Область изобретения и краткое описание известного уровня техники
Данное изобретение относится к процессу биосинтеза для получения и выделения 7-аминодезацетоксицефалоспорановой кислоты (7-АДЦК).
Беталактамные антибиотики образуют наиболее важную группу антибиотических соединений с длительной историей клинического применения. В этой группе выделяются пеницилины и цефалоспорины. Эти вещества природно продуцируются нитчатыми грибами Penicillium chrysogenum и Acreonium chrysogenum, соответственно.
В результате применения методов улучшения классического штамма уровни продукции антибиотиков Penicillinum chrysogenum и Acreonium chrysogenum заметно увеличены за последние десятилетия. С увеличением знаний о метаболических путях биосинтеза, приводящих к продукции пенициллинов и цефалоспиринов и приходом генной инженерии стали доступны новые средства повышения продуктивности штаммов и получения производных соединений in vivo.
Большая часть ферментов, участвующих в биосинтезе беталактамов, уже идентифицирована, и клонированы соответствующие им гены, что можно найти у Ingolia and Queener, Med. Res. Rev., 9 (1989), 245 - 264 (путь биосинтеза и ферменты), и Aharonowitz, Cohen, and Martin, Ann. Rev. Microbiol., 46 (1992), 461 - 495 (клонирование генов).
Первыми двумя этапами в биосинтезе пенициллина у P.chrysogenum являются конденсация трех аминокислот L-5-амино-5-карбоксипентановой кислоты (L-α-аминоадипиновой кислоты) (A), L-цистеина (C) и L-валина (V) в трипептид LLD-ACV, с последующей циклизацией этого трипептида с образованием изопенициллина N. Это соединение содержит типичную беталактамную структуру.
Третий этап включает замену гидрофильной боковой цепи из 1-5-амино-5-карбоксипентановой кислоты на гидрофобную боковую цепь путем воздействия фермента ацетилтрансферазы (AT). При промышленном способе производства пенициллина G боковой цепью выбора является фенилуксусная кислота (ФУ). В ЕР-А-0532341 раскрыто применение в качестве сырья для производства адипата (5-карбоксипентаноата). Включение этого субстрата приводит к образованию пенициллинового производного с 5-карбоксипентаноиловой боковой цепью, а именно 5-карбоксипентаноил-6-аминопенициллановой кислоты. Это включение происходит благодаря тому факту, что ацетилтрансфераза обладает доказанной широкой субстратной специфичностью (Behrens et al. , J. Biol. Chem. 175 (1948), 751 - 809; Cole, Process. Biochem. 1 (1966), 334 - 338; Ballio et al. . Nature 185 (1960), 97 - 99). Ферментативная реакция обмена, опосредуемая AT, происходит внутри клеточной органеллы, микротельца, как описано в ЕР-А-0448180.
Цефалоспорины значительно более дорогостоящи, чем пенициллины. Одна из причин этого состоит в том, что некоторые цефалоспорины (например, цефалексин) получают из пенициллина путем ряда химических превращений. Другой причиной является то, что до сих пор ферментации могут подвергаться только цефалоспорины с D-5-амино-5-карбоксипентаноильной боковой цепью цефалоспорин C, несомненно, наиболее важный исходный материал в этом отношении, очень хорошо растворим в воде при любом pH, что подразумевает длительные и дорогие процессы выделения с использованием громоздкой и дорогостоящей колоночной технологии. Цефалоспорин С, полученный этим путем необходимо превращать в терапевтически применяемые цефалоспорины с помощью ряда химических и ферментативных превращений.
Промежуточная 7-АДЦК в настоящее время производится с помощью химического получения производных пенициллина G. Необходимые этапы химического процесса получения 7-АДЦК включают увеличение 5-членной структуры пенициллинового кольца до 6-членной структуры цефалоспоринового кольца. Однако, фермент экспандаза из нитчатой бактерии Streptomyces clavuligerus может осуществлять подобные расширения кольца. При введении в P.chrysogenum он может превращать пенициллиновую кольцевую структуру в цефалоспориновую кольцевую структуру, как описано у Cantwell et al., Proс. R. Sос. Lond. в 248 (1992), 283 -289 и в EP-A-0532341 и ЕР-А-0540210. Фермент экспандаза хорошо охарактеризован (ЕР-А-0366354) как биохимически, так и функционально, так же как и соответствующий ему ген. Были описаны как физические карты гена cefE (ЕР-А-0233715), последовательность ДНК, так и исследования по трансформации P. chrysogenum с помощью cefE.
Еще одним источником подходящего фермента расширения кольца является нитчатая бактерия Nocardia lactamdurans (первоначально Streptomices lactamdurans). Описаны как биохимические свойства этого фермента, так и последовательность ДНК гена (Cortes, et al., J. Gen. Microbiol. 133 (1987), 3165 - 3174; and Coque et al., Mol. Gen. Genet. 236 (1993), 453 - 458, соответственно).
Более конкретно в ЕР-А-0532341 раскрывается использование in vivo фермента экспандазы в P.chrysogenum в сочетании с 5-карбоксипентаноиловой боковой цепью в качестве сырьевого источника, который используется как субстрат для ацетилтрансферазного фермента в P.chrysogenum. Это приводит к образованию 5-карбоксипентаноил-6-АПК, которая превращается с помощью фермента экспандазы, введенного в штамм P.chrysogenum, с получением 5-карбоксипентаноил-7-АДЦК. В конце предполагается удаление 5-карбоксипентаноиловой боковой цепи дающее 7-АДЦК в качестве конечного продукта. В патентной заявке ЕР-А-0540210 описывается сходный процесс получения 7-АДЦК, включающий дополнительные этапы превращения 3-метильной боковой цепи кольца АДЦК в 3-ацетоксиметильную боковую цепь АЦК. Однако в вышеуказанных патентных заявках не раскрывается эффективный и экономически эффективный процесс, потому что прежде всего не решена проблема своевременной экспрессии фермента экспандазы в клетке, сопутствующей экспрессии фермента ацетилтрансферазы.
В отличие от этого данное изобретение представляет эффективный способ продукции 7-АДЦК, при котором экспандаза и ацетилтрансфераза экспрессируются одновременно.
Кроме того, применение нового предшественника боковой цепи, а именно, 3,3'-тиодипропионовой кислоты, также раскрывается в этом изобретении. Этот предшественник очень эффективно включается P.chrysogenum в соответствующие пенициллины, кольцо которых расширяется с помощью последующего действия фермента экспандазы.
В дополнение к этому, до сих пор не был описан эффективный способ выделения производного 7-АДЦК из ферментационной среды перед его деацилированием. Данное изобретение представляет эффективную процедуру экстракции растворителем для выделения производного 7-АДЦК.
С помощью данного изобретения обеспечивается эффективный процесс получения 7-АДЦК, включающий этапы реакции, не раскрытые и не предложенные в прототипах до сих пор.
Также, путем применения данного изобретения и аналогично описанию, данному в ЕР-А-0540210, по эффективному завершенному процессу таким путем можно получать 7-АЦК.
