[1] Данное изобретение в целом относится к биологии и медицине, а более конкретно к антителам (Ат, Ab), которые связывают и тем самым нейтрализуют комплексы ангиопоэтин-подобных белков (ANGPTL) 3/8 человека. Такие Ат могут повышать активность липопротеинлипазы (ЛПЛ, LPL) и тем самым снижать уровень триглицеридов (ТГ, TG) в сыворотке, в результате чего их можно использовать для лечения заболеваний и нарушений, связанных с метаболизмом липидов и глюкозы.
[2] ANGPTL представляют собой семейство белков, которые регулируют ряд физиологических и патофизиологических процессов. Особый интерес в данном документе представляет роль ANGPTL3 и ANGPTL8 в метаболизме липидов и глюкозы.
[3] Сведения подтверждают роль ANGPTL3 как основного регулятора метаболизма липопротеинов, и то, что он может регулировать клиренс ТГ путем ингибирования ЛПЛ и ингибирования эндотелиальной липазы (ЭЛ, EL). См., Chi et al. (2017) Mol. Metab. 6:1137-1149. Дефицит, инактивация или потеря ANGPTL3 может привести к низким уровням холестерина липопротеинов низкой плотности (ХС-ЛПНП, LDL-C), холестерина липопротеинов высокой плотности (ХС-ЛПВП, HDL-C) и ТГ. ANGPTL3 также может влиять на чувствительность к инсулину, тем самым играя роль в модуляции не только метаболизма липидов, но и метаболизма глюкозы. См., Robciuc et al. (2013) Arterioscler. Thromb. Vasc. Biol. 33:1706-1713. Нуклеотидные и аминокислотные последовательности ANGPTL3 человека известны. Например, одна нуклеотидная последовательность может быть найдена в Референтной Последовательности NCBI № NM_014495 (SEQ ID NO:1), и одна аминокислотная последовательность может быть найдена в Референтной Последовательности NCBI № NP_055310 (SEQ ID NO:2).
[4] ANGPTL8 высоко экспрессируется в печени и жировой ткани и, как сообщается, ингибирует ЛПЛ, образуя комплекс с ANGPTL3 и тем самым активируя его. См. Chi, цитировано выше. ANGPTL8 человека, по-видимому, индуцируется приемом пищи. Нуклеотидные и аминокислотные последовательности ANGPTL8 человека известны. Например, одна нуклеотидная последовательность может быть найдена в Референтной Последовательности NCBI № NM_018687 (SEQ ID NO:3), и одна аминокислотная последовательность может быть найдена в Референтной Последовательности NCBI № NP_061157 (SEQ ID NO:4).
[5] Существуют комплексы ANGPTL3/8, которые имеют один или более ANGPTL3, связанных с одним или более ANGPTL8. Данные свидетельствуют о том, что эти комплексы более эффективно опосредуют ингибирование ЛПЛ по сравнению с только ANGPTL3 или ANGPTL8. Более того, комплексы ANGPTL3/8 могут быть получены in vitro путем коэкспрессии ANGPTL8 и ANGPTL3 в системе экспрессии млекопитающих. См. Chi, цитировано выше.
[6] Известны антитела, которые связываются либо с ANGPTL3, либо с ANGPTL8, и их можно использовать отдельно или в комбинации друг с другом для лечения заболеваний и нарушений, связанных с метаболизмом липидов и глюкозы. Например, публикация Международной заявки на патент № WO 2012/174178 раскрывает полностью человеческое моноклональное Aт и его антигенсвязывающие фрагменты, которые связываются с ANGPTL3 и препятствуют его активности. Также известны другие терапевтические антитела против ANGPTL3. См., например, публикация Международной заявки на патент № WO 2008/073300 и Патент США № 7,935,796. Аналогичным образом, публикация Международной заявки на патент № WO 2017/027316 раскрывает полностью человеческое моноклональное Aт или его антигенсвязывающие фрагменты, которые связываются с ANGPTL8 и препятствуют его активности. Кроме того, публикация Международной заявки на патент № WO 2017/177181 раскрывает комбинированную терапию с использованием Ат против ANGPTL3 и Ат против ANGPTL8.
[7] К сожалению, существующие Ат, которые связываются только с ANGPTL3 или ANGPTL8, не подавляют полностью действие данных ANGPTL и/или комплексов ANGPTL3/8 на метаболизм липидов и/или глюкозы. См., например, Dewey et al. (2017) N. Engl. J. Med. 377:211-221; и Gusarova et al. (2017) Endocrinology 158:1252-1259. В связи с этим существует потребность в дополнительных Ат, особенно в Ат против комплекса ANGPTL3/8, для лечения заболеваний и нарушений, связанных с метаболизмом липидов и глюкозы, при этом такие Ат обладают улучшенными фармакологическими ингибирующими и/или регулирующими свойствами для модуляции метаболизма липидов и/или глюкозы.
[8] Чтобы удовлетворить данную потребность, для модифицированного комплекса ANGTPL3/8 предложены нуклеотидные и аминокислотные последовательности. Соответственно, в данном документе описаны последовательности нуклеиновых кислот, кодирующие один или более модифицированных ANGPTL3 и модифицированных ANGPTL8 (т.е. слитые белки). В некоторых случаях последовательности нуклеиновых кислот содержат полинуклеотидную последовательность, кодирующую слитый белок ANGPTL3, имеющий аминокислотную последовательность SEQ ID NO:17. В других случаях последовательности нуклеиновых кислот содержат полинуклеотидную последовательность, кодирующую слитый белок ANGPTL8, имеющий аминокислотную последовательность SEQ ID NO:18. В других случаях последовательности нуклеиновой кислоты содержат полинуклеотидную последовательность, кодирующую SEQ ID NO:17 и 18.
[9] Кроме того, предложены конструкции нуклеиновых кислот, которые содержат полинуклеотидную последовательность, кодирующую слитый белок ANGPTL3, как описано в данном документе, слитый белок ANGPTL8, как описано в данном документе, или и то, и другое, при этом такие конструкции могут быть кассетой или вектором экспрессии.
[10] Ввиду вышеизложенного предложены клетки-хозяева, которые включают в себя одну или более кассет или векторов экспрессии, как описано в данном документе. В некоторых случаях клетки-хозяева представляют собой эукариотические клетки. В некоторых случаях полинуклеотидные последовательности слитых белков ANGPTL3 и ANGTPL8 находятся в отдельных кассетах или векторах экспрессии, тогда как в других случаях они могут находиться в одной кассете или векторе экспрессии.
[11] Также предложены слитые белки ANGPTL3, которые содержат аминокислотную последовательность SEQ ID NO:17 или 19, а также их активные варианты или фрагменты. Аналогичным образом предложены слитые белки ANGPTL8, которые содержат аминокислотную последовательность SEQ ID NO:18 или 20, а также их активные варианты или фрагменты.
