОГЕН TCI, СОДЕРЖАЩИЙ МНОЖЕСТВЕННЫЕ CTL-ЭПИТОПЫ ОСНОВНЫХ АНТИГЕНОВ ВИЧ-1, ИСКУССТВЕННЫЙ ГЕН TCI, КОДИРУЮЩИЙ ПОЛИЭПИТОПНЫЙ БЕЛОК-ИММУНОГЕН TCI | 2002 |
|
RU2238946C2 |
| SERGEI I | |||
| BAZHAN, Designing and engineering of | |||