Кратное описание рисунков
Рис. 1: Функциональная карта плазмиды pMcTNE
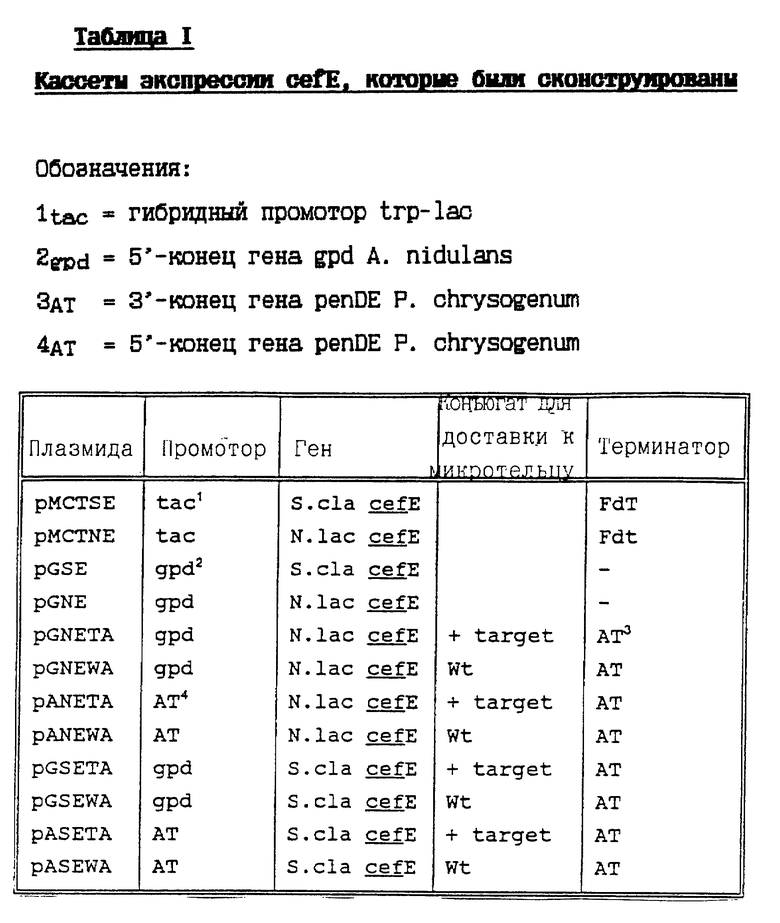
Рис. 2: Функциональная карта плазмиды pMcTSE
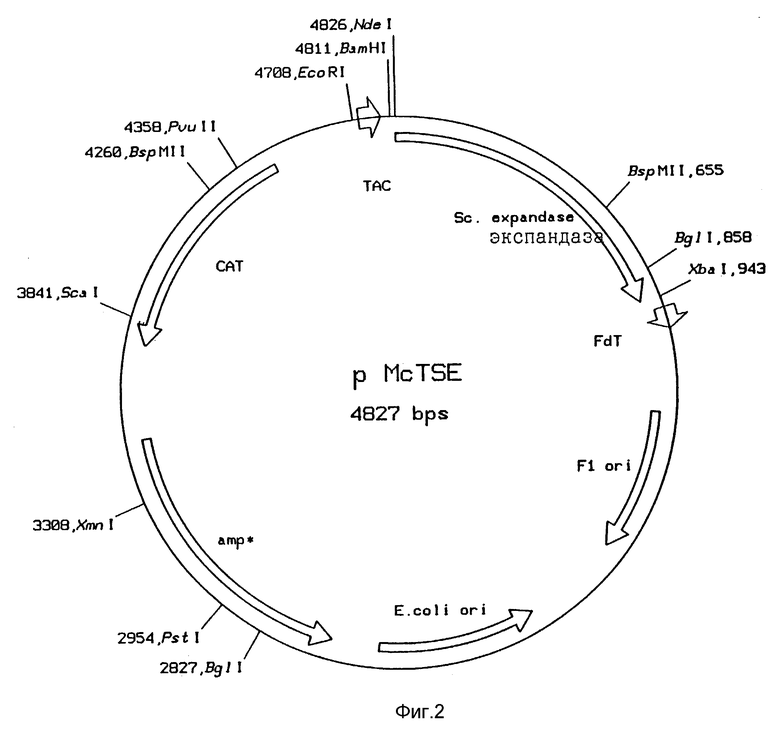
Рис. 3: Функциональная карта плазмиды pMcTNde
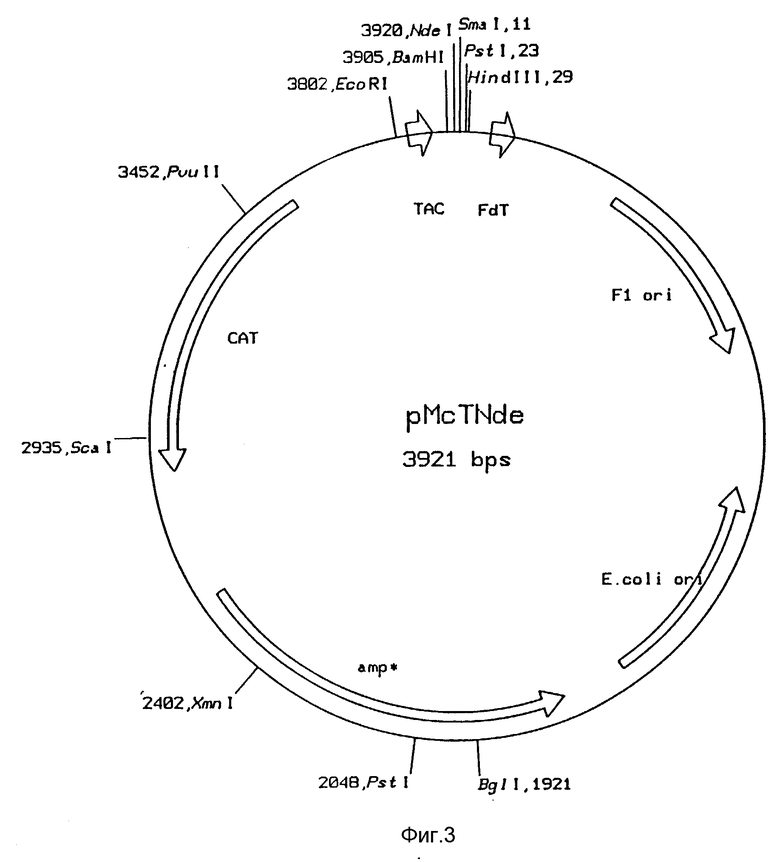
Рис. 4: Функциональная карта плазмиды pGNETA
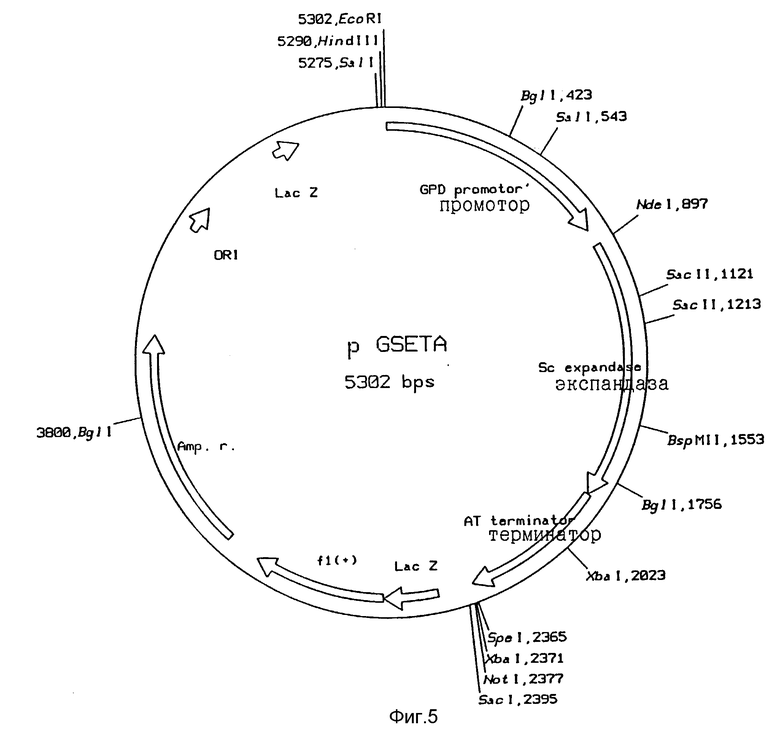
Рис. 5: Функциональная карта плазмиды pGNETA
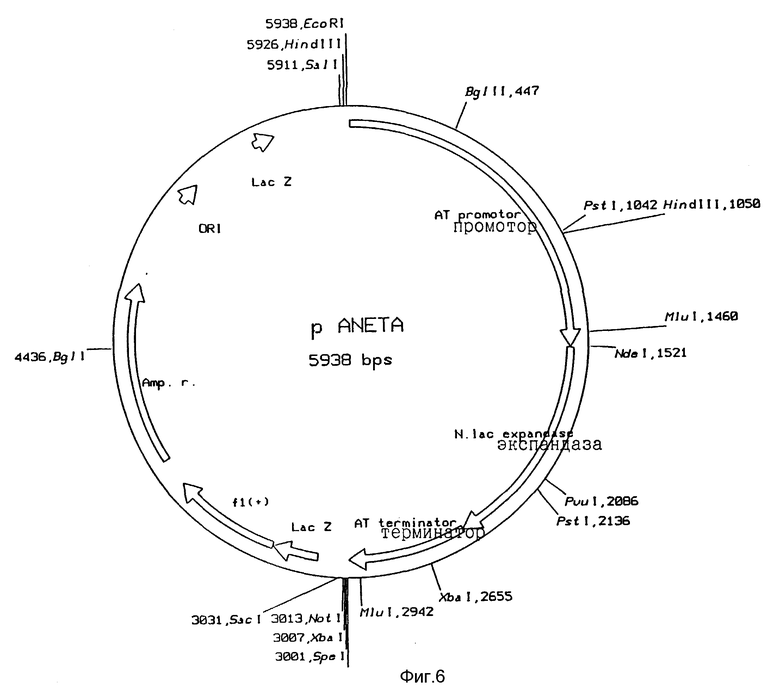
Рис. 6: Функциональная карта плазмиды pANETA
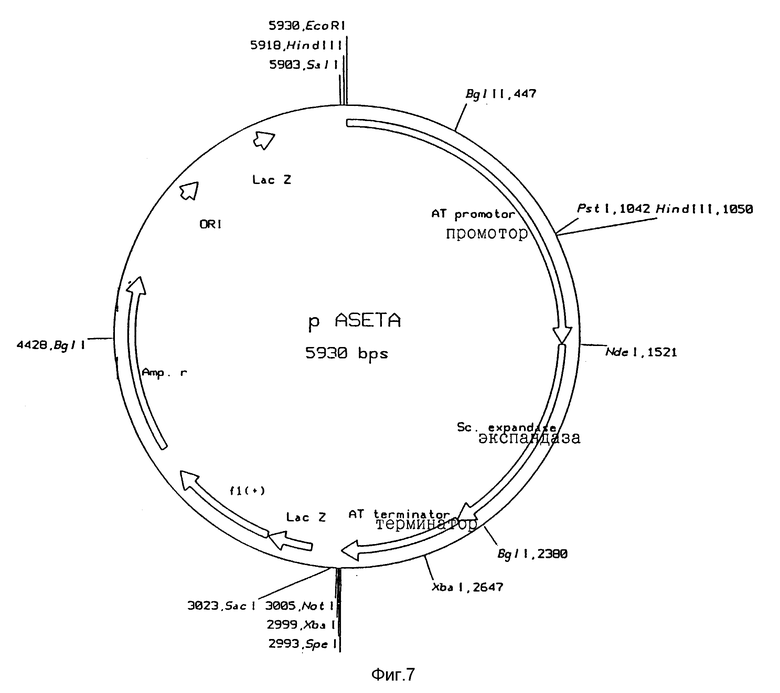
Рис. 7: Функциональная карта плазмиды pASETA
Рис. 8: Последовательность ДНК Nocardia lactamdurans cefE (Coque et al. выше) (нижние строки) в сравнении с последовательностью продукта 1 ПЦР (верхние строки).
Краткое описание перечня последовательностей
Последовательности ИД NN с 1 по 13: олигонуклеотиды, использованные в конструировании кассеты экспрессии P.chrysogenum для генов cefE Streptomyces clavuligerus и Nocardia lactamdurans.
Последовательность ИД N 14: последовательность ДНК cefE Nocardia lactamdurans (Coque et al., выше).
Краткое описание изобретения
Данное изобретение, таким образом представляет способ получения и выделения 7-аминодезацетоксицефалоспорановой кислоты (7-АДЦК) путем:
а) трансформации штамма Penicillium chrysogenum с помощью гена экспандазы под транскрипционной и трансляционной регуляцией сигналов экспрессии нитчатого гриба;
b) ферментирования указанного штамма в культурной среде и добавления к указанной культуральной среде 3,3'-тиодипропионовой кислоты или ее соли или эфира, подходящего для получения 3-(карбоксиэтилтио)пропионил-6-аминопенициллановой кислоты (3-(карбоксиэтилтио)пропионил-6-АПК), кольцо которых in situ расширяется с образованием 3-(карбоксиэтилтио)пропионил-7-АДЦК;
с) выделения 3-(карбоксиэтилтио)пропионил-7-АДЦК из ферментационного бульона;
d) дезацетилирования указанной 3-(карбоксиэтилтио)пропионил-7-АДЦК и
е) выделения кристаллической 7-АДЦК.
Предпочтительно, стадия (е) является стадией фильтрации.
