Настоящее изобретение относится к области фармацевтической и органической химии, в частности, к новым соединениям и фармацевтическим композициям, которые могут быть использованы для увеличения экспрессии инсулина из островковых В-клеток поджелудочной железы млекопитающего и для лечения ювенильного сахарного диабета у млекопитающих.
Внутренняя секреция островков Лангерганса находится под комплексным контролем, осуществляемым не только посредством метаболитов крови /глюкозы, аминокислот, катехоламинов и т.п./, но и также посредством локальной паракринной регуляции. Главные гормоны островков Лангерганса (глюкагон, инсулин и соматотропин) взаимодействуют друг с другом в их специфических клетках (А-, B- и D-клетках, соответственно), модулируя секреторный ответ, опосредованный вышеуказанными метаболитами. Хотя секреция инсулина регулируется, главным образом, уровнями глюкозы в крови, однако, глюкозо-опосредованная секреция инсулина ингибируется соматостатином. Кроме того, помимо предполагаемой межостровковой паракринной регуляции секреции инсулина имеются основания предположить о существовании инсулинотропных факторов в кишечнике. Это предположение было сделано на основании наблюдений, которые показали, что глюкоза, введенная перорально, является более сильным стимулятором секреции инсулина, чем то же самое количество глюкозы, введенное внутривенно.
Гормон человека, глюкагон, продуцируемый А-клетками поджелудочной железы, представляет собой пептид, состоящий из 29 аминокислот. Этот гормон принадлежит к мультигенному семейству структурно родственных пептидов, таких как секретин; пептид, угнетающий секрецию желудка; вазоактивный интестинальный пептид; и глицентин. Указанные пептиды по-разному регулируют углеводный обмен, гастроинтестинальную подвижность, и секреторное процессирование. Однако главной из известных функций глюкагона поджелудочной железы является стимулирование гликогенолиза и глюконеогенеза в печени, что приводит к повышению уровней сахара в крови. С этой точки зрения действие глюкагона является противоположным регулирующему действию инсулина и стимулирует гипергликемию, являющуюся симптомом сахарного диабета (Lund, P.K. и др. Proc.Natl.Acad.Acad.Sci.U.S.A., 79: 345-349 (1982)).
Было установлено, что глюкагон может связываться со специфическими рецепторами, расположенными на поверхности клеток, продуцирующих инсулин. Связываясь с этими рецепторами, глюкагон стимулирует быстрый синтез сАМР этими клетками. Было также установлено, что сАМР, в свою очередь, стимулирует экспрессию инсулина [Korman, L.Y. и др. Diabetes, 34: 717-722 (1985)], а инсулин способствует подавлению синтеза глюкагона [Ganong, W.F., Review of Medical Physiology, Lange Publication, Los Altos, California, p.273 (1979)]. Таким образом, экспрессия глюкагона строго регулируется инсулином и в конечном счете уровнем глюкозы в сыворотке.
Первоначальная трансляция гена глюкагона происходит из предшественника, имеющего 360 пар оснований, и в результате этой трансляции синтезируется полипептид препроглюкагона (Lund и др., Proc.Natl.Acad.Sci.U.S.A., 79: 345-349 (1982)). Затем этот полипептид подвергается посттрансляционному процессингу с образованием проглюкагона. После этого, как было показано Patzelt C. и др. (Nature, 282: 260-266 (1979)), образовавшийся проглюкагон расщепляется на глюкагон и второй полипептид. В своих последующих работах Lund P.K. и др., Lopez L. C. и др., Proc.Natl. Acad.Sci.U.S.A., 80: 5485-5489 (1983), и Bell G. L. и др. Nature 302: 716-718 (1983) продемонстрировали, что молекула проглюкагона расщепляется на участке, расположенном непосредственно после дипептидных остатков лизина-аргинина. Исследования проглюкагона, продуцируемого кошачьим сомиком (Ictalurus punctata), показали, что глюкагон, полученный от этого животного, также был протеолитически отщеплен на участке, расположенном непосредственно после дипептидных лизин-аргининовых остатков [Andrews P. C. и др., J.Biol.Chem., 260: 3910-3914 (1985), Lopez.C. и др., Proc. Natl.Acad.Sci.U.S.A., 80: 5485-5489 (1983)]. Bell G.I. и др. (см.выше) обнаружили, что проглюкагон млекопитающего расщепляется у лизин-аргининовых или аргинин-аргининовых дипептидов, и показали, что молекула проглюкагона состоит из трех дискретных и высокогомологичных пептидных молекул, которым были даны следующие названия: глюкагон, глюкагоноподобный пептид 1 (GLP-1) и глюкагоноподобный пептид 2 (GLP-2). На основании этого Lopez и др. был сделан вывод, что глюкагоноподобный пептид 1 имеет длину в 37 аминокислотных остатков, а глюкагоноподобный пептид 2 имеет длину в 34 аминокислотных остатков. Аналогичные исследования структуры препроглюкагона крыс выявили сходную картину протеолитического расщепления, происходящего на участке между смежными дипептидными лизин-аргининовыми аргинин-аргининовыми остатками, и приводящего к образованию глюкагона, GLP-1 и GLP-2 (Heinrich G. et al., Endocrinol. , 115: 2176-2181 (1984)). Было установлено, что у человека, крысы, быка и хомячка последовательности GLP-1 являются идентичными (Ghiglione M., et al., Diabetologia, 27: 599-600 (1984)).
Вывод относительно размера последовательности GLP-1, высказанный Lopez и др. , был подтвержден работой Uttenthal L.O., и др. (J.Clin.Endocrinol. Metabol., 61: 472-479, (1985)), Uttenthal и др. исследовали молекулярные формы GLP-1, которые присутствуют в поджелудочной железе человека. Результаты этого исследования показали, что GLP-1 и GLP-2, присутствующие в поджелудочной железе, представляют собой пептиды, состоящие, соответственно, из 37 и 34 аминокислот.
В более ранних исследованиях, исходя из аналогии между GLP-1 и глюкагоном, было сделано предположение, что GLP-1 также может обладать биологической активностью. И хотя некоторыми исследователями было установлено, что GLP-1 индуцируется клетками головного мозга крысы с последующим синтезом сАМР (Hoosein N.M. et al., Febs. Lett.178: 83-86, (1984)), однако, другим исследователям не удалось выявить какую-либо физиологическую роль GLP-1 (Lopez L. C., et al.). Эта безуспешная попытка идентифицировать какие-либо физиологические функции GLP-1 поставила исследователей перед решением важной задачи, а именно является ли на самом деле GLP-1 гормоном, и имеется ли действительно родство между глюкагоном и GLP-1 либо предполагаемое родство является лишь артефактом.
