Область изобретения
Изобретение относится к реконструированным человеческим анти-НМ 1.24 антителам и химерным анти-НМ 1.24 антителам, к кодирующим их генам, к способам получения указанных антител и к использованию указанных антител. Реконструированные человеческие антитела и химерные антитела по настоящему изобретению применимы в качестве терапевтических агентов, например для лечения миеломы.
Предпосылки изобретения
Человеческие В-клетки проходят через различные процессы, которые классифицируются на основании типа экспрессируемых поверхностных антигенов, и в итоге созревают в продуцирующие антитела плазмоциты. На этой конечной стадии своей дифференцировки В-клетки, с одной стороны, приобретают способность продуцировать цитоплазматические иммуноглобулины, и, с другой стороны, исчезают связанные с В-клетками антигены, такие как иммуноглобулины клеточной поверхности, HLA-DR, CD20, Fc-рецепторы, С3-рецепторы системы комплемента и т. п. (Ling, N.R. et al., Leucocyte Typing III (1986), p. 320, Oxford, UK, Oxford).
К настоящему времени имеются сообщения о таких моноклональных антителах, как анти-РСА-1 (Anderson, К.С. et al., J. Immunol. (1983) 130, 1132), анти-РС-1 (Anderson, К. С. et al., J. Immunol. (1983) 132, 3172), анти-ММ4 (Tong, A.W. et al., Blood (1987) 69 238) и т.п., которые распознают антигены на клеточной мембране плазмоцитов. Однако анти-СD38 моноклональное антитело все еще используют для определения плазмоцитов и миеломных клеток (Epstein, J. et al. , N. Engl. J. Med. (1990) 322, 664, Terstappen, L.W.M.M. et al., Blood (1990) 76, 1739, Leo, R. et al., Ann. Hematol. (1992) 64, 132, Shimazaki, С. et al. , Am. J. Hematol. (1992) 39, 159, Hata, H. et al., Blood (1993) 81, 3357, Harada, H. et al., Blood (1993) 81, 2658, Billadeau, D. et al., J. Exp. Med. (1993) 178, 1023).
Однако анти-СD38 моноклональное антитело является антигеном, связанным скорее с активацией Т-клеток, нежели антигеном, связанным с дифференцировкой В-клеток, и экспрессируется на различных клетках помимо В-клеток. Кроме того, хотя CD38 не экспрессируется на некоторых лимфоплазмацитоидах, он сильно экспрессируется на клетках-предшественниках гемопоэза. По этой причине считают, что анти-СD38 моноклональное антитело не пригодно для исследования дифференцировки и созревания человеческих В-клеток или для лечения заболеваний, связанных с плазмоцитами.
Goto, Т. et al. сообщали о мышином анти-НМ 1.24 моноклональном антителе, которое распознает антиген с молекулярной массой от 29 до 33 кДа, который специфически экспрессируется на линиях В-клеток (Blood (1994) 84, 1922-1930). На основании того факта, что антиген, распознаваемый анти-НМ 1.24 моноклональным антителом, считается связанным с окончательной дифференцировкой В-клеток (Goto, Т. et al., Jpn. Clin. Immun. (1992) 16, 688-691), и что введение анти-НМ 1.24 моноклонального антитела мышам с трансплантированной плазмацитомой приводит к специфическому накоплению этого антитела в опухоли (Shuji Ozaki et al., The Program of General Assembly of the 19th Japan Myeloma Study Meeting, general presentation 3), было высказано предположение, что меченое радиоизотопом анти-НМ 1.24 моноклональное антитело, можно использовать для диагностики локализации опухоли, для направленной терапии, такой, как радиоиммунотерапия и т.п.
Кроме того, в указанной выше статье, опубликованной в Blood, описано, что анти-НМ 1.24 моноклональное антитело обладает комплементзависимой цитотоксической активностью по отношению к миеломной клеточной линии человека RPMI8226.
Миелома представляет собой неопластическое заболевание, характеризующееся накоплением моноклональных плазмацитов (миеломных клеток) в костном мозге. Миелома является заболеванием, при котором окончательно дифференцированные В-клетки, которые продуцируют и секретируют иммуноглобулины, или плазмоциты, моноклонально пролиферируют, главным образом, в костном мозге, и соответственно, моноклональные иммуноглобулины или составляющие их компоненты, L-цепи или Н-цепи, определяются в сыворотке (Masaaki Kosaka et al., Nippon Rinsho (1995) 53, 91-99).
Обычно для лечения миеломы использовали химеотерапевтические средства, но не было найдено эффективных терапевтических средств, которые могли бы привести к ремиссии у больных миеломой и к продлению срока жизни больных миеломой. Поэтому давно существует необходимость в создании лекарств, которые оказывали бы терапевтическое воздействие на миелому.
Мышиные моноклональные антитела обладают высокой иммуногенностью (иногда называемой "антигенностью") у людей. Соответственно, медицинская терапевтическая ценность мышиных моноклональных антител для применения у людей ограничена. Так, например, мышиное антитело, введенное человеку, может метаболизироваться как чужеродное вещество, так что срок полураспада мышиного антитела в организме человека относительно мал, и поэтому оно не может полностью продемонстрировать ожидаемый эффект. Кроме того, человеческие антимышиные антитела, которые вырабатываются против введенного мышиного антитела, могут запустить иммунологические реакции, которые неблагоприятны и опасны для пациентов, такие как заболевания сыворотки, другие аллергические реакции или т.п. Поэтому мышиное моноклональное антитело нельзя часто вводить людям.
Для решения этих проблем был разработан способ снижения иммуногенности антител, полученных не от человека, таких как полученные из мышей моноклональные антитела. Одним из таких примеров является способ получения химерного антитела, в котором вариабельный участок (V-участок) антитела получен из исходной мыши, а его константный участок (С-участок) получен из подходящего человеческого антитела.
Так как полученное таким образом химерное антитело содержит вариабельный участок исходного мышиного антитела в интактной форме, ожидается, что оно будет связываться с антигеном со специфичностью, идентичной специфичности исходного мышиного антитела. Кроме того, в химерном антителе количество аминокислотных последовательностей, полученных из организма, отличного от человеческого, существенно снижено, и поэтому ожидается, что такое антитело будет обладать пониженной иммуногенностью по сравнению с исходным мышиным антителом. Химерное антитело может связываться с антигеном таким же образом, как и исходное мышиное моноклональное антитело, и может включать иммунологические реакции против вариабельного участка мышиного антитела, несмотря на то, что его иммуногенность понижена (LoBuglio A.F. et al., Proc. Natl. Acad. Sci. USA, 86, 4220-4224, 1989).
Второй способ снижения иммуногенности мышиного антитела, хотя и гораздо более сложный, позволяет еще более снизить потенциальную иммуногенность мышиного антитела. В этом способе лишь участок, определяющий комплементарность (CDR) вариабельного участка мышиного антитела трансплантируют в вариабельный участок человеческого антитела, получая вариабельный участок "реконструированного" человеческого антитела.
Однако для того чтобы сделать структуру вариабельного участка реконструированного человеческого антитела как можно более близкой к структуре исходного мышиного антитела, необходимо, чтобы часть аминокислотной последовательности каркасного участка (FR), который поддерживает CDR, можно было бы трансплантировать из вариабельного участка мышиного антитела в вариабельный участок человеческого антитела. Затем этот V-участок гуманизированного реконструированного человеческого антитела связывают с константным участком человеческого антитела. Часть, которая получена из отличной от человеческой аминокислотной последовательности в окончательно реконструированном гуманизированном антителе является CDR, и составляет лишь часть FR. CDR состоит из гипервариабельной аминокислотной последовательности, которая не содержит видоспецифических последовательностей. Таким образом, гуманизированное антитело, содержащее мышиный CDR, не должно быть более иммуногенным, нежели природное человеческое антитело, содержащее CDR человеческого антитела.
Относительно гуманизированного антитела см. Riechmann, L. et al., Nature, 332, 323-327, 1988; Verhoeye, M. et al., Science, 239, 1534-1536, 1988; Kettleborough, С.A. et al., Protein Engng., 4, 773-783, 1991; Meada, H. et al., Human Antibodies and Hybridoma, 2, 124-134, 1991; Groman, S.D. et al. , Proc. Natl. Acad. Sci. USA, 88, 4181-4185, 1991; Tempest, P.R. et al., Bio/Technology, 9, 266-271, 1991; Co, M.S. et al., Proc. Natl. Acad. Sci. USA, 88, 2869-2873, 1991; Carter, P. et al., Proc. Natl. Acad. Sci. USA, 89, 4285-4289, 1992; Co, M.S. et al., J. Immunol., 148, 1149-1154, 1992; и Sato, К. et al. Cancer Res., 53, 851-856, 1993.
Queen et al. (публикация международной заявки WO 90-07861) раскрывает способ получения гуманизированного антитела из антитела против рецептора IL-2 Anti-Tac. Однако трудно полностью гуманизировать все антитела, даже следуя способу, раскрытому в WO 90-07861. Так, WO 90-07861 не раскрывает общий способ гуманизации антител, а просто описывает способ гуманизации антитела Anti-Tac, которое является одним из антител против рецептора IL-2. Кроме того, даже если полностью следовать способу WO 90-07862, оказывается затруднительным получить гуманизированное антитело, которое обладает активностью, полностью идентичной активности исходного мышиного антитела.
Вообще, аминокислотные последовательности CDR/FR отдельных антител отличаются. Соответственно, определение аминокислотного остатка, который следует заменить для создания гуманизированного антитела, и выбор аминокислотного остатка, которым следует заменить указанный аминокислотный остаток, меняются для каждого отдельного антитела. Поэтому способ получения гуманизированных антител, представленный ранее в WO 90-07861, нельзя применить для гуманизации всех антител.
Queen et al. Proc. Natl. Acad. Sci. USA, (1989) 86, 10029-10033 сделал то же самое открытие, что изложено в WO 90-07861. В этой ссылке указано, что только одна треть активности исходного мышиного антитела достигается для гуманизированного антитела, полученного по способу, изложенному в WO 90-07861. Другими словами это показывает, что способ WO 90-07861 сам по себе не может привести к продуцированию полностью гуманизированного антитела, которое обладало бы активностью, равной активности исходного мышиного антитела.
Со et al.. Cancer Research (1996) 56, 1118-1125 опубликовано группой вышеуказанных Queen et al. В этой ссылке указано, что гуманизированное антитело, обладающее активностью, равной активности исходного мышиного антитела, нельзя сконструировать даже способом получения гуманизированных антител, как это изложено в WO 90-07861. Этот факт не только свидетельствует, что способ WO 90-07861 сам по себе не может привести к получению полностью гуманизированного антитела, обладающего активностью, равной активности исходного мышиного антитела, но что этот способ конструирования гуманизированного антитела, как это представлено далее в WO 90-07861, нельзя применить к гуманизации всех антител.
Ohtomo et al., Molecular Immunology (1995) 32, 407-416 описывает гуманизацию мышиного ONS-M21 антитела. Из этой ссылки следует, что аминокислотный остаток, который предлагался для гуманизации анти-Тас антитела в WO 90-07861, не имеет отношения к активности, и способ в том виде, как он изложен, в WO 90-07861, не применим.
Kettleborough et al. Protein Eng. (1991) 4, 773-783 указывает, что было сконструировано несколько гуманизированных антител из мышиного антитела в результате замены аминокислотных остатков. Однако при этом требуется замещение большего числа остатков, нежели предлагается в способе гуманизации анти-Тас антитела, как указано в WO 90-07861.
В приведенных выше ссылках указано, что способ получения гуманизированных антител, как предложено далее в WO 90-07861, представляет собой методику, которая применима только к описанному там анти-Тас антителу, и что даже использование указанной методики не приведет к достижению активности, равной активности исходного мышиного антитела.
Исходные мышиные антитела, описываемые в этих ссылках, содержат аминокислотные последовательности, отличающиеся от последовательности анти-Тас антитела, описанного в WO 90-07861. Соответственно, этот способ конструирования гуманизированного антитела, который можно было применить к анти-Тас антителу, нельзя применить к другим антителам. Аналогично, так как мышиное анти-НМ 1.24 моноклональное антитело по настоящему изобретению имеет аминокислотную последовательность, отличающуюся от последовательности анти-Тас антитела, нельзя применить способ конструирования гуманизированного антитела для анти-Тас антитела. Кроме того, успешно сконструированное гуманизированное антитело по настоящему изобретению имеет аминокислотную последовательность, которая отличается от последовательности гуманизированного анти-Тас антитела, описанного в WO 90-07861. Этот факт также указывает на то, что один и тот же способ нельзя использовать для гуманизации антител с различными CDR-FR последовательностями.
Таким образом, даже если исходное гуманизированное мышиное антитело известно, идентичность CDR-FR последовательности гуманизированного антитела, обладающего активностью, подтверждается только методом проб и ошибок. В WO 90-07861 не упоминается о FR последовательности, которая объединена в гуманизированном антителе, сконструированном в настоящем изобретении, а также о том факте, что активное гуманизированное антитело можно получить из комбинации с FR, гораздо меньшей последовательности CDR.
Как было указано выше, гуманизированные антитела, как ожидают, должны быть полезны для терапевтических целей, но гуманизированные анти-НМ 1.24 моноклональные антитела не известны и даже не предложены. Кроме того, не существует доступного стандартного способа, который можно было бы применить к любому из антител для получения гуманизированного антитела, и необходимы различные ухищрения для конструирования и гуманизации антитела, которое демонстрировало бы достаточную активность связывания, активность ингибирования связывания и нейтрализующую активность (например, Sato, К. et al., Cancer Res., 53, 851-856, 1993).
Описание изобретения
В настоящем изобретении предложены реконструированные антитела анти-НМ 1.24 антитела. Далее в настоящем изобретении предложены человеческие/мышиные химерные антитела, которые можно использовать в процессе конструирования указанных реконструированных антител. В настоящем изобретении предложены фрагменты реконструированных антител. Далее в настоящем изобретении предложена экспрессионная система для получения химерных антител, реконструированных антител и их фрагментов. В настоящем изобретении предложены способы получения химерных антител из анти-НМ 1.24 антитела и его фрагментов, а также способы получения реконструированных антител из анти-НМ 1.24 антитела и его фрагментов.
Более конкретно, в настоящем изобретении предложены химерные антитела и реконструированные антитела, которые специфически распознают полипептид с аминокислотной последовательностью, представленной далее в Последовательности ИД 103 кДНК, которая кодирует указанный полипептид, была встроена между XbaI сайтами расщепления вектора pUC19, и таким образом, была получена как плазмида pRS38-pUC19. Escherichia coli, которая содержит эту плазмиду pRS38-pUC19, была интернационально депонирована 5 октября 1993 года National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, MITI (Higashi 1-Chome 1-3, Tsukuba city, Ibalaki prefecture, Japan) как Escherichia coli DH5α (pRS38-pUC19) под регистрационным номером FERM BP-4434 в соответствии с Будапештским соглашением (см. японскую патентную публикацию без экспертизы (Kokai) 7-196694).
В качестве одного из осуществлений таких химерных антител или реконструированных антител, упоминается химерное анти-НМ 1.24 антитело или реконструированное человеческое анти-НМ 1.24 антитело. Подробное описание химерного анти-НМ 1.24 антитела или реконструированного человеческого анти-НМ 1.24 антитела будет приведено далее.
Так, в настоящем изобретении предложены также химерные L-цепи, включающие константный участок (С-участок) человеческой легкой (L-) цепи и вариабельный (V-) участок L-цепи анти-НМ 1.24 антитела, и химерную Н-цепь, включающую константный участок человеческой тяжелой (Н-) цепи и V-участок тяжелой (Н-) цепи анти-НМ 1.24 антитела.
Далее в настоящем изобретении предложены химерные антитела, включающие:
(1) L-цепь, включающую С-участок человеческой L-цепи и V-участок L-цепи анти-НМ 1.24 антитела; и
(2) Н-цепь, включающую С-участок человеческой Н-цепи и V-участок Н-цепи анти-НМ 1.24 антитела.
Далее в настоящем изобретении предложен V-участок реконструированной человеческой L-цепи анти-НМ 1.24 антитела, включающий:
(1) каркасный участок (FR) V-участка человеческой L-цепи, и
(2) CDR V-участка L-цепи анти-НМ 1.24 антитела; и V-участок реконструированной человеческой Н-цепи анти-НМ 1.24 антитела, включающий:
(1) FR V-участка человеческой Н-цепи, и
(2) CDR V-участка Н-цепи анти-НМ 1.24 антитела.
Далее в настоящем изобретении предложена реконструированная человеческая L-цепь анти-НМ 1.24 антитела, включающая:
(1) С-участок человеческой L-цепи, и
(2) V-участок L-цепи, включающий FR человеческой L-цепи, и CDR L-цепи анти-НМ 1.24 антитела; и
реконструированная человеческая Н-цепь анти-НМ 1.24 антитела, включающая:
(1) С-участок человеческой Н-цепи, и
(2) V-участок Н-цепи, включающий FR человеческой Н-цепи, и CDR Н-цепи анти-НМ 1.24 антитела.
Далее в настоящем изобретении предложено реконструированное человеческое антитело анти-НМ 1.24 антитела, включающее:
(A) L-цепь, включающую:
(1) С-участок человеческой L-цепи; и
(2) V-участок L-цепи, включающий FR человеческой L-цепи и CDR L-цепи анти-НМ 1.24 антитела; и
(B) Н-цепь, включающую:
(1) С-участок человеческой Н-цепи, и
(2) V-участок Н-цепи, включающий FR человеческой Н-цепи и CDR Н-цепи анти-НМ 1.24 антитела.
Настоящее изобретение предлагает далее ДНК, кодирующую V-участок L-цепи анти-НМ 1.24 антитела, и ДНК, кодирующую V-участок H-цепи анти-HM 1.24 антитела.
Далее настоящее изобретение предлагает ДНК, кодирующую химерную L-цепь, включающую:
(1) С-участок человеческой L-цепи; и
(2) V-участок L-цепи анти-НМ 1.24 антитела, и ДНК, кодирующую химерную Н-цепь, включающую:
(1) С-участок человеческой Н-цепи; и
(2) V-участок Н-цепи анти-НМ 1.24 антитела.
Настоящее изобретение предлагает далее ДНК, кодирующую V-участок реконструированной человеческой L-цепи анти-НМ 1.24 антитела, включающий:
(1) FR V-участка человеческой L-цепи; и
(2) CDR V-участка L-цепи анти-НМ 1.24 антитела; и ДНК, кодирующую V-участок реконструированной человеческой Н-цепи анти-НМ 1.24 антитела, включающий:
(1) FR V-участка человеческой Н-цепи; и
(2) CDR V-участка Н-цепи анти-НМ 1.24 антитела.
В настоящем изобретении предложена ДНК, кодирующая реконструированную человеческую L-цепь анти-НМ 1.24 антитела, включающую:
(1) С-участок человеческой L-цепи; и
(2) V-участок L-цепи, включающий FR человеческой L-цепи и CDR L-цепи анти-НМ 1.24 антитела; и
ДНК, кодирующая реконструированную человеческую Н-цепь анти-НМ 1.24 антитела, включающую:
(1) С-участок человеческой Н-цепи; и
(2) V-участок Н-цепи, включающий FR человеческой Н-цепи и CDR Н-цепи анти-НМ 1.24 антитела.
Далее в настоящем изобретении предложен вектор, включающий любую из указанных выше ДНК.
В настоящем изобретении предложена клетка-хозяин, трансформированная вышеуказанным вектором.
В настоящем изобретении предложены способы получения химерного антитела анти-НМ 1.24 антитела, включающие стадии культивирования клетки-хозяина, которая была котрансформирована экспрессирующим вектором, включающим ДНК, кодирующую указанную химерную L-цепь, и с вектором экспрессии, включающим ДНК, кодирующую указанную Н-цепь, и выделения желаемого антитела.
Далее в настоящем изобретении предложены способы получения реконструированного человеческого антитела анти-НМ 1.24 антитела, включающие стадии культивирования клетки-хозяина, которая была котрансформирована экспрессирующим вектором, включающим ДНК, кодирующую указанную реконструированную человеческую L-цепь, и с вектором экспрессии, включающим ДНК, кодирующую указанную реконструированную человеческую Н-цепь, и выделения желаемого антитела.
Далее в настоящем изобретении предложены фармацевтические композиции, особенно терапевтические агенты для лечения миеломы, включающие указанное химерное антитело или реконструированное человеческое антитело.
Далее в настоящем изобретении предложены фармацевтические композиции, которые содержат в качестве активного ингредиента химерное антитело, специфически распознающее полипептид с аминокислотной последовательностью, представленной в Последовательности ИД 103, и фармацевтические композиции, которые содержат в качестве активного ингредиента реконструированное человеческое антитело, специфически распознающее полипептид с аминокислотной последовательностью, представленной в Последовательности ИД 103. В качестве фармацевтической композиции конкретно предложен терапевтический агент для лечения миеломы.
Краткое объяснение рисунков
Фиг. 1 представляет график, демонстрирующий, что в FCM анализе с использованием клеточной линии миеломы человека КРММ2, интенсивность флуоресценции химерного анти-НМ 1.24 антитела сдвигается аналогично сдвигу для мышиного анти-НМ 1.24 антитела, при сравнении с контрольным антителом.
Фиг. 2 представляет график, демонстрирующий, что в клеточном ELISA с использованием WISH-клеток, химерное анти-НМ 1.24 антитело по аналогии с мышиным анти-НМ 1.24 антителом дозозависимо ингибирует связывание биотинилированного мышиного анти-НМ 1.24 антитела с WISH-клетками.
Фиг.3 представляет график, демонстрирующий, что контрольный человеческий IgG1 или мышиное анти-НМ 1.24 антитело не обладают цитотоксичностью, тогда как химерное анти-НМ 1.24 антитело демонстрирует повышенную цитотоксичность в отношении RPMI 8226 клеток при возрастании отношения Е/Т.
Фиг. 4 представляет схему способа конструирования L-цепи реконструированного человеческого анти-НМ 1.24 антитела в результате трансплантации CDR методом ПЦР.
Фиг. 5 представляет схему способа сборки олигонуклеотидов RVH1, RVH2, RVH3 и RVH4 методом ПЦР при создании Н-цепи реконструированного человеческого анти-НМ 1.24 антитела.
Фиг. 6 представляет схему способа конструирования V-участка Н-цепи человеческого/мышиного гибридного анти-НМ 1.24 антитела методом ПЦР.
Фиг. 7 представляет схему способа конструирования V-участка Н-цепи мышиного/человеческого гибридного анти-НМ 1.24 антитела методом ПЦР.
Фиг.8 представляет график, демонстрирующий, что вариант а L-цепи реконструированного человеческого анти-НМ 1.24 антитела обладает антигенсвязывающей активностью, равной активности химерного анти-НМ 1.24 антитела. -1 и -2 указывают на то, что это различные группы.
Фиг.9 представляет график, демонстрирующий антигенсвязывающую активность реконструированного человеческого анти-НМ 1.24 антитела, в котором вариант а L-цепи и вариант а, b, f или h Н-цепи были объединены, и химерного анти-НМ 1.24 антитела.
Фиг. 10 представляет график, демонстрирующий активность связывания реконструированного человеческого анти-НМ 1.24 антитела, в котором вариант b L-цепи и вариант а, b, f или h Н-цепи были объединены, и химерного анти-НМ 1.24 антитела.
Фиг. 11 представляет график, демонстрирующий активность ингибирования связывания реконструированного человеческого анти-НМ 1.24 антитела, в котором вариант а L-цепи и вариант a, b, f или h Н-цепи были объединены, и химерного анти-НМ 1.24 антитела.
Фиг. 12 представляет график, демонстрирующий активность ингибирования связывания реконструированного человеческого анти-НМ 1.24 антитела, в котором вариант b L-цепи и вариант а, b, f или h Н-цепи были объединены, и химерного анти-НМ 1.24 антитела.
Фиг. 13 представляет график, демонстрирующий активность связывания антигена вариантов а, b, с и d Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, и химерного анти-НМ 1.24 антитела.
Фиг. 14 представляет график, демонстрирующий активность связывания антигена вариантов а и е Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, и химерого анти-НМ 1.24 антитела. -1 и -2 указывают на то, что они относятся к разным группам.
Фиг. 15 представляет график, демонстрирующий активность ингибирования связывания вариантов а, с, р и r Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, и химерного анти-НМ 1.24 антитела.
Фиг. 16 представляет график, демонстрирующий активность связывания антигена человеческого/мышиного гибридного анти-НМ 1.24 антитела, мышиного/человеческого гибридного анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 17 представляет график, демонстрирующий активность связывания антигена вариантов а, b, с и f Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 18 представляет график, демонстрирующий активность связывания антигена вариантов a и g Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 19 представляет график, демонстрирующий активность ингибирования связывания вариантов а и g Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 20 представляет график, демонстрирующий активность связывания антигена вариантов h и i Н-цепи реконструированного Н человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 21 представляет график, демонстрирующий активность связывания антигена вариантов f, h и j Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 22 представляет график, демонстрирующий активность ингибирования связывания вариантов h и i Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 23 представляет график, демонстрирующий активность ингибирования связывания вариантов f, h и j Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 24 представляет график, демонстрирующий активность связывания антигена вариантов h, k, l, m, n и o Н-цепи реконструированного человеческого lнти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 25 представляет график, демонстрирующий активность связывания антигена вариантов а, h, р и q Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 26 представляет график, демонстрирующий активность ингибирования связывания с WISH-клетками вариантов h, k, 1, m, n и о Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 27 представляет график, демонстрирующий активность ингибирования связывания вариантов а, h, р и q Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 28 представляет график, демонстрирующий активность связывания антигена вариантов а, с, p и r Н-цепи реконструированного человеческого анти-НМ 1.24 антитела и химерного анти-НМ 1.24 антитела.
Фиг. 29 представляет график, демонстрирующий, что вариант s реконструированного человеческого анти-НМ 1.24 антитела обладает антигенсвязывающей активностью, равной активности варианта r реконструированного человеческого анти-НМ 1.24 антитела.
Фиг. 30 представляет график, демонстрирующий, что вариант s реконструированного человеческого анти-НМ 1.24 антитела обладает активностью ингибирования связывания, равной активности варианта r реконструированного человеческого анти-НМ 1.24 антитела.
Фиг. 31 представляет график, демонстрирующий, что очищенное реконструированное человеческое анти-НМ 1.24 антитело обладает активностью связывания антигена, равной активности химерного анти-НМ 1.24 антитела.
Фиг. 32 представляет график, демонстрирующий, что очищенное реконструированное анти-НМ 1.24 антитело обладает активностью ингибирования связывания, равной активности химерного анти-НМ 1.24 антитела.
Фиг.33 представляет график, демонстрирующий, что введение химерного анти-НМ 1.24 антитела вызывает продление срока жизни по сравнению с введением контрольного человеческого IgG1 у мышей с трансплантированными клетками миеломы человека.
Фиг.34 представляет график, демонстрирующий, что если клетки, полученные из периферической крови здорового человека, используют в качестве эффекторных клеток, контрольный человеческий IgG1 не демонстрирует цитотоксичности по отношению к клеткам КРММ2, и что мышиное анти-НМ 1.24 антитело также обладает слабой цитотоксичностью, тогда как реконструированное человеческое анти-НМ 1.24 антитело демонстрирует высокую цитотоксичность в отношении клеток КРММ2.
Фиг.35 представляет график, демонстрирующий, что если используют клетки, полученные из периферической крови здорового человека в качестве эффекторных клеток, контрольный человеческий IgG1 не демонстрирует цитотоксичности по отношению к клеткам ARH-77, и что мышиное анти-НМ 1.24 антитело также обладает слабой цитотоксичностью, тогда как реконструированное человеческое анти-НМ 1.24 антитело демонстрирует высокую цитотоксичность в отношении клеток ARH-77.
Фиг. 36 представляет график, демонстрирующий, что если в качестве эффекторных клеток используют клетки, полученные из костного мозга мышей SCID, контрольный человеческий IgG1 не демонстрирует цитотоксичности по отношению к клеткам КРММ2, тогда как реконструированное человеческое анти-НМ 1.24 антитело демонстрирует повышающуюся цитотоксичность в отношении клеток КРММ2 с возрастанием концентрации антител.
Фиг. 37 представляет график, демонстрирующий, что у мышей с трансплантированными клетками миеломы человека уровень человеческого IgG в сыворотке повышается после введения контрольного человеческого IgG1 по сравнению с уровнем до его введения, тогда как введение реконструированного человеческого анти-НМ 1.24 антитела ингибирует повышение уровня человеческого IgG в сыворотке.
Фиг. 38 представляет график, демонстрирующий, что у мышей с трансплантированными миеломными клетками человека введение реконструированного человеческого анти-НМ 1.24 антитела вызывает пролонгирование срока жизни по сравнению со сроком после введения контрольного IgG1 человека.
Фиг. 39 представляет график, демонстрирующий, что у мышей с трансплантированными миеломными клетками человека уровень человеческого IgG в сыворотке повышается после введения мелфалана и контрольного человеческого IgG1 по сравнению с уровнем до введения, тогда как введение реконструированного человеческого анти-НМ 1.24 антитела ингибирует повышение уровня человеческого IgG в сыворотке.
Фиг. 40 представляет график, демонстрирующий, что у мышей с трансплантированными миеломными клетками человека введение реконструированного человеческого анти-НМ 1.24 антитела вызывает пролонгирование периода выживания по сравнению с периодом после введения мелфалана или контрольного человеческого IgG1.
Способ осуществления изобретения
1. Конструирование химерного антитела
(1) Клонирование ДНК, кодирующей V-участок мышиного анти-НМ 1.24 моноклонального антитела
Получение мРНК
Для клонирования ДНК, кодирующей V-участок мышиного анти-НМ 1.24 моноклонального антитела, получают полную РНК из выделенной гибридомы, используя известный способ, такой как способ с использованием гуанидин-ультрацентрифугирования (Chirgwin, J. M. et al., Biochemistry (1979), 18, 5294-5299), способ AGPC (Chomczynski, P. et al, (1987), 162, 156-159), и т.д., и мРНК получают, используя спай-колонку с олиго(дТ)-целлюлозой, дополненной с набором для очистки мРНК (mRNA Purification Kit, Pharmacia) и т.д. Кроме того, используя QuickPrep mRNA Purification Kit (Pharmacia), мРНК можно получить без стадии выделения полной РНК.