Предпочтительно, экспрессия гена экспандазы находится под транскрипционной и трансляционной регуляцией соответствующих контролирующих элементов АТ-гена, что обеспечивает одновременную координацию экспрессии указанных генов.
Предпочтительно, 3-(карбокоиэтилтио)пропионил-7-АДЦК выделяют из жидкой ферментационной среды путем экстрагирования фильтрата среды органическим растворителем, несмешиваемым с водой, при pH ниже примерно 4,5 и обратным экстрагированием тех же самых веществ водой при pH от 4 до 10.
Кроме того, представлены вектор рекомбинантной ДНК, содержащей ДНК, кодирующую экспандазу, функционально связанную с транскрипционными и трансляционными контролирующими элементами АТ-гена P.chrysogenum или гена gpdA A. nidulans, и клетки-хозяева, трансформированные такой ДНК.
Детальное описание изобретения
Данное изобретение относится к использованию функциональных генных конструкций в P.chrysogenum для расширения in vivo структуры пенициллинового кольца в сочетании c использованием нового субстрата для ферментов биосинтеза для образования производного ключевого промежуточного соединения в биосинтезе цефалоспоринов, 7-аминодезацетоксицефалоспорановой кислоты или 7-АДЦК. Это производное имеет такое химическое строение, которое дает возможность эффективной экстракции растворителем, обеспечивая таким образом экономически привлекательный процесс выделения.
Трансформация P. chrysogenum может, в принципе, достигаться с помощью различных средств доставки ДНК, такими, как опосредуемое PEC-Ca поглощение протопластов, электропорация или методы обстрела частицами и селекцией трансформантов. Смотрите, например. Van den Hondel en Punt, Gene Transfer and Vector Development for Filamentous Fungi, in: Applied Molecular Genetics of Fungi (Peberly, Laten, Ogden, Bennet, eds.), Cambridge University Press (1991). Применение доминантных и недоминантных маркеров селекции было описано (Van den Hondel, выше). Были описаны маркеры селекции как гомологичного (происходящие из P.chrysogenum), так и гетерологичного (происходящие не из P.chrysogenum) происхождения.
