Изобретение относится к новой вакцине против бешенства.
Вирус бешенства является рабдовирусом, содержащим пять протеинов, в котором один внешний протеин, гликопротеин, инициирует синтез нейтрализующих антител у привитых животных. Введение очищенного гликопротеина защищает животное от внешней инфекции. Наиболее используемые штаммы вируса бешенства, а именно штаммы CVS и SAD, которые происходят от штаммов SAD, таких как SAD Berne и SAD B19, описаны в "Rabies Viruses", H.F. Clark и T.I. Wictor-Strains of Human Viruses, изд. Majer and Plotkin Karger, Bale, 1972, стр. 177 - 182. Последовательность аминокислот гликопротеина штамма CVS описана Yelverton и др. в "Rabies virus glucoprotein analogs: biosynthesis in Escherichia coli" Science 219, 614 - 620.
Этот гликопротеин содержит два основных антигенных различных сайта, связанных с нейтрализацией вируса /сайты II и III/. Сайт III в положении 330 - 340 содержит аргинин 333, который определяет вирулентность этого штамма.
Последовательность аминокислот гликопротеина штамма SAD Berne не установлена полностью. Однако можно определить, что антигенный сайт III гликопротеина штамма SAD Berne идентичен этому же сайту гликопротеина штамма CVS.
Используемые в настоящее время вакцины против бешенства являются либо вакцинами, полученными из инактивированного вируса, либо вакцинами, состоящими из вирусных штаммов, вирулентность которых уменьшена, либо рекомбинантными вирусами /например, вакцина/.
Инактивация вируса может быть осуществлена различными способами, в частности химическими способами, такими как обработка формолом или β-пропиолактоном.
Главным недостатком способа получения вакцины является работа с вирулентными штаммами, что требует точного соблюдения условий работы, и создает риск отравления для работающего персонала.
С другой стороны, инактивированные вакцины, вводимые оральным путем, не обладают защитными свойствами.
Снижение вирулентности вирусных штаммов основано на хорошо известной методике; оно может быть осуществлено, например, последовательным пропусканием вирусных штаммов через хозяина другого вида штамма /кролика или мыши, например/ или через клеточную культуру. Получают таким образом штаммы, несвойственные первоначальному хозяину, и, следовательно, менее патогенные при сохранении их вакцинирующей способности.
Штаммы SAD, такие как SADB19 и SAD Berne, являющиеся доступными, являются ослабленными штаммами и были испытаны в Европе для вакцинации лисиц. Они могут быть смешаны с приманкой для орального введения. Однако эти штаммы проявляют патогенность на других видах животных и на человеке. Следовательно, они создают потенциальный риск отравления, что значительно уменьшает интерес к этим штаммам для оральной вакцинации.
Действительно, например, оральное введение штамма SAD Berne группе из 23 диких грызунов, выбранных среди Apodemus flavicolus et sylvaticus, Arvicola terrestris, Clethrionomys clareolus, Minotus agresti, вызывает смерть от бешенства двух животных.
Испытания на мышах также показывают, что штамм SAD Berne является патогенным для этого типа животных, как при внутричерепном, так и при внутримышечном введении, как это показывают кривые на фиг. 1a и 1b, на которых соответственно показано:
фиг. 1a - кривая смертности взрослых мышей в зависимости от доз, вводимых интрацеребральным путем (в единицах образующих пятна, ЕОП);
фиг. 1b - кривая смертности взрослых мышей в зависимости от доз, введенных внутримышечным путем (в единицах, образующих пятна).
При оральном пути введения у мышей отмечается 10%-ная смертность при дозе 105,5 ЕОП.
Штамм SAD Berne, применяемый как таковой в вакцинационной кампании на лисицах, представляет опасность для дикой фауны и человека. Это же относится и к штамму SAD B19.
Для устранения этого недостатка уже предлагалась авирулентная вакцина против бешенства. Эта вакцина, описанная в заявке на патент EP 350398, содержит авирулентный мутант штамма SAD вируса бешенства, который характеризуется тем, что вместо аргинина 333 гликопротеина он имеет аминокислоту, отличающуюся от лизина, например глицин, изолейцин или серин.