Получение и амплификация кДНК
Из мРНК, полученной на приведенной выше стадии получения мРНК, каждую кДНК для V-участков L-цепи и Н-цепи синтезируют, используя обратную транскриптазу кДНК V-участка L-цепи синтезируют, используя набор для синтеза первой цепочки кДНК AMV обратной транскриптазы (AMV Reverse Transcriptase First-Strand cDNA Synthesis Kit). Для амплификации синтезированной кДНК используют соответствующий праймер, который гибридизуется с лидерной последовательностью и С-участком гена антитела (например, праймер MKV, нуклеотидная последовательность которого представлена Последовательностями ИД 29-39, и праймер МКС нуклеотидная последовательность которого представлена Последовательностью ИД 40).
Синтез и амплификацию кДНК V-участка Н-цепи можно осуществить с помощью ПЦР (полимеразной цепной реакции) методом 5'-RACE (Frohman, M.A. et al., Proc. Natl. Acad. Sci. USA, 85, 8998-9002, Belyavsky, A. et al., Nucleic Acids Res. 17, 2919-2932, 1989), используя набор 5'-Ampli FINDER RACE (CLONTECH). С 5'-концом синтезированной ранее кДНК лигируютAmpli FINDER Anchor, а в качестве праймера для амплификации V-участка Н-цепи, можно использовать праймер, который специфически гибридизуется с праймером Anchor (Последовательность ИД 77) и константным участком (Cγ-участок) мышиной Н-цепи (например, праймер МНС2а, нуклеотидная последовательность которого представлена последовательностью ИД 42).
Выделение ДНК и определение ее нуклеотидной последовательности
Продукт ПЦР подвергают электрофорезу в агарозном геле, используя известные методики, для вырезания нужного ДНК фрагмента, и ДНК выделяют из него и очищают, а затем лигируют с векторной ДНК.
ДНК можно очистить, используя коммерческий набор (например, GENECLEAN II; BI0101). Известную векторную ДНК (например, pUC19, Bluescript и т.д.) можно использовать для сохранения фрагментов ДНК.
Вышеуказанную ДНК и вышеуказанный вектор лигируют, используя известный набор для лигирования (производитель Таkara Shuzo) для получения рекомбинантного вектора. Затем полученный рекомбинантный вектор вводят в Escherichia coli JM109, после чего устойчивые к ампициллину колонии отбирают, и векторную ДНК получают на основании известного метода (J. Sambrook, et al., "Molecular Cloning", Cold Spring Harbor Laboratory Press, 1989).
После расщепления вышеуказанной векторной ДНК ферментами рестрикции, известным способом определяют нуклеотидную последовательность желаемой ДНК (например, "дидезокси" методом) (J. Sambrook, et al., "Molecular Cloning", Cold Spring Harbor Laboratory Press, 1989). В соответствии с настоящим изобретением можно использовать автоматическую систему секвенирования (DNA Sequencer 373А; изготовитель ABI Co. Ltd.).
Участок, определяющий комплементарность
V-участок Н-цепи и V-участок L-цепи образуют антигенсвязывающий сайт, причем их полные структуры обладают аналогичными свойствами. Так, каждый из четырех каркасных участков (FR) был лигирован с тремя гипервариабельными участками, т. е. участками, определяющими комплементарность (CDR). Аминокислотные последовательности FRs были относительно устойчиво консервативны, тогда как среди аминокислотных последовательностей CDR участков вариации встречаются чрезвычайно часто (Kabat, E.A. et al., "Sequence of Proteins of Immunological Interest", US Dept. Health and Human Services, 1983).
Многие части вышеуказанных четырех FRs принимают структуру β-листа, что приводит к образованию трех CDR-петель. Иногда CDR могут составлять часть структуры β-листа. Три CDR стерически сохраняются в тесной близости друг к другу, и образуют антигенсвязывающий сайт с тремя CDR участками спаривания.
На основании этих фактов аминокислотная последовательность вариабельного участка мышиного анти-НМ 1.24 антитела подогнана к данным для аминокислотных последовательностей антител, полученных Kabat et al. "Sequence of Proteins of Immunological Interest", US Dept. Health and Human Services, 1983), для исследования гомологии и, тем самым, для определения CDR участков.
(2) Конструирование векторов экспрессии для химерного антитела
После того, как клонированы фрагмент ДНК, кодирующий V-участки мышиной L-цепи и Н-цепи мышиного моноклонального антитела, можно получить химерное анти-НМ 1.24 антитело за счет связывания этих мышиных V-участков с ДНК, кодирующей константный участок человеческого антитела, и последующей их экспрессии.
Основной способ конструирования химерного антитела включает связывание мышиной лидерной последовательности и последовательности V-участка, присутствующей в клонированной кДНК, с последовательностью, кодирующей С-участок человеческого антитела, уже присутствующего в экспрессирующем векторе для клеток млекопитающих. В другом варианте он включает связывание мышиной лидерной последовательности с последовательностью V-участка, уже присутствующей в клонированной кДНК, с последовательностью, кодирующей С-участок человеческого антитела, что потом связывают с вектором экспрессии для клеток млекопитающих.
С-участок человеческого антитела может быть С-участком любой Н-цепи и С-участком любой L-цепи. Так, можно, например, упомянуть Cγ1, Cγ2, Cγ3, или Cγ4 человеческой Н-цепи, или Cλ или Cκ L-цепи.
Для получения химерного антитела конструируют два типа экспрессирующих векторов: они представляют собой экспрессирующий вектор, включающий ДНК, кодирующую V-участок мышиной L-цепи и С-участок человеческой L-цепи под контролем участка, регулирующего экспрессию, такого как система энхансер/промотор, и экспрессирующий вектор, включающий ДНК, кодирующую V-участок мышиной Н-цепи, и С-участок человеческой Н-цепи под контролем участка, регулирующего экспрессию, такого, как система энхансер/промотор. Затем, используя эти экспрессирующие векторы, подвергают котрансформации такие клетки хозяина, как клетки млекопитающих, и эти трансформированные клетки культивируют in vitro или in vivo, получая химерное антитело (например, WO 91-16928).
В другом варианте, ДНК, кодирующую мышиную лидерную последовательность и V-участок L-цепи и С-участок человеческой L-цепи, и ДНК, кодирующую мышиную лидерную последовательность, и V-участок Н-цепи и С-участок человеческой Н-цепи, представленные в клонированной кДНК, вводят в один экспрессирующий вектор (см. публикацию международной заявки WO 94-11523), и клетку-хозяин трансформируют, используя указанный вектор. Затем эту трансформированную клетку-хозяин культивируют in vitro или in vivo, получая целевое химерное антитело.
1) Конструирование химерной Н-цепи
Вектор экспрессии для Н-цепи химерного антитела можно получить в результате введения кДНК, кодирующей V-участок мышиной Н-цепи, в соответствующий экспрессирующий вектор экспрессии, содержащий геномную ДНК или кДНК, кодирующую С-участок Н-цепи человеческого антитела. В качестве С-участков Н-цепи можно указать, например, Cγ1, Cγ2, Cγ3, или Cγ4.
Конструирование экспрессирующего вектора для химерной Н-цепи, содержащего геномную ДНК для Cγ1
В качестве экспрессирующего вектора, содержащего геномную ДНК для Cγ1, в качестве С-участка Н-цепи, можно использовать, например, HFE-PMh-gγ1 (публикации международной заявки WO 92/19759), или DHFR-ΔE-RVh-PM1f (публикация международной заявки WO 92/19759).
Для встраивания кДНК, кодирующей V-участок мышиной Н-цепи в эти экспрессирующие векторы, можно методом ПЦР ввести подходящие нуклеотидные последовательности. Эти подходящие нуклеотидные последовательности можно ввести методом ПЦР, используя ПЦР-праймер, сконструированный так, чтобы он содержал распознающую последовательность для подходящего фермента рестрикции 5'-конца, и консенсусную последовательность Kozak непосредственно перед стартовым кодоном, и ПЦР-праймер, сконструированный так, чтобы он включал с 3'-конца распознающую последовательность для соответствующего фермента рестрикции, и сплайсинговый донорный сайт, где первичный транскрипт геномной ДНК подвергается соответствующему сплайсингу, для превращения в мРНК.
Сконструированную таким образом кДНК, кодирующую V-участок мышиной Н-цепи, обрабатывают подходящими ферментами рестрикции, встраивают в вышеуказанный экспрессирующий вектор, и можно сконструировать химерную Н-цепь-экспрессирующий вектор, включающий ДНК Cγ1.
Конструирование экспрессируюшего вектора для кДНК химерной Н-цепи
Экспрессирующий вектор, содержащий кДНК Cγ1 в качестве С-участка Н-цепи, можно сконструировать следующим образом. Так, его можно сконструировать, получая мРНК из СНО-клетки, в которой были интегрированы экспрессирующий вектор DHFR-ΔE-RVh-PM1f (публикация международной заявки WO 92/19759), кодирующий геномную ДНК V-участка Н-цепи гуманизированного РМ1 антитела и С-участок Cγ1 Н-цепи человеческого антитела (N. Takahashi, et al., Cell 29, 671-679, 1982), и экспрессирующий вектор RVl-PM1a (публикация международной заявки WO 92/19759), кодирующий геномную ДНК V-участка L-цепи гуманизированного РМ1 антитела и С-участок κ-цепи гуманизированного РМ1 антитела, и С-участок κ-цепи L-цепи человеческого антитела; клонируя кДНК, включающую V-участок Н-цепи гуманизированного РМ1 антитела и С-участок Cγ1 Н-цепи человеческого антитела методом ОТ-ПЦР; и лигируя в подходящий экспрессирующий вектор для клеток животных, используя подходящие сайты для ферментов рестрикции.
Для непосредственного лигирования кДНК, кодирующей V-участок мышиной Н-цепи с кДНК, содержащей С-участок Cγ1 Н-цепи человеческого антитела, можно ввести подходящие нуклеотидные последовательности методом ПЦР.
Так например, эти подходящие нуклеотидные последовательности можно ввести методом ПЦР, используя ПЦР-праймер, сконструированный таким образом, что он содержит распознающую последовательность для подходящего фермента рестрикции с 5'-конца и консенсусную последовательность Kozak непосредственно перед стартовым кодоном, и ПЦР-праймер, сконструированный таким образом, что он содержит распознающую последовательность для подходящего фермента рестрикции, используемого для прямого лигирования C-участка Cγ1 Н-цепи у 3'-конца.
Экспрессирующий вектор, содержащий кДНК химерной Н-цепи, можно сконструировать, обрабатывая сконструированную таким образом кДНК, кодирующую V-участок мышиной Н-цепи, подходящим ферментом рестрикции, лигируя с вышеуказанной кДНК, содержащей С-участок Cγ1 Н-цепи, и встраивая в такой экспрессирующий вектор, как pCOS1 или pCHOl.
2) Конструирование L-цепи химерного антитела
Экспрессирующий вектор для L-цепи химерного антитела можно получить, связывая кДНК, кодирующую V-участок мышиной L-цепи с геномной ДНК или кДНК, кодирующей С-участок L-цепи человеческого антитела, а затем вводя ее в подходящий экспрессирующий вектор. В качестве С-участка L-цепи можно указать, например, κ-цепь или λ-цепь.
Конструирование экспрессирующего вектора для κ-цепи кДНК химерной L-цепи
Для конструирования экспрессирующего вектора, содержащего кДНК, кодирующую V-участок мышиной L-цепи, можно ввести подходящую нуклеотидную последовательность, используя метод ПЦР. Так, например, эти подходящие нуклеотидные последовательности можно ввести методом ПЦР, используя ПЦР-праймер, сконструированный таким образом, чтобы он содержал распознающую последовательность для подходящего фермента рестрикции и консенсусную последовательность Kozak у 5'-конца, и ПЦР-праймер, сконструированный таким образом, чтобы он содержал распознающую последовательность для подходящего фермента рестрикции 3'-конца.
С-участок κ-цепи человеческой L-цепи для связывания с V-участком мышиной L-цепи можно сконструировать, например, из HEF-PMlk-gκ (публикация международной заявки WO 92/19759), содержащей геномную ДНК. Вектор экспрессии для κ-цепи L-цепи кДНК химерного антитела можно сконструировать, вводя распознающие последовательности подходящих ферментов рестрикции с 5'-конца или с 3'-конца ДНК, кодирующей С-участок κ-цепи L-цепи методом ПЦР, лигируя сконструированный таким образом V-участок мышиной L-цепи с С-участком κ-цепи L-цепи, а затем встраивая в такой экспрессирующий вектор, как pCOS1 или pCHOl.
2. Конструирование реконструированного человеческого антитела
(1) Конструирование V-участка реконструированного человеческого анти-НМ 1.24 антитела
Для конструирования реконструированного человеческого антитела, в котором CDR мышиного моноклонального антитела был трансплантирован в человеческое антитело, желательно, чтобы существовала гомологичность между FR мышиного моноклонального антитела и FR человеческого антитела. Так, V-участки L-цепи и Н-цепи мышиного анти-НМ 1.24 антитела сравнивают с V-участками всех известных антител, структуры которых были установлены, используя банк данных Protein Data Bank.
V-участок L-цепи мышиного анти-НМ 1.24 антитела наиболее близок с консенсусной последовательностью подгруппы IV V-участка L-цепи человеческого антитела (HSGIV) с гомологичностью 66,4%. С другой стороны, он демонстрирует гомологичность в 56,9%, 55,8% и 61,5% с HSGI, HSGII и HSGIII соответственно.
V-участок L-цепи мышиного анти-НМ 1.24 антитела при сравнении с V-участком L-цепи известных человеческих антител демонстрирует гомологичность 67,0% с V-участком L-цепи человеческого антитела REI, одного из подгруппы I V-участка L-цепи человеческого антитела. Поэтому FR из REI используют в качестве исходного материала для конструирования V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела.
Конструируют вариант а V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела. В этом варианте FR человеческого антитела делают идентичным FR на основе REI, присутствующего в реконструированном человеческом САМРАТН-1Н антителе (см. Riechmann, L. et al., Nature 322, 21-25 (1988), FR содержащийся в варианте а V-участка L-цепи реконструированного человеческого РМ-1 антитела, описанного в публикации международной заявки WO 92/19759), и мышиный CDR делают идентичным с CDR в V-участке L-цепи мышиного анти-НМ 1.24 антитела.
V-участок Н-цепи мышиного анти-НМ 1.24 антитела наиболее близок консенсусной последовательности V-участка Н-цепи человеческого антитела (HSGI) с гомологичностью 54,7%. С другой стороны, он демонстрирует гомологичность в 34,6% и 48,1% с HSGII и HSGIII соответственно. При сравнении V-участка Н-цепи мышиного анти-НМ 1.24 антитела с V-участком Н-цепи известных человеческих антител, оказывается, что FR1 до FR3 наиболее близки с V-участком Н-цепи человеческого антитела HG3, одного из подгруппы I V-участка человеческой Н-цепи (Rechavi, et al., Proc. Natl. Acad. Sci. USA, 80, 855-859) с гомологичностью 67,3%.
Поэтому FR человеческого антитела HG3 используют в качестве исходного материала для конструирования V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела.
Однако, так как аминокислотная последовательность FR4 человеческого антитела HG3 не была описана, в качестве FR4 используют аминокислотную последовательность FR4 человеческого антитела JH6 (Ravetch, J.V. et al., Cell, 27, 583-591), которая демонстрирует наивысшую гомологичность с FR4 мышиного анти-НМ 1.24 антитела. FR4 JH6 имеет ту же самую аминокислотную последовательность, что и FR4 Н-цепи мышиного анти-НМ 1.24 антитела, за исключением только одной аминокислоты.
В первом варианте а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, FR1-FR3 делают идентичными FR1-FR3 человеческого антитела HG3, за исключением того, что аминокислоты в положении 30 в человеческого FR1 и в положении 71 человеческого FR3 делают идентичными аминокислотам мышиного анти-НМ 1.24 антитела, и CDR делают идентичным CDR в V-участке Н-цепи мышиного анти-НМ 1.24 антитела.
(2) Конструирование V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела.
L-цепь реконструированного человеческого анти-НМ 1.24 антитела конструируют, используя трансплантацию CDR методом ПЦР. Этот метод схематически представлен на фиг. 4. Восемь ПЦР-праймеров используют для конструирования реконструированного человеческого анти-НМ 1.24 антитела (вариант а), содержащего FR, полученный из человеческого антитела REI. Внешние праймеры А (Последовательность ИД 47) и Н (Последовательность ИД 48) создают для гибридизации с последовательностью ДНК HEF экспрессирующего вектора HEF-VL-gκ.
CDR трансплантированные праймеры L1S (Последовательность ИД 49), L2S (Последовательность ИД 50) и L3S (Последовательность ИД 51) содержат смысловую последовательность ДНК. CDR трансплантированные праймеры L1A (Последовательность ИД 52), L2A (Последовательность ИД 53) и L3A (Последовательность ИД 54) содержат антисмысловую последовательность ДНК, причем каждый содержит последовательность ДНК (20-23 п.н.), комплементарную последовательности ДНК у 5'-конца праймеров L1S, L2S и L3S соответственно.
На первой стадии ПЦР осуществляют четыре реакции: A-L1A, L1S-L2A, L2S-L3A и L3S-H, и каждый продукт ПЦР очищают. Четырем продуктам ПЦР из первой ПЦР дают возможность связаться друг с другом в соответствии с их собственной комплементарностью (см. WO 92-19759). Затем добавляют внешние праймеры А и Н для амплификации полной длины ДНК, кодирующей V-участок L-цепи реконструированного человеческого анти-НМ 1.24 антитела (вторая ПЦР). В вышеуказанной ПЦР в качестве матрицы можно использовать плазмиду HEF-RVL-M21a (публикация международной заявки WO 95-14041), кодирующую вариант а V-участка L-цепи реконструированного человеческого ONS-M21 антитела, основанного на FR, полученного из человеческого антитела REI.
На первой стадии ПЦР используют матричную ДНК и каждый из праймеров.
Продукты ПЦР A-L1A (215 п.н.), L1S-L2A (98 п.н.), L2S-L3A (140 п.н.) и L3S-H (151 п.н.) очищают, используя 1,5% агарозный гель с низкой температурой плавления, и соединяют во второй ПЦР. Во второй ПЦР используют каждый продукт первой ПЦР и каждый внешний праймер (А и Н).
Фрагмент ДНК длиной 516 п.н., полученный во второй ПЦР, очищают, используя 1,5% агарозный гель с низкой температурой плавления, расщепляют BamHI и HindIII, и полученные таким образом фрагменты клонируют в HEF экспрессирующий вектор HEF-VL-gκ. После определения последовательности ДНК плазмиду, содержащую фрагмент ДНК с правильной аминокислотной последовательностью V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела, называют плазмидой HEF-RVLa-AHM-gκ. Аминокислотная последовательность и нуклеотидная последовательность V-участка L-цепи, содержащиеся в этой плазмиде HEF-RVLa-AHM-gκ, представлены в Последовательности ИД 9.
Вариант b V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела, можно сконструировать с помощью мутагенеза, используя ПЦР. Праймеры мутагенеза FTY-1 (Последовательность ИД 55) и FTY-2 (Последовательность ИД 56) сконструированы таким образом, чтобы осуществить мутацию фенилаланина в положении 71 на тирозин.
После того, как вышеуказанные праймеры амплифицированы с использованием плазмиды HEF-RVLa-AHM-gκ в качестве матрицы, конечный продукт очищают. Расщепляя BamHI и HindIII, полученные фрагменты ДНК клонируют в HEF экспрессирующий вектор HEF-VL-gκ для получения плазмиды HEF-RVLb-AHM-gκ. Аминокислотная последовательность и нуклеотидная последовательность V-участка L-цепи, содержащиеся в этой плазмиде HEF-RVLb-AHM-gκ, представлены в Последовательности ИД 10.
(3) Конструирование V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела
3-1. Конструирование вариантов а-е V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела
ДНК, кодирующую V-участок Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, можно сконструировать следующим образом. Связывая последовательность ДНК, кодирующую FR 1-3 человеческого антитела HG3, и FR4 человеческого антитела JH6 с последовательностью ДНК, кодирующей CDR V-участка Н-цепи мышиного анти-НМ 1.24 антитела, можно сконструировать ДНК полной длины, кодирующую V-участок Н-цепи реконструированного человеческого анти-НМ 1.24 антитела.
Затем HindIII распознающий сайт/консенсусную последовательность KOZAK и BamHI распознающий сайт/сплайсинговую донорную последовательность, соответственно, присоединяют к 5'-концу и 3'-концу этой последовательности ДНК, с тем, чтобы обеспечить встраивание экспрессирующего вектора HEF.
Сконструированную таким образом последовательность ДНК разделяют на 4 олигонуклеотида. Затем олигонуклеотиды, которые потенциально препятствуют сборке этих нуклеотидов, подвергают компьютерному анализу в отношении вторичной структуры.
Последовательности четырех олигонуклеотидов RVH1-RVH4 представлены далее в Последовательностях ИД 57-60. Длина этих нуклеотидов составляет 119-144 основания, и они имеют перекрывающийся участок в 25-26 оснований. Среди этих нуклеотидов RVH2 (Последовательность ИД 58) и RVH4 (Последовательность ИД 60) имеют смысловую последовательность ДНК, a RVH1 (Последовательность ИД 57) и RVH3 (Последовательность ИД 59) имеют антисмысловую последовательность. Способ сборки этих четырех олигонуклеотидов с помощью ПЦР представлен на фиг.5.
ПЦР осуществляют, используя четыре олигонуклеотида и RHP1 (Последовательность ИД 60) и RHP2 (Последовательность ИД 62) в качестве внешних праймеров.
Амплифицированный фрагмент ДНК длиной 438 оснований очищают, расщепляют HindIII и BamHI, а затем клонируют в экспрессирующий вектор HEF-VH-gγl. После определения нуклеотидной последовательности плазмиду, содержащую фрагмент ДНК, кодирующий правильную аминокислотную последовательность V-участка Н-цепи называют HEF-RVHa-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащаяся в этой плазмиде HEF-RVHa-AHM-gγl, представлены в Последовательности ИД 11.
Каждый из вариантов b, с, d и е V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, конструируют следующим образом. При конструировании каждого из вариантов b и далее V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, трехмерную структурную модель V-участка мышиного анти-НМ 1.24 антитела можно сконструировать, чтобы предсказать положение аминокислотного остатка, который следует заменить в молекуле антитела.
Используя праймер мутагенеза BS (Последовательность ИД 63) и ВА (Последовательность ИД 64), сконструированные для осуществления мутации аргинина в положении 66 в лизин, и в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγl с помощью метода ПЦР, вариант b амплифицируют для получения плазмиды HEF-RVHb-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHb-AHM-gγl, представлены в последовательности ИД 12.
Используя праймер мутагенеза CS (Последовательность ИД 65) и СА (Последовательность ИД 66), сконструированные для осуществления мутации треонина в положении 73 в лизин, и в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγl методом ПЦР, вариант с амплифицируют для получения плазмиды HEF-RVHc-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHc-AHM-gγl, представлены в Последовательности ИД 13.
Используя праймер мутагенеза DS (Последовательность ИД 67) и DA (Последовательность ИД 68), сконструированные для осуществления мутации аргинина в положении 66 в лизин и треонина в положении 73 в лизин, и в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγl с помощью ПЦР, вариант d амплифицируют для получения плазмиды HEF-RVHo-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHd-AHM-gγl, представлены в Последовательности ИД 14.
Используя в качестве праймера мутагенеза ES (Последовательность ИД 69) и ЕА (Последовательность ИД 70), сконструированные для осуществления мутации валина в положении 67 в аланин и метионина в положении 69 в лейцин, и в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγl, вариант е амплифицируют для получения плазмиды HEF-RVHe-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHe-AHM-gγl, представлены в Последовательности ИД 15.
3-2. Конструирование гибридного V-участка Н-цепи
Конструируя гибридный V-участок Н-цепи, можно определить, какой FR V-участка гуманизированного антитела вносит вклад в активность связывания и в активность ингибирования связывания. Из двух сконструированных последовательностей, аминокислотные последовательности FR1 и FR2 получены из мышиного анти-НМ 1.24 антитела, тогда как FR3 и FR4 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела (мышиное/человеческое гибридное анти-НМ 1.24 антитело) в одном, и аминокислотные последовательности FR1 и FR2 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, а FR3 и FR4 получены из мышиного анти-НМ 1.24 антитела (человеческое/мышиное гибридное анти-НМ 1.24 антитело) в другом. Все аминокислотные последовательности CDR участков получены из мышиного анти-НМ 1.24 антитела.
Два гибридных V-участка Н-цепи конструируют методом ПЦР. Этот метод схематически представлен на фиг.6 и 7. Для конструирования двух гибридных V-участков Н-цепи можно использовать четыре праймера. Внешние праймеры а (Последовательность ИД 71) и h (Последовательность ИД 72) создают для гибридизации с последовательностью ДНК вектора экспрессирующего вектора HEF HEF-VH-gγl. Праймер HYS гибридной конструкции Н-цепи (Последовательность ИД 73) конструируют таким образом, чтобы он имел смысловую последовательность ДНК, а гибридный праймер HYA Н-цепи (Последовательность ИД 74) имел антисмысловую последовательность ДНК так, чтобы эти последовательности ДНК были бы комплементарны друг другу.
Для конструирования гибридного V-участка Н-цепи, в котором аминокислотные последовательности из FR1 и FR2 получены из мышиного анти-НМ 1.24 антитела, a FR3 и FR4 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, ПЦР с использованием плазмиды HEF-1.24-gγl в качестве матрицы, внешнего праймера а, и гибридного праймера Н-цепи HYA, и ПЦР с использованием плазмиды HEF-RVHa-AHM-gγl в качестве матрицы, гибридного праймера HYA Н-цепи, внешнего праймера h, осуществляют на первой стадии ПЦР, и каждый продукт ПЦР очищают.
Двум продуктам первой ПЦР дают возможность соединиться за счет из собственной комплементарности (публикация международной заявки WO 92-19759). Затем, добавляя внешние праймеры а и h, полной длины ДНК, кодирующую гибридный V-участок Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из мышиного анти-НМ 1.24 антитела, а последовательности FR3 и FR4 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, амплифицируют на второй стадии ПЦР.
Для конструирования гибридного V-участка Н-цепи, в котором аминокислотные последовательности из FR1 и FR2 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, а последовательности из FR3 и FR4 получены из мышиного анти-НМ 1.24 антитела, ПЦР с использованием плазмиды HEF-RVHa-AHM-gγl в качестве матрицы, внешнего праймера а, и гибридного праймера Н-цепи HYA, ПЦР с использованием плазмиды HEF-1.24-gγl в качестве матрицы, гибридного праймера HYS Н-цепи, и внешнего праймера h, осуществляют на первой стадии ПЦР, и каждый продукт ПЦР очищают.
Двум очищенным продуктам первой ПЦР дают возможность соединиться за счет их собственной комплементарности (публикация международной заявки WO 92-19759). Затем, добавляя внешние праймеры а и h, полученную ДНК полной длины, кодирующую гибридный V-участок Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из варианта а Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, а последовательности FR3 и FR4 получены из мышиного анти-НМ 1.24 антитела, амплифицируют на второй стадии ПЦР.
Методы первой ПЦР, очистки продуктов ПЦР сборки, второй ПЦР и клонирования в HEF экспрессирующий вектор HEF-VH-gγl осуществляют в соответствии со способом, представленным в примере 9 "Конструирование V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела". После определения последовательности ДНК плазмиду, которая содержит фрагмент ДНК, кодирующий правильную аминокислотную последовательность гибридного V-участка Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из мышиного анти-НМ 1.24 антитела, а последовательности FR3 и FR4 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, называют HEF-MH-RVH-AHM-gγl.
Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-MH-RVH-AHM-gγl, представлены в Последовательности ИД 75. Также плазмиду, которая содержит фрагмент ДНК, кодирующий правильную аминокислотную последовательность гибридного V-участка Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из варианта а реконструированного человеческого анти-НМ 1.24 антитела, а последовательности FR3 и FR4 получены из V-участка Н-цепи мышиного анти-НМ 1.24 антитела, называют HEF-HM-RVH-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-HM-RVH-AHM-AHM-gγl, представлены в последовательности ИД 76.
3-3. Конструирование вариантов f-s V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела
Каждый из вариантов f, g, h, i, j, k, 1, m, n, o, p, q, r и s V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела конструируют следующим образом. При конструировании каждого из вариантов f и далее V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела можно сконструировать трехмерную структурную модель V-участка мышиного анти-НМ 1.24 антитела, как указано ранее, для того, чтобы предсказать положение аминокислотного остатка, который нужно заместить в молекуле антитела.
Используя праймер мутагенеза FS (Последовательность ИД 78) и FA (Последовательность ИД 79), сконструированные так, чтобы осуществить мутацию треонина в положении 75 на серин, и валина в положении 78 на аланин, и в качестве матричной ДНК плазмиду HEF-RVHe-AHM-gγl, методом ПЦР вариант f амплифицируют до получения плазмиды HEF-RVHf-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHf-AHM-gγl, представлены в Последовательности ИД 16.
Используя праймер мутагенеза GS (Последовательность ИД 80) и GA (Последовательность ИД 81), сконструированные так, чтобы осуществить мутацию аланина в положении 40 на аргинин, и в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγl, вариант g амплифицируют до получения плазмиды HEF-RVHg-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHg-AHM-gγl, представлены в последовательности ИД 17.
Используя праймер мутагенеза FS и FA и в качестве матричной ДНК плазмиду HEF-RVHb-AHM-gγl, вариант h амплифицируют до получения плазмиды HEF-RVHh-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHh-AHM-gγl, представлены в Последовательности ИД 18.