Применение различных маркеров селекции трансформантов, гомологичных или гетерологичных, в присутствии или в отсутствии векторных последовательностей, физически связанных или нет с неселектируемой ДНК, при селекции трансформантов хорошо известно.
Предпочтительно, для селекции трансформантов P.chrysogenum используется гомологичный маркер селекции, чтобы ограничить количество гетерологичной ДНК, вводимой в P.chrysogenum. Наиболее предпочтительно использовать доминантный маркер селекции, который может селективно удаляться из трансформированного штамма, например, ген amdS A.nidulans или других нитчатых грибов (Европейская патентная заявка N 94201896.1). Эти предпочтительные характеристики маркера селекции трансформанта P.chrysogenum очень благоприятны для процесса и процедур регистрации продукта, так как в процессе не участвуют маркеры, связанные о антибиотикоусотойчивостью, или они не будут вводиться в окружающую среду.
При наиболее предпочтительном воплощении маркер селекции amdS, который может селективно удаляться из штамма, дает возможность повторных циклов трансформации с использованием той же самой доминантной селекции снова и снова. Это свойство освобождения от маркера селекции новых штаммов P. chrysogenum, экспрессирующих экспандазу, является определяющим для быстрой разработки высокопродуктивных штаммов по программе улучшения промышленных штаммов.
Способность к реакции расширения кольца, опосредуемая ферментом экспандазой, вводится и экспрессируется таким путем в P.chrysogenum, например, штамм Wisconsin 54-1255. Эта реакция расширения кольца происходит также и в их мутантах, дающих увеличенный выход беталактамов. Должно быть понятно, что в этом случае характеристики среды должны быть слегка адаптированы для получения эффективного роста.
Кроме того, ген cefE помещается под транскрипционный и трансляционный контроль соответствующих, контролирующих ген, элементов нитчатого гриба, предпочтительно происходящих из ацетилтрансферазного (AT) гена P.chrysogenum, что дает возможность их экспрессии в оптимальных временных рамках, синхронизированных с действием самого фермента ацетилтрансферазы. Эти меры являются решающими для эффективности реакции расширения кольца в пенициллиновой молекуле.
В дополнение к синхронизированной экспрессии генов, кодирующих экспандазу и трансферазу, могло быть благоприятным для разработки экономичного процесса производства совместное расположение части экспандазных ферментов с ацетилтрансферазой в микротельцах (внутриклеточная локализация ацетилтрансферазы). При этих предпочтительных осуществлениях будет очень сильно снижаться количество пенициллиновых побочных продуктов, которые не допускаются в конечном продукте 7-АДЦК регистрационными органами.
Резюмируя, в данном изобретении раскрывается, как действие фермента экспандазы, введенного в P.chrysogenum, может быть нацелено на расширение пенициллинового кольца на основе синхронизированной экспрессии.
По этому изобретению промежуточные беталактамные соединения 3-(карбоксиэтилтио)пропионил-7-АДЦК продуцируются P.chrysogenum с помощью добавления 3,3'-тиодипропионовой кислоты или ее соли или эфира. Подходящими солями, например, являются соли натрия или калия. Они же эффективно выделяются из среды путем простой экстракции растворителем, например, следующим образом.
Жидкая питательная среда отфильтровывается, и к фильтрату добавляется несмешиваемый с водой органический растворитель. pH доводится так, чтобы проэкстрагировать цефалоспорин из водного слоя. Диапазон pH должен находиться ниже 4,5; предпочтительно в интервале от 4 до 1, более предпочтительно - от 2 до 1. Таким образом цефалоспорин отделяется от многих посторонних примесей, присутствующих в ферментационной среде. Предпочтительно используется небольшой объем органического растворителя, что дает концентрированный раствор цефалоспорина и таким образом достигается снижение объемных скоростей потоков. Вторая возможность состоит в полной экстракции жидкой питательной среды при pH, равном 4 или ниже. Предпочтительно, жидкая питательная среда экстрагируется в интервале от 4 до 1 органическим растворителем, несмешиваемым с водой.
Может использоваться любой растворитель, который не взаимодействует с молекулой цефалоспорина. Подходящими растворителями, например, являются бутилацетат, этилацетат, метилизобутилкетон, спирты, подобные бутанолу, и т. д. Предпочтительно используются 1-бутанол или изобутанол.
Затем цефалоспорин экстрагируется обратно водой при pH от 4 до 10, предпочтительно от 6 до 9. И снова конечный объем сильно снижается. Выделение может проводиться при температурах от 0 до 50oC, и предпочтительно при температурах окружающей среды.
Водный раствор цефалоспорина, полученный таким образом, обрабатывается соответствующим ферментом для удаления 3-(карбоксиэтилтио)пропиониловой боковой цепи и получения желаемой 7-АДЦК.
Предпочтительно используется иммобилизированный фермент, чтобы было возможно использовать фермент повторно. Методология получения таких частиц и иммобилизации ферментов была подробно описана в ЕР-А-0222462. pH водного раствора должен иметь значение, равное, например, от pH 4 до pH 9, при котором реакция деградации цефалоспорина минимизирована, а желаемое превращение с помощью фермента оптимально. Таким образом, фермент добавляется к водному раствору цефалоспорина при поддержании в то же время pH на соответствующем уровне с помощью, например, добавления неорганического основания, такого, как раствор гидроксида калия, или применения катионообменной смолы. Когда реакция завершается, иммобилизированный фермент удаляется фильтрованием. Другая возможность состоит в применении иммобилизованного фермента в колонке с фиксированным или ожиженным слоем, или использовании фермента в растворе и удалении продуктов с помощью мембранной фильтрации. Впоследствии реакционную смесь подкисляют в присутствии органического растворителя, несмешиваемого с водой.
Подходящие ферменты, например, получаются из микроорганизма Pseudomonas SY77, имеющего мутацию в одной или более позиций: 62, 177, 178 и 179. Также могут использоваться ферменты из других микроорганизмов Pseudomonas, предпочтительно Pseudomonas SE83, имеющих, произвольно, мутацию одной или более позиций, соответствующих 62, 177, 178 и 179 у Pseudomonas SY77. После доведения pH до примерно от 0,1 до 1,5 слои разделяются и pH водного слоя доводится до примерно 2 - 5. Кристаллическая 7-АДЦК затем отфильтровывается.
Деацилирование может также выполняться химически, известным образом, например, через образование иминохлоридной боковой цепи путем добавления пентахлорида фосфора при температуре ниже 10oC и затем изобутанола при температуре окружающей среды или ниже.
Следующие примеры предлагаются в качестве иллюстрации, но не для ограничения.
Пример 1
Экспрессия гена cefE Streptomyces и Nocardia в Penicillium chrysogenum
а. Общие процедуры клонирования гена и генной трансформации
В данной заявке используются обычные методики, используемые в процедурах клонирования гена. Эти методики включают полимеразные цепные реакции (ПЦР), синтез синтетических олигонуклеотидов, анализ нуклеотидной последовательности ДНК, ферментативное лигирование и рестрикцию ДНК, субклонирование вектора в Е. coli, трансформацию и селекцию трансформантов, выделение и очистку ДНК, характеристику ДНК с помощью блот-анализа по Саузерну и меченых 32P зондов, мечение ДНК 32P путем случайного примирования. Все эти методики очень хорошо известны в этой области технологии и соответственно описаны во многих источниках. Смотрите, например, Sambrook et al.. Molecular Cloning:, a Laboratory Manual, Cold Spring Harbor, U.S.A. (1989), Innes et al., PCR protocols, a Guide to Metods and Applications, Academic Press (1990), and McPherson et al., PCR, a Practical Approach, IRL Press (1991).