[12] Кроме того, предложены функциональные комплексы ANGPTL3/8, в особенности комплексы ANGPTL3/8 человека, при этом фрагмент ANGPTL3 комплекса представляет собой нативный (полноразмерный или усеченный) ANGPTL3 или слитый белок ANGPTL3, как описано в данном документе, и при этом фрагмент ANGPTL8 комплекса представляет собой слитый белок ANGPTL8, как описано в данном документе. В некоторых случаях слитый белок ANGPTL3 содержит аминокислотную последовательность SEQ ID NO:19. Аналогичным образом и в некоторых случаях слитый белок ANGPTL8 содержит аминокислотную последовательность SEQ ID NO:20. Более того, и в некоторых случаях комплексы могут иметь соотношение 1:1 фрагмента ANGPTL3 к фрагменту ANGPTL8. В других случаях комплексы могут иметь соотношения, отличные от 1:1, такие как соотношение 1:2, 1:3, 2:1 или 3:1 фрагмента ANGPTL3 к фрагменту ANGPTL8, соответственно.
[13] Также предложены способы получения рекомбинантных комплексов ANGTPL3/8. Способы могут включать, по меньшей мере, этап экспрессии одной или более полинуклеотидных последовательностей для фрагмента ANGPTL3 и фрагмента ANGPTL8, как описано в данном документе, в клетке-хозяине, такой как в системе экспрессии млекопитающих, для получения из них комплексов ANGPTL3/8. В некоторых случаях фрагменты ANGPTL3 и ANGPTL8 предложены в отдельных конструкциях или векторах экспрессии. В других случаях ANGPTL3 и ANGPTL8 предложены в одной конструкции или векторе экспрессии. Способы также могут включать этап очистки полученных комплексов ANGPTL3/8, который может включать не только концентрирование комплексов ANGPTL3/8, но также удаление одной или более меток, линкеров и сывороточного альбумина из фрагмента ANGPTL3 и/или фрагмента ANGPTL8. Способы также могут включать этап концентрирования комплексов ANGPTL3/8 до и/или после этапа очистки.
[14] Во-вторых, предложено Ат к комплексу ANGPTL3/8, а также его применение, которое включает лечение заболеваний и нарушений, связанных с метаболизмом липидов и глюкозы, путем связывания и ингибирования активности комплекса ANGPTL3/8.
[15] Эффективное количество Ат против комплекса ANGPTL3/8, описанного в данном документе, или его фармацевтически приемлемой соли можно использовать для увеличения активности ЛПЛ, снижения уровня ТГ и лечения заболеваний или нарушений, связанных с метаболизмом липидов и/или глюкозы, у индивидуума, нуждающегося в этом.
[16] Описанное в данном документе Ат против комплекса ANGPTL3/8 связывает растворимый комплекс ANGPTL3/8, тем самым увеличивая активность ЛПЛ и снижая уровни ТГ в сыворотке. Индивидуумы с более низким уровнем триглицеридов имеют более низкий риск развития сердечно-сосудистых заболеваний. Преимущественно, описанное в данном документе Ат против комплекса ANGPTL3/8 связывается только с комплексом ANGPTL3/8, а не только с ANGPTL3 или только с ANGPTL8 в соответствующих концентрациях. Считается, что Ат против комплекса ANGPTL3/8 увеличивает катаболизм ТГ-богатых липопротеинов (ТГБЛП, TRL), что снижает ТГ и/или не-ХС-ЛПВП (non-HDL-C), тем самым улучшая факторы риска дислипидемии при атеросклеротическом сердечно-сосудистом заболевании (АССЗ, ASCVD), не устраняемого современными способами лечения. Более того, поскольку описанное в данном документе Ат против комплекса ANGPTL3/8 не связывается только с ANGPTL3 или ANGPTL8, другие воздействия данных ANGPTL не ингибируются, что может приводить к меньшему количеству нежелательных эффектов in vivo, таких как снижение дерепрессии ЭЛ.
[17] В частности, Ат против комплекса ANGPTL3/8 представляет собой Ат против комплекса ANGPTL3/8 человека. В некоторых случаях Ат против комплекса ANGPTL3/8 может отменять, блокировать, ингибировать, затруднять, нейтрализовать или снижать активность in vivo комплекса ANGPTL3/8, в особенности его ингибирующую активность ЛПЛ. В некоторых случаях Aт против комплекса ANGPTL3/8 может быть полноразмерным или может быть только антигенсвязывающим фрагментом (например, фрагментом Fab, F(ab')2 или scFv). Желательные свойства Ат против комплекса ANGPTL3/8 включают снижение уровня ТГ при низких дозах Ат, которое сохраняется в течение, по меньшей мере, 21 дня.
[18] В некоторых случаях Ат против комплекса ANGPTL3/8 связывается с комплексом ANGPTL3/8 человека и содержит области легкой цепи, определяющие комплементарность - LCDR1, LCDR2 и LCDR3, и области тяжелой цепи, определяющие комплементарность - HCDR1, HCDR2 и HCDR3, при этом LCDR1 имеет аминокислотную последовательность RSSQSLLDSDDGNTYLD (SEQ ID NO:11), LCDR2 имеет аминокислотную последовательность YMLSYRAS (SEQ ID NO:12), и LCDR3 имеет аминокислотную последовательность MQRIEFPLT (SEQ ID NO:13), и при этом HCDR1 имеет аминокислотную последовательность TFSGFSLSISGVGVG (SEQ ID NO:14), HCDR2 имеет аминокислотную последовательность LIYRNDDKRYSPSLKS (SEQ ID NO:15), и HCDR3 имеет аминокислотную последовательность ARTYSSGWYGNWFDP (SEQ ID NO:16).
[19] Дополнительно предложено Ат, содержащее вариабельную область легкой цепи (LCVR), при этом LCVR имеет аминокислотную последовательность SEQ ID NO:9; или Aт, содержащее вариабельную область тяжелой цепи (HCVR), при этом HCVR имеет аминокислотную последовательность SEQ ID NO:10. В некоторых случаях Aт содержит LCVR с аминокислотной последовательностью SEQ ID NO:9 и HCVR с аминокислотной последовательностью SEQ ID NO:10. В некоторых случаях Aт содержит легкую цепь (LC) и тяжелую цепь (HC), при этом LC имеет аминокислотную последовательность SEQ ID NO:5 или HC имеет аминокислотную последовательность SEQ ID NO:6. Альтернативно Aт содержит легкую цепь (LC) и тяжелую цепь (HC), при этом LC имеет аминокислотную последовательность SEQ ID NO:5, и HC имеет аминокислотную последовательность SEQ ID NO:6. В некоторых случаях Aт представляет собой изотип IgG4.
[20] В некоторых случаях Aт против комплекса ANGPTL3/8 может представлять собой вариант Aт, описанный выше, в особенности LC-вариант, имеющий мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22), мутацию D33T (SEQ ID NO: 23); мутацию M56T (SEQ ID NO: 24), мутацию E99Q (SEQ ID NO: 25) или их комбинацию (например, мутации D33T и M56T или мутации D33A и M56T) в отношении LC, имеющей аминокислотную последовательность SEQ ID NO:5.
[21] Кроме того, предложено Aт, которое получают путем культивирования клетки млекопитающего, содержащей молекулу кДНК, при этом молекула кДНК кодирует полипептиды, имеющие аминокислотные последовательности SEQ ID NO:5 и 6, в таких условиях, при которых полипептиды экспрессируются, и извлечения Ат. В альтернативном варианте предложено Aт, которое получают путем культивирования клетки млекопитающего, содержащей две молекулы кДНК, при этом первая молекула кДНК кодирует полипептид, имеющий аминокислотную последовательность SEQ ID NO:5, а вторая молекула кДНК кодирует полипептид, имеющий аминокислотную последовательность SEQ ID NO:6, в таких условиях, при которых полипептиды экспрессируются, и извлечения Aт. В некоторых случаях Aт против комплекса ANGPTL3/8 может представлять собой вариант описанного выше Aт, в особенности LC-вариант, имеющий мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22), мутацию D33T (SEQ ID NO: 23); мутацию M56T (SEQ ID NO: 24), мутацию E99Q (SEQ ID NO: 25) или их комбинацию в отношении LC, имеющей аминокислотную последовательность SEQ ID NO:5.
