Данное изобретение относится к области способов рекомбинантных ДНК. Более конкретно, данное изобретение относится к области получения цефалоспоринов с использованием генетически модифицированных организмов-хозяев, к организмам-хозяевам, модифицированным таким образом, а также к ДНК для применения в них. Кроме того, данное изобретение относится к модифицированным ферментам, экспрессируемым из рекомбинантной ДНК в указанных организмах-хозяевах.
Антибиотики ряда β-лактама составляют наиболее важную группу антибиотических соединений с длительной историей клинического применения. Среди этой группы наиболее известными являются пенициллины и цефалоспорины. Эти соединения природно продуцируются нитеобразными грибками, такими как Penicillium chrysogenum и Acremonium chrysogenum соответственно.
Как результат классических способов улучшения штаммов уровни продуцирования этих антибиотиков в Р. chrysogenum и А. chrysogenum увеличились драматически на протяжении последних десятилетий. С увеличением знания биосинтетических путей, ведущих к пенициллинам и цефалоспоринам, и появлением технологии рекомбинантных ДНК стали доступны новые инструменты для улучшения производственных штаммов и для дериватизации in vivo этих соединений.
Большинство ферментов, участвующих в биосинтезе β-лактама, были идентифицированы, и их соответствующие гены были клонированы, как можно узнать из Ingоlia and Queener, Med. Res. Rev. 9 (1989), 245-264 (пути и ферменты биосинтеза), и Aharonowitz, Cohen and Martin, Ann. Rev. Microbiol. 46 (1992), 461-495 (клонирование генов).
Две первые стадии в биосинтезе пенициллина в Р. chrysogenum являются конденсацией трех аминокислот L-5-амино-5-карбоксипентановой кислоты (L-α-аминоадипиновой кислоты) (А), L-цистеина (С) и L-валина (V) в трипептид LLD-ACV, с последующей циклизацией этого трипептида с образованием изопенициллина N. Это соединение содержит типичную структуру β-лактама.
Третья стадия включает в себя замену гидрофильной боковой цепи L-5-амино-5-карбоксипентановой кислоты гидрофобной боковой цепью под действием фермента ацилтрансферазы. Эта ферментативная обменная реакция, опосредованная ацилтрансферазой, имеет место в клеточной органелле, микротельце, как было описано в Европейском патенте ЕР-А-0448180.
Цефалоспорины являются гораздо более дорогостоящими, чем пенициллины. Одной из причин этого является то, что некоторые цефалоспорины (например, цефалексин) получают из пенициллинов с использованием ряда химических превращений. Другой причиной является то, что до сих пор только цефалоспорины с D-5-амино-5-карбоксипентаноильной боковой цепью могли подвергаться ферментативному превращению. Цефалоспорин С, несомненно, наиболее важный исходный материал в этом отношении, является очень хорошо растворимым в воде при любом рН, что подразумевает длительные и дорогостоящие процессы выделения с использованием громоздкой и дорогостоящей колоночной технологии. Цефалоспорин С, полученный таким путем, должен быть превращен в терапевтически применимые цефалоспорины с использованием ряда химических и ферментативных превращений.
Способы, предпочитаемые в настоящее время в промышленности для получения промежуточного продукта 7-ADCA, включают в себя сложные химические стадии, приводящие к расширению и дериватизации пенициллина G. Одна из необходимых химических стадий для получения 7-ADCA включает в себя расширение 5-членной кольцевой структуры пенициллина до 6-членной кольцевой структуры цефалоспорина (см., например, патент США US 4003894). Эта сложная химическая переработка является как дорогой, так и вредной для окружающей среды.
Вследствие этого существует большая потребность в замене подобных химических процессов ферментативными реакциями, такими как ферментативный катализ, предпочтительно в процессе in vivo. Ключом к замене химического процесса расширения ферментативными процессами и процессами in vivo является центральный фермент в пути биосинтеза цефалоспорина, дезацетоксицефалоспорин С-синтетаза (DAOCS), также называемая "экспандазой".
Было обнаружено, что фермент экспандаза из бактерии Streptomyces clavuligerus производит в некоторых случаях расширение кольца пенициллина. При введении в Р. chrysogenum он может превращать структуру кольца пенициллина в структуру кольца цефалоспорина in vivo, как описано в Cantwell et al., Proc. R. Soc. Lond. B. 248 (1992), 283-289. Фермент экспандаза хорошо охарактеризован (ЕР-А-0366354) как биохимически, так и функционально, так же, как и соответствующий ему ген cefE. Были описаны как физические карты гена cefE (EP-A-0341892), так и последовательность ДНК, а также исследования по трансформации Р. chrysogenum геном cefE.
Другим источником фермента расширения кольца являются, например, бактерия Nocardia lactamdurans (ранее Streptomyces lactamdurans} и грибок Acremonium chrysogenum (ранее Cephalosporium acremonium). Экспандаза из Cephalosporium является бифункциональным ферментом, имеющим как экспандазную (расширение кольца), так и 3-гидроксилирующую активность. Были описаны как биохимические свойства этого фермента, так и последовательность ДНК этого гена (Cortes et al., J. Gen. Microbiol. 133 (1987), 3165-3174 и Coque et al., Mol. Gen. Genet. 236 (1993), 453-458 соответственно).
Поскольку экспандаза катализирует расширение 5-членного тиазолидинового кольца пенициллина N до 6-членного дигидротиазинового кольца DAOC, этот фермент был бы логическим кандидатом для замены стадий расширения кольца этого химического процесса. К сожалению, этот фермент работает на промежуточном продукте пенициллина N пути биосинтеза цефалоспорина, но не функционирует или функционирует относительно неэффективно на легкодоступных недорогих пенициллинах, продуцируемых Р. chrysogenum, таких как пенициллин V или пенициллин G. Пенициллин N не является коммерчески доступным, и даже когда он расширен, его D-5-амино-5-карбоксипентаноильная боковая цепь не может быть легко удалена пенициллинацилазами.
Недавно было обнаружено, что экспандазный фермент способен расширять пенициллины с не встречающимися в природе боковыми цепями до соответствующих 7-АDСА-производных. Эти боковые цепи включают в себя адипат (5-карбоксилпентановую кислоту) и различные другие соединения. Это свойство экспандазы было использовано в способе получения in vivo адипоил-7-ADCA в Penicillium chrysogenum, описанном в ЕР-А-0532341. Штаммы Penicillium chrysogenum трансформируют экспандазой Streptomyces clavuligerus для продуцирования адипоил-6-аминопенициллановой кислоты (адипоил-6-АПК), при подаче в среду этим трансформантам адипиновой кислоты. Затем адипоил-6-АПК "расширяют" до адипоил-7-ADCA. Получение различных других 7-ацилированных цефалоспоринов было описано в WO 95/04148 и WO 95/04149. В WO 95/04148 и WO 95/04149 было описано, что добавление 3'-карбоксиметилтиопропионовой кислоты и 3,3'-тиодипропионовой кислоты к среде давало пенициллины, которые являются субстратами для экспандазы, приводящими к синтезу 2-(карбоксиэтилтио)ацетил-7-ADCA и 3-(карбоксиметилтио)пропионил-7-ADCA соответственно.