Используя праймер мутагенеза IS (Последовательность ИД 82) и IA (Последовательность ИД 83), сконструированные так, чтобы осуществить мутацию аргинина в положении 83 на аланин и серина в положении 84 на фенилаланин, и в качестве матричной ДНК плазмиду HEF-RVRh-AHM-gγl, вариант i амплифицируют до получения плазмиды HEF-RVHi-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-учаcтка Н-цепи, содержащегося в плазмиде HEF-RVHi-AHM представлены в Последовательности ИД 19.
представлены в Последовательности ИД 19.
Используя праймер мутагенеза JS (Последовательность ИД 84) и JA (Последовательность ИД 85), сконструированные таким образом, чтобы осуществить мутацию аргинина в положении 66 в лизин, и в качестве матричной ДНК плазмиду HEF-RVHf-AHM-gγl, вариант j амплифицируют до получения плазмиды HEF-RVHj-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHj-AHM-gγl, представлены в Последовательности ИД 20.
Используя праймер мутагенеза КS (Последовательность ИД 86) и КА (Последовательность ИД 87), сконструированные таким образом, чтобы осуществить мутацию глутаминовой кислоты в положении 81 в глутамин, и в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγl, вариант k амплифицируют до получения плазмиды HEF-RVHk-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHk-AHM-gγl, представлены в последовательности ИД 21.
Используя праймер мутагенеза LS (Последовательность ИД 88) и LA (Последовательность ИД 89), сконструированные таким образом, чтобы осуществить мутацию глутаминовой кислоты в положении 81 на глутамин и серина в положении 82 на изолейцин, и в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγl, вариант 1 амплифицируют до получения плазмиды HEF-RVH1-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHI-AHM-gγl, представлены в Последовательности ИД 22.
Используя праймер мутагенеза MS (Последовательность ИД 90) и МА (Последовательность ИД 91), сконструированные таким образом, чтобы осуществить мутацию глутаминовой кислоты в положении 81 в глутамин, серина в положении 82 в изолейцин и треонина в положении 87 в серин, и в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγl, вариант m амплифицируют до получения плазмиды HEF-RVHm-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHm-AHM-gγl, представлены в Последовательности ИД 23.
Используя праймер мутагенеза NS (Последовательность ИД 92) и NA (Последовательность ИД 93), сконструированные таким образом, чтобы осуществить мутацию серина в положении 82 в изолейцин, и в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγl, вариант n амплифицируют до получения плазмиды HEF-RVHn-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHn-AHM-gγl, представлены в Последовательности ИД 24.
Используя праймер мутагененза OS (Последовательность ИД 94) и ОА (Последовательность ИД 95), сконструированные таким образом, чтобы осуществить мутацию треонина в положении 87 в серии, и в качестве матричной ДНК плазмиду HEF-RVHn-AHM-gγl, вариант о амплифицируют до получения плазмиды HEF-RVHo-AHM-gγl. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHo-AHM-gγl, представлены в Последовательности ИД 25.
Используя праймер мутагенеза PS (Последовательность ИД 96) и РА (Последовательность ИД 97), сконструированные таким образом, чтобы осуществить мутацию валина в положении 78 в аланин, и в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, вариант р амплифицируют с помощью ПЦР до получения плазмиды HEF-RVHp-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHp-AHM-gγ1, представлены в Последовательности ИД 26.
Используя праймер мутагенеза QS (Последовательность ИД 98) и QA (Последовательность ИД 99), сконструированные таким образом, чтобы осуществить мутацию треонина в положении 75 в серин, и в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, вариант q амплифицируют с помощью ПЦР до получения плазмиды HEF-RVHq-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHq-AHM-gγ1, представлены в последовательности ИД 27.
Используя праймер мутагенеза CS (Последовательность ИД 65) и СА (последовательность ИД 66) и в качестве матричной ДНК плазмиду HEF-RVHp-AHM-gγ1, вариант r амплифицируют с помощью ПЦР до получения плазмиды HEF-RVHr-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHr-AHM-gγ1, представлены в Последовательности ИД 28.
Используя праймер мутагенеза SS (Последовательность ИД 100) и SА (Последовательность ИД 101), сконструированный таким образом, чтобы осуществить мутацию метионина в положении 69 в изолайцин, и в качестве матричной ДНК плазмиду HEF-RVHr-AHM-gγ1, вариант s амплифицируют до получения плазмиды HEF-RVHs-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в плазмиде HEF-RVHs-AHM-gγ1, представлены в Последовательности ИД 102.
Сконструированные аминокислотные последовательности V-участка L-цепи представлены в табл. 1, а аминокислотные последовательности V-участка Н-цепи представлены в табл. 2-4.
3. Получение химерного антитела и реконструированного человеческого антитела.
Для получения химерного антитела или реконструированного человеческого антитела для каждого из них конструируют два экспрессирующих вектора, что включает экспрессирующий вектор, включающий ДНК, кодирующую V-участок мышиной Н-цепи и С-участок человеческой Н-цепи под контролем регулирующего экспрессию участка, такого как энхансер/промоторная система, и ДНК, кодирующую V-участок мышиной L-цепи и С-участок человеческой L-цепи под контролем регулирующего экспрессию участка, такого как энхансер/промоторная система, или экспрессирующий вектор, включающий ДНК, кодирующую V-участок гуманизированной Н-цепи и С-участок человеческой Н-цепи под контролем участка регулирующего экспрессию, такого как энхансер/промоторная система, и ДНК, кодирующую V-участок гуманизированной L-цепи и С-участок человеческой L-цепи под контролем участка регулирующего экспрессию, такого как энхансер/промоторная система.
Затем клетку-хозяин, такую как клетка млекопитающего, подвергают котрансформации, используя эти векторы, и трансформированные клетки культивируют in vitro или in vivo для получения химерного антитела или реконструированного человеческого антитела (см, например, публикацию международной заявки WO 91-16928). Более того, ген антитела вводят млекопитающему, такому как коза, для получения трансгенного животного, из молока которого можно получать химерное антитело или реконструированное человеческое антитело.
Кроме того, гены V-участка Н-цепи, С-участка Н-цепи, V-участка L-цепи и С-участка L-цепи лигируют в один вектор для трансформации клетки подходящего хозяина, и в результате получают антитела. Так, для экспрессии химерных антител, ДНК, кодирующую мышиную лидерную последовательность и V-участок Н-цепи и С-участок человеческой Н-цепи, присутствующие в клонированной кДНК, и ДНК, кодирующую мышиную лидерную последовательность и V-участок L-цепи и С-участок человеческой L-цепи вводят в один экспрессирующий вектор (публикация международной заявки WO 94-11523).
Для экспрессии реконструированного человеческого антитела ДНК, кодирующую V-участок гуманизированной Н-цепи и С-участок человеческой Н-цепи, и ДНК, кодирующую V-участок гуманизированной L-цепи и С-участок человеческой L-цепи вводят в один экспрессирующий вектор (публикация международной заявки WO 94-11523). Клетки-хозяева, трансформируют, используя указанный вектор, и трансформированные клетки-хозяева культивируют in vivo или in vitro, получая желаемое химерное антитело или реконструированное человеческое антитело.
Трансформант, который был трансформирован, как указано выше, геном, кодирующим желаемое химерное антитело или реконструированное человеческое антитело, культивируют, и полученное химерное антитело или реконструированное человеческое антитело можно выделить изнутри или извне клеток и очистить до гомогенности.
Выделение и очистку желаемого белка по настоящему изобретению химерного антитела или реконструированного человеческого антитела можно осуществить, используя аффинную колонку. В качестве колонки, в которой используют протеин А, например, можно указать HyperD, POROS, Sepharose F. F, и т.д. В другом варианте можно использовать методики обычного выделения и очистки, используемые для белков, и выбор метода ничем не ограничивается. Так, например, сочетания различных хроматографических методик, ультрафильтрации, высаливания, диализа и т.п. могут позволить выделить и очистить химерное антитело или реконструированное человеческое антитело.
Для получения химерного анти-НМ 1.24 антитела или реконструированного человеческого анти-НМ 1.24 антитела по настоящему изобретению можно использовать любой способ экспрессии, включающий, например, такие эукариотические клетки, как клетки животных, систему стабилизированной клеточной линии млекопитающего, систему клеток насекомых, систему грибковых клеток и систему дрожжевых клеток, и такие прокариотические клетки, как бактериальные клетки, такие как клетки Escherichia coli, и т.п. Предпочтительно, чтобы химерное антитело или реконструированное человеческое антитело по настоящему изобретению можно было экспрессировать в клетки COS, клетки СНО, клетки Hela, клетки Vero, миеломные клетки или клетки ВНК.
В этих случаях можно использовать обычные промоторы, которые используют для экспрессии клеток млекопитающих. Так, например, можно использовать человеческий цитомегаловирусный немедленно-ранний промотор (HCMV). Примеры экспрессирующих векторов, содержащих HCMV промотор, включают НСМV-VН-HCγ1, HCMV-VL-HCκ и т.д. и те, которые получены из pSV2neo (публикация международной заявки WO 92/19759).
Кроме того, в качестве промотора для экспресси гена в клетках млекопитающих для использования в настоящем изобретении можно использовать вирусные промоторы, такие как ретровирус, вирус полиомы, аденовирус, вирус обезьян 40 (SV40), и т.д., и промоторы, полученные из клеток млекопитающих, такие как фактор удлинения полипептидной цепи 1α(HTF-1α), и т.д. Так, например, если используют промотор SV40, экспрессию можно легко осуществить, используя метод Mulligan et al. (Nature 277, 108(1979)), а если используют промотор HTF-1α, можно использовать метод Mizushima, S. et al. (Nucleic Acids Research, 18, 5322, 1990).
В качестве источника репликации можно использовать источник, полученный из SV40, вируса полиомы, аденовируса, вируса папилломы крупного рогатого скота (BPV) и т.п., и для амплификации числа копий гена в клеточной системе хозяина, экспрессирующий вектор может включать, в качестве селективного маркера, ген аминогликозидфосфотрансферазы (АРН), ген тимидинкиназы (ТК), ген ксантингуанинфосфорибозилтрансферазы Е. coli (Ecogpt), ген дигидрофолатредуктазы (DHRF), и т.п.
4. Активность ингибирования связывания химерного антитела или реконструированного человеческого антитела
(1) Измерение концентрации антител
Концентрацию очищенных антител можно измерить с помощью ELISA, или измеряя поглощение.
Планшеты ELISA для определения концентрации антител можно подготовить следующим образом. Каждую лунку 96-луночного планшета для ELISA (например, Maxisorp, изготовитель NUNC) иммобилизуют 100 мкл козьего антитела против человеческого IgG в концентрации 1 мкг/мл.
После блокирования 100 мкг/мл разбавленного буфера (например, 50 мМ Трис-HCl, 1 мМ MgCl2, 0,15 М NaC1, 0,05% Tween 20, 0,02% NaN3, 1% бычьего сывороточного альбумина (BSA), рН 8,1), в каждую ячейку добавляют серийные разведения надосадочной культуральной жидкости клеток, в которых экспрессировано химерное антитело, гибридное антитело или реконструированное человеческое антитело, например, культуральной надосадочной жидкости клеток COS или клеток СНО, или очищенного химерного антитела, гибридного антитела или реконструированного человеческого антитела. Затем добавляют 100 мкл конъюгированного с щелочной фосфатазой козьего антитела против человеческого IgG, добавляют 1 мг/мл субстратного раствора (Sigma 104, п-нитрофенилфосфата, производства SIGMA), а затем измеряют поглощение на 405 нм, используя ридер планшетов для микротитрования (BioRad). В качестве стандарта для измерения концентрации можно использовать человеческий IgG1κ (производство The Binding Site). Значение концентрации очищенных антител получают, измеряя поглощение при 280 нм и принимая величину 1 мг/мл как 1,35 ОП.
(2) Активность связывания
Активность связывания можно измерить с помощью Cell-ELISA, используя человеческую амниотическую клеточную линию WISH (ATCC CCL25). Планшеты для клеточного ELISA можно получить следующим образом. Подготовленные клетки WISN в соответствующей концентрации в PRMI 1640 среде, дополненной 10% околоплодной сывороткой теленка, добавляют в 96-луночные планшеты, инкубируют в течение ночи, и после двукратного промывания PBS(-), фиксируют 0,1% глутаральдегидом (производство Nakalai tesque).
После блокирования добавляют 100 мкл серийных разведений культуральной надосадочной среды клеток, в которых химерное анти-НМ 1.24 антитело, гибридное анти-НМ 1.24 антитело или реконструированное человеческое анти-НМ 1.24 антитело экспрессировано, например, культуральной надосадочной жидкости клеток COS или клеток СНО, или очищенного химерного анти-НМ 1.24 антитела, гибридного анти-НМ 1.24 антитела или реконструированного человеческого анти-НМ 1.24 антитела в каждую лунку, инкубируют при комнатной температуре в течение двух часов, а затем добавляют меченное пероксидазой кроличье антитело против человеческого IgG (производство DAKO).
После инкубирования в течение 1 ч при комнатной температуре добавляют раствор субстрата и снова инкубируют. Затем реакцию останавливают, добавляя 50 мкл 6 н. серной кислоты, а затем измеряют поглощение при 490 нм, используя ридер MICROPLATE READER Model 3550 (BioRad).
(3) Измерение активности ингибирования связывания
Активность ингибирования связывания биотинилированным мышиным анти-НМ 1.24 антителом определяют с помощью клеточного ELISA, используя человеческую амниотическую клеточную линию WISH (ATCC CCL25). Планшет для клеточного ELISA можно приготовить в соответствии с вышеуказанным (2). Клетки WISH, подготовленные в соответствующей концентрации в PRMI 1640 среде, дополненной 10% околоплодной сывороткой теленка помещают в 96-луночный планшет, инкубируют в течение ночи, и после двукратной промывки PBS(-) фиксируют 0,1% глутаральдегидом (Nacalai tesque).
После блокирования в каждую лунку добавляют 50 мкл серийных разведений культуральной надосадочной среды клеток, в которых химерное анти-НМ 1.24 антитело, гибридное анти-НМ 1.24 антитело или реконструированное человеческое анти-НМ 1.24 антитело экспрессировано, например, культуральной надосадочной жидкости клеток COS или клеток СНО, или очищенного химерного анти-НМ 1.24 антитела, гибридного анти-НМ 1.24 антитела или реконструированного человеческого анти-НМ 1.24 антитела, и одновременно добавляют 50 мкг 2 мкг/мл биотинилированного мышиного анти-НМ 1.24 антитела, затем инкубируют при комнатной температуре в течение двух часов и, после промывки, добавляют меченный пероксидазой стрептавидин (производство DAKO).
После инкубирования при комнатной температуре в течение 1 ч и после промывки добавляют раствор субстрата, а затем инкубируют. Затем реакцию останавливают, добавляя 50 мкл серной кислоты, и затем измеряют поглощение на 490 нм, используя ридер MICROPLATE READER Model 3550 (BioRad).
Измерение ADCC-активности
ADCC-активность химерного антитела или реконструированного человеческого антитела по настоящему изобретению можно измерить следующим образом. Во-первых, мононуклеарные клетки выделяют из периферической человеческой крови или костного мозга методом центрифугирования в градиенте плотности и приготавливают как эффекторные клетки. Миеломные клетки человека приготавливают как клетки-мишени, осуществляя введение 51Сr метки в RPMI 8226 клетки (АТСС CCL155). Затем химерное антитело или реконструированное человеческое антитело, которое подлежит определению на ADCC-активность, добавляют к меченым клеткам-мишеням, инкубируют, а затем добавляют соответствующее количество эффекторных клеток к клеткам-мишеням и инкубируют.
После инкубации надосадочную жидкость отбирают для измерения радиоактивности, используя счетчик гамма-излучения. В это время 1% NP-40 можно использовать для измерения максимальной испускаемой радиоактивности. Цитотоксичность (%) можно рассчитать как (А-С)/(В-С)х(100, где А представляет испускаемую радиоактивность (имп/мин) в присутствии антитела, В представляет испускаемую радиоактивность (имп/мин), выделенную за счет NP-40, и С представляет радиоактивность (имп/мин), испускаемую одной только культуральной жидкостью без антитела.
Если ADCC-активность или CDC-активность ожидается зарегистрировать в С-участке антитела, в качестве С-участка антитела можно использовать человеческий Cγ1 или человеческий Cγ3. Более того, добавляя, изменяя или модифицируя часть аминокислот С-участка антитела, можно индуцировать более высокую активность ADCC или активность CDC.
Так, например, существует IgM-подобная полимеризация IgG путем замещения аминокислот (Smith, R.I.F. & Morrison, S.L, BIO/TECHNOLOGY (1994) 12, 683-688), IgM-подобная полимеризация IgG путем добавления аминокислот (Smith, R. I.F. et al., J. Immunol. (1995) 154, 2226-2236), экспрессия путем тандемного связывания генов, кодирующих L-цепь (Shuford, W. et al., Science (1991) 252, 724-727), димеризация IgG путем замещения аминокислот (Саrоn, Р.С. et al., J. Exp. Med. (1992) 176, 1191-1195, Shopes, B.J. Immunology (1992) 148 2918-2922, димеризация IgG в результате химической модификации (Wolff E.A. et al., Cancer Res. (1993) 53, 2560-2565), и введение эффекторной функции в результате изменения аминокислот в шарнирном участке антител (Norderhaug, L. et al., Eur. J. Immunol (1991) 21, 2379-2384). Это можно осуществить сайтнаправленным мутагенезом олигомеров, используя праймеры, добавляя последовательности оснований, используя сайты расщепления ферментов рестрикции, и химические модификаторы, которые индуцируют ковалентное связывание.
In vivo диагностика миеломы
Химерное анти-НМ 1.24 антитело или реконструированное человеческое анти-НМ 1.24 антитело по настоящему изобретению можно использовать в качестве in vivo диагностического средства для миеломы путем связывания его с меченым соединением, таким как радиоизотоп и т.п.
Кроме того, фрагменты химерного анти-НМ 1.24 антитела или реконструированного человеческого анти-НМ 1.24 антитела, такие как Fab, F(ab')2, Fv, или одноцепочечный Fv (scFv), где Fv или Fv Н-цепи и L-цепь связаны подходящим линкером, который был связан с таким меченым соединением, как радиоизотоп и т.п., могут быть использованы в качестве in vivo диагностических средств для миеломы.
Конкретно, эти фрагменты антител можно получить, конструируя ген, кодирующий эти фрагменты антител, вводя их в экспрессирующий вектор, а затем экспрессируя в клетках подходящего хозяина, или расщепляя химерное анти-НМ 1.24 антитело или реконструированное человеческое анти-НМ 1.24 антитело, подходящим ферментом.
Вышеуказанные in vivo диагностические средства для миеломы можно системно водить парентерально.
Фармацевтическая композиция и лекарственное средство для лечения миеломы
Для подтверждения терапевтического действия химерного анти-НМ 1.24 антитела или гуманизированного анти-НМ 1.24 антитела по настоящему изобретению, указанные антитела вводят животному, которому трансплантированы миеломные клетки, и оценивают противоопухолевое действие.
В качестве миеломных клеток, которые нужно трансплантировать животным, предпочтительно использовать человеческие миеломные клетки, и можно в качестве примера указать КРММ2 (публикация заявки на патент Японии без экспертизы (Kokai) 7-236475), RPMI8226 (АТСС CCL 155), ARH-77 (ATCC CRL 1621) и S6B45 (Suzuki, H. et al., Eur. J. Immunol. (1992) 22, 1989-1993). В качестве животных, которым можно трансплантировать указанные клетки, предпочтительны животные с ослабленными иммунными функциями, или животные без иммунных функций, и в качестве примера можно указать мышей nude, SCID мышей, мышей beige и крыс nude.
Кроме того, противоопухолевое действие, которое нужно оценить, можно подтвердить по изменению количества человеческих иммуноглобулинов в сыворотке путем изменения объема и/или веса опухоли, по изменению веса белков Веnсе Jones в моче, по продолжительности выживания животных или т.п.
Фармацевтические композиции или лекарственные средства для лечения миеломы, которые содержат в качестве активного ингредиента химерное анти-НМ 1.24 антитело или реконструированное человеческое анти-НМ 1.24 антитело по настоящему изобретению можно системно или местно вводить парентерально. Так, например, можно выбрать внутривенные инъекции, такие как капельное введение, внутримышечные инъекции, внутрибрюшинные инъекции или подкожные инъекции, и выбрать соответствующий дозовый режим в зависимости от возраста и состояния здоровья пациента.
Эффективную дозу выбирают в интервале от 0,01 мг до 1000 мг/кг веса тела/на дозу. В другом варианте можно выбрать дозу в 5 мг/кг веса тела, предпочтительно от 50 до 100 мг/кг веса тела.
Фармацевтические композиции или терапевтические агенты для лечения миеломы, которые содержат в качестве активного ингредиента химерное анти-НМ 1.24 антитело или реконструированное человеческое анти-НМ 1.24 антитело по настоящему изобретению, могут содержать фармацевтически приемлемые носители или добавки в зависимости от выбранного способа введения.
В качестве примеров таких носителей или добавок можно указать воду, фармацевтически приемлемые органические растворители, коллаген, поливиниловый спирт, поливинилпирролидон, карбоксивиниловый полимер, натрийкарбоксиметилцеллюлозу, натрийполиакрилат, альгинат натрия, водорастворимый декстран, натрийкарбоксиметилкрахмал, пектин, метилцеллюлозу, этилцеллюлозу, ксантангам, гуммиарабик, казеин, желатин, агар, диглицерин, глицерин, пропиленгликоль, полиэтиленгликоль, вазелин, парафин, стеариловый спирт, стеариновую кислоту, человеческий сывороточный альбумин (HSA), маннит, сорбит, лактозу, фармацевтически приемлемые поверхностно-активные агенты и т.п. Возможные для использования добавки можно выбрать из вышеперечисленного или их комбинаций (не ограничиваясь вышеперечисленным).
ПРИМЕРЫ
Далее настоящее изобретение будет объяснено более подробно.
Пример 1. Клонирование кДНК, кодирующей V-участок мышиного анти-НМ 1.24 антитела
1. Выделение матричной РНК (мРНК)
Используя набор для выделения Fast Track mRNA Isolation Kit Version 3.2 (производитель Invitrogen) в соответствии с прилагаемыми инструкциями мРНК выделяют из 2•108 гибридомных клеток (FERM BP-5233), которые продуцируют мышиное анти-НМ 1.24 антитело.
2. Амплифцикация гена, кодирующего вариабельный участок антитела методом ПЦР
ПЦР осуществляют, используя термореактор Thermal Cycler (производство Perkin Elmer Cetus).
2-1. Амплификация и фрагментация гена, кодирующего V-участок мышиной L-цепи
Из выделенной таким образом мРНК синтезируют одноцепочечную кДНК, используя набор для синтеза AMV Reverse Trans-criptase First-strand cDNA Synthesis Kit (изготовитель Life Science), и используют для ПЦР. В качестве праймеров для ПЦР были использованы MKV (Mouse Kappa Variable) праймеры (Jones, S.T. et al. Bio/Technology, 9, 88-89 (1991), представленные в Последовательностях ИД 29-39, которые гибридизуются с лидерной последовательностью мышиной L-цепи каппа-типа.
100 мкл ПЦР раствора, содержащего 10 мМ Трис-НСl (рН 8,3), 50 мM КС1, 0,1 мМ дНТФ (дАТФ, дГТФ, дЦТФ, дТТФ), 1,5 мМ MgCl2, 5 Ед ДНК-полимеразы Ampli Taq (производство Perkin Elmer Cetus), 0,25 мМ праймеров MKV, представленных в Последовательностях ИД 29-39, 3 мМ праймера МКС, представленного в Последовательности ИД 40, и 100 нг одноцепочечной кДНК покрывают 50 мкл минерального масла, а затем нагревают сначала при температуре 94oС в течение 3 мин, затем при 94oС в течение 1 мин, и при 55oС в течение 1 мин, и при 72oС в течение 1 мин в указанном порядке. После повторения этого цикла 30 раз реакционную смесь инкубируют при 72oС в течение 10 мин. Амплифицированный фрагмент ДНК выделяют с помощью агарозного геля с низкой температурой плавления (производство Sigma), и расщепляют XmaI (производство New England Biolabs) и SalI (производство Takara Shuzo) при 37oС.
2-2. Амплификация и фрагментация кДНК, кодирующей V-участок мышиной Н-цепи
Ген, кодирующий V-участок мышиной Н-цепи, амплифицируют по способу 5'-RACE (Rapid Amplification of cDNA ends; Forhman, M.A. et al., Proc. Natl. Acad. Sci. USA, 85 8998-9002, (1988), Edwards, J.B.D.M., et al, Nucleic Acids Res. , 19, 5227-5232, (1991)). После того, как синтезирована кДНК, с использованием праймера Р1 (Последовательность ИД 41), который специфически гибридизуется с константным участком мышиного IgG2a, кДНК, кодирующую V-участок мышиной Н-цепи, амплифицируют с помощью набора 5'-AmpliFINDER RACE KIT (производство CLONTECH), используя праймер МНС2а (Последовательность ИД 42), который специфически гибридизуется с константным участком мышиного IgG2a и якорным праймером (Последовательность ИД 77), прилагаемым к набору. Амплифицированный ДНК фрагмент очищают с помощью агарозного геля с низкой температурой плавления (производство Sigma) и расщепляют EcoRI (производство Takara Shuzo) и XmaI (производство New England Biolabs) при 37oС.
3. Лигирование и трансформация
Фрагмент ДНК, включающий ген, кодирующий V-участок мышиной L-цепи каппа-типа, полученной, как указано выше, лигируют с вектором pUC19, полученным в результате расщепления SalI и XmaI, путем взаимодействия в реакционной смеси, содержащей 50 мМ Трис-НСl (рН 7,6), 10 мМ MgCl2, 10 мМ дитиотрейтола, 1 мМ АТФ, 50 мг/мл полиэтиленгликоля (8000) и одну Ед. Т4-лигазы (изготовитель GIBCO-BRL) при 16oС в течение 2,5 ч. Аналогично, фрагмент ДНК, включающий ген, кодирующий V-участок мышиной Н-цепи, подвергают взаимодействию и лигируют с вектором pUC19, полученным в результате расщепления EcoRI и XmaI при температуре 16oС в течение трех часов.
Затем 10 мкл вышеуказанной смеси лигирования добавляют к 50 мкл компетентных клеток Escherichia coli DH5α, оставляют на льду на 30 мин, выдерживают одну минуту при 42oС, и снова на льду в течение 1 мин.
Затем добавляют 400 мкл 2 х YT среды (Molecular Cloning: A Laboratory Manual, Sambrook et al., Cold Spring Harbor Laboratory Press, (1989)), инкубируют при 37oС в течение 1 ч, а затем Е. coli высевают на агарную 2 х YT среду (Molecular Cloning: A Laboratory Manual, Sambrook et al., Cold Spring Harbor Laboratory Press, (1989)), содержащую 50 мкг/мл ампициллина, а затем инкубируют в течение ночи при 37oС, получая в результате Е. coli-трансформант.
Этот трансформант культивируют в течение ночи при 37oС в 10 мл 2 х YT среды, содержащей 50 мкг/мл ампициллина, а затем из этой культуры получают плазмидную ДНК, используя щелочной метод (Molecular Cloning: A Laboratory Manual, Sambrook et al., Cold Spring Harbor Laboratory Press, (1989)).
Полученную таким образом плазмиду, содержащую ген, кодирующий V-участок мышиной L-цепи каппа-типа, полученной из гибридомы, которая продуцирует анти-НМ 1.24 антитело, называют pUCHMVL9. Полученную вышеуказанным способом плазмиду, содержащую ген, кодирующий V-участок мышиной Н-цепи, полученной из гибридомы, которая продуцирует анти-НМ 1.24 антитело, называют pUCHMVHR16.
Пример 2. Определение нуклеотидной последовательности ДНК
Нуклеотидную последовательность кДНК, кодирующей участок в вышеуказанной плазмиде, определяют, используя автоматический ДНК-секвенатор (изготовитель Applied Biosystem Inc. ) в соответствии с протоколом, предлагаемым изготовителем.
Нуклеотидная последовательность гена, кодирующего V-участок L-цепи мышиного анти-НМ 1.24 антитела, содержащегося в плазмиде pUCHMVL9, представлена в Последовательности ИД 1. Нуклеотидная последовательность гена, кодирующего V-участок Н-цепи мышиного анти-НМ 1.24 антитела, содержащегося в плазмиде pUCHMVHR16, представлена в Последовательности ИД 2.
Пример 3. Определение CDR
Полные структуры V-участков L-цепи и Н-цепи сходны друг с другом в том, что четыре каркасные участка связаны тремя гипервариабельными участками, т. е. участками, определяющими комплементарность (CDR). Аминокислотная последовательность каркасного участка относительно высоко консервативна, но вариация в последовательности аминокислот CDR чрезвычайно высока (Kabat, E.A., et al. , "Sequences of Proteins of Immunological Interest", US Dept. Health and Human Services, 1983).
На основании этих фактов аминокислотную последовательность вариабельного участка анти-НМ 1.24 антитела сравнивают с аминокислотной последовательностью антител в базе данных для изучения гомологичности, и участок CDR определяют, как представлено в табл. 5.
Пример 4. Подтверждение экспрессии клонированной кДНК (Конструирование химерного анти-НМ 1.24 антитела)
1. Конструированием экспрессирующего вектора
Для конструирования экспрессирующего вектора, который экспрессирует химерное анти-НМ 1.24 антитело, кДНК клоны PUCHMVL9 и pUCHMVHRl6 кодирующие V-участки L-цепи и Н-цепи мышиного анти-НМ 1.24 антитела соответственно модифицируют методом ПЦР, а затем вводят в экспрессирующий вектор HEF (публикация международной заявки WO 92-19759).