Этот мутант получают заменой одного нуклеотида в кодоне аргинина 333.
К сожалению, этот мутант может при простой обратной мутации вновь дать родительский штамм.
Следовательно, эта вакцина, используемая оральным путем, не всегда безопасна для животных других видов.
Настоящее изобретение относится к эффективной вакцине, которая позволяет устранить указанные недостатки.
Вакцина по изобретению включает авирулентный мутант штамма SAD вируса бешенства, который характеризуется тем, что вместо аргинина 333 гликопротеина он содержит природную аминокислоту, кодон которой отличается от кодонов, кодирующих аргинин двумя нуклеотидами.
Изобретение относится также и к способу получения авирулентного мутанта, указанного выше. Этот способ включает:
1) отбор, исходя из штамма SAD вируса бешенства, мутантов, которые не нейтрализованы моноклональными антителами, нейтрализующими вышеупомянутый штамм SAD, но не нейтрализующими штамм TAG1, описанный ниже;
2) отделение путем секвенирования области 333 гликопротеина мутантов, отобранных на стадии 1) от мутанта, содержащего в положении 333 лизин;
3) получение моноклонального антитела, которое одновременно нейтрализует вышеупомянутый штамм SAD и мутант, полученный на стадии 2), но не нейтрализует штамм TAG1;
4) проведение повторного отбора из мутантов, полученных на стадии 1) с помощью моноклонального антитела, полученного на стадии 3).
Моноклональные антитела, используемые при отборе мутантов по изобретению, получают слиянием клеток миеломы и клеток, производящих антивирусные антитела по методике гибридизации, описанной KOHLER et MILSTEIN, в журнале NATURE, 256, 495 - 497 /1975/, эта методика широко известна в настоящее время специалистам.
Согласно этой методике можно подвергать слиянию клетки, происходящие от различных видов, однако предпочтительными являются клетки животного одного и того же вида. Например, используют преимущественно, с одной стороны, клетки миеломы мыши и, с другой стороны, клетки селезенки мыши, предварительно иммунизированные штаммом вируса бешенства согласно протоколу, приведенному ниже.
В основном, этот способ гибридизации, описанный для клеток мышей, включает следующие стадии:
1) иммунизация мышей заданным количеством вируса, инактивированного β-пропиолактоном;
2) извлечение селезенки у иммунизированной мыши и отделение спленоцитов;
3) слияние полученных таким образом спленоцитов с клетками миеломы мыши в присутствии стимулятора слияния;
4) культивирование полученных гибридных клеток в избирательной среде, в которой не развиваются неслившиеся клетки миеломы, в присутствии усваиваемых элементов;
5) отбор клеток, вырабатывающих желаемые антитела и клонирование этих клеток.
Протокол иммунизации включает внутрибрюшинное введение мышам Balb-C 100 мкг инактивированного β-пропиолактоном вируса CV5 с адъювантом Фрейда и повторную инъекцию внутривенным путем за 4 дня до слияния клеток после периода отдыха в течение 1 месяца.
Спленоциты иммунизированных мышей собирают после извлечения селезенки классическим способом.
Клетки миеломы мышей, используемые для получения нейтрализующих моноклональных антител, являются клетками миеломы мышей Balb-C, происходящих из колонии SP2O. Эти клетки миеломы были отобраны с точки зрения их чувствительности к аминоптерину и культивированы в соответствующей среде, такой как основная среда Игла, модифицированная DULBECCO (Dulbecco Modified Eagle medium), называемая далее среда DMEM, к которой добавлено 15% сыворотки жеребенка.
Слияние клеток миеломы со спленоцитами осуществляют путем смешивания 5 • 107 клеток миеломы с 5 • 107 клеток селезенки иммунизированной мыши в присутствии стимулятора слияния, такого как, например, полиэтиленгликоль.
После инкубации при 37oC клетки промыват в среде DMEM и вновь суспензируют, затем культивируют в избирательной среде, предназначенной исключительно для роста гибридных клеток. Эта среда содержит гипоксантин, аминоптерин и тимидин.
Отбирают затем супернатанты культур, спустя 7 - 20 дней после слияния клеток, приводя во взаимодействие супернатанты с суспензией вируса CVS и удерживая антитела, которые нейтрализуют вышеупомянутую суспензию.