Общие процедуры, использованные при трансформации нитчатых грибов и селекции трансформантов включает получение протопластов грибов, перенос ДНК и режим регенерации протопластов, очистку и определение свойств трансформантов. Все эти процедуры хорошо известны в этой области знаний и очень хорошо документированы в: Finkelstein and Ball (eds), Bio- technology of Filamentous Fungi, technology and products, Butterworth-Heinemann (1992); Bennet and Lasure (eds). More Gene Manipulations in Fungi, Academic Press (1991); Turner, in Puhler (ed.), Biotechnology, second completely revised edition, VCH (1992).
Более конкретные применения технологии клонирования генов и генной трансформации в отношении Penicillinum chrysogenum очень хорошо документированы у Bennet and Lasure (выше) и Finkelstein and Ball (выше).
- Синтетические ДНК олигонуклеотидов синтезированы с использованием коммерческого синтезатора ДНК (Applied Biosistems, CA, U.S.A.) в соответствии с инструкциями производителя.
- ПЦР выполняется с использованием коммерческого автоматического аппарата для ПЦР (Perkin Elmer, U.S.A.) и ДНК-полимеразы (Perkin Elmer) по инструкции производителя.
- Процедуру hGC PCR (Dutton et al., Nucleic Acids Res. 21, (no 12) (1993) 2953 - 2954) использовали, чтобы сделать возможной амплификацию кодирующих областей cefE хромосомной ДНК N. lactamdurans и S. clavuligerus.
- Ферменты рестрикции и другие ферменты для модификации ДНК получены из BRL (MD, U.S.A.) и использовались в соответствии с инструкциями производителя.
- Вектор pBluescript® E.coli получен от Stratagene (CA, U.S.A.).
- Другие использованные химреактивы все имеют аналитическую чистоту и получены от разных поставщиков.
- Анализ нуклеотидной последовательности ДНК выполняли с использованием аппарата для автоматического анализа последовательности ДНК (Applied Biosistems), основанного на детекции специфичной для последовательности флюоресцентной метки в соответствии с инструкциями производителя.
b. Культивирование микроорганизмов
Streptomyсes clavuligerus ATCC 27064 выращивается в трипсинизированном соевом бульоне (Difco). Хромосомная ДНК этого штамма использована для выделения гена cefE (Kovacevic et al., J. Bacteriol. (1989), 754 - 760).
Nocardia lactamdurans ATCC 27382 также выращивается в трипсинизированном соевом бульоне (Difco). Хромосомная ДНК этого штамма использована для выделения гена cefE (Coque et. al., выше).
Penicillium chrysogenum Wisconsin 54-1255 (ATCC 28089) выращивается в полной среде YPD (YPD; 1% дрожжевой экстракт, 2% пептона, 2% глюкозы). Хромосомная ДНК этого штамма использована для выделения 5' и 3' регуляторных областей гена penDE, необходимых для экспрессии гена cefE. Penecillium chrуsogenun ATCC 28089 также используется в качестве хозяина для экспериментов с трансформацией геном cefE. Пригодны также другие штаммы Penicillium chrisogenum, включая мутанты штамма Wisconsin 54-1255, дающие увеличенный выход беталактамов. В зависимости от использованного маркера селекции трансформанта могут использоваться штаммы P.chrysogenum, содержащие мутации в pyrG, niaD или facA гене. Эти мутантные штаммы могут быть получены с помощью методов, хорошо известных в этой области технологии (Cantoral, Bio/Technol. 5 (1987), 494 - 497; Gouka et al., J. Biotechn. 20 (1991), 189 - 200; and Gouka et al, Appl. Microbiol. Biotechnol. (1993), 514 - 519).
Культивирование P.chrysogenum для образования протопластов, используемых при трансформации, также производилось в среде YPD.
Хорошо известно, что процедуры образования протопластов и регенерации могут слегка отличаться в зависимости от конкретного использования штамма Penicillium chrysogenum и примененной процедуры селекции трансформанта.
Е. coli WK6 (Zell and Fritz, EMBO J. 6 (1987), 1809 - 1815), XLI-Blue (Stratagene) и HB101 (Boyer and Roulland-Dussoix, J. Mol. Biol., 41 (1969), 459; Bolivar and Backman, Messages Ensymol. 68 (1979), 2040) сохранялись и культивировались с помощью использования стандартной среды для культивирования Е. coli (Sambrook, выше).
с. Культивирование кассет экспрессии cefE
Кассеты экспрессии cefE перечислены в таблице 1, которая также объясняет номенклатуру, которая была использована для этих плазмид.
Опубликованные нуклеотидные последовательности гена cefE S.clavuligerus (Kovacevic, выше); гена cefE N.lactamdurans (Coque, выше); гена gpdA A.nidulans (Punt et al, Gene 69 (1988)); и гена penDE P.chrysogenum (Barredo et al. , Gene 83 (1989), 291 - 300; Dies et al., Mol. Gen. Genet. 218 (1989), были использованы для создания синтетических олигонуклеотидов, перечисленных в таблице II.
c1.. Конструирование плазмид экспрессии cefE E.coli pMCTSE и pMCTNE
ПЦР, 1: cefE N.lactamdurans
В первой ПЦР с использованием хромосомной ДНК N.lactamdurans и олигонуклеотидов 1 и 2, была получена открытая рамка считывания cfeE N.lactamdurans в виде продукта ПЦР 0,9 kb, содержащего уникальный сайт рестрикции Ndel на 5'-конце и уникальный сайт Xbal на 3'-конце.
ПЦР, 2: cefE S.clavuligerus
Во второй ПЦР с использованием хромосомной ДНК S.clavuligerus и олигонуклеотидов 3 и 4 была получена открытая рамка считывания cefE S.clavuligerus в виде продукта ПЦР в 0,9 kb, также содержащего уникальный сайт рестрикции NdeI на 5'-конце и уникальный сайт рестрикции Xbal на 3'-конце.
С целью получения экспрессии генов cefE и E.coli и определения характеристик продуктов ПЦР с помощью анализа последовательности ДНК, продукты ПЦР 1 и 2 клонировали в векторе pMCTNde, производный от рMC-5 (Stanssens et al., Nucleic Acids Res, 17 (1989), 4441), Плазмида pMCTNde получена из рМC5-8 (Европейская патентная заявка N 0351029) путем встройки фрагмента, кодирующего tac-промотор с последующим сайтом RBS и клонирующим сайтом Ndel (рис . 3).
Продукты ПЦР 1 и 2 расщепляли с помощью NdeI и XbaI и лигировали в расщепленный NdeI-XbaI вектор pMCTNde. Смесь лигирования использовали для трансформации E. coli WK6. Трансформанты отбирали по резистентности к хлорамфениколу. Эти транcформанты использовали для изоляции плазмидной ДНК. Вставку кассет экспрессии cefE сначала анализировали с помощью расщепления реcтрикционным ферментом на предсказанную генерацию рестрикционных фрагментов. Плазмиды, содержащие предсказанные сайты для рестрикционных ферментов окончательно проанализированы с помощью автоматического анализа последовательности ДНК,
Последовательность ДНК открытой рамки считывания cefE S.clavuligerus в плазмиде pMCTSE (рис .2) была на 100% идентична опубликованной последовательности (Kovacevic, выше).
Последовательность ДНК (рис. 8) всех клонов, которые были проанализированы, содержащая открытую рамку считывания cefE N.lactamdurans, отличалась от опукликованной последовательности (Coque, выше).