Даже хотя экспандаза проявляет активность на пенициллинах с отличающимися боковыми цепями, расширение этих новых пенициллинов происходит менее эффективно в сравнении с расширением природного субстрата, пенициллина N. Это мнение было основанием для модификации экспандазы с целью изменения активности на адипоилпенициллановой кислоте. Использование мутагенизированной экспандазы было описано в WO 98/02551.
Конечная стадия в синтезе пенициллинов, замена α-аминоадипоильной боковой цепи IPN альтернативной боковой цепью, происходит в микротельце, где локализована ацилтрансфераза. Таким образом, считается, что для оптимального расширения пенициллина предпочтительной является локализация экспандазы в микротельце. Это основано не только на том факте, что субстрат для экспандазы, как принято считать, продуцируется в микротельце, но также на предположении, что в микротельце экспандаза должна быть лучше защищена от протеолитической деградации, которая является известным препятствием в применении Р. chrysogenum для получения ферментов (см., например, Theilgaard et al., 1997; Purification and characterisation of δ-(L-α-aminoadipyl)-L-cysteinyl-D-valine synthetase from Penicillium chrysogenum; Biochem. J. 327, 185-191).
Модификация клеточной локализации ферментов, участвующих в продуцировании бета-лактамного антибиотика, предлагалась в ЕР 0448180. Например, на странице 7 в предпоследнем абзаце предлагается локализация эпимеразы и предпочтительно последующих ферментов биосинтеза цефалоспорина, таких как экспандаза/гидроксилаза из Acremonium chrysogenum или экспандаза из Streptomyces spec., в микротельцах для улучшения эффективности продуцирования цефалоспорина или промежуточных продуктов в Penicillium chrysogenum. Здесь показано также, что удаление таргетирующего на микротельца сигнала ацилтрансферазы, фермента, участвующего в удалении боковой цепи ацил-6-АПК, нарушает одновременно продуцирование пенициллина. Это является серьезным основанием для предположения, что ферменты, участвующие в образовании бета-лактама, должны быть локализованы в микротельцах, если не по причинам локализации субстрата, то по меньшей мере по причинам стабильности.
Данное изобретение связано с ДНК, гетерологичными клетками-хозяевами, способными транскрибировать, транслировать или экспрессировать указанную ДНК, и способами с использованием таких клеток-хозяев и их культур для улучшенного получения in vivo ацилированных цефалоспоринов с высоким выходом. Согласно данному изобретению обеспечена клетка-хозяин, содержащая фермент, имеющий экспандазную активность, который преимущественно локализован в цитозоле (в противоположность ферменту, локализованному в основном в пероксисомах или микротельцах или с пероксисомами или микротельцами) клетки-хозяина.
В предпочтительном варианте данное изобретение связано с выделенным фрагментом ДНК, кодирующим модифицированную экспандазу, которая после экспрессии указанного фрагмента ДНК в Penicillium chrysogenum в качестве хозяина обнаруживается преимущественно в цитозоле этого Penicillium-хозяина. Такой ДНК-фрагмент, кодирующий модифицированную экспандазу, может быть, например, получен модификацией ДНК, кодирующей экспандазу, как обнаружено с использованием Nocardia lactamdurans (ранее Streptomyces lactamdurans) и грибка Acremonium chrysogenum и других родственных (т.е. продуцирующих цефалоспорин/цефамицин) микроорганизмов.
Данное изобретение связано с выделенным фрагментом ДНК, кодирующим модифицированную экспандазу, которая предпочтительно локализуется в цитозоле, а не в микротельцах гетерологичного хозяина. В предпочтительном варианте данного изобретения такой фрагмент ДНК может быть получен из ДНК Streptomyces clavuligerus, кодирующей экспандазу, и модифицирован для получения фрагмента ДНК, кодирующего экспандазу, который при экспрессии в хозяине Penicillium chrysogenum обнаруживается преимущественно в цитозоле указанного Penicillium-хозяина.
Такая модификация включает в себя клонирование рекомбинантными способами нуклеотидной последовательности, кодирующей указанную экспандазу, но предпочтительно включает в себя модификацию рекомбинантными способами последовательности, кодирующей нацеливающий на микротельца или на пероксисомы сигнал в указанной экспандазе. Модификация может предусматривать единственную мутацию или множественные мутации (такие как делеция, вставка или реверсия) последовательности нуклеиновой кислоты, кодирующей экспандазу, предпочтительно последовательности, кодирующей карбокси-концевую аминокислотную последовательность указанной экспандазы.
На эукариотических организмах таргетирование белков на пероксисомы или микротельца было исследовано относительно хорошо в не-нитеобразных дрожжах. В этих организмах белки, предназначенные для пероксисомы или микротельца, обычно содержат С-концевую таргетирующую последовательность, аминокислотная последовательность которой обычно представляет собой S (серин) К (лизин) L (лейцин) или, более часто, S/C/A/ - K/H/R - L. Такая таргетирующая последовательность, по меньшей мере, в гомологичном контексте обеспечивает необходимый сигнал для белка, такого как фермент, для направления его к микротельцу, где он находит свою конечную локализацию.
Для филаментных грибков, таких как Penicillium chrysogenum, таргетирование белков на пероксисомы или микротельца исследовалось относительно мало, и только очень мало известно о таргетировании белков в гетерологичном контексте, когда организм, такой как дрожжи или грибок, используют в качестве клетки-хозяина для трансляции или даже экспрессии гетерологичного белка. Это, в частности, имеет место, когда гетерологичный белок получают из прокариотического организма, такого как бактерия, так как бактерии не имеют органелл, подобных пероксисомам или микротельцам, и, следовательно, не используют таргетирующий на пероксисомы сигнал в гомологичном контексте. Обычно считается, что для того, чтобы гетерологично экспрессируемый белок или фермент хорошо функционировал в клетке-хозяине, соответствующие таргетирующие сигналы должны быть по меньшей мере похожими на сигналы, используемые этим хозяином, для того, чтобы найти сравнимую субклеточную локализацию.
