Изобретение относится к новому промотору, который является функциональным в растениях, более конкретно, к промотору, который контролирует экспрессию целевого гена предписанным образом. Оно также включает различные последовательности в направлении вверх от вышеупомянутого промотора, которые можно использовать в конструкции гибридных промоторов, имеющих целевую активность.
Предпосылки настоящего изобретения
Гены теплового шока (hsp - гены) известны в самых разнообразных организмах, включая дрожжи, дрозофиллы и некоторые растительные виды. Один класс генов теплового шока экспрессирует протеин в ответ только на тепловой стресс. Другой класс генов теплового шока имеет низкий исходный уровень активности, который значительно увеличивается при индукции нагреванием.
При создании гетерологического гена при помощи приемов генной инженерии в растениях, селекция промотора часто является решающим фактором. В то время, как может оказаться желательным экспрессировать некоторые гены только в ответ на определенное воздействие (стимул) или локализовать их экспрессию в некоторых тканях, может потребоваться, чтобы другие гены экспрессировались конститутивно, т.е. во всем растении в течение всего времени и в большинстве тканей. Ранее использовали промотор 35S из Мозаичного Вируса Цветной Капусты (CAMV) для конструктивной экспрессии гетерологических генов. Для регулирования и по другим причинам было бы желательно регулировать экспрессию гетерологического гена промотором, который не имел бы патогенной природы. Кроме того, использование растительного промотора может изменять уровень активности в конкретных тканях и может изменять спектр тканей, в которых достигают экспрессии, по сравнению с вирусными промоторами.
Описание изобретения
Настоящее изобретение относится в конструктивному промотору из цветной капусты (Brassica oleracca cv. "Delira") т. е. промотору, который будет экспрессировать гены все время в большинстве тканей и органов. Этот промотор может быть оперативно связан с любым целевым геном и он будет направлять экспрессию этого гена. Этот промотор можно отличить от промоторов ранее описанных hsp - генов тем, что он имеет высокий исходный уровень конститутивной экспрессии и немного более высокую экспрессию при тепловой индукции.
Этот промотор был назван "промотором hsp 80", так как он является конструктивным промотором с совпадающим элементом теплового шока и берется из гена, который обладает некоторой гомолочностью относительно семейства hsp 80 генов из других видов. Промотор hsp 80 управляет продуцированием протеинов теплового шока с высоким исходным уровнем при нормальных температурах (20 - 25oC) и демонстрирует несколько более высокую экспрессию при тепловой стрессе (35 - 40oC).
Описание чертежей
На фиг. 1 представлены плазмиды PZ 0217 и конструкция плазмид pZ 0601 и pZ 0601BS.
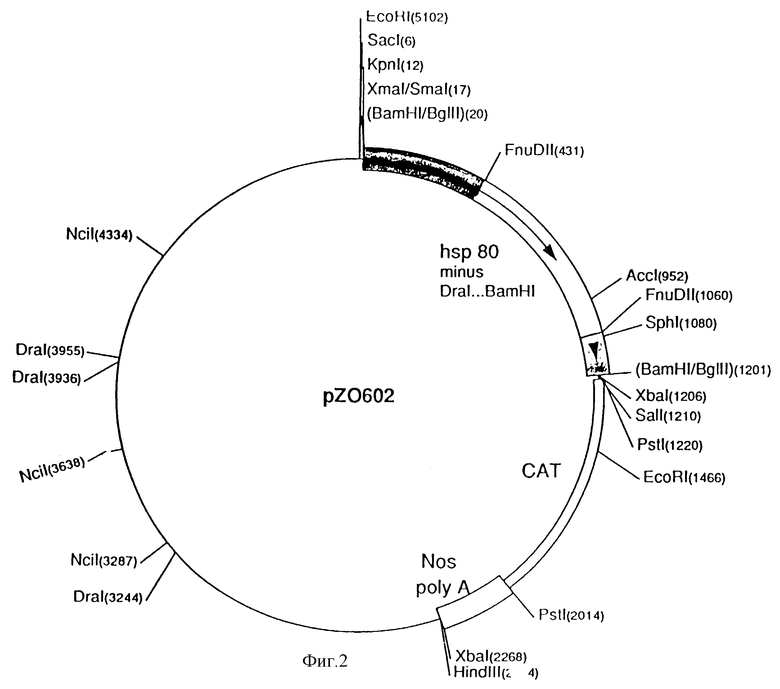
На фиг. 2 представлена плазмида pZ 0602.
Последовательность полного промотора приведена в таблице 1 (SEQ. ID. N 1). Полная нативная последовательность содержит 1568 пар оснований. Нуклеотиды занумерованы в отрицательном порядке от сайта начала трансляции (ATG) для нативного протеина теплового шока. Следующие участки были идентифицированы в этой последовательности:
A) Блок "ТАТА" является последовательностью из восьми пар оснований "ТАТАТАТА", расположенной от -97 до -90, включительно.
B) Имеется последовательность лидера и РНК в 61 п.о., которая расположена от -61 до -1.