Кроме того, были также получены и исследованы варианты GLP-1 (7-37) и их аналоги. Такими вариантами и аналогами являются, например, Gln9-GlP-1 (7-37), D-Gln9-GLP-1 (7-37), ацетил-Lys9-GLP-1 (7-37), Thr16-Lys18-GLP-1 (7-37), Lys18-GLP-1 (7-37) и т.п., а также их производные, включая кислые аддитивные соли, карбоксилатные соли, низшие алкиловые сложные эфиры, и амиды (см., например, WO 91/11457). Известно, что, в основном, вышеуказанные формы стимулируют секрецию (инсулинотропное действие) и образование сАМР (см., например, Mojsov S., Int. J. Peptide Protein Reseach, 40, 333-343 (1992)).
При этом важно отметить, что многими исследователями была проиллюстрирована связь между лабораторными экспериментами и инсулинотропными ответами млекопитающих, в частности человека, на введение GLP-1, а конкретно GLP-1 (7-36) NH2 и GLP-1 (7-37) (см., например, Nauck M.A. et al., Diabetologia, 36:7 41-744 (1993); Gutniak M., et al., New England J. of Medicine, 326(20): 1316-1322 (1992); Nauck M.A., et al., J. Clin. Invest., 91: 301-307 (1993); и Thorens B., et al., Diabetes, 42: 1219-1225 (1993)).
В частности, было показано, что главными нарушениями, вызывающими гипергликемию при заболевании ювенильным диабетом, являются уменьшение секреции эндогенного инсулина и резистентность к действию инсулина в мышцах и печени (Galloway J.S., Diabetes Care, 13: 1209-1239 (1990)). Последнее из указанных нарушений приводит к чрезмерному продуцированию глюкозы в печени. Так, например, у здорового индивидуума, высвобождение глюкозы происходит со скоростью примерно 2 мг/кг/мин, тогда как у пациентов с ювенильным диабетом количество высвобождаемой глюкозы обычно превышает 2,5 мг/кг/мин, что приводит к избыточному продуцированию глюкозы, составляющему, по крайней мере, 70 грамм глюкозы за 24 часа. Факт существования исключительно высокой степени корреляции между продуцированием глюкозы в печени, содержанием глюкозы в крови натощак и полным метаболическим контролем, как показали измерения гликогемоглобина (Galloway J.A., см.выше; и Galloway J.A. и др., Clin. Therap. , 12: 460-472 (1990)), со всей очевидностью свидетельствует о том, что регулирование уровня глюкозы в крови натощак является необходимым условием для достижения полной нормализации обмена веществ, которая является достаточной для предупреждения осложнений гипергликемии. Однако, если учитывать тот факт, что существующие формы инсулина редко нормализуют продуцирование глюкозы в печени, не стимулируя при этом значительной гиперинсулинемии и гипогликемии (Galloway J.A., и Galloway J.A. и др., см. выше), то необходимо разработать другие альтернативные способы регулирования уровней глюкозы в крови.
Внутривенные вливания GLP-1 (7-36)NH2 с продуцированием сывороточных концентраций, вдвое превышающих нормальную концентрацию, показали результаты, представленные в нижеприведенной таблице 1.
Однако стабильность GLP-1, а в частности стабильность GLP-1 как компонента фармацевтической композиции, предназначенной для введения млекопитающим, не является достаточно долговременной. И действительно, при хранении при температуре 4oC, уже через 11 месяцев после получения образцов были обнаружены побочные продукты GLP-1 (7-37) (Mojsov S., см. выше). Поэтому совершенно очевидно, что необходимо получить более стабильное соединение GLP-1, которое можно было бы без всякого риска вводить млекопитающему, нуждающемуся в таком лечении.
Кроме того, биологическое время полужизни молекул GLP-1, а в частности молекул, подвергающихся воздействию дипептидил-пептидазы IV (DPP IV), является очень непродолжительным. Например, биологическое время полужизни GLP-1 (7-37) составляет лишь 3-5 минут, и, кроме того, после его парентерального введения млекопитающему он быстро абсорбируются. Поэтому желательно также получить такое соединение GPL-1, абсорбция которого происходит через более длительный промежуток времени после его введения млекопитающему.
В соответствии с вышеуказанным необходимо отметить, что настоящее изобретение позволяет разрешить проблемы, связанные с нестабильностью нативных молекул GLP-1, присутствующих в сыворотке, а также с их непродолжительным временем полужизни. Кроме того, соединения настоящего изобретения после их парентерального введения, обладают замедленной абсорбцией, а значит, и более продолжительным биологическим временем полужизни. Настоящее изобретение также относится к фармацевтическим композициям, содержащим соединения настоящего изобретения, а также к способам использования этих соединений.
Настоящее изобретение относится к комплексу, состоящему из катиона двухвалентного металла, ассоциированного и способного к совместной преципитации с соединением формулы
R1-X-Glu-Gly-Thr-Phe-Thr-Ser-Asp-Val-Ser-Ser-Tyr-Leu-Y-Gly-Gln-Ala-Ala-Lys-Z-Phe-Ile-Ala-Trp-Leu-Val-Lys-Gly-Arg-R2
где R1 выбирают из группы, включающей в себя L-гистидин, D-гистидин, дезамино-гистидин, 2-амино-гистидин, β-гидрокси-гистидин, гомогистидин, альфа-фторометил-гистидин, и альфа-метил-гистидин;
X выбирают из группы, включающей в себя Ala, Gly, Val, Thr, Ile, и альфа-метил-Ala;
Y выбирают из группы, включающей в себя Glu, Gln, Ala, Thr, Ser, и Gly;
Z выбирают из группы, включающей в себя Glu, Gln, Ala, Thr, Ser, и Gly;
R2 выбирают из группы, включающей в себя NH2, и Gly-OH, при условии, что указанное соединение имеет изоэлектрическую точку в диапазоне от около 6,0 до около 9,0, и при условии, что если R1 является His, X является Ala, Y является Glu, а Z является Glu, то R2 должен представлять собой NH2.
Настоящее изобретение также относится к фармацевтической композиции, содержащей соединение настоящего изобретения в комбинации с одним или несколькими фармацевтически приемлемыми носителями, разбавителями или наполнителями.