Экспандаза из бактерии S. clavuligerus обычно содержит С-концевую последовательность, SKA (серин-лизин-аланин), которая, по-видимому, функционирует в качестве таргетирующей на пероксисомы последовательности при экспрессии этого белка в гетерологичном контексте в эукариотическом хозяине. Например, штаммы Penicillium chrysogenum, трансформированные экспандазой S. clavuligerus (как, например, обсуждаемые в ЕР 0532341 А1), обнаруживают локализацию соответствующей экспандазы в пероксисоме.
Было обнаружено, что экспандаза дикого типа из Streptomyces clavuligerus при продуцировании рекомбинантно в Penicillium chrysogenum преимущественно накапливается в микротельцах. Вышеуказанная последовательность трех аминокислот, Ser-Lys-Ala, служит, очевидно, в качестве таргетирующего на микротельца сигнала в Penicillium chrysоgenum, хотя она несколько отличается от консенсусной последовательности для таргетирующих на микротельца сигналов в дрожжах.
Когда сигналы пероксисомного таргетирования, а также сигналы, которые должны предотвращать локализацию в пероксисомах, были сконструированы на карбокси-конце экспандазы S. clavuligerus, или для этой же цели на карбокси-конце экспандазы/гидролазы Cephalosporium acremonium, эта тенденция локализоваться в пероксисоме не изменялась, независимо от различных карбокси-концевых последовательностей аминокислот, которые были добавлены (Verburg et al., Р.35, page 108. In: 7th Int. Symp. Gen. Industr. Microorg. 26 June - 1 July 1994, Montreal Canada).
По-видимому, этот фермент несет эффективный сигнал таргетирования, заставляющий его локализоваться, по меньшей мере, преимущественно в микротельце гетерологичной клетки-хозяина Penicillium chrysоgenum. Однако в одном варианте данного изобретения теперь обеспечивается эффективная модификация экспандазы, изменяющая тенденцию локализоваться в микротельце и приводящая к преимущественной локализации этой экспандазы в цитозоле хозяина.
Тенденция локализации при микротельце или пероксисоме управляется N-концевыми или С-концевыми сигналами пероксисомного таргетирования (PTS), причем модификация указанных PTS (будь то N-концевые или С-концевые PTS) по существу предотвращает такую локализацию в микротельцах. Примером такой модификации является модификация в последовательности нуклеотидов, кодирующей карбокси-концевой трипептид X1 - Х2 - Х3, например такая, в которой X1 является одним аминокислотным остатком, выбранным из группы, состоящей из серина, цистеина и аланина, Х2 является одним аминокислотным остатком, выбранным из группы, состоящей из лизина, гистидина и аргинина, и Х3 является лейцином или аланином. Такие трипептидные последовательности называются также SKL-подобными пептидами, и существенной последовательностью этого сигнала может быть небольшая аминокислота в начале, основной остаток на предпоследнем месте и большой неполярный остаток в С-концевом положении. Данное изобретение обеспечивает фрагмент ДНК, кодирующий экспандазу, причем указанный фрагмент, кодирующий SKL-подобный трипептид, был модифицирован или выбран таким образом, чтобы по существу предотвращать локализацию этой экспандазы в микротельцах и, следовательно, заставлять экспандазу локализоваться преимущественно в цитозоле клетки-хозяина при экспрессии в указанной гетерологичной клетке.
В предпочтительном варианте данного изобретения указанный фрагмент ДНК получают из ДНК, кодирующей экспандазу S. clavuligerus, первоначально содержащей последовательность нуклеотидов, кодирующую карбокси-концевой трипептид серин-лизин-аланин, которая модифицирована таким образом, чтобы заставлять экспандазу локализоваться преимущественно в цитозоле клетки-хозяина при экспрессии в ней. Примером такой модификации является, в частности, фрагмент ДНК данного изобретения, в котором последовательность нуклеотидов, кодирующая серин - лизин - аланин, была модифицирована в последовательность, кодирующую серин - лизин - аспарагиновую кислоту, или в последовательность, где аминокислота в последнем положении была делетирована или заменена, например, другим негидрофобным остатком.
Еще одна модификация содержит серин - серин - аспарагиновую кислоту или последовательность, где аминокислота в предпоследнем положении была делетирована или заменена остатком, не заряженным положительно.
Другие модификации включают в себя делении или модификации в первом положении этого трипептида, которые изменяют направление модифицированного таким образом белка в гетерологичном хозяине и заставляют его преимущественно локализоваться в цитозоле используемой клетки-хозяина.
Другие модификации включают в себя делеции или модификации любой из комбинаций этик трех аминокислот или кодонов, экспрессирующих эти карбокси-концевые аминокислоты.
С одной стороны, неожиданно было обнаружено, что преднамеренная модификация с использованием способов рекомбинантных ДНК, трипептидной последовательности SKA (или ее функциональных вариантов, как объяснено выше), которая, по-видимому, функционирует в качестве таргетирующего на микротельца сигнала, заставляет гетерологично экспрессируемый фермент накапливаться преимущественно в цитозоле клетки-хозяина, предпочтительнее, чем в микротельцах.
Однако, с другой стороны, вариации белков по их природе имеются в большом количестве, и в настоящее время можно найти микроорганизмы, которые содержат фермент, такой как экспандаза, который при экспрессии указанного фермента в гетерологичном хозяине будет преимущественно локализоваться в цитозоле этого хозяина. В частности, белки или ферменты, которые имеют С-концевую последовательность, отклоняющуюся от вышеописанной идентифицированной таргетирующей последовательности, являются вероятными кандидатами для испытания на цитозольную локализацию и отбора для дальнейшего использования в улучшении выходов при получении цефалоспорина. В конкретном варианте данное изобретение обеспечивает последовательность нуклеотидов, кодирующую экспандазу, которая при экспрессии в клетке-хозяине Penicillium chrysogenum локализуется преимущественно в цитозоле, улучшая тем самым выход цефалоспорина по сравнению с выходами клетки-хозяина, экспрессирующей сравнимый фермент внутри или вблизи микротелец.
Совершенно вопреки ожиданию фермент, обеспечиваемый данным изобретением, является, по-видимому, стабильным вне микротелец. Экспандаза, обеспечиваемая данным изобретением, все еще способна расширять пенициллины, даже пенициллины с не встречающимися в природе боковыми цепями (см., например, ЕР 97201196.9 и ЕР 97201197.7), до соответствующих 7-ADCA-производных.