C) Выступающий сайт идентифицировали в позиции - 61.
D) Имеется область от -604 до 488, которая, как оказалось, содержит расположенную в направлении вверх активирующую последовательность для этого промотора. Эту область обозначают через UASI. Было замечено, что эта область наделяет конститутивной активностью при осуществлении переходного анализа и ранее ее именовали "конститутивной зоной".
E) Имеется область о -1000 до -604, которая, как оказалось, содержит дополнительную, расположенную по направлению вверх, активирующую последовательность. Эту область обозначают UAS2.
F) Имеется область от -488 до -120, которая, оказывается, содержит дополнительную, расположенную по направлению вверх, активирующую последовательность. Эту область обозначали через UAS3.
G) Имеется два направленных повтора от -779 до -741 и от -740 до -702. Кроме того, последовательность, содержащая часть направленного повтора, простирается от -701 до -677.
H) Один совпадающий элемент теплового шока расположен в области от -131 до -120, а другой такой элемент расположен в области от -244 до -237.
Таким образом, один аспект настоящего изобретения состоит в том, чтобы предложить конструкцию ДНК, содержащую промотор hsp 80 Brassica, оперативно соединенный с гетерологическим геном. Так как представляется, что небольшие изменения можно осуществить в этой ДНК-последовательности без существенного нарушения активности промотора, настоящее изобретение также включает ДНК-последовательности, которые "функционально эквиваленты" промотору hsp 80 Brassica, и конструкции, содержащие такие ДНК-последовательности, оперативно связанные с гетерологическим геном.
На протяжении патентного описания и формулы изобретения используются следующие определения:
"Функциональный эквивалент" относится к любой ДНК-последовательности, которая является комплементарной к ДНК-последовательности, которая при строгих условиях гибридизации будет гибридизироваться с упомянутой последовательностью, и обладает промоторной активностью, аналогичной промотору hsp 80 Brassica.
"Строгими условиями гибридизации" являются условия, при которых гибридизация осуществляется при температуре 60oC в 2,5X соляном цитратном буфере (SSC-буфере) с последующей единственной промывкой при 37oC при сниженной концентрации буфера, которая не может нарушить гибридизацию, которая имеет место.
"Гетерологический ген" является ДНК-последовательностью, которая кодирует любой пептид или протеин, отличный от протеина hsp 80 Brassica.
"Промотором с делецией" называется любой промотор hsp 80 Brissica, который содержит "пробелы", но при этом сохраняет активность.
"Функциональным эквивалентом промотора с делецией" называется промотор с делецией, который имеет другие делеции, но все-таки сохраняет по существу эквивалентную активность при сравнении с промотором с делециями.
"Регулируемым промотором" является любой промотор, активность которого нарушается цис- или транс-действующим фактором (факторами).
"Конструктивным промотором" является любой промотор, который является активным в большинстве тканей или органов в течение большей части времени.
Промотор hsp 80 (или его функциональный эквивалент) можно использовать для осуществления конструктивной экспрессии любого целевого гетерологического гена. Примеры соответствующих гетерологических генов включают (но ими не исчерпывается полный список): инсектицидальные токсины (такие, как токсины из Bacillus thuringiensis), гены стойкости к гербицидам, антимикробные гены, антигрибковые гены, антивирусные гены и антипитающие гены.
В предпочтительном варианте конструкцию гетерологического гена hsp 80 вставляют в вектор и этот вектор используют для трансформации эукариотного хозяина. Эукариотным хозяином в предпочтительном варианте является растительная клетка или растительный протопласт. Предпочтительные векторы, разумеется, могут варьироваться в зависимости от выбранного хозяина. Для двудольных растений вектор может быть введен в протопласт при помощи электропорации или этот вектор может быть производным Ti-плазмиды Agrobacterium fumifaciens (A. t. ), который инфицирует клетку или протопласт, и может быть использован в трансформации с участием A.t., включающей так называемые бинарные приемы. (См., например, Gasser, C. и др., 1989, Science т. 244, стр. 1293-1299). Однопольные в предпочтительном варианте трансформирует с использованием так называемой "баллистической" техники (Gasser и др., см. выше) или они могут быть трансформированы с использованием протопластов.
В том и другом случае соответствующие векторы и протоколы трансформации хорошо известны в этой области техники. Трансформированные клетки или протопласты выращивают в соответствующей культурной среде и трансформированное растение регенерируется. Трансформированное растение экспрессирует гетерологический ген конструктивно.
Кроме того, неожиданным образом было установлено, что в этом промоторе могут быть сделаны различные делеции и полученный в результате промотор с делециями, как было установлено, обладает либо a) увеличенной активностью; либо b/ по существу той же активностью, что и нативный промотор; либо c/ замедленной активностью.
Таким образом, в соответствии с еще одним аспектом настоящего изобретения предлагается ДНК-последовательность, содержащая промотор с делециями промотора hsp 80 Brassica. Еще одним аспектом настоящего изобретения является ДНК-конструкция, содержащая промотор с делециями, оперативно связанный с гетерологическим геном. Кроме того, в этот аспект изобретения включаются функциональные эквиваленты промоторов с делециями и конструкции, содержащие функционально эквивалентный промотор с делециями и гетерологический ген.