Кроме того, настоящее изобретение относится к способу увеличения экспрессии инсулина, заключающемуся в обеспечении островковых B-клеток поджелудочной железы эффективным количеством соединения настоящего изобретения, а также к способу лечения ювенильного диабета, заключающемуся во введении млекопитающему, нуждающемуся в таком лечении, эффективного количества соединения настоящего изобретения.
В одном из вариантов своего осуществления настоящее изобретение относится к комплексу, состоящему из молекулы GLP-1, имеющей изоэлектрическую точку в диапазоне от около 6,0 до около 9,0, и из катиона двухвалентного металла.
Используемый в настоящем описании термин "молекула GLP-1" относится к натуральным глюкагоноподобным пептидам 1 (GLP-1)(7-36)NH2 и GLP-1 (7-37), к натуральным или ненатуральным функциональным аналогам и производным этих пептидов, и к их солям. Аминокислотная последовательность пептида GLP-1 (7-36)NH2 хорошо известна специалистам, однако для удобства читателя эта последовательность приводится ниже:
His7-Ala-Glu-Gly10-Thr-Phe-Thr-Ser-Asp15-Val-Ser-Ser-Tyr-Leu20-Glu-Gly-Gln-Ala-Ala25-Lys-Glu-Phe-Ile-Ala30- Trp-Leu-Val-Lys-Gly35-Arg-NH2.
В случае GLP-1 (7-37) карбокси-концевая амидная функциональная группа Arg36 заменена на Gly в 37-положении молекулы GLP-1 (7-36)NH2.
Кроме того, существование и получение многочисленных защищенных, незащищенных и частично защищенных натуральных и ненатуральных функциональных аналогов и производных молекул GLP-1 (7-36)NH2 и GLP-1 (7-37) раскрываются в литературе (см., например, патенты США NN 5120712 и 511866, которые вводятся в настоящее описание посредством ссылки; и Orskov C., и др., J.Biol. Chem., 264(22): 12826-12829 (1989), и WO 91/11457 (Buckley, D.I., и др., опубл. 8 авг. 1991)).
Известно, что аминокислотные остатки могут существовать в защищенной форме, в которой обе амино- и карбоксигруппы имеют соответствующие защитные группы; в частичнозащищенной группе, в которой либо амино-, либо карбоксигруппа имеет соответствующую защитную группу; или в незащищенной форме, в которой ни амино-, ни карбоксигруппа не имеет соответствующей защитной группы. В литературе описано множество реакций, используемых для образования и удаления защитных групп: см., например, такие работы, как "Protective Groups in Organic Chemistry", Plenum Press (London and New York, 1973); Green, T.H. , "Protective Groups in Organic Synthesis", Wiley (New Vork, 1981); и "The Peptides", Vol. 1, Schroder & Lubke, Academic Press (London and New York, 1965).
Характерными аминозащитными группами являются, например, формил, ацетил, изопропил, бутоксикарбонил, флуоренилметоксикарбонил, карбобензилокси и т.п.
Характерными карбоксизащитными группами являются, например, бензиловый сложный эфир, метиловый сложный эфир, этиловый сложный эфир, т-бутиловый сложный эфир, п-нитрофениловый сложный эфир и т.п.
Помимо защищенных форм, в которых амино- и карбоксигруппы имеют соответствующие защитные группы, термин "защищенный" также относится к таким молекулам GLP-1, в которых нейтрализуется или ингибируется активность дипептидил-пептидазы IV (см., например, Mentlein, R., и др., Eur.J. Biochem., 214: 829-835 (1993)). Помимо GLP-1 (7-36)NH2 предпочтительными являются молекулы, которые защищены от активности DPP IV, а более предпочтительными являются GLy8-GLP-1(7-36)NH2, Val8-GLP-1(7-37)OH, α-метил-Ala8-GLP-1(7-36)NH2 и Gly8-Gln21-GLP-1(7-37)OH.
Производными природных молекул GLP-1 являются такие пептиды, которые могут быть получены путем фрагментации нативной последовательности или путем синтеза, проводимого на основании имеющихся данных о природной аминокислотной последовательности из генетического материала (ДНК или РНК), кодирующего эту последовательность. Термин "производные" также относится к химическим модификациям натуральной или ненатуральной молекулы GLP-1. Способы получения таких производных хорошо известны специалистам в области органической и пептидной химии (см., например, WO 91/11457, см. выше).
К молекулам GLP-1 настоящего изобретения также относятся аналоги GLP-1 (7-36)NH2 и GLP-1(7-37), где одна или несколько аминокислот, не присутствующих в исходной последовательности, являются добавленными; либо одна или несколько аминокислот являются делетированными; а также производные указанных аналогов. В частности, в вышеуказанных молекулах, предпочтительно, если R1 представляет собой His и дезамино-гистидин, при условии, что изоэлектрическая точка всей молекулы находится в диапазоне от около 6 до 9. Кроме того, предпочтительно, если в положении "X" присутствуют Ala, Gly и Val, при условии, что изоэлектрическая точка всей молекулы находится в диапазоне от около 6 до 9. Аналогичным образом, предпочтительно, если в положение "Y" присутствуют Glu и Gln, при условии, что изоэлектрическая точка всей молекулы находится в диапазоне от около 6 до 9. Также предпочтительно, если в положении "Z" присутствуют Glu и Gln, при условии, что изоэлектрическая точка всей молекулы находится в диапазоне от около 6 до 9. И, наконец, предпочтительно, если R2 представляет собой Gly-OH, при условии, что изоэлектрическая точка всей молекулы находится в диапазоне от около 6 до 9.