[22] Кроме того, предложено Aт, которое связывается с комплексом ANGPTL3/8 человека и нейтрализует его в стандартном анализе активности ЛПЛ с EC50 0,5 нМ или менее. Также предложено Aт, которое связывается с комплексом ANGPTL3/8 человека с константой диссоциации менее или равной 1×10-6 M. Вместе с тем предложено Aт, которое связывается с ANGPTL3 человека и ANGPTL8 человека с константой диссоциации более чем 1×10-6 M. Кроме того, предложено Aт, которое связывается с комплексом ANGPTL3/8 человека с сигналом, более чем в 3 раза превышающим фоновый сигнал, характерный при отсутствии связывания, при измерении с помощью анализа ИФА с одноточечной калибровкой, но не связывается только с ANGPTL3 человека или только с ANGPTL8 с сигналом, более чем в 3 раза превышающим фоновый сигнал, характерный при отсутствии связывания, при измерении с помощью анализа ИФА с одноточечной калибровкой. Аналогичным образом предложено Aт, которое снижает ТГ in vivo, по меньшей мере, на 50% по сравнению с контролем IgG в дозе 10 мг/кг в момент времени через 14 дней после введения дозы.
[23] В-третьих, предложена фармацевтическая композиция, которая содержит Aт, описанное в данном документе, или популяцию Ат, описанных в данном документе, и приемлемый носитель, разбавитель или эксципиент. Также предложена клетка млекопитающего, содержащая молекулу ДНК, которая содержит полинуклеотидную последовательность, кодирующую полипептиды, имеющие аминокислотные последовательности SEQ ID NO:5 и SEQ ID NO:6, при этом клетка способна экспрессировать Aт, описанное в данном документе. Дополнительно предложена клетка млекопитающего, содержащая первую молекулу ДНК и вторую молекулу ДНК, при этом первая молекула ДНК содержит полинуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность SEQ ID NO:5, и при этом вторая молекула ДНК содержит полинуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность SEQ ID NO:6, и при этом клетка способна экспрессировать Aт, описанное в данном документе. В некоторых случаях Aт против комплекса ANGPTL3/8 может представлять собой вариант описанного выше Aт, в особенности LC-вариант, имеющий мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22), мутацию D33T (SEQ ID NO: 23); мутацию M56T (SEQ ID NO:24), мутацию E99Q (SEQ ID NO:25) или их комбинацию в отношении LC, имеющей аминокислотную последовательность SEQ ID NO:5.
[24] В-четвертых, предложен способ получения Aт, который включает культивирование клетки млекопитающего с молекулой ДНК, имеющей полинуклеотидную последовательность, кодирующую полипептиды, имеющие аминокислотные последовательности SEQ ID NO:5 и SEQ ID NO:6, при этом клетка способна экспрессировать Aт, описанное в данном документе, в условиях, в которых происходит экспрессия Ат, и выделение экспрессированного Ат. В некоторых случаях полинуклеотидная последовательность, кодирующая полипептид, имеющий аминокислотную последовательность SEQ ID NO:5, может кодировать мутацию D31S, мутацию D33A, мутацию D33T, мутацию M56T, мутацию E99Q или их комбинацию. В данном документе также предложен способ лечения АССЗ, хронической почечной недостаточности (ХПН, CKD), диабета, гипертриглицеридемии, неалкогольного стеатогепатита (НАСГ, NASH), ожирения или их комбинации, при этом способ включает введение индивидууму, нуждающемуся в этом, эффективного количества Aт, описанного в данном документе. Дополнительно предложен способ снижения уровня ТГ, который включает введение индивидууму, нуждающемуся в этом, эффективного количества Ат, описанного в данном документе.
[25] В-пятых, предложено Aт для использования в терапии. В частности, Aт предназначено для лечения АССЗ, ХПН, диабета, гипертриглицеридемии, НАСГ, ожирения или их комбинации. Также предложена фармацевтическая композиция для применения при лечении АССЗ, ХПН, диабета, гипертриглицеридемии, НАСГ, ожирения или их комбинации, которая содержит эффективное количество Ат, описанного в данном документе.
[26] Преимущества, эффекты, особенности и объекты, отличные от изложенных выше, станут более очевидными при рассмотрении подробного описания, приведенного ниже. В таком подробном описании делается ссылка на следующий графический материал(ы), при этом:
[27] На Фиг. 1 представлено изображение ДСН-ПААГ геля, демонстрирующее анализ комплекса ANGPTL3/8 в восстанавливающих и невосстанавливающих условиях.
[28] Упоминание элемента в единственном числе не исключает возможности присутствия более чем одного из элементов, если контекст явным образом не предполагает присутствия одного и только одного из элементов. Соответственно, термины в единственном числе обычно означают «по меньшей мере, один».
[29] Определения
[30] Используемый в контексте данного документа термин «около» означает в пределах статистически значимого диапазона значения или значений, таких как, например, установленная концентрация, длина, молекулярная масса, pH, сходство последовательности, временные рамки, температура, объем и т. д. Такое значение или диапазон может быть в пределах порядка, обычно в пределах 20%, более типично в пределах 10% и даже более типично в пределах 5% от данного значения или диапазона. Допустимое отклонение, охватываемое термином «около», будет зависеть от конкретной исследуемой системы и может быть легко оценено специалистом в данной области техники.
[31] Используемый в контексте данного документа термин «аффинность» означает силу связывания Ат с эпитопом на комплексе ANGPTL3/8.
[32] Используемый в контексте данного документа термин «ангиопоэтин-подобный белок 3» или «ANGPTL3» означает белок, имеющий аминокислотную последовательность, содержащую SEQ ID NO:2.
[33] Используемый в контексте данного документа термин «ангиопоэтин-подобный белок 8» или «ANGPTL8» означает белок, имеющий аминокислотную последовательность, содержащую SEQ ID NO:4.
[34] Используемый в контексте данного документа термин «комплекс ANGPTL3/8» означает мультибелковый комплекс одного или более соединений ANGPTL3, которые связаны с одним или более соединениями ANGPTL8.
[35] Используемый в контексте данного документа термин «Ат против комплекса ANGPTL3/8» или «комплексное Ат против ANGPTL3/8» означает Ат, которое одновременно распознает и связывается с областью как на ANGPTL3, так и на ANGPTL8, в особенности в случае, когда они находится в форме ANGPTL3/8 комплекса. Как правило, Ат против комплекса ANGPTL3/8 обычно не связывается с другими членами семейства ANGPTL (например, ANGPTL1, ANGPTL2, ANGPTL4, ANGPTL5, ANGPTL6 или ANGPTL7). Более того, как отмечалось в другом месте данного документа, Ат против комплекса ANGPTL3/8 также не будет связываться только с ANGPTL3 или только с ANGPTL8 при определенных концентрациях, как описано ниже в анализе ИФА с одноточечной калибровкой.
[36] Используемый в контексте данного документа термин «связывать» или «связывает» означает способность белка образовывать тип химической связи или силы притяжения с другим белком или молекулой, как это определяется обычными способами, известными в данной области техники. Связывание можно охарактеризовать равновесной константой диссоциации (KD) около 1×10-6 M или менее (т.е. меньшее значение KD означает более плотное связывание). Способы определения, связываются ли две молекулы, хорошо известны в данной области техники и включают, например, равновесный диализ, поверхностный плазмонный резонанс и т.п. Как указано в данном документе Aт против комплекса ANGPTL3/8 связывается только с комплексом ANGPTL3/8 и не связывается отдельно только с ANGPTL3 или только с ANGPTL8. Связывается ли Ат только с комплексом ANGPTL3/8, а не отдельно только с ANGPTL3 или только с ANGPTL8, можно определить с помощью стандартных анализов ИФА в одноточечном формате, как описано ниже, а связывание можно охарактеризовать с помощью технологии Biacore, как описано ниже. Несмотря на то, что описанные в данном документе Ат являются человеческими, они могут, однако, проявлять перекрестную реактивность с другими комплексами ANGPTL3/8 других видов, например комплексом ANGPTL3/8 яванской макаки, комплексом ANGPTL3/8 мыши или комплексом ANGPTL3/8 крысы.
[37] Используемый в контексте данного документа термин «эффективное количество» обозначает количество или дозу соединения или фармацевтической композиции, содержащей его, которое при однократном или многократном введении доз индивидууму будет вызывать биологическую или терапевтическую реакцию, или желаемый терапевтический эффект в ткани, системе, организме животного, млекопитающего или человека, что находиться под наблюдением исследователя, ветеринара, врача или другого лечащего персонала. В некоторых случаях эффективное количество соединения, описанного в данном документе, или композиций, включающих его, для индивидуума, нуждающегося в этом, может привести к увеличению активности ЛПЛ. Доза может включать в себя более высокую начальную нагрузочную дозу с последующей более низкой дозой. Дозу можно вводить с любым терапевтически эффективным интервалом, например, несколько раз в день, один раз в день, через день, три раза в неделю, два раза в неделю, один раз в неделю, один раз в две недели, один раз в месяц, один раз каждые два месяца и т. д. Доза, составляющая эффективное количество, может составлять от 0,01 мг/кг до 100 мг/кг.
[38] Используемый в контексте данного документа термин «равновесная константа диссоциации» или «KD» означает количественное измерение аффинности Aт к конкретному взаимодействию антигена, такой как аффинности Aт к комплексу ANGPTL3/8, в особенности мере склонности конъюгата Aт/комплекс ANGPTL3/8 к обратимому разделению на его составные части. Аналогичным образом, как здесь используется, «равновесной константой ассоциации» или «Ka» означает величину, обратную KD.
[39] Используемый в контексте данного документа термин «функциональный» означает, что слитый белок ANGPTL3, слитый белок ANGPTL8 или комплекс ANGPTL3/8 обладают биологической активностью, аналогичной активности природного ANGPTL3, природного ANGPTL8 или природного комплекса ANGPTL3/8, включая, например, ингибирование ЛПЛ или действуя как антиген, к которому может быть создано и направлено Ат.
[40] Используемый в контексте данного документа термин «заболевание или нарушение, связанное с метаболизмом глюкозы» означает диабет и тому подобное.
[41] Используемый в контексте данного документа термин «заболевание или нарушение, связанное с метаболизмом липидов» означает состояние, связанное с аномальным метаболизмом липидов, такое как дислипидемия, гиперлипидемия и гиперлипопротеинемия, включая гипертриглицеридемию, гиперхолестеринемию, хиломикронемию, смешанную дислипидемию (ожирение, метаболический синдром, диабет и т. д.), липодистрофию и липоатрофию. Данный термин также охватывает некоторые сердечно-сосудистые заболевания, такие как атеросклероз и коронарная болезнь сердца, острый панкреатит, НАСГ, ожирение и тому подобное.
[42] Используемый в контексте данного документа термин «полумаксимальная эффективная концентрация» или «ЕС50» означает концентрацию Aт (обычно выраженную в молярных единицах (M)), которая вызывает ответ на полпути между исходным уровнем и максимумом по прошествии заранее определенного периода времени. EC50, описанная в данном документе, в идеале составляет 3,0 нМ или менее.
[43] Используемый в контексте данного документа термин «конструкция нуклеиновой кислоты» или «кассета экспрессии» означает молекулу нуклеиновой кислоты, имеющую, по меньшей мере, регуляторную последовательность, функционально связанную с кодирующей последовательностью. Таким образом, регуляторная последовательность, такая как промотор, находится в рабочем взаимодействии с последовательностями нуклеиновой кислоты, кодирующими, по меньшей мере, один представляющий интерес полипептид, такой как слитые белки ANGPTL3, описанные в данном документе, и/или слитые белки ANGPTL8, описанные в данном документе. Такие конструкции нуклеиновых кислот могут быть в форме кассеты экспрессии или переноса. Конструкция нуклеиновой кислоты может включать олигонуклеотид или полинуклеотид, состоящий из дезоксирибонуклеотидов, рибонуклеотидов или их комбинаций, содержащих нуклеотидные последовательности для одного или более представляющих интерес полипептидов.
[44] Используемый в контексте данного документа термин «функционально связанный» означает, что элементы конструкции нуклеиновой кислоты сконфигурированы так, чтобы выполнять свои обычные функции. Таким образом, регуляторные последовательности (т.е. промоторы), функционально связанные с кодирующей последовательностью, способны влиять на экспрессию кодирующей последовательности. Регуляторные последовательности не обязательно должны контактировать с кодирующей последовательностью, если они функционируют так, чтобы направлять ее экспрессию (т.е. поддерживать правильную рамку считывания). Таким образом, например, промежуточные нетранслируемые, но еще транскрибируемые последовательности могут присутствовать между промотором и кодирующей последовательностью, и промоторная последовательность по-прежнему может считаться «функционально связанной» с кодирующей последовательностью.
[45] Используемый в контексте данного документа термин «регуляторная последовательность» или «регуляторные последовательности» означает промоторы, сигналы полиаденилирования, последовательности терминации транскрипции и трансляции, регуляторные домены, расположенные до кодирующей последовательности, точки начала репликации, области внутренней посадки рибосомы («IRES»), энхансеры и тому подобное, которые вместе обеспечивают репликацию, транскрипцию и трансляцию кодирующей последовательности в реципиентной клетке-хозяине. Не все эти регуляторные последовательности должны присутствовать обязательно, если выбранная кодирующая последовательность способна реплицироваться, транскрибироваться и транслироваться в соответствующей клетке-хозяине.
[46] Используемый в контексте данного документа термин «кодирующая последовательность» или «кодирующие последовательности» означает последовательность нуклеиновой кислоты, которая кодирует один или более представляющих интерес полипептидов, и представляет собой последовательность нуклеиновой кислоты, которая транскрибируется (в случае ДНК) и транслируется (в случае РНК) в полипептид in vitro или in vivo при размещении под контроль соответствующих регуляторных последовательностей. Границы кодирующей последовательности(ей) определяются стартовым кодоном на 5’ (амино) конце и стоп-кодоном трансляции на 3' (карбокси) конце. Кодирующая последовательность может содержать, но не ограничивается этим, последовательности вирусной нуклеиновой кислоты, кДНК из прокариотической или эукариотической мРНК, последовательности геномной ДНК из прокариотической или эукариотической ДНК или даже синтетические последовательности ДНК.
[47] Что касается регуляторных и кодирующих последовательностей, то они могут быть нативными/аналогичными по отношению к клетке-хозяину или друг другу. Альтернативно, регуляторная и кодирующая последовательности могут быть гетерологичными по отношению к клетке-хозяину или друг другу.
[48] Используемый в контексте данного документа термин «промотор» означает нуклеотидную область, состоящую из регуляторной последовательности нуклеиновой кислоты, при этом регуляторная последовательность получена из гена или создана синтетически и способна связывать РНК-полимеразу и инициировать транскрипцию расположенной после него (3'-направление) кодирующей последовательности. В конструкциях нуклеиновых кислот можно использовать ряд промоторов, включая нативный промотор для одного или более представляющих интерес полипептидов. Альтернативно, промоторы могут быть выбраны на основании желаемого результата. Такие промоторы могут включать, но не ограничиваются ими, индуцируемые промоторы, репрессируемые промоторы и конститутивные промоторы.
[49] Используемый в контексте данного документа термин «вариант» означает полинуклеотид или полипептид, имеющий одну или более модификаций, таких как добавление, делеция, вставка и/или замена одной или более специфических нуклеиновых кислот или аминокислотных остатков по сравнению с референтной нуклеотидной или аминокислотной последовательностью. Таким образом, вариант включает одно или более изменений по сравнению с референтной нуклеотидной или аминокислотной последовательностью. Как указано в данном документе Aт против комплекса ANGPTL3/8 может иметь вариацию LC или HC. В частности, Aт может быть LC-вариантом, имеющим мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22), мутацию D33T (SEQ ID NO:23); мутацию M56T (SEQ ID NO:24) или мутацию E99Q (SEQ ID NO:25) в отношении LC, имеющей аминокислотную последовательность SEQ ID NO:5. Аналогичным образом, LC-вариант может представлять собой комбинацию любых двух из вышеперечисленных мутаций, таких как, например, мутаций D31S и D33A, мутаций D31S и D33T, мутаций D31S и M56T, мутаций D31S и E99Q, мутаций D33A и M56T, мутаций D33A и E99Q, мутаций D33T и M56T, мутаций D33T и E99Q, а также M56T и E99Q, опять же в отношении LC, имеющей аминокислотную последовательность SEQ ID NO:5. Более того, LC-вариант может представлять собой комбинацию любых трех из вышеперечисленных мутаций, таких как, например, мутаций D31S, D33A и M56T, мутаций D31S, D33A и E99Q, мутаций D31S, D33T и M56T, мутаций D31S, D33T и E99Q, D33A, мутаций M56T и E99Q, и мутаций D33T, M56T и E99Q, опять же в отношении LC, имеющей аминокислотную последовательность SEQ ID NO: 5. Кроме того, LC-вариант может представлять собой комбинацию любых четырех из вышеперечисленных мутаций, таких как, например, мутаций D31S, D33A, M56T и E99Q; и мутаций D31S, D33T, M56T и E99Q, опять же в отношении LC, имеющей аминокислотную последовательность SEQ ID NO:5.
[50] Используемый в контексте данного документа термин «вектор» означает репликон, такой как плазмида, фаг или космида, к которому может быть присоединена другая последовательность нуклеиновой кислоты, такая как кассета экспрессии так, чтобы вызвать репликацию присоединенной последовательности. Вектор способен переносить молекулы нуклеиновой кислоты в клетки-хозяева. Векторы обычно содержат один или небольшое количество сайтов распознавания эндонуклеазой рестрикции, при этом интересующая нуклеотидная последовательность может быть вставлена определенным образом без потери основной биологической функции вектора, а также селективный маркер, который можно использовать для идентификации и отбора клеток-хозяев, трансформированных вектором. Поэтому вектор может быть способен переносить последовательности нуклеиновой кислоты в клетки-мишени.
[51] Используемый в контексте данного документа термин «лечение» или «лечить» означает ведение индивидуума и уход за индивидуумом, имеющим патологию, для которой рекомендуется введение Ат против комплекса ANGPTL3/8 с целью борьбы с симптомами, или облегчения симптомов и осложнений таких патологий. Лечение включает в себя введение индивидууму, нуждающемуся в этом, соединения или композиции, содержащей соединение, описанное в данном документе, для предотвращения появления симптомов или осложнений, облегчения симптомов или осложнений, или устранения заболевания, патологии или нарушения. Лечение включает введение индивидууму, нуждающемуся в этом соединения или композиций, содержащих описанное в данном документе, приводящее к увеличению активности ЛПЛ и снижению ТГ. Индивидуум, подлежащий лечению, является животным, в особенности человеком.
[52] Используемый в контексте данного документа термин «пациент», «субъект» и «индивидуум» используются в данном документе взаимозаменяемо и означают животное, в особенности человека. В некоторых случаях индивидуум представляет собой человека и, кроме того, характеризуется заболеванием, нарушением или состоянием, которые могут улучшиться от введения Aт против комплекса ANGPTL3/8.
[53] Используемый в контексте данного документа термин «антитело» или «Aт» и тому подобное означает полноразмерное Aт, содержащее две тяжелые цепи и две легкие цепи, имеющие межцепочечные и внутрицепочечные дисульфидные связи. Аминоконцевая часть каждой из четырех полипептидных цепей содержит вариабельную область, в первую очередь отвечающую за распознавание антигена. Каждая HC содержит N-концевую HCVR и константную область HC (HCCR). Каждая легкая цепь содержит вариабельную область легкой цепи (LC)- (LCVR) и константную область LC - (LCCR). Как указано в данном документе, Aт представляет собой Aт типа иммуноглобулина G (IgG), и изотип IgG можно дополнительно разделить на подклассы (например, IgG1, IgG2, IgG3 и IgG4). Области HCVR и LCVR могут быть дополнительно подразделены на области гипервариабельности, называемые областями, определяющими комплементарность («CDR»), перемежающиеся с областями, которые являются более консервативными и называются каркасными областями («FR»). Каждая LCVR и HCVR содержит три CDR и четыре FR, расположенных от N-конца до C-конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В данном документе три CDR тяжелой цепи обозначены как HCDR1, HCDR2 и HCDR3, а три CDR легкой цепи обозначены как LCDR1, LCDR2 и LCDR3. CDR содержат большинство остатков, которые образуют специфические взаимодействия с антигеном, таким как комплекс ANGPTL3/8. Присвоение остатков различным CDR может быть выполнено с помощью таких алгоритмов, как, например, по Чотиа, по Кабату или по Норсу. Определение CDR по Норсу основано на кластеризации распространения аффинности с большим количеством кристаллических структур (North et al. (2011) J. Mol. Bio. 406:228-256). В данном документе CDR лучше всего обозначаются последовательностями, перечисленными в Списке Последовательностей, которые основаны на комбинации нескольких определений, включая по Норсу.
[54] Выделенная ДНК, кодирующая область HCVR, может быть преобразована в полноразмерный ген тяжелой цепи путем функционального соединения ДНК, кодирующей HCVR, с другой молекулой ДНК, кодирующей константные области тяжелой цепи. Последовательности генов константной области тяжелой цепи человека, а также других млекопитающих известны в данной области техники. Фрагменты ДНК, охватывающие эти области, можно получить, например, с помощью стандартной амплификации ПЦР.
[55] Выделенная ДНК, кодирующая область LCVR, может быть преобразована в полноразмерный ген легкой цепи путем функционального соединения ДНК, кодирующей LCVR, с другой молекулой ДНК, кодирующей константную область легкой цепи. Последовательности генов константной области легкой цепи человека, а также других млекопитающих, известны в данной области техники. Фрагменты ДНК, охватывающие эти области, можно получить, например, с помощью стандартной амплификации ПЦР. Константная область легкой цепи может представлять собой константную область каппа или лямбда.
[56] Описанные в данном документе Aт могут содержать часть Fc IgG4-PAA. Fc-часть IgG4-PAA имеет мутацию Ser на Pro в положении 231 (S231P), мутацию Phe на Ala в положении 237 (F237A) и мутацию Leu на Ala в положении 238 (L238A), согласно нумерации по абсолютному положению в SEQ ID NO:6. Мутация S231P является шарнирной мутацией, которая предотвращает образование полу-Ат (явление динамического обмена полумолекул в Ат IgG4). Мутации F237A и L238A дополнительно уменьшают эффекторную функцию и без того низкого изотипа IgG4 человека. Однако предполагается, что Ат, описанные в данном документе, альтернативно могут содержать другую Fc-часть.
[57] С целью уменьшения потенциальной индукции иммунного ответа при введении дозы человеку, некоторые аминокислоты могут потребовать обратных мутаций, чтобы соответствовать последовательностям зародышевой линии Ат.
[58] Фармацевтические композиции, содержащие описанные в данном документе соединения, могут быть парентерально введены индивидууму, нуждающемуся в таком лечении. Такой индивидуум может иметь АССЗ или иметь высокий риск развития АССЗ. Данные индивидуумы могут иметь острый коронарный синдром, инфаркт миокарда (ИМ, MI) в анамнезе, стабильную или нестабильную стенокардию, коронарную или другую артериальную реваскуляризацию, инсульт, транзиторную ишемическую атаку (ТИА, TIA), аневризму грудной или брюшной аорты или заболевание периферических артерий, предположительно атеросклеротического происхождения. Индивидуумы с высоким риском развития АССЗ дополнительно могут иметь сахарный диабет 2-го типа (СД2, T2D), ХПН или семейную гиперхолестеринемию (СГ, FH).
[59] Парентеральное введение может осуществляться путем подкожной, внутримышечной или внутривенной инъекции с помощью шприца, необязательно шприца-ручки, или механического инъектора. Альтернативно, парентеральное введение может осуществляться с помощью инфузионной помпы. В некоторых случаях фармацевтические композиции, подходящие для введения индивидууму, содержат терапевтически эффективное количество соединения, описанного в данном документе, и один или более фармацевтически приемлемых наполнителей. Такие фармацевтические композиции могут быть получены любым из множества способов с применением традиционных эксципиентов для фармацевтических препаратов, которые хорошо известны в данной области техники. См., например, Remington, “The Science and Practice of Pharmacy” (D.B. Troy ed., 21st Ed., Lippincott, Williams & Wilkins, 2006).
[60] Соединения, описанные в данном документе, могут быть использованы в одновременной, раздельной или последовательной комбинации с одним или более дополнительными терапевтическими агентами, полезными для модуляции активности ЛПЛ, лечения заболеваний или нарушений, связанных с метаболизмом липидов, или лечения заболеваний или нарушений, связанных с метаболизмом глюкозы, включая любое из нарушений, перечисленных выше. Неограничивающие примеры дополнительных терапевтических агентов, которые можно комбинировать с заявленными соединениями, включают, но не ограничиваются ими, антидиабетические агенты, такие как инсулин или аналоги инсулина, бигуаниды, сульфонилмочевины, тиазолидиндионы, ингибиторы дипептидилпептидазы-4 («ДПП-4», «DPP- 4») или ингибиторы натрий-зависимого переносчика глюкозы (SGLT2); инкретиновые соединения, такие как глюкагоноподобный пептид-1 (GLP-1) или аналоги GLP-1, желудочный ингибиторный полипептид (GIP) или аналоги GIP, оксинтомодулин (OXM) или аналоги OXM; аспирин; антитромбоцитарные средства; блокаторы Н2-рецепторов; ингибиторы протонной помпы; гипотензивные средства; липид-модифицирующие препараты, такие как ингибиторы редуктазы HMG-CoA, ингибиторы PCSK9, ингибиторы абсорбции холестерина, фибраты, ниацин, агонисты LXR, агонисты RXR, агонисты ROR или модуляторы обратного транспорта холестерина; препараты для лечения сердечной недостаточности, такие как ингибиторы ACE, ингибиторы рецепторов ангиотензина/неприлизина, ARB или антагонисты B-адренорецепторов; препараты для противовоспалительной терапии; препараты для лечения гипертонии, препараты для лечения фибрилляции предсердий; препараты для лечения нейродегенерации; противоопухолевые препараты; препараты для лечения диабетической кардиомиопатии, диабетической ретинопатии, диабетической нейропатии, диабетической нефропатии, для снижение веса, заживление ран; препараты для лечения нефропатии; препараты для лечения БПА (PAD) или комбинации любых из вышеуказанных агентов. Ат против комплекса ANGPTL3/8 и один или более дополнительных терапевтических агентов могут быть введены либо вместе с помощью одинакового пути доставки и средства, такого как одна пилюля, капсула, таблетка или инъекционная лекарственная форма; либо введены раздельно, либо в одно и тоже время различными средствами доставки или путями; либо введены последовательно.
[61] Получение и Очистка Комплексов ANGPTL3/8
[62] Одним из аспектов раскрытия являются комплексы ANGPTL3/8, которые можно использовать для создания Ат, которое связывается только с комплексом и не связывается отдельно только с ANGPTL3 или только с ANGPTL8. Не смотря на то, что Ат против ANGPTL3 и Ат против ANGPTL8 уже известны, существует проблема в синтезе достаточных количеств функциональных комплексов ANGPTL3/8, в особенности комплексов ANGPTL3/8 человека, для создания Ат против комплекса ANGPTL3/8. Дополнительно или альтернативно такие комплексы ANGPTL3/8 можно использовать в анализах для оценки свойств Ат, направленных на ANGPTL3, ANGPTL8 и/или комплексы ANGPTL3/8.
[63] Таким образом, в раскрытии также описаны способы создания ANGPTL8 путем превращения его в N- или C-концевой сывороточный альбумин (например, человека, мыши или кролика). Функциональные комплексы ANGPTL3/8 затем могут быть получены путем коэкспрессии слитого белка ANGPTL8 с нативным слитым белком ANGPTL3 или ANGPTL3 в системе экспрессии млекопитающих.
[64] Как отмечалось выше, известны нуклеотидные и аминокислотные последовательности нативного ANGPTL3 человека и нативного ANGPTL8 человека (см., например, SEQ ID NO:1-2 и 3-4, соответственно). Однако ANGPTL3 или ANGPTL 8, как описано в данном документе, являются модифицированными (т.е. рекомбинантными/синтетическими) и поэтому отличаются от нативных последовательностей содержанием дополнительных аминокислотных последовательностей для улучшения получения, секретирования и/или образования комплексов ANGTPL3 и ANGTPL8.
[65] Например, ANGPTL3 можно модифицировать путем включения в него одного или более линкеров и меток, известных в данной области техники. Как указано в данном документе ANGPTL3 человека (SEQ ID NO:2) модифицирован путем включения линкера и FLAG-метки, так что слитый белок ANGPTL3 имеет аминокислотную последовательность SEQ ID NO:17. В некоторых случаях линкер может содержать от около 1 до около 10 аминокислот, например, 3 аминокислоты, в особенности 3 остатка Ala. Линкер и FLAG-метки могут быть размещены на N- или C-конце последовательности ANGPTL3, в особенности на C-конце, как в SEQ ID NO:17, при этом остатки 1-460 соответствуют ANGPTL3, а остатки 461-471 соответствуют линкеру 3-Ala и FLAG-метке.
[66] Аналогичным образом, ANGPTL8 можно модифицировать путем включения одного или более линкеров и меток в дополнение к последовательности для сывороточного альбумина, в особенности человеческого сывороточного альбумина. Как указано в данном документе ANGPTL8 человека (SEQ ID NO:4) модифицирован путем включения линкера, сигнального пептида каппа-цепи IgG, полигистидиновой (His)-метки, зрелого человеческого сывороточного альбумина человека, линкера и сайта расщепления PreScission®, таким образом, что слитый белок ANGPTL8 имеет аминокислотную последовательность SEQ ID NO:18. В некоторых случаях линкер может содержать от около 1 до около 10 аминокислот, например, жесткий полипролиновый повтор, в особенности линкер Ala-Pro (AP)-10. Сигнальный пептид, His-метка, зрелый ЧСА (HSA), линкер и сайт расщепления PreScission® могут быть размещены на N- или C-конце последовательности ANGPTL8, в особенности на N-конце как в SEQ ID NO:18, при этом остатки 1-20 соответствуют сигнальному пептиду каппа-цепи IgG, остатки 21-27 соответствуют His-метке, остатки 28-612 соответствуют ЧСА, остатки 613-632 соответствуют линкеру AP-10, остатки 633-643 соответствуют сайту расщепления PreScission®, а остатки 644-820 соответствуют ANGPTL8.
[67] Способы конструирования конструкций нуклеиновых кислот для экспрессии слитого белка ANGPTL3 и/или слитого белка ANGPTL8, как описано в данном документе, хорошо известны в данной области техники и могут быть найдены, например, в Balbás & Lorence, “Recombinant Gene Expression: Reviews and Protocols” (2nd Ed. Humana Press 2004); Davis et al., “Basic Methods in Molecular Biology” (Elsevier Press 1986); Sambrook & Russell, “Molecular Cloning: A Laboratory Manual” (3rd Ed. Cold Spring Harbor Laboratory Press 2001); Tijssen, “Laboratory Techniques in Biochemistry and Molecular Biology - Hybridization with Nucleic Acid Probes” (Elsevier 1993); и “Current Protocols in Molecular Biology” (Ausubel et al. eds., Greene Publishing and Wiley-Interscience 1995); а также патентах США № 6,664,387; 7,060,491; 7,345,216 и 7,494,805. Поскольку ANGPTL8 не содержит дисульфидных связей, можно использовать системы экспрессии млекопитающих. Как указано в данном документе системы экспрессии HEK293 или системы экспрессии CHO могут быть использованы для создания слитых белков ANGPTL3 и/или ANGPTL8, в особенности путем коэкспрессии.
[68] Помимо экспрессии слитого белка ANGPTL3 и/или слитого белка ANGPTL8, способы также могут включать очистку полученных слитых белков на основе конкретной метки, используемой для каждого слитого белка, которые хорошо известны в данной области техники. Что касается слитых белков ANGPTL3 и ANGTPL8, описанных в данных документах, очистка может привести к ряду усечений после удаления меток, таким образом, что слитый белок ANGPTL3 имеет аминокислотную последовательность SEQ ID NO:19 (при этом остатки 1-444 соответствуют ANGPTL3, а остатки 445-455 соответствуют линкеру 3-Ala и FLAG-метке), а слитый белок ANGPTL8 имеет аминокислотную последовательность SEQ ID NO:20 (при этом остатки 1-4 соответствуют остаткам расщепления из сайта расщепления PreScission®, а остатки 5-182 соответствуют фрагменту ANGPTL8), которые легко связываются друг с другом с образованием функциональных комплексов ANGPTL3/8.
[69] Получение и Очистка Антител
[70] Ат против комплекса ANGPTL3/8 могут быть получены в системе экспрессии клеток млекопитающих с использованием линии клеток CHO - GSKO. Нокаут гена глутаминсинтетазы (ГС, GS) обеспечивает жесткую строгость отбора путем устранения эндогенной фоновой активности ГС, которая может позволить выживание низко- или непроизводительных клеток в условиях отбора. Гены, кодирующие HC и LC Aт, описанного в данном документе, могут быть субклонированы в отдельные ГС-содержащие плазмиды экспрессии для котрансфекции, или обе цепи могут быть субклонированы в одну ГС-содержащую экспрессионную плазмиду. Последовательность кДНК, кодирующая HC или LC, слита в рамке считывания с кодирующей последовательностью сигнального пептида, который может представлять собой лидерную последовательность каппа-цепи мыши, для усиления секреции желаемого продукта в среду для культивирования клеток. Экспрессия управляется вирусным промотором цитомегаловируса (ЦМВ, CMV).
[71] Клетки CHO - GSKO - стабильно трансфицируют с использованием электропорации и соответствующего количества рекомбинантной плазмиды для экспрессии HC и LC, и трансфицированные клетки поддерживают в суспензионной культуре при подходящей плотности клеток. Отбор трансфицированных клеток осуществляют путем выращивания без глутамина в бессывороточной среде, содержащей 25 мкМ метионинсульфоксимина (МСК, MSX), и инкубируют при 32°C-37°C и 5-7% CO2. Ат секретируются в среду из клеток CHO. Ат могут быть очищены с помощью аффинной хроматографии с протеином A с последующей анионообменной хроматографией или хроматографией с гидрофобным взаимодействием (или другими подходящими способами), и можно использовать эксклюзионную хроматографию для дальнейшей очистки.
[72] Ат из собранной среды захватываются на смоле MabSelect SuRe Protein A (GE Healthcare). Затем смолу быстро промывают рабочим буфером, таким как фосфатно-солевой буфер (ФСБ, PBS) pH 7,4 или рабочим буфером, содержащим Трис, для удаления неспецифически связанного материала. Затем Ат элюируют из смолы раствором с низким pH, таким как 20 мМ уксусная кислота/5 мМ лимонная кислота. Фракции, содержащие Ат против ANGPTL3/8, объединяют и могут удерживаться при низком pH для инактивации потенциальных вирусов. Уровень pH может быть увеличен, по мере необходимости, добавлением основания, такого как 0,1 М Трис pH 8,0. Ат против ANGPTL3/8 могут быть дополнительно очищены с помощью хроматографии с гидрофобным взаимодействием (HIC) с использованием смол, таких как Phenyl HP (GE Healthcare). Ат против ANGPTL3/8 могут быть элюированы из колонки HIC с использованием градиента сульфата натрия в 20 мМ Трис, pH 8,0. Ат против ANGPTL3/8 могут быть дополнительно очищены с помощью эксклюзионной хроматографии с использованием колонки Superdex 200 (GE Healthcare) с изократическим элюированием в ФСБ, pH 7,4.