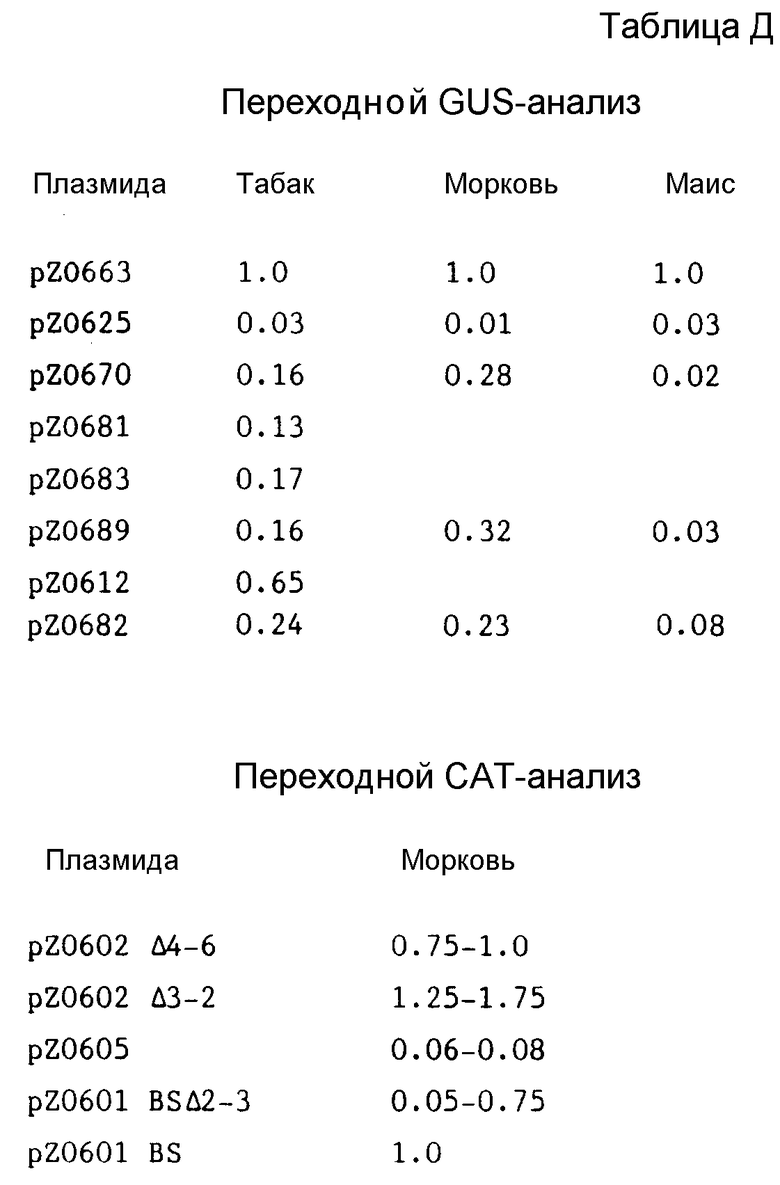
Различные делеции и гибридные промоторы получают, как это подробно описано в примерах. Первая совокупность промоторов с делециями образует серию 601BS. Эти промоторы отличаются тем, что содержат делецию, которая включает пары оснований от -118 до -246. Было установлено, что один промотор из этой серии 601BS Δ 2-3, который содержит пробел от -493 до -118, сохраняет приблизительно 50-75% активности по сравнению с интактным промотором.
Вторая совокупность промоторов с делециями образует серию 602. Все эти промоторы обладают пробелом от по крайней мере -488 до -134 и могут иметь пробел на 5'-конце различной длины, как это кратко описано в примере 3. Удивительным образом некоторые делеции увеличивают активность.
Промотор с делециями 603 имеет пробел, простирающийся от -1125 до -134. Этот промотор сохраняет лишь примерно 10% активности интактного промотора. Промотор с делециями 604 содержит пробелы от -1568 до -1125 и от -496 до -134, сохраняя приблизительно 25% активности. Промотор 605 имеет пробелы из всех пар оснований, расположенных в направлении вверх от -488, а соответствующее снижение активности составляет только примерно 6-8%.
Одна особенно важная область для активности лежит между -134 и -120. Промотор с делециями 601BS /BSph/ имеет удаленной только эту небольшую область, но его активность падает только до примерно 50-75% интактного промотора. Предпочтительные промоторы в соответствии с настоящим изобретением, таким образом, содержат по крайней мере эту короткую последовательность.