Аминокислотная последовательность, произведенная из опубликованного гена cefE N.lactamdurans имеет пролин в положении аминокислоты 41 (смотрите Посл. ИД N 14). Этот пролин опущен в клонах, которые получены при ПЦР 1. Эта плазмида названа pMCTNE (рис. 1).
с2. Конструирование плазмид экспрессии cefE P. chrysogenum
ПЦР, 3: промотор gpdA
При этой третьей ПЦР с использованием пламидной ДНК pAN7-1 (Punt et al., Gene 56 (1987) 117 - 124), содержащей ген hph E.coli под контролем промотора gpdA A. nidulans и олигонуклеотиды 5 и 6, был проучен промотор gpdA в виде продукта ПЦР 0,9 kb, содержащий единственный сайт рестрикции EcoRI на 5'-конце и единственный сайт NdeI на 3'-конце.
ПЦР, 4: промотор AT
В четвертой ПЦР хромосомная ДНК P.chrysogenum и олигонуклеотиды 7 и 8 были использованы для получения AT промоторного фрагмента, равного 1,5 kb, который также содержит единственный сайт рестрикции EcoRI на 5'-конце и единственный сайт NdeI на 3'-конце.
ПЦР, 5: AT терминатор и 3'-конец гена cefE
N.lactamdurans.
В пятой ПЦР была получена 0,5 kb терминаторная область penDE (AT) с использованием хромосомной ДНК P.chrysogenum и олигонуклеотидов 9 и 10, и 11 и 10, соответственно. Эти продукты ПЦР содержат, таким образом, 3'-концевую последовательность гена cefE с или без сигнала нацеливания к микротельцу, состоящего из C-концевой аминокислотной последовательности ARL (Muller et al., Biochimica et Biophysica Acta 1116 (1992), 210 - 213).
Олигонуклеотиды сконструированы таким образом, что единственный сайт BspEl встраивается на 5'-конце продукта ПЦР и единственный сайт SpeI вводится на 3'-конце продукта ПЦР.
ПЦР, 6: AT терминатор и 3'-конец гена cefE S.clavuligerus
В этой шестой ПЦР была получена 0,5 kb терминаторная область penDE (AT) с использованием хромосомной ДНК P.chrysogenum и олигонуклеотидов 12 и 10, и 13 и 10, соответственно. Эти продукты ПЦР, таким образом, содержат 3'-концевую последовательность гена cefE S.clavuligerus с или без сигнала нацеливания к микротельцу, состоящего из C-концевой аминокислотной последовательности SKL (De Hoop et al., Biochem. J. 286 (1992), 657 - 669).
Олигонуклеотиды сконструированы так, что единственный сайт рестрикции BqlI вводится на 5'-конце продукта ПЦР и единственный сайт SpeI получается на 3'-конце этого продукта ПЦР.
С целью получения экспрессии генов cefE в P.chrysogenum промотор gpdA и фрагмент промотора AT лигировали к фрагментам cefE из плазмид pMCTNE и pMCTSE. Эти лигированные фрагменты клонировали в вектор pBluescript II KS.
Продукт ПЦР 3 расщепляли EcoRI и Ndel. pMCTNE и pMCTSE расщепляли с помощью NdeI и XbaI. Фрагменты рестрикции разделяли с помощью электрофореза в агарозном геле. Кодирующие фрагменты 0,9 kb cefE очищали из агарозного геля. Промоторный фрагмент EcoRI-Ndel лигировали вместе с фрагментами NdeI-XbaI cefE в расщепленный EcoRI-XbaI вектор pBluescript II KS. Таким образом были получены следующие плазмиды: pGSE и pGNE.
Для получения оптимальной экспрессии генов cefE в P.chrysogenum мы выбрали клонирование сигнальной последовательности терминации AT после генов cefE в плазмидах экспрессии Penicillium, упомянутых выше.
pGNETA-pGNEWA
Продукты ПЦР 5 расщепляли с помощью BspEI и SpeI и лигировали в расщепленный BspEI и SpeI вектор pGNE. Смеси лигирования использовали для трансформации E. coli НВ101. Трансформанты отбирали по устойчивости к ампициллину. Плазмиды, выделенные из этих трансформантов, охарактеризовывали с помощью анализа фрагментов рестрикции и затем с помощью анализа последовательности ДНК. Таким образом были получены следующие плазмиды: pGNEWA и pGNETA (рис. 4).
pGSETA-pGSEWA
Продукты ПЦР 6 расщепляли с помощью BqlI и SpeI и лигировали в расщепленный BqlI и SpeI вектор pGSE. Смеси лигирования использовали для трансформации E. coli НВ101. Трансформанты отбирали по устойчивости к ампициллину. Выделенные из этих трансформантов плазмиды также охарактеризовывали с помощью анализа фрагментов рестрикции и затем анализа последовательности ДНК. Таким образом были получены следующие плазмиды: pGSWA и pGSETA (рис. 5).
pANETA, pANEWA и pASETA и pASEWA
Плазмиды pGNETA, pGNEWA, pGSETA и pGSEWA расщепляли с помощью EcoRI и NdeI. Фрагменты рестрикции разделяли с помощью электрофореза в агарозном геле и из геля очищали фрагменты 4.5 kb.
Продукт ПЦР 4 расщепляли с помощью EcoRI и NdeI и лигировали с очищенными фрагментами, упомянутыми выше. После трансформации лигированных смесей в E.coli НВ101, трансформанты отбирали по резистентности к ампициллину.
Трансформанты подращивали и выделяли их плазмиды и характеризовали с помощью анализа фрагментов рестрикции и окончательно - с помощью анализа последовательности ДНК. Таким образом были получены желаемые конструкции, а именно pANETA (рис. 6), pANEWA, pASETA (рис. 7) и pASEWA.
d. Трансформация P.chrysogenum
Использована процедура трансформации протопластов, опосредуемой Ca-PEG.
По методам, описанным у Cantoral (смотрите выше), Gouka et al. (J.Biotechn., смотрите выше) и Gouka et al. (Appl. Miсrobiol. Biotechnol., смотрите выше) для трансформации штамма P.chrysogenum использовали целую плазмиду или очищенный полигенный экспрессирующий кластер cefE (лишенный векторных последовательностей E.coli) с генами pyrG, niaD, facA или amdS (Beri et al., Curr. Genet. 11 (1987) 639 - 641), соответственно, в качестве маркеров селекции.
Путем использования гомологичных маркеров селекции pyrG, niaD или facA, в чистой форме, лишенных векторных последовательностей E.coli, были получены трансформированные штаммы P.chrysogenum, которые не содержат бактериальных генов резистентности.
В Европейской патентной заявке N 94201896.1 описывается метод получения рекомбинантных штаммов, лишенных генов маркеров селекции. Этот метод был успешно использован на трансформантах P.chrysogenum, содержащих ген A.nidulans amdS в качестве доминантного маркера селекции.
В таком случае единственными элементами гетерологического происхождения являются кодирующая область cefE 0,9 kb и, произвольно, промоторная область 0,9 kb gpdA.
е. Анализ трансформантов
Трансформанты P. chrysogenum очищаются путем повторного культивирования на селективной среде. Единичные устойчивые колонии использованы для приготовления агаровых косяков для получения спор и для проверки на трансформанты, содержащие кассету экспрессии cefE. Кипячение фрагмента свежего мицелия из трансформантов на агаровой пластинке использовали для получения достаточного количества матричной ДНК для эффективной проверки сотен трансформантов на присутствие гена cefE с использованием методики ПЦР. (Seth, Fungal Genetis Conference, Asilomar (1991), Fungal Genetics Newsletter 38, 55). Действуя таким образом установили эффективность трансформации.