Кроме того, настоящее изобретение включает в себя солевые формы молекулы GLP-1. GLP-1-молекулы настоящего изобретения могут иметь достаточно кислотные, достаточно основные, или кислотные и основные функциональные группы; и в соответствии с этим указанные молекулы могут вступать в реакцию с любыми из соответствующих неорганических оснований и неорганических и органических кислот с образованием соли. Для образования кислых аддитивных солей обычно используют неорганические кислоты, такие как соляная кислота, бромистоводородная кислота, иодистоводородная кислота, серная кислота, фосфорная кислота, и т. п.; и органические кислоты, такие, как п-толуолсульфоновая кислота, метансульфоновая кислота, щавелевая кислота, п-бромофенилсульфоновая кислота, угольная кислота, янтарная кислота, лимонная кислота, бензойная кислота, уксусная кислота и т.п. Примерами таких солей могут служить сульфат, пиросульфат, бисульфат, сульфит, бисульфит, фосфат, монобифосфат, дибифосфат, метафосфат, пирофосфат, хлорид, бромид, иодид, ацетат, пропионат, деканоат, каприлат, акрилат, формат, изобутират, капроат, гептаноат, пропионат, оксалат, малонат, сукцинат, суберат, себацат, фумарат, малеат, бутин-1,4-диоат, гексин-1,6-диоат, бензоат, хлоробензоат, метилбензоат, динитробензоат, гидроксибензоат, метоксибензоат, фталат, сульфонат, ксилолсульфонат, фенилацетат, фенилпропионат, фенилбутират, цитрат, лактат, гамма-гидроксибутират, гликолат, тартрат, метансульфонат, пропансульфонат, нафталин-1-сульфонат, нафталин-2-сульфонат, манделат и т.д. Предпочтительными являются кислые аддитивные соли, образованные минеральными кислотами, такими как хлористоводородная и бромистоводородная кислота, а предпочтительно хлористоводородная кислота.
Основными аддитивными солями являются соли, образованные неорганическими основаниями, такими как гидроксиды, карбонаты, и бикарбонаты аммония, щелочных металлов и щелочноземельных металлов и т.п. Основаниями, которые могут быть использованы для получения солей настоящего изобретения, являются гидроксид натрия, гидроксид калия, гидроксид аммония, карбонат калия и т.п. Соединения настоящего изобретения в виде соли являются особенно предпочтительными.
Если соединения настоящего изобретения используются в фармакотерапевтических целях, то само собой разумеется, что эти соединения также могут быть получены в виде соли, но эти соли должны быть фармацевтически приемлемыми.
Так, например, помимо вышеуказанных GLP-1-молекул настоящее изобретение также включает в себя молекулы GLP-1, которые обладают функциональной инсулинотропной активностью. Термин "инсулинотропная активность" означает способность данного соединения стимулировать или индуцировать стимуляцию синтеза или экспрессии инсулина.
Инсулинотропные свойства соединения могут быть определены путем обработки клеток животного данным соединением или путем инъецирования данного соединения животному с последующим наблюдением высвобождения иммунореактивного инсулина (IRI) в среду или в систему кровообращения животного, соответственно. Присутствие IRI может быть обнаружено с помощью радиоиммуноанализа, позволяющего осуществлять специфическую детекцию инсулина.
Хотя в целях настоящего изобретения может быть использован любой радиоиммунный анализ, способный обнаруживать присутствие IRI, однако, предпочтительно использовать модификацию аналитического метода Albano, J.D.M., et al. (Acta Endocrinol., 70: 487-509 (1972)). Эта модификация предусматривает применение фосфатно-альбуминового буфера с pH 7,4. Среду для инкубации приготавливают путем последовательного добавления 500 мкл фосфатного буфера, 50 мкл перфузируемого образца или стандартную норму крысиного инсулина в перфузате, 100 мкл антиинсулиновой антисыворотки (Wellcome Laboratories; разведение 1: 40000), и 100 мкл [125I]-инсулина; в результате чего получают полный объем 750 мкл в стеклянной пробирке (10 х 75 мм) для одноразового использования. После инкубации в течение 2-3 дней при 4oC свободный инсулин отделяют от инсулина, связанного с антителом, методом разделения с использованием активированного угля. Чувствительность анализа составляет 1-2 мкЕд/мл. Для оценки высвобождения IRI в культуральную среду клеток, выращиваемых в тканевой культуре, предпочтительно вводить в проинсулин радиоактивную метку. В этих целях может быть использована любая радиоактивная метка, способная к мечению полипептида; однако, для получения меченного проинсулина предпочтительно использовать 3H-лейцин. Мечение может быть осуществлено в течение любого периода времени, достаточного для образования обнаруживаемого меченного пула проинсулиновых молекул; но тем не менее инкубирование клеток в присутствии радиоактивной метки предпочтительно проводить в течение 60 минут.
Для оценки инсулинотропных свойств соединения могут быть использованы многие клеточные линии, способные экспрессировать инсулин, однако предпочтительно использовать клетки инсулиномы крысы, а особенно клетки инсулиномы крысы RIN-38. Эти клетки могут быть культивированы в любой подходящей среде, но тем не менее предпочтительной является DME-среда, содержащая 0,1% BSA и 25 мМ глюкозы.
Инсулинотропная способность соединения может быть также определена путем панкреатического вливания. In situ-выделенную перфузируемую поджелудочную железу крысы получали в соответствии с модифицированным методом Penhos J.C. et al. Diabetes, 18: 733-738 (1969). Белых крыс-самцов штамма Charles River, весом 350-600 г, находящихся на голодной диете, анестезируют путем внутрибрюшинной инъекции амиталнатрия (Amytal Sodium) (Eli Lilly & Co., 160 нг/кг). Почки, надпочечник, желудок и кровеносные сосуды нижней части толстой кишки лигируют. Весь кишечник удаляют за исключением примерно 4 см двенадцатиперстной кишки, нисходящей ободочной и прямой кишки. Поэтому лишь небольшая часть кишечника подвергается перфузии, что минимизирует возможное влияние кишечного материала на глюкагоноподобную реактивность. Перфузат представляет собой модифицированный по способу Кребсарингера бикарбонатный буфер, содержащий 4% декстрана Т70 и 0,2% альбумина бычьей сыворотки (фракция V), и барботированный 95% O2 и 5% CO2. При этом используют 4-канальный перистальтический насос, дающий непульсирующий поток (Buchler polystatic, Buchler Instruments Division, Nuclear-Chicago Corp.), а переключение с одного источника перфузата на другой осуществляют с помощью трехходового запорного крана. Перфузию, слежение и анализ осуществляют по методу Weir G. C. et. al. , J. Clin. Inestigat., 54: 1403-1412 (1974), который вводится в настоящее описание посредством ссылки.
При этом необходимо, чтобы GLP-1-молекулы настоящего изобретения имели в своем амино-конце гистидиновые функциональные группы, GLP-1-молекулы настоящего изобретения могут также иметь модифицированные гистидиновые функциональные группы вместо требуемых гистидиновых функциональных групп.
Термин "модифицированный гистидин" относится к гистидиновой функциональной группе, которая была изменена химическим или биологическим способом, либо к уже измененной гистидиновой функциональной группе, которая была синтезирована de novo, но которая при этом сохранила свою способность связываться с металлом.