Данное изобретение обеспечивает также выделенный фрагмент ДНК, кодирующий модифицированный иди отобранный фермент, обеспечиваемые данным изобретением, причем этот фрагмент дополнительно содержит регуляторные районы для экспрессии указанной ДНК в эукариотической клетке-хозяине. Например, в рамках квалификации специалиста в данной области является обеспечение фрагмента ДНК с регуляторными районами, функциональными в одной или нескольких клетках-хозяевах, происходящих из различных организмов, таких как животное, или растение, или дрожжи, или грибок. В предпочтительном варианте данное изобретение обеспечивает фрагмент ДНК, кодирующий модифицированный фермент, обеспечиваемый данным изобретением, содержащий регуляторные районы, пригодные для регуляции экспрессии в клетке-хозяине, происходящей из микроорганизма, такого как дрожжи или грибок.
Неожиданно было обнаружено, что в клетке-хозяине (например, Penicillium chrysogenum), трансформированной экспандазой, накапливающейся в основном в цитозоле, в противоположность образованию в микротельцах, продуцирование цефалоспориновых антибиотиков (например, адипил-7-ADCA) скорее увеличивается, чем уменьшается. То же самое имеет место даже в сравнении с клеткой-хозяином, которая была трансформирована ДНК, кодирующей сравнимую во всех других отношениях локализованную в микротельцах экспандазу.
Данное изобретение обеспечивает также клетку-хозяин или организм-хозяин (или его потомство), обеспеченные модифицированным или выбранным фрагментом ДНК, кодирующим локализованный в цитозоле фермент, обеспечиваемый данным изобретением. Такой организм-хозяин включает в себя, например, в результате трансформации этого организма-хозяина или предка этого организма-хозяина фрагмент ДНК, необязательно содержащий регуляторные районы или последовательности, содержащие последовательность нуклеотидов, кодирующую модифицированный фермент, обеспеченный данным изобретением. Такой хозяин предпочтительно выбран из группы, состоящей из растений и микроорганизмов, причем предпочтительно он является грибком, выбранным из группы, состоящей из Penicillium chrysogenum, Aspergillus nidulans или A. niger и Cephalosporium acremonium.
Данное изобретение обеспечивает также культуры клеток-хозяев или организмов-хозяев, обеспечиваемых данным изобретением. Такая культура содержит клетки, в которых функциональная экспандаза, способная расширять пенициллины с (не встречающимися в природе) боковыми цепями до соответствующих производных 7-ADCA, преимущественно обнаруживается в цитозоле. В предпочтительном варианте данное изобретение обеспечивает культуру для крупномасштабной ферментации, в которой модифицированная экспандаза, локализованная в цитозоле, способствует более высоким уровням производства адипоил-7-ADCA, чем в сравнимой крупномасштабной ферментационной культуре, использующей экспандазу дикого типа сравнимого происхождения.
Еще в одном варианте данное изобретение обеспечивает клетку-хозяин и ее культуры, обеспеченные рекомбинантным способом, с экспандазой, отобранной среди прочих по ее способности или тенденции преимущественно локализоваться в цитозоле гетерологичного хозяина, независимо от того, локализуется ли данная экспандаза в гомологичном контексте в микротельце или нет. Такая клетка-хозяин может быть выбрана путем обеспечения клетки-хозяина фрагментом ДНК, кодирующим экспандазу, и испытания того, локализуется ли эта экспандаза преимущественно в цитозоле. В предпочтительном варианте испытание на локализацию указанного фермента выполняют при помощи электронной микроскопии, как объясняется в экспериментальной части описания.
Данное изобретение обеспечивает также способ получения клетки- или организма-хозяина, обеспеченных модифицированным или отобранным фрагментом ДНК, кодирующим модифицированный или выбранный фермент, обеспечиваемый данным изобретением, предусматривающий контактирование клеток в условиях трансформации с фрагментом ДНК, обеспечиваемым данным изобретением, и отбор клеток, получивших эту ДНК. Трансформация клеток-хозяев, например Penicillium chrysogenum или других грибков, в общем достигается различными способами доставки ДНК, такими как опосредованное ПЭГ-Са поглощение протопластов, электропорация или способы бомбардировки частицами, и отбором трансформантов. См., например, Van den Hondel en Punt, Gene and Transfer and Vector Development for Filamentous Fungi, in: Applied Molecular Genetics of Fungi (Peberdy, Laten, Ogden, Bennett, eds.), Cambridge University Press (1991). Было описано применение доминантных и недоминантных селектируемых маркеров (Van den Hondel, выше). Селектируемые маркеры как гомологичного (полученные из Р. chrysogenum), так и гетерологичного (не происходящие из Р. chrysogenum) происхождения были описаны (Gouka et al., J. Biotechnol. 20 (1991) 189-200).
Применение различных маркеров для отбора трансформантов, гомологичных или гетерологичных, в присутствии или в отсутствие векторных последовательностей, физически связанных или не связанных с неселектируемой ДНК, в отборе трансформантов хорошо известно в данной области.
Данное изобретение обеспечивает также способ экспрессии фрагмента ДНК, кодирующего экспандазу в клетке-хозяине, в котором указанный фрагмент ДНК кодирует экспандазу, заставляя эту экспандазу продуцироваться и локализоваться преимущественно в цитозоле. В предпочтительном варианте данного изобретения такой способ предусматривает обеспечение указанного фрагмента ДНК с модификацией в нуклеотидной последовательности, кодирующей карбокси-концевую последовательность аминокислот, предпочтительно последовательность трипептида, такую как SKA, обнаруженную в исследованиях с экспандазой Streptomyces clavuligerus. Такой способ, обеспеченный данным изобретением, используют предпочтительно с таким хозяином, как клетка Penicillium chrysogenum.
Данное изобретение обеспечивает также способ расширения 5-членного кольца соединения бета-лактама с образованием шестичленного соединения цефема, предусматривающий стадии выращивания хозяина, способного продуцировать указанное соединение бета-лактама, в условиях, способствующих этому, и экспрессии гетерологичной экспандазы, полученной рекомбинантными способами и способной расширять 5-членное кольцо указанного соединения бета-лактама, из фрагмента ДНК, кодирующего ее, таким образом, чтобы образовать указанное 6-членное соединение цефема, отличающийся тем, что указанная экспандаза экспрессируется из фрагмента ДНК, который был модифицирован или отобран для кодирования экспандазы, которая продуцируется или по меньшей мере локализуется преимущественно в цитозоле клетки-хозяина.
В предпочтительном варианте данное изобретение обеспечивает способ расширения 5-членного кольца соединения бета-лактама модифицированной экспандазой, в котором эта экспандаза локализуется в цитозоле клетки-хозяина вследствие модификации во фрагменте ДНК, кодирующем ее, посредством которой был модифицирован сигнал таргетирования на микротельце, предпочтительно в котором был модифицирован таргетирующий сигнал, содержащий аминокислотную последовательность трипептида, такой как полученный из экспандазы из Streptomyces clavuligerus.