Как было упомянуто выше, область в 116 по, простирающаяся от -604 до -488 (UAS 1), или ее часть оказывается отвечает за сообщение конститутивной активности в область тканей. UAS 2 и UAS 3, как описано выше, оказывается сообщают активность другим тканям. Таким образом, в соответствии с еще одним аспектом настоящего изобретения конститутивная активность сообщается, вообще говоря, неконститутиному промотору (такому, как промотор, который в общем случае является индуцируемым или, в противном случае, регулируемым) при помощи оперативного соединения с индуцируемым или регулируемым промотором или вставки внутрь него одной или нескольких, расположенных в направлении вверх, активирующих областей, ибо только они обеспечивают активность в некоторых - большинстве органов/тканей и соединенные вместе дают так называемую конститутивную активность. Настоящее изобретение включает также ДНК-конструкции, содержащие такие промоторы с сообщенной конститутивной активностью, оперативно соединенные со структурным геном, способы, например, трансформации растительных клеток и протопластов, используя эти конструкции, и растительные клетки и протопласты, трансформированные этой конструкцией.
Должно быть ясно, что возможно, что области, меньшие, чем UAS 1, 2 и 3 области, будут достаточны для того, чтобы сообщить конститутивную активность. Это можно испытать, делая делеции в этих областях, используя приемы, хорошо известные в этой области техники. Промоторы с делециями в UAS-областях могут быть затем подвергнуты анализу на сохранение конститутивной активности. Такие анализы также доступны любому специалисту в этой области техники.
Области UAS 1, 2 и 3 отдельно или вместе можно также использовать для того, чтобы восстановить активность промотора, который был дезактивирован при помощи удалений и/или мутации. Это также составляет еще один аспект настоящего изобретения. Один пример такого применения может доставить промотор CAMY 35S, который был подвергнут делеции так, что после этого уже не функционировал. Вставка элементов UAS 1, 2 и 3 в комбинации или отдельно будет восстанавливать активность промотора в некоторых или во всех тканях.
Настоящее изобретение далее иллюстрируется при помощи примеров, которые не следует рассматривать в качестве его ограничения.
Пример 1. Выделение промотора hsp 80
Геномную библиотеку Brassica oleracea (cv. "Delira") строили в ламбда фаге Чарон 35 (Charon 35) и клетках K802, используя приемы, по существу описанные в книге: Maniatis и др., 1982 Molecular Clonuig, изд. Cold Spring Harbor Laboratory, стр. 282-283, которая здесь включена в качестве ссылки. Эту библиотеку просеивали с PvuI-StuI-фрагментом из гена hsp 83 Drosophila (Hacketl, R. W., и др., 1983, Nucl. Acids Res., т. 11/20/, стр. 7011-7030). Извлекали двадцать рекомбинантов с очевидной гомологичностью относительно гена Drosophila и осуществляли анализ на блотирование Саутерна, используя фрагмент гена hsp 83 Drosophila в качестве зонда. Фрагмент HindIII в 5,8 ко выбирали для субклонирования в вектор pUC9 и именовали как pZ217. Эта плазмида представлена на фиг. 1.
Далее, ген Хлорамфеникол ацетил трансферазы (CAT) (фирма Pharmacia) вставляется в сайт PstI известного вектора pUC19. Затем терминатор NOS [Bevan, M. и др., 1983. "Structura and Transcription of the Nopaline Synthase Gene Region of T-DNA", Nucl. Acids Res., т. 11(2)] вставляли в сайт PstI-HindIII. Полученную в результате плазмиду обозначали pZ030. BglII-фрагмент из pZ0217 отделяли и субклонировали в сайт BamHI из pZ030. Это дает конструкцию промотор-CAT ген-NOS терминатор, которая переносится в виде EcoRI-HindIII-фрагмента в производимый промышленностью вектор pTZ18R (фирма Pharmacia), образуя pZ0601, который представлен на фиг. 1.
Пример 2. Конструкции плазмид
In vitro мутагенез промотора hsp 80 в pZ0601 осуществляли с использованием системы in vitro мутагенеза, направленного на олигонуклеотиды, производимой фирмой Amersham. Два олигонуклеотида синтезировали в соответствии с инструкциями фирмы-производителя, чтобы образовать два единственных сайта рестрикции внутри промотора в направлении вверх от блока TATA, BamHI в -134 и SphI в -120. (Все позиции нуклеотидов даны относительно сайта начала трансляции). Эту новую плазмиду обозначали через pZ0601BS, и она также представлена на фиг. 1.
Плазмида pZ0602 и pZ0603
Стадия A) pZ0601BS переваривали при помощи BamHI, а концы заполняли, используя фрагмент Кленоу ДНК-полимеразы. Затем ДНК переваривали KpnI и полученный в результате фрагмент без промотора отделяли на агарозовом геле с низкой температурой точки плавления.
Стадия B) pZ0601BS переваривали при помощи BamHI и KpnI и hsp80-промотор отделяли на агарозовом геле с низкой температурой точки плавления. Этот фрагмент очищали, используя ELUTIP (фирма Schleicher and Schnell) и переваривали либо при помощи DraI, либо при помощи FnuDII.
Стадия C) DraI-фрагмент от -1568 до -488 со стадии B) подвергали лигации во фрагмент без промотора со стадии A. Полученную в результате плазмиду обозначали через pZ0602, и она представлена на фиг. 2.