Многие из таких модифицированных гистидиновых функциональных групп, а также способы их получения, являются известными, например, D-гистидин (WO 91/11457), дезамино-гистидин (WO 92/18531), 2-амино-гистидин (Levine-Pinto H. , et al. , Biochem. Biophys. Res. Commun., 103 (4): 1121-1130 (1981)), β-гидрокси-L-гистидин (Owa T. , et al. , Chemistry Letters pp. 1873-1874 (1988), L-гомогистидин (Altman J., et al., Synthetic Commun., 19 (11&12): 2069-2076 (1989)), α-фторометил-гистидин (патент США N 4347374), и α- метилгистидин (O'Donnell, M.J., Synthetic Commun., 19 (78): 1157-1165 (1989)).
Кроме того, необходимо, чтобы GLP-1-молекулы настоящего изобретения имели изоэлектрическую точку в диапазоне от около 6,0 до около 9,0. В литературе описаны многие молекулы GLP-1, имеющие изоэлектрическую точку в указанном диапазоне, например:
GLP-1 (7-46)NH2
Gly8-GLP-1 (7-36)NH2
Gln9-GLP-1 (7-37)
D-GLn9-GLP-1 (7-37)
ацетил-Lys9-GLP-1 (7-37)
Thr9-GLP-1 (7-37)
D-Thr9-GLP-1 (7-37)
Asn9-GLP-1 (7-37)
D-Asn9-GLP-1 (7-37)
Ser22-Arg23-Arg24-Gln26-GLP-1 (7-37)
Thr16-Lys18-GLP-1 (7-37)
Lys18-GLP-1 (7-37)
Arg23-GLP-1 (7-37)
Arg24-GLP-1 (7-37), и т. п. (например, WO 91/11457, см. выше). Кроме того, если GLP-1-молекулы настоящего изобретения вместо гистидиновых функциональных групп имеют вышеуказанные модифицированные гистидиновые функциональные группы, то их изоэлектрическая точка должна находиться в вышеуказанном диапазоне. Способы вычисления или экспериментального определения изоэлектрической точки других молекул GLP-1 хорошо известны специалистам.
Способы получения GLP-1-молекул настоящего изобретения также хорошо известны любому специалисту в области пептидной химии.
В одном из таких способов молекулы GLP-1 получают в соответствии с хорошо известными процедурами твердофазного пептидного синтеза, описанного Merrifield J. M., Chem. Soc., 85: 2149 (1962), и Stewart & Young, Solid Phase Peptide Synthesis, pp. 24-66, Freeman (Сан-Франциско, 1969). Однако отдельные фрагменты проглюкагонового полипептида или GLP-1 (1-37) могут быть также получены путем фрагментации нативной аминокислотной последовательности с использованием, например, протеолитического фермента. Кроме того, нужные фрагменты проглюкагонового пептида или GLP-1 (1-37) могут быть получены с использованием техники рекомбинантных ДНК, описанной Maniatis T., et al., Molecular Biology: A Laboratory Manual. CSH (Cold Spring Harbor, 1982).
Более того, пользуясь достижениями современной молекулярной биологии, каждый специалист может использовать и другие средства для получения соединений настоящего изобретения. И хотя, как указывалось выше, для получения соединений настоящего изобретения могут быть использованы методы твердофазного пептидного синтеза или рекомбинантные методы, однако, благодаря возможности получить более высокие выходы, рекомбинантные методы могут оказаться предпочтительными. Рекомбинантный метод продуцирования включает в себя следующие стадии:
a) выделение натуральной ДНК-последовательности, кодирующей GLP-1-молекулу, или конструирование синтетической или полусинтетической ДНК-последовательности, кодирующей GLP-1-молекулу;
b) введение кодирующей последовательности в экспрессирующий вектор, подходящий для экспрессии белков либо в изолированном виде, либо в виде гибридных белков;
c) трансформация соответствующей эукариотической или прокариотической хозяйской клетки с использованием экспрессирующего вектора;
d) культивирование трансформированной хозяйской клетки в условиях, благоприятствующих экспрессии молекулы GLP-1; и
e) выделение и очистка рекомбинантно продуцированной молекулы GLP-1.
Как было указано выше, кодирующие последовательности могут быть полностью синтезированы либо они могут быть получены в результате модификации более крупной нативной глюкагон-кодирующей ДНК. ДНК-последовательность, кодирующая препроглюкагон, приводится в работе Lund и др. (Proc. Natl. Acad. Sci. U.S. A., 79: 345-349 (1982)) и может быть использована в качестве исходного материала в полусинтетическом продуцировании соединений настоящего изобретения путем модификации нативной последовательности с получением нужных результатов.
Синтетические гены, которые посредством in vitro или in vivo-транскрипции и трансляции продуцируют молекулу GLP-1, могут быть сконструированы известными способами. Каждому специалисту известно, что вследствие естественного вырождения генетического кода может быть сконструировано вполне определенное число ДНК-последовательностей, кодирующих молекулы GLP-1.
Методика конструирования синтетического гена хорошо известна специалистам (см. , например, Brown и др. (1979) Methods in Enzymology, Academic Press, N. Y., Vol. 68, pp. 109-151). ДНК-последовательности, кодирующие молекулу GLP-1, могут быть сконструированы исходя из аминокислотных последовательностей, раскрытых в настоящем описании. После осуществления генетических построений сама последовательность может быть генерирована с использованием стандартной аппаратуры для синтеза ДНК, например, такой, как ДНК-синтезаторы модели 380A или 380B (PE-Applied Biosystems, Inc., 850 Lincoln Center Drive, Foster City, CA 94404).
Для осуществления экспрессии молекулы GLP-1 сконструированную синтетическую ДНК-последовательность вставляют в один из многих экспрессирующих векторов, подходящих для экспрессии рекомбинантной ДНК, с использованием соответствующих рестриктирующих эндонуклеаз (см., например, Maniatis и др. (1989) Molecular Cloning; A Laboratory Manual, Cold Springs Harbor Laboratory Press, N. Y. , Vol. 1-3). Сайты рестрикции эндонуклеазы конструируют на любом конце ДНК, кодирующей молекулу GLP-1, для облегчения ее выделения из известных амплифицирующих и экспрессирующих векторов, и интеграции в эти векторы. Выбор конкретных эндонуклеаз может быть осуществлен исходя из рестрикционной карты используемого исходного вектора экспрессии. Рестрикционные сайты выбирают так, чтобы кодирующая последовательность была правильно ориентирована по отношению к регуляторным последовательностям для осуществления правильного считывания с сохранением рамки и экспрессии нужного белка. Кодирующая последовательность должна быть вставлена с сохранением рамки считывания по отношению к промотору и сайту связывания с рибосомой экспрессирующего вектора, которые являются функциональными в хозяйской клетке, предназначенной для экспрессии белка.