Отбор трансформантов также выполняли, применяя биологическое исследование. Трансформанты подращивали на агаризованной среде, которая содержала выбираемые предшественники боковой цепи. E.coli ESS2231 использовали в качестве индикаторной бактерии в кроющем слое агара, который также содержал Бактопеназу, чтобы было возможно отдифференцировать продукцию пенициллина и цефалоспорина по методам, хорошо известным в этой области науки и описанным, например, у Guttierez et al., Mol. Gen. Genet. 225 (1991), 56 - 64.
Споры использовали для посева P.chrysogenum в культуральную среду, как описано в разделе d. После 72 часов культивирования (при 25oC) из мицелия выделяют хромосомную ДНК. ДНК расщепляют рестрикционным ферментом с распознаваемой последовательностью в 6 п.о., подобным EcoRI или PstI.
Фрагменты ДНК разделяли с помощью электрофореза в агарозном геле и переносили на нейлоновые мембраны Gene screen (New England Nuclear). Блоттинг по Саузерну проводили гибридизацией с меченным 32P продуктом ПЦР 2 в качестве пробы для последовательностей гена cefE. Мечение 32P очищенного продукта ПЦР 2 достигается с помощью мечения случайным примированием в присутствии α32P дЦТФ при использовании коммерческого набора для мечения (Boeringer Mannheim).
Трансформанты, содержащие кодирующую последовательность cefE, испытаны на экспрессию продукта гена cefE, называемую здесь экспандазной активностью.
Отобранные транcформанты культивируются в среде для продукции пенициллина (смотрите пример 2).
В эксперименте по оценке протекания процесса во времени образцы мицелия - отбирали через 48, 72 и 96 часов ферментации. Готовятся экстракты мицелия, и экспандазная активность определяется в неочищенных экстрактах, как описано у Rollins et al., Can. J. Microbiol. 34 (1988), 1196 - 1202. Трансформанты с экспандазной активностью испытывали на ацилтрансферазную активность также методами описанными у Alvares et al., Antimicrob. Agent Chem. 31 (1987), 1675 - 1682).
По этим анализам отбирали трансформанты с различными уровнями ацитрансферазной и экспандазной ферментативной активности для ферментативной продукции производных 7-АДЦК.
Пример 2
Ферментативное получение 3-(карбоксиэтилтио)пропионил-7-АДЦК и ее выделение
P. chrysogenum штамм Wisconsin 54-1255 (ATCC 28089) трансформируется с помощью одной из конструкций ДНК, как описано в примере 1 и засевается в концентрации 2•106 конидий/мл в посевную среду, состоящую из (г/л): глюкозы, 30; (NH4SO4, 10; KH2PO4, 10; раствора микроэлементов I (MgSO4•7H2O, 25; FeSO4•7H2O, 10; CuSO4••5H2O, 0,5; ZnSO4•7H2O, 2; Na2SO4, 50; MnSO4•H2O, 2; CaCl2•2H2O, 5), 10 (мл/л) (pH до стерилизации 6,5).
Посевную культуру инкубировали в течение 48 - 72 часов при 25-30oC и затем используют для засева 10-20 объемов производственной среды, содержащей (г/л) лактозы, 80; мальтозы, 20; CaSO4, 4; мочевины, 3; MgSO4•7H2O, 2; KH2PO4, 7; NaCl, 0,5; (NH4)2SO4, 6; FeSO4•7H20, 0,1; 3,3'-тиодипропионовой кислоты, 5; раствор микроэлементов II (CuSO4•5H2O, 0,5; ZnSO4•7H20, 2; MnSO4•H2O, 2; Na2SO4, 50), 10 (мл/л) (pH до стерилизации 5,5-6,0). Затем инкубация продолжается в течение еще 96-120 часов.
В конце производственной ферментации мицелий отделяется центрифугированием или фильтрованием, и 3-(карбоксиэтилтио)пропионил-7-АДЦК анализируют с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ) на колонке с обращенной фазой. Использован аппарат для ВЭЖХ Beckman System Gold, состоящий из программируемой системы для растворителей 168, автоматического пробоотборника модели 507, диодно-сетчатого детектора модели 168 и системы показателей System Gold (5.10). В качестве стационарной фазы использовали две (2) катриджных колонки Chromspher C18 (100 х 3 мм, Chrompack) в ряд. Подвижная фаза состоит из линейного градиента от 100% 0.07М фосфатного буфера pH 4,8 до 25% ацетонитрила и 75% фосфатного буфера pH 4,8 через 15 минут при скорости потока, равной 0,5 мл/мин. Продукция 3-(карбоксиэтилтио)пропионил-7-АДЦК определяется количественно при 260 нм с использованием 3-(карбоксиэтилтио)пропионил-7-АДЦК в качестве вещества для сравнения.
Идентичность пика подтверждается путем сравнения кривых ультрафиолетового и ЯМР спектров.
После фильтрования жидкой питательной среды к фильтрату добавляется примерно 0,1 объема 1-бутанола. Значение pH доводится до 2 с помощью разбавленной хлористоводородной кислоты, и смесь перемешивается в течение 5 минут при комнатной температуре. После разделения органический слой или выпаривается или используется далее для химического дезацилирования (пример 3) или обратно экстрагируется объемом воды, равным 0,33 с pH 8 и далее используется при ферментативном дезацилировании (пример 4).
Пример 3
Дезацилирование 3-(карбоксиэтилтио)пропионил-7-АДЦК
К смеси 3 г (8 ммолей) 3-(карбоксиэтилтио)пропионил-7-АДЦК, 3,5 мл (36 ммолей) N,N-диметиланилина, 13 мл метиленхлорида и при комнатной температуре добавляется 2,6 мл (12 ммоль) триметилхлорсилана. После перемешивания в течение 30 минут реакционную смесь охлаждают до примерно -50oC и добавляют 1,8 г (8,5 ммоля) пентахлорида фосфора, все сразу. Температура поддерживается на уровне -40oC в течение двух часов и затем реакционную смесь охлаждают до -65oC. Затем ее обрабатывают 12 мл (137 ммолей) изобутанола с такой скоростью, чтобы температура не повышалась свыше -40oC. После дополнительного перемешивания в течение двух часов раствор выливают в 15 мл воды и потом сразу добавляют 5 мл 4,5 N аммиака. pH доводится до 4 путем медленного добавления твердого бикарбоната аммония. После охлаждения до 5oC смесь фильтруют и кристаллическую 7-АДЦК промывают 5 мл водного ацетона (1:1) и отделяют.
Пример 4
Ферментативное деацилирование 3-(карбоксиэтилтио)пропионил-7-АДЦК с использованием мутантной ацилазы Pseudomonas SY77
Превращение 3-(карбоксиэтилтио)пропионил-7-АДЦК выполняется в одну энзиматическую стадию с использованием специфической ацилазы, которая производится из ацилазы Pseudomonas SY77 путем направленного на определенную область мутагенеза. Конструирование и идентификация мутантной ацилазы Pseudomonas SY77 с улучшенной активностью в отношении 3-(карбоксиэтилтио)пропионильной боковой цепи было описано в EP-A-0453048. В мутанте тирозин в положении 178 в альфа-субъединице ацилазы Pseudomonas SY77 заменен на гистидин. Мутантная ацилаза продуцируется E.coli. Клетки собираются центрифугированием и снова суспендируются в 10 мМ фосфатном буфере pH 7,4, содержащем 140 мМ NaCl. Затем клетки разрушают ультразвуком. После удаления клеточных остатков собирали супернатанты, обладающие ацилазной активностью. Дальнейшую очистку ацилазы выполняли с помощью ряда этапов хроматографии:
(1) ионообменной хроматографии на Q-сефазоре с быстрым протеканием при pH 8,8;
(2) хроматография гидрофобного взаимодействия на фенилсефарозе; и
(3) гельпроникающей хроматографии на колонке с сефакрилом.