Стадия D) FnuDII-фрагмент от -1568 до -1125 подвергали лигации во фрагмент без промотора со стадии A. Полученную в результате плазмиду обозначали через pZ0603.
Плазмида pZ0604
Стадия A) pZ0601BS переваривали при помощи BamHI и SmaI, а сайт BamHI заполняли ДНК-полимеразой T4 и деоксинуклеазой. Полученный в результате фрагмент без промотора отделяли на агарозовом геле с низкой температурой точки плавления.
Стадия B) pZ0601BS переваривали при помощи FnuDII. FnuDII-фрагмент от -1125 до -496 подвергали лигации во фрагмент без промотора со стадии A, чтобы получить pZ0604.
Плазмида pZ0605
Стадия A) pZ0601BS переваривали фрагментами BamHI и SmaI, а полученный в результате фрагмент без промотора отделяли на агарозовом геле с низкой температурой точки плавления.
Стадия B) pZ0601BS переваривали DraI. DraI-фрагмент от -488 до -134 подвергали лигации во фрагмент без промотора со стадии A, чтобы получить pZ0605.
Мутанты делеций
Строили из плазмиды pZ0602 серии 5'-делеций в промоторе hsp 80. pZ0602 переваривали ферментами SacI и SmaI, чтобы образовать субстрат для переваривания Экзонуклеазой III (Стратаген). После обработки Экзонуклеазой III в течение различных промежутков времени, полученные в результате ДНК превращали в ДНК с липкими концами, используя Нуклеазу Фасоли Золотистой /фирма Boehringer Mannheim/. ДНК отделяли, используя электрофорез на агаровом геле с низкой температурой точки плавления. Яркие полосы вырезали, разбавляли и подвергали лигации. После трансформации мутанты делеции выбирали и исследовали последовательность до точки соединения.
Строили из pZ0601В серию 3'-делеций в промоторе hsp 80 при помощи переваривания с использованием BamHI и SphI, а затем следовали той же процедуре, что была описана при создании 5'-делеций.
Пример 3 - Биоанализы
Поддержание линии клеток моркови
Культуру суспензии дикой моркови Редвуд Сити (RCWC) (полученную из Стэнфордского универститета) поддерживали в следующей среде суспензии моркови: 1 x MS соли, 1 мг/л никотиновой кислоты, 1 мг/л пиридоксина HCl, 1 мг/л тиамина, 100 мг/л инозита, 0,1 мг/л 2,4 - D, 30 г/л сахарозы и финальное значение pH 5,8 обеспечивали при помощи KOH. Культуру поддерживали при помощи разбавления в свежей среде каждые 7 дней.
Образование протопластов
Культуру суспензии RCWC разбавляли в отношении 1 : 10 за четыре дня до использования. 50 мл культуры (приблизительно 5 мл объема упакованных клеток) подвергали центрифугированию в течение 10 минут со скоростью 500 g. Затем клетки снова суспендировали в 50 мл следующего фильтрованного Раствора Фермента Моркови: 10 г/л Целлюлизина (Фирма Calbiochem), 5 г/л Розима (фирма Genecor), 0,4 М маннита, 50 мМ CaCl2, 10 мМ NaOAc, pH 5,8. Клетки спокойно перемешивали в течение двух часов перед перевариванием. Протопласты дважды промывали и снова суспендировали в Культурную Среду Моркови (CCM), которая представляет собой Среду Суспензии Моркови, описанную выше, с добавлением 0,4 М маннита. Протопласты подсчитывали на гемоцитометре при разбавлении 1 : 10, чтобы определить концентрацию.
Электропорация
Во всех электрофорезах использовали Прогенитор II типа PG 200 (фирма Hoefer) с круговым электродом. Образцы подвергали электропорации в стерильных микротитрованных чашках с 24 углублениями при разности потенциалов 200 Вольт в течение 100 мс.
Каждую плазмиду, содержащую CAT (фирма Pharmacia) - конструкцию, испытывали в 3 или 4 копиях в нескольких экспериментах. В каждом эксперименте в качестве контрольной использовали от 30 до 50 мкг плазмиды pZ060IBS. Все другие плазмиды испытывали с использованием эквивалентного молярного количества ДНК относительно pZ060IBS (см. табл. А).
Порции в приблизительно 106 протопластов помещали в 1,5 мл пробирки, центрифугировали в течение двух минут со скоростью 500 g, и большую часть верхнего слоя удаляли. В 75 мкл 2M KCl добавляли каждый ДНК. Объем доводили до 1 мл добавлением ССМ (pH обеспечивали на уровне 8,0) и эту смесь подвергали электропорации и сразу разбавляли 5 мл ССМ при pH 5,8 в чашке Петри. Разбавленные образцы хранили в темном шкафу в течение 1 - 2 дней перед отбором проб для CAT-анализа.
Пример 4. Конструктивная экспрессия в полном растении
Табак трансформировали с использованием следующего протокола.