Термин "антитело, нейтрализующее вирус" обозначает такие антитела, которые при помещении их в суспензию данного вируса ингибируют его вирулентность.
Нейтрализующая способность моноклональных антител, полученных выше, определяется классическим способом, хорошо известным специалисту. Этот способ состоит в смешивании 100 мкл суспензии вируса с содержанием вируса 1000 ЕОП и 100 мкл супернатанта культуры гибридомы, в инфицировании культуры клеток этой смесью и, после 4 дней инкубирования, в подсчете лизированных пятен на агаре по методу, описанному BUSSEREAU et al., 1982, J. Virol. Meth. Vol. 4, стр. 277 - 282. Антитела являются нейтрализаторами, если они ингибируют образование пятен на агаре в вышеупомянутых условиях.
Среди этих антител отбирают затем антитела, которые не нейтрализуют авирулентный мутант TAG1, производный штамма CVS, представленный в Национальной Коллекции Культур Микроорганизмов (C.N.C.M. - Collection Nationale de Cultures de Micro-organismes) Института Пастера - Франция 12 апреля 1985 под N 1-433. Полученные моноклональные антитела, которые нейтрализуют штамм CVS, но не нейтрализуют авирулентный мутант TAG1, позволяют селекционировать авирулентные мутанты исходя из любого штамма вируса бешенства, который нейтрализуется этими моноклональными антителами.
Моноклональные антитела, полученные таким образом, являются антителами, нейтрализующими также штаммы SAD вируса бешенства. Они подходят также для осуществления первичного отбора по способу изобретения.
Секвенирование области 333 гликопротеина мутантов, отобранных на стадии 1) осуществляют по классической методике, хорошо известной специалисту [SANGER et al., 1977, Proc. Nat. Acad.Sci. USA, vol. 74, p. 5463 - 5467].
Это секвенирование позволяет изолировать мутант, который имеет в положении 333 гликопротеина лизин; этот мутант именуется далее "мутант SK".
Кодон /AAA/ этой аминокислоты /лизина/ отличается от кодона аргинина в положении 333 штамма SAD единственным нуклеотидом, причем кодон аргинина в положении 333 имеет строение AGA.
Далее переходят к получению моноклонального антитела, нейтрализующего одновременно штамм SAD и мутант, полученный выше.
Это моноклональное антитело получают удерживанием среди моноклональных антител, нейтрализующих SAD и не нейтрализующих TAG1, такого, который нейтрализует равным образом лизиновый мутант или мутант SK, полученные выше.
Это моноклональное антитело позволяет осуществить второй отбор /стадию 4/ способа по изобретению.
Патогенная возможность мутантов, полученных после второй селекции, определяется путем интрацеребральной инъекции 105 ЕОП взрослым мышам. Мутанты, которые не убивают при этой дозе и при этом методе введения, рассматриваются как авирулентные.
Мутанты по изобретению являются двойными авирулентными мутантами штамма SAD, в частности штамма SAD Berne, получаемыми двумя последовательными селекциями с помощью моноклональных антител, определенных выше.
Мутанты по изобретению могут, кроме того, заключать в себе другие мутации, как например мутацию, придающую устойчивость к моноклональным специфичным антителам антигенного сайта II, что позволяет распознать возможный ревертант вирулентности штаммов SAD, используемых для оральной вакцинации лис.
Мутанты по изобретению могут быть размножены на клетках почки новорожденного хомяка BHK 21, в присутствии среды GEM (основная среда минимум-модификации Глазгоу, выпускаемая фирмой FLOW) и 2%-ной сыворотки теленка при 33oC во влажной атмосфере с 5%-ным содержанием CO2.
Их охарактеризовывают обычными методами, например, определяя летальную дозу 50 /ЛД50/ на мышах, иммунофлуоресценцией или обсчитывая лизированные пятна на агаре. Они могут храниться при -70oC.
Анализ последовательности нуклеотидов гликопротеина этих мутантов показывает, что кодон аминокислоты в положении 333 отличается по крайней мере двумя нуклеотидами от всех кодонов, возможных для аргинина. Среди мутантов, которые отвечают этим условиям, предпочтительным особенно является мутант, имеющий в 333 глутаминовую кислоту, так как он хорошо размножается на клеточной культуре, менее патогенен для новорожденных мышей, и он обладает высокой защитной способностью.