Для эффективного осуществления транскрипции синтетического гена этот ген должен быть правильно присоединен к области промотора-оператора. Поэтому область промотора-оператора синтетического гена помещают в той же самой ориентации по отношению к инициирующему кодону ATG синтетического гена.
Для трансформации прокариотических и эукариотических клеток могут быть использованы различные экспрессирующие векторы, хорошо известные специалистам (см. , например, The Promega Biological Research Products Catalogue (1982) (Promega Corp., 2800 Woods Hollow Road, Madison, W1, 53711-5399); и The Stratagene Cloning Systems Catalogue (1992) (Stratagene Corp., 11011 North Torrey Pines Road, La Jolla, CA, 92037)). Кроме того, в патенте США N 4710473 описаны трансформирующие кольцевые ДНК-плазмидные векторы, используемые для экспрессии экзогенных генов в E. coli с высокими уровнями экспрессии. Указанные плазмиды могут быть использованы в качестве трансформирующих векторов при продуцировании рекомбинантных ДНК, и
(a) несут в себе способность к автономной репликации в клетке-хозяине;
(b) регулируют автономную репликацию плазмиды при температуре, при которой содержатся культуры клеток-хозяев;
(c) обеспечивают сохранение плазмиды в популяциях клеток-хозяев;
(d) регулируют синтез белкового продукта, что свидетельствует о сохранении плазмиды в популяции клеток-хозяев;
(e) обеспечивают распознавание серией рестриктирующих эндонуклеаз сайтов, уникальных для данной плазмиды; и
(f) обеспечивают терминацию транскрипции мРНК.
Указанные кольцевые ДНК-плазмиды могут быть использованы в качестве векторов при продуцировании рекомбинантных ДНК для обеспечения высоких уровней экспрессии экзогенных генов.
После того, как вектор для экспрессии молекулы GLP-1 будет сконструирован, его необходимо ввести в соответствующую клетку в целях создания рекомбинантной хозяйской клетки для экспрессии полипептида. Техника трансформации клетки с использованием рекомбинантных ДНК-векторов хорошо известна специалистам и описана в таких фундаментальных работах, как Maniatis и др. (см. выше). Клетки-хозяева могут быть сконструированы из эукариотических или прокариотических клеток.
В основном прокариотические клетки-хозяева продуцируют более высокие уровни белка и легче поддаются культивированию. Белки, продуцируемые в бактериальных системах с высоким уровнем экспрессии, обычно представляют собой агрегаты типа гранул или тел включения, которые содержат высокие уровни сверхпродуцированного белка. Указанные белковые агрегаты, как правило, должны быть солюбилизированы, денатурированы и подвергнуты повторной укладке с использованием известной техники (см., например, Kreuger et al., (1990), "Protein Folding" Gierachs & King, eds., pp. 136-142, American Association for the Advancement of Science Publication N 89-185, Washington, D.C., и патент США N 4923967).
После получения нужной молекулы GLP-1, имеющей изоэлекрическую точку в пределах от около 6,0 до около 9,0, изготавливают комплексы настоящего изобретения, содержащие указанную GLP-1-молекулу и катион двухвалентного металла, с использованием хорошо известной методики комплексообразования. Примерами указанных катионов двухвалентного металла могут служить Zn++, Mn++, Fe++, Co++, Cd++, Ni++ и т.п. Из этих металлов предпочтительным является Zn++.
Обычно нужную GLP-1-молекулу, имеющую требуемую изоэлектрическую точку, объединяют со смесью соответствующего буфера и соответствующей формой катиона металла.
Подходящими для этой цели являются такие буферы, которые способны поддерживать pH смеси в пределах от около 6,0 до около 9,0, и которые, кроме того, не оказывают неблагоприятного действия на реакцию. Предпочтительными являются буферы Гуда, в частности HEPES, а также Трис- и Трис-ацетатный буферы.
Подходящими формами катионов металлов являются любые формы катиона двухвалентного металла, которые способны образовывать комплексы с GLP-1-молекулой настоящего изобретения. Для этих целей предпочтительно использовать избыточное количество катионной соли двухвалентного металла, такой как хлорид цинка, в молярном соотношении, составляющем примерно до 50 молекул катиона двухвалентного металла на каждую молекулу GLP-1-субстрата.
Температура, используемая на этой стадии, должна быть достаточной для полного завершения реакции. Обычно такую реакцию проводят при комнатной температуре.
Продукт, полученный в результате этой реакции и представляющий собой кристаллическую или аморфную суспензию, выделяют и очищают с использованием стандартной техники.
Настоящее изобретение также относится к фармацевтическим композициям, содержащим соединение настоящего изобретения в комбинации с фармацевтически приемлемым носителем, разбавителем или наполнителем. Такие фармацевтические композиции могут быть получены в соответствии со стандартной техникой, обычно используемой в фармацевтической практике, и введены пациенту либо отдельно, либо в сочетании с другими терапевтическими средствами, предпочтительно парентерально. Особенно предпочтительным является внутримышечное и подкожное введение.
Суточная парентеральная доза, предпочтительно, разовая суточная доза составляет от около 1 пг/кг до около 1000 мкг/кг веса тела, хотя могут быть также использованы меньшие или более высокие дозы. Требуемая доза зависит от тяжести состояния пациента, а также от таких факторов, как рост пациента, его вес, пол, возраст и история болезни.
При изготовлении композиций настоящего изобретения активный ингредиент, содержащий, по крайней мере, одно соединение настоящего изобретения, обычно смешивают с наполнителем или разводят наполнителем. Если наполнитель используют в качестве разбавителя, то этот наполнитель может быть твердым, полутвердым или жидким материалом, действующим как растворитель, носитель, или среда для активного ингредиента.
При изготовлении композиции может оказаться необходимым измельчить активное соединение в целях получения нужных размеров частиц перед смешиванием этого соединения с другими ингредиентами. Если активное соединение является, в основном, нерастворимым, то обычно его измельчают до образования частиц размером приблизительно менее чем 200 меш. Если активное соединение является в основном водорастворимым, то это соединение размалывают с получением размеров частиц, имеющих однородный гранулометрический состав в данной композиции, и составляющих, например, около 40 меш.