Растительная ткань
Экспланты листьев табака получали из выращенных в стерильных условиях растений табака. Стерильные растения табака вегетативно размножали через интервалы времени и в приблизительно 1 месяц при помощи удаления верхних узлов существующего растения и рекультивирования их в пористые сосуды, содержащие 75 мл отвержденной агаром, не содержащей гормонов, среды (0/0 среды), содержащей соли Мурашиге-Скуга, 1 мл/л 100 мг/л мио-инозита, 5 мл/л витаминной смеси (которая обеспечивает 0,5 мг/л пиридоксина HCl, 0,5 мг/л никотиновой кислоты и 1,0 мг/л тиамина) и 3% сахарозы. Растения табака поддерживали при постоянном освещении средней интенсивности. Сосуды плотно закрывали зеленой бумажной тесьмой или их можно оставить незакрытыми, чтобы обеспечить газовый обмен.
Агробактериальный вектор
Систему бинарного вектора использовали для того, чтобы трансформировать Agrobacterium fumifaciens, а затем использовали для того, чтобы трансформировать клетки табака.
Общие процедуры
Векторы Agrobacterium хранили при температуре -70oC. За приблизительно 18 часов по трансформации 25 мл стерилизованной жидкой среды LB3 (см. ниже) содержащей соответствующие антибиотики, прививали 100 - 500 микролитрами культуры Agrobacterium. Культуру выращивали на вибраторе при скорости 250 об/мин при 28oC в течение ночи.
Среда LB3 (для 1 литра)
Триптон - 10 г
Экстракт дрожжей - 5 г
NaCl - 4 г
KCl - 1 г
MgSO4 • 7H2O - 3 г
Добавляли воду до 1 литра и распределяли порциями по 25 мл на колбу.
Антибиотиками являются 25 мкг/мл стрептомицина и 50 млг/мл канамицина. Трансформацию в общем случае осуществляли, когда клетки находились в своей log-фазе; О. П. в диапазоне 600 нм в предпочтительном варианте содержится в области 0,50 - 1,0. Культура должна быть разбавлена в концентрации 108 клеток/мл в 50 мл жидкой, не содержащей гармонов 0/0 среды.
Экспланты табачных листьев погружали на 3 минуты в культуру с разбавлением 108 клеток/мл трансформированных Agrobacterium, полученную выше. Экспланты удаляли и сушили блоттированием на стерильной фильтровальной бумаге Ватман. Затем их помещали на регенерационную среду без антибиотиков на два дня. На 3 день обработанные экспланты переносили на регенерационную среду, содержащую соответствующие антибиотики. Экспланты оставались на этих пластинах в течение 3 - 4 недель или до тех пор, пока соответствующим образом трансформированные корни не становились достаточно большими, чтобы их можно было перевести в среду для развития корня. Средой для ускорения является полупрочная 0/0 среда, содержащая 250 мг/л карбенициллина и либо 100 мг/л канамицина, либо 50 мг/л гигромицина, в зависимости от конструкции. Спустя приблизительно две недели веточки укоренялись и начинали зеленеть. Затем их переносили в банки и каждому растению давали собственный номер. Далее, осуществляли анализы на различных тканях, чтобы подтвердить конститутивную активность промотора.
Векторы трансформации
pZ 0639
Плазмиду строили по аналогии с pZ 0601 за тем исключением, что вместо содержащейся кассеты hsp 80-CAT-ген-NOS терминатор, она содержит кассету hsp-80-GUS-NOS терминатор. Эту плазмиду обозначали через pZ 06012.
Плазмиду pBin 19 (фирма Clonefech) расщепляли ферментами EcoRI и Hind III. Плазмиду pZ 0612 расщепляли ферментами ScaI, EcoRI и Hind III. Оба продукта переваривания разделяли на агарозе с низкой температурой точки плавления. Фрагмент вектора pBin 19 в 12 ко и фрагмент hsp 80-NOS в приблизительно 3,6 ко выделяли и, далее, подвергали вместе лигации. Полученную в результате плазмиду, обозначенную через pZ0639, можно идентифицировать при помощи переваривания ферментом Pst I.
pZ 0640
Следуя процедуре, описанной выше, строили pZ 0640. Он отличается от pZ 0639 тем, что содержит усеченный hsp 80-фрагмент/0,624 ко против полного промотора в 1,56 ко).
pZ 0641
Следуя процедуре, описанной выше, строили pZ 0641. pZ 0641 содержит 0,252 ко hsp 80-промотора, имеющего полную длину 1,56 ко).
pZ 0642
Следуя процедуре, описанной выше, строили pZ 0642. Вместо hsp 80 или производного промотора, pZ 0642 содержит фрагмент CaMY 35S промотор-GUS-NOS.