Двойной мутант, несущий глутаминовую кислоту, кодон которой: GAA на месте аргинина в положении 333, полученный по вышеприведенному способу; исходя из штамма SAD Berne, называемый далее SAG2, представлен в Национальную Коллекцию Культур Микроорганизмов /C.N.C.M. - Collection Nationale de Cultures de Micro-organismes/ Института Пастера - Франция 9 июля 1992 под N 1-1238. Этот мутант содержит другую мутацию, а именно устойчивость к моноклональному специфическому антителу антигенного сайта II, что является дополнительным маркером штамма.
Далее представлено детальное описание изобретения, на примере мутанта SAG2, что однако не ограничивает изобретение только этим мутантом.
A - Исследование генетической стабильности штамма при проведении пассажей на мозге мышат.
103 ЕОП SAG2 вводят 6 мышатам в возрасте 4 дней. Когда животные заболевают /6/, их умерщвляют. Готовят 6 отдельных измельченных образцов; каждый образец оттитровывают и 30 мкл разбавленного 1/10 образца вводят 3 взрослым мышам /патогенный контроль/ и мышонку /следующий пассаж/, 6 мышат позволяют сделать 6 независимых серий из 3 пассажей.
Титры мозговых веществ в ЕОП /мл даны в таблице.
Все инъектированные взрослые особи /54 мыши/ после 1-го, 2-го и 3-го пассажей остались живы и показали отсутствие реверсии.
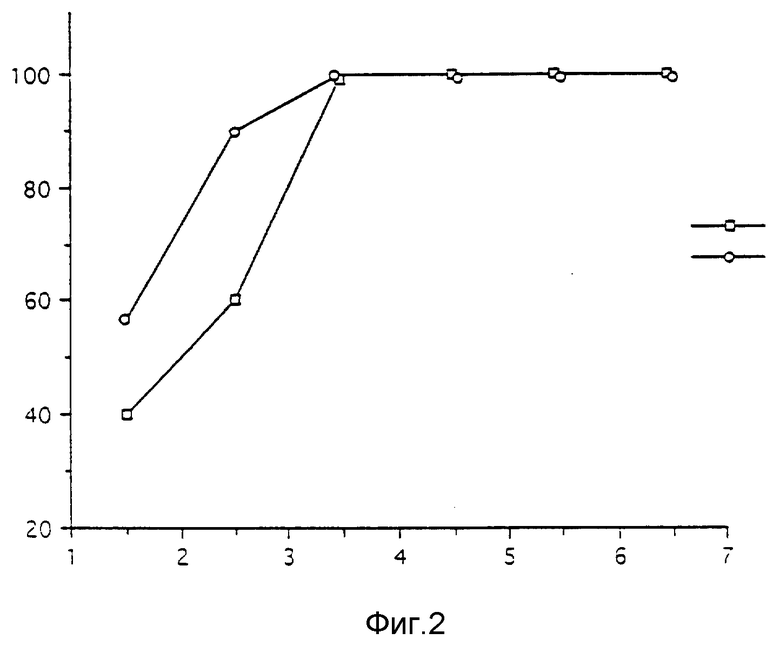
B - Защитная способность SAG2.
Мутант SAD2 вводят интрацеребральным путем мышам и определяют защитную способность мутанта в опыте внутримышечного введения 100 ЛД50 штамма CVS.
Полученные результаты представлены на фиг. 2, где графически показана защитная способность /%/ как функция количества введенного мутанта, выраженного в (ЕОП) UFP/мышь (log). Повторяют то же исследование с мутантом SK, имеющим в положении 333 лизин.