Примерами подходящих наполнителей являются лактоза, декстроза, сахароза, трегалоза, сорбит и маннит. Композиции настоящего изобретения могут быть изготовлены в виде препаратов с быстрым, замедленным или пролонгированным высвобождением активного ингредиента после введения этих препаратов пациенту, и способы изготовления таких композиций хорошо известны специалистам.
Предпочтительно, если композиции настоящего изобретения изготавливают в виде разовой унифицированной лекарственной формы, которая обычно содержит от около 50 мкг до около 100 мг, в основном от около 1 мг до около 10 мг активного ингредиента. Термин "разовая унифицированная лекарственная форма" относится к физически дискретным единичным формам, используемым в качестве разовых лекарственных доз для введения человеку и другим млекопитающим; причем каждая из указанных единичных лекарственных форм содержит заранее определенное количество активного ингредиента, достаточное для продуцирования нужного терапевтического эффекта, в сочетании с подходящим фармацевтическим наполнителем.
Композиции, предназначенные для парентерального введения и содержащие соединение настоящего изобретения, предпочтительно смешивают с дистиллированной водой, а pH доводят до значения, составляющего от около 6,0 до около 9,0.
Для регулирования продолжительности высвобождения могут быть использованы дополнительные методы фармацевтической практики. Препараты с регулируемым высвобождением могут быть получены с использованием полимеров для образования комплексов с соединениями настоящего изобретения или для абсорбции соединений настоящего изобретения. Регулируемая доставка активного ингредиента к нужным органам или тканям может быть обеспечена посредством выбора соответствующих макромолекул (например, сложных полиэфиров, полиаминокислот, поливинилпирролидона, этиленвинилацетата, метилцеллюлозы, карбоксиметилцеллюлозы и протаминсульфата), концентрации этих макромолекул, а также методов включения.
Другим возможным методом получения препаратов с пролонгированным высвобождением является введение соединения настоящего изобретения в полимерные частицы таких материалов, как сложные полиэфиры, полиаминокислоты, гидрогели, сополимеры поли молочной кислоты) и сополимеры этилена и винилацетата.
Альтернативно, вместо введения в указанные полимерные частицы, соединения настоящего изобретения могут быть введены в микрокапсулы, полученные, например, с использованием техники консервации или межфазной полимеризации, а именно, в гидроксиметилцеллюлозные или желатиновые капсулы, соответственно; либо соединения настоящего изобретения могут быть введены в виде коллоидальных систем доставки лекарственного средства, например, таких как липосомы, альбуминовые микросферы, микроэмульсии, наночастицы, и нанокапсулы; или в виде макроэмульсий. Техника изготовления таких препаратов описана в Remington's Pharmaceutical Sciences (1980).
Соединения настоящего изобретения обладают инсулинотропной активностью. Поэтому в другом варианте своего осуществления настоящее изобретение относится к способу увеличения экспрессии инсулина, заключающемуся в том, что B-клетки панкреатических островков млекопитающего обрабатывают эффективным количеством соединения настоящего изобретения.
Аналогично настоящее изобретение относится к способу лечения ювенильного сахарного диабета у млекопитающих, предпочтительно у человека, заключающемуся в том, что млекопитающему, нуждающемуся в таком лечении, вводят эффективное количество соединения или композиции настоящего изобретения.
Ниже, в целях иллюстрации настоящего изобретения, приводятся соответствующие примеры. Однако эти примеры не должны рассматриваться как некое ограничение объема изобретения.
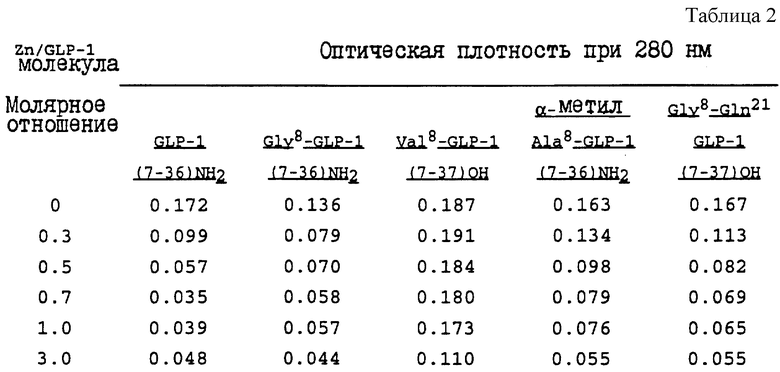
Пример 1
Отдельные аликвоты 5 различных GLP-1-молекул получали хорошо известным способом твердофазного пептидного синтеза, а затем лиофилизовали в небольших сосудах. К этим аликвотам добавляли части 0,1 М HEPES-буферов (N-2-гидроксиэтилпиперазин-N'-2-этансульфоновая кислота) (pH 7,4), содержащие различные количества хлорида цинка, с получением концентрации белка около 0,1 мг/мл. Полученные образцы смешивали и хранили при комнатной температуре (22oC) в течение приблизительно 18 часов. Затем смеси центрифугировали (Fisher Model 235C micro-centrifuge) в течение 5 минут. Прозрачные супернатанты пипетировали из пробирок. Содержание белка в супернатантах оценивали путем измерения их оптической плотности при 280 нм при помощи спектрофотометра (Gilford 260). Теоретическое значение оптической плотности при этой длине волны для 0,1 мг/мл раствора GLP-1-молекул в 1-см кюветах составляет 0,207. Результаты этого эксперимента представлены в таблице 2.
Этот пример проиллюстрировал, что для образования комплекса требуется лишь небольшое количество цинка и что из указанных разведенных растворов осаждается значительная часть молекул GLP-1.
Пример 2
5 мг GLP-1 (7-36) NH2 полностью растворяли в 2,5 мл 0,1 М HEPES-буфера (pH 7,4), не содержащего цинка. Затем быстро добавляли еще 2,5 мл 0,1 М HEPES-буфера (pH 7,4), содержащего 0,6 мМ хлорида цинка. В полученном образце молярное отношение цинка к GLP-1 (7-36)NH2 составляло приблизительно 1: 1. После добавления раствор сразу становился мутным, и тотчас стал образовываться осадок. Смесь хранили при комнатной температуре (22oC) в течение 18 часов.
Образовавшийся осадок прочно прикреплялся ко дну стеклянного сосуда. Супернатант полностью декантировали с помощью пипетки. Затем осадок целиком растворяли в 5,0 мл 0,01 н. соляной кислоты. Для супернатанта и для растворов вновь растворенного осадка определяли оптическую плотность при 280 нм. Содержание цинка в этих растворах определяли с помощью атомно-абсорбционной спектрофотометрии. Результаты этого эксперимента представлены в таблице 3.
В этом примере показано, что при добавлении цинк-содержащего HEPES-раствора из раствора осаждается большая часть GLP-1 (7-36)NH2. Значение оптической плотности при 280 нм, равное 1,932, указывают на то, что концентрация GLP-1 (7-36)NH2 во вновь растворенном преципитате составляет 0,933 мг/мл или 283 мкМ. Концентрация цинка в этом же самом растворе, составляющая 13,3 частей на миллион, эквивалентна концентрации цинка 203 мкМ. Поэтому молярное отношение цинка к GLP-1 (7-36)NH2 в преципитате составляет 0,717:1.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения рекомбинантной плазмидной ДНК и способ получения зимогенной формы белка С человека | 1991 |
|
SU1838411A3 |
| АНАЛОГ ИНСУЛИНА, ОБЛАДАЮЩИЙ АКТИВНОСТЬЮ СНИЖЕНИЯ УРОВНЯ ГЛЮКОЗЫ В КРОВИ | 1991 |
|
RU2109749C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЗИМОГЕННОЙ ФОРМЫ ЧЕЛОВЕЧЕСКОГО БЕЛКА С | 1989 |
|
RU2018535C1 |
| Производное инсулинотропного пептида с модифицированным N-концевым зарядом | 2014 |
|
RU2673179C2 |
| СПОСОБ ВВЕДЕНИЯ МОЛЕКУЛ GLP-1 | 2003 |
|
RU2332229C2 |
| Ацилированное производное GLP-1 | 2019 |
|
RU2773242C2 |
| ПРОИЗВОДНОЕ ИНСУЛИНОТРОПНОГО ПЕПТИДА, СОДЕРЖАЩЕЕ МОДИФИЦИРОВАННУЮ N-КОНЦЕВУЮ АМИНОКИСЛОТУ | 2008 |
|
RU2442792C2 |
| ПРОИЗВОДНЫЕ ПОЛИПЕПТИДА, ОБЛАДАЮЩИЕ ИНСУЛИНОТРОПНОЙ АКТИВНОСТЬЮ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБЫ УСИЛЕНИЯ ДЕЙСТВИЯ ИНСУЛИНА, СПОСОБЫ ЛЕЧЕНИЯ ДИАБЕТА | 1993 |
|
RU2128663C1 |
| АНТИТЕЛА ПРОТИВ КОМПЛЕКСА ANGPTL3/8 И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2019 |
|
RU2791034C2 |
| Способ конструирования рекомбинантной плазмидной ДНК, кодирующей зимоген С человека | 1988 |
|
SU1739854A3 |


Описывается новый Glp-1-молекулярный комплекс, состоящий из катиона двухвалентного металла, ассоциированного и способного к совместному осаждению с соединением формулы
R1-X-Glu-Gly-Thr-Phe-Thr-Ser-Asp-Val-Ser-Ser-Tyr-Leu-Y-Gly-Gln-Ala-Ala-Lys-Z-Phe-Ile-Ala-Trp-Leu-Val-Lys-Gly-Arg-R2,
где R1 выбирают из группы, включающей в себя L-гистидин, D-гистидин, дезамино-гистидин, 2-амино-гистидин, β-гидроксигистидин, гомогистидин, альфа-фторометил-гистидин, и альфа-метил-гистидин, Х выбирают из группы, включающей в себя Ala, Gly, Val, Thr, Ile, и альфа-метил-Ala; Y выбирают из группы, включающей в себя Glu, Gln, Ala, Thr, Ser, и Gly; Z выбирают из группы, включающей в себя Glu, Gln, Ala, Thr, Ser, и Gly; R2 выбирают из группы, включающей в себя NH2, и Gly - ОН, при условии, что указанное соединение имеет изоэлектрическую точку в диапазоне 6,0 - 9,0, и при условии, что, если R1 является His, Х является Ala, Y является Glu, a Z является Glu, то R2 должен представлять собой NH2 в молярном соотношении 0,1-3:1. Он может быть использован для увеличения экспрессии инсулина из островковых β-клеток поджелудочной железы млекопитающего и для лечения ювенильного сахарного диабета у млекопитающих. Описываются фармацевтическая композиция и способ получения комплекса. 4 с. и 8 з.п. ф-лы, 3 табл.
R1-X-Glu-Gly-Thr-Phe-Thr-Ser-Asp-Val-Ser-Ser-Tyr-Leu-Y-Gly-Gln-Ala-Ala-Lys-Z-Phe-Ile-Ala-Тrр-Leu-Val-Lys-Gly-Arg-R2,
где R1 выбирают из группы, включающей в себя L-гистидин, D-гистидин, дезамино-гистидин, 2-амино-гистидин, β-гидроксигистидин, гомогистилин, альфа-фторометил-гистидин и альфа-метилгистидин;
X выбирают из группы, включающей в себя Ala, Gly, Val, Thr, Ile, и альфа-метил-Ala;
Y выбирают из группы, включающей в себя Glu, Gln, Ala, Thr, Ser и Gly;
Z выбирают из группы, включающей в себя Glu, Gin, Ala, Thr, Ser и Gly;
R2 выбирают из группы, включающей в себя NH2 и Gly-OH, при условии, что указанное соединение имеет изоэлектрическую точку в диапазоне 6,0 - 9,0, и при условии, что, если R1 является His, X является Ala, Y является Glu, а Z является Glu, то R2 должен представлять собой NH2, в молярном соотношении металл: GLP-1, равном 0,1 - 3 : 1.
H2N-His-Val-Glu-Gly-Thr-Phe-Thr-Ser-Asp-Val-Ser-Ser-Tyr-Leu-Gly-Glu-Gln-Ala-Lys-Glu-Phe-Ile-Ala-Trp-Leu-Val-Lys-Gly-Arg-Gly-OH
в эффективном количестве в сочетании с одним или несколькими фармацевтическими приемлемыми носителями, разбавителями или наполнителями.
| SU 913939 А, 1982 | |||
| Устройство для измерения напряжений в массиве горных пород | 1976 |
|
SU587255A1 |
| JP 54024868 А, 1979. | |||