Пример 5. Активность промотора
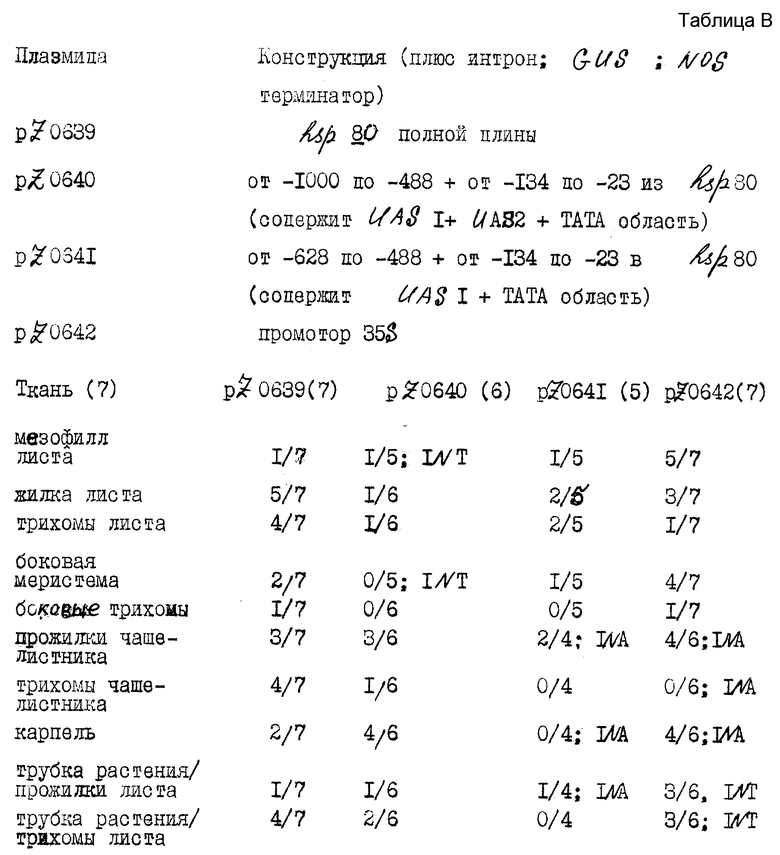
Растения, полученные из процедуры в примере 4, испытывали на экспрессию гетерологического гена GUS. Брали образцы ткани и анализировали на присутствие GUS. GUS-активность обнаруживали во всех образцах ткани. Контрольные ткани (полученные из нетрансформированных растений, которые подвергали тем же процедурам культивирования и регенерации) также анализировали на GUS-активность; таковой не обнаруживали. Число в круглых скобах обозначает общее число растений, из которых брали образцы. Результаты приведены в таблю В, в виде числа положительных образцов (наблюдается голубое окрашивание). "NT" обозначает "не испытывали"; "NA" обозначает "не доступный"; "M" означает мутант.
Эти данные показывают, что промотор hsp 80 является активным до некоторой степени во всех испытываемых тканях, хотя интенсивность окрашивания в большинстве случаев была меньшей, чем для промотора 35S. В общем случае, мезофилл листа не окрашивается до вакуумной инфильтрации, но затем большое число клеток окрашивалось, хотя их внешний вид напоминает скорее внешний вид трихом, чем стандартных мезофилловых клеток.
Пример 6. Гибридные промоторы
Различные области, расположенные в направлении вверх от промотора hsp 80, подвергали лигации с неактивным гетерологическим минимальным промотором, чтобы определить, придает ли эта область активность. Фрагменты, указанные ниже, клонировали в направлении вверх от усеченного промотора CaМУ 35S, расположенного от позиции -46 относительно сайта начала транскрипции СаМУ до позиции +131 (блок TATAAA занимает позиции от -31 до -25); эту область в дальнейшем именуют как "-46 35S -промотор". [NB. нумерация, используемая здесь для идентификации усеченного CaMY35S, дана относительно самого CaMY35S, и она не совпадает с нумерацией, используемой в дальнейшем для hsp 80 и его фрагментов] . Плазмида pZ 0625 содержит промотор -46 35 S, итрон 6 из гена ADHIS маиса, область, кодирующую β- глюкуронидазу (GUS) и терминатор NOS в pT7T3 18U (предоставленной фирмой Pharmacia. Следующие фрагменты hsp 80 Brassica подвергали лигации в направлении вверх от -46 35S фрагмент промотора (см. табл. С).
Гибридные промоторы содержащиеся в плазмидах (см. табл. С), именуются как гибридные промоторы 670, 681, 682, 683 и 689, соответственно.
Кроме того, подвергали испытаниям также pZ0612 (которая совпадает с pZ 0601 за тем исключением, что CAT-ген заменяли на GUS-ген) и содержит hsp 80-промотор полной длины, контролирующий GUS, с терминатором NOS (но не итрон) в p218R.
Протопласты получали из моркови, как это было описано ранее, Черного Мексиканского Сладкого (BMS) маиса и табака. Протопласты подвергали электропорации с целевой плазмидой, извлекаемой за день до этого, а затем экстрагировали, а экстракты анализировали на GUS-активность. GUS-активность измеряли спектрофотометрическим способом. Результаты приведены в виде средних значений, нормализованных для конструкции полной длины промотора 35S, pZ0663 (35S от -366 до +131, интрон 6, GUS, терминатор NOS). Промотор 35S полной длины описан у Frank и др., Cell, т. 21, стр. 285-294 (1980).