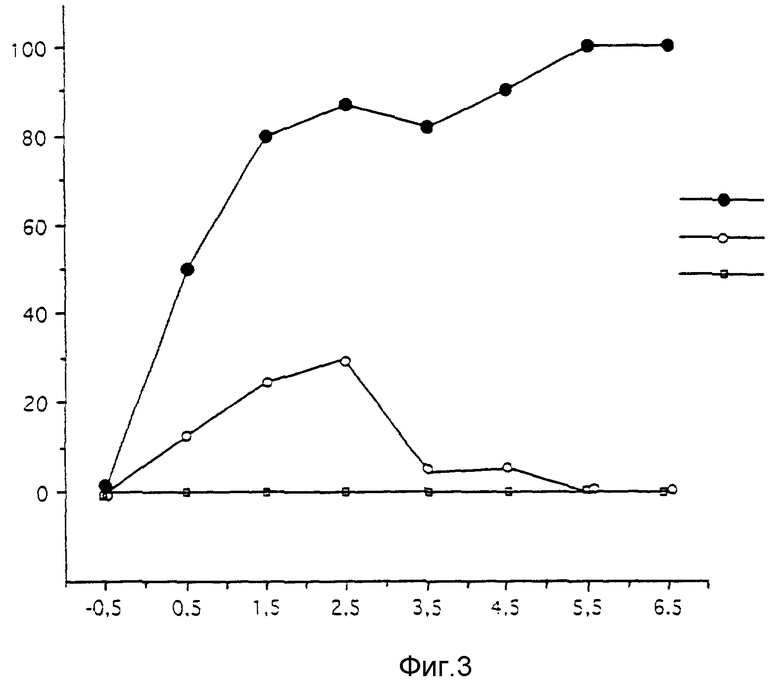
C - Патогенность SAG2 при интрацеребральном введении.
Вводят мутант SAG2 мышам в дозах от 10-0,5 до 106 ЕОП/мышь и не отмечают смертности в течение периода времени 28 дней.
Параллельно, осуществляют тот же опыт с вирусом SAD Berne или мутантом SK.
Результаты представлены на фиг. 3, где дан график зависимости % смертности /ось ординат/ от количества введенного мутанта мышам /ось абсцисс/.
Установлено, что мутант SAG2 не вызывает никакой смертности, тогда как мутант SK имеет слабую остаточную патогенность.
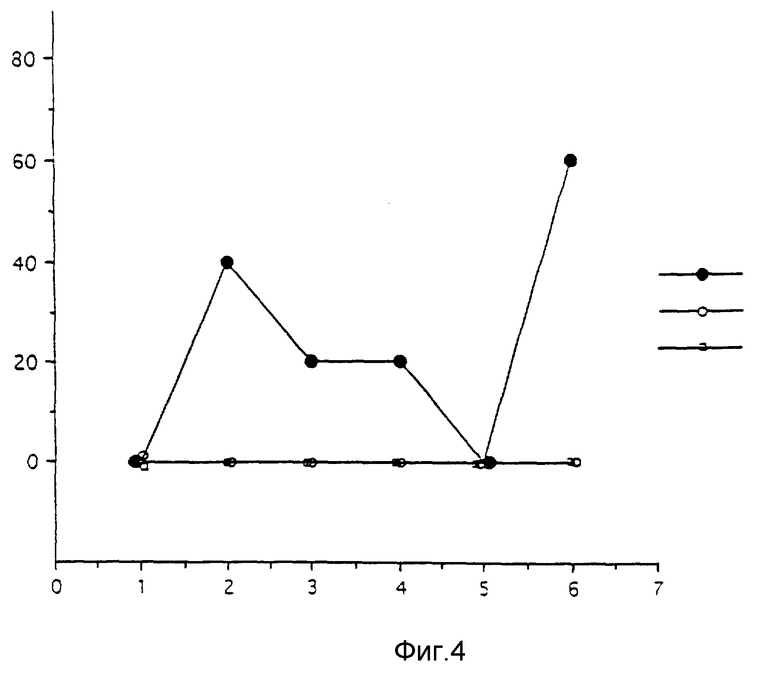
D - Патогенность SAG2 при внутримышечном введении.
Повторяют испытание, приведенное выше, но при внутримышечном введении. Полученные результаты представлены на фиг. 4, где приведен % смертности /ось ординат/ в зависимости от введенной дозы в ЕОП/мышь /ось абсцисс/.
Видно, что SAG2 и мутант SK не вызывают никакой смертности.