В экспериментальной части этого описания приведен пример способа, обеспеченного данным изобретением, в котором указанной клеткой-хозяином является Penicillium chrysogenum.
В предпочтительном варианте данного изобретения обеспечен способ, в котором указанное соединение бета-лактама выбрано из группы, состоящей из фенилацетил-6-АПК, феноксиацетил-6-АПК, альфа-аминоадипоил-6-АПК, адипоил-6-АПК, глутарил-6-АПК или из суберил-6-АПК, пимелил-6-АПК, транс-β-гидромуконил-6-АПК или сравнимых соединений. Кроме того, данное изобретение обеспечивает способ получения соединения цефема, предусматривающий стадии расширения 5-членного (пенамного) кольца указанного соединения бета-лактама в процессе или способе в соответствии с данным изобретением и извлечение образованного таким образом цефема. В предпочтительном варианте данное изобретение обеспечивает способ получения соединения цефема в культуре клетки-хозяина для крупномасштабной ферментации, в котором экспандаза, которая преимущественно локализована в цитозоле указанной клетки-хозяина, способствует получению более высоких уровней продуцирования адипоил-7-ADCA, чем в способе получения соединения цефема с использованием сравнимой крупномасштабной ферментационной культуры, использующей экспандазу (дикого типа или модифицированной формы) сравнимого происхождения, которая преимущественно локализована в микротельцах.
В нижеследующих 4 примерах данное изобретение иллюстрируется более подробно.
Описание фигур
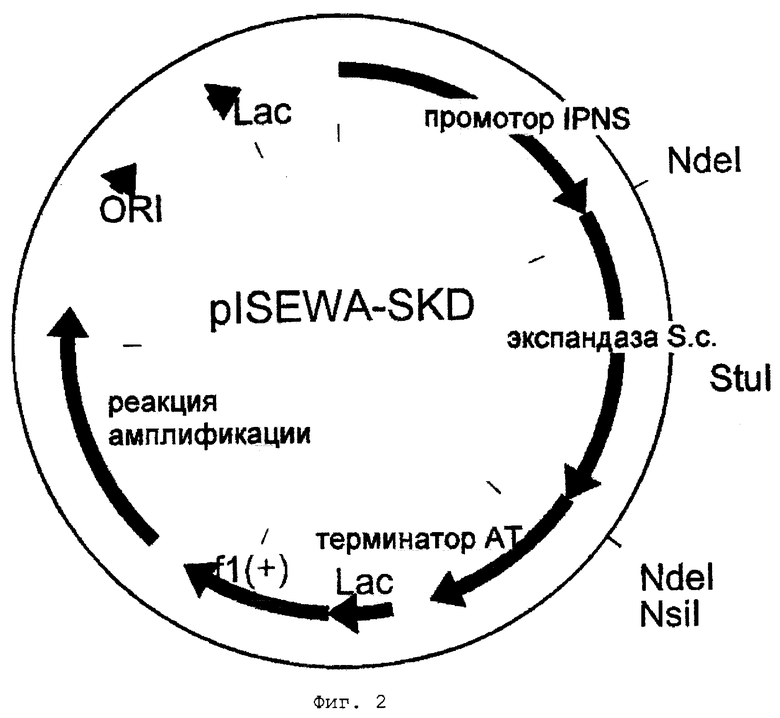
Фиг.1 - схематическое представление плазмиды plSEWA.
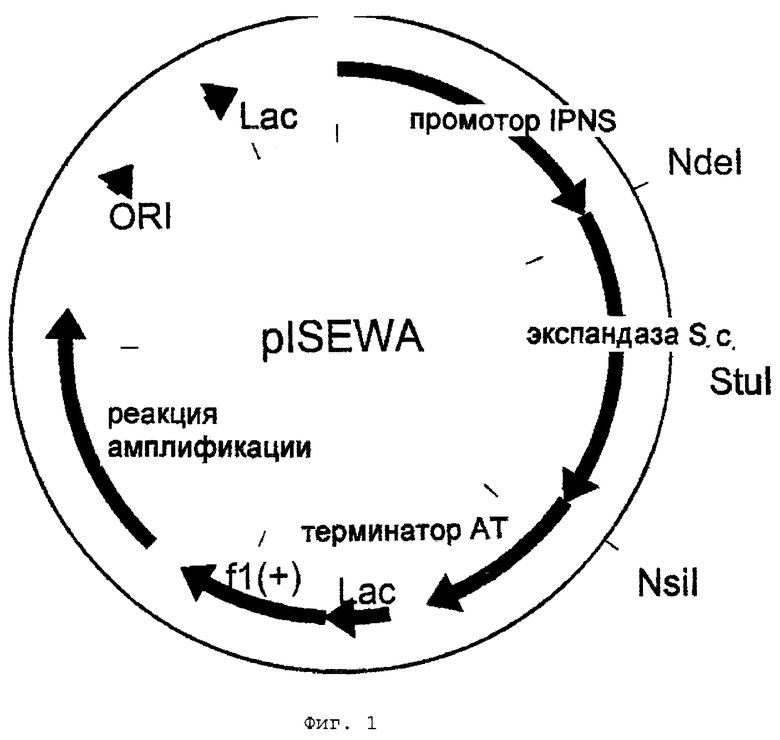
Фиг.2 - схематическое представление плазмиды plSEWA-SKD.
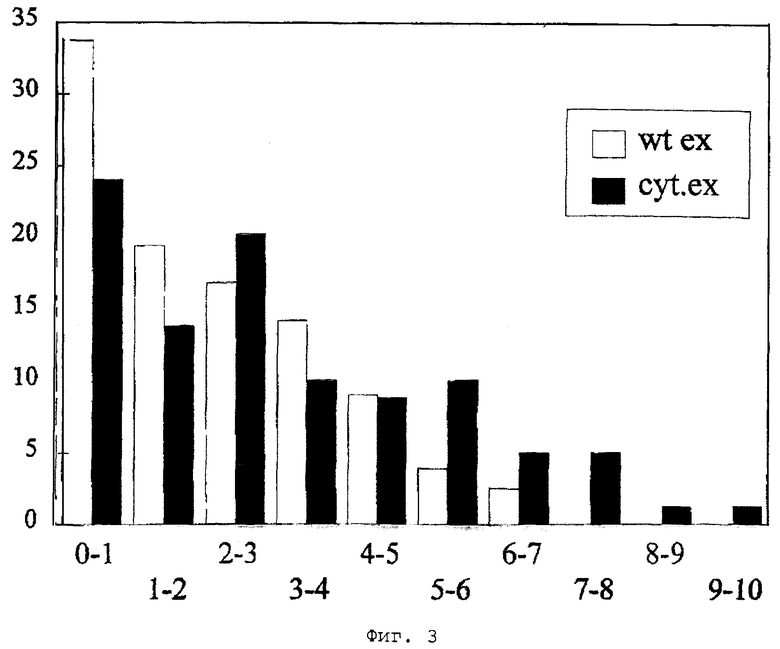
Фиг.3 - продуктивности адипоил-7-ADCA большого числа (около 80) трансформантов, полученных из трансформаций плазмидами plSEWA (белые столбцы) и plSEWA-SKD (черные столбцы). Ось X: класс продуктивности (произвольные единицы); ось Y: % трансформантов на класс.