Обратный праймер ONS-L722S (Последовательность ИД 43) для V-участка L-цепи и обратный праймер VHR16S (Последовательность ИД 44) для V-участка Н-цепи конструируют таким образом, чтобы они гибридизовались с ДНК, кодирующей начало лидерной последовательности V-участка каждого, и чтобы они имели консенсусную последовательность Kozak (Kozak, M. et al., J. Mol. Biol., 196, 947-950, (1987)), и сайт распознавания для фермента рестрикции HindIII. Обратный праймер VL9A (Последовательность ИД 45) для V-участка L-цепи и обратный праймер VHR16A (Последовательность ИД 46) для V-участка Н-цепи конструируют таким образом, чтобы они гибридизовались с последовательностью ДНК, кодирующей конец J-участка, и чтобы они имели сплайсинговую донорную последовательность и сайт распознавания для фермента рестрикции BamHI.
100 мкл реакционной смеси ПЦР, содержащей 10 мМ Трис-НСl (рН 8,3), 50 мМ KC1, 0,1 мМ дНТФ, 1,5 мМ MgCl2, 100 пМ каждого из праймеров, 100 нг матричной ДНК (pUCHMVL9 или pUCHMVHR16), и 5 Ед. Ampli Taq фермента покрывают 50 мкл минерального масла, а затем, после начального денатурирования при 94oС, нагревают при 94oС в течение 1 мин, при 55oС в течение 1 мин, и при 72oС в течение 1 мин, и так 30 циклов, и в конце инкубируют при 72oС в течение 10 мин.
Продукт ПЦР очищают в 1,5% агарозном геле с низкой температурой плавления и расщепляют HindIII и BamHI, а затем клонируют в HEF-VL-gκ для V-участка L-цепи, и в HEF-VH-gγ1 для V-участка Н-цепи. После определения последовательности ДНК, плазмиды, содержащие фрагмент ДНК, который содержит правильную последовательность ДНК, называют HEF-1.24L-gκ и НЕF-1.24Н-gγ1, соответственно.
Участки, кодирующие соответствующий вариабельный участок из вышеуказанных плазмид HEF-1.24L-gκ и НЕF-1.24Н-gγ1, расщепляют ферментами рестрикции HindIII и BamHI для создания рестрикционных фрагментов, которые встраивают в HindIII сайт и в BamHI сайты плазмидного вектора pUC19, и их называют pUC19-1.24L-gκ и pUC19-1.24H-gγ1, соответственно.
Escherichia coli, содержащие соответствующие плазмиды pUC19-1.24L-gκ и pUC19-l. 24H-gγ1, называют Escherichia coli DH5α(pUC19-1.24L-gκ) и Escherichia coli DH5α(pUC19-1.24Hgγ1); они были интернационально депонированы 29 августа 1996 года National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, MITI (Higashi 1-Chome 1-3, Tsukuba city, Ibalaki prefecture, Japan) под регистрационными номерами FERM-5646 и FERM-5644, соответственно, в соответствии с Будапештским Соглашением.
2. Трансфекция клеток COS-7
Для наблюдения временной экспрессии химерного анти-НМ 1.24 антитела, вышеуказанные экспрессирующие векторы были тестированы в клетках COS-7 (ATCC CRL-1651). HEF-1.24L-gκ и HEF-1.24H-gγ1 котрансформируют в клетки COS-7 с помощью электропорации, используя прибор Gene Pulser (изготовитель BioRad). Каждую ДНК (10 мкг) добавляют к 0,8 мл аликвот 1•107 клеток/мл в PBS, и обрабатывают импульсами при 1500 В и емкости 25 мкФ.
После периода восстановления 10 мин при комнатной температуре подвергшиеся электропорации клетки добавляют к 30 мл DMEM культуральной жидкости (GIBCO), содержащей 10% околоплодную сыворотку теленка без гамма-глобулина. После инкубирования в течение 72 ч в инкубаторе с CO2 BNA120D (производства TABAI), надосадочную культуральную жидкость собирают, и осколки клеток удаляют центрифугированием, что потом используют в дальнейших экспериментах.
3. Проточный цитометрический анализ (FCM)
Активность связывания антигена химерным анти-НМ 1.24 антителом исследуют с помощью FCM (проточная цитометрия), используя клетки КРММ2. После того, как 4,7•105 клеток КРММ2 (публикация заявки на патент Японии без экспертизы (Kokai) 7-236475) промывают PBS(-), добавляют 50 мкл культуры клеток COS-7, которые продуцируют вышеуказанное химерное анти-НМ 1.24 антитело, и 50 мкл FACS буфера (PBS(-), содержащий 2% околоплодной сыворотки теленка и 0,1% азида натрия), или 5 мкл 500 мкг/мл очищенного мышиного анти-НМ 1.24 антитела и 95 мкл FACS буфера, и инкубируют на льду в течение одного часа.
В качестве контроля добавляют 50 мкл 2 мкг/мл химерного SK2 (публикация международной заявки WO 94-28159) и 50 мкл FACS буфера, или 5 мкл 500 мкг/мл очищенного мышиного IgG2aκ (UPC10) (изготовитель CAPPEL) вместо очищенного мышиного анти-НМ 1.24 антитела, и 95 мкл FACS буфера, и инкубируют аналогичным образом. После промывки FACS буфером добавляют 100 мкл 25 мкг/мл конъюгированного с ФИТЦ козьего антитела против человеческого антитела(GAH) (изготовитель CAPPEL), или 10 мкг/мл конъюгированного с ФИТЦ козьего антитела против мышиного антитела (GAM) (изготовитель Becton Dickinson), и инкубируют при температуре льда в течение 30 мин. После промывки FACS буфером все это суспендируют в 1 мл FACS буфера, и измеряют интенсивность флуоресценции каждой лунки с помощью FACScan (изготовитель Becton Dickinson).
Как видно на фиг.1, выявлено, что химерное анти-НМ 1.24 антитело связывается с клетками КРММ2, так как пик интенсивности флуоресценции сдвигается вправо для клеток, к которым добавлено химерное анти-НМ 1.24 антитело, по сравнению с контролем, аналогично случаю, когда добавляют мышиное анти-НМ 1.24 антитело. Это подтверждает тот факт, что клонированная кДНК кодирует вариабельный участок мышиного анти-НМ 1.24 антитела.
Пример 5. Создание клеточной линии СНО, которая стабильно продуцирует химерное анти-НМ 1.24 антитело
1. Конструирование экспрессирующего вектора для химерной Н-цепи
После расщепления вышеуказанной плазмиды НЕF-1.24Н-gγ1 ферментами рестрикции PvuI и BamHI фрагмент около 2,8 т.п.н., содержащий EF1 промотор и ДНК, кодирующую V-участок Н-цепи мышиного анти-НМ 1.24 антитела, выделяют, используя 1,5% агарозный гель с низкой температурой плавления, затем вышеуказанный фрагмент ДНК встраивают во фрагмент длиной примерно 6 т.п.н., полученный при расщеплении экспрессирующего вектора, использованного для экспрессирующего вектора человеческой Н-цепи, DHFR-ΔE-Rvh-PM1f (публикация международной заявки WO 92/19759), содержащего DHFR ген и ген, кодирующий константный участок человеческой Н-цепи с помощью PvuI и BamHI, для конструирования экспрессирующего вектора DHFR-ΔE-HEF-1.24H-gγ1 для Н-цепи химерного анти-НМ 1.24 антитела.
2. Введение гена в клетки СНО
Для создания системы, стабильно продуцирующей химерное анти-НМ 1.24 антитело, гены вышеуказанных экспрессирующих векторов HEF-1.24L-gκ и DHFR-ΔE-HEF-1.24H-gγ1, которые были линеаризованы за счет расщепления PvuI, одновременно вводят в СНО клетку DXB11 (полученную из Medical Research Council Collaboration Center) методом электропорации в условиях, аналогичных вышеуказанным вышеуказанная трансфекция в клетки COS-7.
3. Амплификация гена с помощью МТХ
Среди клеток СНО с введенным геном, только те клетки СНО, в которые были введены как экспрессирующий вектор L-цепи, так и экспрессирующий вектор Н-цепи, могут выжить в α-MEM культуральной жидкости, не содержащей нуклеозида (производство GIBCO-BRL), к которой добавляют 500 мкг/мл G418 (производство GIBCO-BRL) и 10% околоплодную сыворотку теленка, и таким образом производят отбор. Затем к этой культуральной жидкости добавляют 10 нМ МТХ (производство Sigma). Среди растущих клонов отбирают те, которые продуцируют химерное анти-НМ 1.24 антитело в больших количествах. В результате получают клоны 8-13, которые демонстрируют эффективность продуцирования около 20 мкг/мл химерного антитела, и называют клеточными линиями, продуцирующими химерное анти-НМ 1.24 антитело.
Пример 6. Конструирование химерного анти-НМ 1.24 антитела
Химерное анти-НМ 1.24 антитело конструируют следующим образом. Вышеуказанные клетки СНО, продуцирующие химерное анти-НМ 1.24 антитело, непрерывно культивируют в течение 30 суток, используя среду Дюльбекко, модифицированную Исковым (производство GIBCO-BRL), содержащую 5% сыворотку новорожденного теленка, не содержащую гамма-глобулина (производство GIBCO-BRL), с помощью прибора для клеточной культуры высокой плотности Verax system 20 (производство CELLEX BIOSCIENCE Inc.).
На 13, 20, 23, 26 и 30 сутки после начала культивирования культуральную жидкость извлекают, используя фильтрующее устройство под давлением SARTOBRAN (производство Sartorius), а затем химерное анти-НМ 1.24 антитело аффинно очищают, используя крупномасштабную систему сбора антитела Afi-Prep System (производство Nippon Gaishi) и колонку Super Protein А (объем слоя: 100 мл, производство Nippon Gaishi), используя PBS в качестве абсорбирующего/промывочного буфера и 0,1 М натрийцитратный буфер (рН 3) в качестве элюирующего буфера в соответствии с прилагаемой инструкцией. рН элюированных фракций доводят до 7,4, немедленно добавляя 1 М Трис-НС1 (рН 8,0). Концентрацию антител измеряют по поглощению на 280 нм и рассчитывают, принимая величину 1,35 ОП, соответствующую 1 мкг/мл.
Пример 7. Определение активности химерного анти-НМ 1.24 антитела.
Содержание химерного анти-НМ 1.24 антитела оценивают по следующей активности ингибирования связывания.
1. Измерение активности ингибирования связывания
1-1. Конструирование биотинилированного анти-НМ 1.24 антитела
После того, как мышиное анти-НМ 1.24 антитело разбавляют 0,1 М бикарбонатным буфером до 4 мг/мл, 4 мкл добавляют 50 мг/мл биотин-N-гидроксисукцинимида (производство EY LABS Inc.), и реакцию ведут при комнатной температуре в течение 3 ч. После этого добавляют 1,5 мл 0,2 М раствора глицина, инкубируют при комнатной температуре в течение 30 мин для остановки реакции, а затем биотинилированные IgG фракции собирают, используя PD-10 колонку (производство Pharmacia Biotech).
1-2. Измерение активности ингибирования связывания
Активность ингибирования связывания биотинилированным мышиным анти-НМ 1.24 антителом измеряют с помощью клеточного ELISA, используя человеческую амниотическую мембранную клеточную линию WISH клеток (АТСС CCL 25). Планшеты для клеточного ELISA подготавливают следующим образом. В 96-луночный планшет добавляют 4•105 клеток/мл, приготовленных в PRMI 1640 среде, дополненной 10% околоплодной сывороткой теленка, инкубируют в течение ночи, и после двукратной промывки PBS(-) иммобилизуют 0,1% глутаральдегидом (изготовитель Nakalai tesque).
После блокирования 50 мкл серийных разведений химерного анти-НМ 1.24 антитела или мышиного анти-НМ 1.24 антитела, полученного аффинной очисткой, добавляют в каждую лунку, и одновременно добавляют 50 мкл 2 мкг/мл биотинилированного мышиного анти-НМ 1.24 антитела, инкубируют при комнатной температуре в течение 2 ч, а затем добавляют меченный пероксидазой стрептавидин (изготовитель DACO). После инкубирования при комнатной температуре в течение 1 ч с последующей промывкой добавляют раствор субстрата. После того, как реакцию останавливают, добавляя 50 мкл 6 н. серной кислоты, измеряют поглощение на 490 нм, используя ридер MICROPLATE READER Model 3550 (производство BioRad).
Представленный на фиг. 2 результат, свидетельствует, что химерное анти-НМ 1.24 антитело обладает активностью ингибирования связывания с биотинилированным мышиным анти-НМ 1.24 антителом, идентичной активности мышиного анти-НМ 1.24 антитела. Это указывает на то, что химерное антитело имеет тот же самый V-участок, что и мышиное анти-НМ 1.24 антитело.
Пример 8. Измерение АРСС-активности химерного анти-НМ 1.24 антитела
ADCC (клеточная цитотоксичность, зависящая от антитела) активность измеряют по способу, изложенному в Current Protocols in Immunology, Chapter 7. Immunologic Studies in humans, Editor. John E, Coligan et al., John Wiley & Sons, Inc., 1993.
1. Получение эффекторных клеток
Моноциты выделяют из периферической крови или костного мозга здоровых людей и пациентов с множественной миеломой путем центрифугирования в градиенте плотности. Так, равные количества PBS(-) добавляют к периферической крови или костному мозгу здоровых людей и пациентов с множественной миеломой, которые помещены на Ficoll (изготовитель Pharmacia) Conrey (изготовитель Daiichi Pharmaceutical Co. Ltd.) (удельная плотность 1,077), и центрифугируют при 400 g в течение 30 мин. Слой моноцитов собирают, дважды промывают RPMI 1640 (изготовитель Sigma), дополненной 10% околоплодной сывороткой теленка (изготовитель Witaker), и приготавливают при плотности клеток 5•106/мл в той же самой культуральной жидкости.
2. Получение клеток-мишеней
В клеточную линию миеломы человека RPMI 8226 (АТСС CCL 155) вводят радиометку путем инкубирования в RPMI 1640 (изготовитель Sigma), дополненной 10% околоплодной сывороткой теленка (изготовитель Witaker), вместе с 0,1 мКи 51Сr-натрий хроматом при 37oС в течение 60 мин. После введения радиометки клетки трижды промывают сбалансированным раствором Хэнкса (Hanks) (HBSS) и доводят до концентрации 2•105/мл.
3. ADCC анализ
В 96-луночный планшет с U-образным дном (изготовитель Corning) помещают 50 мкл 2•105 клеток-мишеней/мл, 1 мкг/мл аффинно очищенного химерного анти-НМ 1.24 антитела и мышиного анти-НМ 1.24 антитело, или контрольный человеческий IgG (изготовитель Serotec), и реакцию ведут при 4oС в течение 15 мин.
Затем к этому добавляют 100 мкл 5•106 эффекторных клеток/мл, и культивируют в инкубаторе с СО2 в течение 4 ч, когда отношение (Е:Т) эффекторных клеток (Е) к мишеневым клеткам (Т) устанавливается равным 0:1, 5:1, 20:1 или 50:1.
100 мкл надосадочной жидкости отбирают и радиоактивность, испускаемую культуральной надосадочной жидкостью, измеряют с помощью счетчика гамма-излучения (ARC361, изготовитель Aloka). Для измерения максимальной радиоактивности используют 1% NP-40 (изготовитель BRL). Цитотоксичность (%) рассчитывают по формуле (А-С)/(В-С) х 100, где А представляет радиоактивность (имп/мин), испускаемую в присутствии антитела, В представляет радиоактивность (имп/мин), испускаемую за счет NP-40, а С представляет радиоактивность (имп/мин), испускаемую культуральной жидкостью без антитела.
Как представлено на фиг.3, если химерное анти-НМ 1.24 антитело добавляют по сравнению с контрольным IgG1, цитотоксичность возрастает с повышением отношения Е:Т, что указывает на то, что это химерное анти-НМ 1.24 антитело обладает ADCC-активностью. Кроме того, так как цитотоксичность не наблюдается, даже если добавляют мышиное анти-НМ 1.24 антитело, было показано, что Fc-часть человеческого антитела необходима для достижения ADCC-активности, если эффекторными клетками являются клетки, полученные от человека.
Пример 9. Конструирование реконструированного человеческого анти-НМ 1.24 антитела
1. Создание V-участка реконструированного человеческого анти-НМ 1.24 антитела
Для конструирования реконструированного человеческого антитела, в которое был трансплантирован CDR мышиного моноклонального антитела, предпочтительно, чтобы существовала высокая гомологичность между FR мышиного антитела и FR человеческого антитела. Так, V-участки L-цепи и Н-цепи мышиного анти-НМ 1.24 антитела сравнивают с V-участками всех известных антител, структура которых была установлена, используя банк данных Protein Data Bank.
V-участок L-цепи мышиного анти-НМ 1.24 антитела наиболее близок к V-участку консенсусной последовательности подгруппы IV (HSGIV) человеческой L-цепи с гомологичностью 66,4%. С другой стороны, было показано, что он гомологичен на 56,9%, 55,8% и 61,5% с HSGI, HSGII и HSGIII соответственно.
Если V-участок L-цепи мышиного анти-НМ 1.24 антитела сравнивают с V-участком L-цепи известных человеческих антител, было показано, что существует гомологичность 67,0% с V-участком REI человеческой L-цепи, одного из подгруппы I V-участка человеческой L-цепи. Поэтому FR REI используют в качестве исходного материала для конструирования V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела.
Создают вариант а V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела. В этом варианте человеческий FR делают идентичным FR на основе REI, присутствующему в реконструированном человеческом САМРАТН-1Н антителе (см. Riechmann, L. et al., Nature 322, 21-25 (1988), FR, содержащийся в варианте а реконструированного человеческого РМ-1, описанного в публикации международной заявки WO 92-19759), и мышиный CDR делают идентичным CDR в V-участке L-цепи мышиного анти-НМ 1.24 антитела.
V-участок Н-цепи мышиного анти-НМ 1.24 антитела наиболее близок к V-участку консенсусной последовательности HSGI человеческой Н-цепи с гомологичностью 54,7%. С другой стороны, было показано, что он гомологичен на 34,6%, и 48,1% с HSGII и HSGIII соответственно. Если V-участок Н-цепи мышиного анти-НМ 1.24 антитела сравнивают с V-участком Н-цепи известных человеческих антител, то FR1-FR3 оказываются наиболее близкими к V-участку Н-цепи человеческого антитела HG3, одного из подгруппы I V-участка человеческой Н-цепи (Rechavi, G. et al., Proc. Natl. Acad. Sci. USA, 80, 855-859) с гомологичностью 67,3%.
Поэтому FR человеческого антитела HG3 используют в качестве исходного материала для конструирования V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела. Однако так как аминокислотная последовательность FR4 человеческого HG3 не была описана, используют аминокислотную последовательность FR4 человеческого антитела JH6 (Ravetch, J.V. et al., Cell, 27, 583-591), которая демонстрирует наивысшую гомологичность с FR4 Н-цепи мышиного анти-НМ 1.24 антитела. FR4 из JH6 имеет ту же самую аминокислотную последовательность, что и FR4 Н-цепи мышиного анти-НМ 1.24 антитела, за исключением одной аминокислоты.
В первом варианте а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела FR1-FR3 делают идентичными FR1-FR3 человеческого HG3, и CDR делают идентичным CDR V-участка Н-цепи мышиного анти-НМ 1.24 антитела, за исключением того, что аминокислоты в положении 30 в человеческом FR1 и в положении 71 в человеческом FR3 делают идентичными аминокислотам в мышином анти-НМ 1.24 антителе.
2. Конструирование V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела
L-цепь реконструированного человеческого анти-НМ 1.24 антитела конструируют трансплантацией CDR способом ПЦР. Этот способ представлен на фиг.4. Восемь ПЦР-праймеров используют для конструирования реконструированного человеческого анти-НМ 1.24 антитела (вариант а), содержащего FR, полученный из человеческого антитела REI. Внешние праймеры А (Последовательность ИД 47) и Н (Последовательность ИД 48) конструируют для гибридизации с последовательностью ДНК экспрессирующего вектора HEF-VL-gκ.
Праймеры трансплантации CDR L1S (Последовательность ИД 49), L2S (Последовательность ИД 50) и L3S (Последовательность ИД 51) имеют смысловую последовательность ДНК. Праймеры трансплантации CDR L1A (Последовательность ИД 52), L2A (Последовательность ИД 53) и L3A (Последовательность ИД 54) имеют антисмысловую последовательность ДНК, причем каждый содержит последовательность ДНК (20-23 п.н.), комплементарную последовательности ДНК у 5'-конца праймеров L1S, L2S и L3S соответственно.
На первой стадии ПЦР проводят четыре реакции A-L1A, L1S-L2A, L2S-L3A и L3S-H для очистки продукта ПЦР. Четырем продуктам из первой ПЦР дают возможность осуществить сборку друг с другом за счет их собственной комплементарности (публикация международной заявки WO 92-19759). Затем добавляют внешние праймеры А и Н для амплификации полной длины ДНК, кодирующей V-участок L-цепи реконструированного человеческого анти-НМ 1.24 антитела, (вторая ПЦР). В вышеуказанной ПЦР плазмиду HEF-RVL-M21a (публикация международной заявки WO 95-14041), кодирующую вариант а V-участка L-цепи реконструированного человеческого ONS-M21 антитела на основе FR, полученной из человеческого антитела REI, используют в качестве матрицы.
На первой стадии ПЦР используют ПЦР-смесь, содержащую 10 мМ Трис-НС1 (рН 8,3), 50 мМ КС1, 0,1 мМ дНТФ, 1,5 мМ MgCl2, 100 нг матричной ДНК, 100 пмоль каждого праймера и 5 Ед. Ampli Taq. Каждую ампулу ПЦР покрывают 50 мкл минерального масла. Затем после первой денатурации в результате нагревания при 94oС, проводят цикл реакций при 94oС 1 мин, при 55oС 1 мин и при 72oС 1 мин, а затем инкубируют при 72oС в течение 10 мин.
Продукты ПЦР A-L1A (215 п.н.), L1S-L2A (98 п.н.), L2S-L3A (140 п.н.) и L3S-H (151 п.н.) очищают, используя 1,5% агарозный гель с низкой температурой плавления, и осуществляют сборку во второй ПЦР. Во второй ПЦР 98 мкл ПЦР-смеси, содержащей по 1 мкг каждого из продуктов первой стадии ПЦР и 5 Ед. Ampli Taq, инкубируют в течение 2 циклов по 94oС 2 мин, 55oС 2 мин и 72oС 2 мин, а затем добавляют по 100 пмоль каждого из внешних праймеров (А и Н). ПЦР-ампулы покрывают 50 мкл минерального масла и осуществляют 30 циклов ПЦР в тех же условиях, что и раньше.
Фрагмент ДНК в 516 п.н., полученный во второй ПЦР, очищают, используя 1,5% агарозный гель с низкой температурой плавления, расщепляют BamHI и HindIII, и полученные таким образом фрагменты клонируют в HEF экспрессирующий вектор HEF-VL-gκ. После определения последовательности ДНК плазмиду, содержащую фрагмент ДНК с правильной аминокислотной последовательностью V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела называют плазмидой HEF-RVLa-АНМ-gκ. Аминокислотная последовательность и нуклеотидная последовательность V-участка L-цепи, содержащегося в этой плазмиде HEF-RVLa-AHM-gκ представлены в Последовательности ИД 9.
Вариант b V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела конструируют с помощью мутагенеза, используя ПЦР. Праймеры мутагенеза FTY-1 (Последовательность ИД 55) и FTY-2 (Последовательность ИД 56) сконструированы таким образом, чтобы подвергнуть мутации фенилаланин в положении 71, заменив его на тирозин.
После того, как вышеуказанные праймеры амплифицируют, используя плазмиду HEF-RVLa-AHM-gκ в качестве матрицы, конечный продукт очищают и расщепляют BamHI и HindIII. Полученные таким образом фрагменты ДНК клонируют в HEF экспрессирующий вектор HEF-VL-gκ, получая плазмиду HEF-RVLb-AHM-gκ. Аминокислотная последовательность и нуклеотидная последовательность V-участка L-цепи, содержащегося в этой плазмиде HEF-RVLb-AHM-gκ, представлены в Последовательности ИД 10.
3. Конструирование V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела
3-1. Конструирование вариантов а-е V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела
ДНК, кодирующую V-участок Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, конструируют следующим образом. Связывая последовательность ДНК, кодирующую FR1 до 3 человеческого антитела HG3, и FG4 человеческого антитела JH6 с последовательностью ДНК, кодирующей CDR V-участка Н-цепи мышиного анти-НМ 1.24 антитела, конструируют полной длины ДНК, кодирующую V-участок Н-цепи реконструированного человеческого анти-НМ 1.24 антитела.
Затем к 5'-концу и 3'-концу этой последовательности ДНК присоединяют HindIII распознающий сайт/KOZAK консенсусную последовательность и BamHI распознающий сайт/сплайсинговую донорную последовательность, соответственно, таким образом, чтобы обеспечить встраивание HEF экспрессирующего вектора.
Сконструированную таким образом последовательность ДНК разделяют на 4 олигонуклеотида. Затем олигонуклеотиды, которые потенциально затрудняют сборку этих олигонуклеотидов, подвергают компьютерному анализу в отношении вторичной структуры. Последовательности четырех олигонуклеотидов RVH1-RVH4 представлены в Последовательностях ИД 57-60. Эти олигонуклеотиды имеют длину от 119 до 144 оснований и содержат 25-26 п.н. перекрывающиеся участки. Среди этих олигонуклеотидов, RVH2 (Последовательность ИД 58) и RVH4 (Последовательность ИД 60) имеют смысловую последовательность ДНК, a RVH1 (Последовательность ИД 57) и RVH3 (Последовательность ИД 59) имеют антисмысловую последовательность. Способ сборки этих четырех олигонуклеотидов с помощью ПЦР представлен на фиг.5.
ПЦР-смесь (98 мкл), содержащую 100 нг каждого из четырех олигонуклеотидов и 5 Ед. Ampli Taq, вначале денатурируют, нагревая при 94oС в течение 2 мин, а затем осуществляют 2 цикла инкубирования, включающих 94oС в течение 2 мин, 55oС в течение 2 мин и 72oС в течение 2 мин. После того, как 100 пмоль каждого из RHP1 (Последовательность ИД 61) и RHP2 (Последовательность ИД 62) добавляют в качестве внешних праймеров, ПЦР ампулу покрывают 50 мл минерального масла. Затем вначале осуществляют денатурацию, нагревая при 94oС в течение 1 мин, а затем проводят 38 циклов, состоящих из 94oC в течение 1 мин, 55oС в течение 1 мин и 72oС в течение 1 мин, а затем инкубируют при 72oС в течение 10 мин.
Фрагмент ДНК 438 п.н. очищают, используя 1,5% агарозный гель с низкой температурой плавления, расщепляют BamHI и HindIII, а затем клонируют в HEF экспрессирующий вектор HEF-VH-gγ1. После определения нуклеотидной последовательности плазмиду, которая содержит фрагмент ДНК, который кодирует аминокислотную последовательность правильного V-участка Н-цепи, называют HEF-RVHa-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHa-AHM-gγ1, представлены в Последовательности ИД 11.
Каждый из вариантов b, с, d, и e V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела конструируют следующим образом.
Используя праймеры мутагенеза BS (Последовательность ИД 63) и ВА (Последовательность ИД 64), сконструированные для осуществления мутации аргинина в положении 66 на лизин, а в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, методом ПЦР вариант b амплифицируют, получая плазмиду HEF-RVHb-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащиеся в этой плазмиде HEF-RVHb-AHM-gγ1, представлены в Последовательности ИД 12.
Используя праймеры мутагенеза CS (Последовательность ИД 65) и СА (Последовательность ИД 66), сконструированные для осуществления мутации треонина в положении 73 на лизин, а в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, методом ПЦР вариант с амплифицируют, получая плазмиду HEF-RVHc-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHc-AHM-gγ1, представлены в Последовательности ИД 13.
Используя праймеры мутагенеза DS (Последовательность ИД 67) и DA (Последовательность ИД 68), сконструированные для осуществления мутации аргинина в положении 66 на лизин и треонина в положении 73 на лизин, а в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, методом ПЦР вариант d амплифицируют, получая плазмиду HEF-RVHd-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHd-AHM-gγ1, представлены в Последовательности ИД 14.
Используя праймеры мутагенеза ES (Последовательность ИД 69) и ЕА (Последовательность ИД 70), сконструированные для осуществления мутации валина в положении 67 на аланин и метионина в положении 69 на лейцин, а в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, вариант e амплифицируют, получая плазмиду HEF-RVHe-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHe-AHM-gγ1, представлены в Последовательности ИД 15.
3-2. Конструирование гибридного V-участка Н-цепи
Конструируют два гибридных V-участка Н-цепи. Один является мышиным/человеческим гибридным анти-НМ 1.24 антителом, в котором аминокислотные последовательности FR1 и FR2 получены из мышиного анти-НМ 1.24 антитела, а последовательности FR3 и FR4 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, а другой является человеческим/мышиным гибридным анти-НМ 1.24 антителом, в котором аминокислотные последовательности FR1 и FR2 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, а последовательности FR3 и FR4 получены из мышиного анти-НМ 1.24 антитела. Все аминокислотные последовательности CDR участков получены из мышиного анти-НМ 1.24 антитела.
Методом ПЦР конструируют два гибридных V-участка Н-цепи. Этот метод схематически представлен на фиг.6 и 7. Для конструирования двух гибридных V-участков Н-цепи используют четыре праймера. Внешние праймеры а (Последовательность ИД 71) и h (Последовательность ИД 72) сконструированы таким образом, чтобы они гибридизовались с последовательностью ДНК HEF экспрессирующего вектора HEF-VH-gγ1. Праймер HYS гибридной конструкции Н-цепи (Последовательность ИД 73) сконструирован таким образом, что содержит смысловую последовательность ДНК, а гибридный праймер Н-цепи HYA (Последовательность ИД 74) имеет антисмысловую последовательность ДНК, так, что эти последовательности ДНК комплементарны друг другу.
