1 Область изобретения
Изобретение относится к библиотеке биоолигомеров, иммобилизованных на твердофазных подложках, где каждая твердофазная подложка связана только с одной биоолигомерной формой, и все возможные комбинации мономерных субъединиц, составляющих указанные биоолигомеры, включены в эту библиотеку. В качестве биоолигомера предлагаемого изобретения можно использовать пептид, олигонуклеотид или химерную пептид-олигонуклеотидную конструкцию. Изобретение относится также к методу синтеза указанной библиотеки.
Изобретение относится, кроме того, к использованию биоолигомеров предлагаемой библиотеки для идентифицирования и характеристики лигандов, обладающих способностью связывать акцепторную молекулу или передавать исследуемую биологическую активность. Биоолигомеры предлагаемой библиотеки могут также ускорять химическую реакцию.
2. Предпосылки создания изобретения
При распознавании и связывания лигандов осуществляется регуляция почти всех биологических процессов, таких как иммунное распознавание, передача сигнала клеточным рецепторам, транскрипция и трансляция, внутриклеточная передача сигнала и катализ, то есть ферментативных реакций. Длительное время в данной области знаний интерес представляет проблема, связанная с распознаванием акцепторных молекул, выступающих в качестве агонистов или способных к супресссии и ингибированию биологической активности лигандов, например, как гормоны, факторы роста и нейромедиаторы; индуцирующих B-клеточный (антитело-опосредованный) или T-клеточный (клеточно-опосредованный) иммунитет; обладающих способностью катализировать химические реакции; и регулирующих экспрессию генов на уровне транскрепции или трансляции.
Особенный интерес представляют белковые и пептидные лиганды. К ним относится большая часть гормонов, факторов роста, нейрореактивные молекулы и иммуннодоминантные детерминанты. Более того, как указывалось ранее, основные усилия по созданию антагонистов или агонистов рецепторно-опосредованной биологической активности, либо антител или T-клеточных антигенных детерминант сконцентрированы на пептидах. Созданию лекарственных веществ, распознающих участки связывания рецептора, однако, препятствовала в значительной степени трудность в определении последовательности пептидных лигандов. Количество вариаций и множество таких пептидных последовательностей сделало невозможным достижение указанной цели любыми методами, за исключением использования трудоемкой методики, заключающейся в том, что выделяют специфичный комплекс, идентифицируют месторасположение эпитопа и секвенируют указанный эпитоп. Решение указанной задачи затрудняется также тем, что указанная антигенная детерминанта состоит из аминокислотных остатков, которые не совпадают в первичной последовательности.
Некоторые исследователи в данной области попытались преодолеть этот трудоемкий процесс путем определения аминокислотной последовательности на основе последовательности нуклеотидов его комплемента. Белки представляет собой высокомолекулярные пептиды, состоящие из аминокислоты; каждую аминокислоту копируют один или несколько кодонов, состоящих из трех остатков нуклеиновых кислот. Так, например, пептид A, содержащий глутамин, кодирует кодон из трех остатков нуклеиновых кислот: цитозина, аденина и гуанина. Комплементом к указанному кодону является гуанин (связываемый с цитозином), тимин (связываемый с аденином) и цитозин, кодирующий аминокислоту в пептиде B. В соответствии с теорией комплементарности указанный пептид B связан с пептидом A. В частности, Bost and Blalock (1989, Hethods in Enzymology, вып. 168, стр. 16-28) полагают, что любой заданный пептид связывается с пептидом, кодирующим комплементарную последовательность остатков нуклеиновых кислот, и на основе этой информации они предугадали аминокислотную последовательность любого комплементарного пептида. Эти авторы использовали указанную последовательность в пептидном синтезе и в исследованиях способности пептида к связыванию.
Этот метод, однако, не обеспечивает решение указанной задачи вследствие того, что степень сродства связывания между комплементарными пептидами обычно невысока и необходимо наличие комплементарных пептидов, содержащих более 15 остатков нуклеиновых кислот. Более того, для этого метода необходима информация либо об аминокислотной последовательности, либо о последовательности связующего партнера анализируемого белка. Этот метод, кроме того, не эффективен для антигенных детерминант, состоящих из аминокислотных остатков, которые не совпадают в первичной последовательности. В последнее время появилось несколько сообщений о получении библиотек пептидов и их использовании для идентифицирования пептидных лигандов, которые могут связываться с акцепторами. В одном способе используют рекомбинантный бактериофаг для создания крупных библиотек. При использовании "фагового метода" (Scott и Smith, 1990, Science, вып. 249, стр. 386-390; Cwirla и др., 1990, Proc Natl. Acad Sci. , вып. 87, стр. 6378-6382; Devlin и др., 1990, Science, вып. 249, стр. 404-406) можно сконструировать очень крупные библиотеки (106-108 химических единиц), но генетический код и биосистема накладывают жесткие ограничения на универсальность и разнообразие их конструкции. Во второй методологии используют, главным образом, химические методы синтеза, в качестве примеров которых можно указать метод Гейсена (Geysen и др., 1986, Molecular Immunology, вып. 23, стр. 709-715; Geysen и др., 1987, J. Immunologic Method, вып. 102, стр. 259-274) и самый современный метод Fodor и др., Science, вып. 251, стр. 767-772, 1991). Методологии Geysen и др. предусматривает возможность синтеза ограниченного количества пептидов на полиэтиленовых штырях за несколько дней. В методе Fodor и др. используют технологию "светоиндуцируемого пространственно адресуемого параллельного химического синтеза. Эта технология также имеет ограниченные возможности из-за относительной нехватки разработок фотохимических методов пептидного синтеза.
Разработаны также технологии крупномасштабного параллельного конкурентного пептидного синтеза. В публикации Houghton сообщается о синтезировании одновременно сотен аналогичных пептидов в полипропиленовых сетчатых пакетах (метод чайного мешочка (Houghton, 1985, Proc Natl. Acad. Sci. USA, вып. 82, стр. 5131-1535; Berg и др., 1989, J. Am. Chem. Soc., вып. 111, стр. 8024-8026) сообщает о новой подложке на основе пленки из полистирол-привитого полиэтилена, пригодной для проведения параллельного пептидного синтеза. В обоих технологиях использована стандартная Boc-аминокислотная смола и традиционные операции снятия защиты, нейтрализации, связывания и промывания согласно оригинальной методике твердофазного синтеза Merrifield (1963, J. Am. Chem. Soc., вып. 85, стр. 2149-2154).
В публикации Furka и др. (1988, 14 th International Congress of Biochemistry, x. 5, Abstract FR:OB) описан способ получения пептидной смеси связыванием по отдельности каждой из трех различных аминокислот с последующим смешиванием всех трех различных аминокислот, содержащих указанную смолу. В раскрываемой методике Glysen и др. не предлагается никакого эффективного способа для выделения исследуемого пептида из множества полученных пептидов.
Несмотря на практическую пригодность, химические методы синтеза Glysen, Fodor, Houghton, Berg, Furka и соавторов позволяют синтезировать и анализировать одновременно только пептиды, содержащие от нескольких сотен до менее тысячи аминокислотных последовательностей. Указанные методы весьма ограничены в свете миллионов возможных пептидных последовательностей, одна или несколько из которых должны соответствовать сайтам связывания между анализируемыми структурными единицами. В случае пептидов из 20 аминокислот, в любой последовательности из пяти аминокислот, имеется 205 или примерно 3,2•106 возможных аминокислотных комбинаций. Ни один из указанных методов не позволяет осуществлять синтез за один раз такого большого количества пептидов. Аналогично этому, общеизвестные технологии пептидного синтеза, например, которую описывают Stewart и Goung (1984, Solid Phase Synthesis, Second Edition, Pierce chemical Co. Rock ford, IN) не предлагают метода синтеза от нескольких тысяч до нескольких миллионов пептидов за один раз.
Более того, ни один из других известных методов пептидного синтеза не предлагает получение библиотеки пептидов, иммобилизованных на твердофазном носителе, которая абсолютно рандомизирована. Абсолютно рандомизированная пептидная библиотека представляет собой банк с хорошим статическим распределением всех молекулярных структур, в результате чего указанная библиотека содержит примерно эквимолярные соотношения всех индивидуальных видов пептидов.
Синтез абсолютно рандомизированного пептида обычно нельзя обеспечить в условиях одновременного введения различных аминокислот в одностадийный реактор из-за того, что скорость связывания различных аминокислот существенно отличается в процессе проведения твердофазного пептидного синтеза (ТФПС) (Ragnarsoon и др., Acta Chem. Scand. вып. 25, стр. 1487, 1489; Ragnarsson и др. , 1974, J. Org. Chem, вып. 39, стр. 3837-2842). Так, например, скорость связывания Fmoc-глицина с растущей петидной цепью гораздо быстрее по сравнению с Fmos-валином возможно из-за стерической затрудненности, обусловленной объемистой боковой цепью валина. В случае, когда приходилось смешивать с полимером все 20 активированных L-аминокислот эукариот в течение каждого цикла связывания, то в указанный пептид в первую очередь включались наиболее быстро вступающие в реакцию аминокислоты, и при этом нельзя было получить эквимолярного соотношения каждой пептидной формы. Кроме того, каждый из возможных нуклеофилов обычно имеет разную реакционную способность.
Более того, ни один из известных методов пептидного синтеза не обеспечивает создание библиотеки из более 105 пептидов, где только один вид пептида иммобилизован на одном твердофазном носителе. Иммобилизация только одного вида пептида на носителе обычно значительно повышает эффективность существующих в настоящее время методик выделения пептидов.
Таким образом, имеется необходимость в данной области знаний в создании библиотеки, состоящей из абсолютно неупорядоченных пептидных и олигонуклеотидных последовательностей, то есть последовательностей биоолигомеров, где только одну олигомерную форму можно выделить легко и быстро из остальных компонентов этой библиотеки. Кроме того, существует готовность в данной области знаний в создании метода для проведения быстрого и дешевого синтеза от нескольких тысяч до нескольких миллионов указанных абсолютно рандомизированных биоолигомерных последовательностей.
3. Краткое изложение сущности изобретения
Изобретение относится к библиотеке биоолигомеров, содержащей все возможные комбинации субъединиц, методом синтеза указанной библиотеки, заключающимся в том, что многократно проводят операции для получения, по крайней мере, двух аликвот твердофазного носителя; к указанным аликвотам твердофазного носителя прибавляют по отдельности набор субъединиц; связывают полностью субъединицу со всеми сайтами твердофазного носителя с образованием комбинации из новой субъединицы и твердофазного носителя, оценивают полноту связывания, и при необходимости продолжают реакцию до полного ее завершения; тщательно перемешивают аликвоты полученной комбинации из новой субъединицы и твердофазного носителя и после повторения вышеуказанных операций необходимое количество раз удаляют защитные группы, в результате чего полученный биоолигомер остается иммобилизованным на твердофазном носителе. В другом варианте осуществления изобретения в качестве соответствующей субъединицы можно использовать аминокислоту, а в качестве биоолигомера пептид. В другом варианте в качестве субъединицы можно использовать нуклеозид, а биоолигомер может быть представлен олигонуклеозидом. Еще в одном варианте осуществления изобретения в качестве нуклеозида используют ДНК, а в еще одном аспекте в качестве нуклеозида используют РНК. В другом варианте осуществления изобретения в качестве субъединицы может быть аминокислота или нуклеозид, а в качестве биоолигомера химерный пептид-олигонуклеотидный комплекс.
Изобретение предлагает метод определения последовательности биоолигомерного лиганда акцепторной молекулы, заключающийся в том, что осуществляют синтез рандомизированной библиотеки биоолигомеров, иммобилизованных на твердофазных носителях, в которой каждый твердофазный носитель связан только с одной биоолигомерной формой, причем все возможные комбинации мономерных субъединиц, из которых состоят указанные биоолигомеры входят в состав этой библиотеки; вводят в указанную рандомизированную библиотеку требуемые акцепторную или субстратную молекулу, в результате чего обеспечивается распознавание и иммобилизация одной или нескольких биоолигомерных субъединиц на твердофазной подложке в пределах указанной библиотеки, либо указанная субстратная молекула претерпевает химическую реакцию, катализированную одной или несколькими комбинациями из биоолигомера и твердофазного носителя; выделяют комбинацию из биоолигомера и твердофазного носителя, проявляющую требуемую способность, и секвенируют биоолигомер выделенной комбинации биоолигомера и твердофазного носителя. В другом варианте осуществления изобретения биоолигомерный фрагмент биоолигомера выделяют из комбинации биоолигомера и твердофазного носителя in situ с последующим детектированием in situ исследуемой биологической активности. В другом варианте осуществления изобретения к биоолигомеру относится пептид. Еще в одном варианте осуществления изобретения в качестве биоолигомера используют олигонуклеотид, в частности ДНК или РНК. И еще в одном варианте осуществления изобретения в качестве биоолигомера используют химерный пептид-олигонуклеотидный комплекс.
Изобретение также предлагает лекарственные и диагностические средства, содержащие биоолигомерные последовательности, установленные в соответствии с вышеуказанными методами.
4. Краткое описание чертежей
На фиг. 1 приведена схема синтеза пептидов, состоящих из рандомизированных последовательностей, с использованием расщепленного метода синтеза неупорядоченного трипептида со вставкой триптофана на в концевом участке цепи: X-X-X W (где X означает S, A или Y; возможно наличие 33 или 27 комбинаций).
На фиг. 2 показаны в схематическом виде циклические пептиды, n = 0, 1, 2, 3, ... и m = 1, 2, 3, ...; n и m могут иметь, хотя не обязательно, одинаковые значения. Сплошные линии показывают связи линейного пептида; штриховые линии означают перекрестные сшивки. Пары специфично сшиваемых субъединиц обозначены A и B. А имеет перекрестные сшивки только с A, а B перекрестно сшивается только с B.
(a) "корзинообразная форма" структуры;
(b) "лестничная" форма;
(c) "лариатная" форма.
На фиг. 3 - хроматограммы (обращеннофазовая ВЭЖХ, система Вилак) неупорядоченных тетрапептидов (X-X-X-W, где X означает S, A или Y), синтезированных: (A) - новым методом (см. текст описания) и (B) - стандартным методом твердофазного пептидного синтеза. Хроматограмма получена при элюировании колонки в линейном градиенте ацетонитрила. Растворитель A:0,1% трифторуксусная кислота и 5% ацетонитрил; растворитель B:0,1% трифторуксусная кислота и 100% ацетонитрил.
На фиг. 4 - снимок пептида/сикросфер с "длинной V-mos последовательностью", меченных антителом анти-V-mos и вторым антителом.
На фиг. 5 - снимок смеси из микросфер с "длинной V-mos последовательностью" и "короткой V-mos последовательностью", меченных антителом анти-V-mos и вторым антителом.
На фиг. 6 - снимок из микросфер с "длинной V-mos последовательностью" и "короткой V-mos последовательностью", меченных антителом анти-V-mos и вторым антителом.
На фиг. 7 - микроснимок скрининга библиотеки типичного пептидного лиганда, где можно легко идентифицировать позитивную микросферу (темно-голубого цвета) на фоне множества тысяч негативных (бесцветных) микросфер.
На фиг. 8 - микроснимок, где показан зависимый от концентрации ингибирующий эффект биотина при окрашивании стрептавидин-щелочной фосфатазой LHPQF-полимер покрытые мимеотопные микросферы. A: 100 нмоль; B: 10 нмоль; C: 1 нмоль и D: 0,1 нмоль биотина. Контрольные микросферы (полимерная композиция на основе бета-Ala-аминокапроновой кислоты) перемешивают в соотношении 1:1 с LHPQF-смолой до инкубации со стрептавидин-щелочной фосфатазой для использования их в качестве внутреннего контроля негатива.
5. Подробное описание изобретения
Далее в описании будут подробно описаны предпочтительные варианты осуществления изобретения.
Используемый в данном описании термин "библиотека" означает банк фактически неупорядоченных биоолигомеров. Используемый в данном описании термин "биоолигомер" означает полимер, содержащий менее примерно 100 субъединиц. В качестве биоолигомера изобретения могут быть представлены любой пептид, состоящий из аминокислотных остатков, или олигонуклеотид, состоящий из нуклеовидов, либо химерный пептид-олигонуклеотидный комплекс.
5.1. Методы получения неупорядоченной библиотеки биоолигомеров.
Как указывалось выше, изобретение относится к методу получения биоолигомерной библиотеки путем синтеза биоолигомеров, состоящих из неупорядоченных последовательностей мономерных субъединиц. Используемый в данном описании термин "неупорядоченная последовательность мономерных субъединиц" означает последовательность, в которой любая мономерная субъединица предшествует или следует за любой другой мономерной субъединицей.
В одном варианте осуществления изобретения в качестве мономерной субъединицы можно использовать аминокислоту, любой аминокислотный аналог или любой пептидный минеотоп. Используемый в данном описании термин "пептидный минеотоп" означает молекулу, которая схожа по структуре и химическому составу с пептидом, состоящим из двух или нескольких аминокислот. В другом варианте осуществления изобретения в качестве мономерной субъединицы можно использовать любой нуклеозид; в качестве нуклеозида можно использовать ДНК или РНК. Еще в одном варианте изобретения в качестве мономерных субъединиц можно использовать аминокислоты и нуклеозиды. В качестве биоолигомера можно использовать пептид (состоящий из аминокислот), олигорибонуклеотид (содержащий дезоксирибонуклеозиды), химерный ДНК-РНК олигонуклеотидный комплекс или любую пептид-олигонуклеотидную химерную структуру. Библиотеку, содержащую пептиды, олигонуклеотиды или пептид-олигонуклеотидные химерные структуры можно получить в соответствии с методом, включающим многократное проведение стадии из операций, заключающихся в том, что:
(i) получают, по крайней мере, две аликвоты твердофазного носителя для рандомизированных последовательностей субъединиц;
(ii) вводят в указанные аликвоты твердофазного носителя по отдельности набор из субъединиц;
(iii) связывают полностью указанные субъединицы практически со всеми сайтами твердофазного носителя, в результате чего образуется новая комбинация субъединиц, иммобилизованных на твердофазном носителе;
(iv) оценивают степень полноты связывания и, при необходимости, продолжают указанную реакцию до полного ее завершения;
(v) тщательно перемешивают аликвоты новой комбинации субъединиц, иммобилизованных на твердофазном носителе; и после повторения операций
(I)-(V) требуемое количество раз проводят заключительную операцию (VI), удаляя защитные группы, в результате чего полученный биоолигомер остается иммобилизованным на твердофазном носителе. В другом варианте осуществления изобретения неупорядоченную биоологомерную библиотеку можно получить таким образом, что в течение, по крайней мере, одной стадии синтеза одна и та же субъединица связывается со всеми участками твердофазных подложек и за, по крайней мере, одну другую стадию, по меньшей мере, две субъединицы иммобилизуются на твердофазной подложке. Неупорядоченная биоолигомерная библиотека может быть синтезирована однократным проведением вышеуказанных операций (i)-(V); в еще одном варианте осуществления предлагаемого изобретения неупорядоченную биоолигомерную библиотеку можно получить многократным проведением вышеуказанных операций (i)-(V). Любая твердофазная подложка может быть получена в комбинации с одной или несколькими предварительно связанными субъединицами.
Биоолигомерная библиотека может состоять из данного, ограниченного количества субъединиц. В другом варианте осуществления изобретения неупорядоченная биоолигомерная библиотека может быть составлена из всех имеющихся в наличии субъединиц.
Еще в одном варианте осуществления настоящего изобретения анализируемый биоолигомер можно идентифицировать в процессинге путем создания первоначально библиотеки и последующей идентификации биоолигомерной последовательности, проявляющей исследуемую способность. В результате получают твердофазную подложку, содержащую идентифицированную, таким образом, биоолигомерную последовательность. Новый сегмент мономерной субъединицы вводят в идентифицированную ранее последовательность, после чего распознают полученный фрагмент, состоящий из известной последовательности и неупорядоченной последовательности, проявляющей исследуемую способность. Такая последовательная стратегия оптимизации-рандомизации позволяет проводить быструю идентификацию анализируемого биоолигомера.
Биоолигомеры предлагаемой библиотеки могут, хотя и не обязательно, содержаться в ней в эквимолярных количествах. Специалисту общеизвестно, что мольное количество означает концентрацию, при которой одну грамм-молекулу любого вещества растворяют в растворителе, достаточном, чтобы получить один литр раствора. Используемый в данном описании термин "практически эквимолярные количества" биоолигомеров относятся к отдельным видам мономерных субъединиц, которые содержатся примерно в одной и той же концентрации. Так, например, если содержание биоолигомера A в наборе, содержащем 150000 биоолигомеров, составляет 200 нмоль/л, то все оставшиеся из 150000 биоолигомерных форм будут присутствовать в нем в концентрации примерно 200 пмоль/л. Однако термин "практически эквимолярное количество", используемый в данном описании, имеет толкование для оценки гетерогенности поверхности твердофазных носителей. Гетерогенность твердофазного носителя приводит к изменению количества биоолигомера, которое может быть иммобилизовано на данном носителе.
В предлагаемом методе используют, по крайней мере, две аликвоты твердофазного носителя, где количество твердофазных носителей в указанных аликвотах предпочтительно соответствует, по крайней мере, количеству синтезируемых биоолигомеров. Это позволяет создавать библиотеку, где каждый твердофазный носитель содержит только один биоолигомерный вид, то есть один биоолигомер - одна микросфера. Используемый в данном описании термин "аликвота" означает часть, представляющую собой определенную фракцию от суммарного количества твердофазных носителей.
5.2. Рандомизированные пептидные библиотеки
В конкретном варианте осуществления настоящего изобретения предлагаемая рандомизированная биоолигомерная библиотека может включать в свой состав пептиды. Термин "пептид" в данном описании в самом широком понятии для обозначения соединения, состоящего из двух или более аминокислот, аминокислотных аналогов или пептидомиметиков в качестве субъединиц. Указанные субъединицы могут быть связаны пептидными связями. В другом варианте осуществления изобретения используемая субъединица может быть связана другими связями, например, сложноэфирной, простой эфирной и т.д. Используемый в данном описании термин "аминокислота" означает как природные и/или неприродные, так и синтетические аминокислоты, в том числе глицин или обе D- или L-формы оптических изомеров, а также аминокислотные аналоги и пептидомиметики. Любой пептид, состоящий из трех или более аминокислот, обычно называют олигопептидом в случае короткой пептидной цепи. При наличии длинной пептидной цепи такой пептид общепринято называют полипептидом или белком.
Настоящее изобретение в своей основе базируется на химии синтетических пептидов и не полагается на любые системы живых организмов для амплификации или скрининга. Пептидные библиотеки настоящего изобретения могут включать в свой состав органические аминокислоты. Так, например, предлагаемые пептиды могут содержать D-аминокислоты, смесь D- и L-аминокислот и различные аминокислотные производные (например, бета-метиламинокислоты, CL-метиламинокислоты, Nα-метиламинокислоты и т.п.) для передачи особых свойств пептидам в предлагаемой библиотеке. Кроме того, при переносе специфической способности аминокислот на стадиях сайт-специфического связывания можно получить пептидные библиотеки, содержащие пептиды альфа-спиральной, бета-изогнутой, бета-складчатой, гамма-изогнутой и циклической конформации.