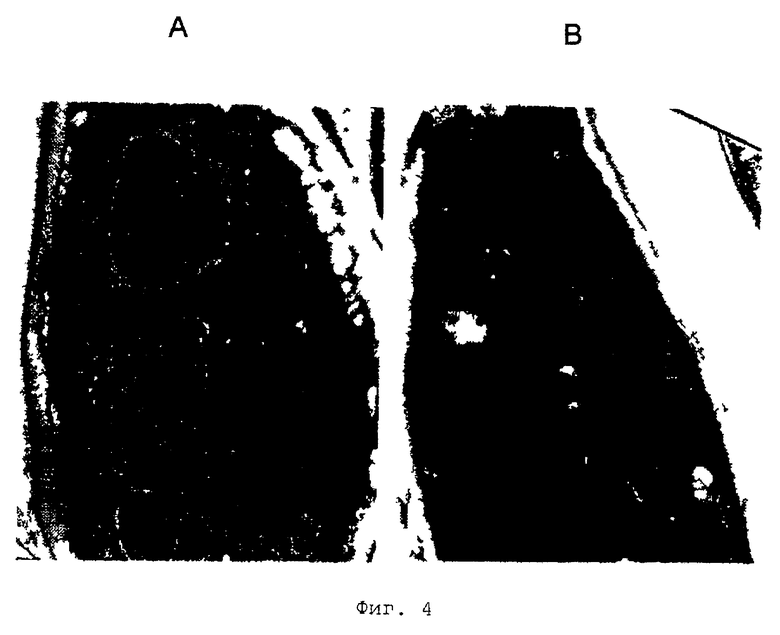
Фиг.4 - электронная микроскопия (ЭМ) трансформанта, экспрессирующего экспандазу дикого типа (панель А), и трансформанта, экспрессирующего экспандазу с нетаргетирующим сигналом (SKA→SKD, панель В). Частицы золота указывают на присутствие экспандазы. N: ядро, М: митохондрия; Р: пероксисома (микротельце).
Экспериментальная часть
Трансформация клеток-хозяев, например Р. chrysogenum или других грибков, может быть достигнута в общем различными способами доставки ДНК, такими как опосредованное ПЭГ-Са поглощение протопластов, электропорация или способы бомбардировки частицами, и отбором трансформантов. См., например, Van den Hondel en Punt, Gene and Transfer and Vector Development for Filamentous Fungi, in: Applied Molecular Genetics of Fungi (Peberdy, Lacen, Ogden, Bennett, eds.), Cambridge University Press (1991). Было описано применение доминантных и не-доминантных селектируемых маркеров (Van den Hondel, выше). Селектируемые маркеры как гомологичного (полученные из Р. chrysogenum), так и гетерологичного (не происходящие из Р. chrysogenum) происхождения были описаны (Gouka et al., J. Biotechnol. 20 (1991) 189-200).
Применение различных маркеров для отбора трансформантов, гомологичных или гетерологичных, в присутствии или в отсутствие векторных последовательностей, физически связанных или не связанных в неселектируемой ДНК, в отборе трансформантов хорошо известно в данной области.
Реакция расширения кольца, опосредованная модифицированным или отобранным экспандазным ферментом, вводится в Р. chrysogenum, например в штамм Wiskonsin 54-1255 (депонированный при АТСС под номером доступа 28089), и осуществляется в нем. Пригодны также другие штаммы Р. chrysogenum, в том числе мутанты штамма Wiskonsin 54-1255, имеющие улучшенный выход бета-лактама.
Кроме того, модифицированный ген cefE помещают под транскрипционный или трансляционный контроль регуляторных элементов грибкового гена. Эти элементы могут быть получены из клонированных грибковых генов, таких как ген IPNS или ген рсbС Р. chrysogenum, ген β-тубулина, ген gpdA Aspergillus nidulans или ген glcA Aspergillus niger.
В соответствии с данным изобретением промежуточный продукт β-лактама адипоил-7-ADCA образуется в трансформантах Р. chrysogenum, экспрессирующих экспандазу, в результате добавления адипиновой кислоты или ее соли или эфира к среде. Подходящими солями являются, например, соли натрия или калия. Адипоил-7-ADCA эффективно извлекается из этой среды простой экстракцией растворителем, например, следующим образом.
Бульон фильтруют и к фильтрату добавляют органический растворитель, не смешивающийся с водой. рН корректируют для экстракции цефалоспорина из водного слоя. Диапазон рН должен быть ниже 4,5; предпочтительно между 4 и 1, более предпочтительно между 2 и 1. Таким путем цефалоспорин отделяют от многих других примесей, присутствующих в ферментационном бульоне. Предпочтительно используют небольшой объем органического растворителя, дающий более концентрированный раствор цефалоспорина, достигая таким образом снижения объемных скоростей потока. Второй возможностью является экстракция всего бульона при рН 4 или ниже. Предпочтительно бульон экстрагируют между рН 4 и 1 органическим растворителем, не смешивающимся с водой.
Может быть использован любой растворитель, который не нарушает молекулу цефалоспорина. Подходящими растворителями являются, например, бутилацетат, этилацетат, метилизобутилкетон, спирты, такие как бутанол, и т.д. Предпочтительно используют 1-бутанол или изобутанол.
После этого цефалоспорин обратно экстрагируют водой при рН между 4 и 10, предпочтительно между 6 и 9. Опять уменьшают конечный объем. Извлечение можно проводить при температурах между 0 и 50°С и предпочтительно при температурах окружающей среды.
Водный раствор полученного таким образом цефалоспорина обрабатывают подходящим ферментом для удаления адипоильной боковой цепи и получения целевой 7-ADCA.