Для конструирования гибридного V-участка Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из мышиного анти-НМ 1.24 антитела, а последовательности FR3 и FR4 из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, осуществляют ПЦР, используя в качестве матрицы плазмиду НЕF-1.24Н-gγ1, внешний праймер а, и гибридный праймер Н-цепи HYA, и ПЦР, используя в качестве матрицы плазмиду HEF-RVHa-AHM-gγ1, и гибридный праймер Н-цепи HYS (Последовательность ИД 73), и внешний праймер h (Последовательность ИД 72), осуществляют на первой стадии ПЦР, и каждый продукт ПЦР очищают. Двум продуктам первой ПЦР дают возможность осуществить сборку за счет собственной комплементарности (публикация международной заявки WO 92-19759).
Затем, добавляя внешние праймеры а (Последовательность ИД 71) и h (Последовательность ИД 72) на второй стадии ПЦР амплифицируют полной длины ДНК, кодирующую гибридный V-участок Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из мышиного анти-НМ 1.24 антитела, а последовательности FR3 и FR4 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела.
Для конструирования гибридного V-участка Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, а последовательности FR3 и FR4 из мышиного анти-НМ 1.24 антитела, проводят ПЦР, используя в качестве матрицы плазмиду HEF-RVHa-AHM-gγ1, внешний праймер а, и гибридный праймер HYA Н-цепи, и ПЦР, используя плазмиду HEF-1.24H-gγ1 в качестве матрицы, гибридный праймер HYS Н-цепи, и внешний праймер h, осуществляют на первой стадии ПЦР, и каждый продукт ПЦР очищают. Двум очищенным продуктам первой ПЦР дают возможность осуществить сборку за счет собственной комплементарности (публикация международной заявки WO 92-19759).
Затем, добавляя внешние праймеры а и h, полной длины ДНК, кодирующую гибридный V-участок Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, а последовательности FR3 и FR4 из мышиного анти-НМ 1.24 антитела, амплифицируют на второй стадии ПЦР.
Способы осуществления первой ПЦР, очистки продуктов ПЦР, сборки, осуществления второй ПЦР и клонирования в HEF вектор HEF-VH-gγ1 осуществляют в соответствии со способами, представленными в примере 9. "Конструирование V-участка L-цепи реконструированного человеческого анти-НМ 1.24 антитела".
После определения последовательности ДНК плазмиду, которая содержит фрагмент ДНК, кодирующий правильную аминокислотную последовательность гибридного V-участка Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из мышиного анти-НМ 1.24 антитела, а последовательности FR3 и FR4 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, называют HEF-MH-RVH-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-MH-RVH-AHM-gγ1, представлены в Последовательности ИД 75. Также плазмиду, которая содержит правильную аминокислотную последовательность гибридного V-участка Н-цепи, в котором аминокислотные последовательности FR1 и FR2 получены из варианта а V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, а последовательности FR3 и FR4 из мышиного анти-НМ 1.24 антитела, называют HEF-HM-RVH-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-MH-RVH-AHM-gγ1, представлены в Последовательности ИД 76.
3-3. Конструирование вариантов f-s V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела
Каждый из вариантов f, g, h, i, j, k, 1, m, n, o, p, q, r, и s V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела конструируют следующим образом.
Используя праймеры мутагенеза FS (Последовательность ИД 78) и FA (Последовательность ИД 79), сконструированные для осуществления мутации треонина в положении 75 на серин и валина в положении 78 на аланин, а в качестве матричной ДНК плазмиду HEF-RVHe-AHM-gγ1, с помощью ПЦР вариант f амплифицируют, получая плазмиду HEF-RVHf-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHf-AHM-gγ1, представлены в Последовательности ИД 16.
Используя праймеры мутагенеза GS (Последовательность ИД 80) и GA (Последовательность ИД 81), сконструированные для осуществления мутации аланина в положении 40 на аргинин, в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, вариант g амплифицируют, получая плазмиду HEF-RVHg-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHg-AHM-gγ1, представлены в Последовательности ИД 17.
Используя праймеры мутагенеза FS (Последовательность ИД 78) и FA (Последовательность ИД 79), а в качестве матричной ДНК плазмиду HEF-RVHb-AHM-gγ1, вариант h амплифицируют, получая плазмиду HEF-RVHh-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHh-AHM-gγ1, представлены в Последовательности ИД 18.
Используя праймеры мутагенеза IS (Последовательность ИД 82) и IA (Последовательность ИД 83), сконструированные для осуществления мутации аргинина в положении 83 на аланин и серина в положении 84 на фенилаланин, а в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγ1, вариант i амплифицируют, получая плазмиду HEF-RVHi-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHi-AHM-gγ1, представлены в Последовательности ИД 19.
Используя праймеры мутагенеза JS (Последовательность ИД 84) и JA (Последовательность ИД 85), сконструированные для осуществления мутации аргинина в положении 66 на лизин, а в качестве матричной ДНК плаэмиду HEF-RVHf-AHM-gγ1, вариант j амплифицируют, получая плазмиду HEF-RVHj-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHj-AHM-gγ1, представлены в Последовательности ИД 20.
Используя праймеры мутагенеза КS (Последовательность ИД 86) и КА (Последовательность ИД 87), сконструированные для осуществления мутации глутаминовой кислоты в положении 81 на глутамин, а в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγ1, вариант k амплифицируют, получая плазмиду HEF-RVHk-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHk-AHM-gγ1, представлены в Последовательности ИД 21.
Используя праймеры мутагенеза LS (Последовательность ИД 88) и LA (Последовательность ИД 89), сконструированные для осуществления мутации глутаминовой кислоты в положении 81 на глутамин и серина в положении 82В на изолейцин, а в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγ1, вариант 1 амплифицируют, получая плазмиду HEF-RVHI-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVH1-АНМ-gγ1, представлены в Последовательности ИД 22.
Используя праймеры мутагенеза MS (Последовательность ИД 90) и MА (Последовательность ИД 91), сконструированные для осуществления мутации глутаминовой кислоты в положении 81 на глутамин, серина в положении 82B на изолейцин, и треонина в положении 87 на серин, а в качестве матричной ДНК плазмиду HEF-RVHh-AHM вариант m амплифицируют, получая плазмиду HEF-RVHm-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHm-AHM-gγ1, представлены в Последовательности ИД 23.
вариант m амплифицируют, получая плазмиду HEF-RVHm-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHm-AHM-gγ1, представлены в Последовательности ИД 23.
Используя праймеры мутагенеза NS (Последовательность ИД 92) и NA (Последовательность ИД 93), сконструированные для осуществления мутации серина в положении 82В на изолейцин, а в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγ1, вариант n амплифицируют, получая плазмиду HEF-RVHn-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHn-AHM-gγ1, представлены в Последовательности ИД 24.
Используя праймеры мутагенеза OS (Последовательность ИД 94) и ОА (Последовательность ИД 95), сконструированные для осуществления мутации треонина в положении 87 на серии, а в качестве матричной ДНК плазмиду HEF-RVHh-AHM-gγ1, вариант о амплифицируют, получая плазмиду HEF-RVHo-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHo-AHM-gγ1, представлены в Последовательности ИД 25.
Используя праймеры мутагенеза PS (Последовательность ИД 96) и РА (Последовательность ИД 97), сконструированные для осуществления мутации валина в положении 78 на аланин, а в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, вариант p амплифицируют методом ПЦР, получая плазмиду HEF-RVHp-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHp-AHM-gγ1, представлены в Последовательности ИД 26.
Используя праймеры мутагенеза QS (Последовательность ИД 98) и QA (Последовательность ИД 99), сконструированные для осуществления мутации треонина в положении 75 на серин, а в качестве матричной ДНК плазмиду HEF-RVHa-AHM-gγ1, вариант q амплифицируют методом ПЦР, получая плазмиду HEF-RVHq-AHM-gγ1. Аминокислотная последовательность и нуклеотидная
последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHq-AHM-gγ1, представлены в Последовательности ИД 27.
Используя праймеры мутагенеза CS (Последовательность ИД 65) и СА (Последовательность ИД 66), а в качестве матричной ДНК плазмиду HEF-RVHp-AHM-gγ1, вариант r амплифициру-ют методом ПЦР, получая плазмиду HEF-RVHr-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHr-AHM-gγ1, представлены в Последовательности ИД 28.
Вариант s V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела конструируют с помощью мутагенеза, используя ПЦР. Праймеры мутагененза SS (Последовательность ИД 100) и SA (Последовательность ИД 101) конструируют для осуществления мутации метионина в положении 69 на изолейцин.
После того, как вышеуказанный праймер амплифицируют, используя в качестве матрицы плазмиду HEF-RVHr-AHM-gγ1, конечный продукт очищают, расщепляют BamHI и HindIII, и полученный фрагмент ДНК клонируют в HEF экспрессирующий вектор HEF-VH-gγ1, получая плазмиду HEF-RVHs-AHM-gγ1. Аминокислотная последовательность и нуклеотидная последовательность V-участка Н-цепи, содержащегося в этой плазмиде HEF-RVHs-AHM-gγ1, представлены в Последовательности ИД 102.
Участки, кодирующие вариабельный участок каждой из вышеуказанных плазмид HEF-RVLa-AHM-gκ и и HEF-RVHr-AHM-gγ1, расщепляют для получения рестрикционных фрагментов ферментами рестрикции HindIII и BamHI.
Их встраивают в HindIII и BamHI сайты плазмидного вектора pUC19. Каждую плазмиду называют pUC19-RVLa-AHM-gκ и pUCl 9-RVHr-AHM-gγ1.
Eschericia coli, которые содержат каждую из плазмид pUC19-RVLa-AHM-gκ и pUC19-RVHr-AHM-gγ1, называют Eschericia coli DH5α (pUC19-RVLa-AHM-gκ) и Eschericia coli DH5α (pUC19-RVHr-AHM-gγ1), соответственно, и они были интернационально депонированы 29 августа 1996 г. the National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, MITI (Higashi 1-Chome 1-3, Tsukuba city, Ibalaki prefecture, Japan) под регистрационными номерами FERM BP-5645 и FERM ВР-5643, соответственно, в соответствии с Будапештским соглашением.
Участки, кодирующие вариабельный участок вышеуказанной плазмиды HEF-RVHs-AHM-gγ1, расщепляют для получения рестрикционных фрагментов ферментами рестрикции HindIII и BamHI. Их встраивают в HindIII и BamHI сайты плазмидного вектора pUC19. Полученную плазмиду называют pUC19-RVHs-AHM-gγ1.
Eschericia coli, которая содержит плазмиду pUC19-RVHs-AHM-gγ1, называют Eschericia coli DH5α (pUC19-RVHs-AHM-gγ1), и она была интернационально депонирована 29 сентября 1997 г., the National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, MITI (Higashi 1-Chome 1-3, Tsukuba city, Ibalaki prefecture, Japan) под регистрационным номером FERM BP-6127, в соответствии с Будапештским соглашением.
4. Конструирование реконструированного человеческого анти-НМ 1.24 антитела, химерного анти-НМ 1.24 антитела и антитела с гибридной Н-цепыо
Для оценки каждой из цепей реконструированного человеческого анти-НМ 1.24 антитела предоставляют возможность экспрессироваться реконструированному человеческому анти-НМ 1.24 антителу и химерному анти-НМ 1.24 антителу в качестве позитивного контроля. При конструировании каждого из вариантов b и далее V-участка Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, Н-цепи гибридного антитела предоставляют возможность экспрессироваться для того, чтобы определить, какую аминокислотную последовательность в FR следует заместить. Кроме того, ее экспрессируют в сочетании с химерной Н-цепью для того, чтобы оценить вариант а L-цепи реконструированного человеческого анти-НМ 1.24 антитела.
4-1. Экспрессия реконструированного человеческого анти-НМ 1.24 антитела
10 мкг каждого из экспрессирующих векторов (HEF-RVHa-AHM-gγ1 до HEF-RVHr-AHM-gγ1 для Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, и экспрессирующего вектора (НЕF-RVLa-АНM-gκ или HEF-RVLb-АНM-gκ для L-цепи реконструированного человеческого анти-НМ 1.24 антитела котрансформируют в COS-7 клетки с помощью электропорации, используя прибор Gene Pulser (изготовитель BioRad). Каждую ДНК (10 мкг) добавляют к 0,8 мл аликвотам 1•107 кл/мл в PBS, и обрабатывают импульсами при 1500 В и емкости 25 мкФ.
После 10 минутного восстановительного периода при комнатной температуре подвергшиеся электропорации клетки добавляют к 30 мл DMEM культуральной среды (изготовитель GIBCO), содержащей 10% околоплодной сыворотки теленка без гамма-глобулина. После 72 ч инкубирования в инкубаторе СО2 BNA120D (изготовитель TABAI) в условиях 37oС и 5% СO2 надосадочную культуральную жидкость собирают, осколки клеток удаляют центрифугированием при 1000 об/мин в течение 5 мин в центрифуге 15PR-22 (изготовитель HITACHI), снабженной центрифужным ротором 03 (изготовитель HITACHI), и микроконцентратором (Centricon, изготовитель Amicon), и подвергают ультрафильтрации, используя центрифугу J2-21 (изготовитель BECKMAN), снабженную центрифужным ротором JA-20.1 (изготовитель BECKMAN), и используют для клеточного ELISA.
Экспрессия реконструированного человеческого анти-НМ 1.24 антитела (2)
10 мкг каждого из экспрессирующих векторов (HEF-RVHs-AHM-gγ1) для варианта "s" Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, и экспрессирующий вектор (HEF-RVLa-AHM-gκ) для L-цепи реконструированного человеческого анти-НМ 1.24 антитела котрансформируют в COS-7 клетки с помощью электропорации, используя прибор Gene Pulser (изготовитель BioRad). Каждую ДНК (10 мкг) добавляют к 0,8 мл аликвотам 1•107 кл/мл в PBS и обрабатывают импульсами при 1500 В и емкости 25 мкФ.
После 10 минутного восстановительного периода при комнатной температуре, подвергшиеся электропорации клетки добавляют к 30 мл DMEM культуральной среды (изготовитель GIBCO), содержащей 10% околоплодной сыворотки теленка без гамма-глобулина. После 72 ч инкубирования в инкубаторе СО2 BNA120D (изготовитель TABAI) в условиях 37oС и 5% СО2 надосадочную культуральную жидкость собирают, осколки клеток удаляют центрифугированием при 1000 об/мин в течение 5 мин в центрифуге 05PR-22 (изготовитель HITACHI), снабженной центрифужным ротором 03 (изготовитель HITACHI), и микроконцентратором (Centricon, изготовитель Amicon), и концентрируют с помощью ультрафильтрации, используя центрифугу J2-21 (изготовитель BECKMAN), снабженную центрифужным ротором JA-20.1 (изготовитель BECKMAN), и стерилизуют фильтрованием, используя фильтр Millex GV13mm (изготовитель Millipore), и используют для клеточного ELISA.
4-2. Экспрессия химерного анти-НМ 1.24 антитела
Используя 10 мкг каждого из экспресирующих векторов НЕF-1.24Н-gγ1 для Н-цепи химерного анти-НМ 1.24 антитела и экспрессирующий вектор HEF-1.24L-gκ для L-цепи химерного анти-НМ 1.24 антитела, химерное анти-НМ 1.24 антитело, которое нужно использовать для клеточного ELISA, приготавливают в соответствии с вышеуказанным способом для экспрессии реконструированного человеческого анти-НМ 1.24 антитела.
4-3. Экспрессия анти-НМ 1.24 антитела, включающего вариант а гуманизированной L-цепи и химерную Н-цепь
Используя 10 мкг каждого из экспресирующих векторов HEF-1.24H-gγ1 для Н-цепи химерного анти-НМ 1.24 антитела и экспрессирующий вектор HEF-RVLa-AHM-gκ для варианта а L-цепи реконструированного человеческого анти-НМ 1.24 антитела, анти-НМ 1.24 антитело, которое включает вариант а гуманизированной L-цепи и химерную Н-цепь, которое нужно использовать для клеточного ELISA, получают в соответствии с вышеуказанным способом для экспрессии реконструированного человеческого анти-НМ 1.24 антитела.
4-4. Экспрессия антитела с гибридной Н-цепью
Используя 10 мкг каждого из экспрессирующих векторов HEF-MH-RVH-AHM-gγ1 или HEF-HM-RVH-AHM-gγ1) для V-участка гибридной Н-цепи и экспрессирующий вектор HEF-RVLa-AHM-gκ для L-цепи реконструированного человеческого анти-НМ 1.24 антитела, антитело с гибридной Н-цепью, которое нужно использовать для клеточного ELISA, получают в соответствии с вышеуказанным способом для экспрессии реконструированного человеческого анти-НМ 1.24 антитела.
4-5. Определение концентрации антител
Концентрацию полученных антител определяют с помощью ELISA. Каждую из лунок 96-луночного планшета ELISA (Махisorp, изготовитель NUNC) иммобилизуют, добавляя 100 мкг козьего античеловеческий IgG антитела (изготовитель BIO SOURCE), приготовленного в концентрации 1 мкг/мл в покрывающем буфере (0,1 М NаНСО3, 0,02% NaN3, pH 9,6) и инкубируют при комнатной температуре в течение 1 ч. После блокирования 100 мкл буфера для разбавления (50 мМ Трис-HCl, 1 мМ MgCl2, 0,15 М NaC1, 0,05% Tween 20, 0,02% NaN3, 1% бычьего сывороточного альбумина (BSA), pH 8,1), 100 мкл каждого из разбавлений культуральной надосадочной жидкости клеток COS-7 секретирующих реконструированное человеческое анти-НМ 1.24 антитело, химерное анти-НМ 1.24 антитело, или антитело с гибридной Н-цепью, которые были сконцентрированы ультрафильтрацией, добавляют в каждую лунку и инкубируют при комнатной температуре в течение одного часа. Затем после промывки добавляют 100 мкл козьего антитела против человеческого IgG, меченного щелочной фосфатазой (изготовитель DACO).
После инкубирования при комнатной температуре в течение 1 ч и промывки добавляют 100 мкл субстратного раствора в концентрации 1 мкг/мл (Sigma 104, п-нитрофенилфосфат, SIGMA), растворенного в субстратном буфере (50 мМ NаНСО3, 10 мМ MgCl2, pH 9,8), а затем измеряют поглощение на 405 нм, используя ридер MICROPLATE READER Model 3550 (изготовитель BioRad). В качестве стандарта для измерения концентраций используют человеческий IgG1κ (изготовитель The Binding Site).
5. Создание клеточной линии СНО, которая стабильно продуцирует реконструированное человеческое анти-НМ 1.24 антитело.
5-1. Конструирование экспрессирующего вектора для Н-цепи реконструированного человеческого анти-НМ 1.24 антитела
Расщепляя плазмиду HEF-RVHr-AHM-gγ1 ферментами рестрикции Pvul и BamHI, фрагмент около 2,8 т.п.н., содержащий ДНК, кодирующую промотор EFI и V-участок Н-цепи реконструированного человеческого анти-НМ 1.24 антитела, выделяют, используя 1,5% агарозный гель с низкой температурой плавления. Затем вышеуказанный фрагмент ДНК встраивают в примерно 6 т.п.н. фрагмент, который получен в результате расщепления экспрессирующего вектора, использованного для экспрессирующего вектора человеческой Н-цепи, DHFR-ΔE-RVh-PM1f (публикация международной заявки WO 92-19759), содержащего ген DHFR и ген, кодирующий константный участок человеческой Н-цепи, с помощью Pvul и BamHI для конструирования экспрессирующего вектора DHFR-ΔE-HEF-RVHr-AHM-gγ1 для Н-цепи реконструированного анти-НМ 1.24 антитела.
5-2. Введение гена в клетки СНО
Для создания системы, стабильно продуцирующей реконструированное человеческое анти-НМ 1.24 антитело, гены вышеуказанных экспрессирующих векторов, DHFR-ΔE-HEF-RVHr-AHM-gγ1 и HEF-RVLa-AHM-gκ, которые были линеаризованы в результате расщепления Pvul, одновременно вводят в клетки СНО DXB-11 методом электропорации в условиях, аналогичных вышеуказанным (трансфекция вышеуказанных клеток COS-7).
5-3. Амплификация генов с помощью МТХ
Среди клеток СНО с введенными генами только те клетки СНО, в которые были введены экспрессирующие векторы как L-, так и Н-цепей, могут выживать в α-MEM культуральной среде без нуклеозида (изготовитель GIBCO-BRL), к которой добавлены 500 мкг/мл G418 (изготовитель GIBCO-BRL) и 10% околоплодной сыворотки теленка, и это позволяет произвести отбор. Затем 10 нМ МТХ (изготовитель Sigma) добавляют к вышеуказанной культуральной среде. Среди разрастающихся клонов отбирают те клоны, которые продуцируют реконструированное человеческое анти-НМ 1.24 антитело в больших количествах. В результате получают клон 1, который демонстрирует эффективность продуцирования около 3 мкг/мл реконструированного человеческого анти-НМ 1.24 антитела, и его называют клеточной линией, продуцирующей реконструированное человеческое анти-НМ 1.24 антитело.
5-4. Конструирование реконструированного человеческого анти-НМ 1.24 антитела
Реконструированное анти-НМ 1.24 антитело конструируют следующим образом. Вышеуказанные клетки СНО, которые продуцируют реконструированное человеческое анти-НМ 1.24 антитело, культивируют в течение 10 дней, используя в качестве среды α-MEM культуральную среду без нуклеозида (изготовитель GIBCO-BRL), к которой добавлены 500 мкг/мл G418 (изготовитель GIBCO-BRL) и 10% околоплодной сыворотки теленка, используя СО2 инкубатор BNAS120D (изготовитель TABAI) в условиях 37oС и 5% СO2. На 8 и 10 сутки после начала культивирования культуральную жидкость извлекают, клеточные осколки удаляют центрифугированием в течение 10 мин при 2000 об/мин, используя центрифугу RL-500SP (изготовитель Tomy Seico), снабженную ротором TS-9, а затем стерилизуют на фильтре, используя фильтр, оканчивающийся колбой (изготовитель FALCON) с мембраной с диаметром отверстий 0,45 мкм.
После того, как равные количества PBS(-) добавляют к культуральной жидкости клеток СНО, продуцирующих реконструированное человеческое анти-НМ 1.24 антитело, реконструированное человеческое анти-НМ 1.24 антитело аффинно очищают, используя высокоскоростную систему очистки антител ConSep LC100 (изготовитель MILLIPORE) и колонку Hyper D Protein A (изготовитель Nippon Gaishi), используя PBS(-) в качестве абсорбирующего/промывочного буфера и 0,1 М натрийцитратный буфер (рН 3) в качестве элюирующего буфера в соответствии с прилагаемыми инструкциями. рН элюированных фракций доводят примерно до 7,4, немедленно добавляя 1 М Трис-НС1 (рН 8,0), а затем осуществляют, используя ультрафильтрационный концентратор с центрифугой Centriprep-10 (изготовитель MILLIPORE), концентрирование и замещение на PBS(-), стерилизуют на фильтре, используя мембранный фильтр MILLEX-GV (изготовитель MILLIPORE) с размером пор 0,22 мкм, получая очищенное реконструированное человеческое анти-НМ 1.24 антитело. Концентрацию антител измеряют по поглощению на 280 нм, принимая концентрацию 1 мг/мл соответствующей 1,35 ОП.
Пример 11. Определение активности реконструированного человеческого анти-НМ 1.24 антитела
Проводят оценку активности связывания антигена и активности ингибирования связывания реконструированного человеческого анти-НМ 1.24 антитела.
1. Способ измерения активности связывания антигена и активности ингибирования связывания
1-1. Измерение активности связывания антигена
Активность связывания антигена измеряют с помощью клеточного ELISA, используя клетки WISH. Планшеты клеточного ELISA подготавливают, как указано выше в примере 7.1-2.
После блокирования в каждую лунку добавляют 100 мкл серийных разбавлений реконструированного человеческого анти-НМ 1.24 антитела, которое было получено из концентрата культуральной надосадочной жидкости клеток COS-7, или выделено из культуральной надосадочной жидкости клеток СНО. После 2 ч инкубирования при комнатной температуре и промывки добавляют кроличье антитело против человеческого IgG, меченное пероксидазой (изготовитель DAKO). После инкубирования в течение 1 ч при комнатной температуре и промывки в каждую лунку добавляют 100 мкл субстратного раствора. После инкубирования реакцию останавливают 50 мкл 6 н серной кислоты, и поглощение при 490 нм измеряют, используя ридер MICROPLATE READER Model 3550 (изготовитель BioRad).
1-2. Измерение активности ингибирования связывания
Активность ингибирования связывания мышиным анти-НМ 1.24 антителом, меченным биотином, измеряют с помощью клеточного ELISA, используя клетки WISH. Клеточный ELISA подготавливают как указано выше в примере 7.1-2. После блокирования в каждую лунку добавляют 50 мкл серийных разбавлений реконструированного человеческого анти-НМ 1.24 антитела, которое было получено из концентрата культуральной надосадочной жидкости клеток COS-7, или выделено из культуральной надосадочной жидкости клеток СНО, и одновременно добавляют мышиное анти-НМ 1.24 антитело, меченное биотином. После 2 ч инкубирования при комнатной температуре и промывки, добавляют меченный пероксидазой стрептавидин (изготовитель DAKO). После инкубирования в течение 1 ч при комнатной температуре и промывки в каждую лунку добавляют 100 мкл субстратного раствора. После инкубирования реакцию останавливают 50 мкл 6 н. серной кислоты, и поглощение на 490 нм измеряют, используя ридер MICROPLATE READER Model 3550 (изготовитель BioRad).
2. Оценка реконструированного человеческого анти-НМ 1.24 антитела
2-1. L-цепь
Вариант а L-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивают, как указано для измерения активности связывания антигена. Как представлено на фиг.8, если вариант а L-цепи экспрессируется в сочетании с химерной Н-цепью, она демонстрирует аналогичный уровень активности связывания антигена. Однако с учетом дальнейшего повышения активности и совместимости с Н-цепью был сконструирован вариант b L-цепи. Варианты а и b L-цепи были оценены вместе на предмет активности связывания антигена и активности ингибирования связывания при объединении с вариантами а, b, f, или h Н-цепи. Как видно на фиг.9, 10, 11 и 12 вариант а L-цепи обладает более высокой активностью, нежели вариант b в отношении обеих активностей для всех вариантов a, b, f и h Н-цепи. Поэтому для следующих экспериментов был использован вариант а L-цепи реконструированного человеческого анти-НМ 1.24 антитела.
2-2. Варианты а-е Н-цепи
Варианты а-е Н-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивали в сочетании с вариантом а L-цепи по данным измерения активности связывания антигена и активности ингибирования связывания. Результаты показывают, как видно на фиг.11, 13, 14 и 15, что все варианты оказались слабее в отношении обеих активностей по сравнению с химерным анти-НМ 1.24 антителом, что предполагает необходимость дальнейшего замещения аминокислот.
2-3. Антитело с гибридной Н-цепью
Антитело с гибридной H-цепью оценивают, как было указано для измерения активности связывания антигена. Результаты, как видно на фиг.16, показывают, что человеческое/мышиное гибридное анти-НМ 1.24 антитело демонстрирует активность, аналогичную активности химерного анти-НМ 1.24 антитела в отношении активности связывания антигена, тогда как мышиное/человеческое гибридное анти-НМ 1.24 антитело обладает более слабой активностью, нежели химерное анти-НМ 1.24 антитело. Это указывает на то, что для конструирования реконструированного человеческого анти-НМ 1.24 антитела, обладающего активностью связывания антигена, аналогичной активности химерного анти-НМ 1.24 антитела, необходимо превратить аминокислоты, включенные в FR3 или FR4, среди тех, которые содержатся в V-участке Н-цепи.
2-4. Варианты f-r Н-цепи
Варианты f Н-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивают, как указано для измерения активности связывания антигена. Результат, как представлено на фиг.17, показывает, что его активность связывания антигена снижается по сравнению с активностью химерного анти-НМ, 1.24 антитела, но повышается по сравнению с вышеуказанными вариантами а-с, что предполагает, что любая из четырех аминокислот в положении 67, 69, 75 и 78, которые были вновь превращены в этой версии, ответственна за активность реконструированного человеческого антитела.
Вариант g Н-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивают, как указано для измерения активности связывания антигена. Результат, как представлено на фиг.18 и 19, показывает, что данный вариант демонстрирует уровень активности самое большее аналогичный уровню активности вышеуказанного варианта а, что предполагает, что как показано ранее для Н-цепи человеческого/мышиного гибридного антитела, аминокислота в положении 40, которая была превращена в данном варианте, не ответственна за активность реконструированного человеческого антитела.
Варианты h-j Н-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивают, как указано для измерения активности связывания антигена и активности ингибирования связывания. Результат, как представлено на фиг.20, 21, 22 и 23, указывает, что все варианты были слабее в отношении обеих активностей по сравнению с активностями химерного анти-НМ 1.24 антитела и были аналогичны активностям вышеуказанного варианта f, что предполагает, что аминокислоты в положении 67 и 69 среди четырех вновь измененных аминокислот в варианте f, не ответственны за повышение активности реконструированного человеческого антитела.
Варианты k-p Н-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивают, как указано для измерения активности связывания антигена и активности ингибирования связывания. Результат, как представлено на фиг.24, 25, 26 и 27 указывает, что все варианты были слабее в отношении обеих активностей по сравнению с активностями химерного анти-НМ 1.24 антитела и были аналогичны активностям вышеуказанного варианта h, что предполагает, что аминокислоты в положении 80 и далее после тех, которые были вновь превращены в этих шести вариантах, не ответственны за повышение активности реконструированного человеческого антитела.
Вариант q Н-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивают, как указано для измерения активности связывания антигена и активности ингибирования связывания. Результат, как представлено на фиг.25 и 27, указывает, что данный вариант был слабее в отношении обеих активностей по сравнению с активностями вышеуказанного варианта h или варианта р, и была аналогична активностям вышеуказанного варианта а, что предполагает, что замещение аминокислоты в положении 78 и существенно для повышения активности реконструированного человеческого антитела.