Очищенную ацилазу иммобилизировали на частицах, состоящих из смеси желатина и хитозана. Частицы обрабатывали глутаральдегидом непосредственно перед добавлением фермента.
Превращение 3-(карбоксиэтилтио)пропионил-7-АДЦК выполняли в реакторе с мешалкой. Сначала в реактор добавляют водный раствор цефалоспорина. Затем температура раствора доводится до 30oC при постоянном перемешивании и pH фиксируется на 8 с помощью гидроксида калия. Затем добавляется иммобилизированный фермент и превращение начинается. Во время превращения pH в реакторе непрерывно контролируется и поддерживается на 8. 3,3'-тиодипропионовая кислота, которая освобождается при реакции, титруется КОН. Количество КОН, которое добавляется, суммируется и регистрируется на записывающем устройстве с плоским стендом. Превращение контролируется путем отбора образцов из реактора, которые анализируются на 3-(карбоксиэтилтио)пропионил-7-АДЦК и 7-АДЦК с помощью ВЭЖх, как описано в примере 2.
Когда реакция завершена, иммобилизированный фермент удаляется фильтрованием и pH фильтрата доводится до 1, тогда как фильтрат содержит бутилацетат. Слои разделяются, и pH водной фазы, которая содержит 7-АДЦК, доводится до 3. Кристаллическая 7-АДЦК затем отфильтровывается.
Пример 5
Ферментативное деацилирование 3-(карбоксиэтилтио)пропионил-7-АДЦК с использованием ацилазы Pseudomonas SE83
Превращение 3-(карбоксиэтилтио)пропионил-7-АДЦК выполняется, как и в примере 4, однако при использовании в качестве ацилазы ацилазы Pseudomonas SE83 с получением того же самого результата.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЭФФЕКТИВНОГО ПРОИЗВОДСТВА 7-АДЦК ЧЕРЕЗ 2-(КАРБОКСИЭТИЛТИО) АЦЕТИЛ-7-АДЦК И 3-(КАРБОКСИМЕТИЛТИО)ПРОПИОНИЛ-7-АДЦК | 1994 |
|
RU2139349C1 |
| СПОСОБ ПОЛУЧЕНИЯ 7-АДЦК ПУТЕМ ВОЗДЕЙСТВИЯ АКТИВНОСТИ ЭКСПАНДАЗЫ НА ПЕНИЦИЛЛИН G | 1996 |
|
RU2192471C2 |
| СПОСОБ ПОЛУЧЕНИЯ 7-АМИНОЦЕФАЛОСПОРАНОВОЙ КИСЛОТЫ, ВЕКТОР ЭКСПРЕССИИ, РЕКОМБИНАНТНЫЙ ШТАММ | 1992 |
|
RU2202616C2 |
| СПОСОБ РАСШИРЕНИЯ 5-ЧЛЕННОГО КОЛЬЦА СОЕДИНЕНИЯ БЕТА-ЛАКТАМА ДО 6-ЧЛЕННОГО ЦЕФЕМА | 1999 |
|
RU2230790C2 |
| СПОСОБ ПОЛУЧЕНИЯ N-ДЕАЦИЛИРОВАННОГО ЦЕФАЛОСПОРИНА | 1998 |
|
RU2208644C2 |
| СПОСОБ ПОЛУЧЕНИЯ 7-АМИНОДЕЗАЦЕТОКСИЦЕФАЛОСПОРАНОВОЙ КИСЛОТЫ (7-ADCA) | 1992 |
|
RU2178808C2 |
| СПОСОБ ПОЛУЧЕНИЯ N-ДЕАЦИЛИРОВАННОГО ЦЕФАЛОСПОРИНА | 1998 |
|
RU2208643C2 |
| СПОСОБ ПОЛУЧЕНИЯ ШТАММА BACILLUS, СОДЕРЖАЩЕГО ДВЕ КОПИИ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК, КОДИРУЮЩЕЙ ФЕРМЕНТ С ГИДРОЛАЗНОЙ АКТИВНОСТЬЮ | 1988 |
|
RU2091487C1 |
| СПОСОБ ПОЛУЧЕНИЯ СЕРИНОВОЙ ПРОТЕАЗЫ, ШТАММ ЩЕЛОЧЕФИЛЬНЫХ BACILLUS-ПРОДУЦЕНТ СЕРИНОВОЙ ПРОТЕАЗЫ | 1988 |
|
RU2023723C1 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ ЦЕФАЛОСПОРИНА | 1998 |
|
RU2203323C2 |



Изобретение относится к биотехнологии. Представлен завершенный эффективный процесс получения и выделения 7-аминодезацетоксицефалоспорановой кислоты (7-АДЦК) через 3-(карбоксиэтилтио)пропионил-7-АДЦК с использованием трансформантного штамма Penicillium chrysogenum, экспрессирующего экспандазу в сочетании с ацилтрансферазой. В способе применяется новый предшественник боковой цепи -3,3'-тиодипропионовая кислота. Способ позволяет повысить продуктивность штаммов для получения 7-АДЦК, являющейся промежуточным соединением при производстве цефалоспоринов. 4 з.п.ф-лы, 8 ил., 2 табл.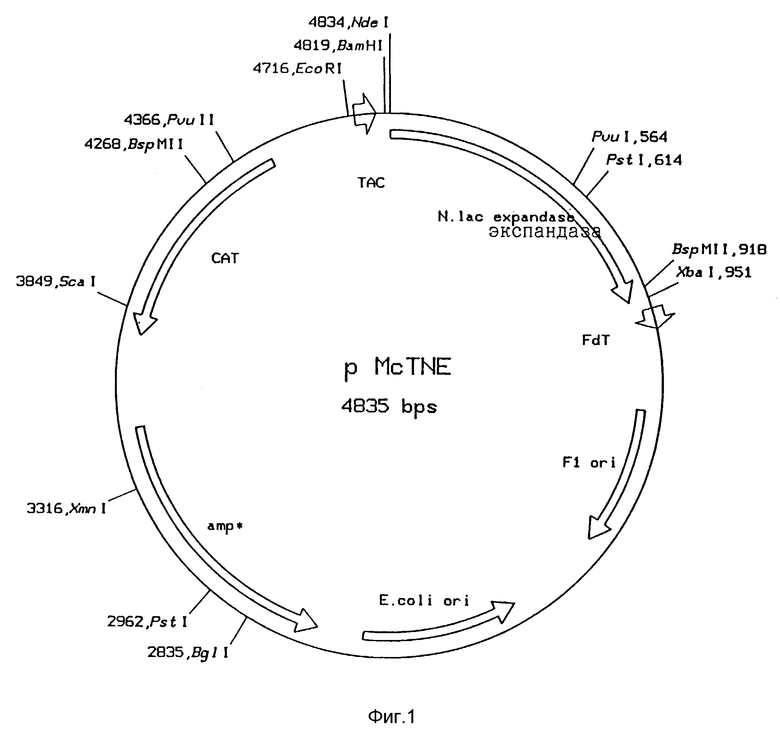
| Способ выделения фермента, катализирующего превращение креатинина в креатин и фермента, катализирующего превращение креатина в саркозин и мочевину | 1972 |
|
SU532341A3 |
| Потенциметрический титрограф | 1975 |
|
SU540210A1 |
| Способ получения фентиасилинов | 1972 |
|
SU448180A1 |