В состав предлагаемой библиотеки пептидов входит любая возможная комбинация аминокислот, из которых состоят указанные пептиды. Используя в качестве примера пептид, образованный из двух аминокислот, глицина и пролина, возможно существование четырех комбинаций: глицин-глицина, глицин-пролина, пролин-глицина и пролин-пролина и, следовательно, в состав предлагаемой неупорядоченной библиотеки будут входить все эти четыре комбинации.
Набор первых аминокислот вводят по отдельности в каждую аликвоту. Обычно для пептидного синтеза в качестве аминокислот используют основно-подвижные аминокислоты с защитной 9-флуоренилметоксикарбонилом (Fmoc) Nα-аминогруппы, описанные впервые Carpino и Han (1972, J.org Chem, 37, стр. 3403-3409). В предлагаемом методе можно также использовать Boc-аминокислоты (с защищенной Nα-третбутилоксикарбонилом Nα-аминогруппы). Оба Fmos и Boc-аминокислоты можно получать у Fluka, Bachem, Advanced Chemtech, Sigma, Cambredge Research Biochemical, Bachem или Pеninsula Labs, либо в других химических фирмах, известных специалистам в данной области. Кроме того, в предлагаемом методе можно использовать другие известные специалистам Nα-защитные группы.
Возвращаясь к раскрываемому выше примеру дипептида, первый набор интродуцируемых аминокислот обычно содержит глицин и пролин; каждая аликвота принимает либо Nα-Fmos-глицин, либо Nα-Fmos-пролин.
После введения указанный набор первых аминокислот полностью связывается со всеми сайтами твердофазных носителей. Используемый в данном описании признак "полное связывание" означает, что реакция связывания протекает до ее завершения независимо от различий скорости связывания отдельных аминокислот. Более того, указанные аминокислоты связываются практически со всеми имеющимися на твердофазном носителе сайтами связывания, в результате чего каждый твердофазный носитель содержит только один вид пептида. Реакция полного связывания приводит к образованию комбинаций из набора первых аминокислот и твердофазных носителей. При использовании в качестве примера вышеописанного дипептида после завершения связывания получают комбинацию иммобилизованного на микросфере глицина и комбинацию иммобилизованного на микросфере пролина.
Реакцию связывания указанных аминокислот можно осуществлять известными специалистам методами, предлагаемыми, например, Sfewart и Young, 1984, Solid Phase Synthesis, Second Edition, Pierce Chemicol Co, Kockfd, IL. Как известно специалистам в данной области, в технологию пептидного синтеза на твердофазных носителях обычно входит построение пептида из карбоксила или конца C-терминальной области, где C-терминальная аминокислота с защищенной альфа-аминогруппой иммобилизована на твердофазном полимере. Защитную группу затем удаляют, и следующая аминокислота, также с защитой, связывается посредством пептидной связи с альфа-аминогруппой аминокислоты, иммобилизованной на твердофазном носителе. Цикл снятия защиты предыдущей аминокислоты и связывания следующей аминокислоты повторяют вплоть до окончания синтеза пептида. Любые реакционноспособные боковые цепи защищают химическими группами, которые могут выдерживать операции связывания и Nα-разблокировки, но которые можно удалить в конце синтеза.
Для связывания любой аминокислоты с растущей синтетической цепью необходимо провести активирование карбоксильной группы блокированной аминокислоты. В практике изобретения можно использовать различные методы активации, которые включают, например, предварительно приготовленные симметрические антигидриды (in situ), предварительно приготовленную ангидридную смесь (РМА), хлорангидриды, реакционноспособные сложные эфиры и активацию карбоновой кислоты по методике, описанной Fields и Noble, 1990, "Solid Phese Peptide Syntbesis utilizing 9-флуоренилметоксикарбонил аминокислоту" (твердофазный пептидный синтез с использованием 9-флуоренилметоксикарбонил аминокислот) Int. J. Pept. Protech. Res, 35, 161-214.
Необходимо оценивать полноту связывания. Специалисты в данной области хорошо знают известные реакции при проведении количественного анализа с использованием, например, нингидрина (проба Кайзера), пикриновой кислоты, 2,4,6-тринитробензолсульфокислоты (TNBS), фторэскамина и хлоранила, в основе которых лежит взаимодействие реактивов со свободными аминогруппами с образованием хромофора. В случае использования иминокислот (например, Pro и Hyp) количественный контроль с использованием изатиновой реакции предпочтительный метод анализа (см. выше Fields и Noble). Количественный анализ или оценки степени завершенности реакции связывания можно проводить в течение всего времени синтеза, например, как описывают Salisbury и др. (заявка по договору о патентной кооперации PCT N 91/03485).
При проведении Fmos-аминокислотного синтеза предпочтительно использование пробы Кайзера. В тексте Кайзера образец из каждой пробирки можно проанализировать при использовании нингидринового реагента, поставляемого фирмой Pierce Chemicol, по методике Savine и др. (1981, Anal. Biochem, 117, стр. 147-157).
Если реакция связывания произошла неполностью, как установлено в результате этого теста, то указанную реакцию можно довести до ее завершения несколькими методами, которые известны специалистам, в том числе (a) проведение второй реакции связывания с использованием пятикратного избытка защищенной аминокислоты, (b) аддитивного связывания с использованием других или дополнительных растворителей (например, трифторэтана) или (c) прибавление хаотропных солей, например, NaClO4 или hcBr (Klis и Stewart, 1990, "Peptides: Chemistry, Structure and Biology", Rivier and Harshall, eds., ESCOM Publ., стр. 904-906).
После полного связывания аликвоты комбинаций первых аминокислот твердофазного носителя тщательно перемешивают. Тщательного перемешивания достигают при условии, когда получают гомогенную смесь аликвот предпочтительно при перемешивании указанных аликвот в одностадийном реакторе. Хотя любой метод тщательного перемешивания входит в объем настоящего изобретения, а специалисты хорошо знают множество других средств, однако к предпочтительным можно отнести, например, вихревое или ротационное перемешивание в любом имеющемся в продаже роторном шюттль-аппарате или барботирование инертным газом, например, азотом или аргоном.
Полученную смесь делят, по крайней мере, на две аликвотные части. Эти аликвотные части равны по объему и, если достигнута достаточная гомогенность, должны содержать практически равные количества комбинаций первых аминокислот на твердофазных носителях. В случае дипептида, используемого в качестве примера, каждая аликвота содержит фактически равные количества комбинаций микросфера-глицин и микросфера-пролин.
В каждую аликвоту вводят по отдельности второй набор аминокислот. Этот второй набор может состоять из (a) тех же самых аминокислот, которые были введены в первом наборе, то есть глицина или пролина; (b) другого набора аминокислот, например, триптофана или лейцина; (c) только одного типа аминокислот, например, изолейцина.
Как и в случае первого аминокислотного набора, второй набор аминокислот полностью связывается по отдельности с комбинацией первых аминокислот, иммобилизованных на твердофазных носителях из каждой аликвоты с образованием пептидов, содержащих первую аминокислоту и вторую аминокислоту. Так же, как и в случае предыдущей реакции связывания, следующее связывание можно выполнять любым методом, используемым в данной области для проведения таких реакций. Используя в качестве примера вышеобсуждаемый дипептид:
(a) при введении тех же самых аминокислот полученные пептиды представляют комбинации как глицин-глицин, глицин-пролин, пролин-глицин, так и пролин-пролин;
(b) при наборе других аминокислот полученные пептиды представляют комбинации либо Gly-Trp, Gly-Leu, Pro-Trp, либо Pro-Leu;
(c) при введении одного типа аминокислот полученные пептиды представляют Gly-ILe или Pro-ILe.
Эту методику связывания можно повторять столько же раз, сколько приходится вводить аминокислот. Если интересующий пептид представляет собой тетрапептид X-X-X-T, где Х означает либо валин, серин, либо аланин, например, то указанную методику связывания можно проводить трехкратно для получения тетрапептида со структурной цепи X-X-X-Trp. При первом, втором и третьем введениях аминокислот либо Nα-Fmos-валин, Nα-Fmos-серин (O-Bu'), либо Nα-Fmos-аланин добавляется к аликвотам твердофазного носителя с выходом 27 различных пептида фактически в эквимолярных количествах (фиг. 1). Если необходимо получение гексапептида, указанную методику связывания повторяют шесть раз. Если гексапептид должен включать в свой состав 5 разных аминокислот, то указанный метод можно осуществлять при использовании 5 аликвот, каждая из которых содержит другую кислоту на каждой стадии связывания. Если, однако, целевой гексапептид должен содержать любую комбинацию из главной цепи из аминокислот, указанный метод можно проводить при использовании 20 аликвот на каждой стадии связывания.
Предлагаемый метод пептидного синтеза можно использовать с твердофазными носителями, на которых любая аминокислота либо уже иммобилизована, либо еще нет. Кроме того, возможно использование линкера, который уже связан с твердофазным носителем. В качестве одной общей подложки, на которой уже иммобилизована любая аминокислота, можно использовать бета-аланин-полиакриламидную смолу (поставляемую фирмой Bachem Biochemical). Такие смолы можно получить из многочисленных коммерческих источников или получить в лаборатории любым специалистом в области пептидного синтеза.
Если смесь аминокислот, иммобилизованных на твердофазном носителе или линкер на твердофазной подложке используют в качестве исходного реагента, то ее разделяют, по крайней мере, на две аликвоты, каждая из которых содержит любую аминокислоту из первого набора аминокислот. Как указывалось выше, первый набор аминокислот полностью связывается практически со всеми участками связывания иммобилизованной на твердофазном носителе смеси аминокислот, либо на твердофазном линкере, после чего аликвоты, содержащие эти заново введенные аминокилсоты, тщательно перемешивают. Как указывалось ранее, полученную смесь делят, по крайней мере, на две аликвоты, при этом каждая аликвота получает одну аминокислоту из второго набора аминокислот, и реакцию связывания проводят повторно с образованием растущего пептида. Как уже говорилось ранее, указанный процесс можно повторять столько раз, сколько это необходимо для получения требуемых пептидов.
Указанный метод можно использовать для синтеза неупорядоченных пептидов, а также для синтеза пептидной библиотеки, содержащей заранее установленные последовательности. Синтез заранее установленных последовательностей включает использование специфических Nα-Boc, Nα-Fmos или других защищенных надлежащих образом аминокислот во время операций сайт-специфического связывания. Например, любой может подобрать аминокислоты на стадиях сайт-специфического связывания так, чтобы целевые пептиды имели вероятность и предпочтительность получения особой вторичной структуры, например, как бета-складка, альфа-спираль, бета-изгиб и т. п. Так, например, альфа-спираль предпочтительна, если Glu, Ala, Leu, His, Trp используют в качестве предпочтительных аминокислот; с другой стороны бета-складки обычно предпочтительны в случае использования Val, ILе, Tyr и Met. В альтернативном варианте, если используют Gly, Asp, Ser, Pro, Asp, то предпочтительна бета-изогнутая структура. В качестве других примеров можно указать, например, кислые аминокислоты у N-концевой области и основные аминокислоты у C-концевой области, которые стабилизируют альфа-спираль. D-аминокислоты могут стабилизировать некоторые изогнутые структуры, и многочисленные другие структурные формы могут быть включены в предлагаемую библиотеку (см. ниже Разделы 5.2.1 и 5.2.2). Вполне даже вероятен синтез библиотеки циклических пептидов с дисульфидными, лактамными, лактоновыми или другими замыкающими цикл остатками (см. ниже Раздел 5.2.1).
Следует подчеркнуть, что предлагаемый метод позволяет проводить синтез пептидов так, что каждая твердофазная подложка, например, как полимерная микросфера, содержит только один вид пептида. Указанный метод обеспечивает контакт отдельной полимерной микросферы только с одной Fmos-аминокислотой во время каждого цикла связывания и протекание указанной реакции вплоть до полного ее завершения. Указанный метод синтеза одна микросфера - один пептид позволяет повысить чувствительность и эффективность выделения пептида, который специфичен к субъединице, с которой он связывается.
Предлагаемый метод можно легко реализовать для синтеза рандомизированного пептидного пула, содержащего от 105 до 107 различных пептидных форм.
В одном аспекте предлагаемого изобретения пептиды, входящие в состав библиотеки, могут содержать специальный аминокислотный остаток у C-концевого участка с либо COO H, либо CONH боковой цепью, предназначенную для имитирования свободного глицина или глицин-амидной группы. Более того, этот специальный аминокислотный остаток обычно используют в виде D- или L-аминокислотного аналога с боковой цепью, содержащей линкер или связь к микросфере. В другом варианте указанный псевдосвободный аминокислотный остаток у C-концевого участка можно использовать в виде оптического изомера D- или L-конфигурации; в еще одном варианте - в виде рацеамической смеси, содержащей D- и L-изомеры.
В еще одном варианте осуществления изобретения пептиды предлагаемой библиотеки могут содержать пироглутамат в качестве N-концевого аминокислотного остатка. Хотя пироглутамат не поддается секвенированию методом деградации Эдмана, однако при ограничении замены пептидов с N-концевым пироглутаматом только до 50% на любой заданной микросфере, достаточное количество несодержащего пироглутамат пептида остается на указанной микросфере для секвенирования. Любому специалисту должно быть ясно, что указанный метод можно использовать для секвенирования любого пептида, который содержит аминокислотный остаток в N-концевой области, устойчивый к деградации Эдмана. Другие методы для характеристики индивидуальных пептидов, проявляющих требуемую активность, описаны подробно ниже в описании. Специфическая активность пептида, содержащего защищенную N-концевую группу, например, пироглутамат, если такая N-концевая группа присутствует в 50% указанных пептидов, может быть легко продемонстрирована при сравнивании активности полностью защищенного (100%) пептида с незащищенным (0%) пептидом.
В другом варианте предлагаемого изобретения выбирают субъединицы пептидов, несущие полезные химические и структурные свойства. Так, например, пептиды, содержащие L-аминокислоты устойчивы к L-амино-кислотоспецифичным протеазам in vivo. Кроме того, настоящее изобретение рассматривает проблему получения библиотек с более четко установленными структурными характеристиками, а также использования пептидомиметиков и пептидомимеотические связи, например, как сложноэфирные связи для получения пептидных библиотек с новыми свойствами. В другом варианте осуществления изобретения можно создать пептидную библиотеку, включающую редуцированную пептидную связь, то есть Ri-CH2-NH-R2, где R1 и R2 означают аминокислотные остатки или последовательности. Редуцированную пептидную связь можно вводить в виде дипептидной субъедицины. Такая молекула обычно резистентна к расщеплению пептидной связи, например, протеазной активности. Такие библиотеки создают лиганды с уникальной функциональной способностью и активностью, например, как пролонгированный полупериод жизни in vivo, обусловленный устойчивостью к метаболическому распаду или протеазная активность. Более того, общеизвестно, что в некоторых биологических системах конформационно ограниченные пептиды проявляют повышенную функциональную активность (Hruby, 1982, Kife Sciences вып. 31, стр. 189 -; Hruby и др., 1990, Biochem J., 268, стр. 249-262) настоящее изобретение предлагает способ для получения конформационно-ограниченного пептида, содержащего неупорядоченные последовательности во всех других положениях.
Ограниченные и циклические пептиды.
Любой ограниченный, циклический или рандомизированный пептид можно получить в соответствии с вышеописанным методом при условии, что, по крайней мере, в два положения в последовательности любого из пептидов библиотеки вставлена аминокислота или аминокислотный аналог, что дает химическую функциональную группу, обладающую способностью к сшиванию, приводящему к конформационному ограничению, циклизации и жесткому каркасу пептида после обработки с образованием поперечных связей. Реакция циклизации будет протекать благоприятно при введении любой изгиб-индуцирующей аминокислоты. К примерам аминокислот, которые могут сшивать пептид, относятся цистеин с образованием дисульфидов, аспарагиновая кислота, образующая лактон или лактам и любой халатирующий агент, например, гамма-карбоксил-глутаминовая кислота (61a) (производимая фирмой Bachem), дающая хелатный комплекс переходного металла с образованием мостиковой связи. Защищенную гамма-карбоксил-глютаминовую кислоту можно получить модифицированием технологии синтеза, которую описывают Zee-Cheng и Olson (1980, Biophys. Biochem. Res. Commun., вып. 1128, стр. 1128-1132). Любую пептидную библиотеку, в которой указанная пептидная последовательность содержит, по крайней мере, две аминокислоты, обладающих способностью сшивания, можно обработать, например, окислением цистеиновых остатков с образованием дисульфида, или введением иона металла с образованием хелатного комплекса, в результате чего происходит сшивание указанного пептида, при этом образуется конформационно ограниченный, циклический пептид или пептид с жестким скелетом.
Настоящее изобретение предлагает несколько основных жестких скелетов для использования их в синтезе предлагаемых библиотек. В одном варианте осуществления изобретения, показанного на фиг. 2, две пары сшивающих остатков располагают в таком порядке, что образуется каркас типа "корзина". Такой "корзинообразный" скелет можно использовать, в частности, в качестве каталитического кармана. В другом варианте осуществления изобретения из двух пар сшивающих аминокислотных остатков можно построить "лестничный" скелет, который показан на фиг. 2. При попеременном использовании D- и L-аминокислот в "лестничной" основе можно получить пептид, где каждая из боковых цепей ориентирована на одной поверхности аналогично бета-складчатой конформации, установленной в грамицидине. Такая поверхность может потенциально образовывать уникальный каталитический центр. Если в одном варианте осуществления изобретения можно создать простой "веревочный" скелет, в котором два остатка образуют мостиковую связь, как показано на фиг. 2. В дополнение к полученной пептидной петле, укороченный "веревочный" скелет приводит обычно к образованию конформационно ограниченного линейного пептида и, следовательно, стабилизации вторичной структуры, например, альфа-спирали.
В предлагаемом изобретении также установлено, что межпептидные сшивки можно получать с образованием жесткого пептидного каркаса.
Настоящее изобретение предлагает стратегии для системного получения мостиковых связей. Так, например, при введении цистеиновых остатков в последовательность такого пептида, можно использовать различные защитные группы (HisKey, 1981, In The Peptides: Analysis, Synthesis, Biology, v. 3, Gross и Meienhofer, изп. Academic Press: New York, стр. 137-167; Ponsanti и др., 1990, Tetrahedron, вып. 46, стр. 8255-8266). С первой пары цистеиновых остатков можно снять защиту и подвергнуть окислению, а затем можно разблокировать и окислить вторую пару. В результате такой методики можно образовать несколько дисульфидных мостиков. В альтернативном варианте пару цистеинов и пару хелатообразующих аминокислотных аналогов можно вводить в такую пептидную последовательность, в результате чего образуются мостиковые связи разной химической природы.
5.2.2. Нетрадиционные аминокислоты, вызывающие конформационные ограничения
Следующие нетрадиционные аминокислоты можно включать в состав рандомизированной пептидной библиотеки с целью введения характерных конформационных каркасов: 1,2,3,4-тетрагидроизохинолин-3-карбоксилат (Kazmierski и др. , 1991, J.Am.Chem.Soc., вып. 113, стр. 2275-2283); (2S, 3S)-метилфенилаланин, (2S, 3R)-метилфенилаланин, (2R, 3S)-метилфенилаланин и (2R, 3R)-метилфенилаланин (Kazmiercki и Hruby, 1991, Tetrahedron Lett). 2-аминотетрагидронафталин-2-карбоновая кислота (Landis, 1989, Ph.D.Nhesis, University of Arisona), гидрокси-1,2,3,4-тетрагидроизохинолин-3-карбоксилат (Miyake и др., 1989, J.Takeda Res. Labs, вып. 43, стр. 53-76); бета-карболин (D- и L-формы) (Kazmierski, 1988, Ph. D.Thesis, University of Arisona); HIC (гистидин-изоколин-карбоновая кислота) (Zechel и др., 1991, Int. J. Pep. Perotein Res. 43) и HIC (гистидинцикломочевина) (Dharanipragada).
Следующие аминокислотные аналоги и пептидомиметики, которые приводят к образованию или выгодной организации специфических вторичных структур, можно включать в такую рандомизированную библиотеку: LL-Acp (LL-3-амино-2-пропенидон-6-карбоновая кислота), бета-изгиб-индуцирующий дипептидный аналог (Kemp и др., 1985, J.Org.Chem., вып. 50, стр. 5384-5838); аналоги, индуцирующие бета-складчатую конформацию (Kemp и др., 1988, Tetrahedron Lett, вып. 29, стр. 5081-5082); аналоги, индуцирующие бета-изогнутую конформацию (Kemp и др. , 1988, Tetrahedron Lett, вып. 29, стр. 5057-5060); альфа-спираль-индуцирующие аналоги (Kemp и др., 1988, Tetrahedron Lett, 29, стр. 4935-4938); гамма-изгиб-индуцирующие аналоги (Kemp и др., 1989, J.Org.Chem. вып. 54, стр. 109-115); и аналоги, раскрываемые в следующих ссылках: Nagai и Sato, 1985, Tetrahedron Lett, 26, стр. 647-650; DiMaio и др., 1989, J.Chem. Soc. Perkin Trans. , стр. 1687; кроме того аналог Gly-Ala- изогнутой конформации (Kahn и др., 1989, Tetrahedron Lett, вып. 30, стр. 2317); изостеры с амидной связью (Jones и др. , 1988, Tetrahedron Lett., вып. 29, стр. 3853-3856); третразол (Zabrocki и др., 1988, J.Am. Chem. Soc., вып. 58, стр. 5875-5880); DTC (Samanen и др., 1990, Int. J. Protein Pep. Res., вып. 35, стр. 501-509); и аналоги, раскрываемые Olson и др., 1990, J.Am. Chem. Sci, вып. 112, стр. 323-333 и
Garvey и др., 1990, J.Org. Chem., вып. 56, стр. 436. Хотя вышеуказанные нетрадиционные пептиды и пептидомиметики возможно не способны к секвенированию общепринятым методом деградации Эдмана, для определения структуры пептида с исследуемой активностью можно использовать сочетание первоначального проведения деградации Эдмана с последующим анализом аминокислот оставшейся цепи. В альтернативном варианте можно использовать масс-спектральный анализ.
5.2.3. Дериватизированные и модифицированные пептиды
Настоящее изобретения также предусматривает модификацию пептидов или получение их производных в библиотеке. Специалисты хорошо знакомы с приемами модификации пептидов, включая фосфорилирование, карбоксиметилирование и ацилирование.
В другом варианте осуществления изобретения можно получить гликозилированные или жирные ацилированные пептидные производные. Известно получение гликозилированных или жирных ацилированных пептидов, которое приведено в следующих ссылках:
1. Garg and Jeanloz, 1985, in Advances in Cerbohydrate Chemistry and Biochemistry, Academic Press.
2. Kunz, 1987, in Ang. Chem. Int. Ed. English 26: 294-308.
3. Horvat et al., 1988, Int. J. Pept. Protein Res. 31: 499-507.
4. Bardaji et al., 1990, Ang. Chem. Int. Ed. English, 23: 231.
5. Toth et al. , 1990, in Peptides: Chemistry, Structure and Biology, Rivier and Marshal. eds., ESCOM Publ., Leiden, pp. 1078-1079.
6. Torres et al., 1989, Experientia 45: 574-576.
7. Torres et al., 1989, EMBO J. 8: 2925-2932.
8. Hordever and Musiol, 1990, in Peptides: Chemistry, Structure and Biology, loc. cit., pp. 811-812.
9. Zee-Cheng and Olson, 1989, Biochem, Biophys, Res. Commun. 94: 1128-1132.
10. Merki et al., 1977, Helv. Chem. Acta., 60:807.
11. Fuji et al. 1987, J. Chem. Soc. Chem. Commun., pp. 163-164.
12. Ponsati et al., 1990, Peptides 1990, Giralt and Andreu, eds., ESCOM Publ., pp. 238-240.
13. Fuji et al., 1987, 1988, Peptides: Chemistry and Biology, Merahall, ed., ESCOM Publ., Leiden. pp. 217-219.
Существует два главных класса пептид-углеводных связей. Во-первых, сложноэфирные группы соединяют гидроксил серина или треонина с гидроксильной группой углевода. Во-вторых, амидные связи соединяют карбоксильные группы глутамата или аспартата с аминогруппой у углевода. В частности, в вышеуказанных ссылках 1 и 2 описаны методы получения пептид-углеводных простых эфиров и амидов. Ацетальные и кетальные связи могут также связывать углевод с пептидом.
Более того, можно также получить жирные ацилированные пептиды. Так, например, но не в коей мере не ограничивая объем, в свободную аминогруппу (в N-конце или лизил) можно ввести ацильную группу, например, ниристоил. В альтернативном варианте аминокислоту, содержащую алифатическую боковую цепь структуры (CH2)n(CH3) можно вводить в пептиды предлагаемой библиотеки. Этот и другие конъюгаты пептидов и жирными кислотами, пригодными для использования в настоящем изобретении, раскрыты в патенте Великобритании GB-8809162.4; Международной патентной заявке PCT/AU 89/00166 и противопоставленном источнике 5, см. выше.
5.3. Рандомизированные олигонуклеотидные библиотеки
Метод получения селективной библиотеки, состоящей из нуклеиновых кислот, может брать за основу твердофазный синтез ДНК по фосфорамидатному механизму, предложенному впервые Caruthers (1985, Science, вып. 230, стр. 281; Caruthers и др., 1987, Methods in Ensymology, вып. 154, стр. 287-313).
Как нерастворимый полимерный номитель на основе кремнезема, так и защитные дезоксинуклеозиды поставляются на рынок (например, Penin Sula Laboratories, Jnc. , California, Applied Biosystems. Jnc.). В качестве защищенных дезоксинуклеозидов можно использовать 5'-0-диметокситритилдезокситимидин, 5'-0-диметокситритил-4-N-бензоселдезоксиаденозин и 5'-0-диметокситритил-N-изобутилдезоксигуанозин. Другие специфические защитные группы можно использовать в зависимости от применения. Можно синтезировать соответствующие дезоксинуклеозид-3-фосфорамидиты с последующим их связыванием с твердофазным носителем по методике Caruthers и др., 1987, см. выше. Первый дезоксинуклеозид может быть иммобилизован, например, в виде дезоксиаденозина. После снятия тритильной группы и промывания вначале дихлорметаном, затем ацетонитрилом, полученный продукт-носитель разделяют на четыре равных аликвоты и переносят в четыре отдельных реактора. Затем в эти четыре реактора вводят по отдельности четыре дезоксинуклеозид-3-фосфорамида. После завершения реакции связывания указанные продукты-носители, извлеченные из четырех реакторов, смешивают вместе, тщательно промывают, а затем подвергают окислению при использовании смеси, состоящей из йода, воды, лутидина и тетрагидрофурана. После окисления полученный продукт-носитель тщательно промывают ацетонитрилом и вышеуказанный цикл повторяют. После завершения синтеза неупорядоченной полидезоксинуклеотидной цепи (например, после 11 циклов связывания) остатки сложного метилового эфира расщепляют тиофенолом, а диметокситритил - трихлоруксусной кислотой. Разблокированные полинуклеотидные цепи могут оставаться ковалентно прикрепленными к твердофазному носителю (при подборе соответствующих линкеров), готовому для использования в приводимой ниже стратегии избирательного скрининга.
Настоящее изобретение предусматривает возможность использования олигонуклеотидов с другими, чем фосфодиэфирное связями. Так, например, олигонуклеотид может включать фосфортионатную связь. Известны модифицированные фосфодиэфирные связи или их аналоги. Такие модифицированные связи, как известно, обладают устойчивостью к экзонуклеазной и эндонуклеазной активности.
Поскольку имеется только четыре дезоксирибо- или рибонуклеозидов на цикл связывания, то в библиотеке с 12 нуклеозидными основаниями будет содержаться 412 возможных полинуклеотидных последовательностей, что есть в сумме будет составлять 1,68 • 107 возможных последовательностей. Олигонуклеотид можно также синтезировать при использовании как дезоксирибо- и рибонуклеозидов. Любой специалист должно быть также знает, что, кроме, мажорных нуклеозидов, можно использовать необычные (минорные) и модифицированные нуклеозиды. В качестве минорных и модифицированных нуклеозидов можно использовать нуклеозиды, содержащие метилированные пурины, производные уридина и 2-метилрибоза, которые могут образовываться в комбинации с любым рибонуклеозидом.
5.4. Твердофазные носители и ликеры, предназначенные для использования в рандомизированной биоолигомерной библиотеке.
Твердофазный носитель, предназначенный для использования в настоящем изобретении, должен быть инертен к реакционным условиям биоолигомерного синтеза, например, пептидного или олигонуклеотидного синтеза, либо к каждому из них. Твердофазный носитель изобретения должен иметь реактивные группы для прикрепления мономерной субъединицы или прикрепления линкера или спейсера, которые могут служить в качестве начальной точки связывания мономерной субъединицы. В другом варианте предлагаемого изобретения твердофазный носитель может быть пригоден для in vivo применения, то есть его можно использовать в качестве носителя для непосредственного его использования, либо в качестве подложки при применении непосредственно в биоолигомерной библиотеке (например, Tentagel, Rapp. Polymerg, Tubingen, Германия: см. ниже раздел 5.8). В конкретном варианте указанный твердофазный носитель может поступать через слизистую оболочку ротовой полости или его можно вводить перорально. В другом варианте осуществления настоящего изобретения твердофазный носитель предлагаемого изобретения можно использовать в качестве хроматографической подложки.
Твердофазный носитель, как он используется в изобретении, не ограничивает объем какого-либо конкретного типа носителя. Имеется довольно большое количество различных носителей, и они известны любому специалисту в данной области. В качестве твердофазных носителей можно использовать силикагели, полимерные смолы, пленки, полученные на основе пластиков, стеклянные бусы, хлопок, полимерные микросферы, алюмогели. Подходящий твердофазный носитель можно подобрать с учетом целевого назначения и его пригодности к различным режимам синтеза. Так, например, в качестве твердофазного носителя для пептидного синтеза можно использовать смолы, такие как полистирол (например, полиакриламидная смола, поставляемая Bachem Inc., Peninsula Laboratories и т.д.), смола Polyhipe ® (поставляемая фирмой Aminotech, Канада), полиамидная смола (поставляемая фирмой Peninsula Laboratories), полиэтиленгликоль - привитая полистирольная смола (Tentagel ®, Rapp. Polymex, Tubingen, Германия) или полидиметилакриламидная смола (поставляемая фирмой Milligene/Biosecarch, Калифорния). В предпочтительном варианте для пептидного синтеза в качестве твердофазного носителя используют полидиметилакриламидную смолу.
Твердофазные носители предлагаемого изобретения могут также содержать линкер. Как используется в данном описании термин линкер означает любую молекулу, обеспечивающую пространственный промежуток между носителем и синтезируемым пептидом. Линкеры могут быть ковалентно прикреплены к твердофазному носителю перед связыванием с ним Nα-Boc- или Nα-Fmoc-аминокислот, либо других надлежаще защищенных аминокислот. Различные линкеры можно использовать для иммобилизации на нем полученного олигомера. К примерам линкеров предлагаемого изобретения относятся аминокапроновая кислота, аминомасляная кислота, 7-аминогептановая кислота и 8-аминокаприловая кислота. Fmos - аминокапроновая кислота поставляется на рынок фирмой Bachem Biochem и наиболее предпочтительна для использования. В другом варианте осуществления изобретения линкеры могут дополнительно включать один или несколько бета-аланинов в качестве спейсеров. Более того, указанный твердофазный носитель можно модифицировать для удовлетворения особых требований, предъявляемых при проведении биологического анализа или распознавания конкретного объекта. Модификацию твердофазного носителя можно произвести при введении специфического линкера. Так, например, модифицированный твердофазный носитель может быть выполнен в виде носителя, чувствительного к кислотам, основаниям, нуклеофилам, электрофилам, свету, реакции окисления или восстановления.
Кроме вышеуказанных линкеров можно использовать селективно расщепляемые линкеры. Использование УФ-чувствительного линкера, а именно infra, приведено ниже в разделе 12 настоящего описания (см. Barany и Albenicia, 1985, J.Am. Chem. Soc., вып. 107, стр. 4936-4942). Для других расщепляемых линкеров необходимо проведение гидрогенолиза и фотолиза. Примеры светочувствительных (расщепляемых фотолизом) линкеров можно найти у Wang (1976, J.Org. Chem., вып. 41, стр. 32-58), Hammer и др. (1990, Int.J.Pept. Protein, вып. 36, стр. 31-45) и Kreib Cordonier и др. (1990, Peptides-Chemistry, Structure and biology, River and Marshall, второе издание, стр. 895-897). (1977, Meth. Enzym., вып. 47, стр. 145-149) использовал водный раствор муравьиной кислоты для расщепления Asp-Pro-линкерной группы: указанный метод использован для характеристики T-клеточных эпитонов в комбинации с синтезом на шпильках по методике Van der Zee и др. (1989, Eur. J.Jmmunol, вып. 191, стр. 43-47). К другим потенциальным линкерным группам, расщепляемым при использовании оснований, относятся линкерные группы на основе пара-(гидроксиметил)бензойной кислоты (Athirton и др., 1981, J.Chem. Soc. Perkin, часть 1, стр. 538-546) и оксиуксусной кислоты (Baleaux и др., 1986, Int. J.Pept. Protein Res, вып. 28, стр. 22-28). Geysen и др. (1990, J.Immunol. Methods, вып. 134, стр. 23-33) сообщают о пептидном расщеплении по дикетопиперазиновому механизму. Любой фермент может специфично расщеплять любой линкер, содержащий последовательность, которая чувствительна к или является субстратом для ферментативного гидролиза, например, протеазное расщепление пептида; эндонуклеазное расщепление олигонуклеотида. В некоторых случаях можно получать полимерную смесь, 10-50% в которой составляет расщепляемый линкер, а остальные 50-90% - нерасщепляемый линкер, что обеспечивает достаточное количество пептида, остающегося для секвенирования после расщепления линкера. Можно также использовать смеси из расщепляемых линкеров, что обеспечивает последовательное отщепление от только одной микросферы.
Твердофазный носитель, предназначенный для использования в настоящем изобретении может дополнительно содержать анализируемый биоолигомер, в который может быть введена рандомизированная последовательность субъединиц. Преиммобилизованный биоолигомер можно выбрать по описанным в данном описании методикам, либо он может содержать известную последовательность с требуемыми свойствами.
В синтезе олигонуклеотидов предпочтительно использование твердофазного носителя на двуокиси кремния. Как уже указывалось в разделе 5.3, такие твердофазные носители можно получить на коммерческой основе (например, у фирмы Peninsula, Laboratories Inc. и Applied Biosystems Inc.).
5.5. Методы детектирования и идентификации исследуемых биоолигомеров
Кроме создания достоверно рандомизированных библиотек биоолигомеров и методов их синтеза, настоящее изобретение также предлагает методы скрининга любой биоолигомерной библиотеки с идентифицированием биоолигомеров внутри указанной библиотеки, которые проявляют исследуемую биологическую активность, например, связывающую, ингибирующую, стимулирующую, цитотоксическую и т. п. Другие биоолигомерные библиотеки можно скринировать по методикам, описанным ниже, на ферментативную активность, фермент-ингибирующую активность и физико-химические свойства, которые представляют интерес.
Для исследуемых биоолигомеров, установленных в первом скрининге, не требуется их представление в форме лигандов на заключительном этапе. Фактически, предпочтительно синтезировать вторую библиотеку на основе обычных последовательностей лигандов, выбранных в первом скрининге. Следуя этой методике, можно идентифицировать лиганды даже с более высокой активностью при условии, что второй скрининг проводят при более жестких условиях.
5.5.1. Методы определения связывающей способности
Настоящее изобретение позволяет осуществлять идентификацию биоолигомерных лигандов, связывающих акцепторные молекулы. Термин "акцепторная молекула", как используется в данном описании, означает любое вещество, которое связывается с биоолигомерным лигандом. В качестве акцепторных молекул можно использовать биомакромолекулу, например, но нисколько не ограничивая объема, антитела, рецептуры или вирусы. Кроме того, в качестве акцепторных молекул можно использовать любое химическое соединение, например, но не ограничивая объема, белки, углеводы, нуклеиновые кислоты, липиды, лекарственные вещества, металлы или небольшие молекулы
Предлагаемая биоолигомерная библиотека может эффективно взаимодействовать с множеством различных акцепторных молекул. В результате идентифицирования определенного биоолигомерного лиганда, с которым связывается специфичная акцепторная молекула, становится возможным физически выделить исследуемый биоолегомерный лиганд.
Поскольку удается извлечь только небольшое количество микросфер в течение каждого цикла скрининг-детектирование-выделение, то большая часть микросфер остается в указанном пуле. Таким образом, указанную рандомизированную биоолегомерную библиотеку можно скринировать многократно. В случае использования индивидуальных схем окрашивания или идентификации для разных акцепторных молекул (например, при использовании флуорогенов, например, как флуоресцин (зеленый), красный Техаса (красный) и DAPI (синий), нанесенных на указанные акцепторы) и при наличии соответствующих фильтров возбуждения в флуоресцентном микроскопе или флуоресцентном детекторе в любую пептидную библиотеку можно вводить различные акцепторы (рецепторы) и одновременно проводить оценку, способствуя тем самым быстрому скринингу специфических лигандов. Такая методология не только снижает стоимость, но также увеличивает количество акцепторных молекул, которые можно скринировать.
В предлагаемом методе анализируемую акцепторную молекулу вводят в биоолигомерную библиотеку, в которой она распознает и связывается с одним или несколькими биоолигомерными лигандами внутри указанной библиотеки. Каждый биоолигомерный лиганд, с которым связывается акцепторная молекула, распознается только на одном твердофазном носителе, в результате чего можно легко идентифицировать и выделить указанный носитель, а следовательно, и требуемый биоолигомер.
Указанный биоолигомер можно выделить любым общеизвестным специалисту методом, однако изобретение не ограничивает объема до метода выделения. Так, например, но ни в коей мере не ограничивая объем, можно физически выделить биоолигомер, иммобилизованный на твердофазном носителе, проявляющий самое сильное физико-химическое взаимодействие со специфичной акцепторной молекулой. В одном варианте осуществления изобретения, основанной на физико-химическом взаимодействии, раствор, содержащий специфический акцептор, прибавляют к неупорядоченной пептидной библиотеке, которая эквивалентна примерно 105 - 107 твердофазных носителей. Акцепторную молекулу затем инкубируют с указанной смолой в течение промежутка времени, достаточного для связывания пептида с антителом, например, в течение одного часа при температуре 22oC. После этого акцепторную молекулу, покрытую твердофазным носителем, с биоолигомером выделяют. Более конкретные варианты осуществления настоящего изобретения раскрыты в нижеследующих методах, где описано использование моноклонального антитела в виде растворимой акцепторной молекулы. Специалисту станет очевидным, что эти методы можно легко приспособить для определения связывающей способности любой акцепторной молекулы. Более того, хотя нижеследующее касается библиотеки пептидов, следует понимать, что можно также проводить анализ олигонулеотидных библиотек и пептид-олигонуклеотидных химерных комплексов.
(i) Моноклональное тело вначале метят флуорогеном или вводят флуоресциновую метку по методикам, которые составляют рутинную работу специалиста в данной области. Антитело при концентрации 1 мкг/мл вводят в библиотеку пептидов, и после осторожного перемешивания при температуре 22oC в течение одного часа твердофазные носители промывают. Полученные комплексы пептидов с флуоресцирующими антителами, иммобилизованные на твердофазных носителях, идентифицируют и отделяют с помощью клеточного сортера с флуоресцентным возбуждением. В альтернативном варианте полученные комплексы пептидов с флуоресцирующими антителами, иммобилизованные на твердофазных носителях, идентифицируют и физически отделяют под препаративным микроскопом с флуоресцентной приставкой с помощью микроманипулятора. Относительная интенсивность флуоресценции обычно пропорциональна аффинности пептид-лиганда к анализируемому антителу.
(ii) Моноклональное антитело вначале конъюгируют на микрогранулах на основе ферромагнитного материала по рутинным методикам. Полученный антительный конъюгат при концентрации 1 мкг/мл затем инкубируют вместе с пептидной библиотекой в течение одного часа при температуре 22oC. Магнитные микросферы образуют розетку вокруг анализируемого пептида на твердофазном носителе, которую затем можно выделить сильным магнитом.
(iii) Моноклональное антитело вначале конъюгируют с ферментом, например, щелочной фосфатазой по рутинным методикам. Полученный антитело-ферментный конъюгат затем инкубируют вместе с рандомизированной пептидной библиотекой в течение промежутка времени от 30 минут до 1 часа при температуре 22oC. После промывания все содержимое библиотеки выливают в чашку Петри, содержащую субстрат на щелочную фосфатазу, например, 5-атом-4-хлор-3-индоил-фосфат (BCIP) и нитросиний тетразолий (NBT). После инкубирования в течение нескольких минут антитело-нагруженный пептидный комплекс на твердофазном носителе меняет свою окраску (становится синим) из-за осаждения трансформированного субстрата на твердофазном носителе, что позволяет легко идентифицировать его и физически выделить под препаративным микроскопом при использовании микроманипулятора. Относительная интенсивность цветной реакции обычно пропорциональна аффинности пептида на анализируемое антитело.
(iv) Моноклональное антитело вначале конъюгируют с ферментом, например, как пероксидаза хрена по рутинным методикам. Полученный антитело-ферментный конъюгат затем инкубируют в рандомизированной пептидной библиотекой в течение промежутка времени от 30 минут до 1 часа при температуре 22oC. После промывания все содержимое библиотеки выливают в чашку Петри, содержащую субстрат на пероксилазу, например, 3,3', 4,4'-диаминобензидин (DAB); 3,3', 5,5-тетраметилбензидин (TMB) или 4-хлор-1-нафтол (4ON). После инкубации в течение нескольких минут полученный антитело-пептидный комплекс на твердофазном носителе меняет свою окраску и его можно идентифицировать и изолировать физически под препаративным микроскопом с помощью микроманипулятора. Относительная интенсивность цветной реакции обычно пропорциональна аффинности пептида на анализируемое моноклональное антитело.
(v) Моноклональное антитело вначале метят биотином, то есть вводят биотиновую метку по рутинным методикам. После этого биотинилированное антитело инкубируют вместе с рандомизированной пептидной библиотекой в течение промежутка времени от 30 минут до 1 часа при температуре 22oC. После промывания прибавляют стрептазидин-щелочной фосфатазный комплекс или комплекс стрептазидина с пероксидазой хрена и инкубируют в течение 30 минут. Полученную подложку затем промывают, и цветную реакцию создают, как описано в вышеуказанном пункте (iii), в комбинации с иммунноферментным методом. Анализируемый носитель с иммобилизованным пептидом выделяют по методике, описанной ранее.
Кроме использования растворимых акцепторных молекул, в другом варианте изобретения можно детектировать биоолигомеры, которые связывают с рецепторами клеточной поверхности с использованием интактных клеток. Интактные клетки предпочтительно использовать с мультивалентными или подвижными рецепторами, либо с рецепторами, для которых необходимо наличие функционального липидного домена клеточной мембраны. Используемые в данной методике клетки могут быть как жизнеспособными, так и иммобилизованными. Обычно указанные клетки инкубируют вместе с пептидной библиотекой, которые, связываясь с некоторыми пептидами, образуют "розетку" между клетками-мишенями и релевантным пептидом, иммобилизированным на твердом носителе. Розетку можно затем выделить методом дифференциального центрифугирования, либо извлечь физически при использовании препаративного микроскопа.
В альтернативном варианте можно провести скрининг библиотеки при использовании пэннингового метода с клеточными линиями, например, как (i) родительская клеточная линия, где отсутствует на своей клеточной поверхности анализируемый рецептор и (ii) рецептор-позитивная клеточная линия, например, клеточная линия, образованная трансфекцией родительской линии геном, кодирующим исследуемый рецептор. После этого можно скринировать библиотеку, используя нижеследующую стратегию:
(i) вначале удаляют из библиотеки неспецифические микросферы, связывающиеся с клетками, не имеющими указанного рецептора посредством введения монослоя родительской клеточной линии по стандартному методу пэннинга, оставляя рецептор-специфические не связанные микросферы, или нерелевантные несвязанные микросферы;
(ii) удаляют несвязанные микросферы, к которым относятся как рецептор-специфические, так и нерелевантные микросферы и наносят их на монослой рецептор-позитивной клеточной линии, где указанная рецептор-специфическая микросфера связывается с рецептор-позитивной клеточной линией;
(iii) удаляют остальные нерелевантные несвязанные микросферы путем осторожного промывания и декантирования и
(iv) удаляют рецептор-специфическую микросферу (микросферы) при использовании микроманипулятора. В качестве альтернативного варианта интактную клетку анализируют на наличие иммобилизованных на мембране рецепторов и рецепторов, требующих функционального липидного домена клеточной мембраны, причем рецепторные молекулы можно заключить в липосомы, к которым могут быть прикреплены сигнальная группа или фермент. Хотя вышеуказанные примеры относятся к пептидным лигандам, любой из биоолигомеров, описанных ранее в разделах 5.1, 5.2 и 5.3, можно использовать в практике настоящего изобретения. Таким образом, акцепторная молекула может связываться с неклассическими, циклическими, конформационно подвижными или структурно ограниченными пептидами, олигонуклеотипами или пептид-олигонуклеотидными химерными комплексами.
В еще одном варианте осуществления предлагаемого изобретения в акцепторную молекулу можно ввести метку. В еще одном варианте осуществления изобретения можно использовать меченный второй реагент для детектирования участка прикрепления акцепторной молекулы к твердофазному носителю, содержащему исследуемый биоолигомер. Сайт связывания можно определить по in situ образованию хромофора с помощью ферментной метки. К подходящим ферментам для использования, но не ограничивая их объем, относятся щелочная фосфатаза и пероксидаза хрена. Еще в одном варианте осуществления изобретения можно использовать метод двухцветной реакции с использованием двух хромогенных субстратов с двумя ферментными метками на различных анализируемых акцепторных молекулах. С помощью указанной двухцветной пробы можно идентифицировать перекрестно-реактивные и монореактивные лиганды.
В качестве других методов в предлагаемом изобретении можно использовать цветные латексные микросферы, магнитные микрогранулы, флуоресцентные метки (например, флуоресцирующий изоцианат (FITC), фикоэритрин (PE), красный Техаса (TR), родамин, свободные или хелатообразующие соли лантанидов, особенно Eus, некоторые флуорофоры), хемилюминесцентные молекулы, радиоизотопы и метки на основе магнитограмм. Двухцветные пробы можно проводить при использовании двух или нескольких окрашенных латексных микросфер или флуорофором, которые излучают на разных длинах волн. Меченные микросферы можно выделять вручную или механическими средствами. В качестве механических средств можно использовать сортирование с активацией флуоресценции, например, аналогично клеточным сортерам с активацией флуоресценции и микроманипуляторные средства для выделения биовеществ.
В конкретных примерах осуществления изобретения, приводимых далее в описании, используют фермент-хромогенные метки и метки на основе флуоресцирующего изотиоционата (FITC).
Реактивные микросферы можно выделять на основе интенсивности метки, например, яркости, интенсивности флуоресценции, напряженности магнитного поля или радиоактивности. Наиболее интенсивно меченные микросферы можно выбирать и секвенировать, или в противном случае можно проводить анализ для определения структуры, например, масс-спектральный анализ. В другом варианте осуществления изобретения, при случайном отборе микросфер с повышенной интенсивностью метки можно выбирать и секвенировать любой произвольный фрагмент. Можно эффективно использовать самые современные методы микроскопии с визуальной детекцией для количественного определения интенсивности цветной реакции, а следовательно, и более корректного определения относительной аффинности лиганда к акцепторной молекуле до секвенирования микросферы. Аналогичным образом, в количественном анализе можно использовать метод иммуннофлуоресцентной люминесценции, если акцепторная молекула снабжена флуоресцентной меткой. Еще в одном варианте настоящего изобретения микросферы, показывающие некоторую напряженность метки, выбирают из библиотеки для компонентного анализа, например, определения аминокислотного состава. Можно получить ревизированную библиотеку, содержащую ограниченный набор мономерных субъединиц, которые по данным анализа химического состава, идентифицированы как важные, а затем провести ее скрининг.
В другом варианте осуществления настоящего изобретения биоолигомер(ы) с самой высокой аффинностью связывания, то аффинный константной, можно идентифицировать путем серийного разведения раствора исследуемой акцепторной молекулы до точки детектирования ее связывания только с несколькими твердофазными носителями, в альтернативном варианте можно обеспечить более жесткие условия связывающего раствора, либо в случае нуклеиновых кислот гибридизации с целевой нуклеиновой кислотой, то есть акцепторной молекулой. Любой специалист обычно знает, что более жесткие условия связывания можно обеспечить путем (i) повышения ионной силы раствора; (ii) повышения концентрации денатурирующих соединений, например, мочевины; (iii) повышения или снижения pH до нейтрального состояния (pH 7,0); в случае нуклеиновых кислот доведения до температуры плавления (Tп). Известны другие средства для изменения титра раствора для ограничения связывания по механизму высокой аффинности. Высокий тетр раствора акцепторной молекулы и связывание ее в строгих условиях с биоолигомером, иммобилизованным на твердофазном носителе, можно использовать для детектирования анализируемого лиганда в рандомизированной библиотеке, содержащей все или почти все возможные комбинации мономерных субъединиц, или в ревизированной библиотеке с ограниченным набором мономерных субъединиц.
В другом варианте осуществления изобретения биоолигомеры, показывающие низкоаффиное связывание, могут также представлять интерес. Их можно выбрать из библиотеки, первоначально извлекая все биоолигомеры с высокой аффинностью к связыванию, с последующим проведением детектирования в условиях пониженной жесткости и более низкого титра раствора.
В предпочтительном варианте можно использовать пробу с двойной меткой. Первую метку можно использовать для детектирования неспецифического связывания исследуемой акцепторной молекулы с микросферами в присутствии растворимого лиганда. Меченные микросферы затем извлекают из библиотеки, а затем удаляют растворимый лиганд. После этого детектируют акцепторную молекулу, показывающую специфическое связывание с остальными микросферами библиотеки. Биоолигомеры, иммобилизованные на таких микросферах, как можно предположить, связывают акцепторную молекулу в том же самом сайте связывания, что и исследуемый лиганд и, следовательно, имитируют этот лиганд. Анализ с использованием двойной метки имеет преимущество в том, что отпадает необходимость очистки исследуемой акцепторной молекулы, так как первая стадия анализа позволяет выделять неспецифические позитивно-отвечающие микросферы.
5.5.2. Методы определения биологической активности
Настоящее изобретение предлагает также методы для определения биологической активности любого биоолигомера из библиотеки путем ее обработки, в результате которой удаляются любые остающиеся от синтеза токсичные молекулы, например, методом нейтрализации с тщательным промыванием растворителем, стерильной водой и культуральной средой. Тесты для определения биологической активности можно проводить на токсичность, киллерную активность, стимулирующую и рост-стимулирующую активность и эффект, вызывающий физиологическую перестройку организма.
В предпочтительном варианте осуществления настоящего изобретения биоолигомеры предлагаемой библиотеки способны к избирательному высвобождению из твердофазного носителя, называемого также в данном описании как "микросфера". В одном варианте осуществления изобретения микросферы получают в такой форме, что только одна фракция биоолигомеров может избирательно высвобождаться. Селективно высвобождаемые биоолигомеры, линкеры и микробусы рассмотрены выше в разделе 5.4. Библиотеку биоолигомеров обрабатывают расщепляющими средствами, в результате чего происходит высвобождение одной фракции биоолигомеров. В качестве расщепляющих средств можно, но не ограничивая их объема, использовать, например, УФ-излучение, кислоту, основание, фермент или катализатор. В одном варианте осуществления изобретения библиотеку обрабатывают таким образом, что высвобождается от 10 до 90% биоолигомеров. В более предпочтительном варианте выделяется от 25 до 50% биоолигомеров. В случае, когда все биоолигомеры способны к высвобождению, можно провести неселективное расщепление путем ограничения действия расщепляющего агента. В одном варианте осуществления изобретения можно ограничить время экспозиции и интенсивность УФ-излучения. В другом варианте ограничивают концентрацию расщепляющего агента. После обработки с обеспечением расщепления библиотеку можно обработать повторно, например, реакцией нейтрализации для обеспечения биологической совместимости с требуемой пробкой. При практической реализации любой специалист может легко определить надлежащие условия гидролиза для частичного расщепления, если все биоолигомеры библиотеки иммобилизованы на твердофазном носителе посредством расщепляемых линкеров или мостиковых групп. Любой специалист также должен знать, что на относительную концентрацию высвобождаемого биоолигомера оказывает влияние изменение условий гидролиза.
В результате иммобилизации микросфер на твердой фазе создается градиент концентрации конкретного биоолигомера. Высокие концентрации биоолигомера отмечают, как правило, вблизи расположения микросферы, из которой он высвобождается. Таким образом, установление биологической активности вблизи расположения анализируемой микросферы позволяет осуществлять идентифицирование и выделение указанной микросферы и, следовательно, секвенирование и определение других характеристик биоолигомера. Идентифицирование биоолигомера возможно вследствие того, что достаточное его количество остается на микросфере после частичного гидролиза для его секвенирования и определения других характеристик. В другом варианте осуществления изобретения микросферы можно распределить по пунктам микротитровального планшета (например, 10 микросфер на лунку), при этом часть биоолигомера выделяют и анализируют на биологическую активность, тем самым устраняя возможные трудности, связанные с диффузией. Как будет описано ниже, различные фракции биоолигомера могут быть иммобилизованы на твердофазном носителе, либо микросфере с помощью различных способных к расщеплению линкеров для проведения секвенирования. В объеме указанных примеров под термином "микросфера" подразумевается твердофазный носитель. Следующие примеры предназначены для иллюстрации методов определения биологической активности, но не ограничивая объема.
(i) Популяцию клеток в моноклональной суспензионной культуре наносят послойно на жидкую среду или полутвердую матрицу, содержащую рандомизированную биоолигомерную библиотеку. В одном варианте осуществления настоящего изобретения эту процедуру выполняют в 96-луночном микрокультуральных планшетах с одной или несколькими микросферами на лунку вместе с клеточной суспензией. В другом варианте осуществления настоящего изобретения барьерной матрикс или "cookie-cutter" наносят на суспензию клеток и микросферы библиотеки с созданием отдельных камер. Аликвотное количество пептида на каждой микросфере ковалентно связывают с водорасщепляемым (например, как дикетопиперазин) или светорасщепляемым линкером. В результате можно выделить фракцию пептида, достаточную для оказания биологического эффекта, в то время как достаточное количество пептида остается иммобилизованным на микросфере для проведения секвенирования. Суспензия клеток может быть получена в виде раствора, либо полутвердой матрицы. После соответствующего инкубационного периода клеточную популяцию анализируют на ростовую и пролиферативную способность, например, путем идентификации колоний. В другом варианте осуществления настоящего изобретения можно прибавить к клеточной культуре тетразолиевую соль MMT (3-(4,5-диметилтриазол-2-ил)-2,5-дифенилтетразолийбромид) (Mossor, 1983, J. Jmmunol. Methods, 65: 55-63; Niks и Otto, 1990, J. Jmmunol Methods, 130: 140-151). Сукцинат-дегидрогеназа, обнаруженная в митохондрии живых клеток, превращает MMT в формазан голубой. Таким образом, насыщенная голубая окраска свидетельствует о наличии метаболически активных клеток. В другом варианте осуществления настоящего изобретения можно проводить количественный анализ с введением радиоактивной метки, например, тритий-меченного тимидина для определения пролиферирующей активности клеток. Аналогичным образом можно визуально представить пептидный синтез при введении 35S-метионина. Высвобождаемый из микросфер пептид, проявляющий как промотирующую, так и ингибирующую активность относительно роста клеток, выделяют и секвенируют, при этом идентифицированные пептиды последовательности повторно анализируют в растворе с тест-культурами против клеток-индикаторов.
(ii) В другом варианте осуществления изобретения, вытекающего из вышеуказанного пункта (i) микросферы библиотеки распределяют по пунктам микротитрационного планшета из расчета примерно 10 микросфер на лунку. Микросферы суспендируют в жидкой фазе. Из каждой микросферы высвобождается пептид в количестве, достаточном для проявления биологической активности, хотя достаточное количество пептида для секвенирования остается на микросфере. Супернатант, содержащий выделившийся пептид, переносят в микропланшет для репликации клеток или оставляют в лунках вместе с микросферами. Затем определяют биологическую активность, например, способность к росту или пролиферации клеточной линии. Микросферы из лунок, показывающие биологическую активность секвенируют с последующим приготовлением и анализом каждого фрагмента для выявления фрагментов, проявляющих исследуемую биологическую активность.
(iii) В еще одном варианте осуществления настоящего изобретения, вытекающего из вышеуказанного пункта (ii), биоолигомеры прикреплены к микросферам так, что примерно 1/3 биоолигомера может выделиться на первой стадии, примерно 1/3 его - на второй стадии и остальная 1/3 его удерживается на микросфере. Последовательное высвобождение биоолигомера обусловлено использованием двух разных расщепляемых линкеров, либо является результатом ограничения действия расщепляющего агента, что приводит к высвобождению только части биоолигомера на каждой стадии. Для ограничения такого действия предпочтительно использование регулируемого облучения светорасщепляемого линкера, хотя частичный гидролиз можно обеспечить при тщательно заданной длительности облучения в отношении расщепляющего агента химическими и ферментативными методами. Синтезируют библиотеку последовательно расщепляемых биоолигомеров и распределяют по лункам микротитрационных планшетов таким образом, что каждая лунка содержит более 50, предпочтительнее от примерно 50 до 250 микросфер/лунка. Затем микросферы обрабатывают с расщеплением примерно 1/3 указанных биоолигомеров. Полученный супернатант затем анализируют на биологическую активность путем репликации. Выделенные из лунок микросферы, показывающие биологическую активность, затем суспендируют и распределяют по лункам микротетрационного планшета из расчета на то, что каждая лунка содержит примерно от 1 до 10 микросфер. Полученные микросферы затем обрабатывают с высвобождением еще 1/3 части биоолигомера, и супернатант анализируют на биологическую активность. Микросферы из лунок, проявляющие биологическую активность выделяют и иммобилизованный биоолигомер секвенируют. В случае, где обнаружено более 1 микросферы, получают все идентифицированные последовательности, которые анализируют по отдельности на биологическую активность. Такая двухстадийная обработка биопробы приводит к созданию высокоэффективного метода скрининга очень крупных библиотек биоолигомеров со специфической биологической активностью.
(iv) Стимулирующую активность высвобождения цитокина можно определить путем прибавления суспензионной клеточной культуры, иммобилизованной на полутвердой матрице, например, агарозном геле. Если биоолигомер предлагаемого изобретения индуцирует высвобождение цитокина, например, лимфокина, фактора роста, гормона и т.д., то присутствие цитокина можно обнаружить по активности индикаторной клеточной линии. Специфические пробы с индикаторной клеточной линией можно провести по методике, раскрываемой выше в п. (i). В другом варианте осуществления настоящего изобретения цитокин, высвобождаемый при стимуляции клеток, может быть блоттирован на мембране, например, нитроцеллюлозной мембране, и наличие цитокина определяют методом иммунноанализа или рецептор-связывающего анализа.
(v) В другом варианте осуществления настоящего изобретения можно наблюдать цитотоксическую активность биоолигомера. Зоны или колонии, содержащие нерастущую, например, трансформированную или раковую клеточную линию, покрывающую биоолигомерную библиотеку, свидетельствуют о цитотоксичной активности. В конкретном варианте осуществления изобретения, в полутвердой матрице две популяции клеток могут покрывать одна другую. В этом случае можно идентифицировать цитотоксичный биоолигомер к клеткам-мишеням и не проявляющий цитотоксичности к любой клетке с антигенным фенотипом. Такой метод анализа позволяет быстро идентифицировать биоолигомеры, которые можно использовать в качестве химикотерапевтических средств. К цитотоксичным биоолигомерам относятся токсичные пептиды и анти-смысловые олигонуклеотиды.
(vi) Можно также детектировать физиологические изменения. В одном варианте осуществления настоящего изобретения суспензионную культуру миокардных клеток накладывают на биоолигомерную библиотеку. Можно наблюдать "биение" клеток, стимулированных биоолигомером. В другом варианте осуществления настоящего изобретения можно определить активацию конкретного фермента путем детектирования повышения активности специфичного фермента при наличии соответствующего субстрата, например, хромогена (например, ММТ, см. выше п. (i)), флуорофора или хемилюминесцентного агента. В альтернативном варианте активацию фермента можно определить методом иммунологического анализа. Еще в одном варианте осуществления настоящего изобретения гистологические исследования могут также указывать на физиологические и морфологические изменения, обусловленные биоолигомером библиотеки.
(vii) Настоящее изобретение предлагает метод определения активности любого биоолигомера в библиотеке на поляризованных клетках, например, клетках с базальной и инерстрициальной мембранной поверхностью. Полярные клеточные культуры можно приготовить на полупроницаемой мембране, соответствующей указанному интерстрицию. Библиотеку вводят в полутвердую матрицу на интерстициальную или базальную мембранную поверхность. Можно определить различные эффекты биоолигомера настоящего изобретения, например, как полярный транспорт, пролиферацию, межклеточное взаимодействие и т.д. В частности, при введении метки в биоолигомер, например, радиоактивной метки или флуорофора, можно идентифицировать транспортную способность биоолигомеров. Длительное время существует проблема создания специфически абсорбируемых молекул. В частности, такие молекулы можно было бы использовать для орального и назального введения лекарственных средств, в случае, где необходим транспорт молекул от интерстрициальной к базальной поверхности эпителия.
Можно также определить биологическую активность нерасщепляемых биоолигомеров. После чего можно провести скрининг биологической активности всех биоолигомер-покрытых микросфер. В одном варианте любую библиотеку предлагаемого изобретения можно выделить из специфической ткани. Можно выделить микросферы, которые специфически абсорбируются после орального, назального или подкожного введения. В предпочтительном варианте настоящего изобретения намагничены или имеют какую-либо другую идентифицирующую метку, и таким образом их легко выделить из ткани.
Любому специалисту должно быть ясно, что все из вышеуказанных методов определения биологической активности применимы к биоолигомерам, включая пептиды, олигонуклеотиды или пептид-нуклеотидные химеры. Известно использование пептидов или пептидных аналогов в качестве промоторов фактора роста, ингибиторов фактора роста и регуляторных молекул. Пептиды могут действовать в качестве генных регуляторов путем связывания их с регуляторным участком на любом гене, например, путем усиления или ингибирования эффектов промоторных, энхансерных и регуляторных белков. Аналогичным образом в качестве ингибиторов могут выступать нуклеиновые кислоты на индукторах генной экспрессии на уровне транскрипции (например, путем связывания или блокирования промоторов, энхансоров, сайтов терминации транскрипции и т.д.), процессинга и трансляции. Общеизвестно использование олигонуклеотида или олигонуклеотидного аналога для ингибирования трансляции специфичных нРНК. Любую и каждую из библиотек, описанных ранее в разделах 5.1 - 5.3, можно проанализировать на биологическую активность.
Специалист также должен понимать, что любую клетку, поддерживаемую в тканевой культуре, в течение как длительного, так короткого промежутка времени можно использовать в анализе на биологическую активность. Термин "клетка", как использован в данном описании, должен означать прокариотические (например, бактериальные) и эукариотические клетки, дрожжи, плесень или грибок. Можно использовать сохраняемые в культуре первичные клетки или клеточные линии. Более того, заявитель предусматривает, что биопробы на вирусы можно осуществлять путем инфицирования или трансформации клеток вирусом. Так, например, но нисколько не ограничивая объема, можно определить способность биоолигомера ингибировать лигогенную активность бактериофага лямбда путем идентифицирования инфицированных колоний E.coli, не образующих прозрачных бляшек при заражении.
Предлагаемые методы определения биологической активности любого биоолигомера библиотеки не ограничиваются вышеуказанными примерами; заявители полагают, что любую систему анализа можно модифицировать и включить в раскрываемое здесь изобретение и такие модификации должны находиться в пределах объема данного изобретения.
5.5.2.1. Биопроба на эритропоэтиновый агонист
В конкретном варианте осуществления настоящего изобретения предложен метод определения биоолигомерного агониста эритропойэтина. Следует иметь в виду, что раскрываемый в данном изобретении метод предлагает эффективную стратегию идентифицирования любого агониста, например, агониста ростовых факторов, гормонов, цитокинов, лимфокинов и других межклеточных переносчиков, которые описаны ранее в разделе 5.5.2.
В данном примере биоолигомерная библиотека может состоять из пентапептидов, полученных с 19 обычными аминокислотами (кроме цистеина). Расчетное количество четко выраженных пептидов составляет 2 476 099. Библиотеку можно синтезировать из 19 циклов, что облегчает проведение скрининга. Скрининг такой библиотеки проводят путем отбора только одной аминокислоты C-конца на каждой стадии, что приводит к упорядочению только положений других четырех аминокислот. Таким образом, последовательность каждой стадии соответствует XXXXY-линкер-смола, где Y выбирают для данной стадии, а X означает произвольное введение аминокислот. Указанный метод уменьшает количество возможных пептидов каждой стадии до 130 321. Если вносить в каждую лунку 96 луночного микротитрационного планшета по 10 микросфер (пептидов), то потребуется 136 микротитрационных планшет (одна дополнительная планшета необходима в качестве стандартов и контролей биопроб, так что общее их количество составляет 137). Указанный метод позволяет осуществлять распределение полного цикла создания библиотеки по указанному количеству микроитрационных планшет за один рабочий день.
Большую часть (50-80%) синтезируемых на микросферах пептидов можно расщепить для использования в анализе на биологическую активность, в результате чего, по крайней мере, 50 пмоль постоянно высвобождается из каждой такой микросферы. В качестве расщепляемого линкера предпочтительно использование дикетопиперазиновой
мостиковой группы, хотя светорасщепляемый линкер можно использовать также. Эта группа достаточно устойчива в мягких кислых условиях (например, 0,1 - 1,0 ммоль HCl), что позволяет распределять микросферы за 6-8 часов. Расщепление ускоряют (pH 8,5). Расщепление проходит в течение ночи (12-18 часов) при сохранении конечного объема лунки. Степень упаривания можно регулировать при выдерживании планшет в увлажненной камере.
Пептидный раствор, полученный в результате гидролиза микросфер библиотеки, должен отвечать требованиям асептики, если нет стерильности. Соблюдение асептических требований необходимо, так как раствор составляет 25% от объема конечного продукта, при этом время культивирования составляет, по крайней мере, 24 часа. Асептический режим можно обеспечить (1) гидратированием микросфер в стерильной воде после завершения синтеза, (2) разведением исходной суспензионной культуры микросфер в подкисленной стерильной воде и (3) при использовании стерильной техники для распределения указанных микросфер. В конечной суспензионной культуре микросфер присутствует не менее 20% ДМСО для облегчения солюбилизации гидрофобных пептидов. ДМСО (DMSO) следует вводить, если необходимо, в начале гидратации для облегчения солюбилизации пептидов. Конечная концентрация суспензионной культуры микросфер составляет 10 микросфер/50 мкл. Удержание микросфер в суспензии во время пипетирования можно обеспечить прибавление в нее примерно от 0,8 до 3,2% метилцеллюлозы. Использование метилцеллюлозы позволяет уменьшить действие ДМСО, который используют для ускорения солюбилизации. Конечная концентрация метилцеллюлозы в культурах составляет менее 0,8, чтобы не влиять на биоанализ.
Выделившиеся пептиды можно перенести в микробиологические планшеты в виде 50 мкл образцов, обеспечивая точное соответствие между чашками с образцами и культуральными планшетами. Во время указанного переноса необходимо обеспечит стерильные условия. Рекомбинантный EPO человека можно прибавлять к выбранным лункам планшетов для использования в качестве позитивного контроля (каждый планшет) и для построения калибровочных кривых (первый и последний планшет). Контрольные лунки могут содержать 0,1 М ед. EPO, при этом калибровочные кривые, полученные из шести наборов дубликатных лунок, содержат от 1,0 до 100 миллиединиц EPO (D' Andrea и др., 1991, Moll.Cell.Biol., 11: 1980-1087).
Биопробу можно получить с Ba/F3-T рекомбинантной клеточной линией, экспрессирующей эритропоэтиновый рецептор (EPO-рецептор). Указанные клетки зависят от присутствия либо интерлейкина-3 (IL-3), либо EPO. Культуры указанных клеток, содержащие IL-3 (поставляемые в виде 10% (об/об) WEHI - кордиционированной культуральной среды) предотвращает возможное влияние EPO в среде с биопробой. В качестве базальной культуральной среды используют среду RPMI 1640, содержащую 2,0 г/л NaHCO3, 10% фетальной бычьей сыворотки (об/об), 1 х пенициллин-стрептомицина, 5 мкл бета-меркаптоэтанола и/или 10 мМ HEPES-буфера (конечная концентрация 7,0). В указанную среду необходимо добавить 10% WEHI - кондиционированную среду (об/об), прибавленную к IL-3 WEHI - кондиционированную среду получают культивированием WEHI-клеток с их лизисом в той же самой среде. После этого кондиционированную среду центрифугируют для удаления клеток, пропускают через 22 мкм фильтр и хранят в замороженном состоянии. Ba/F3-T-клетки культивируют с выходом 1,31 и 107 клеток/лунка, которые распределяют по пунктам в количестве 1 • 103 клеток/лунка в объеме 150 мкм по методике, описанной в (Joshimura и др., 1990, Proc. Natl. Acad. Sei U.S.A., 87:4139-4143).
Ba/F3-T-клетки переносят из роллер-флаконов в большие (250 мл) стерильные роллерные колбы. Клетки собирают центрифугированием при скорости примерно 500 • об/5 мин. После этого клетки ресуспендируют в 200 мл свежей базальной среды без присутствия JL-3 для подсчета клеток. Конечный объем доводят до получения титра 6,67 •103 клеток/мл (1•103 клеток/150 мкл) при добавлении указанной среды. После этого необходимо разделить полученную суспензионную клеточную культуру на 4-8 аликвот для хранения в инкубаторе во время длительного процесса распределения клеток. Количество клеток и их жизнеспособность следует определять по образцам, взятым в начале и в конце процесса распределения, для обеспечения присутствия такого же самого количества жизнеспособных клеток в первом и последнем культуральном планшете. Полученные клетки распределяют по культуральным планшетам, содержащим супернатант из выделившихся пептидов, с последующим их инкубированием в течение 3 дней (Joshimura и др., см.выше).
Титр биопробы в конечной точке соответствует количеству живых клеток, присутствующих в каждой лунке с клеточной культурой. Его можно определить при использовании пробы Мосманна на МТТ (1983, J.Jmmunol. Methods 65:55-63), модифицированного Никсон и Отто (1990, J. Jmmunol. Methods, 130:140-151). Указанная модифицированная методика позволяет определять количество жизнеспособных клеток без их удаления из культуральной среды.
МТТ (3(4,5-диметилтриазол-2-ил)-2,5-дифенилтетразолийбромид получают в виде 5 мг/мл раствора в PBS (около 270 мкл). В каждую лунку планшета с биопробой пипетируют 20 мкл указанного раствора с последующим инкубированием в течение 4 часов при температуре 37oC. После этого в каждую лунку прибавляют 100 мкл экстракционного раствора и планшет помещают в ультразвуковой гомогенизатор на 120 минут. Раствор для экстрагирования содержит 50% (об/об) N, N-диметилформамида в 20% (мас./об) раствора, содержащего додецилсульфат натрия (SDS), доведенного до pH 4,7 при введении смеси уксусной и соляной кислоты по методике, описанной Mansen и др. (1989, J.Jmmunol. Methods, 119: 203). При такой обработке происходит солюбилизация формазанового продукта метаболизма МТТ для определения оптической плотности на длине волны 570 нм при использовании планшет-рипера.
Полученные данные оптической плотности биопробы усредняют по всем лункам и определяют 95% интервал достоверности усредненного значения. Значения оптической плотности каждой лунки, выходящие за пределы этого интервала используют определения степени достоверности по методу Стьюдента. Показатели достоверности сравнивают с калибровочной кривой EPO и получают расчетные данные в виде зависимости M. ед.-EPO. Калибровочную кривую определяют методом нелинейного регрессионного анализа с использованием логического уравнения. Данные и полученные из двух калибровочных кривых анализируют вместе с по отдельности, для определения, если имеется значительная разница, показатели реакции, снятой у обоих концов биопробы (тест для определения F-индекса). Сравнительный анализ контрольных значений, полученных для каждого планшета по каждому образцу, используют для определения, если имеется постоянное изменение в реакции биопробы.
Необходимо указать, что на клетках Ba/F3-T присутствуют рецепторы двух факторов роста (рецептор IL-3 и рецептор EPO), и что активация любого из них продуцирует позитивную реакцию биопробы. Таким образом, пробу на биологическую активность, описанную ранее, выбирают как для агониста рецептора JL-3, так и агониста EPO-рецептора. Имеются два пути решения указанной проблемы. Первый заключается в использовании штамма разных клеток или возможно клеток селезенки, взятых у мышек, которым вводили фенилгидразин (Krystal, 1983, Exp.Hematol., II, 649-660). Второе решение проблемы заключается в исследовании синтетических пептидов на биологическую активность относительно EPO и IL-3 путем проведения второго биоанализа, либо используя методы связывания с радиоактивномеченным лигандом.
5.5.3 Фермент-имитаторы и ферментные ингибиторы
Настоящее изобретение также включает в свой объем биоолигомеры, катализирующие реакции, в частности библиотеку ферментов, а также выступающие в качестве коферментов или ингибирующие ферментативные реакции. В соответствии с этим настоящее изобретение предлагает методы для определения ферментной и коферментной активности или ингибирования активности ферментов.
Активность ферментов можно наблюдать при образовании детектируемого реакционного продукта. В конкретном варианте осуществления настоящего изобретения биоолигомер библиотеки катализирует ферментативную реакцию субстрата на основе щелочной фосфатазы, например, 5-бром-4-хлор-3-индолил-фосфат (BCIP) и образует голубой несолюбилизированный продукт реакции на твердофазном носителе (см. ниже пример 13).
В другом варианте осуществления настоящего изобретения на полутвердой матрице можно получить зону видимого продукта, например, окрашенного или флуоресцирующего. Библиотеку получают послойно в полутвердой матрице, например, агарозном геле, а затем прибавляют хромофор или другой индикаторный субстрат. На участке, где иммобилизованный на твердофазной подложке биоолигомер проявляет анализируемую ферментную активность, образуется зона видимого продукта. Например, но ни в коей мере не ограничивая объема, биоолигомерный аналог пероксидазы хрена можно идентифицировать при добавлении раствора, содержащего аминоантипирен (0,25 мг/мл; Kodac), фенол (8 мг/мл) и воду (0,005%) в 0,1 М фосфатном буфере, pH 7,07. Микросферы с ферментативной активностью образуют пурпурную зону окраски. В другом варианте осуществления настоящего изобретения биоолигомеры/микросферы с протеазной активностью можно идентифицировать путем добавления хорошо известных специалистам колориметрических протеазных субстратов.
Коферментную активность можно наблюдать на пробе, анализуемого на коэнзим-опосредованную ферментную активность, где отсутствует природный или связанный кофермент.
Ферментингибирующую активность можно обнаружить в случае частично-выделившегося биоолигомера. Высвобождение биоолигомеров рассмотрено ранее в разделах 5.4 и 5.5.2. В одном варианте осуществления настоящего изобретения, но не ограничивая объема, библиотеку получают послойно в полутвердой матрице, которая содержит любой фермент. Библиотеку обрабатывают с частичным высвобождением биоолигомера. На участке, где биоолигомер ингибирует ферментную активность, можно идентифицировать зону невидимого продукта. В одном варианте осуществления настоящего изобретения, где в качестве ферментного субстрата используют хромогенный субстрат, образуется окрашенный продукт. Таким образом, наличие ферментного ингибитора приводит к образованию невидимой зоны. В другом варианте осуществления настоящего изобретения ингибирование протеолиза гемоглобина или фермента-индикатора, например, щелочной фосфатазы, можно установить по наличию непрозрачной зоны в полутвердой матрице. Иначе говоря, наличие протеазного ингибитора препятствует деградации гемоглобина и индикаторного фермента.
Любой специалист хорошо знает, что любой биоолигомер, проявляющий ферментную и коферментную активность или ингибирующий активность ферментов, может относится к пептиду, олигонуклеотиду, либо пептид-олигонуклеотидным химерным комплексам. Особенный интерес представляют ограниченные и циклические пептиды, которые продуцируют уникальный каталитический центр или поверхность связывания (см. выше раздел 5.2.1). Более того, пептид-олигонуклеотидные химерные комплексы, как предполагают, могут проявлять специфическую химическую способность, например, ферментную или коферментную активность, вследствие уникального расположения вблизи друг с другом соответствующих функциональных групп. Кроме того, твердофазный носитель с иммобилизованным на нем биоолигомером, как полагают, может проявлять активность фермента или кофермента, в то время как несвязанный биоолигомер может не иметь никакой активности. Такая высокая концентрация биоолигомера вблизи расположения соответствующих функциональных групп может передать уникальные химические свойства биоолигомерному продукту на твердофазном носителе. Предполагают также, что любой биоолигомер может проявлять ферментную или коферментную активность при высвобождении из микросферы. Общеизвестные коферменты (кофакторы), как полагают, можно включать химическим методом в конформационно ограниченный биоолигомер для стимуляции простого или сложного фермента, включая, например, электронтранспортную цепь.
5.6 Методы характеристики биоолигомеров
Настоящее изобретение предполагает средство для определения структуры и последовательности биоолигомера сразу же после выделения из библиотеки микросферы, содержащей исследуемый биоолигомер по любому из методов, описанных ранее в разделе 5.5.
В случае, когда биоолигомер относится к пептиду, в качестве предпочтительного метода для секвенирования используют способ деградации Эдмана. В особенности предпочтительном методе секвенирования используют белковый секвенатор модели 477 А Applied Biosystem. Аминокислотную последовательность пептидов можно также определить либо методом масс-спектроскопии с быстрой атомной бомбардировкой (PAB-MS), либо другими методами аналитического анализа.
Полученные пептиды можно секвенировать путем иммобилизации их на твердофазном носителе, либо при отщеплении их от твердофазного носителя. Для отщепления пептида выделенные микросферы с пептидом обрабатывают традиционными расщепляющими агентами, которые используют специалисты, для отделения полимера от твердофазных подложек. Подбор подходящих расщепляющих агентов зависит от используемого твердофазного носителя. Так, например, для снятия пептидов со смолы Wung предпочтительно использовать 50% трифторуксусную кислоту (TPA) в дихлорметане.
В альтернативном варианте осуществления настоящего изобретения можно выделить только один твердофазный носитель, например, микросферу, с прикрепленными к нему биоолигомерами с последующим переносом его в секвенатор без предварительного отщепления биоолигомеров от микросферы. Так, например, если исследуемые биоолигомеры относятся к пептиду, в отдельной микросфере на основе полимерной смолы с диаметром 100 мкм, что равноценно 0,5 мЭк/грамм смолы, содержится примерно 200 пмоль пептида. В одной 250 мкм-микросфере на основе полиакриламидной смолы из расчета 0,5 мЭк/грамм смолы содержится 3125 пмоль пептида. При использовании традиционного пептидного секвенатора необходимо только 5-10 пмолей пептида для корректного секвенирования. Таким образом, одна микросфера на основе ПАМ-смолы диаметром 100 мкм стандартного размера содержит пептид в количестве более, чем это необходимо для секвенирования.
В случае, где пептиды аминокислоты или пептид-инициаторы (псевдопептиды), которые невозможно анализировать методом Эдмановской деградации, можно получить такую микросферу при условии, что 10-50% ее пептидов и не должны содержать последовательность с несеквенируемым аминокислотным остатком. Можно определить структуру оставшейся аминокислотной последовательности, а затем на ее основе провести экстраполирование последовательности, содержащей несеквенируемый остаток.
Другой метод для определения структуры несеквенируемых остатков заключается во временном кэппинге части указанного пептида до введения несеквенируемого остатка во время синтеза библиотеки. В процессе последующей структурной идентификации можно использовать деградацию Эдмана вплоть до несеквенируемого остатка с последующим снятием временной "шапочки" и возобновлением секвенирования отдаленной области (например, С-концевой области) в направлении несеквенируемого остатка.
В случае олигонуклеотидов, секвенирование можно проводить на автоматическом олигонуклеотидном секвенаторе (например, фирмы Applied Вiosystems). В альтернативном варианте предпочтительно использование метода Максама и Гильберта (1977, Proc. Natl. Acad. Sci USA, 75:560-564). Можно также использовать другие известные в данной области методы секвенирования олигонуклеотидов.
Масс-спектроскопия с быстрой ионной бомбардировкой обеспечивает возможно наиболее эффективный структурный анализ. При детектировании фрагментов, а также самих биоолигомерных форм можно реконструировать любую последовательность. Можно также получить данные о структуре и последовательности олигонуклеотидов при использовании масс-спектрометрии высокого разрешения с электростатическим напылением (Finnigan, МАТ).
Сразу же после определения последовательности выбранного биоолигомера можно синтезировать химическим способом большое количество пептидов при использовании автоматического синтезатора пептидов или других средств, используемых в био- или химическом синтезе. Более того, сразу же как только установлена биоолигомерная последовательность, субъединичные аналоги можно заменить для усиления активности специфического биоолигомера.
5.7. Терапевтические диагностические средства на основе рандомизированных биоолигомерных библиотек
После того, как установлена исследуемая биоолигомерная последовательность, настоящее изобретение предлагает использование молекул, содержащих указанную биоолигомерную последовательность в терапевтических или диагностических целях. Указанная последовательность биоолигомера может сама дать диагностическое или терапевтическое средство, либо ее можно включать в более крупную молекулу. Любую молекулу, содержащую любую последовательность биоолигомера с биологической или связывающей активностью можно назвать "эффекторной молекулой. Изобретение также предлагает библиотеки, предназначенные для различных применений. Эффекторная функция такой эффекторной молекулы может быть любой из описываемых в данном описании или известных в данной области.
Раскрываемый в данном описании метод предлагает не только новый инструмент для поиска специфичных лигандов с потенциальным диагностическим и терапевтическим действием, но также представляет важную информацию о ряде лигандов, которые очень сильно отличаются по первичной структуре и химическому составу, но тем не менее могут взаимодействовать физически с одной и той же акцепторной молекулой. Интеграция такой информации с методами молекулярного моделирования и современной компьютерной техникой вероятно приведет к созданию новой фундаментальной концепции лиганд-рецепторных взаимодействий.
Лекарственные средства предлагаемого изобретения содержат эффекторные молекулы, связывающиеся с биологически активным центром цитокинов, факторов роста или гормонов, и тем самым усиливают или нейтрализуют их действие, а также ингибирующие или промотирующие транскрипцию и/или трансляцию.
К лекарственным средствам предлагаемого изобретения относятся, например, эффекторные молекулы, которые связываются с рецептором интересующего фармакологического действия, например, как рецепторы факторов роста, рецепторы нейромедиаторов или рецепторы гормонов. Указанные эффекторные молекулы можно использовать в виде либо агонистов или антагонистов лиганд-рецепторного взаимодействия.
Другое применение эффекторных молекул, которые связываются с рецепторами, заключается в использовании связывания для блокирования прикрепления вирусов или микробов, приобретающие доступ к клетке при прикреплении к нормальному клеточному рецептору и внедряются в него. В качестве примера такого феномена можно указать связывание ВИЧ с рецептором СД4, а также вируса герпеса вульгарного с рецептором фактора роста фибробласта. Эффекторные молекулы, внедряющиеся в рецептор, можно было бы использовать в качестве лекарственных веществ для ингибирования вирусной инфекции клето-мишеней. Паразитная инвазия клеток может быть аналогичным образом ингибирована после идентификации соответствующих эффекторных молекул согласно настоящему изобретению.
В другом варианте осуществления настоящего изобретения эффекторной молекулой, содержащей последовательность биоолигомеров, связывающуюся с любой интересующей акцепторной молекулы, можно использовать для целенаправленной доставки лекарственного средства или токсина. В предпочтительном варианте осуществления изобретения в качестве указанной акцепторной молекулы может быть рецептор или антиген, идентифицированный на поверхности опухолевой клетки, паразит животного или микроб, например, бактерии, вирус, моноклеточный паразит или патоген, грибок или плесень.
Возможно также, что несколько из миллионов биоолигомеров в пуле могут представлять последовательности, которые обладают биологической активностью. Например, можно выделить биоолигомеры, которые обладают противоопухолевой, антипаразитарной или антимикробной активностью, например, антифунгицидной, антибактериальной активностью против моноклеточного паразита или патогена, или антивирусной активностью. Более того, некоторые из указанных биоолигомеров могут действовать как агонисты или антагонисты факторов роста, например, эритропойэтина, эпидермального фактора роста, фибробластного фактора роста, опухолевых факторов роста, а также гормонов, нейромедиаторов или других регуляторных молекул. В одном варианте осуществления изобретения в качестве таких биоолигомеров используют пептиды.
Терапевтические средства настоящего изобретения включают также в свой объем эффекторные молекулы, содержащие биоолигомерную последовательность с высокой аффинностью к лекарственным препаратам, например, как дигоксин, героин, кокаин или теофилин. Такие пептиды можно использовать в качестве антидотов при сверхдозировках таких указанных лекарств. Аналогичным образом, к терапевтическим средствам настоящего изобретения относятся эффекторные молекулы, конъюгированные с небольшими молекулами или ионами металлов, в том числе ионами тяжелых металлов. Биоолигомерные последовательности с высокой аффинностью на билирубин можно использовать при лечении новорожденных с повышенным содержанием билирубина в крови.
Более того, настоящее изобретение предлагает методы для идентификации биоолигомерных последовательностей, используемых для терапии заболеваний или патологий, например, которые перечислены в Указателе категорий разрешенных биопродуктов настольного Справочника врача (PDR, 1991, 45 издание, Medical Economics Data: Oradell N 1, стр. 201 - 202). Так, например, можно идентифицировать эффекторную молекулу с активностью любого из противопаразитного, противокоагулянтного антагониста, противодиабетического, противосудорожного, антидепрессивного, противопоносного, антипотного, антигонадотропного, антигистаминного, противогипертонического, противовоспалительного, противозудного, противомигреневого, противопаркинсонного, антитромбоцитного, противорвотного, жаропонижающего, токсиннейтрализующего, бронхорасширяющего, сосудорасширяющего, хелатообразующего, противозачаточного, миорелаксантного, противоглаукомного или седативного средства.
Лекарственные средства предлагаемого изобретения могут также содержать соответствующие фармацевтически приемлемые носители, разбавители и адъюванты. В качестве таких фармацевтических носителей можно использовать стериальные жидкости, например, воду и масла, в том числе нефтяные, животные, растительные или синтетические масла, например, арахисовое, соевое, минеральное, кунжутное масло и тому подобное. В качестве предпочтительного носителя используют воду, если лекарственный препарат вводят внутривенно. В качестве жидких носителей можно также использовать физиологические растворы и водные растворы пекстрозы и глицерина, особенно растворы для вливаний. В качестве фармацевтически приемлемых наполнителей можно использовать крахмал, глюкозу, лактозу, сахарозу, желатин, мальтозу, рисовую муку, мел, силикагель, карбонат магния, стеарат магния, стеарат натрия, моностеарат глицерина, тальк, хлористый натрий, сухое обезжиренное молоко, глицерин, пропиленгликоль, воду, этанол и тому подобное. Указанные фармацевтические композиции можно готовить в виде растворов, суспензий, таблеток, пилюль, капсул, порошков, лекарственных форм с пролонгированным действием и тому подобное. Фармацевтически приемлемые носители описывает E.W. Martin в журнале Renington's Pharmaceutical Sciences. Такие композиции обычно содержат эффективное количество активного соединения в смеси с соответствующим количеством носителя в форме, пригодной для введения больному. Хотя внутривенное вливание является очень эффективной формой для введения лекарственного препарата, можно использовать другие формы, например, путем инъекции, или введением перорально, назально или парентерально.
Молекулу, содержащую биоолигомерную последовательность, идентифицированную согласно настоящему изобретению, можно также использовать для получения диагностических средств. Диагностический препарат может быть приготовлен на основе одной или нескольких последовательностей биоолигомеров предлагаемого изобретения, например, более одной пептидной или олигонуклеотидной последовательности. Кроме того, диагностическое средство может содержать любой из вышеуказанных фармацевтически приемлемых носителей.
Используемый в данном описании термин "диагностическое средство" означает любое вещество, которое можно использовать для детектирования заболеваний, например, но нисколько не ограничивая объем, рака, например, как T- или B-клеточно-опосредованная лимфома и инфекционных болезней, о которых говорилось выше. Термин "детектирование" используют в данном описании в самом широком смысле, охватывающем подтверждение наличия болезни, месторасположение пораженного органа или показатель тяжести болезни. Так, например, пептид-иммунопероксидазный комплекс или аналогичное иммуногистохимическое средство можно использовать для обнаружения и анализа специфических рецепторов или молекул антител в тканях, сыворотке или общей воды в организме. Диагностические средства можно использовать in vitro или in vivo. Более конкретно, настоящее изобретение предлагает эффективные диагностические реактивы для использования в иммунноанализах, гибридизации методом саузерн- или назерн-блоттинга и in situ анализах.
Более того, предлагаемое диагностическое средство может содержать один или несколько маркеров, например, как, но не ограничивая их объема, радиоизотопные, флуоресцентные метки, маркеров на основе парамагнетиков или других веществ, усиливающих изображение. Специалисты в данной области обычно знают множество маркеров и методы их введения для получения диагностических агентов.
Лекарственные и диагностические средства настоящего изобретения можно использовать для лечения и/или диагностики заболеваний у животных, более предпочтительно у млекопитающих, в том числе людей, а также экспериментальных млекопитающих, например, собак, кошек, лошадей, коров, свиней, морских свинок, мышей и крыс. Лекарственные и диагностические средства можно также использовать для лечения и диагностики болезней растений.
Заболевания и синдромы, которые можно лечить или диагносцировать с помощью биоолигомеров, найденных согласно настоящему изобретению, также же многочисленные и варьируют в широком диапазоне, как и вариации структур в рандомизированной биоолигомерной библиотеке. Проводимые ниже примеры предназначены только для иллюстрации, а не для ограничения объема изобретения.
5.7.1. Цитотоксичные композиции
Молекула, содержащая биоолигомерную последовательность, может проявлять специфическую цитотоксичную активность по отношению к себе самой. Более того, биоолигомер, связывающий исследуемый акцептор, можно модифицировать рутинными методами, применяемыми в данной области, например, путем конъюгирования с цитотоксичными веществами, например, лекарственными препаратами или радионуклидами с образованием цитотоксичной молекулы-носителя. Такой конъюгат цитотоксичного вещества с биоолигомером, например, пептидом, может обеспечивать целенаправленную поставку указанного вещества и специфически разрушать клетки, представляющие конкретную акцепторную молекулу. Так, например, такие цитотоксичные пептидные конъюгаты могут непосредственно разрушать нежелательные В-клеточные популяции, В-клеточные лимфомы, Т-клеточные популяции или Т-клеточные лимфомы у больного. В качестве возможного клинического применения таких конъюгатов можно указать их использование для лечения аутоиммунных заболеваний, лимфом и в иммуносупрессорной терапии при трансплантации органов. Можно также лечить по такой методике другие раковые формы, в которых раковые клетки проявляют рецептор-опосредованное связывание лиганда, например, рак молочной груди или матки, где рецепторы эпидермального ростового фактора (EGP), как полагают, играют важную ролью
Цитотоксичные препараты, специфичные к клетке-мишени, например, раковый или вирус-инфицированной клетке, но не приводящие к гибели иммунофенотипных клеток, тканей или органов, можно получать по предлагаемым в настоящем изобретении методам. Кроме целенаправленной поставки цитотоксичных препаратов в клетки-мишени, например, раковые, указанные специфические токсины можно использовать в качестве противомикробных средств. В частности, указанные лекарственные средства могут проявлять бактериостатическую, бактерицидную, противовирусную, противопаразитную или фунгицидную активность. Аналогичным образом можно идентифицировать токсины, проявляющие инсектицидную или гербицидную активность.
В одном варианте осуществления настоящего изобретения в качестве биоолигомера можно использовать пептид. Такой пептид может выступать в качестве носителя с иммобилизованным на нем любым токсином. Указанный пептид может сам поражать клетку и выступать в качестве токсина. В другом варианте осуществления настоящего изобретения в качестве такого биоолигомера можно использовать олигонуклеотид. Указанный олигонуклеотид может передавать свой токсичный эффект на уровне транскрипции и трансляции гена, ответственного за жизнеспособность клеток
5.7.2. Иммуномодуляторы
Настоящее изобретение предлагает молекулы и композиции для использования в качестве иммуномодуляторов. Термин "иммуномодулятор" молекулы или соединения, способные осуществлять изменения в иммунной системе. В частности, иммуномодуляторы могут стимулировать или ингибировать Т-клеточные иммунные реакции, гуморальный иммунный ответ, и неспецифические иммунные реакции, например, которые опосредованы действием макрофагов, нейтрофилов, полиморфно-ядерных фагоцитов, гранулоцитов и других клеток миелоидной линии дифференцировки. Эффективные молекулы можно использовать для лечения следующих заболеваний, связанных с нарушением иммунной системы: (1) различных аутоиммунных болезней, в том числе миастенин gravis, рассеянного склероза, болезни Грейвса, равматоидного артрита, системной красной волчанки (SLF), обыкновенной пузырчатки, аутоиммунной гемолитической анемии, иммунного тромбоцитоза, (2) болезни Ходжкина и различные другие раковые формы, (3) аллергии, (4) иммунных симптомокомплексов, (5) состояний, связанных с отторжением трансплантируемых органов, (6) инфекционных болезней и (7) сахарного диабета.
Иммуномодуляторы могут стимулировать иммунный эффект путем имитирования активности стимулирующего лимфокина, например, (IL)-1, (IL)-2, (IL)-3, (IL)-4, (IL)-6, гранулоцит-колониестимулирующего фактора (CSF), макрофаг-колониестимулирующего фактора и гранулоцит-макрофаг-колониестимулирующего фактора. Такое стимулирование может обеспечить пептид, связывающий лиганд с лимфокиновым рецептором, или олигонуклеотид, выступающий в качестве мениатора активации клеточной регуляции на уровне транскрепции. Иммуномодулятор может проявлять активность при связывании с лейкоцитом или лимфоцитом, например, как Гс-рецептор, LAP-1, LAP-2 и т.д., в том числе фагоцитирующую активность и активность высвобождения цитоксинов. Эффекторная молекула предлагаемого изобретения может действовать в качестве хемотаксина.
В предпочтительном варианте осуществления настоящего изобретения молекула, содержащая биоолигомер, может имитировать антиген. В такой форме указанный биоолигомер можно использовать в качестве вакцины для проявления Т-клеточной или В-клеточной реакции, специфичной к конкретному патогену.
Предлагаемые эффекторные молекулы, как полагают, имеют эффективное действие в высвобожденной форме. Однако отдельный биоолигомер может проявлять больший эффект, если остается иммобилизованным на твердофазном носителе. В частности, имитатор с высокой плотностью антигенных детерминант может более эффективно стимулировать гуморальный иммунный ответ при связывании и кэпировании мембранного иммуноглобулина или другой рецептор-опосредованной иммунной реакции.
Кроме того, ограниченную библиотеку предлагаемого изобретения, как полагают, можно использовать в качестве вакцины против патогена, представленного множеством эпитопов. Известно, например, что первичная структура (последовательность) VSG (гликопротеина с вариабельной мембраной) трипаносомы меняется непрерывно во время инфицирования. Пори изменении эпитопа VSG трипаносома теряет иммуннологическое распознавание. Аналогичным образом кровососущие паразиты, как установлено, выражают множественные антигенные эпитопы между видами на различных стадиях жизненного цикла, и внутри подвидов. Таким образом, пептидную библиотеку с ограниченной вариабельностью можно было бы использовать для иммунизации против кариабельного антигенного разнообразия, представленного трипаносомными или кровососущими паразитами. Ограниченная библиотека может найти применение в качестве вакцины в любом случае, где необходим иммунитет к ряду антигенов.
Эффекторные молекулы предлагаемого изобретения, как полагают, могут также ингибировать иммунный ответ путем (i) блокирования специфического иммуннологического распознавания на уровне антитела или рецептора Т-клеточного антигена; (ii) блокирования Fc-фрагмента или других иммунных рецепторов; или (iii) связывания с лимфокинами и цитокинами и ингибирования их активности; (iv) создания негативных обратных сигналов на иммунокомпетентные клетки. Эффекторные молекулы можно использовать для толеризации иммунной системы, в частности, для аутоантигенов. Так, например, иммунную толерантность в отношении ДНК можно обеспечить любым олигонуклеотидом или олигонуклеотидной библиотекой, и при лечении системной красной волчанки. Более того, ингибирование иммунной реакции можно обеспечить по механизмам, описанным выше в разделе 5.7.1.
Лекарственные средства предлагаемого изобретения могут также включать выбранные синтетические антигенные пептиды, с помощью которых можно манипулировать иммунной системой так, чтобы лишить иммуногенности этот антиген и, следовательно, подавить или излечить соответствующее аутоиммунное заболевание. Аналогичным образом при использовании специфических синтетических антигенных пептидов можно ингибировать образование мультимерных иммунных комплексов и, следовательно предупредить возникновение специфических иммунокомплексных заболеваний.
Пептиды, связывающиеся с опухолевоспецифичными моноклональными антителами, можно выделить, секвенировать и синтезировать для использования в качестве иммуногена, индуцирующего активную иммунную реакцию против этого новообразования.
Специфические пептиды, проявляющие высокую аффинность к рецепторам Fc-фрагмента, можно использовать в качестве лекарственных средств для ингибирования Fc-рецепторов ретикулоэндотелиальной системы, которые должны быть особенно эффективны для больных с аутоиммунными болезнями, например, как идиопатическая тромбоцитопения и аутоиммунная гемолитическая анемия.
Потенциал терапии при использовании таких пептидов может быть даже более значительным. Пептиды, которые схожи с эпитопами инвазивных организмов, можно использовать для блокирования инфекции с помощью такого организма. Так, например, последние исследования заболевания, связанных с приобретенным иммунодефицитом (СПИД), показали, что инфицирование вирусом СПИД начинается с узнавания и связывания между специфическим гликопротеином (др 120) на оболочке СПИД-вируса - СД4 - поверхностным рецептором на клетке. Введение пептида, имитирующего белок др 120, может ингибировать в значительной степени СД4-рецептор, в результате чего вирус СПИД теряет способность связывания с указанной клеткой и инфицировать ее. Аналогичным образом можно подавлять паразитную инвазию и инфекцию.
5.7.3. Нейрореактивные агонисты и антагонисты
Эффекторные молекулы предлагаемого изобретения, как полагают, могут действовать в качестве агонистов (имитаторов) или антагонистов (ингибиторов) эффектов гормонов, нейромедиаторов, анальгетиков, противопсихотических средств, антидепрессантов или наркотиков. Такие эффекторные молекулы следует использовать в поиске регуляторов аппетита, психиатрических средств, регуляторов внимания и усвоения и анамнестических средств. Настоящее изобретение предлагает также заменители вкусовых и ароматических добавок, например, заменители сладостей, соли и ароматических добавок.
5.8. Ограниченные библиотеки
Кроме того, ограниченную библиотеку предлагаемого изобретения можно использовать в качестве смешанной ароматической добавки, например, как пряности, при ее низкой стоимости и без возможных аллергических реакций от таких ароматиков. В этом случае можно получить недорогие заменители ароматических добавок, например, как шафран. В другом варианте осуществления настоящего изобретения можно получить новый класс ароматических веществ.
Еще в одном варианте осуществления настоящего изобретения ограниченную библиотеку можно использовать в качестве носителя для хроматографии. Библиотека биоолигомеров, например, пептидов, которые проявляют такие же химические свойства, но имеющие большое разнообразие последовательностей, должны, как полагают, быть более эффективной хроматографической подложкой по сравнению с имеющимися в настоящее время носителями. Такой хроматографический носитель обладает большей избирательностью по сравнению с ионообменным или обращеннофазовым носителем. Еще в одном аспекте настоящего изобретения очищаемую акцепторную молекулу можно без труда элюировать из указанного носителя в более мягких условиях, чем обычно могут использовать, например, высокоаффинного носителя, что снижает вероятность денатурации. В одном варианте осуществления настоящего изобретения указанный носитель получают на основе состава или структуры биоолигомеров, которые, как установлено, либо проявляют аффинность интермендиата, например, интенсивность мечения промежуточной формы, либо связываются только при высокой концентрации специфической акцепторной молекулы. Более того, высокоизбирательный носитель, используемый в условиях пониженной жесткости, можно быстро идентифицировать без его очистка (см. выше раздел 5.5.1).
В другом варианте осуществления настоящего изобретения можно подобрать низкоаффинные микросферы и на основе состава отобранных микросфер можно получить ограниченную библиотеку. В еще одном варианте настоящего изобретения традиционные низкоаффинный или высокоаффинный носитель, содержащий одну или несколько биоолигомерных последовательностей, идентифицированные из миллионов последовательностей, полученных согласно настоящему изобретению, можно использовать для хроматографии.
Приводимые далее примеры дополнительно поясняют данное изобретение и предназначены только для иллюстративных целей.
6. Пример: Синтез библиотеки тетрапептидов
Предлагаемый метод используют для синтеза семейства тетрапептидов формулы X-X-X-Trp, где X может означать либо валин, серин, либо аланин, при условии, что в качестве первой аминокислотой всегда используют триптофан. Триптофан введен в C-конец для облегчения спектрофотометрического анализа при ОД 280.
Na-Fmos-триптофан-алкоксиметил-полистирол, который описывает Wang (1973, J. Amer. Chem., 95:1328-1333), получают у фирмы Bachem. Inc. Torrence, Ca и помещают в обычный твердофазный пептидный синтезатор. В качестве вводимых аминокислот используют также Fmos-аминокислоты, поставляемые фирмой Bachem Inc. Другие используемые реагенты практически такие же, которые используют в рутинном методе твердофазного пептидного синтеза, описываемого Austem (1986, Peptide Synthesis: Methods in Moleculae Biology, стр. 311-331).
Биореактор с тефлоновым покрытием используют для реакции связывания пептидов; стандартный биореактор для твердофазного синтеза белков используют в качестве смесительной камеры, где после завершения связывания перемешивают аликвоты.
Примерно 0,5 г Fmos-Trp-алкоксиметил-полимера выдерживают для набухания в диметилхлорметане. Затем указанный биополимер промывают дважды дихлорметаном, один раз смесью дихлорметилформамид, 1:1 и трижды только диметилформамидом. После тщательного промывания биополимера с удаленными защитными группами диметилформамидом (3 раза), дихлорметаном (3 раза), смесью диметилхлорметан-диметилформамид, 1: 1, указанный биополимер делят на три отдельные аликвоты примерно по 2,5 мл каждая и помещают в три серии конъюгационных пробирок.
Количество вводимой защищенной аминокислоты рассчитывают исходя из количества молей триптофана, уже иммобилизованного на указанном биополимере. Для каждой вводимой аминокислоты пятикратный избыток указанной аминокислоты вводили в каждый биореактор, в которые уже ввели аликвоты промытого биополимера. В каждый биореактор таким образом введено пятикратный избыток конкретной аминокислоты. После этого каждый биореактор подвергают перемешиванию шейкерным методом в течение 2 минут и прибавляют диизопропилкарбодиимид в пятикратном избытке в 3 мл дихлорметана (ДХМ) с последующим перемешиванием в шейкере в течение 1 часа.
Для проверки степени завершенности связывания пробу из каждой пробирки подвергают реакции на нонгидрин, поставляемый фирмой Pcerce Chemical по методике, описанной Sarin и др. (1981, Anal. Biochem., 117:147-157), специально включенную в данное описание в качестве ссылки. Если реакция связывания не закончена полностью, как показывают данные указанной пробы, то реакцию доводят до полного завершения следующими методами, которые известны специалистам, в том числе (a) проведение второго связывания при использовании защищенной аминокислоты в 1-5-кратном избытке; (b) проведения дополнительного связывания при использовании разных или дополнительных растворителей (например, трифторэтанола) или (c) прибавлении хаотропных солей, например, NaClO4 или LiBr (Klis и Stewart, 1990, Peptides: Chemistry, Structure and biology, River and Marchal, 2-е издание, стр. 904-906).
После завершения связывания биополимеры из трех конъюгационных пробирок осторожно переносят и смешивают в гомосмесителе. Полученную смолу промывают дважды смесью ДХМ/ДМФ, 1:1, трижды ДХМ, трижды ДМФ и защитные группы снимают 20% раствором пиперидина в ДМФ, об/об. После тщательного промывания с использованием ДХМ и ДМФ по вышеуказанной методике полученную смесь делят на три аликвоты и помещают в три отдельные биореактора. Затем прибавляют вторую серию аминокислот. После полного связывания полученный биополимер сначала разблокировывают 20% раствором ДХМ и ДМФ по вышеуказанной методике. Аналогичным образом прибавляют третью серию аминокислот.
Для отщепления пептидов от твердофазных носителей к указанному биополимеру прибавляют 30 мл 50%, об/об трифторуксусной кислоты, 5% анизол и 0,9% (об/об) этандитиола в ДМХ. Полученную смесь перемешивают в шейкере в течение 4 часов, и пептидный супернатант собирают. После этого указанный супернатант пептидов упаривают в роторном испарителе, и пептиды осаждают в простом эфире. После тщательного промывания пептидный преципитат сушат и подготавливают для последующего анализа. Лиофилизированный пептид (в порошкообразной форме) хранят в замороженном виде.
7. Пример: Сравнительный анализ заявляемого метода и традиционного метода синтеза пептидов
Библиотеку рандомизированных пептидов получают по методике, описанной в примере 6 выше. Более того, библиотеку тетрапептидов получают при использовании стандартного твердофазного пептидного синтеза (упоминаемого далее как "SPPS"), описываемый Austen (см. выше). Na-Fmos-триптофан-алкоксиметил-биополимер, поставляемый фирмой Bachem, Inc., используют в виде комбинации твердофазный носитель - аминокислота. Эквимолярные количества пятикратного избытка Na-Fmos-валина, Na-Fmos-серина (O-Bu) и Na-Fmos-аланина прибавляют в биореактор во время каждой стадии связывания. После трех поочередных стадий связывания полученные тетрапептиды расщепляют в смеси, содержащей 50% (об/об) трифторуксусную кислоту, 5% (об/об) анизол и 0,9% (об/об) этандитиол в ДХМ, по вышеописанной методике Austen.
7.2 Результаты
Обе пептидные библиотеки анализируют методом C-18 обращеннофазной ВЭЖХ (Vydac) для определения количества пептидных видов (по количеству пиков), относительной концентрации пептидов (пиковой зоне и относительной гидрофильной природы указанных пептидов (по раннему или позднему элюированию из колонки). Полученные результаты представлены на фиг. 3). Хроматограмма в верхней панели показывает образец, полученный вместе с библиотекой пептидов, синтезированную по методу согласно настоящему изобретению, а хроматограмма в нижней панели представляет образец, полученный стандартным методом твердофазного пептидного синтеза.
Обе хроматограммы демонстрируют 21 отчетливых пиков, что свидетельствует о присутствии, по крайней мере, 21 разных пептидных вида в пределах каждой библиотеки. Картина, полученная SPPS-методом, однако, показывает значительно большие пики у # 1, 2, 3, 4, 5, 6 и 7, свидетельствуя о том, что библиотека, полученная SPPS-методом, содержит большую концентрацию пептидов 1-7 по сравнению с пептидами 8-21. Повышенное содержание пептидов 1-7 показывает, что указанные пептиды имеют предпочтительное преимущество синтезирования по сравнению с остальными 21 пептидами. Более того, указанные выступающие пики элюированы первыми, то есть указанные пептиды демонстрируют более короткое время удержания в колонке, что свидетельствует о более гидрофильной природе этих пептидов.
Такой результат нельзя ожидать при использовании SPPS-метода. Известно, что валин является гидрофобом и объемным веществом и имеет значительно медленную скорость связывания (как предполагают из-за стерической затрудненности), чем это установлено в случае как с валином, так и серином. Таким образом, в процессе традиционного синтеза неупорядоченных пептидов, проводимого в данном изобретении, при котором валин "конкурирует" с аланином и серином за сайты связывания, синтезируемые пептиды являются валин-обедненные, и получаемая пептидная библиотека не демонстрирует эквимолярного распределения неупорядоченных пептидов.
В противоположность этому хроматограмма библиотеки неупорядоченных пептидов, создаваемой согласно методу настоящего изобретения, подтверждает эквимолярное их распределение. Хотя пики 3, 6, 12, 13 и 18 примерно в два раза превышают зону других пиков, что свидетельствует о присутствии двух пептидов на том участке, при этом остальные 16 пиков имеют почти идентичную картину. Более того, все 21 пика лежат в диапазоне времени удержания, что также свидетельствует об эквимолярном распределении этих пептидов. Секвенирование образцов с заданными пиками также подтверждает этот результат. Образцы с небольшими пиками 8, 9 и 21 и образец 6 с большим пиком секвенировали на белковом секвенаторе 477A фирмы Applied Biosysmems, как следует ниже
# 8 = Val-Ala-Ser-Trp
# 9 = Val-Ser-Ala-Trp
# 21 = Val-Val-Val-Trp
# 6 = (Ser-Val)-(Ser-Ala)-(Ser-Ala)-Trp
Указанные валин-содержащие последовательности подтверждают возможность использования метода предлагаемого изобретения для рандомизированного синтеза пептидов даже в случае, когда используют известные медленно связывающиеся аминокислоты.
Последовательность самого большого образца 6 не была подтверждена, по-видимому, из-за присутствия нескольких пептидов у этого пика. Наиболее вероятно, что самые главные пики относятся к последовательности Ser-Ala-Trp и Val-Ser-Ser-Trp.
7.3. Выводы
Полученные результаты свидетельствуют о том, что предлагаемый метод синтеза неупорядоченных пептидов позволяет получить библиотеку рандомизированных пептидов в практически эквимолярных количествах в противоположность стандартному SPPS-методу, в которой приобладает набор пептидов, содержащих аминокислоты с более быстрой скоростью связывания.
8. Примеры: Выделение пептидного лиганда, связывающегося с рецепторной молекулой
С целью подтверждения использования метода настоящего изобретения для выделения конкретного пептида, синтезируют 12-аминокислотный пептид с заданной последовательностью на основе V-mos-генного продукта. V-mos-генный продукт относится к онкогену, выделенному из мышиной саркомы, и относится к вирусу саркомы Молони мышей. Известно, что V-mos-
генный продукт проявляет серин/треонин-киназную активность.
8.1.Материалы и методы
Последовательность Leu-Gly-Ser-Gly-Gly-Phe-Ser-Val-Tyr-Lys-Ala синтезируют на полиакриламидной макросфере (примерно 300 мкм-диаметра) с использованием Na-Fmos-химии и стандартных реактивов и методов для проведения твердофазного пептидного синтеза. Защитные группы боковой цепи снимают при использовании 50% раствора трифторуксусной кислоты, и оставшийся ковалентно прикрепленным к полиакриламидной смоле через линкер пептид на выходе имеет следующую структуру: Leu-Gly-Ser-Gly-Gly-Phe-Ser-Val-Tyr-Lys-Ala-аминокапроновая кислота - этилендиаминовый полимер (далее называемый как "длинная" V-mos-микросфера). Указанная пептидная последовательность соответствует аминокислотным остатком 100-111 V-mos-генного продукта.
Используя эту же методику, укороченный пептид, соответствующий аминокислотным остаткам 106-111 V-mos-генного продукта (Gly-Ser-Val-Tyr-Lys-Ala) на полиакриламидной микросфере с тем же самым линкером (далее называемый так "короткая" V-mos-микросфера). Этот пептид используют в качестве негативного контроля.
Гибридомная клеточная линия, продуцирующая мышиное моноклональное антитело против "длинного" V-mos-пептида, известное как анти-V-mos (Гибридома N-165-28E7, SCRF 354, Лот N 165-119) поступает в продажу из фирмы Biological Associates Inc. , штат Мериленд. В твердофазном иммуноферментном анализе (ELISA - указанное антитело распознает гомологичную последовательность из V-mos, MOS, Neu HER-1, HER-2-генных продуктов. Известно, что указанное антитело проявляет ничтожно низкую аффинность к короткому V-mos-пептиду. Вторичный козий-анти-мышиный JgG, меченный щелочной фосфатазой, получают у фирмы Sigma.
Используя традиционные методы продуцирования моноклонального антитела, как описано, например, в журнале Methods in Enzymology, вып. 121, 1986, моноклональные антитела получают в асцитной форме и затем очищают на G-белковой колонке, полученной у Pharmacia.
8.2. Результаты
Длинные V-mos-микросферы смешивают 1000-кратным избытком коротких -mos-микросфер 2 мл очищенного анти-V-mos моноклонального антитела (1 мкг/мл) в забуферном фосфатом физиологическом растворе с 0,1% Твин 20 прибавляют к полученной смеси микросфер и инкубируют при комнатной температуре в течение одного часа при осторожном перемешивании. После этого микросферы промывают на небольшой съемной полипропиленовой колонке (получают из фирмы Isolab), где они удерживаются фриттой. Затем микросферы перемешивают с 2 мл второго антитела при разведении 1:2000 в течение 1 часа. После промывания микросферы выливают в полистироловые чашки Петри и дают осесть. Полученный супернатант удаляют и прибавляют раствор, содержащий 5-бром-4-хлор-3-индолил фосфат и нитросиний тетразолий, в качестве субстрата.
После инкубации при комнатной температуре в течение 15 минут длинные V-mos-микросферы становятся пурпурными в противоположность коротким V-mos-микросферам, которые остаются неокрашенными. Это позволяет сразу же обнаружить только одну темную микросферу в газоне из тысяч неокрашенных микросфер. Отличия между микросферами показаны на фиг. 4-6, каждый из которых является фотографией микросфер в чашках Петри, при 40-кратном увеличении. На фиг. 4 показан газон длинных V-mos-микросфер, меченных моноклональным антителом; на фиг. 5 показана смесь из длинных и коротких V-mos-микросфер, меченных моноклональным антителом против V-mos, и на фиг. 6 показана единственная синяя микросфера после детектирования в газоне неокрашенных микросфер. Соответственно, микросферы. содержащие исследуемую пептидную, требуемую пептидную последовательность, легко распознаются и выделяются из других микросфер в указанной библиотеке.
После выделения для определения N-концевых аминокислотных последовательностей единственной микросферы с длинной V-mos-используют белковый секвенатор 477A фирмы Applied Biosystems.
8.3. Выводы
Указанный пример показывает мощный инструмент настоящего изобретения для отбора микросферы, содержащей биоолигомерный лиганд, в данном случае пептид, который выявляют из тысячекратного избытка несвязывающих, нерелевантных микросфер. Более того, в указанном примере продемонстрирована возможность выделения реактивной микросферы и определения последовательности пептида.
9. Пример: Выделение укороченного пептидного лиганда, связывающегося с рецепторной молекулой
Для дальнейшей иллюстрации использования предлагаемого метода для выделения конкретного пептида синтезирован гексапептид с заданной последовательностью Gly-Phe-Gly-Ser-Val-Tyr на стандартной 100 мкм полиакрилметакрилатной смоле с использованием Na-Fmos-химия и других реагентов из традиционного твердофазного пептидного синтеза.
9.1. Материалы и методы
Реакцию связывания проводят по методике, описанной выше в разделе 8.1. Альфа-аминозащитные группы удаляют 20% пиперидином, защитные группы боковой цепи снимают 50% раствором трифторуксусной кислоты, при этом оставшийся ковалентно прикрепленным к полиакриламидной смоле через линкер пептид на выходе имеет следующую структуру: Gly-Phe-Gly-Ser-Var-Tyr-аминокапроновая кислота-бета-Ala-олигомер. Указанная пептидная последовательность соответствует аминокислотным остаткам 104-109 V-mos-генного продукта, описанного выше в примере 8.
Аналогично примеру 8 антитела против V-mos собирают из гибридомной клеточной линии, продуцирующей мышиное моноклональное антитело, специфичное к V-mos-пептиду. Второе меченое щелочной фосфатазой козье антитело против мышиного IgG получают у фирмы Sigma.
9.2. Результаты
Примерно 0,1 мг ПАМ/V-mos-пептида, описанного ранее, перемешивают с 100-кратным избытком N-бета-F-mos-ПАМ-микросфер, полученных у Bachem Inc. Два мл очищенного моноклонального антитела (1 мкг/мл в забуференном фосфатом физиологическом растворе с 0,1% Твин 20 прибавляют к полученной смеси пептид/носитель и инкубируют при комнатной температуре в течение 45 минут при осторожном перемешивании.
После этого микросферы промывают на небольшой съемной полипропиленовой колонке (получают из фирмы Isolab), где они удерживаются на фритте (стеклянном фильтре). Затем микросферы перемешивают с 2 мл второго антитела, меченого щелочной фосфатазой при разведении 1:100, в течение 1 часа. После промывания микросферы раскладывают на кусочке фильтровальной бумаги и просушивают в 2,2'-азинобис(3-этилбензтиозолин-сульфокислотном (ABTS) субстрате с H2O2. После инкубации при комнатной температуре в течение 15 минут ПАМ/V-mos-пептидные микросферы становятся темно-зелеными. Небольшая светло-зеленая полоса образуется на стеклянном фильтре. Большая часть твердофазных носителей, где отсутствует V-mos-пептида, не взаимодействует с моноклональным антителом и, таким образом, не окрашивается. V-mos-микросферы обнаружить не составляет труда.
9.3. Выводы
В указанном примере показано, что анализируемая акцепторная молекула не специфична к твердофазному носителю, а скорее специфична к комбинации носитель/пептид. Аналогично вышеописанному примеру 8 позитивно-реагирующую микросферу можно выделить, а иммобилизованный на ней пептид секвенировать.
10. Пример: Определение лигандов для стрептавидина и МАВ против β-эндорфина
В настоящем примере иллюстрируется совершенного другой подход к идентификации пептидного лиганда настоящего изобретения. В описываемых методах вместо применения биологической системы (например, гибридного нитчатого фага) для генерирования рандомизированной библиотеки эффективно используется синтез огромных пептидных библиотек, где каждый отличающийся от других пептид находится на отдельной микросфере. Затем отдельные специфически связанные с пептидом микросферы физически выделяют по одной микросфере и определяют последовательность связанного с ней пептида.
Этот метод зависит от возможностей химического синтеза огромной рандомизированной пептидной библиотеки и ее связывания для соответствующего обнаружения, выделения и системы определения структуры.
Средство для решения этой проблемы, предоставляемое настоящим изобретением, заключается в том, что полимерные микросферы разделяют на серии отдельных равных аликвот в течение каждого цикла связывания, и каждую аликвоту полимера подвергают реакции вплоть до ее завершения с отдельной активированной аминокислотой. После завершения связывания различные аликвоты полимера тщательно смешивают, промывают, подвергают разблокированию, промывают и снова разделяют на аликвоты для нового цикла связывания. В соответствии с указанной методикой, ни одна полимерная микросфера не подвергается воздействию более, чем одной аминокислоты в любом одном цикле связывания, и при завершении нескольких таких стадий каждая микросфера будет содержать одну единственную пептидную последовательность. Пептидная библиотека, полученная таким образом, теоретически будет абсолютно рандомизированной. Кроме того, в результате такой процедуры будут получены эквимолярные отношения всех видом пептида. Полное число пермутаций, а следовательно, и число пептидов будет зависеть от числа аликвот и аминокислот, выбранных в каждой стадии связывания, и от полного числа стадий связывания при синтезе (длины пептида).
Новый метод одновременного синтеза широкого ряда пептидов не только позволяет получить абсолютно рандомизированную и эквимолярную библиотеку, но и, что более важно, дает возможность получить библиотеку твердофазных пептидных полимерных микросфер, где каждая микросфера содержит только одну уникальную пептидную последовательность. Этот последний из указанных факторов является совершенно бесспорным, поскольку в течение каждого цикла пептидного синтеза каждая микросфера находится в контакте только с одной отдельной аминокислотой в данный момент времени, и каждая реакция связывания протекает вплоть до ее завершения. В основе успеха описываемого метода лежит, главным образом, идея связывания одной микросферы с одним пептидом.
Кроме того, благодаря описываемому методу, может быть синтезирована практически любая пептидная библиотека с хорошо определенным составом. Например, в каждой стадии связывания может быть использовано до 20 натуральных L-аминокислот в отдельных аликвотах, или в некоторых стадиях связывания может быть использована одна или несколько аминокислот.
10.1. Материалы и методы
10.1.1. Синтез пептидной библиотеки
Синтезировали большую библиотеку, имеющую следующую структуру X-X-X-X-X- β-аминокапроновая кислота - этилендиаминполимер (X = 19 из 20 известных аминокислот, кроме цистеина, в каждой стадии связывания). В пептидном синтезе в качестве твердофазных полимерных микросфер использовали полидиметилакриламид (ПДА) (M:11: Gen.Inc., США).
Способ химического пептидного синтеза с использованием полимера осуществляли согласно Atherton (1988, "Твердофазный пептидный синтез. Практические методы", Solid Phase Peptide Synthesis, A.Practicоl Approach, IDL-Press).
Три грамма полимера (приблизительно 2 миллиона микросфер) слегка размешивали в течение ночи с этилендиамино. После тщательного промывания аминокапроновую кислоту, а затем β-аланин связывали с полимером с помощью Fmos-химии, но без использования отщепляемого линкера. Рандомизацию осуществляли с последовательных пяти стадиях связывания, и все 19 Fmos-аминокислота-OPFp, за исключением цистеина, использовали отдельно в течение каждой стадии связывания. После завершения указанных пяти стадий связывания Fmos-группу удаляли в 20% пиперидине (об/об) в ДМФ. Защитные группы боковой цепи удаляли смесью 90% TFA (об/об), 1% анизола (об/об) и 0,9% этандитиола (об/об). Полимер нейтрализовали 10% диизопропилэтиламином (в ДМФ) и хранили в ДМФ при 4oC.
Линкерный β-аланин-аминокапроновая кислота - этилендиамин в целом состоит из 11 атомов C и 4 атомов с максимальной длиной плеча 17,6 А. Поскольку в каждой из пяти стадий рандомизированного связывания использовали 19 различных аминокислот, то теоретическое число пептидов в этой библиотеке составляло 195 или 2476099.
Как указывалось выше, основная идея этого метода заключается в том, что огромную библиотеку рандомизированных пептидов, находящихся на отдельных твердофазных полимерных микросферах, синтезируют таким образом, что каждая полимерная микросфера содержит один вид пептида. Каждая отдельная полимерная микросфера, взаимодействующая с акцепторной молекулой, может быть затем идентифицирована и физически выделена, а аминокислотная последовательность пептидного лиганда может быть затем определена методом деградации Эдмана. Поэтому для успешного осуществления этой методики необходима точная идентификация пептидной последовательности на одной сфере. Используя автоматический пептидный секвенатор (Model 477А-01 Pulsed Ligued Automatic Protein (Peptid Seguencer, Applied. Biosystems, Foster City, Калифорния), из каждой полимерной микросферы обычно выделяли 50-500 пМ пептидов. Кроме того, предварительный анализ (DiMarch и др.. 1990, "Peptids Chemistry, Structure and Biolody", Proccedings of the Eleventh American Peptide Symposium, июль 9-14, 1988, La Solla, Ca ESCOM, Leiden стр.229-230) данных секвенирования показал, что эффективность связывания при синтезе твердофазных пептидов превышала 98%.
10.1.2. Специфическая идентификация и отбор пептидных лигандов из библиотеки
Идентификация и отбор специфических пептидных лигандов из рандомизированной библиотеки могут быть легко осуществлены с помощью иммунологической техники, например, такой как твердофазный иммуноферментный анализ (ELISA), иммунофлуоресценция, или с помощью использования иммуномагнетических микросфер. Для экспериментов, описываемых в настоящем примере, в системе обнаружения использовали иммуногистохимическую технику. В настоящем эксперименте в качестве специфически связывающихся акцепторных молекул использовали (i) биотинсвязывающий белок (стрептавидин) и (ii) моноклональное антитело (МАТ) против β-эндорфина. Используя библиотеку эпитопов гибридного нитчатого фага (Cwirla и др., De Vlin и др., см.выше, часть 2) и обе из указанных акцепторных молекул, были успешно идентифицированы пептидные лиганды.
Для обнаружения стрептавидин-связывающих микросфер была использована иммуногистохимическая техника. Рандомизированную библиотеку осторожно смешивали с постепенно возрастающим количеством добавляемой дистиллированной воды для разведения ДМФ. Затем микросферы тщательно промывали PBS, а для блокирования любого неспецифического связывания использовали желатин (0,1 % мас. /об). Затем в течение 1 часа, слегка размешивая, к этим микросферам добавляли 1: 200000 разведение стрептавидин-щелочной фосфатазы (Pilrce, Rockford, Иллинойс). После этого микросферы тщательно промывали и добавляли стандартный субстрат 5-бромо-4-хлор-3-индолилфосфат/нитросиний тетразолий (BCIP/NBT). Микросферы вместе с субстратным раствором переносили на 15 полистироловых чашек Петри (100 х 20 мм) и проводили реакцию в течение почти 2 часов. Микробусы со связанной стрептавидин-щелочной фосфатазой окрашивались в темно-синий цвет, тогда как большинство микробус в библиотеке оставались неокрашенными.
10.1.3. Определение аффинности пептидных лигандов.
Сродство связывания пептидного лиганда с моноклональным антителом (МАТ) против β-эндорфина определяли в фазе раствора. В анализе связывания анти-β-эндорфинового МАТ измеряли ингибирование пептидным лигандом связывания 5,0 нМ [3H] [Leu] энкефалина (специфическая активность = 39,0 Ки/мМ, New England Nuclear, Бостон, МА) с 125 - 200 нг/мл анти-β-эндорфинового МАТ в 1,0 мл 40 мМ Трис-HCl, 150 мМ NaCl, pH 7,4 буфера, содержащего 1,0 мг/мл альбумина бычьей сыворотки, 0,1% (об/об). Твин-20 и 0,05 % (мас./об.) азида натрия. Специфическое связывание определяли как разность между связыванием, измеренным в присутствии или в отсутствии 1,0 мкМ немеченного [Leu] энкефалина. Связанный радиолиганд осаждали путем добавления 10-кратного избытка Белка-G-Сефарозы (Pharmacia) с последующим инкубированием в течение ночи (23-24oC). Белок-G-Сефарозу собирали путем центрифугирования (13000-х g, в течение 5 минут), и осадок суспендировали в 250 мкл 5% (по объему) уксусной кислоты перед переносом в сосуды для подсчета сцинтилляционной жидкости. Величины K0 (n=3) определяли путем анализа методом насыщения с использованием 5 концентрацией радиолиганда (1,87 - 30 нМ) для [3H] [Leu] энкефалина с дублированием каждого из образцов для полного и неспецифического связывания. Средняя величина K0 составляла 9,79 ± 4,63 нМ. Кривую ингибирования пептидным лигандом строили для восьми концентраций пептида в 400-кретном диапазоне. Данные связывания для насыщения и ингибирования анализировали методом нелинейной регрессии с использованием соответствующих моделей с одним сайтом (Khapp и др., 1990, J.Pharmacol.Exp.Ther.; 255:1278-1282). Ki - величины для констант ингибирующего связывания по сродству рассчитывали по методу Cheng и Prusoff (1973, Biochem Pharmacol, 22:3099-3102). Каждую K - величину рассчитывали по 3-4 независимым измерениям.
10.2 Результаты
Скринировали большую синтетическую неупорядоченную пептидную библиотеку (X-X-X-X-X-полимер, где X = 19 известных аминокислот (кроме цистеина) всего 195 = 2476099 пермутаций). В части скринированной библиотеки присутствовали приблизительно 2 миллиона микросфер. В первой стадии отбора, с использованием лишь стрептавидин-щелочной фосфатазы (см. часть 10.1.2, см.выше), приблизительно 75 микросфер были окрашены с различной интенсивностью цвета; их физически выделяли и удаляли под препаровальной лупой с помощью микроманипулятора. Затем каждую микросферу промывали с 8 М гуанидингидрохлорида для удаления стрептавидин-ферментного конъюгата. После этого, каждую микросферу отдельно помещали на стеклянный фильтр для патрона белкового секвенатора (ABI) Applied Biosystem. В табл. 1 показаны последовательности 28 и 75 микросфер. Все указанные микросферы имели обобщающую типичную последовательность либо HPO, либо HPM. Фотомикрограмма на фиг. 7 иллюстрирует, каким образом положительная (темно-синяя) микросфера может быть легко идентифицирована на фоне многих тысяч отрицательных (бесцветных) микросфер в процессе скрининга библиотеки пептидных лигандов.
Для доказательства того, что типичная обобщающая последовательность действительно связывается с биотин-связывающим сайтом в молекуле стрептавидина, синтезировали LMPQF-β-AIa-аминокапроновая кислота - этилендиаминполимер (LHPQF-полимер). Затем LHPQF-полимер смешивали со стрептавидин-щелочной фосфатазой в присутствии различных концентраций биотина. Результаты представлены на фиг. 8. При 10 нМ биотин полностью блокировал окрашивание LHPQF-полимера. При концентрациях 10 и 1,0 нМ биотин частично ингибировал окрашивание, а при концентрации 0,1 нМ биотин не оказывал никакого влияния на окрашивание LHPQF-полимера стрептавидин-щелочной фосфатазой. Анализ ингибирования показал, что обобщающая типичная последовательность связывается с биотин-связывающим сайтом стрептавидина.
Перед тем, как использовать ту же самую неупорядоченную пептидную библиотеку для скринирования с использованием анти-β-эндорфинового МАТ, все синие микросферы, которые были окрашены лишь одной стрептавидин-щелочной фосфотазой, удаляли. Для удаления любого связанного белка оставшиеся микросферы затем обрабатывали 8 М гуанидингидрохлорида.
Эту рециклизованную библиотеку затем смешивали с биотинилированным анти-β-эндорфиновым МАТ (анти-β-эндорфин, клон 3-Е7, был закуплен у Boehringer Mannhein, Индиана) в течение 16 часов. После экстенсивной промывкой проводили вторую стадию с использованием стрептавидина-щелочной фосфатазы в целях стимулирования реакции окрашивания для ELISA. В этом скрининге были идентифицированы 6 пептидов с типичной обобщающей последовательностью, которые очень схожи с нативным лигандом, YGGFL-энкафалином (YQGM): YGALQ, YGGLS, VGGFA, YGGFT, YGGFQ. Были синтезированы пептидные аналоги вышеуказанных лигандов с различными карбокси-концами, а их аффинности (Ki) определяли, используя [3H] (New - энкафалин (New England Nuclear, Бостон, Массачусетс) в качестве меченного лиганда и немеченные пептиды в качестве конкурентного лиганда (анализ с использованием анти-β-эндорфина, см. часть 10.1.2). Результаты этих исследований систематизированы в табл. 2.
YGGI представляет собой [Leu] энкафалин, нативный лиганд для анти-β-эндорфинового МАТ.
10.3. Обсуждение
При использовании методологии настоящего изобретения ключом к успеху является возможность синтеза отдельных пептидов на каждой микросфере в сочетании с чувствительным и специфическим обнаружением и системой отбора. Эта новая методология называется "селективным способом". Отдельную пептидную микросферу, выбранную из библиотеки и обработанную 8 М гуанидингидрохлоридом (для удаления связанного белка) эффективно очищали, а оставшийся пептид ковалентно связывали с полимером и подготавливали для секвенирования. Стандартные автоматические пептидные секвенаторы пригодны для секвенирования пептида при концентрациях до 5-10 нМ. Кроме того, синий краситель, который необратимо связывается с микросферой, не препятствует секвенированию. Во время каждого цикла рандомизации все усилия были направлены на оптимизацию методологии синтеза, включая использование больших избытков Nα-Fmos-аминокислот. Предварительный анализ ряда данных секвенирования ясно показал, что эффективность связывания в синтетических химических реакциях превышала 98%.
Поскольку окрашенные микросферы отчетливо видны на фоне бесцветных микросфер (например, на фиг. 7), то просмотреть визуально под препаровальной лупой 2 миллиона микросфер в серии 10-15 чашек Петри и отобрать реактивные микросферы для секвенирования почти не составляет усилий. Кроме того, оценить относительную аффинность различных лигандов можно путем оценки относительной интенсивности окрашивания каждой положительной микросферы. Это позволяет отобрать микросферы со специфической интенсивностью окраски для секвенирования.
Devlin и др. (1990, Science 259: 404-406, часть 2, см.выше) указывали на важность типичных обобщающих последовательностей HPQ, HPM и HPN в их 20 стрептавидин-связывающих лигандах, выделенных с помощью гибридного нитчатого фага. Из 20 изолятов 15 имели HPQ, 4 имели HPM и 1 имел HPN. Интересно отметить, что пептидная библиотека дала 28 различных пептидов, из которых 23 имели последовательность HPO, а 5 имели последовательность HPM (табл. 1). Очевидно, что положение последовательности HPQ/HPM в пептапентиде не является важным для связывания со стрептавидином. Из всех идентифицированных последовательностей HPQ или HPM пентапептида только две были повторяющими (TpHPQ и MHPMA). Эти результаты идут вразрез с данными Devlin и др. (см.выше), где среди 20 изолятов имеется множество повторов, что позволяет предположить о наличии стандартной ошибки в селекции по используемому ими биосинтетическому методу.
В системе анти-β-эндорфина были идентифицированы 6 пептидов с последовательностями, очень схожими с нативным лигандом YGGF1 (табл. 2). Эти результаты аналогичны результатам, полученным с использованием гибридного нитчатого фага, где использовалось то же самое моноклональное антитело (клон 3-E7) (Cwirla и др., 1990, Proc. Nate. Acad. Sci. USA 87:6378-6382, см.выше). Хотя данная пептидная библиотека дала меньше лигандных последовательностей, чем было получено Cwirla и др., однако 50% полученных лигандов имели гораздо большее сродство к антителу, чем лиганды, выбранные способом с использованием фага.
11. Пример: Ограниченная пептидная библиотека
В другой серии экспериментов настоящее изобретение тестировали с использованием системы другого антитела, где эпитоп был расположен в середине пептидной цепи, а не в ее N-конце (как в случае β-эндорфина). Используемым антителом было анти-V-mos-пептидное моноклональное антитело (анти-V-mos-MAT) (см. ниже, часть 11.1). Это антитело было получено с помощью иммунизации мышей пептидом из 12 аминокислот (LQSGGFGSYKA), соответствующих остаткам 100-111 онкогенного продукта V-mos. Этот пептид конъюгировали с белком-носителем перед иммунизацией. B ELISA-тесте анти-V-mos-MAT обнаруживает гомологичную последовательность V-mos, mos, mos neu/HER1 и HER-2-генных продуктов.
11.1 Материалы и методы.
Используя коммерчески доступный набор для картирования эпитопов (Cambridge Research Biochenricol, Бостон) в многошпилечной системе (Geysen и др. , 1986, Mol. Ymmunol, 23:709-715) для синтеза перекрывающихся пептидов (серии тетрапептидов, пентапептидов и гексапептидов), картировали эпитоп внутри V-mos-пептида из 12 аминокислот, распознаваемого анти-V-mos-MAT, для последовательности пентапептида (FOSSVV) (т.е. остатков 6-10 V-mos-додекапептида).
В настоящем эксперименте использовали ограниченную рандомизированную библиотеку, где аминокислоты валин и серин, которые присутствуют в V-mos-эпитопе, были намеренно исключены. Эта ограниченная рандомизированная библиотека имела следующий состав: G-X-X-X-X-X- β-Ala-аминокапроновая кислота - этилендиаминполимер, где X = Glu, Pro, Asn, Phe, Leu, Thr, Lys, Leu, Gly, Tyr, Ala, Met, Arg, Trp. Эти 14 аминокислот были выбраны таким образом, что валин и серин были исключены, но при этом все функциональные группы боковых цепей были включены, то есть (i) выбирали Asn, но не Glu; (ii) выбирали Glu, но не Asn; (iii) выбирали Thr, но не Ser; (iv) выбирали Leu, но не Phe или Val; или (V) выбрали Met, но не Cys. Поскольку 14 аминокиcлот выбирали в каждой из пяти стадий рандомизированного связывания, то теоретическое число пептидов составляло 145 или 537824 отдельных пептидов.
Клеточная линия гибридомы, которая продуцирует анти-V-mos-MAT, была закуплена у Microbiological Associatod. Jnc., Maryland (Гибрилома N 165-28Е7, SCRF 354, серия N 165-119).
Сродство пептидного лиганда к анти-V-mos-MAT измеряли посредством связывания в фазе раствора с использованием [3H] ацетил-V-mos-пептида ([3H]-V-mos-) в качестве радиолиганда. Этот радиолиганд получали путем - концевого ацетилирования V-mos-пептида перед разблокированием боковых цепей с использованием эквимолярного количества [3H] ацетата натрия (специфическая активность = 2,52 Ки/мМ, Neu England Nugleur, Бостон, МА). [3H] Ac-V-mos-продукт, который был выделен из непрореагировавшего V-mos-пептида с помощью обращенно-фазовой ЖХВР, имел специфическую активность 2,50 Ки/мМ. Аффинность связывания 3H Ac-V-moc- для анти-V-mos MAT (=10 мкг/мл) измеряли в 1,0 мл PBS-желатинового буфера (0,05% желатина в PBS) при 23-24oC, через три часа после инкубации. Специфическое связывание определяли как разницу между связыванием [3H] Ac-V-mos в присутствии (неспецифическое) и в отсутствии (полное) 1000 мкМ немеченного V-mos-пептида для каждой концентрации [3H] Ac-V-mos]. Связанный радиолиганд выделяли путем центрифугирования, используя 10-кратный избыток (способность к связыванию по отношению к используемому иммуноглобулину) белок-G-Cефарозы (Pharmacia) для осаждения антитела. Анализ связывания методом насыщения по пяти определениям в диапазоне концентраций 125-5000 нМ показал, что [3H]-Ac-V-mos связывался с анти-V-mos MAT с величиной Kd, составляющей 850 ± 160 нМ. Аффинности связывания пептидных лигандов определяли путем анализа ингибирования связывания с использованием семи концентраций пептидного лиганда при конкуренции для 10 мкг/мл анти-V-mos MAT с 20 нМ [3H] Ac-V-mos в описанных выше условиях. Анализ ингибирования показал, что свыше 50% полного связывания было специфическим. Константы связывания при ингибировании и насыщении определяли для связывания с одним сайтом путем анализа методом нелинейной регрессии, как было описано ранее (Knapp и Yamamura, 1990, Life, 46:1457-1463).
11.2. Результаты
Приблизительно 230000 микросфер скринировали с использованием конъюгата "анти-V-mos-щелочная фосфатаза". Поэтому было исследовано менее, чем 43% пермутаций. После инкубации с субстратом около 50 микросфер обнаруживали интенсивную синюю окраску, 4 из этих микросфер физически выделяли и определяли аминокислотную последовательность 11 из них. Кроме того, 17 неокрашенных микросфер выбирали произвольно для секвенирования.
Результаты секвенирования анти-V-mos-лиганда представлены в табл. 3. Поскольку валин и серин были намеренно не включены в эту пептидную библиотеку, то не было ничего неожиданного в том, что ни одна из последовательностей 11 пептидных лигандов не напоминала нативный эпитоп (FCSW). Хотя в этих 11 пептидных лигандах не было повторов, однако их последовательности были нерандомизированными. В этих последовательностях часто встречаются аргинин и тирозин. Кроме того, по крайней мере, один, а иногда два аргинина присутствуют во втором и/или третьем положении каждого из этих пептидных лигандов. С другой стороны, отобранные произвольно отрицательные микросферы не обнаруживали каких-либо общих типов аминокислотных последовательностей. Хотя размер образцов был ограничен, однако статистическое chi-квадратичное соответствие для последовательностей от негативных микросфер было незначительным (x2 = 18,27, df = 13, P = 0,15), что указывает на отсутствие каких-либо свидетельств о неравномерном распределении аминокислот для неокрашенных произвольных пептидных микросфер.
Были синтезированы некоторые из положительных лигандов, а их аффинность для анти-V-mos MAT определяли путем анализа связывания в фазе раствора (см. выше часть 11.1). Результаты этих анализов представлены в табл. 4.
В табл. 4A представлены аффинности связывания V-mos-эпитопа (определенного путем картирования эпитопов) (для анти-V-mos моноклонального антитела (МАТ). Показано также влияние изменения карбокси-конца. В табл. 4B представлены аффинности связывания мимеотопов, идентифицированных из пептидной библиотеки, где отсутствует несколько аминокислот с V-mos-эпитопе. В некоторые из тестируемых лигандов был включен амин β-аланина для моделирования структуры идентифицированного лиганда, с которым связывается антитело на микросфере.
В табл. 4 аффинность наилучшего анти-V-mos-мимеотопа приблизительно в 2,5 раза меньше, чем аффинность эпитопа нативного пептида. Хотя ни один из исследованных пептидных лигандов не имел такую же величину Ki, как нативный V-mos-лиганд, однако полученные результаты ясно показывают, что используя рандомизированную библиотеку, в которой отсутствуют некоторые из аминокислот, присутствующие в нативном эпитопе, могут быть идентифицированы серии структурно отличающихся мимеотопов с аффинностью к акцепторной молекуле, т. е. к анти-V-mos- МАТ.
11.3. Обсуждение
Анти-V-mos МАТ, используемые в настоящем исследовании, имели относительно низкую аффинность к V-mos-пептиду (иммуногену). Серин и валин присутствовали в линейном эпитопе (FGSVY), но были намеренно исключены из библиотеки, используемой для скрининга, тем не менее описываемый способ позволяет определить линейный мимеотоп совершенно другой последовательности и с другим составом аминокислот, но со сравнимой аффинностью к антителу. Отсюда видно, что макромолекулярно-пептидные взаимодействия имеют сложную и многостороннюю природу.
12. Пример: Селективно отщепляемый линкер
Получали и испытывали серию четырех пептидов, включающих в себя УФ-отщепляемый линкер GNb (см. выше, часть 5.4).
12.1. Материалы и методы
Используя технику твердофазного синтеза (например, см. выше части 6 и 7), получали следующие четыре пептида на двух полимерах:
(i) Fmos-Trp-Tyr(OBut)-Phe-ONb-β-Ala-AСA-EDA- Pep-SynK
(ii) TPP-Tyr-Рhe-ONb-β-Ala-ACA-EDA-Pep-SynK
(iii) Fmos-Tyr(OBut)-Phe-ONb-β-Ala-ACA-4-MBHA
(iv) Trp-Tyr-Phe-ONb-β-Ala-ACA-4-MBHA
В течение 1 и 3 часов каждый пептид подвергали ультрафиолетовому (УФ) облучению и облучению видимым светом (ВИЗ) в 0,3 мл воды или 3/7 смеси хлороэтанола и дихлорометана. В целом проводили и анализировали 16 + 16 экспериментов.
После облучения супернатант отфильтровывали, лиофилизовали и снова растворяли до получения равных объемов MeOH (0,3 мл). Продукты анализировали с помощью ЖХВР.
12.2. Результаты
Анализы, проведенные с помощью ЖХВР, ясно показали, что при облучении видимым светом в воде трипептид не выделялся, а при УФ-облучении в течение 1 часа наблюдалось почти полное высвобождение трипептида.
В частности, пептид i в воде расщеплялся до простого продукта. В некоторых случаях два продукта наблюдались до элюирования (ЖХВР). При более длительном экспонировании (УФ) взаимное превращение двух продуктов наблюдалось, по крайней мере, в нескольких случаях.
12.3. Обсуждение
Очевидно, что УФ-чувствительный ленкер способствует высвобождению пептида в водном растворе. Экспонирование в течение 1 часа было бы достаточным для неполного высвобождения пептида. Полученные результаты также показали, что полиамидная смола является стабильной и совместимой с водными системами.
13. Пример: Идентификация имитируемого фермента
13.1. Материалы и методы
Неупорядоченную библиотеку пентапептидов, содержащих 19 и 20 известных аминокислот (за исключением цистеина), получали в соответствии с настоящим изобретением (см. выше, часть 10.1).
Эту пептидную библиотеку подвергали воздействию хромогенного субстрата, нитросинего тетразолийхлорида, в отсутствие экзогенного фермента и в условиях, приемлемых для образования продукта; и идентифицировали положительные микросферы. Эти микросферы отбирали и секвенировали.
13.2. Результаты
Последовательности пяти микросфер, которые, очевидно, катализуют восстановление нитросинего тетразолия до его темно-синего диформазанового продукта, представлены в табл. 5.
13.3. Обсуждение
Полученные результаты позволяют предположить, что пептидная PNNNH-микросфера обладает способностью к восстановлению нитросинего тетразолия (NBT) до темно-синего диформазанового пигмента либо химически, либо ферментативно. Таким образом, пептид или пептид на микросфере продемонстрировали свою активность в качестве "искусственного фермента" или в качестве имитатора фермента.
14. Пример: Скрининг на противораковую пептидную последовательность
Ограниченная библиотека, содержащая пентапептиды, включающие в себя тирозин, за которым следует неупорядоченная последовательность, состоящая из пяти аминокислот, выбранных из группы следующих аминокислот: глутаминовая кислота, серин, валин, глицин, агринин и аспарагин, имеет пептидные последовательности с активностью против раковой клеточной линии (т.е. с противоопухолевой активностью).
14.1. Материалы и методы
Неупорядоченную пептидную библиотеку настоящего изобретения синтезировали с использованием гидролизуемого дикетопиперазинового линкера. В скрининге на противораковые (противоопухолевые) лекарственные средства использовали 96-луночные планшеты.
После разблокирования защитной группы боковой цепи и Nα-Fmos-группы библиотеку пептидных микросфер нейтрализовали DIEA (диизопропилэтиламином) и экстенсивно промывали ДМФ для удаления любых оставшихся, возможно токсичных, химических веществ. Затем библиотеку подвергали постепенной замене в 0,01 М HCl (условия, при которых линкер является стабильным) и, наконец, промывали 0,001 М HCl. После этого приблизительно 10 микросфер библиотеки переносили на каждую лунку планшеты. Затем для нейтрализации pH раствора добавляли 50 мкм RPMI-среды (с 25 мМ HEPES-буфера). При нейтральном или слабощелочном pH пептиды будут высвобождаться (например, через 16-24 часа).
Затем, либо (I) часть или все количество среды переносили на отдельную планшету со специфической клеточной линией рака, либо (2) клеточную линию рака добавляли непосредственно в лунки с микросферами. При этом использовали приблизительно 2500 клеток рака легких на одну лунку. Затем эти планшеты инкубировали 8 дней при 37oC, и проводили MTT-анализ для количественной оценки относительной цитотоксичности выделенных пептидов в каждой лунке.
Для скрининга могут быть использованы различные стандартные клеточные линии карциномы, лимфомы, миеломы человека и лейкемии. В качестве примера может служить следующий перечень опухолей (NCl): лимфолейкоз L 1210, меланома B16, карцинома легких Льюиса, саркома M5076, карцинома толстой кишки 38 и карцинома молочной железы человека MX-1. В качестве других примеров могут быть приведены следующие опухоли: рак молочной железы MCF-7, клеточная линия миеломы 8226, клеточная линия лейкозы (мыши) P388 и клеточная линия немелкоклеточного рака легких Ходкенса.
14.2. Результаты
Скринировали библиотеку, содержащую около 8000 пептидов с последовательностью YXXXXX-микросфера, где Y является Tyr, а X является Glu, Val, Gly, Arg или Aru. В двух экспериментах 3-4 супернатанта, содержащих высвободившийся пептид, показали ингибирование роста линии немелкоклеточного рака легких Ходкенса. Поскольку каждая лунка содержала приблизительно 10 микросфер, то, в основном, все 8000 возможных последовательностей были исследованы, в результате чего были идентифицированы не менее 3 активных пептидов. Аналогичные результаты были получены при прямой инкубации микросфер с клетками и при переносе супернатанта с высвобожденным пептидом.
14.3. Обсуждение
Полученные результаты свидетельствуют о том, что ограниченная (укороченная) библиотека пептидов может содержать несколько пептидных последовательностей, проявляющих цитотоксическую активность. В вышеуказанном примере 0,05% возможных последовательностей демонстрировали активность против раковых клеток. При биоанализе супернатантов, содержащих выделившийся пептид в множестве биопроб, можно установить цитотоксичность против раковых клеток и против зрелых тканевых клеток.
В соответствии с этим, можно идентифицировать пептидные последовательности с широкой цитотоксичной активностью к опухолевым клеткам или с цитотоксичной активностью к специфической опухолевой клеточной линии; последовательности с низкой токсичностью к интактным клеткам предпочтительности использовать в качестве лекарственных средств. Указанный метод можно использовать для скрининга противомикробных, противопаразитных средств и антагонистов факторов роста.
Аналогичный метод скрининга агониста фактора роста используют рост-фактор зависимостей клеточной линии. В такой биопробе пептидные последовательности с активностью агониста ростового фактора стимулируют рост клеток, культивированных при отсутствии главного фактора роста.
Настоящее изобретение не ограничивается этими конкретными вариантами его осуществления, описанных в данном описании. Различные варианты осуществления изобретения, кроме раскрытых здесь, должны находиться в пределах объема изобретения, который определен формулой изобретения.
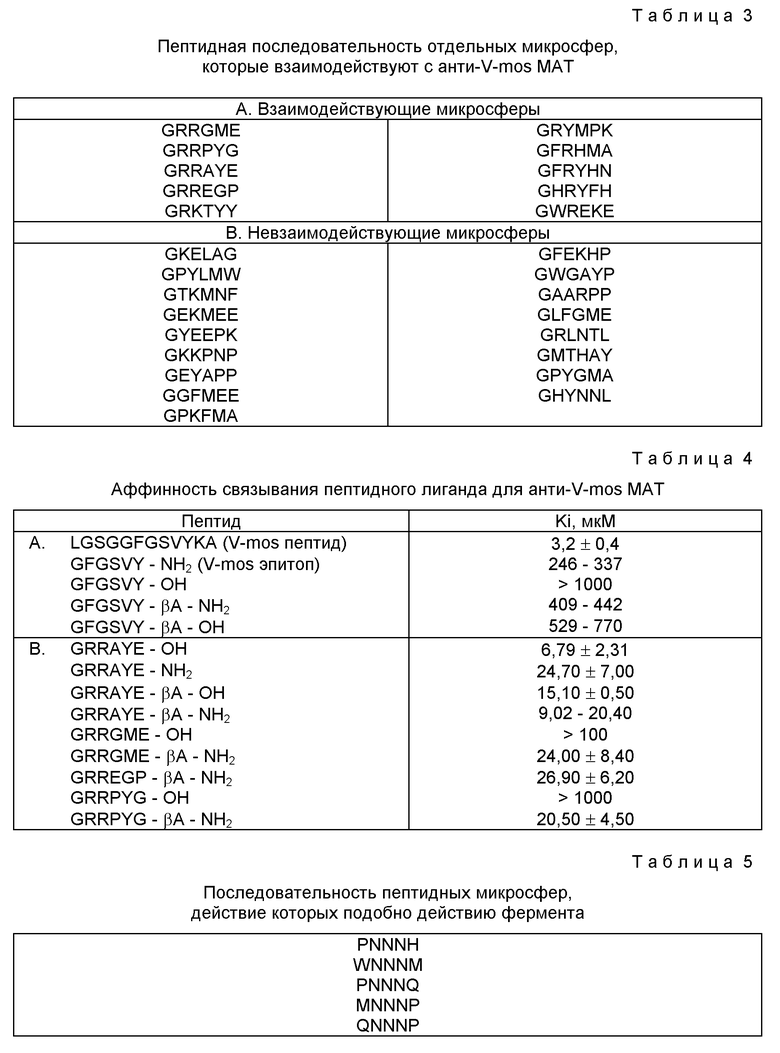
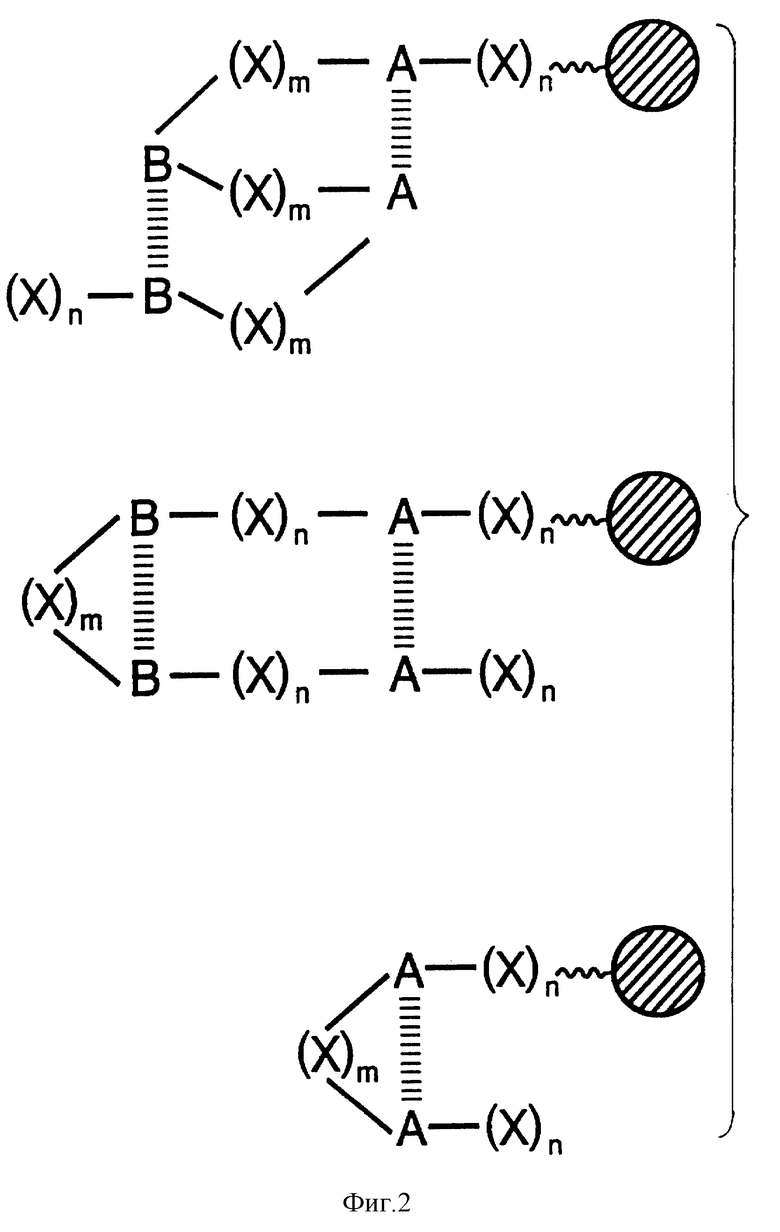





Изобретение относится к биотехнологии и касается библиотеки пептидов, иммобилизованных на твердофазном носителе, способа ее получения и идентификации пептида, синтезированного твердофазным синтезом. Сущность изобретения: библиотека пептидов, иммобилизованных на твердофазных носителях, представляет собой неупорядоченную библиотеку незащищенных пептидов, содержащую многочисленные твердофазные носители, в которой один вид незащищенного пептида связан с каждым твердофазным носителем с включением в библиотеку всех статистически возможных комбинаций субъединиц. Способ получения библиотеки включает введение в алкивоты твердофазного носителя набор из субьединиц таким образом, что в каждую аликвоту вводиться только одна субьединица, имеющая защитную группу, далее осуществляют полностью ковалентное связывание субъединицы со всеми реакционноспособными свойствами твердофазного носителя, удаляют блокирующие группы и удаляют защитные группы. Способ идентификации пептида включает введение в неупорядоченную библиотеку активной молекулы, которая специфически связывается с пептидами библиотеки, выделения полученного комплекса и определяют последовательность пептида этого комплекса. Технический результат заключается в расширении получения различных аминокислотных комбинаций неупорядоченной библиотеки пептидов. 3 с. и 12 з.п. ф-лы, 8 ил., 5 табл.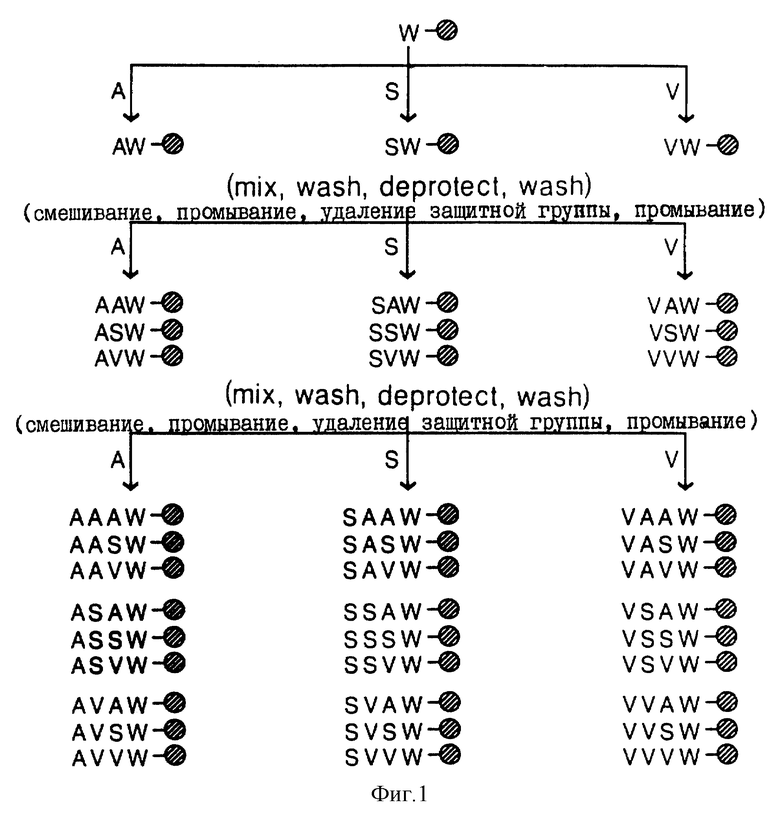
| US 4631211 A, 23.12.86 | |||
| Merrifield R.B | |||
| et al., J.Am.Chem.Soc | |||
| Приспособление к комнатным печам для постепенного сгорания топлива | 1925 |
|
SU1963A1 |
| Прибор для испытания исправности телефонных линий | 1923 |
|
SU2149A1 |
| ANGE WA NDTE CHEMIE, 1985, v.24, N 10 | |||
| Itoh N | |||
| et al., Cell, 1991, v.66, pp | |||
| Крутильный аппарат | 1922 |
|
SU233A1 |