Предпочтительно используют иммобилизованный фермент, чтобы иметь возможность повторного использования фермента. Методология приготовления таких частиц и иммобилизация ферментов была подробно описана в ЕР-А-0222462. рН водного раствора имеет величину, например, рН 4-рН 9, при которой реакция деградации цефалоспорина является минимизированной, а желательное превращение этим ферментом является оптимизированным. Таким образом, этот фермент добавляют к водному раствору цефалоспорина при поддержании рН на подходящем уровне путем добавления, например, неорганического основания, такого как раствор гидроксида калия, или путем применения катионообменной смолы. По завершении реакции иммобилизованный фермент удаляют фильтрованием. Другой возможностью является применение иммобилизованного фермента в фиксированной колонке или колонке с псевдоожиженным слоем или использование фермента в растворе и удаления продуктов фильтрованием через мембрану. Затем реакционную смесь подкисляют в присутствии органического растворителя, не смешивающегося с водой.
Подходящие ферменты получают, например, из микроорганизма Pseudomonas SY77, имеющего мутацию в одном или нескольких положениях 62, 177, 178 и 179. Могут быть также использованы ферменты из других микроорганизмов Pseudomonas, предпочтительно Pseudomonas SE83, необязательно имеющего мутацию в одном или нескольких положениях, соответствующих положениям 62, 177, 178 и 179 в Pseudomonas SY77.
После доведения рН приблизительно до 0,1-1,5 слои разделяют и рН водного слоя доводят до 2-5, более предпочтительно - до 3-4. Затем отфильтровывают кристаллический 7-ADCA.
Деацилирование может проводиться химическим путем, как известно в данной области, например, через образование иминохлоридной боковой цепи, путем добавления пентахлорида фосфора при температуре менее 10°С и затем изобутанола при температурах окружающей среды или более низких температурах.
Следующие примеры предоставляются для иллюстрации, а не для ограничения. Общий подход предусматривает i) идентификацию остатков экспандазы, участвующей в специфичности таргетирования, ii) отбор экспандазных белков или конструирование мутантных экспандазных белков, iii) субклонирование мутантных или отобранных генов экспандазы в экспрессирующих векторах Р. chrysogenum и экспрессию этой экспандазы в Р. chrysogenum, iv) определение продуцирования адипоил-7-ADCA в сравнении с продуцированием a-D-аминоадипоил-7-ADCA и адипоил-6-АПК.
Подобным образом, как было описано для адипоильной боковой цепи, специалист, квалифицированный в данной области, сможет также использовать модифицированный экспандазный фермент в способах, описанных в WO 95/04148 и WO 95/04149, в которых используют 3'-карбоксиметилтиопропионовую кислоту и 3,3'-тиодипропионовую кислоту в качестве боковых цепей, с получением 2-(карбоксиэтилтио)ацетил-7-ADCA и смеси 3-(карбоксиметилтио)пропионил-7-ADCA и 2-(карбоксиэтилтио) ацетил-7-ADCA соответственно.
Пример 1
Сайт-направленный мутагенез гена cefE S. clavuligerus
Способы, используемые в генетическом манипулировании гена экспандазы, хорошо описаны. Источники, которые могут быть использованы в качестве матрицы в ПЦР-реакциях, включают в себя препараты хромосомной ДНК из открыто доступных штаммов Streptomyces clavuligerus (таких как АТСС 27064) или экспрессионные кассеты, такие как pZEx (описанная в РСТ/ЕР97/03879) и plSEWA (описанная в ЕР-02812Р). Кассета экспрессии экспандазы plSEWA (фиг.1) содержит ген экспандазы Streptomyces clavuligerus дикого типа, включающий в себя промотор IPNS и терминатор AT. Для разрушения пероксисомного таргетирования экспандазы последовательность С-конца CefE мутирована из SKA в SKD. Для этой цели с использованием plSEWA в качестве матрицы проводят ПЦР-реакцию с применением праймеров 1 и 2 (см. таблицу). ПЦР проводят с корректирующей полимеразой. После стадии денатурации (2 мин. 98°С) часть гена cefE амплифицируют в 25 циклах (1,15 мин. 94°С, 45 сек. 55°С, 1 мин. 72°С) с последующей стадией удлинения (8 мин. 72°С).
Продукт этой реакции простирается от сайта StuI в cefE (приблизительно 450 т.п.н. от старта трансляции; см. Kovacevic et al.: Cloning, characterisation, and expression in Escherichia coli of the Streptomyces clavuligerus gene encoding desacetoxycephalosporin С synthetase, J. Bacteriol., 171, 754-760, 1989) до сайта NsiI справа (по ходу транскрипции) от стоп-кодона. После подтверждения этой последовательности секвенированием нуклеотидов фрагмент StuI-NsiI используют для замены фрагмента cefE "дикого типа" в plSEWA. Таким образом получают плазмиду plSEWA-SKD (фиг.2).

Пример 2
Трансформация штамма Р. chrysogenum
Способы, используемые в переносе ДНК в протопласты Р. chrysogenum, хорошо известны в данной области и описаны во многих ссылках, в том числе в Finkelstein and Ball (eds.), Biotechnology of filamentous fungi, technology and products, Butterworth-Heinemann (1992); Bennett and Lasure (eds.) More Gene Manipulations in fungi, Academic Press (1991); Turner, in: Puhler (ed), Biotechnology, second completely revised edition, VHC (1992). Опосредованную Са-ПЭГ трансформацию протопластов использовали, как описано в ЕР 635574. Конструкции plSEWA и plSEWA-SKD вводят, следовательно, в Р. chrysogenum Wisconsin 54-1255 (АТСС 28089) посредством котрансформации с GBGLA28 (ЕР 635574), которая позволяет трансформантам Р. chrysogenum расти на селективной среде, содержащей ацетамид в качестве единственного источника азота. Более предпочтительно используют штамм, который имеет более ярко выраженную способность продуцирования пенициллинов. Примерами являются штаммы, такие как CBS 455.95.
Пример 3
Отбор; жидкие культуры
Трансформанты очищали повторяемым культивированием на селективной среде. Одиночные стабильные колонии используют для дальнейшего скрининга на присутствие гена экспандазы при помощи биотеста. Трансформанты выращивают на агаровой среде. Е. coli ESS2231 используют в качестве индикаторной бактерии в верхнем (мягком) слое агара, который также содержит бактопеназу для возможности дискриминировать между продуцированием пенициллина и цефалоспорина, в соответствии со способами, хорошо известными в данной области и описанными, например, в Guttierez et al., Mol. Gen. Genet. 225 (1991), 56-64. Присутствие гена cefE в трансформантах подтверждают при помощи ПЦР, как описано, например, в WO 95/04149, на хромосомной ДНК с использованием праймеров 1 и 2 (таблица). Экспандаза-положительные трансформанты отбирают для дополнительного скрининга на способность продуцировать цефалоспорины, как описано, например, в WO 95/04149.
Трансформанты используют для инокуляции встряхиваемых колб с жидкой средой, как описано, например, в WO 95/04149. Трансформанты инокулируют при 2×106 конидий/мл в среду для засева, состоящую, например, из, г/л: глюкозы 30; (NH4)2SO4 10; KH2PO4 10, раствора микроэлементов I (MgSO4·7H2O 25; FеSO4·7Н2О 10; CuSO4·5H2O 0,5; ZnSO4·7H2O 2; Na2SO4 50; MnSO4·H2O 2; CaCl2·2H2O 5) 10 (мл/л) (рН перед стерилизацией 6,5).
Среду для засева инкубируют в течение 48-72 часов при 25-30°С и затем используют для инокуляции 10-20 объемов производственной питательной среды, содержащей, г/л: лактозу 80; CaSO4 4; мочевину 3; МgSO4·7Н2О 2; КН2РO4 7; NaCl 0,5; (NН4)2SO4 6; FeSO4·7H2O 0,1; адипиновую кислоту 2-5; раствор микроэлементов II (CuSO4·5Н2О 0,5; ZnSO4·7H2O 2; MnSO4·H2O 2; Na2SO4 50) 10 (мл/л) (рН перед стерилизацией 5,5-6,0). Затем инкубацию продолжают еще в течение 96-120 часов. Фильтраты хорошо выращенных культур анализируют при помощи ВЭЖХ и ЯМР на продуцирование адипоил-7-ADCA. Как показывает фигура 3, трансформанты с экспандазой, преимущественно локализованной в цитозоле, имеют более высокую производительность, чем трансформанты с экспандазой, локализованной в микротельцах.
Пример 4
Локализация экспандазы
Для оценки того, приводит ли мутация С-конца экспандазы к блокированию в транспорте экспандазы к пероксисоме, используют электронную микроскопию. Пробы фиксируют при помощи глутарового альдегида в соответствии с процедурой, описанной Waterham et al., J. Cell Biol. 127: 737-749 (1994). Исследования по локализации проводили с использованием иммуноцитохимии с поликлональными антителами против экспандазы. Фиг.4 показывает накопление экспандазы преимущественно в цитозоле или в микротельцах.
Пример 5
Производительность мутантов экспандазы в крупномасштабной ферментации
Для определения производительности трансформантов с мутированной экспандазой в крупномасштабных ферментациях отбирали штаммы из обеих трансформаций для дополнительного анализа. Таким образом, был выбран трансформант мутантной экспандазы, который продуцирует на 10% больше адипоил-7-ADCA, чем трансформант, который был отобран из трансформации геном экспандазы дикого типа. При выращивании штаммов в ферментере на 10 л количество адипоил-7-ADCA в культуре с экспандазой, локализованной преимущественно в цитозоле, является более высоким, чем в ферментации с экспандазой, локализованной преимущественно в микротельцах. Таким образом, экспандаза, преимущественно локализованная в цитозоле, является, очевидно, достаточно стабильной для поддержания стабильной продуктивности и улучшенного выхода в долгосрочной или крупномасштабной ферментации.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЭФФЕКТИВНОГО ПРОИЗВОДСТВА 7-АДЦК ЧЕРЕЗ 2-(КАРБОКСИЭТИЛТИО) АЦЕТИЛ-7-АДЦК И 3-(КАРБОКСИМЕТИЛТИО)ПРОПИОНИЛ-7-АДЦК | 1994 |
|
RU2139349C1 |
| СПОСОБ ПОЛУЧЕНИЯ 7-АМИНОДЕЗАЦЕТОКСИЦЕФАЛОСПОРАНОВОЙ КИСЛОТЫ (7-ADCA) | 1992 |
|
RU2178808C2 |
| СПОСОБ ЭФФЕКТИВНОГО ПРОИЗВОДСТВА 7-АДЦК ЧЕРЕЗ 3-(КАРБОКСИЭТИЛТИО)ПРОПИОНИЛ-7-АДЦК | 1994 |
|
RU2139350C1 |
| СПОСОБ ПОЛУЧЕНИЯ 7-АДЦК ПУТЕМ ВОЗДЕЙСТВИЯ АКТИВНОСТИ ЭКСПАНДАЗЫ НА ПЕНИЦИЛЛИН G | 1996 |
|
RU2192471C2 |
| СПОСОБ ПОЛУЧЕНИЯ 7-АМИНОЦЕФАЛОСПОРАНОВОЙ КИСЛОТЫ, ВЕКТОР ЭКСПРЕССИИ, РЕКОМБИНАНТНЫЙ ШТАММ | 1992 |
|
RU2202616C2 |
| СПОСОБ ПОЛУЧЕНИЯ N-ДЕАЦИЛИРОВАННОГО ЦЕФАЛОСПОРИНА | 1998 |
|
RU2208643C2 |
| СПОСОБ ПОЛУЧЕНИЯ N-ДЕАЦИЛИРОВАННОГО ЦЕФАЛОСПОРИНА | 1998 |
|
RU2208644C2 |
| Способ конструирования рекомбинантной плазмидной ДНК, кодирующей фермент деацетоксицефалоспорин С синтетазу/деацетилцефалоспорин С синтетазу | 1988 |
|
SU1739856A3 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pSVH0106, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ Gl7ACA-АЦИЛАЗЫ В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli BL21(DE3)/pSVH0106-ПРОДУЦЕНТ Gl7ACA-АЦИЛАЗЫ | 2005 |
|
RU2300566C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pZEN16 ДЛЯ ПЕРЕНОСА И ЭКСПРЕССИИ ГЕНОВ В МИЦЕЛИАЛЬНОМ ГРИБЕ ACREMONIUM CHRYSOGENUM | 2009 |
|
RU2434944C2 |
Изобретение относится к биотехнологии, в частности получению цефалоспоринов с использованием генетически модифицированных организмов-хозяев. Для реализации способа используют клетки-хозяина, выбранные из группы Penicillium chrysogenum, Aspergillus nidulans, Asp. niger и Acremonium chrysogenum, имеющей эспандазную активность, локализованную в цитозоле. Фрагмент ДНК, кодирующий эспандазу, получают из Str. clavuligerus и модифицируют так, чтобы обеспечить локализацию фермента в цитозоле клетки-хозяина при экспрессии в ней. Использование способа улучшает получение in vivo ацилированных цефалоспоринов с высоким выходом. 7 з.п. ф-лы, 4 ил., 1 табл.
| Потенциметрический титрограф | 1975 |
|
SU540210A1 |
| Способ выделения фермента, катализирующего превращение креатинина в креатин и фермента, катализирующего превращение креатина в саркозин и мочевину | 1972 |
|
SU532341A3 |
| WO 9504149 А, 1995. | |||