Вариант r Н-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивают указанным выше способом. Результат, как представлено на фиг.15 и 28 указывает, что вариант r обладает уровнями активности связывания антигена и активности ингибирования связывания, аналогичными активностям химерного анти-НМ 1.24 антитела.
Вышеприведенные результаты указывают, что минимальным превращением, необходимым для того, чтобы реконструированное человеческое анти-НМ 1.24 антитело имело бы уровень активности связывания антигена аналогичный уровню активности мышиного анти-НМ 1.24 антитела или химерного анти-НМ 1.24 антитела, является превращение аминокислот в положениях 30, 71 и 78, и кроме того 73.
Активность связывания антигена и активность ингибирования связывания для вариантов а-r Н-цепи реконструированного человеческого анти-НМ 1.24 антитела суммированы в табл. 6.
2-5. Вариант s Н-цепи
Вариант s Н-цепи реконструированного человеческого анти-НМ 1.24 антитела оценивают в комбинации с вышеуказанным вариантом а L-цепи, как указано для измерения активности связывания антигена и активности ингибирования связывания. Результат, как представлено на фиг.29 и 30, показывает, что вариант s обладает активностью связывания антигена и активностью ингибирования связывания, аналогичной активностям варианта r.
Как было указано выше, реконструированное человеческое анти-НМ 1.24 антитело по настоящему изобретению сохраняет способность связывать антиген даже после того, как один или более из аминокислотных остатков был заменен другими аминокислотами. Соответственно, настоящее изобретение включает реконструированное человеческое анти-НМ 1.24 антитело, в котором аминокислотные остатки были заменены другими аминокислотами в вариабельном участке Н-цепи или L-цепи, до тех пор, пока оно сохраняет исходные свойства.
3. Оценка очищенного реконструированного человеческого анти-НМ 1.24 антитела
Очищенное реконструированное человеческое анти-НМ 1.24 антитело оценивают в отношении вышеуказанных активности связывания антигена и активности ингибирования связывания. Результат, как представлено на фиг.31 и 32, показывает, что реконструированное человеческое анти-НМ 1.24 антитело обладает уровнем активности связывания антигена и активности ингибирования связывания аналогичными уровню для химерного анти-НМ 1.24 антитела. Этот факт показывает, что реконструированное человеческое анти-НМ 1.24 антитело обладает уровнем активности связывания антигена таким же, что и мышиное анти-НМ 1.24 антитело.
Пример 12. Противоопухолевое действие химерного анти-НМ 1.24 антитела в отношении миеломы человека в модели на мышах
1. Получение антитела, подлежащего введению
1-1. Получение химерного анти-НМ 1.24 антитела
Очищенное химерное анти-НМ 1.24 антитело, полученное ранее в примере 6, концентрируют, и буферный раствор заменяют PBS(-) используя центрифужный ультрафильтрационный концентратор Centriprep 10 (изготовитель Amicon). Его стерилизуют на фильтре, используя мембранный фильтр MILLEX-GV (изготовитель MILLIPORE) с размером пор 0,22 мкм. Приготавливают концентрацию 200 мкг/мл, используя стерилизованный на фильтре PBS(-), и используют в дальнейших экспериментах. Концентрацию антител измеряют по поглощению на 280 нм, и рассчитывают, принимая оптическую плотность 1.35 ОП, соответствующей 1 мг/мл.
1-2. Очистка контрольного человеческого IgG
Человеческий IgG, который собираются использовать в качестве контроля для химерного анти-НМ 1.24 антитела, очищают следующим образом. После того, как равное количество PBS(-) добавляют к Нu IgG1 степени очистки каппа (изготовитель BINDING SITE), его аффинно очищают, используя высокоскоростную систему очистки антител ConSep LC100 (изготовитель Millipore) и колонку Hyper D Protein А (изготовитель Nippon Gaishi), используя PBS(-) в качестве абсорбирующего буфера, и 0,1 М натрийцитратный буфер (рН 3) в качестве элюирующего буфера в соответствии с прилагаемыми инструкциями. рН элюированных фракций доводят до примерно 7,4, немедленно добавляя 1М Трис-НС1 (рН 8,0), а затем осуществляют концентрированно с помощью центрифужного ультрафильтрационного концентратора Centriprep 10 (изготовитель Amicon) и замену буфера на PBS(-). Его стерилизуют на фильтре, используя мембранный фильтр MILLEX-GV (изготовитель MILLIPORE) с размером пор 0,22 мкм. Приготавливают концентрацию 200 мкг/мл, используя стерилизованный на фильтре PBS(-), и используют в дальнейших экспериментах. Концентрацию антител измеряют по поглощению на 280 нм, и рассчитывают, принимая оптическую плотность 1.35 ОП, соответствующей 1 мг/мл.
2. Способ количественного определения человеческого сывороточного IgG в мышиной сыворотке
Содержание человеческого IgG в мышиной сыворотке, количественно определяют с помощью следующего ELISA. 100 мкл козьего античеловеческого IgG, разбавленного до концентрации 1 мкг/мл в 0,1 М бикарбонатном буфере (рН 9,6), помещают в 96-луночный планшет (изготовитель NUNC) и инкубируют при 4oС в течение ночи, чтобы иммобилизовать антитело. После блокирования добавляют 100 мкл серийно разбавленной мышиной сыворотки или человеческого IgG в качестве стандарта, и инкубируют при комнатной температуре в течение 1 ч. После промывки добавляют 100 мкл 2000-кратно разбавленного античеловеческого IgG, меченного щелочной фосфатазой (изготовитель CAPPEL), и инкубируют при комнатной температуре в течение часа. После промывки добавляют субстратный раствор и инкубируют, а затем измеряют поглощение на 405 нм, используя ридер MICROPLATE READER Model 3550 (изготовитель BioRad).
3. Противоопухолевое действие химерного анти-НМ 1.24 антитела в отношении мышей, которым трансплантированы клетки миеломы человека
3-1. Создание мышей с трансплантированными клетками миеломы человека
Мышей с трансплантированными клетками миеломы человека создают следующим образом. Клетки КРММ2 после пассажей in vivo с использованием мышей SCID (от Nichon CLEA) приготавливают в концентрации 3•107 кл/мл в RPMI 1640 среде, дополненной 10% околоплодной сывороткой теленка (изготовитель GIBCO BRL). 200 мкл суспензии вышеуказанных клеток КРММ2 вводят инъекцией в хвостовую вену SCID мышей (самцы возраста 8 недель от Nichon CLEA), которым накануне внутривенно ввели 100 мкл антиазиало GM1(antiasialo GM1 изготовитель Wako Pure Chemical Industries Co., Ltd.).
3-2. Введение антител
На 12-й день после трансплантации клеток КРММ2 от мышей с трансплантированными клетками миеломы человека получают сыворотку, и содержание человеческого IgG в этой сыворотке количественно определяют, используя ELISA, как указано ранее в 2. Захват клеток КРММ2 в костном мозге подтвержден повышением уровня человеческого IgG в сыворотке. На 14, 21 и 28 сутки после трансплантации КРММ2 клеток 100 мкл каждого из антител, полученных ранее в 1, вводят этим мышам внутривенно.
3-3. Оценка противоопухолевого действия химерного анти-НМ 1.24 антитела на мышей с трансплантированными клетками миеломы человека
Противоопухолевое действие химерного анти-НМ 1.24 антитела оценивают по периоду выживания мышей. Как видно на фиг.33, мыши, которым вводили химерное анти-НМ 1.24 антитело, демонстрируют пролонгированный период выживания по сравнению с мышами, которым вводили контрольный человеческий IgG. Таким образом, подтверждается, что химерное анти-НМ 1.24 антитело обладает противоопухолевым действием в отношении мышей с трансплантированными миеломными клетками человека.
Пример 13. Определение активности реконструированного человеческого анти-НМ 1.24 антитела
ADCC-активность (зависящая от антитела клеточная цитотоксичность) определяют по способу, представленному в Current Protocols in Immunology, Chapter 7, Immunologic studies in humans. Editor, John E, Coligan et al., John Wiley & Sons, Inc., 1993).
1. Получение эффекторных клеток
Мононуклеарные клетки выделяют из периферической крови здоровых людей методом центрифугирования. Так, равные количества PBS(-) добавляют к периферической крови здоровых людей, которая нанесена на Ficoll-Plaque PLUS (изготовитель Pharmacia), и центрифугируют при 400 g в течение 40 мин. Мононуклеарный клеточный слой собирают и четыре раза промывают RPMI 1640 средой, дополненной 10% околоплодной сывороткой теленка (изготовитель GIBCO BRL), и приготавливают при плотности клеток 5•106 кл/мл в той же самой культуральной среде.
LAK (Limphokine Activated Killer Cell-лимфокином активированные киллерные клетки) индуцируют из клеток костного мозга SCID мышей (разведения Nichon CLEA). Так, клетки костного мозга выделяют из бедренной кости мышей и промывают дважды RPMI 1640 средой (изготовитель GIBCO BRL), дополненной 10% околоплодной сывороткой теленка (изготовитель GIBCO BRL), и приготавливают при клеточной плотности 2•105 кл/мл в той же самой культуральной среде. Их инкубируют вместе с 50 нг/мл рекомбинантного человеческого IL-2 (изготовитель R & D SYSTEMS) и 10 нг/мл рекомбинантных мышиных GM-CSP (изготовитель R & D SYSTEMS) в CO2 инкубаторе (изготовитель TABAI) в течение 7 дней. Число клеток доводят до 2•106 кл/мл в той же самой культуральной среде.
2. Получение клеток-мишеней
В клеточную линию миеломы человека КРММ2 (публикация заявки на патент Японии без экспертизы (Kokai) 7-236475) или в плазмоцитарно-лейкозную ARH-77 (ATCC CCL-1621) вводят радиоактивные метки с помощью инкубирования в RPMI 1640 среде (изготовитель GIBCO BRL), дополненной 10% околоплодной сывороткой теленка (изготовитель GIBCO BRL) вместе с 0,1 мкКи 51Сr-хромата натрия (изготовитель ICN) при 37oС в течение 60 мин. После введения радиоактивной метки клетки трижды промывают той же самой культуральной средой, и концентрацию доводят до 2•105 кл/мл.
3. ADCC-анализ
В 96-луночный планшет с U-образным дном (изготовитель Becton Dickinson) добавляют 50 мкл 2•105 клеток-мишеней/мл, 50 мкл реконструированного человеческого анти-НМ 1.24 антитела, мышиного анти-НМ 1.24 антитела, контрольного человеческого IgG (изготовитель THE BINDING SITE) или контрольного мышиного IgG2a (UPC10, изготовитель CAPPEL), и подвергают взаимодействию при 4oС в течение 15 мин.
Затем 100 мкл эффекторных клеток культивируют в СО2 инкубаторе в течение 4 ч, причем отношение (Е:Т) эффекторных клеток (Е) к клеткам-мишеням (Т) устанавливается 0:1, 3,2:1, 8:1, 20:1 или 50:1.
100 мкл надосадочной жидкости отбирают, и измеряют радиоактивность, выделившуюся в культуральную надосадочную жидкость, с помощью счетчика гамма-излучения (ARC-300, изготовитель Aloka). Для измерения максимальной радиоактивности используют 1% NP-40 (изготовитель Nakalai). Цитотоксичность (%) рассчитывают как (А-С)/(В-С) х 100, где А представляет радиоактивность (имп/мин), испускаемую в присутствии антитела, В представляет радиоактивность (имп/мин), испускаемую за счет NP-40, а С представляет радиоактивность (имп/мин), испускаемую только в культуральной среде без антител.
На фиг.34 представлен результат, полученный в случае, когда клетки, полученные из периферической крови здорового человека, используют в качестве эффекторных клеток, а клетки КРММ2 используют в качестве клеток-мишеней. Фиг. 35 демонстрирует результат, полученный в случае, когда клетки, полученные из периферической крови здорового человека используют в качестве эффекторных клеток, a ARH-77 используют в качестве клеток-мишеней. Если добавляют реконструированное человеческое анти-НМ 1.24 антитело, цитотоксичность возрастает с увеличением концентрации по сравнению с контрольным человеческим IgG, что указывает на то, что реконструированное человеческое анти-НМ 1.24 антитело обладает ADCC-активностыо.
Кроме того, если добавляют реконструированное человеческое анти-НМ 1.24 антитело, цитотоксичность заметно повышается по сравнению с мышиным анти-НМ 1.24 антителом, что указывает на то, что реконструированное человеческое анти-НМ 1.24 антитело обладает более высокой ADCC-активностью, нежели мышиное анти-НМ 1.24 антитело. Более того, если КРММ2 используют в качестве клеток-мишеней, добавление реконструированного человеческого анти-НМ 1.24 антитела в концентрации 0,1 мкг/мл или выше не вызывает изменений в цитотоксичности, что указывает на то, что концентрация 0,1 мкг/мл или выше обладает достаточной ADCC-активностыо. Если в качестве клеток-мишеней используют ARH-77, добавление реконструированного человеческого анти-НМ 1.24 антитела в концентрации 1 мкг/мл или выше не вызывает изменений в цитотоксичности, что указывает на то, что концентрация 1 мкг/мл или выше обладает достаточной ADCC-активностью.
На фиг. 36 представлен результат, полученный в случае, когда клетки, полученные из костного мозга мышей SCID используют в качестве эффекторных клеток. Если добавляют реконструированное человеческое анти-НМ 1.24 антитело, цитотоксичность возрастает с увеличением концентрации антител, по сравнению с контрольным человеческим IgG, что указывает на то, что реконструированное человеческое анти-НМ 1.24 антитело обладает ADCC-активностыо. Кроме того, добавление реконструированного человеческого анти-НМ 1.24 антитела в концентрации 0,1 мкг/мл или выше не вызывает изменений в цитотоксичности, что указывает на то, что концентрация 0,1 мкг/мл или выше обладает достаточной ADCC активностью.
Эти результаты показывают, что реконструированное человеческое анти-НМ 1.24 антитело обладает ADCC-активностью, даже если используемые эффекторные клетки получены от людей или мышей.
Пример 14. Противоопухолевое действие реконструированного человеческого анти-НМ 1.24 антитела против миеломы человека в модели на мышах
1. Получение предназначенного для введения антитела
Реконструированное человеческое анти-НМ 1.24 антитело, полученное в результате введения плазмиды HEF-RVLa-AHM-gκ и плазмиды HEF-RVHr-AHM-gγ1 в клетки СНО, приготавливают в концентрации 40, 200 и 1000 мкг/мл, используя стерилизованный на фильтре PBS(-) и контрольный человеческий IgG, полученный в примере 12.1-2, приготовленные в концентрации 200 мкг/мл, с использованием стерилизованного на фильтре PBS(-), которые используют в качестве подлежащих введению антител.
2. Противоопухолевое действие реконструированного человеческого анти-НМ 1.24 антитела против мышей с трансплантированными клетками миеломы человека
2-1. Создание мышей с трансплантированными клетками миеломы человека
Мышей с трансплантированными клетками миеломы человека получают в соответствии с примером 12.3-1. Используют SCID мышей (возраст 5 недель, от Nichon CLEA).
2-2. Введение антител
На 9-е сутки после трансплантации клеток КРММ2 у мышей с трансплантированными клетками миеломы человека, полученными по 2-1, собирают сыворотку, и количество человеческого IgG в этой сыворотке определяют количественно с помощью ELISA, как указано выше в 12.2. Захват клеток КРММ2 на костном мозге подтверждается возрастанием уровня человеческого IgG в сыворотке. На 10-е сутки после трансплантации клеток КРММ2 100 мкл антител, подготовленных как указано ранее в 1, внутривенно вводят этим мышам.
2-3. Оценка противоопухолевого действия реконструированного человеческого анти-НМ 1.24 антитела против мышей с трансплантированными миеломними клетками человека
Противоопухолевое действие реконструированного человеческого анти-НМ 1.24 антитела оценивают по изменению количества человеческого IgG в мышиной сыворотке и по периоду выживания мышей.
Изменение количества человеческого IgG в мышиной сыворотке количественно определяют для сыворотки, собранной на 35 сутки после трансплантации КРММ2 клеток, определяя содержание человеческого IgG с помощью ELISA, как указано а примере 12.2. Результаты, представленные на фиг.37, показывают, что в контрольной группе, которой вводили человеческий IgG1, количество человеческого IgG в сыворотке на 35 день после трансплантации клеток КРММ2, повысилось примерно в 1000 раз по сравнению с количеством на 9-й день (за день до введения антител), тогда как в группе, которой вводили реконструированное человеческое анти-НМ 1.24 антитело, это количество было почти равным или даже ниже, чем на 9-е сутки, для любой дозы, что указывает на то, что реконструированное человеческое анти-НМ 1.24 антитело подавляет рост клеток КРММ2. С другой стороны, для периода выживания, как представлено на фиг.38, пролонгирование наблюдается для группы с введенным реконструированным человеческим анти-НМ 1.24 антителом по сравнению с группой с введенным контрольным человеческим IgG1. Изложенное показывает, что реконструированное человеческое анти-НМ 1.24 антитело обладает противоопухолевым действием в отношении мышей с трансплантированными клетками миеломы человека.
Пример 15. Сравнение противоопухолевого действия реконструированного человеческого анти-НМ 1.24 антитела и существующего лекарства мелфалана в отношении миеломы человека в модели на мышах
1. Получение подлежащих введению лекарств
1-1. Получение подлежащих введению антител
Реконструированное человеческое анти-НМ 1.24 антитело, полученное в результате введения плазмиды HEF-RVLa-AHM-gκ и плазмиды HTF-RVHr-AHM-gγ1 в клетки СНО приготавливают в концентрации 40 и 200 мкг/мл, используя стерилизованный на фильтре PBS(-) и контрольный человеческий IgG, полученный в примере 12.1-2, приготовленные в концентрации 200 мкг/мл, с использованием стерилизованного на фильтре PBS(-), которые используют в качестве подлежащих введению антител.
1-2. Получение мелфалана
Мелфалан (производство SIGMA), который представляет собой существующее лекарство для лечения миеломы, приготавливают в концентрации 0,1 мг/мл, используя 0,2% карбоксиметилцеллюлозу (CMC) изготовитель Daicel Chemical Industries, Ltd.).
2. Противоопухолевое действие реконструированного человеческого анти-НМ 1.24 антитела и мелфалана в отношении мышей с трансплантированными клетками миеломы человека
2-1. Создание мышей с трансплантированными клетками миеломы человека
Мышей с трансплантированными клетками миеломы человека создают по способу примера 14.2-1.
2-2. Введение лекарства
На 9-е сутки после трансплантации клеток КРММ2 у мышей с трансплантированными клетками миеломы человека, полученными по 2-1, собирают сыворотку, и количество человеческого IgG в этой сыворотке определяют количественно с помощью ELISA, как указано выше в 12.2. Захват клеток КРММ2 на костном мозге подтверждается возрастанием уровня человеческого IgG в сыворотке. На 10-е сутки после трансплантации клеток КРММ2 100 мкл каждого из антител, подготовленных, как указано ранее в 1-1, внутривенно вводят этим мышам. Кроме того, 200 мкл 0,2% CMC раствора перорально вводят раз в день в течение 5 дней начиная с дня 10 после трансплантации. С другой стороны, для группы, которой вводят мелфалан, мелфалановый раствор, подготовленный как указано ранее в 1-2, перорально вводят в количестве 100 мкл на 10 г веса тела (1 мг/кг в расчете на мелфалан) раз в день в течение 5 дней, начиная с 10 дня после трансплантации клеток КРММ2.
2-3. Оценка противоопухолевого действия реконструированного человеческого анти-НМ 1.24 антитела в отношении мышей с трансплантированными клетками миеломы человека
Противоопухолевое действие реконструированного человеческого анти-НМ 1.24 антитела оценивают по изменению количества человеческого IgG в мышиной сыворотке и по периоду выживания мышей.
Изменение количества человеческого IgG в мышиной сыворотке количественно определяют для сыворотки, собранной на 35 сутки после трансплантации клеток КРММ2, определяя содержание человеческого IgG с помощью ELISA, как указано в примере 12.2. Результаты, представленные на фиг.39, показывают, что в контрольной группе, которой вводили человеческий IgG1, количество человеческого IgG в сыворотке на 35 день после трансплантации клеток КРММ2, повысилось примерно в 1000 раз по сравнению с количеством на 9 сутки (за день до введения антител), тогда как видно, что клетки КРММ2 росли в организме данных мышей. В группе, которой вводили мелфалан, также количество человеческого IgG в сыворотке возросло более высокого уровня, нежели было до введения лекарства, хотя и не столь сильно, как в контрольной группе, которой вводили человеческий IgG. Этот результат указывает на то, что введение мелфалана не достаточно подавляет рост клеток КРММ2. С другой стороны, количество человеческого IgG в сыворотке группы, которой вводили реконструированное человеческое анти-НМ 1.24 антитело, на 35 сутки было меньше, чем на 9 сутки после трансплантации для любых доз, что указывает на то, что реконструированное человеческое анти-НМ 1.24 антитело подавляет рост клеток КРММ2.
С другой стороны, для периода выживания, как представлено на фиг.40, также наблюдается пролонгирование для группы, которой вводили реконструированное человеческое анти-НМ 1.24 антитело, по сравнению с группой, которой вводили контрольный человеческий IgG1. Из изложенного следует, что реконструированное человеческое анти-НМ 1.24 антитело обладает противоопухолевым действием в отношении мышей с трансплантированными клетками миеломы человека, и что противоопухолевое действие антитела по настоящему изобретению сильнее, нежели действие существующего лекарства мелфалана.
Вышеприведенные результаты показывают, что если используют полученные от человека эффекторные клетки, мышиное анти-НМ 1.24 антитело обладает малой цитотоксичностью в отношении клеток миеломы человека, тогда как реконструированное человеческое анти-НМ 1.24 антитело и химерное анти-НМ 1.24 антитело обладают высокой цитотоксичностью. Этот факт демонстрирует важность гуманизации антитела, и дает надежду на полезность реконструированного человеческого анти-НМ 1.24 антитела для людей.
Реконструированное человеческое анти-НМ 1.24 антитело демонстрирует очень сильное противоопухолевое действие в отношении мышей SCID с трансплантированными клетками миеломы человека. Так как у людей эффекторные клетки получают от людей и обычно присутствуют лимфоциты, ожидается еще более сильное противоопухолевое действие реконструированного человеческого анти-НМ 1.24 антитела.
В модели с миеломой реконструированное человеческое анти-НМ 1.24 антитело демонстрирует сильное противоопухолевое действие по сравнению с существующим лекарством, и поэтому ожидается, что реконструированное человеческое анти-НМ 1.24 антитело может составить эпоху в производстве лекарств для лечения миеломы.
Сравнительный пример 1. Конструирование гибридомы, которая продуцирует мышиное анти-НМ 1.24 моноклональное антитело
Гибридому, которая продуцирует мышиное анти-НМ 1.24 моноклональное антитело, получают по способу, раскрытому у Goto, Т. et al., Blood (1994) 84, 1992-1930.
Плазму клеточной линии КРС-32, негативную по ядерному антигену вируса Эпштейна-Барра (EBNA) (1•107 клеток), полученную из костного мозга пациента с множественной миеломой (Goto, Т. et al., Jpn. J. Clin. Hematol. (1991) 32, 1400) вводят внутрибрюшинно дважды BALB/с мышам (от Charles River) каждые шесть недель.
Для дальнейшей оценки титра продуцирования антител 1,5•106 КРС-32 клеток вводят в селезенку мышам за три дня до умерщвления животных (Goto, Т. et al. , Tokushima J. Exp. Med. (1990) 37, 89). После умерщвления селезенку извлекают, и клетки селезенки, удаленные по способу Groth, de St. & Schreidegger (Cancer Research (1981) 41, 3465) подвергают клеточному слиянию с миеломными клетками SP2/0.
Антитела в надосадочной жидкости гибридомной культуры скринируют с помощью ELISA (Posner, M.R. et al., J. Immunol. Methods (1982) 48, 23), используя планшеты с нанесенными клетками КРС-32. 5•104 клеток КРС-32 суспендируют в 50 мл PBS, и аликвоты помещают в 96-луночный планшет (с U-образным дном. Corning, изготовитель Iwaki). После блокирования PBS, содержащим 1% альбумина бычьей сыворотки (BSA), добавляют надосадочную жидкость гибридомы и инкубируют в течение 2 ч при 4oС. Затем реагируют с козьим антителом против мышиного IgG, меченным пероксидазой (изготовитель Zymed) при 4oС в течение 1 ч, один раз промывают, и реагируют с субстратным раствором о-фенилендиамина (изготовитель Sumitomo Bakelite) при комнатной температуре в течение 30 мин.
После того, как реакцию останавливают 2 н. серной кислотой, измеряют поглощение на 492 нм, используя считывающее устройство ELISA Reader (изготовитель BioRad). Для удаления гибридомы, которая продуцирует антитело против человеческого иммуноглобулина, позитивную гибридомную надосадочную культуральную жидкость предварительно адсорбируют на сыворотку человека и скринируют реакционную способность в отношении других субклеточных компонентов. Позитивные гибридомы отбирают, и исследуют их реакционную способность в отношении различных клеточных линий и образцов человека, используя цитометрию в потоке. Выбранные в итоге гибридомные клоны дважды клонируют, инъектируют в абдоминальную полость обработанных пристанем BALB/c мышей, а затем из них получают асцитную жидкость.
Моноклональное антитело очищают от мышиных асцитов с помощью осаждения аммонийсульфатом и набора для аффинной хроматографии Protein A (Ampure PA, изготовитель Amersham). Очищенное антитело конъюгируют с флуоресцинизотиоцианатом (FITC), используя набор для конъюгации Quick Tag FITC (изготовитель Boechringer Mannheim).
В результате моноклональные антитела, продуцируемые 30 гибридомными клонами, реагируют с КРС-32 и клетками RPMI 8226. После клонирования исследуют реакционную способность надосадочных жидкостей этих гибридом с другими клеточными линиями и мононуклеарными клетками, полученными из периферической крови.
Среди них имеются три клона, которые продуцируют моноклональные антитела, которые специфически взаимодействуют с клетками плазмы. Среди этих трех клонов отбирают гибридомный клон, который продуцирует моноклональное антитело, которое наиболее подходит для цитометрического анализа в потоке и которое обладает комплемент-зависимой цитотоксичностью в отношении клеток RPUI 8226, и называют НМ 1.24. Подкласс моноклональных антител, продуцирующих эту гибридому, определяют с помощью ELISA, используя субкласс-специфическое кроличье антимышиное антитело (изготовитель Zymed). Анти-НМ 1.24 антитело относится к подклассу IgG2aκ. Гибридома, которая продуцирует анти-НМ 1.24 антитело, была интернационально депонирована 14 сентября 1995 года the National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, MITI (Higashi 1-Chome 1-3, Tsukuba city, Ibalaki prefecture, Japan) как под регистрационным номером FERM ВР-5233 в соответствии с Будапештским соглашением.
Сравнительный пример 2. Клонирование кДНК, кодирующей полипептид НМ 1.24 антигена
1. Конструирование библиотеки кДНК
1) Получение полной РНК
кДНК, которая кодирует НМ 1.24 антиген, который представляет собой полипептид, специфически распознаваемый мышиным анти-НМ 1.24 моноклональным антителом, выделяют следующим образом.
Из человеческой клеточной линии рассеянной миеломы КРММ2 полную РНК получают по способу Chirgwin et al. (Biochemistry, 18, 5294 (1979)). Так, 2,2•108 КРММ2 клеток полностью гомогенизируют в 20 мл 4 М гуанидинтиоцианата (изготовитель Nakalai tesque).
Гомогенат наносят на слой 5,3 М хлорида цезия в центрифужной ампуле, а затем центрифугируют при 31000 об/мин при 20oС в течение 24 ч для осаждения РНК, используя ротор Beckman SW40. Осадок РНК промывают 70% этанолом и растворяют в 300 мкл 10 мМ Трис-НС1 (рН 7,4), содержащем 1 мМ ЭДТА и 0,5% ДДС. После добавления к этому проназы (изготовитель Boehringer) до концентрации 0,5 мг/мл все инкубируют при 37oС в течение 30 мин. Полученную смесь экстрагируют фенолом и хлороформом для осаждения РНК. Затем осадок РНК растворяют в 200 мкл 10 мМ Трис-НС1 (рН 7,4), содержащем 1 мМ ЭДТА.
2) Получение поли(А)+РНК
Используя 500 мкг полной РНК, полученной, как указано выше, в качестве сырья, выделяют поли(А)+РНК, используя набор для выделения Fast Track 2,0 m RNA Isolation Kit (изготовитель Invitrogen) в соответствии с инструкциями изготовителя.
3) Конструирование библиотеки кДНК
Используя 10 мкг вышеуказанного поли(А)+РНК в качестве сырья, синтезируют двухцепочечную кДНК, используя набор для синтеза кДНК TimeSaver cDNA Synthesis Kit (изготовитель Pharmacia) в соответствии с инструкциями изготовителя, прилагаемыми к набору, и используя Directional Cloning Toolbox (изготовитель Pharmacia), с ней связывают EcoRI адаптор в соответствии с инструкциями изготовителя, прилагаемыми к набору. Кинецию и обработку ферментом рестрикции NotI EcoRI адаптера осуществляют в соответствии с инструкциями, прилагаемыми к набору. Далее двухцепочечную кДНК с присоединенным адаптером размером около 500 п.н. или выше выделяют и очищают, используя 1,5% агарозный гель с низкой температурой плавления (производство SIGMA), получая около 40 мкл двухцепочечной кДНК с присоединенным адаптером.
Полученную таким образом двухцепочечную кДНК с присоединенным адаптером связывают с pCOS1 вектором (публикация заявки на патент Японии без экспертизы (Kokai) 8-255196), который ранее был обработан ферментами рестрикции EcoRI и NotI и щелочной фосфатазой (производство Takara Shuzo), используя Т4 для конструирования ДНК-лигазы (изготовитель GIBCO BRL) для конструирования кДНК-библиотеки. Сконструированную кДНК-библиотеку трансдуцируют в Eschericia coli штамм DH5α (изготовитель GIBCO BRL), и устанавливают, что полное количество составляет около 2,5•106 независимых клонов.
2. Клонирование за счет прямой экспрессии
1) Трансфекция клеток COS-7
кДНК амплифицируют, культивируя около 5•105 клонов вышеуказанной трансдуцированной Eschericia coli в 2-YT среде (Molecular Cloning: A Laboratory Manual, Sambrook et al., Cold Spring Harbor Laboratory Press, (1989)), содержащей 50 мкг/мл ампициллина, и плазмидную ДНК выделяют из Eschericia coli щелочным методом (Molecular Cloning: A Laboratory Manual, Sambrook et al., Cold Spring Harbor Laboratory Press, (1989)). Полученной плазмидной ДНК трансфицируют клетки COS-7, используя прибор Gene Pulser (изготовитель BioRad).
Затем 10 мкг очищенной плазмидной ДНК добавляют к 0,8 мл клеток COS-7, которые суспендированы в PBS в концентрации 1•107 кл/мл, и обрабатывают импульсами при 1500 В и емкости 25 мкФ. После 10 минутного периода восстановления при комнатной температуре подвергнутые электропорации клетки культивируют в DMEM (изготовитель GIBCO BRL), дополненной 10% околоплодной сывороткой теленка в условиях 37oС и 5% CO2 в течение трех дней.
2) Приготовление кюветы для пэннинга
Кювету для пэннинга с нанесенным мышиным анти-НМ 1.24 антителом приготавливают по способу В. Seed et al. (Proc. Natl. Acad. Sci. USA, 84, 3365-3369 (1987)). Так, мышиное анти-НМ 1.24 антитело добавляют к 50 мМ Трис-HCl, рН 9,5, до концентрации 10 мкг/мл. Три мл раствора антител, приготовленного таким образом, наносят на чашку для культуры тканей с диаметром 60 мм, и инкубируют при комнатной температуре в течение 2 ч. После трехкратной промывки 0,15 М раствором NaC1 и блокирования PBS, содержащим 5% околоплодную сыворотку теленка, 1 мМ ЭДТА и 0,02% NaN3, эти пластины используют для последующего клонирования.
3) Клонирование кДНК-библиотеки
Клетки COS-7, трансфицированные как указано выше, освобождают с помощью PBS, содержащего 5 мМ ЭДТА, а затем промывают один раз PBS, содержащим 5% околоплодную сыворотку теленка. Затем эти клетки суспендируют в PBS, содержащем 5% околоплодную сыворотку теленка 0,02% NaN3 до концентрации около 1•106 кл/мл, которые потом добавляют в кювету для пэннинга, подготовленную как указано выше, и инкубируют при комнатной температуре в течение 2 ч. После трехкратной осторожной промывки PBS, содержащим 5% околоплодную сыворотку теленка, 0,2% NaN3, плазмидную ДНК выделяют из клеток, связанных с кюветой для пэннинга, используя раствор, содержащий 0,6% ДДС и 10 мМ ЭДТА.
Выделенную плазмидную ДНК снова трансдуцируют в Еschericia coli штамм DH5α. После амплификации плазмидной ДНК, как указано выше, ее выделяют щелочным методом. Выделенную плазмидную ДНК трансфицируют в клетки COS-7 методом электропорации, и плазмидную ДНК выделяют из связанных клеток, как указано выше. Эту же самую процедуру повторяют еще раз, и выделенную плазмидную ДНК расщепляют ферментами рестрикции EcoRI и NotI. В результате подтверждается концентрирование вставок с размером около 0,9 т.п.н. Eschericia coli, трансдуцированную с частью выделенной плазмидной ДНК инокулируют в агарные пластины 2-YT, содержащие 50 мкг/мл ампициллина. После культивирования в течение ночи плазмидную ДНК выделяют из одной колонии. Ее расщепляют ферментами рестрикции EcoRI и NotI, и получают клон р 3.19, содержащий вставку 0,9 т.п.н.
Нуклеотидную последовательность этого клона определяют в реакции, используя набор PRISM, Dye Terminator Cycle Sequencing Kit (производство Perkin Elmer) в соответствии с прилагаемыми к набору инструкциями, и осуществляют секвенирование с помощью секвенатора ABI 373А DNA Sequencer (производство Perkin Elmer). Аминокислотная последовательность и последовательность оснований представлены в Последовательности ИД 103.
кДНК, кодирующую полипептид с аминокислотной последовательностью, представленной в последовательности ИД 103, встраивают в XbaI сайт расщепления вектора pUC19 и получают в виде плазмиды pRS38-pUC19. Eschericia coli, которая содержит эту плазмиду pRS38-pUC19, была интернационально депонирована 5 октября 1993 г., как Eschericia coli DH5α (pRS38-pUC19), the National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, MITI (Higashi 1-Chome 1-3, Tsukuba city, Ibalaki prefecture, Japan) как под регистрационным номером FERM ВР-4434 в соответствии с Будапештским соглашением (см. публикацию заявки на патент Японии без экспертизы (Kokai) 7-196694).
Промышленная применимость
Так как химерное анти-НМ 1.24 антитело состоит из вариабельного участка мышиного анти-НМ 1.24 антитела и константного участка человеческого антитела, и реконструированное человеческое анти-НМ 1.24 антитело состоит из участка, мышиного анти-НМ 1.24 антитела, определяющего комплементарность, каркасного участка человеческого антитела и константного участка человеческого антитела, оно обладает слабой антигенностью в отношении человека, и поэтому, как ожидают, может быть использовано в качестве лекарственной композиции, особенно при лечении миеломы.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОНСТРУИРОВАННЫЕ ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-8 | 1995 |
|
RU2159776C2 |
| Н- И L-ЦЕПИ МОНОКЛОНАЛЬНОГО АНТИТЕЛА РМ1 (МОНАТ) К РЕЦЕПТОРУ IL-6R ЧЕЛОВЕКА И ИХ V-ОБЛАСТИ, МОДИФИЦИРОВАННАЯ МОНАТ, ЕГО Н- И L-ЦЕПИ И ИХ V-ОБЛАСТИ, CDR- ПОСЛЕДОВАТЕЛЬНОСТИ, ДНК-ПОСЛЕДОВАТЕЛЬНОСТИ | 1992 |
|
RU2139351C1 |
| МОДИФИЦИРОВАННОЕ АГОНИСТИЧЕСКОЕ АНТИТЕЛО | 2001 |
|
RU2295537C2 |
| ДЕГРАДИРОВАННОЕ АНТИТЕЛО, ЯВЛЯЮЩЕЕСЯ АГОНИСТОМ TPO | 2001 |
|
RU2287534C2 |
| СРЕДСТВО, ПРИМЕНЯЕМОЕ ПРИ ЛЕЧЕНИИ ОПУХОЛЕЙ ЛИМФАТИЧЕСКОЙ ТКАНИ | 1998 |
|
RU2177805C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО, ЗАМЕНЯЮЩЕЕ ФУНКЦИОНАЛЬНЫЕ БЕЛКИ | 2004 |
|
RU2339696C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА, РОДСТВЕННОГО ПАРАЩИТОВИДНОМУ ГОРМОНУ ЧЕЛОВЕКА | 2002 |
|
RU2322453C2 |
| АНТИ-ГЛИПИКАН 3-АНТИТЕЛО, ИМЕЮЩЕЕ МОДИФИЦИРОВАННУЮ САХАРНУЮ ЦЕПЬ | 2005 |
|
RU2451030C2 |
| СПОСОБ И НАБОР ДЛЯ ВЫЯВЛЕНИЯ ГЕНА, КОДИРУЮЩЕГО МЕМБРАНОСВЯЗАННЫЙ БЕЛОК, ВЕКТОР (ВАРИАНТЫ) | 1999 |
|
RU2246538C2 |
| СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ КАРДИОПАТИИ | 2006 |
|
RU2450830C2 |



















































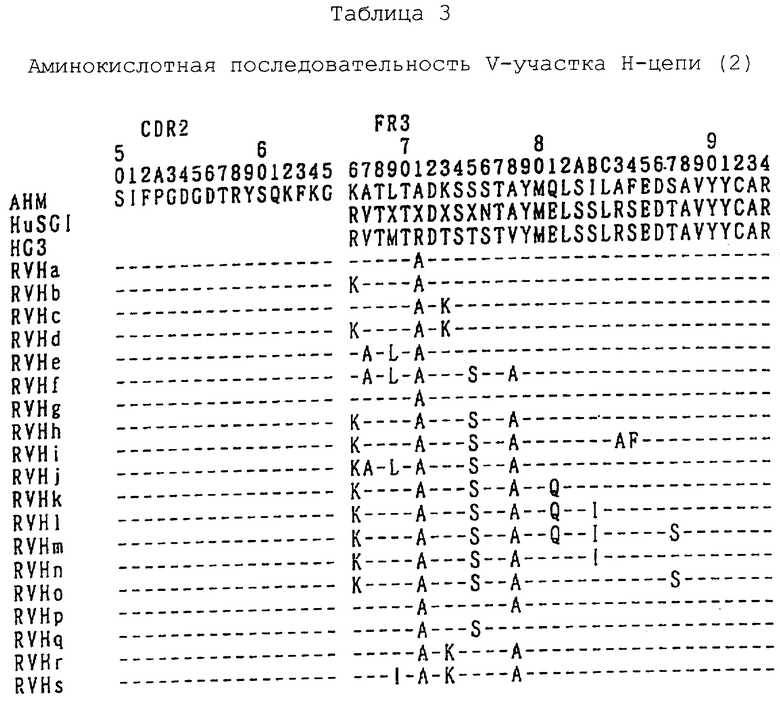



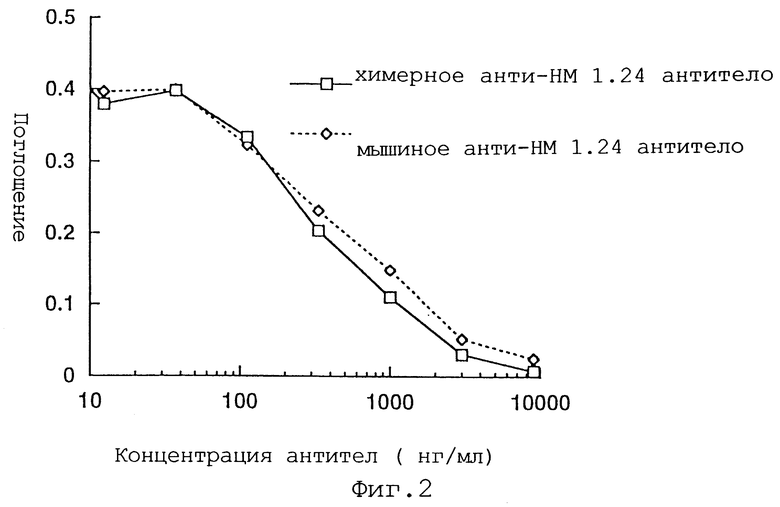
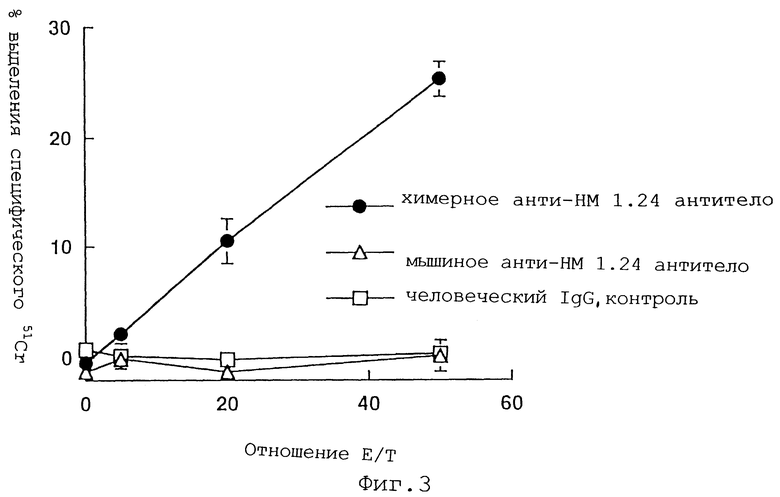
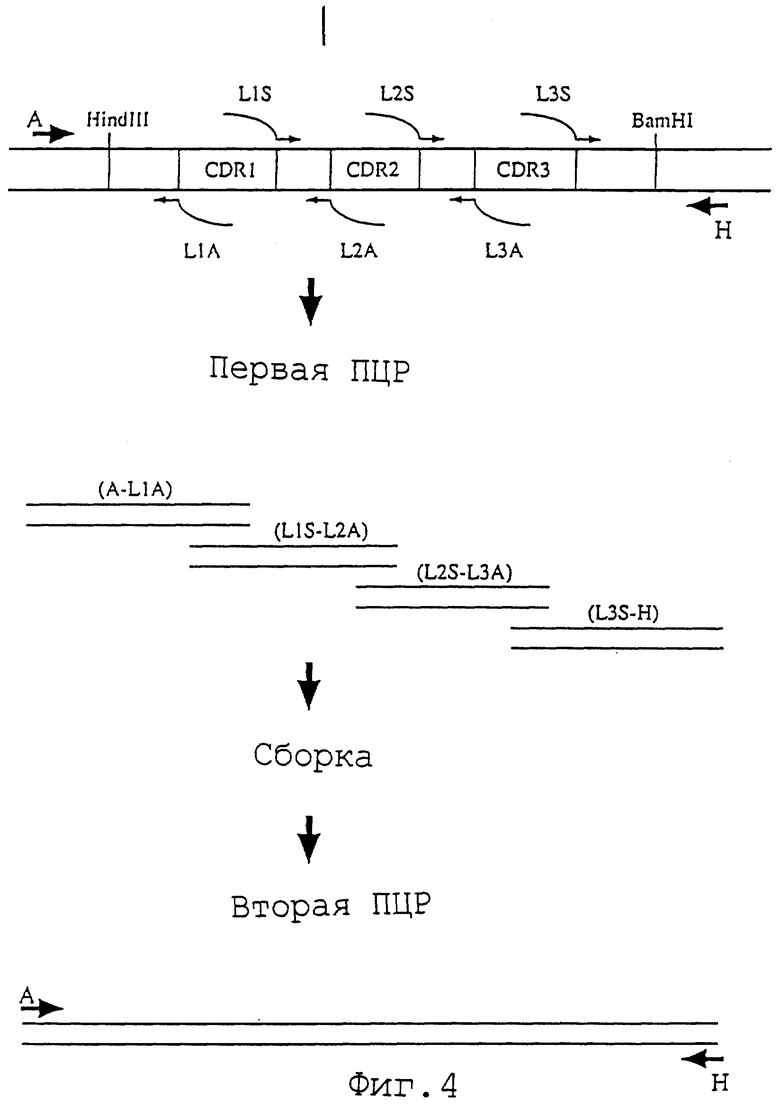
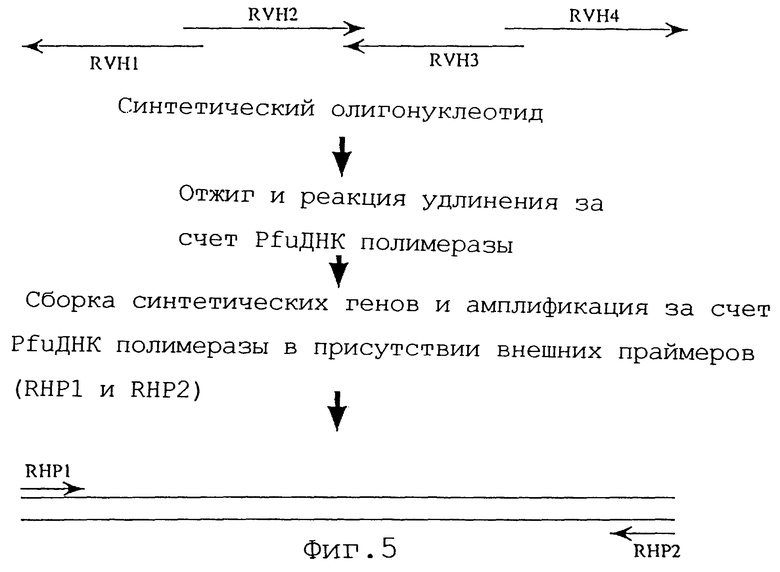
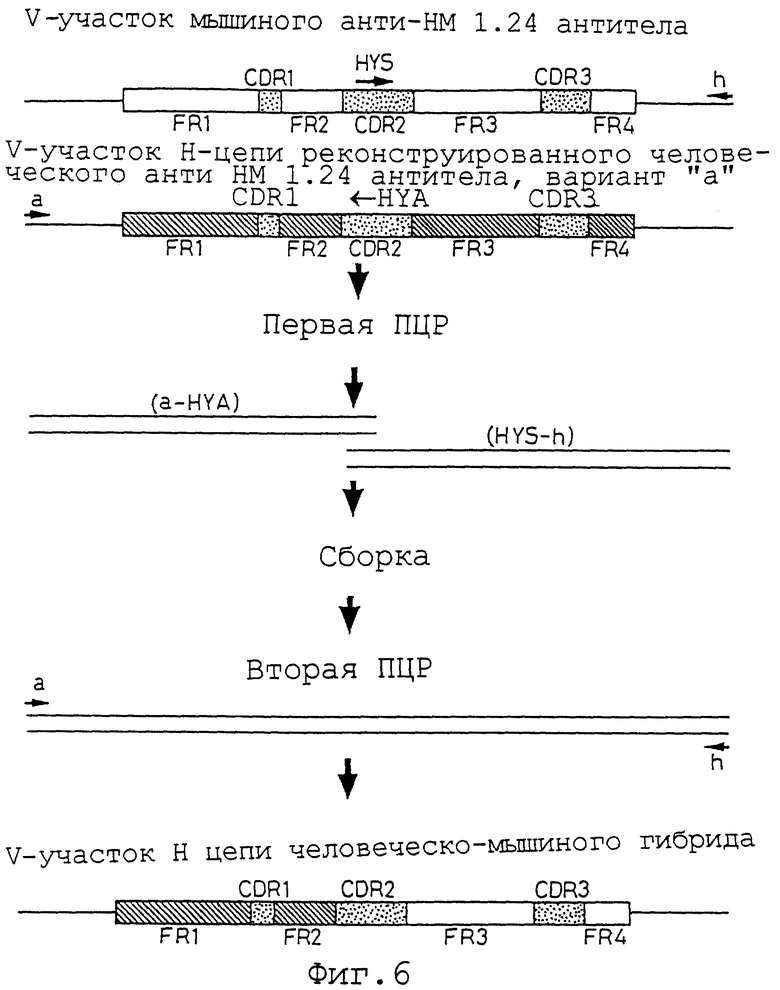
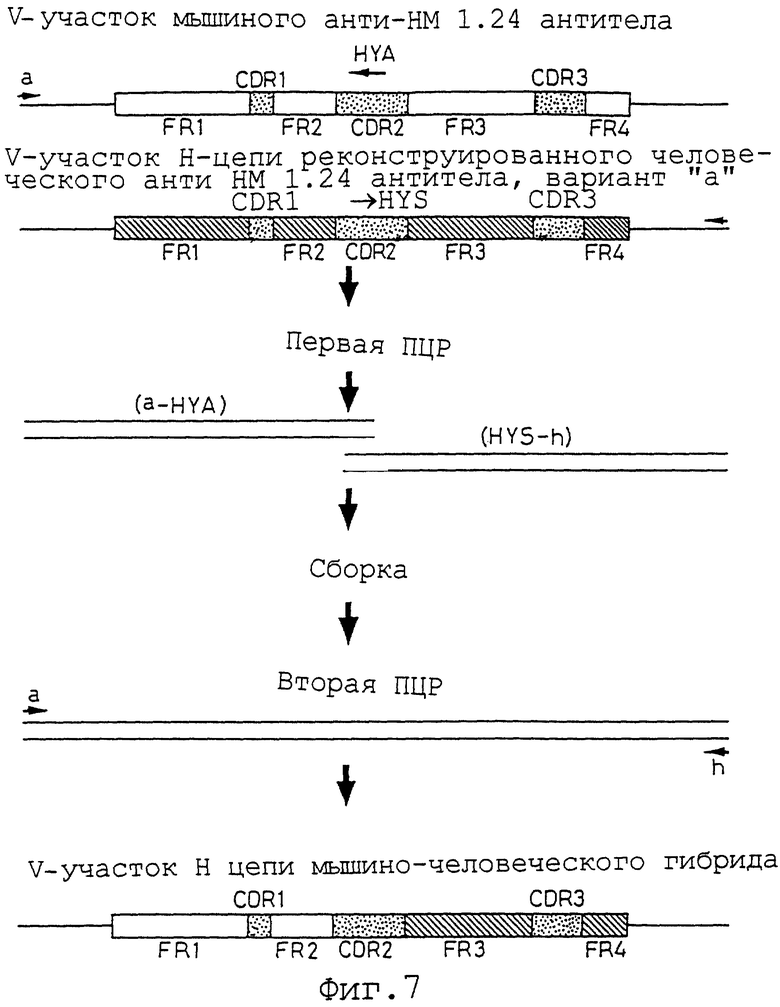
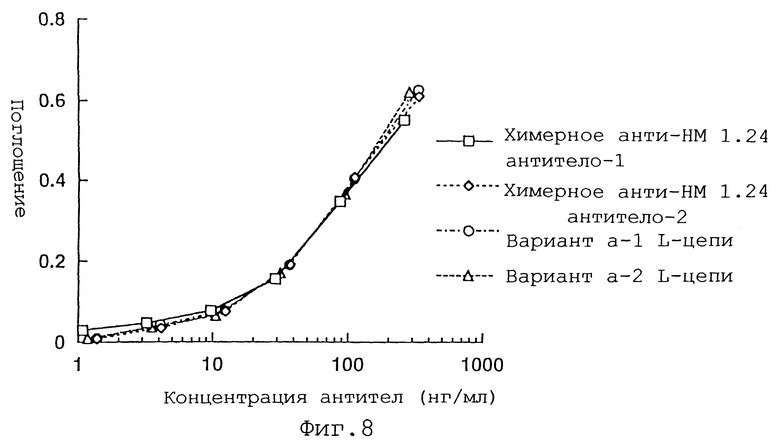

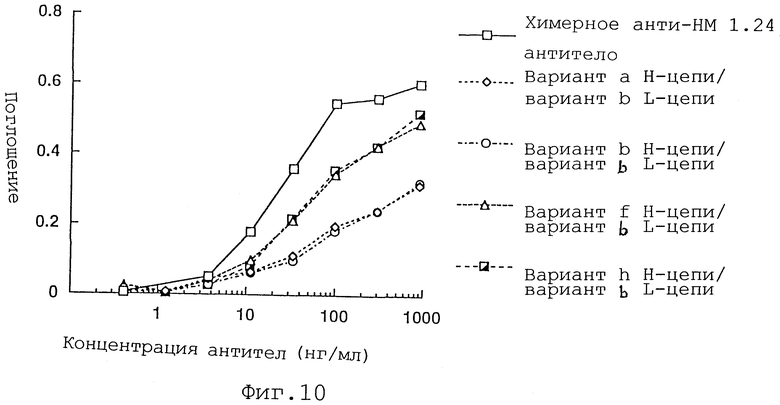
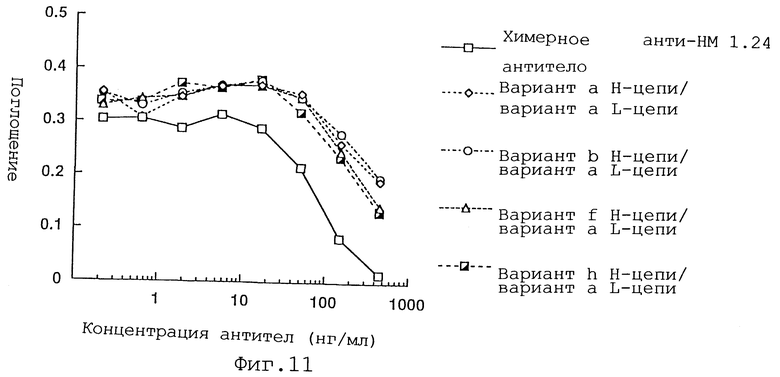
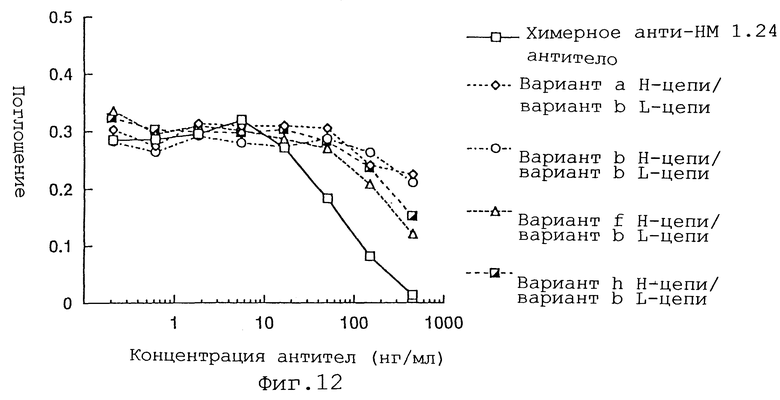
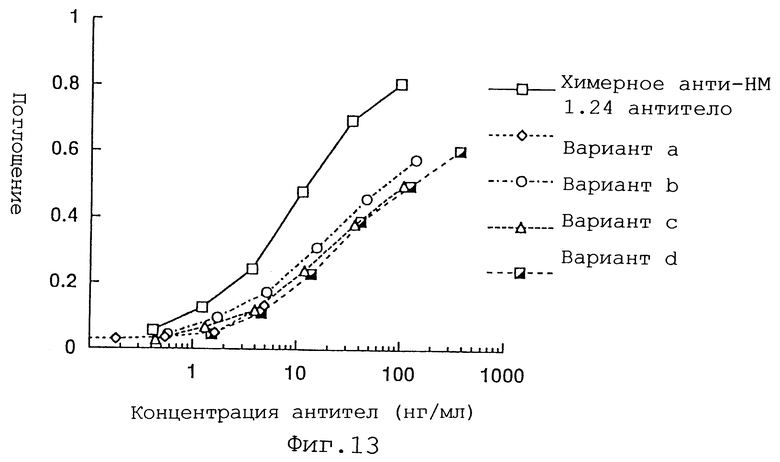
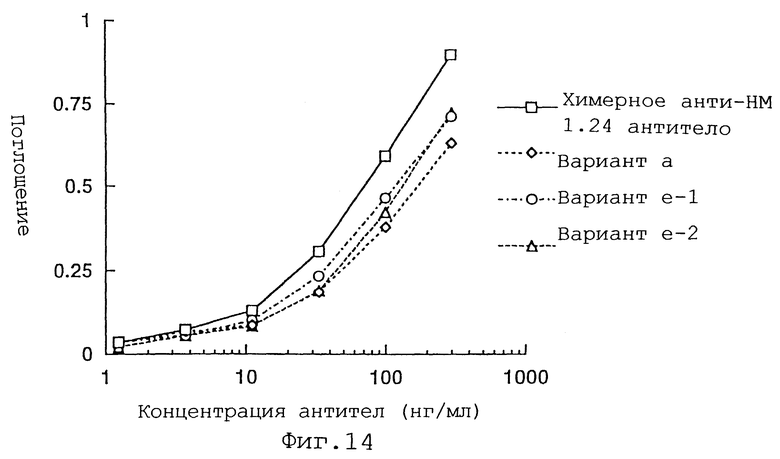
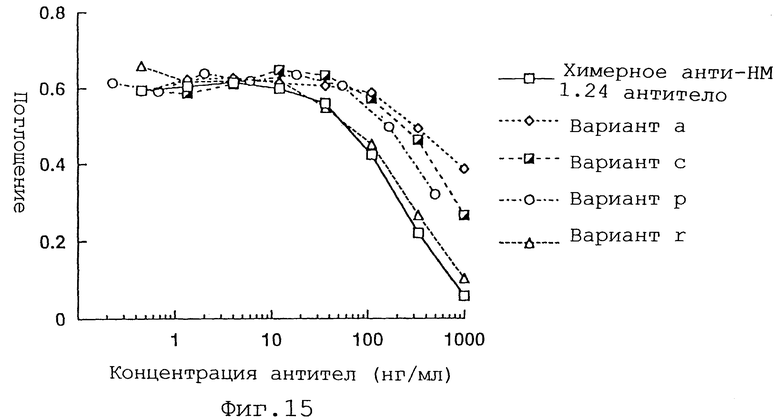
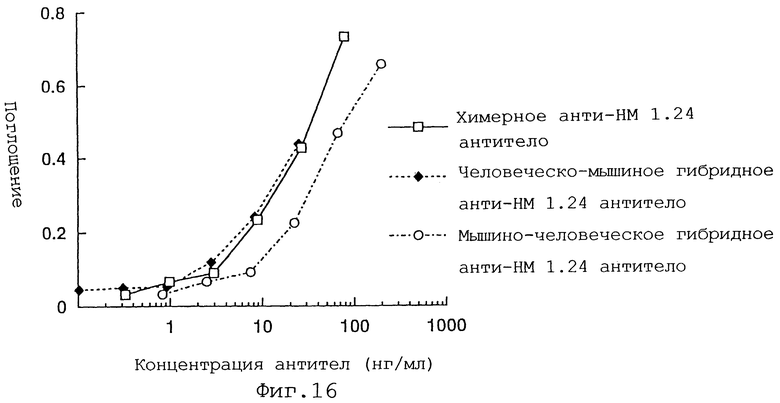
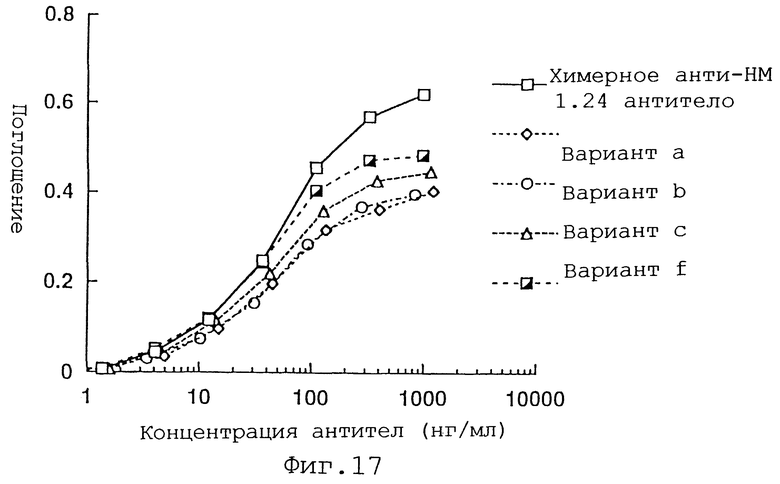
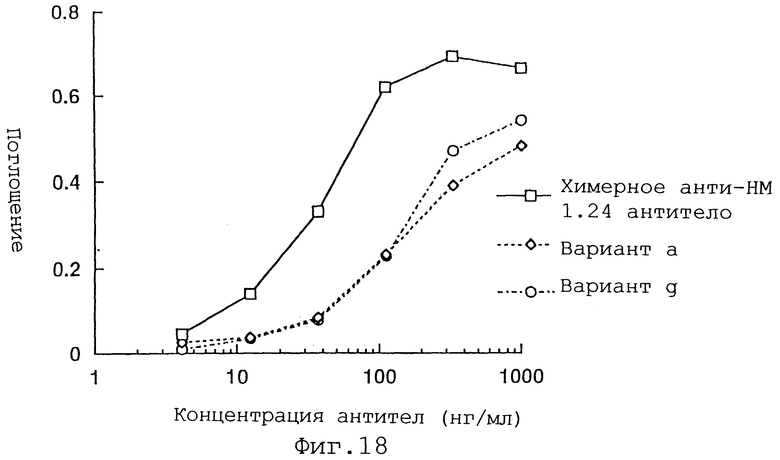
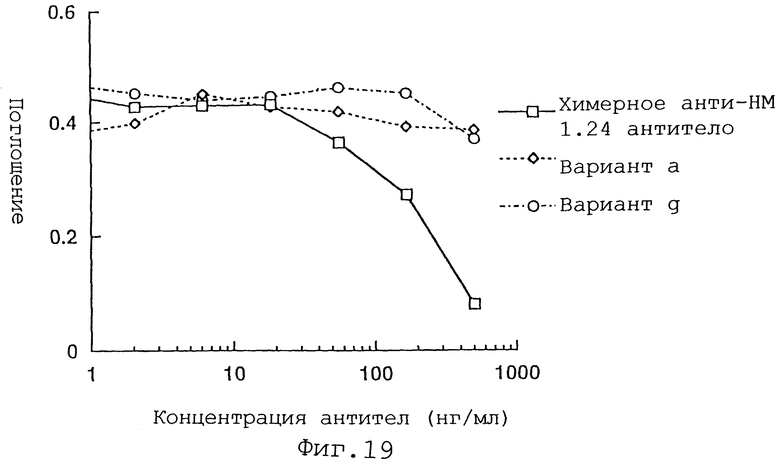
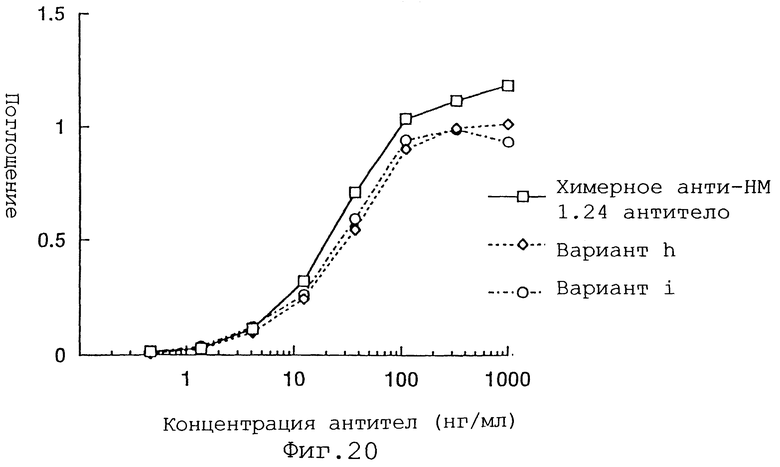
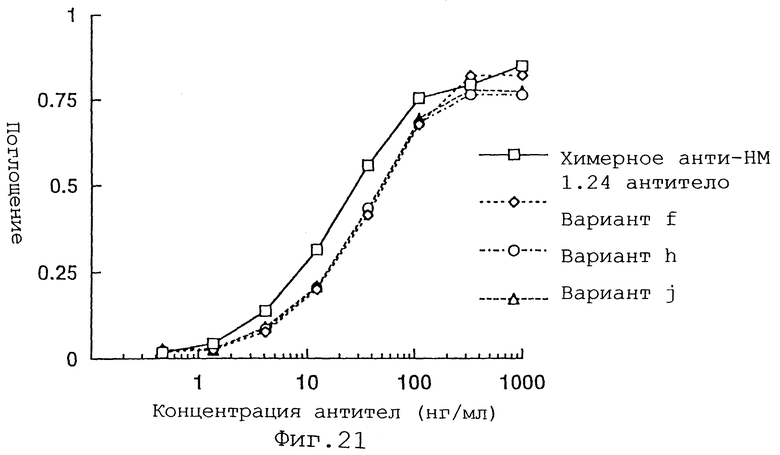
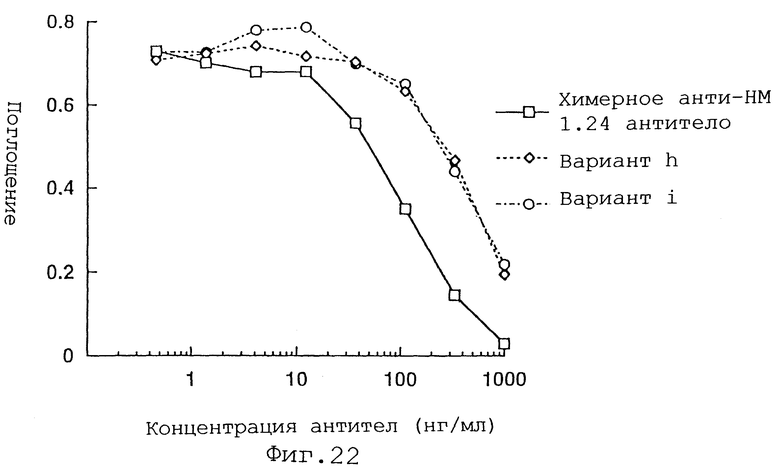
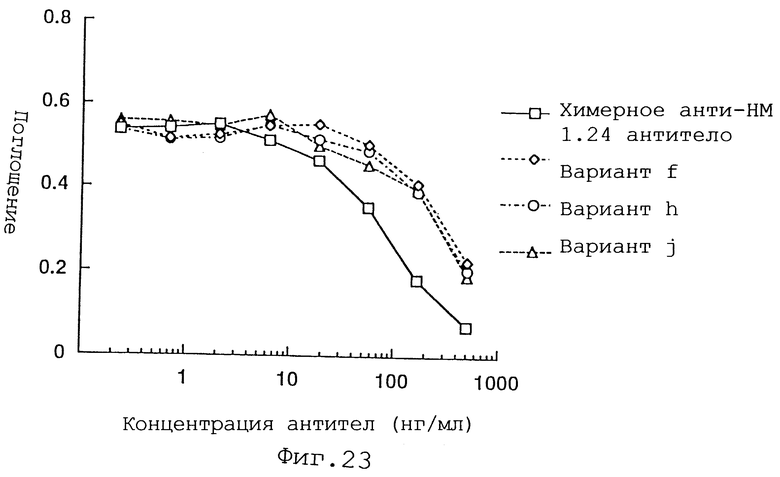
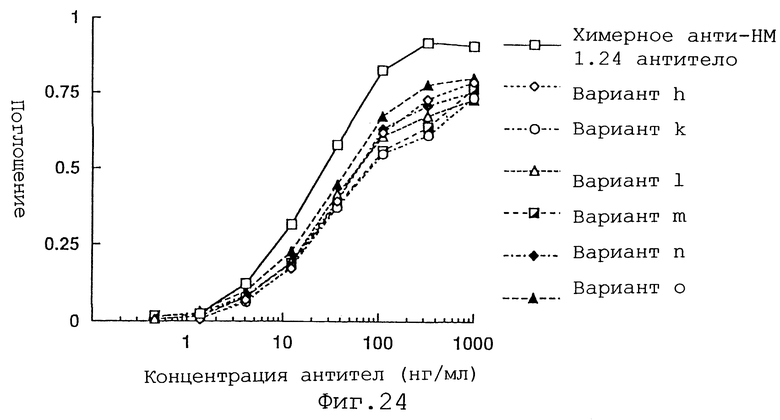
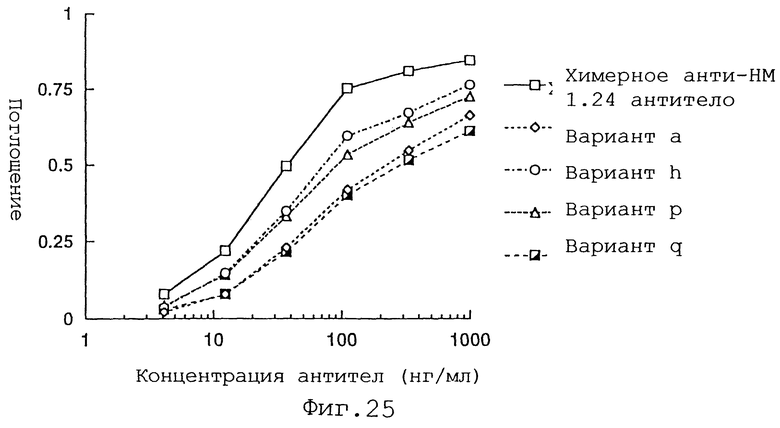
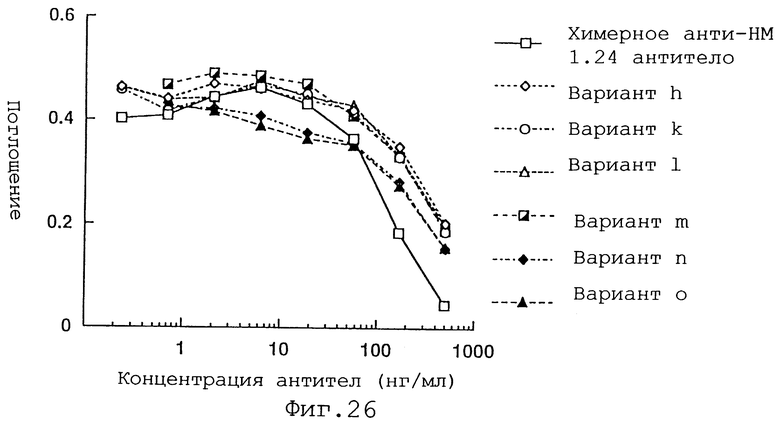
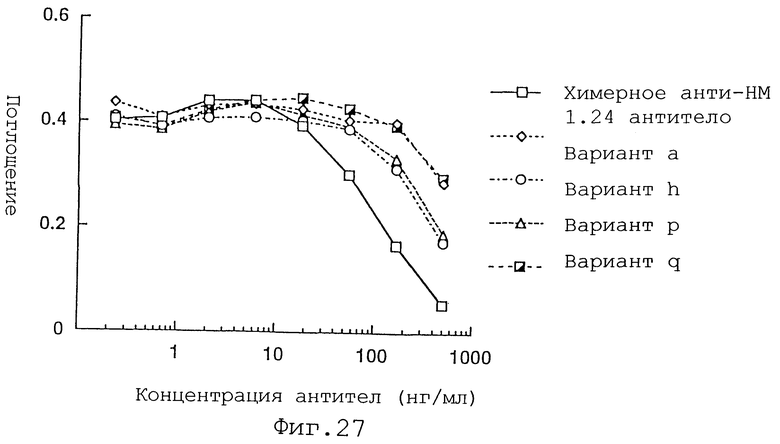
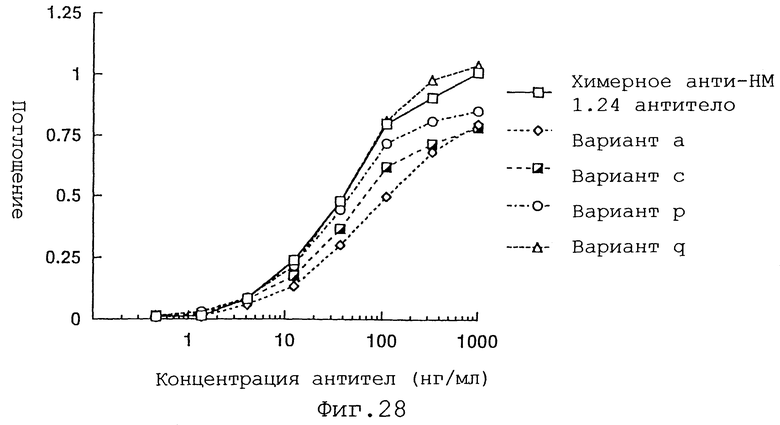


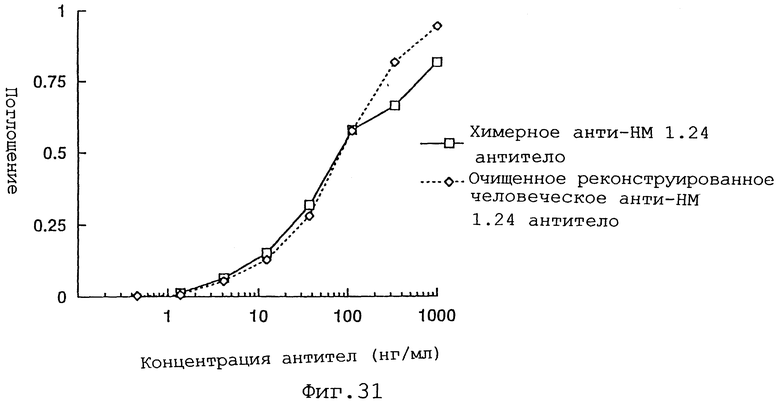
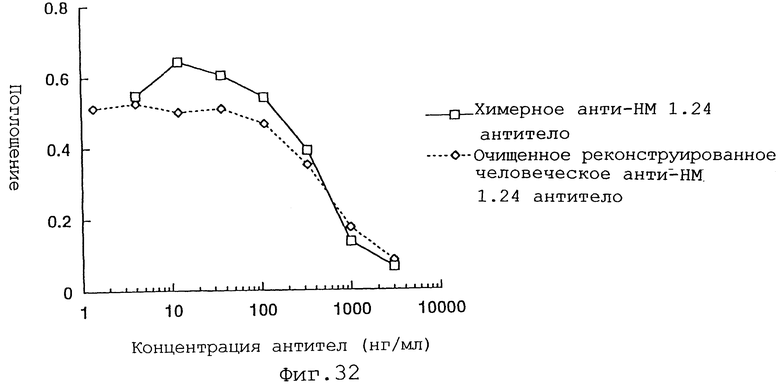
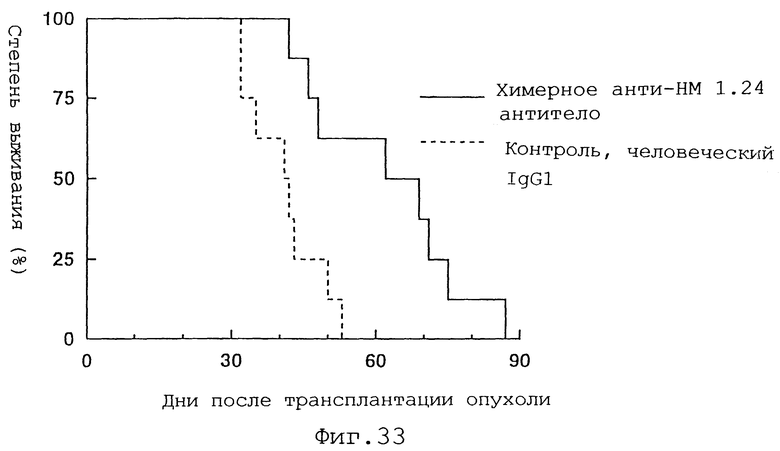
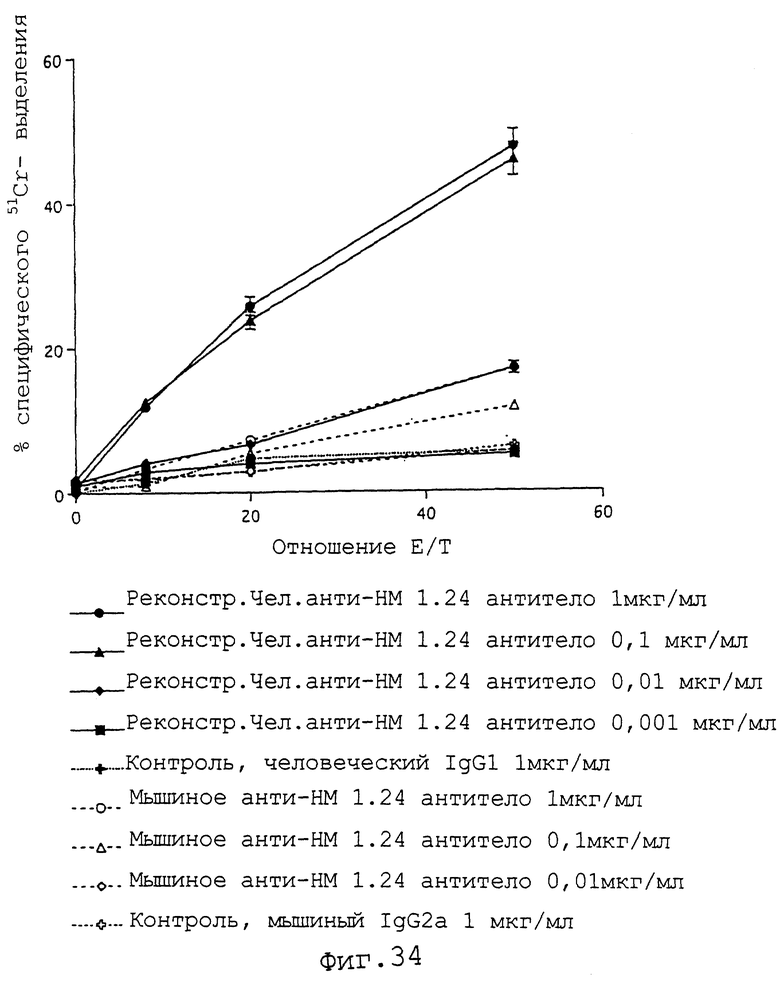
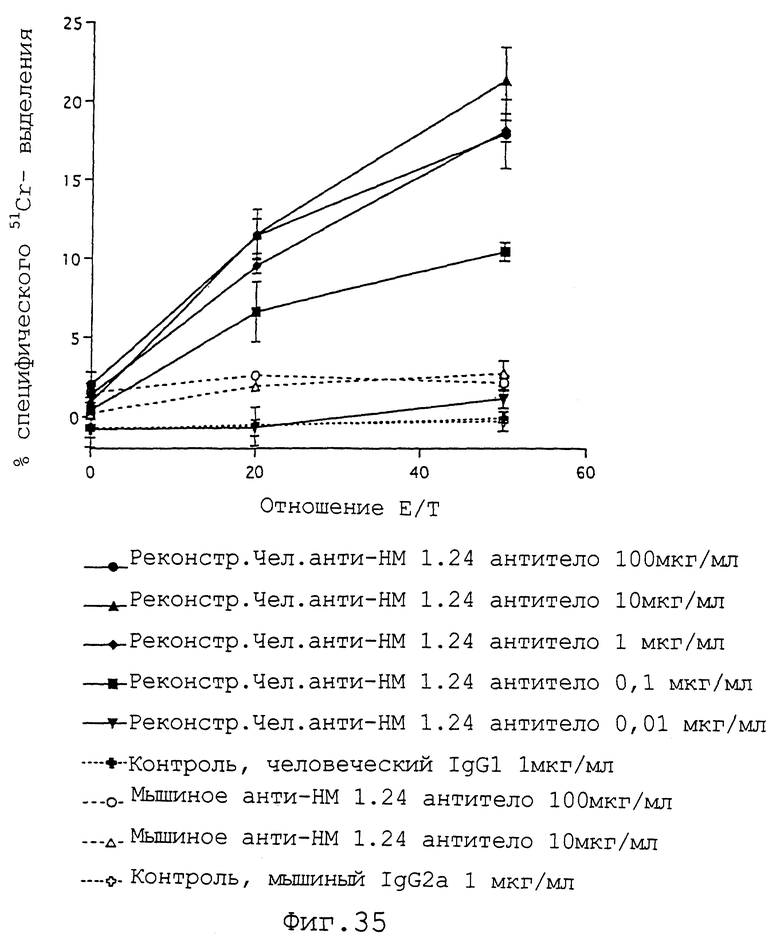
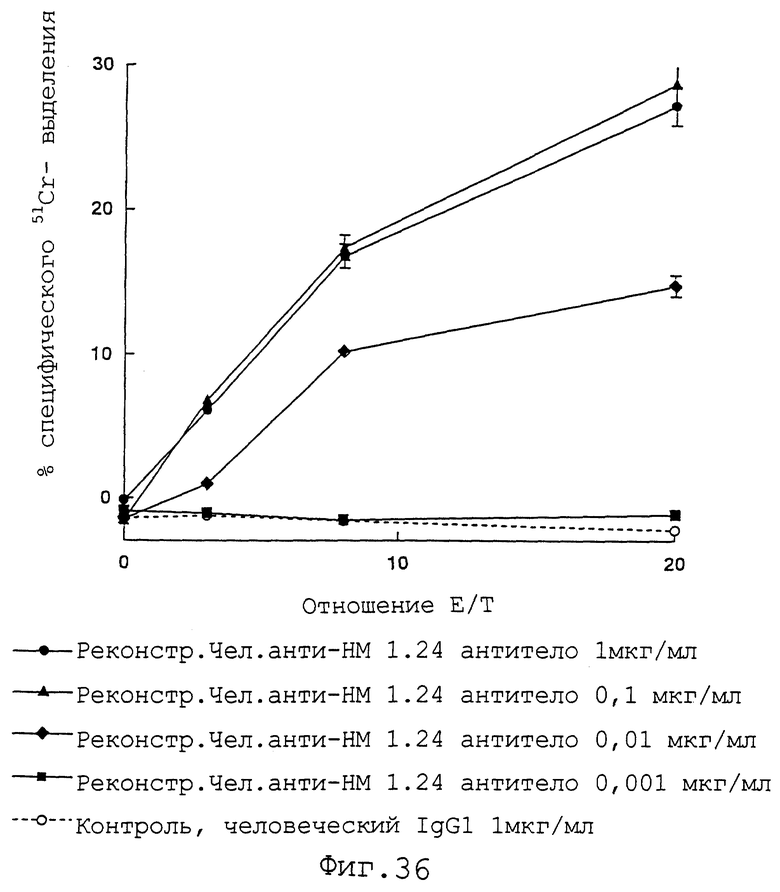
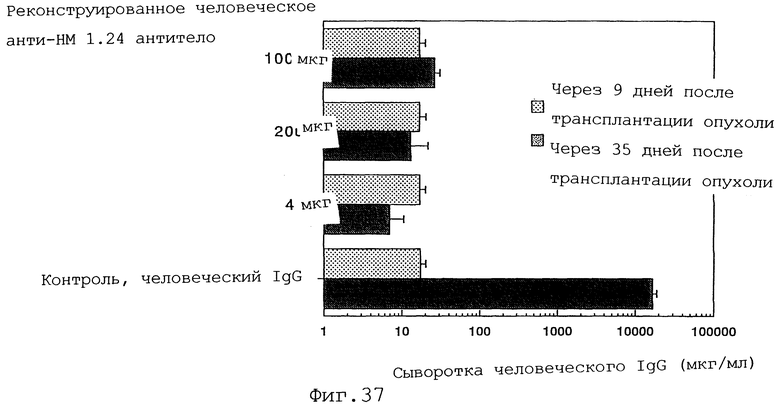
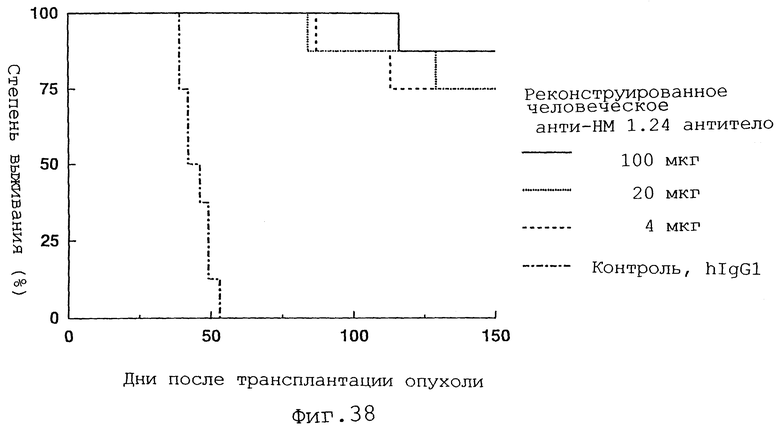
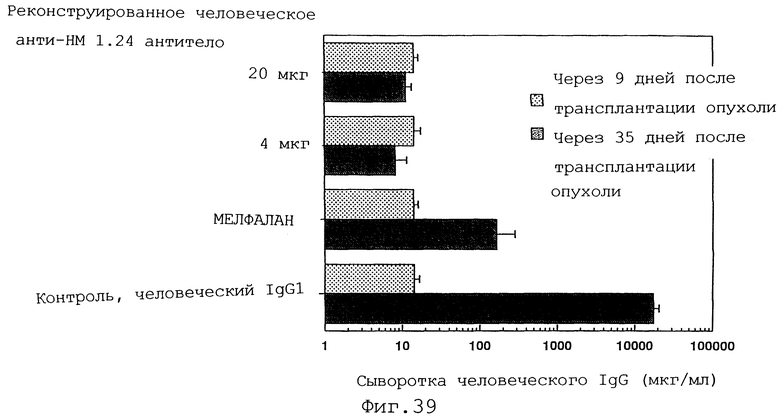
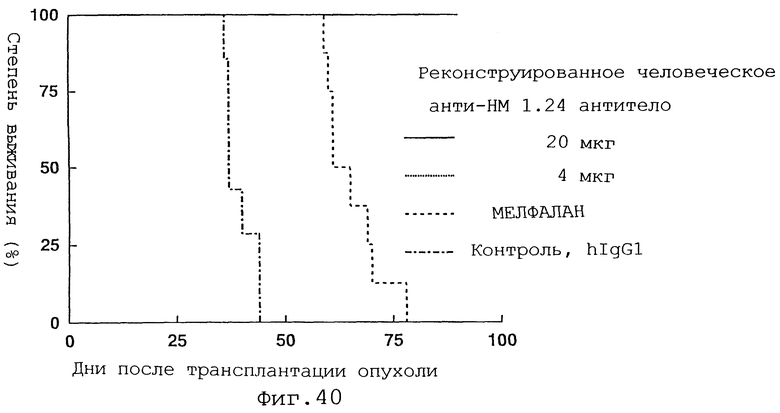
Изобретение относится к области молекулярной биотехнологии. Представлено реконструированное человеческое анти-НМ 1.24 антитело, в котором С-участок Н и L-цепи и V-участок Н и L-цепи включают FR человека, a CDR получают из анти-НМ 1.24 моноклонального антитела. При этом CDR обладает низкой антигенностью. Получают реконструированное человеческое анти-НМ 1.24 антитело путем трансформации клетки-хозяина экспрессирующим вектором, который содержит ДНК, кодирующую антитело. Человеческое анти-НМ 1.24 антитело используется в качестве активного компонента для лечения миеломы. 12 с. и 5 з.п.ф-лы, 40 ил., 6 табл.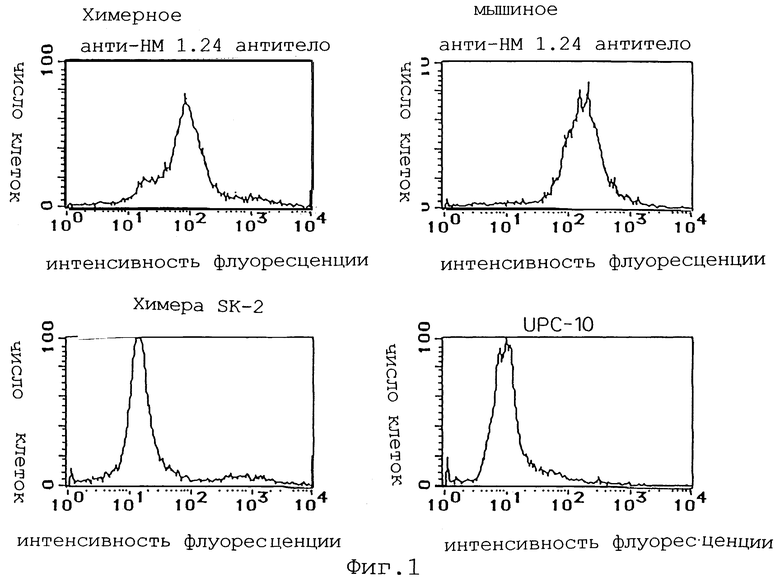
12. Реконструированное человеческое анти-НМ1.24 антитело (FERM ВР-5233), состоящее из реконструированной человеческой L-цепи по п. 10 и реконструированной человеческой Н-цепи по п. 11.
| WO 9219759 А, 24.04.1992. |