Как можно видеть из табл. D, все области, расположенные по направлению вверх, дают по существу одинаковую активность после слияния с 35S -46-промотором и при этом, оказывается, дают слабый дополнительный эффект.
Перечень последовательности
(1) Общая информация:
(i) Заявитель: фирма Sandoz Limited
(ii) Заголовок изобретения: Новый растительный промотор
(iii) Номер последовательностей: 1
(2) Информация для SEQID N 1:
(i) Характеристики последовательности:
(A) Длина: 2042 пар оснований
(B) Тип: нуклеиновая кислота
(C) Тип нити: двойная
(D) Топология: линейная
(ii) Тип молекулы: ДНК (геномная)
(iii) Гипотетическая: нет
(iv) Противоположное направление: отсутствует.






Изобретение предназначено для использования в генетической инженерии растений. Конститутивный промотор гена теплового шока hsp 80 Brassica контролирует экспрессию целевого гетерологичного гена с высоким исходным уровнем при нормальных температурах (20-25°С) и обеспечивает более высокую экспрессию при тепловом стрессе (35-40°). Мутанты промотора с делециями сохраняют активность и/или обладают более высокой активностью. 2 ил., 5 табл.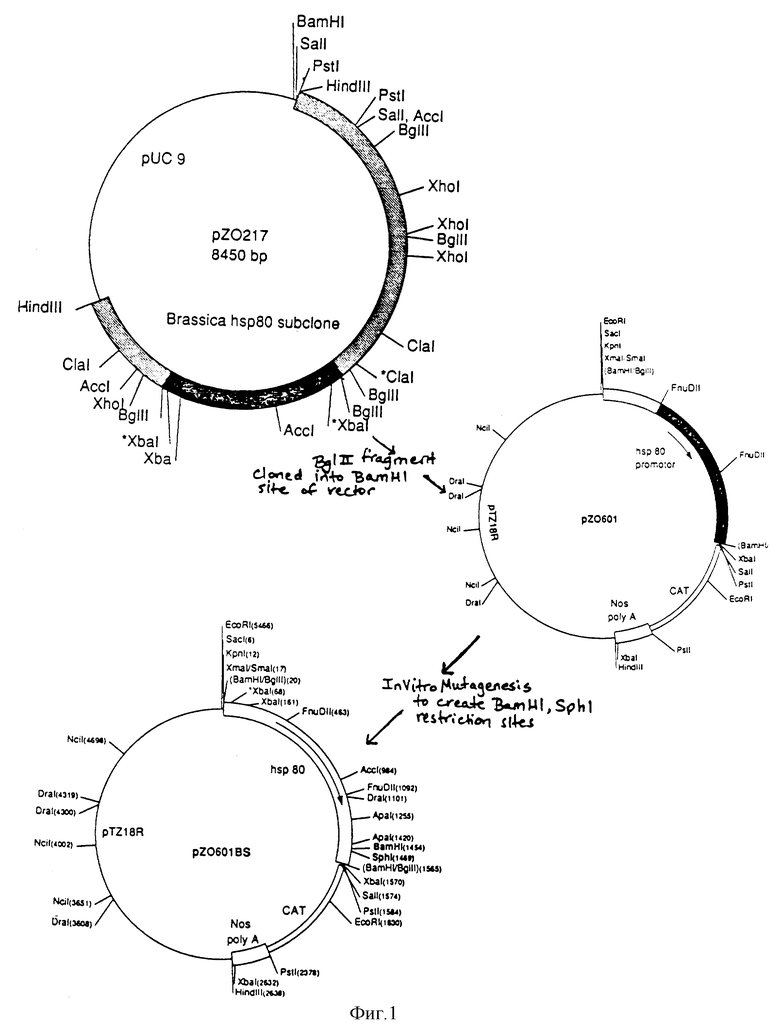
ДНК-последовательность для экспрессии генов в тканях и органах, включающая промотор цветной капусты, названный hsp 80 Brassica, кодируемый последовательностью нуклеотидов 472 - 2039 SeqID N 1, приведенной в описании, или последовательностью, у которой делетированы нуклеотиды 472 - 1040 и 1552 - 1906, или последовательностью, у которой делетированы нуклеотиды 493 - 1412 и 1552 - 1906, операбельно связанный с гетерологичным геном.
| Торфодобывающая машина с вращающимся измельчающим орудием | 1922 |
|
SU87A1 |
| СПОСОБ ПОЛУЧЕНИЯ АНГИДРИДА 1,4-ФЕНИЛЕН-БИС- ДИТИОФОСФОНОВОЙ КИСЛОТЫ | 0 |
|
SU248677A1 |
| Транскрипция и Трансляция | |||
| Пер | |||
| с англ | |||
| /Под ред | |||
| Хеймса Б | |||
| - М.: Мир, 1987, с | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |
