Изобретение касается линии гибридомных клеток, которые продуцируют антитела человека, которые нейтрализуют вирус гепатита B, способов получения клеточных линий, антител, продуцированных клеточными линиями, и применения антител, в особенности, в лечебных целях.
Создание линий гибридомных клеток для цели продуцирования моноклональных антител в настоящее время, вообще, является делом хорошо известным исследователям, работающим в этой области техники. Настоящее изобретение касается получения моноклональных антител человека, эффективных, в частности, против поверхностного антигена гепатита B /HBsAg/, причем антитела получают в соответствии с общепринятым способом, описанным заявителем в Hybridoma, 2(4): 361 (1983), и в заявке на патент Соединенного Королевства 2113715A, опубликованной 10 августа 1983. Конкретнее, обнаружено, что линия клеток гибридомы, содержащая родительскую иммортализирующуюся клетку грызуна, такую как клетка миеломы мыши, например, SP-2, слитую с партнерской клеткой человека, дает в результате иммортализующуюся ксеногенную гибридомную клетку. Эта ксеногенная гибридомная клетка может быть слита с клеткой, способной продуцировать противо-HBsAg антитело человека, давая в результате новую линию триомных клеток, способную генерировать антитело человека, эффективное против такого антигена человека. С другой стороны, когда желательна большая стабильность, создают линию триомных клеток, которая предпочтительно далее не имеет способности продуцировать свое собственное антитело, и эту триому затем сливают с добавочной клеткой, способной продуцировать нечто полезное против упомянутого антигена, и таким образом получают еще более стабильную гибридому /квадрому/, которая продуцирует антитело против антигена.
Обратимся к более ранним публикациям заявителя, чтобы описать получение ксеногенной гибридомы, названной SPAZ4, получаемой из лекарственно-устойчивой линии клеток SP-2, которую можно получить, например, от NIGM Human Genetic Mutant Cell Repository Ref. GM35669A /см. U.S. DHHS 1982 Catalog of Cell Lines/. Получение SPAZ4 сводится к следующему. Линию клеток SP-2 сливают с нормальными лимфоцитами периферической крови человека обычными техническими приемами. Получают большое число гибридом и приблизительно через пять недель отбирают пять клонов, которые быстро растут и не образуют антител. Эти клетки отбирают по устойчивости к 8-азагуанину и с тремя из этих линий можно получить мутанты, которые устойчивы к 20 мкг/мл 8-азагуанина. В то же время эти клетки чувствительны к среде с гипоксантин-аминоптерин-тимидином /ГАТ/, которая показывает, что они потеряли свою способность продуцировать гипоксантин-фосфорибозилтрансферазу. Одной из таких линий является SPAZ4.
Клеточная линия SPAZ4 может быть слита с клетками, полученными из крови лиц, имунизированных вакциной гепатита B, чтобы получить линии клеток гибридомы, которые дают положительные культуры при использовании стандартных методик отбора, включая связывание антител с вирусными антигенами, о которых идет речь. Предпочтительно, чтобы упомянутые положительные культуры снова включались в процесс вторичного отбора, в котором для получения антигена используются различные субтипы вируса. Это обеспечивает благоприятную возможность для того, чтобы точно засечь правильную антигенную детерминанту, распознаваемую антителом.
Клеточные линии, являющиеся результатом слияния ксеногенной гибридомы и человеческой клетки, продуцирующей моноклональное антитело /триома/, пригодны, следовательно, для получения моноклональных антител, способных эффективно действовать при нейтрализации вируса, вызывающего гепатит, и упомянутые антитела могут, следовательно, предотвращать распространение гепатита через, например, переливание крови. Они также могут быть использованы для начального предохранения новорожденных или облученных людей раньше, чем сможет действовать вакцина. Противогепатитные антитела могут использоваться для защиты пациента с угнетенным иммунитетом, включая больных с пересадкой тканей, от повторного гепатита. Это наиболее важно в случае гепатит B - позитивных печеночных реципиентов. Кроме того, антитела могут использоваться при диагностических испытаниях.
Также обнаружено, что фрагменты антител, такие как Fab-фрагменты, могут также связываться с поверхностным антигеном вируса гепатита B. Эти фрагменты также составляют часть настоящего изобретения.
Специфические антитела, которые конструируют в соответствии с настоящим изобретенем, включают PE1-1, ZM1-1, ZM1-2, MD3-4 и L03-3, причем каждое из этих антител является антителом класса IgGi.
Линия клеток, продуцирующая PE1-1, депонирована в American Type Culture Collection 16 октября 1986 под индексом ATCC HB 9234; линия клеток, продуцирующих ZM1-1, депонирована под индексом ATCC HB 9191 4 сентября 1986 и линия клеток, продуцирующих ZM1-2 депонирована под индексом ATCC HB 9192. Адрес American Type Culture Collection - 12301 Parklawn Drive, Rockville, Maryland 20852.
Линии клеток настоящего изобретения все ведут себя как типичные гибридомы (мышь х человек) х человек и продуцируют соответствующие им антитела при концентрациях в интервале до 25 мкг/л в стандартной суспензионной культуре.
На фиг. 1 показаны результаты иммуноферментного анализа прямого связывания, сравнивающие кинетику связывания антитела PE1-1 /показана одиночной линией/ и антитела ZM1-2 /двойная линия/. Подробности даются в примере 4A.
На фиг. 2 показаны сывороточные уровни антитела PE1-1 в сыворотке макаки резуса, определяемые через разное время после дозировки. Подробности приводятся в примере 4C.
Моноклональное антитело и клеточная линия PE1-1 также описаны изобретателем и названы изобретателем OST 577 и 64-577. Подобным образом, моноклольное антитело и клеточная линия ZM1-2 обозначены как 265-695 и моноклональное антитело и клеточная линия L03-3 обозначаются 266-215.
Антитела и фрагменты антител, полученные в соответствии с изобретением, имеют хорошую специфичность для поверхностного антигена гепатита B при анализах связывания ELISA in vitro.
Так как антитела настоящего изобретения имеют человеческое происхождение, они используются преимущественно для лечения людей, так как не вызывают аллергической реакции при повторном лечении, как это имеет место с антителами мышиными и овечьими. Таким образом, другим аспектом настоящего изобретения является способ лечения гепатита B путем введения одного или нескольких вышеупомянутых антител. Обнаружено, что повторные дозы приблизительно в 10 - 40 мг антитела будут существенно снижать количество циркулирующего HBsAg. Найдено, что дополнительные дозы уменьшают количество HBsAg до уровней ниже предела обнаружения в антигенных анализах.
Еще одном аспектом настоящего изобретения является смесь двух или большего числа моноклональных антител. Такая смесь является особенно подходящей для введения пациентам, которые несут недикий тип штамма вируса гепатита B, который не связывается хорошо с данным единичным моноклональным антителом. Например, нуждающемуся в помощи пациенту, страдающему от злокачественной гепатомы и хронического гепатита B, дают антитело PE1-1 до пересадки печени и повторные дозы после пересадки (подробности приводятся ниже в примере 5). Приблизительно через четыре с половиной месяца лечения могут быть определены низкие уровни HBsAg в сыворотке в случае поликлонального антитела, но не в случае PE1-1. Выполняют ДНК-анализ полимеразной цепной реакции /PCR/ участка в 230 пар оснований гена HBsAg, соответствующего предполагаемому домену связывания моноклонального антитела. PCR ДНК клонируют в бактериофаг M13 и секвенируют получающуюся в результате ДНК. Анализ клонов из каждого из образцов сыворотки обнаруживает двухвариантные последовательности при сравнении с PCR ДНК из исходной печени и антитела до лечения. Вариантная ДНК кодирует две различные аминокислоты в S-протеине HBsAg и также кодирует стоп-кодон /UAG/ в вирусном полимеразном гене. Оба вариантных гена содержат аминокислотное изменение, приводящее к замене аргинина глицином в консервативной области пептида.
Так как показано, что моноклональные антитела PE1-1, ZM1-2, ZM1-1, MD3-4 и L03-3 связывают различные эпитопы и обнаружено, что по крайней мере одно из моноклональных антител связывается с каждым вариантом вируса, испытанным до сих пор в достаточной степени, чтобы сделать его клинически полезным. Еще одним аспектом настоящего изобретения является смесь двух или большего числа моноклональных антител, отбираемых из группы, состоящей из PE1-1, ZM1-2, ZM1-1, MD3-4 и L03-3. Особенно предпочтительными являются смеси двух моноклональных антител, особенно смесь PE1-1 и ZM1-2 и смесь PE1-1 и L03-3. Соотношение моноклональных антител, присутствующих в смеси, может изменяться в зависимости от многих факторов, очевидных для специалистов в этой области техники и включающих генотип вируса гепатита или вируса, присутствующего в сыворотке пациента, относительную эффективность связывания выбранных антител, эпитопы, с которыми связываются выбранные антитела, и экономические соображения. Как правило, антитела будут присутствовать при отношении 1:99, типичнее 25:75 и предпочтительно по существу в равных количествах.
Секции PE1-1, ZM1-1, ZM1-2 и MD3-4 секвенируют, используя стандартные технические приемы. Последовательность, полученная для VH-области PE1-1, дается в табл. 8-1 и отмечаются области, соответствующие CDR1, CDR2 и CDR3 /DH и JH4/. Так как CDR-области являются особенно важными участками при определении связывающих свойств антитела, настоящее изобретение включает антитело, которое имеет аминокислотную последовательность CDR1-области, которая, по существу, подобна последовательности PE1-1, как показано в табл. 8-1. Настоящее изобретение также включает антитело, которое имеет аминокислотную последовательность области CDR2, которая, по существу, подобна последовательности PE1-1, как показано в табл. 8-1. Кроме того, настоящее изобретение также включает антитело, которое имеет аминокислотную последовательность области CDR3, которая, по существу, подобна области CDR3 PE1-1, как показано в табл. 8-1.
Подобным образом секвенируют VH-область ZM1-1, как дается в табл. 8-2. Также указываются области, соответствующие его CDR1, CDR2 и CDR3 /DH и JH4/. Настоящее изобретение включает антитело, которое имеет аминокислотную последовательность области CDR1, которая, по существу, подобна последовательности ZM1-1, как показано в табл. 8-2. Кроме того, настоящее изобретение включает антитело, которое имеет аминокислотную последовательность области CDR2, которая, по существу, подобна последовательности ZM1-1, как показано в табл. 8-2, и далее настоящее изобретение также включает антитело, которое имеет аминокислотную последовательность области CDR3, которая, по существу, подобна последовательности ZM1-1, как показано в табл. 8-2.
ДНК-последовательности, которые кодируют области ZM1-2 и MD3-4, даются в таблицах 8-3 и 8-4 соответственно. Настоящее изобретение также включает любое антитело, которое имеет аминокислотные последовательности, которые, по существу, подобны последовательностям областей ZM1-2 и MD3-4, указанным в таблицах 8-3 и 8-4.
Определяют ДНК-последовательности, которые кодируют VH-области PE1-1, ZM1-1, ZM1-2 и MD3-4, и представляют их в таблицах 8-1 и 8-2, 8-3 и 8-4 соответственно. Эти последовательности или соответствующие фрагменты могут быть использованы в клонирующих антителах /или модифицированных антителах/ или в качестве зондов. Антитела, которые получают методами генной инженерии /скорее, чем обычным отбором из гибридом/, могут быть созданы с использованием технических приемов клонирования, которые известны в технике. Для получения молекулы искусственного антитела может быть использована ДНК из других источников, которая сохраняет свойства связывания PE1-1, ZM1-1, ZM1-2 и MD3-4 за счет наличия областей, подобных, по существу, CDR1, CDR2 и/или CDR3. Такие антитела входят в объем настоящего изобретения.
ДНК-последовательности, которые кодируют вариабельные районы VL легкой цепи PE1-1, ZM1-1, ZM1-2 и MD3-4, даются в таблицах 9-1, 9-2, 9-3 и 9-4 соответственно. Настоящее изобретение также включает любое антитело, которое имеет аминокислотные последовательности, которые, по существу, подобны последовательностям областей PE1-1, ZM1-1, ZM1-2 и MD3-4, указанным в таблицах 9-1, 9-2, 9-3 и 9-4.
Также в объем настоящего изобретения входят ДНК-последовательности, которые кодируют VH-область, VL-область, CDR1-области, CDR2-области и/или CDR3-области PE1-1, ZM1-1, ZM1-2 и MD3-4. Также включаются ДНК, которые будут скрещиваться с любой из вышеупомянутых последовательностей в условиях жесткой гибридизации. Такие ДНК, по существу, свободны от другой ДНК млекопитающего-донора и могут содержать интроны или могут быть кДНК.
В описании и формуле изобретения используются определения, которые имеют следующие значения. Аминокислотная последовательность является "по существу, подобной" другой аминокислотной последовательностью, если их аминокислотная гомология составляет по крайней мере 80%. Относительно ДНК "условиями жесткой гибридизации" являются условия, когда гибридизацию осуществляют при 60oC в 2,5 х физиологический раствор - цитратный буфер /SSC/, после чего следует только промывка при 37oC буфером уменьшенной концентрации, который не будет влиять на гибридизации, которые происходят. "Ассоциированная ДНК млекопитающего" означает ДНК, присутствующую у млекопитающего, которая является источником VH-цепи антитела, но которая не включается в кодирование антитела или фрагмента антитела.
Настоящее изобретение дополняется следующими далее примерами, которые не являются ограничивающими.
Пример 1
Получение клеточных линий антител
Добровольцев иммунизируют вакциной гепатита B. Клеточные линии гибридом MD3-4, ZM1-2, ZM1-1 и PE1-1 получают из лимфоцитов лиц, иммунизированных Heptavax® /Merck & Co./. Клеточную линию L03-3 констатируют из клеток лиц, инъецированных несколько раз Heptavax®, и непосредственно перед слиянием - Recombivax® /Merck & Co./. Лимфоциты периферической крови очищают в градиенте плотности на градиенте Перколла /Pharmacia Inc./, плотность 1,085 г/мл. Выделенные лимфоциты промывают три раза сбалансированным солевым раствором Хэнкса и смешивают с равным числом клеток из клеточной линии (мышь х человек) SPAZ-4. Смесь клеток осаждают центрифугированием при комнатной температуре при 400•g в течение 5 минут. После удаления среды клеточный осадок обрабатывают 50% раствором ПЭГ-1000 в минимальной поддерживающей среде (МЕМ) Дульбекко в течение 1 минуты при 37oC, после чего среду слегка разбавляют МЕМ Дульбекко. Клетки собирают центрифугированием и ресуспендируют в МЕМ Дульбекко, содержащей 20% фетальной коровьей сыворотки. Клетки высевают - приблизительно 2•106 клеток на мл - в микроячеистые планшеты. На следующий день добавляют свежую среду, содержащую компоненты ГАТ-среды /среда с гипоксантин-аминоптерин-тимидином/, чтобы отобрать неслитые клетки SPAZ-4. На 4-й день после слияния среду заменяют свежей средой, содержащей только ГТ, так как все клетки, чувствительные к ГАТ-отбору, за это время убиты.
Через 3 - 4 недели, когда в микроскоп наблюдают хороший рост гибридомаподобных клеток, супернатант проверяют на присутствие антитела против поверхностного антигена гепатита B. Применяют ELISA анализ, использующий разбавление 1/100 Heptavax®, на твердой фазе. После инкубирования с супернатантами планшеты обрабатывают набором биотинилированного козьего противочеловеческого иммуноглобулина и пероксидазы хрена, смешанной с авидином /Vestastain®, Vector Laboratories Inc./. Фермент определяют по реакции окрашивания с фенилендиамином. Позитивные культуры пикируют в новые ячейки и часть клеток клонируют путем ограничивающего разбавителя с МЕМ Дульбекко, содержащей 20% фетальной коровьей сыворотки и 107 тимоцитов мыши на миллилитр. Планшеты клонирования проверяют тем же ELISA-методом, как описано выше, и позитивные культуры размножают и замораживают.
Пример 2
Иммунохимическая характеристика
A. Класс/подкласс антитела
Определяют класс иммуноглобулинов антител PE1-1, ZM1-1, ZM1-2, MD3-4 и L03-3, используя методологию ELISA. Каждое антитело улавливают накрытым антигеном планшетом и каждый анализ проводят со специфичным, конъюгированным с пероксидазой противочеловеческим Ig /Tago/. Каждое из антител четко является IgGI.
B. Тип легкой цепи
Используя методы ELISA, подобные описанным выше в A, испытывают антитело с реагентом с анти-k- или анти-λ-легкими цепями /Tago/. Получают следующие результаты.
PE1-1 - лямбда, ZM1-1 - каппа, ZM1-2 - каппа, L03-3 - лямбда, MD3-4 - лямбда.
C. Изоэлектрическое фокусирование /IEF/
Образец антитела L03-3 или PE1-1 вводят в гель. Обнаруживают, что каждый ведет себя как основной протеин.
D. Специфичность
Очищенные субтипы HBsAg adw и ayr закупают у Scripps Laboratories, San Diego, California. Субтип HBsAg ayw получают из Connaught Laboratories (Willowdale, Ontario). Анализы ELISA выполняют, по существу, так, как описано Osterg et al. (1983) Hybridoma 2: 361 - 367.
PE1-1 взаимодействует как с ayr, так и с adw, но с субтипом adw он реагирует несколько лучше. L03-3 реагирует по существу одинаково, как с ayr, так и с adw. Эти результаты подтверждаются для PE1-1 и L03-3 анализом Скэтчарда в твердофазном РИА с адсорбированным в твердой фазе ayr- и adw-антигеном. Таким образом, хотя эти моноклональные антитела явно не связываются с субтиповой детерминантой, на их взаимодействие с HBsAg существенным образом может влиять субтип.
E. Определение аллотипа
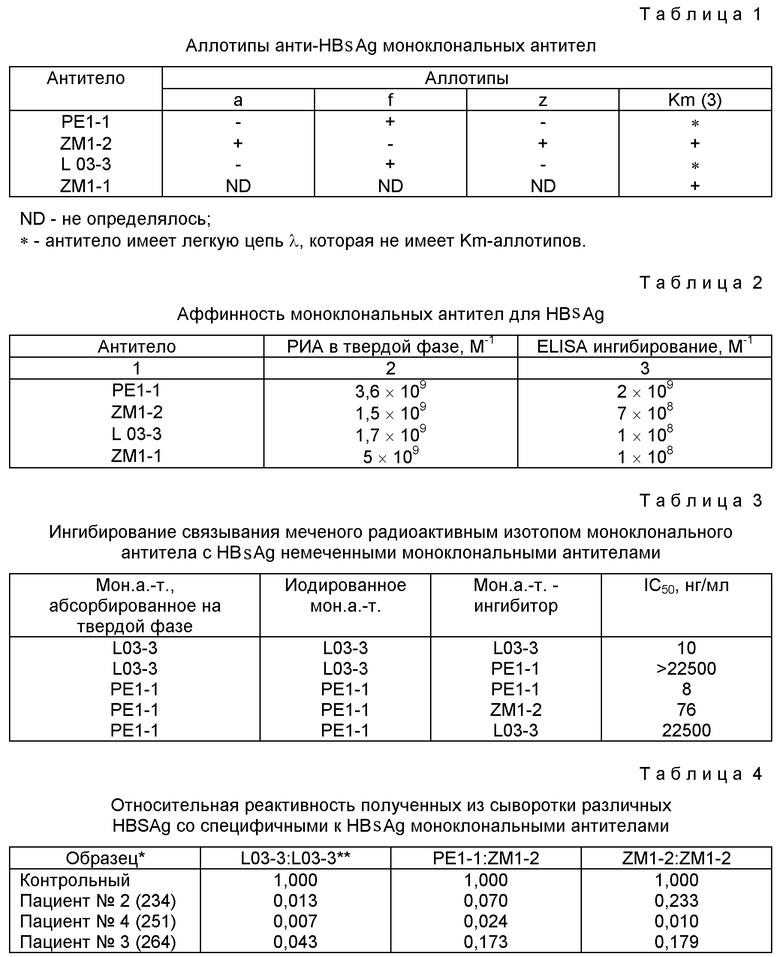
Аллотипы определяют, используя реагенты, поставляемые Central Laboratory of the Netherlands Red Cross Transfusion Service. Используют ELISA ингибирования или ELISA прямого связывания. Результаты приводятся табл. 1. Как можно видеть, не существует явной рестрикции на высокоаффинных анти-HBsAg антителах в отношении легкой цепи или аллотипа.
G. Аффинность
Аффинность для абсорбированного в твердой фазе HBsAg определяют для каждого антитела, используя антитела, меченые радиоактивными изотопами, по существу, так, как описано в Wands et al., (1981) Gastroenterology 80: 225 - 232, что включено в настоящее в качестве ссылки. Антитела метят 125I йодогеном /Pierce/. Для каждого моноклонального тела, за исключением L03-3, абсорбированный в твердой фазе HBsAg представляет собой ayw. L03-3 анализируют как с ayr, так и с adw, по существу, с одинаковыми результатами. Инкубирование антитела-антигена проводят при комнатной температуре.
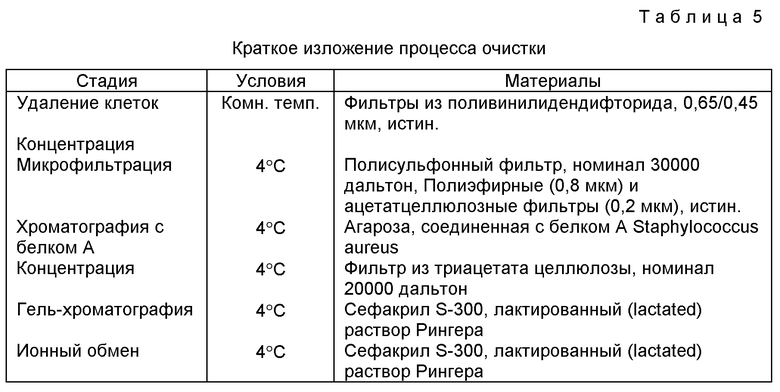
Относительную аффинность определяют также, используя ELISA ингибирования, при котором растворимый HBsAg /субтип ayw/ при изменяющихся концентрациях предварительно инкубируют с моноклональным антителом, и смесь затем инкубируют при 37oC в ячейке микротитратора с нанесенным HBsAg. Результаты приводятся в табл. 2.
Как можно видеть из табл. 2, результаты ELISA как для PE1-1, так и для ZM1-2 приблизительно в два раза ниже, чем результаты РИА, что находится в пределах экспериментальной ошибки. Распределение Скэтчарда результатов РИА, выполненного для ZM1-1, показывает, что возможно существование связывающего центра с низкой аффинностью. Таким образом, возможно, что ELISA замеряет этот центр связывания с низкой аффинностью, так как результаты ELISA в 50 раз ниже, чем РИА. Кроме того, распределение Скэтчарда также показывает, что существуют центры ZM1-1 со значительно менее высокой аффинностью, чем сайты с высокой аффинностью ZM1-2 или PE1-1. Хотя теоретически нет стремления к связыванию, оказывается, что ZM1-1 может иметь самую высокую аффинность к HBsAg из четырех сравниваемых антител, но только к HBsAg с определенным пространственным расположением. Такое расположение обнаруживается только у небольшого процента молекул HBsAg. Также возможно, что это может иметь место благодаря двухвалентному связыванию ZM1-1 и HBsAg, в то время как сайт с низкой аффинностью является одновалентным.
Пример 3
Эффективность PE1-1
Антитело PE1-1 проверяют на эффективность радиоиммунным анализом AUsAB /Abbott/. Испытания осуществляют по отношению к иммуноглобулину гепатита B из Bureau of Biologic Reference и некоторым коммерческим препаратам иммуноглобулина гепатита B /H-Big Immune Globin®, Hep B Gammagee Immune Globin® и Hyper Hep Immune Globin®, все закуплены в фирмах, поставляющих фармацевтические препараты/. Несмотря на то, что препараты иммуноглобулина являются поликлональными, а PE1-1 является моноклональным, данные по связыванию соответствуют критериям Bureau of Biologic для сравнения препаратов иммуноглобулина, т.е. линии параллельны при уровне вероятности, меньшем или равном 0,01.
Определение эффективности проводят следующим образом. Препараты сравнивают по весу /поглощение при 208 нм 1,4 предположительно соответствует 1 мг/мл/. Препараты PE1-1, которые хранились при 5oC, затем сравнивают с вышеназванными поликлональными препаратами, которые также хранились при 5oC. Строят график логарифма 1000, деленной на мкг/мл в препаратах /т.е. логарифма числа, которое обратно пропорционально концентрации иммуноглобулина, подобно логарифму фактора разведения/, от логарифма отсчетов в минуту /среднее из трех измерений/. Гипотезу, что соответствующие прямые являются параллельными, проверяют, используя дисперсионный анализ. Находят, что прямые являются параллельными при уровне вероятности, меньшем или равном 0,01. Прямые для всех препаратов являются параллельными и определяют общий наклон. Из общего наклона вычисляют x-отрезки и различие в отрезках используют для определения различия в эффективности. По этой методике моноклональное антитело PE1-1 является приблизительно в 435 раз более эффективным, чем иммуноглобулин гепатита B от Bureau of Biologic. Так как обнаружено, что коммерческие препараты иммуноглобилуна гепатита B в два /или меньше/ раза более эффективны, чем препарат Bureau of Biologic, PE1-1 является по крайней мере в 200 раз более эффективным, чем коммерческие препараты иммуноглобулина гепатита B, относительно веса.
Пример 4
A. Кинетика связывания
Используют иммуноферментный анализ с прямым связыванием, чтобы сравнить кинетику связывания HBsAg антителами PE1-1 и ZM1-2. На титрационные микропланшеты EI I A наносят Heptavax® с концентрацией 1 мкг/мл. Ячейки затем инкубируют при 37oC с 2% фетальной телячьей сыворотки в забуференом фосфатом физиологическом растворе. Моноклональное антитело PE1-1 или ZM1-2 при содержании 0,5 мкг/мл в 2% фетальной телячьей сыворотки инкубируют в ячейках в течение различного периода времени. В определенное время раствор антитела удаляют и ячейку три раза промывают свежей 2% фетальной телячьей сывороткой. Ячейку затем инкубируют с 2% фетальной телячьей сывороткой до тех пор, пока ячейки в течение 90 минут не показывают содержание 2% фетальной телячьей сыворотки. Раствор затем заменяют пероксидазой, конъюгированной либо с козьими антителами к лямбда - цепи /ячейки с PE1-1/, либо с козьими антителами к каппа-цепи /ячейки с ZM1-2/. Количественное определение конъюгата пероксидазы, связанного с пластиком, осуществляют с добавлением O-фенилендиамина и H2O2. Результаты представлены на фиг. 1, на которой одиночная линия соответствует PE1-1 и двойная линия - ZM1-2.
Как можно видеть из фиг. 1, при концентрации, при которой PE1-1 почти полностью взаимодействует в течение 5 минут, взаимодействие ZM1-2 с адсорбированным на твердой фазе HBsAg не заканчивается за 30 минут и может продолжаться до 90 минут или дольше. Таким образом, при таком анализе PE1-1 связывается с антигеном значительно быстрее. При предположении, что это имеет место и in vivo, PE1-1 является вероятно более эффективным для нейтрализации частиц вируса до того, как они смогут инфицировать печень.
B. Относительное положение эпитопов
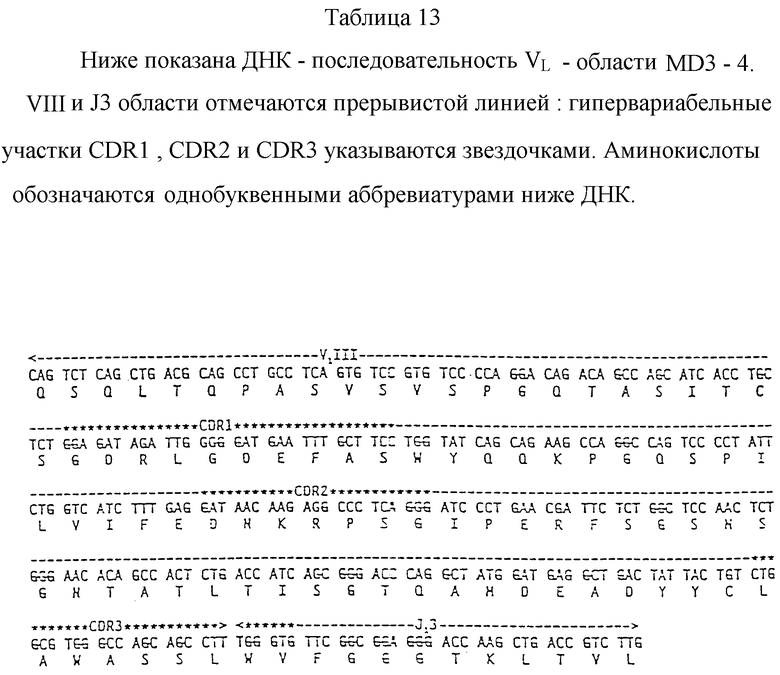
Определяют относительное положение эпитопов антител PE1-1 L03-3 и ZM1-2. Применяют одновременный сэндвич-иммунотест с адсорбированным на твердой фазе моноклональным антителом. Упомянутое антитело метят радиоактивным изотопом и инкубируют в лунке микротитровального планшета с сывороткой больного гепатитом B. Используют Fab-фрагмент меченого PE1-1, в то время как меченый L03-3 представляет собой целый IgG. Результаты приводятся в табл. 3.
Моноклональное антитело ZM1-2 только приблизительно в девять раз менее эффективно при ингибировании связывания 125I-PE1-1 с HBsAg, чем немеченый PE1-1, в то время как L03-3 менее эффективен в тысячи раз. Таким образом, эпитопы ZM1-2 и PE1-1 находятся на молекуле HBsAg вероятно близко один от другого, в то время как эпитоп L03-3 находится вероятно на другой части молекулы. Реципрокный эксперимент - ингибирование PE1-1 меченого L03-3 - служит дополнительным доказательством того, что PE1-1 и L03-3 связываются с эпитопами, которые не перекрываются.
Подобие эпитопов PE1-1 и ZM1-2 и их отличие от L03-3 подтверждаются иммуноанализом с восстановленным и алкилированным HBsAg. L03-3 может связываться с денатурированным антигеном, в то время как ZM1-2 и PE1-1 не могут связываться. Следует отметить, что PE1-1 и ZM1-2 имеют различные эпитопы, так как их взаимодействие с различными субтипами расходится.
C. Фармокинетика PE1-1 у макак резус
Фармокинетику PE1-1 изучают на двух макаках резус. Каждое животное получает единичную внутривенную инъекцию ударной дозы /0,5 мг/кг/ моноклонального антитела PE1-1. Уровни PE1-1 в сыворотке определяют в разное время после дозировки, используя ELISA на основе сэндвич-иммунотеста с Heptavax®, нанесенным на планшеты ELISA и кроличьи антиидиотипические антитела к PE1-1. Результаты приводятся на фиг. 2.
Уровни PE1-1 в сыворотке у двух макак резус характеризуются двумя наклонами /t 1/2 α = 1 и 1,4 дня; t 1/2 β = 11 и 16 дней/ с более коротким временем полужизни, связанным вероятно с фазой распределения моноклонального антитела. Вычисляют объем распределения в устойчивом состоянии /Vdss/, которое составляет 114 - 144% объема плазмы, что наводит на мысль о слабом распространении PE1-1 до тканевого участка у обезьяны, свободной от антигена.
Пример 5
A. Применение PE1-1 из сочувствия к двум пациентам в конечной стадии болезни печени, производной от острого хронического гепатита B и злокачественной гепатомы, переносящим пересадку печени
PE1-1 дают двум пациентам в конечной стадии заболевания печени, которым предстоит пересадка печени. Пациент N 1 - мужчина в возрасте 56 лет, болеющий в течение 20 лет хроническим острым гепатитом, с диагнозом злокачественной гепатомы. Другой пациент - мальчик 10 лет, предполагается, что инфицирован гепатитом B при рождении. У пациента N 2 изначально установлена большая масса правой доли печени, которая, как подтверждает биопсия, является злокачественной гепатомой.
Этим пациентам вводят дооперационные дозы PE1-1 и значительно уменьшают у них уровни циркулирующего HBsAg до процедуры пересадки. Каждый пациент получает также две 20-миллиграммовые дозы PE1-1 во время пересадки. Послеоперационные дозы затем начинают вводить на другой день после хирургического вмешательства.
У пациента N 1 никогда не становится отрицательной реакция на HBsAg, хотя уровень циркулирующего HBsAg у него заметно уменьшается относительно уровня до лечения. У пациента N 2 реакция на HBsAg становится отрицательной, что впервые отмечается на 9-й день после пересадки. Пациент N 1 получает дополнительные дозы PE1-1 в интервале 5 - 40 мг с интервалом в 2 - 20 дней. Пациент N 2 получает либо 5-, либо 10-миллиграмовые дозы в среднем каждые 21 - 28 дней.
Не сообщается о неблагоприятном ходе событий у кого-либо из этих двух пациентов в то время, когда они получают PE1-1. Однако приблизительно через четыре недели после того, как пациент N 1 был выписан из больницы, установлено, что у него имеется метастатическая злокачественная опухоль. Он скончался на 139-й день после операции. Не отмечено явного возвратного гепатита за время после пересадки, несмотря на присутствие заметного количества циркулирующего HBsAg. Хотя ДНК-проба на вирус гепатита B до операции является отрицательной, одно положительное значение определено через 60 дней после пересадки.
На 143-й день после пересадки впервые обнаружено, что пациент N 2 является положительным для HBsAg. Уровень HBsAg непродолжительное время колебался, прежде чем установился на уровне, значительно меньшем уровней до лечения. Изоляты вируса гепатита B этого пациента, полученные до лечения, PE1-1 и в последнее время, анализируют на их способность связывания с PE1-1. Обнаружено, что PE1-1 способен связываться с вариантом вируса, но не настолько хорошо, как с вирусом дикого типа.
Генетический анализ двух вирусных изолятов показывает единичные нуклеотидные различия в высококонсервативной области главного поверхностного белка вируса. Такие различия при сравнении с вирусом до лечения могут потенциально кодировать единичное аминокислотное различие, которое будет уменьшать связывающую способность PE1-1 со связывающей частицей вируса гепатита B.
B. Применение PE1-1 для пациентов с хроническим острым гепатитом B, которые подвергаются пересадке печени /не осложненной злокачественной гепатомой/
Настоящее исследование включает пять пациентов с положительной реакцией на HBsAg /но не имеющих злокачественной гепатомы/ и которые подвергаются пересадке печени. Каждому пациенту в течение трех дней до операции вводят три дневные дозы /10, 20 и 40 мг соответственно/. Пересадку печени осуществляют минимум через два дня и максимум через 32 дня после введения дооперационных доз исследуемого лекарственного препарата. Дополнительную дозу в 40 мг PE1-1 вводят во время операции. Все пять пересадок закончились успешно.
Во время пребывания в больнице у пациентов внимательно контролируют титры HBsAg, фрагменты печени и другие клинические параметры. Продолжаются регулярные наблюдения и введение PE1-1 /приблизительно каждые 1-3 недели/ лечащим врачом каждого пациента. Дозировка и другие параметры меняются от пациента к пациенту.
Двое пациентов /N 5 и 6/ имеют результаты, схожие с результатами у пациента N 2, упоминающимися выше, в том отношении, что вариантный вирус появляется после периода получения отрицательных данных по HBsAg. Сыворотки этих пациентов остаются активными с PE1-1. Анализ последовательностей показывает присутствие отдельных нуклеотидных различий между вариантами из сывороток пациентов и вируса дикого типа. У каждого пациента определяются два варианта. Иммуноанализы и анализ последовательностей показывают, что варианты у каждого пациента различаются и они также отличаются от варианов пациента N 2.
Пациент N 3 - кавказец 39 лет, у которого терминальная стадия заболевания печени в результате 16-летнего заболевания хроническим гепатитом B. Три предоперационные дозы PE1-1, которые вводят пациенту N 3, вызывают существенное снижение его титра HBsAg. На 2-й и 3-й день после пересадки он получает 20 мг PE1-1 и впервые отрицательную реакцию на HBsAg отмечают на 2-й день после пересадки. В течение двух месяцев пациент N 3 получает 10 мг PE1-1 в среднем каждые 1 - 7 дней. Затем он получает 7,5-миллиграммовые или 10-миллиграммовые дозы PE1-1 каждые 14 - 43 дня. Гистопатологическая оценка при биопсии печени, осуществленная в феврале 1989, является отрицательной как на HBsAg, так и на HBcAg. Пациент N 3 остается с отрицательной реакцией на HBsAg в течение 582 дней после пересадки. Кроме PE1-1, он также получает три последовательные ежемесячные инъекции Recombivax® в июле, августе и сентябре 1989 г.
Пациент N 4 - 40-летняя женщина арабского происхождения, у которой терминальная стадия заболевания печени после 10-летнего заболевания хроническим острым гепатитом B. Три дооперационные дозы PE1-1, которые получает пациент N 4, вызывают значительное снижение уровня HBsAg. Пациент N 4 получает 20 мг PE1-1 в 1-й и 2-й день после пересадки, и отрицательную реакцию на HBsAg обнаруживают на 6-й день после пересадки. В течение двух месяцев после этого она получает 10 мг PE1-1 в среднем каждые 3 - 8 дней. Затем она получает 10 мг PE1-1 каждые 5 - 26 дней. Приблизительно через 1 год после пересадки у пациентки развивается гепатитный артериальный тромбоз, но реакция на HBsAg остается отрицательной, и пересадку повторяют. Тремя днями позже, вследствие ишемии, проводят третью пересадку. Через 20 дней проводят четвертую пересадку из-за инфекции. Пациентка скончалась через 18 дней после четвертой пересадки /через 404 дня после первой пересадки/ вследствие печеночной недостаточности и бактериального сепсиса. Гистопатологическая оценка биопсии из печени, пересаженной в первый раз, показывает отрицательную реакцию на HBsAg.
Пациент N 5 - кавказец 38 лет, у которого терминальная стадия заболевания печени после хронического острого гепатита B. Дооперационные дозы PE1-1, вводимые пациенту, существенно снижают у него уровень циркулирующего HBsAg. Пациент N 5 получает 20 мг PE1-1 на 2-й и 3-й день после пересадки и отрицательная реакция на HBsAg обнаруживается на 3-й день после пересадки. В течение первых двух месяцев после пересадки он получает 10 мг PE1-1 в среднем каждые 3 - 7 дней. Позднее он получает 10 мг PE1-1 каждые 9 - 26 дней. У пациента отмечается положительная реакция на HBsAg на 252-й день, хотя уровень антигена у него существенно ниже, чем до пересадки. Гистопатологическая оценка биопсии печени, осуществленная в январе 1990 г, является положительной как на HBsAg, так и на HBcAg.
Пациент N 6 - кавказец 38 лет, у которого терминальная стадия заболевания печени после хронического острого гепатита B и злоупотребления алкоголем. Этот пациент первоначально заразился при переливании крови. До пересадки у него положительная реакция как на HBsAg, так и на HBeAg. Каждая дооперационная доза PE1-1 вызывает снижение у пациента уровня титра HBsAg. Пациент N 6 получает 20 мг PE1-1 на 1-й и 2-й день после пересадки и отрицательная реакция на HBsAg отмечается на 1-й день после пересадки. После этого в течение двух месяцев пациент N 6 получает 10 мг PEI-1 в среднем каждые 3 - 14 дней. Впоследствии он получает 10 мг PE1-1 каждые 7 - 63 дня амбулаторно. Первая положительная реакция на HBsAg отмечается на 251-й день после пересадки и имеет место после самого продолжительного /63 дня/ перерыва между дозами PE1-1. Хотя в настоящее время у пациента N 6 положительная реакция на HBsAg, его титр остается значительно меньшим, чем до пересадки.
Пациент N 7 - 38-летняя женщина кавказского происхождения с IV степенью токсикомании. У этой пациентки терминальная стадия заболевания печени вследствие хронического острого гепатиата B. До пересадки у пациентки положительная реакция как на HBsAg, так и на HBeAg. Каждая дооперационная доза PE1-1 вызывает снижение титра HBsAg у пациентки. В первый месяц после пересадки пациент N 7 получает от 10 до 40 мг PE1-1 в среднем каждые 1 - 7 дней и отмечается отрицательная реакция на HBsAg на 16-й день после пересадки. Затем она получает 10 мг PE1-1 каждые 15 - 29 дней. Гистопатологическая оценка биопсии печени, выполненная в июле 1989, является отрицательной как для HBsAg, так и для HBcAg. У пациента N 7 остается отрицательная реакция на HBsAg на 464-й день после пересадки.
Пример 6
Реактивность с вариантными вирусами
Реактивность моноклональных антител PE1-1, ZM1-2 и L03-3 с различными вирусами гепатита B, выделенными у пациентов, о которых шла речь в примере 5, исследуется. Радиоиммуноанализ осуществляют, определяя радиоактивность границы абсорбированного на твердой фазе антитела. Раствор моноклонального антитела с концентрацией 20 мкг/мл в забуференном фосфатом физиологическом растворе, содержащем 0,02% NaN3, инкубируют по крайней мере в течение 18 часов в ячейках с U-образным дном /Falcon Micro Test III Flexible Assay Plates/. Раствор удаляют из ячеек и ячейки затем промывают три раза дистиллированной водой. Добавляют фетальную телячью сыворотку при концентрации 2% в забуференном фосфатом физиологическом растворе и инкубируют в течение ночи при комнатной температуре с растворами сывороточного HBsAg или контрольных образцов и помеченным 125I антителом /приблизительно 4000 имп. в мин. в 1% фетальной телячьей сыворотке/. Ячейки затем промывают три раза дистиллированной водой. Отдельные ячейки возбуждаются и проводится подсчет. Результаты приводятся в табл. 4.
Примечания к табл. 4:
*Представленные положительные на HBsAg образцы сыворотки взяты от пациентов после пересадки печени и лечения противо-HBsAg лечебным моноклональным антителом PE1-1. Числа в скобках указывают число дней после пересадки.
**L03-3:L03-3 обозначает радиоиммуноанализ, который проводят для моноклонального антитела человека L03-3 как адсорбированного на твердой фазе, так и помеченного радиоактивным изотопом. PE1-1:ZM1-2 обозначает радиоммуноанализ для моноклонального антитела человека PE1-1, адсорбированного на твердой фазе, и помеченного радиоактивным изотопом моноклонального антитела человека ZM1-2, ZM1-2:ZM1-2 обозначает радиоиммуноанализ для моноклонального антитела человека ZM1-2 как адсорбированного на твердой фазе, так и помеченного радиоактивным изотопом. Контрольные образцы положительной на HBsAg сыворотки хорошо взаимодействуют с антителами L03-3, PE1-1 и ZM1-2.
Пример 7
Крупномасштабное производство антител
Чтобы инициировать производственный цикл с клетками, извлекают одну или несколько ампул с замороженными клетками из жидкого азота. После быстрого разогревания на водяной бане при 37oC до тех пор, пока почти весь лед расплавится, ампулу вскрывают в ламинарном боксе. Содержимое ампулы смешивают с 1 мл смеси MEM Дульбекко и Ham's F12 /1:1 по объему/ (DMEM/F12), к которой добавлены соли железа до конечной концентрации Fe+++ 50 мкМ. После перемешивания пробирку наполняют приблизительно до 10 мл той же средой и клетки собирают центрифугированием. Клеточный осадок ресуспендируют в 5 мл вышеупомянутой среды с 20% фетальной коровьей сывороткой и высевают в 1 ячейку 6-ячеечного планшета для культуры тканей. Клетки инкубируют в инкубаторе при 37oC и в атмосфере с 5% CO2. Когда клетки закрепятся в культуре и начинают размножаться и их концентрация становится приблизительно 106/мл, клетки и среду переносят во флакон для культуры тканей с площадью поверхности 80 см2 и разводят до 40 мл, используя DMEM/F12 (без сыворотки). Когда концентрация клеток снова достигнет 106/мл, их и среду переносят во флакон для культуры ткани с площадью поверхности 175 см2 и разбавляют далее до объема 100 мл, используя DMEM/F12. Когда клетки снова достигнут оптимально концентрации, их и среду переносят в роллер-флакон с площадью поверхности 850 см2 и разводят до конечного объема 500 мл. Когда в этом роллер-флаконе достигается оптимальная концентрация клеток, его содержимое делят на 3 части и переносят в новые роллер-флаконы, используя ту же среду, что и прежде. Этот процесс деления содержимого роллер-флаконов продолжают до тех пор, пока не получат количество флаконов, достаточное, чтобы иметь нужное количество клеток для высевания в ферментер Verax System 200.
Verax System 200
Ферментер Verax System 200 представляет собой замкнутую систему культивирования клеток периодического действия, в которой клетки культивируют на утяжеленных нержавеющей сталью микросферах (плотность 1,6 г/мл), состоящих из коровьего коллагена продольно-поперечного типа I. Микросферы загружают в вертикальную прозрачную стеклянную трубку, через которую прокачивают культуральную среду /ту же, что упоминалась выше/, поступающую снизу. Вход в трубу оформляется таким образом, что микросферы будут создавать конфигурацию псевдоожиженного слоя, когда среду прокачивают с соответствующей скоростью. В ходе процесса постоянно добавляют свежую среду и удаляют кондиционированную среду со скоростью, определяемой ростом клеток, который контролируют по расходу глюкозы. Поддерживают температуру 37oC, поддерживают pH 7,1 и также регулируется соотношение кислород/азот.
После загрузки микросфер содержащей 1% фетальной коровьей сыворотки средой ферментер работает по крайней мере в течение трех суток без клеток, чтобы убедиться, что загрузка микросфер не загрязнила систему. В это время в ферментер подают свободную от белка среду, чтобы снизить прайминг-дозу фетальной коровьей сыворотки. Если все системы работают удовлетворительно, в реактор инокулируют клетки из роллер-флаконов.
Verax System 2000
Это оборудование используют тот же тип микросфер, что и System 200, и в ней приемы контроля и работы по существу такие же, как и в меньшей системе. Система 2000 приблизительно в 15 раз больше по сравнению с системой 200.
Контроль выхода антитела из натуральной культуры
Кондиционированную среду контролируют, каждый раз опорожняя сборник, на уровень человеческого имуноглобулина в супернатанте, используя анализ типа ELISA. Результаты подтверждают, используя метод ЖХВР с белком A.
Сбор клеток, выросших в культуральной среде, и формирование собранного фонда
Кондиционированную среду непрерывно выводят из Verax-оборудования в охлаждаемый сборник. Эту среду впоследствии выгружают /используя давление азота в системе Verax/ в передвижной сборник из нержавеющей стали для дальнейшей переработки.
Контрольная клеточная среда
Среда, которую обычно используют, представляет собой смесь 1:1 МЕМ Дульбекко H21 и Ham F12 /Mediatech/. Среду закупают в виде порошка, достаточного для приготовления 50 л конечной среды. Такой порошок среды из двух таких контейнеров вводят в емкость из нержавеющей стали, содержащую приблизительно 190 л воды. Порошок суспендируют при вращающейся мешалке до тех пор, пока он весь не растворится. Добавляют бикарбонат натрия, как рекомендует производитель, и устанавливают pH среды 7,4. Добавляют селенит натрия до конечной концентрации 17,3 мкг/л и доводят объем до 200 л, добавляя воду. Среду также снабжают ионами железа в форме нитрата железа с цитратом натрия до конечной концентрации Fe+++ 50 мкМ. Среду сразу же загружают в емкость для среды в Verax System 200 через встроенный фильтр для стерилизации. В среду не добавляют белок. Никогда не используют никаких антибиотиков.
Очистка моноклонального антитела.
Описание методологии сбора и очистки конечного продукта
Моноклональное антитело получают в клеточной культуре из линии клеток гибридом в отсутствии сыворотки. Это означает, что из конечного продукта необходимо удалить только компоненты клеточного материала. Так как не ожидается, что моноклональные антитела человека сами являются иммуногенными, становится весьма важным удаление всех потенциально иммуногенных компонентов.
Целью операций очистки, использующих аффинную хроматографию, является конечный продукт с чистотой более 99,9%. Все сильно зависит от биологической специфичности аффинной хроматографии. Каждая стадия процесса очистки /кратко показанного в табл.5/ обсуждается подробнее ниже.
Сбор клеток и удаление материала микрочастиц из кондиционированой среды.
Даже хотя большая часть клеток удерживается микросферами, порядочное число частиц присутствует в собираемом супернатанте. Чтобы избежать большого загрязнения среды клеточными компонентами, супернатант фильтруют через поливинилиден-дифторидный фильтр, 0,65 мкм, Prostack® /Миллипор/, сразу же после удаления из сборника установки Verax. Этот тип фильтровального узла работает в режиме тангенциального потока, что позволяет фильтровать большое количество материала с микрочастицами без забивания фильтра. Очищенную среду собирают в емкости из нержавеющей стали с охлаждением.
Концентрация кондиционированной среды
Кондиционированную среду концентрируют, используя поставляемую Миллипором полисульфоновую спирально намотанную мембрану номиналом 30000 дальтон. После концентрирования устанавливают pH 7,9, используя 1M уксусную кислоту. Материал для стерилизации фильтруют через Sartobran-PH, 0,8/0,2 мкм /Сарториус/ /компонент 0,8 микрометровый представляет собой полиэфир, 0,2-мкм компонент представляет собой ацетат целлюлозы/, прежде чем поместить его на хранение при 4oC. Материал подвергают микрофильтрации /0,22 мкм, Millipore/ и разливают в сосуды из полипропилена.
Хроматография с белком A
Стадия чрезвычайно глубокой очистки использует высокую аффиность антитела IgGl человека к белку A Staphylococcus aureus.
Белок A закупают уже связанный ковалентно амидной связью с агарозой. После набивки колонки гелем ее и ее содержимое и соединительные трубки дезинфицируют путем обработки 70% этанолом в воде в течение 24 часов. Колонку затем уравновешивают ЭФР, pH 7,0.
Осуществление аффинно-хроматографического разделения на колонке с белком A включает следующие последовательные стадии
A/ Загрузка. Концентрированную кондиционированную среду загружают в колонку с помощью насоса. Вытекающий из колонки поток собирают и контролируют на присутствие антитела путем ELISA с иммуноглобулином человека. Колонку загружают до такой степени, чтобы через колонку проходило измеряемое количество содержащей антитело жидкости. Перегруженную фракцию извлекают отдельно и возвращают в цикл, если она содержит более 20 мг/мл антитела.
B/ Промывание. Чтобы удалить несвязанные материалы, колонку обильно промывают забуференным фосфатом физиологическим раствором, pH 7, с хлоридом натрия, добавленным до конечной концентрации 0,5М. После такой промывки следует стадия второй промывки с применением буфера из 0,02 M цитрата натрия, pH 5,6, содержащего 0,5 M хлорида натрия. Такая промывка высвобождает небольшие количества антитела человека.
C/ Элюирование. Связанные моноклональные антитела элюируют из колонки, используя буфер, составленный из 0,02 M цитрата натрия, pH 3,0, содержащего 0,5 M хлорида натрия. Элюированный материал непрерывно разбавляют в объеме 1 M трис-HCl, pH 8,0, чтобы быстро восстановить условия, близкие к нейтральному состоянию.
Очистку с белком A осуществляют в замкнутой системе с использованием системы очистки Waters 650 Protein Purification System.
Концентрирование элюата из колонки с белком A
Чтобы сделать следующую стадию очистки более эффективной и удобной, элюат из колонки с белком A концентрируют по крайней мере до 5 мг/мл антитела. Концентрат стерилизуют, фильтруя его через фильтр, 0,2 мкм, и стерильный концентрат хранят при 4oC до тех пор, пока не соберут количество материала, достаточное для следующей стадии очистки.
Разделение по размеру гель-хроматографией на сефакриле S-300 высокого разрешения
Препарат антитела пропускают через гель-сефакрил S-300 высокого разрешения /Pharmacia/, которым заполняют колонку Pharmacia BP113/120 с объемом слоя приблизительно 10 л. Колонку заполняют в Lactated Ringer's Irrigation USP /Travenol Laboratories/. Элюирование колонки контролируют системой Waters 650 Protein Purification Systems.
Целью этой стадии является главным образом не дополнительная очистка, а буферная перестройка. После элюирования из колонки с белком A антитела находятся в комплексном гипертоническом буфере, состоящем из цитрата натрия, хлорида натрия и трис-HCl. Такая буферная смесь не может непосредственно использоваться как носитель для внутривенной инъекции. После этой стадии буфер является подходящим как для внутривенной инъекции, так и для более длительного хранения при пониженной температуре.
Удаление ДНК клетки-хозяина путем пропускания через ионообменнную колонку
Даже после хроматографии с белком A, которая удаляет основную массу ДНК, присутствующей в концентрированном супернатанте, и после обработки с сефакрилом S-300, который удаляет молекулы ДНК, которые либо значительно больше, либо значительно меньше молекул моноклонального антитела, в препарате антитела присутствует небольшое, но заметное количество ДНК. Для удаления этого загрязнения избирается стадия ионного обмена на сильной анионообменной смоле - Q-сефарозе /Pharmacia Inc./. При pH лактированного раствора Рингера белки антитела имеют положительный заряд и отталкиваются анионообменной смолой. Нуклеиновые кислоты, однако, имеют отрицательный заряд при этом pH, и будут связываться в колонке.
Колонку заполняют в соответствии с предложениями изготовителей. После декантирования 20% раствором этанола гель вносят в лактированный раствора Рингера - 100 мл геля суспендируют в 200 мл раствора. Смесь выливают в колонку Pharmacia K50/30 и, когда гель сам уложится до постоянного объема, его дезинфицируют объемом 0,5 N гидроксида натрия, равным объему колонки, затем объемом, равным трем объемам колонки, ЗФР Дульбекко, затем объемом, равным 5 объемам колонки, лактированного раствора Рингера. Непосредственно перед употреблением колонку промывают дополнительным, равным 5 объемам колонки, объемом лактированного раствора Рингера. Затем через колонку пропускают образец и прошедший через колонку материал собирают в стерильную емкость.
Пример 8
Молекулярный анализ PE1-1, ZM1-1, ZM1-2 и MD3-4
Выделяют и секвенируют тяжелую вариабельную цепь /VH/ антител PE1-1, ZM1-1, ZM1-2 и MD3-4. Полную РНК экстрагируют из 107 гибридомных клеток каждой клеточной линии, используя методики, описанные в Sanz et al., 1989, J. Immunol, 142: 883, и эта работа включена в качестве ссылки. Одноцепочечную ДНК синтезируют, используя в качестве фермента AMV-обратную транскриптазу и олиго-dT в качестве праймера. Количество синтезированной одноцепочечной кДНК определяют путем измерения включений 32p-dCTP /дезоксицитидин-5'-трифосфата/.
Цепные полимеразные реакции /PCR/ осуществляют по существу так, как рекомендует изготовитель /Perkin Elmer Cetus, Norwalk, Connecticut/. 1 мкг ДНК добавляют к 200 мкм раствору каждого из dATP, dCTP, dGTP и dTTP со 100 пмолями каждого из праймеров и 5 единицами Taq ДНК-полимеразы. Циклы PCR следующие: денатурирование при 98oC в течение 3 минут, ренатурация при 55oC в течение 2 минут и удлинение при 72oC в течение 3 минут при контроле в ДНК-термоячейке /Perkin Elmer Cetus/.
Амплифицированную ДНК отбирают по размеру на 1,0% геле низкоплавкой агарозы, лигируют в EcoRV-сайт фагемидного вектора BLUES-CRIPT и трансформируют в CaCl2-компетентную бактерию BSJ 72. Для секвенирования выделяют одноцепочечную ДНК из каждого позитивного клона после суперинфекции с M13K07, как описано у Sanz et al., см. выше. Секвенирование выполняют методом терминации дидезоксицепи, как описано у Sanger et al., 1980, J. Mol. Biol. 143:161, за исключением йодифицированной T7 ДНК-полимеразы /секвеназы/, для которой используют метод, описанный Tabor et al., 1987, PNAS (USA) 84:4767. Результаты приводятся в таблицах 6, 7, 8 и 9.
Пример 9
Следуя процедурам примера 8, выделяют и секвенируют легкую вариабельную цепь (VL) антител PE1-1, ZM1-1, ZM1-2 и MD3-4. Результаты приводятся в табл. 10, 11, 12 и13.







Изобретение касается моноклонального антитела, эффективного для диагностики и лечения гепатита В. Моноклональное антитело получают из линии клеток, полученных путем слияния ксеногенной гибридомы, обозначенной SPAZ-4, с клетками крови пациента, иммунизированного вакциной гепатита В. Моноклональные антитела противогепатитного действия применяют для защиты пациентов с угнетенным иммунитетом и для диагностики гепатита В. 3 с. и 2 з.п.ф-лы, 13 табл., 2 ил.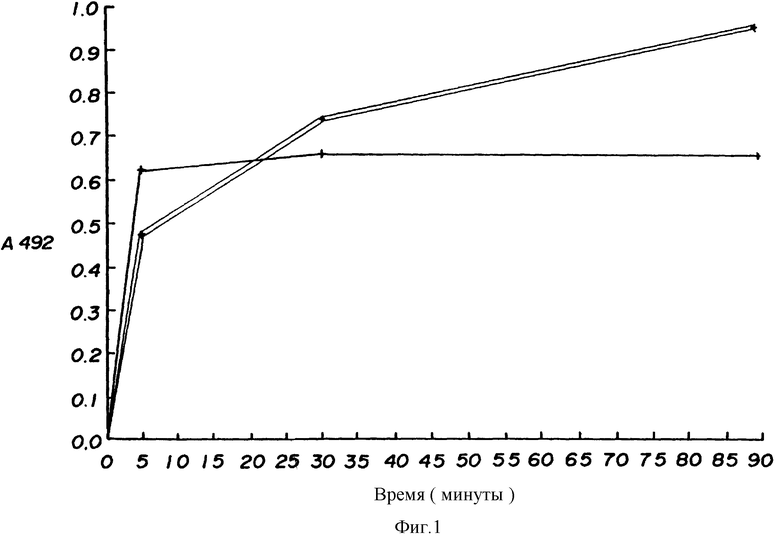
| RU 94038719, 30.05.96 | |||
| RU 94038730, 30.05.96 | |||
| RU 94039347, 30.05.96 | |||
| ЯДЕРНЫЙ МАГНИТОМЕТР СВОБОДНОЙ ПРЕЦЕССИИПРОТОНОВ | 0 |
|
SU179483A1 |
| РСТ 9105059, 18.04.91. | |||