Мутанты по изобретению могут применяться в виде живой вакцины при всех обычно используемых способах введения при вакцинации и особенно при внутримышечном или оральном способе введения. Мутанты предварительно разбавляют в инертном фармацевтически пригодном растворителе, таком как физиологическая сыворотка.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ПЕРОРАЛЬНОЙ ВАКЦИНЫ ПРОТИВ ВИРУСА БЕШЕНСТВА | 2010 |
|
RU2432963C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ ВИРУСА БЕШЕНСТВА И ИХ СМЕСЬ | 2019 |
|
RU2822457C2 |
| ВАКЦИНА ПРОТИВ БЕШЕНСТВА | 2014 |
|
RU2712743C2 |
| ВАКЦИНА ПРОТИВ ВИРУСА БЕШЕНСТВА | 2018 |
|
RU2782350C2 |
| СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ ВИРУСА БЕШЕНСТВА НЕЙТРАЛИЗУЮЩИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА И НУКЛЕИНОВЫЕ КИСЛОТЫ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2001 |
|
RU2272809C2 |
| Ассоциированная вакцина против панлейкопении, калицивироза, вирусного ринотрахеита и бешенства кошек | 2024 |
|
RU2827230C1 |
| ИММУНИЗАЦИЯ ВЕКТОРОМ НА ОСНОВЕ ВИРУСА БЕШЕНСТВА, ЭКСПРЕССИРУЮЩИМ ЧУЖЕРОДНЫЙ БЕЛКОВЫЙ АНТИГЕН | 2013 |
|
RU2660566C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ИММУНОГЕННОЙ АКТИВНОСТИ ВАКЦИНЫ ПРОТИВ БЕШЕНСТВА | 2016 |
|
RU2616898C1 |
| Вакцина антирабическая инактивированная эмульсионная культуральная для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных | 2019 |
|
RU2722868C1 |
| Рекомбинантный слитый белок, специфически связывающий и нейтрализующий вирус бешенства, способ его получения и применение | 2023 |
|
RU2829507C1 |
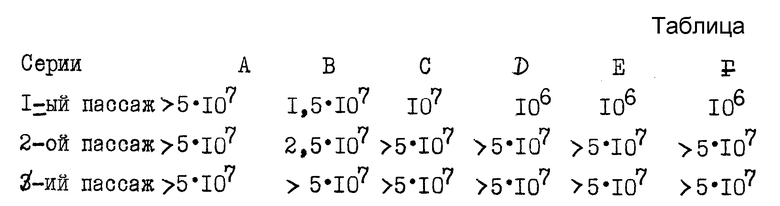
Изобретение предназначено для получения средства специфической профилактики против бешенства. Вакцина против бешенства содержит авирулентный мутант штамма SAD Berne вируса бешенства. Штамм характеризуется тем, что содержит в положении 333 природную глутаминовую кислоту, кодон которой отличается от кодонов аргинина по крайней мере двумя нуклеотидами. Штамм депонирован в Национальной Коллекции Культур Микроорганизмов и имеет регистрационный номер C. N.С.М. 1-1238. Штамм получен путем отбора с использованием моноклональных антител, нейтрализующих штамм SAD и нейтрализующих штамм ТAG 1. Штамм и вакцина на его основе характеризуются повышенной безопасностью и эффективностью. 3 с. и 2 з.п. ф-лы, 1 табл., 4 ил.
1) из штамма вируса бешенства C.N.C.M.No 1-1238 получают мутанты, которые не нейтрализуются моноклональным антителом, нейтрализующим штамм SAD, но не нейтрализующим штамм TAG1;
2) выделяют путем секвенирования положения 333 в гликопротеине мутантов, отобранных на стадии 1), мутант, имеющий в положении 333 лизин;
3) получают моноклональное антитело, которое одновременно нейтрализуют указанный штамм SAD и мутант, полученный на стадии 2), но не нейтрализует штамм TAG 1;
4) осуществляют второй отбор мутантов, полученных на стадии 1), с помощью моноклонального антитела, полученного на стадии 3).
| EP 0350398 A, 10.01.90 | |||
| US 4752474 C, 21.06.88 | |||
| СО АН СССР | 0 |
|
SU202142A1 |