[73] Описанные в данном документе соединения получают данным или подобным способом, что может быть легко определено специалистом в данной области техники.
[74] Мышей, несущих вариабельные домены легкой и тяжелой цепей человека, иммунизируют комплексом ANGPTL3/8, описанным выше, и выделяют отдельные антиген-специфические В-клетки с помощью FACS с использованием ANGPTL3/8 (положительный) и ANGPTL3 (отрицательный) в качестве маркеров. ДНК вариабельной области тяжелой цепи и легкой цепи выделяют из отдельных В-клеток с помощью ПЦР и клонируют в векторах экспрессии IgG. После трансфекции супернатанты клеток СНО исследуют на связывающую активность.
ПРИМЕРЫ
[75] Нижеследующие неограничивающие примеры предложены в целях иллюстрации, а не ограничения.
[76] Пример 1: Создание Комплексов ANGPTL3/8
[77] Экспрессия ANGPTL3 и ANGPTL8: нуклеотидные последовательности, кодирующие аминокислотную последовательность ANGPTL3 человека, линкер и FLAG-метку (SEQ ID NO:17) вставляют в вектор экспрессии млекопитающих, содержащий промотор ЦМВ. Аналогичным образом, нуклеотидные последовательности, кодирующие аминокислотную последовательность ANGTPL8 человека, сигнальный пептид каппа-цепи IgG мыши, HIS-метку, сайт расщепления зрелого человеческого сывороточного альбумина (ЧСА, HSA)-PreScission® (SEQ ID NO:18) вставляют в вектор экспрессии млекопитающих, содержащий промотор ЦМВ. Экспрессия белка осуществляется посредством временной котрансфекции обоих описанных выше векторов экспрессии в клетках HEK293F, культивируемых в бессывороточной среде. Культуральную среду собирают через 5 дней после трансфекции и хранят при 4°C для последующей очистки белка.
[78] Очистка белка: очистку белка проводят при 4°C, при этом 4 л культуральной среды дополняют 1 M Трис-HCl (pH 8,0) и 5 M NaCl до конечной концентрации 25 мМ и 150 мМ, соответственно. Затем среду инкубируют со 150 мл смолы Ni-NTA (Qiagen) в течение ночи. Смолу набивают в колонку и промывают Буфером А (50 мМ Трис-HCl, pH 8,0, 0,3 М NaCl). Белок элюируют градиентом имидазола 0-300 мМ в буфере A. Фракции, содержащие HIS-ЧСА-ANGPTL8/ANGPTL3-Метка, объединяют, концентрируют и загружают в колонку HiLoad® Superdex® 200 (GE Healthcare Biosciences) и элюируют буфером A. Фракции, содержащие HIS-ЧСА-ANGPTL8/ANGPTL3-Метка, снова объединяют, концентрируют и расщепляют протеазой PreScission® (GE Healthcare Biosciences) для удаления ЧСА из слитого белка HIS-ЧСА-ANGPTL8. Образец белка, расщепленного PreScission®, загружают в колонку HiLoad® Superdex® 200 и элюируют буфером для хранения (20 мМ HEPES, pH 8,0, 150 мМ NaCl). Фракции, содержащие комплекс ANGPTL3/8, объединяют и концентрируют, при этом концентрацию белка определяют с использованием способа Брэдфорда. Комплекс ANGTPL3/8 делят на аликвоты и хранят при -80°C до дальнейшего использования.
[79] Анализ активности ЛПЛ. Анализ EnzCheck® LPL проводят в соответствии с инструкциями производителя (ThermoFisher Scientific). Вкратце, комплексы ANGPTL3 и ANGPLT3/8 серийно разводят в питательной среде и заменяют среду ЛПЛ-клетками в течение 1 часа инкубации. Затем к ЛПЛ-экспрессирующим клеткам добавляют липазный субстрат EnzCheck® (ELS) и инкубируют в течение 30 минут. Флуоресценцию измеряют при 482 нм/515 нм (возбуждение/испускание, соответственно). Рассчитывают % ингибирующего воздействия ANGPTL3 и 3/8 на LPL.
[80] Результаты:
[81] В Таблице 1 представлен выход комплексного белка ANGTPL3/8 на различных стадиях очистки/концентрирования.
[82] Таблица 1: Выход Комплексного Белка ANGPTL3/8.
[83] Подобным образом на Фиг. 1 представлен 4-20% TG TGX гель, окрашенный Кумасси, с нанесенным образцом очищенного и концентрированного комплекса ANGPTL3/8, полученного из колонок, что подтверждает, что комплексы формируются/собираются. После очистки и концентрирования ANGTL3 имеет аминокислотную последовательность SEQ ID NO:19, а ANGPTL8 имеет аминокислотную последовательность SEQ ID NO:20.
[84] В анализе ЛПЛ EC50 ANGPTL3 составляет 21,5 нМ, тогда как EC50 комплекса ANGTPL3/8 составляет 0,54 нМ, что подтверждает функциональность комплексов.
[85] Пример 2: Анализы
[86] Анализ ИФА с одноточечной калибровкой (ИФАОТК, SPE): селективность связывания Aт с комплексом ANGPTL3/8, свободным ANGPTL3 или свободным ANGPTL8 первоначально проверяется с использованием стандартных анализов ИФА в одноточечном формате. Вкратце, аналитические планшеты покрывают Ат против Fc человека в концентрации 2 мкг/мл и затем блокируют казеином. Затем IgG, секретируемый в супернатант после экспрессии в клетках СНО, захватывается на аналитическом планшете. Биотинилированный антиген добавляют в концентрации 25 нМ, чтобы обеспечить связывание Ат/антиген. Комплексы Aт/антиген детектируют после добавления нейтравидина, конъюгированного с щелочной фосфатазой, и субстрата щелочной фосфатазы и последующего измерения оптической плотности при 560 нм. Положительное связывание определяется сигналом, более чем в 3 раза превышающим фоновый сигнал, характерный при отсутствии связывания.
[87] Анализ ИФА: селективность связывания Aт с комплексом ANGPTL3/8, свободным ANGPTL3 или свободным ANGPTL8 проверяется с использованием стандартных анализов ИФА. Вкратце, аналитические планшеты покрывают Ат против Fc человека в концентрации 2 мкг/мл и затем блокируют казеином. Затем Ат из супернатанта клеток СНО, после экспрессии в клетках СНО, захватывается на аналитическом планшете, и в результате получается концентрация Ат, равная 2 мкг/мл. Биотинилированный антиген (ANGPTL3, ANGPTL8 или комплекс ANGPTL3/8) добавляют в различных концентрациях путем серийных разведений, чтобы обеспечить связывание Aт/антиген. Комплексы Aт/антиген детектируют после добавления нейтравидина, конъюгированного с щелочной фосфатазой, и субстрата щелочной фосфатазы и последующего измерения оптической плотности при 560 нм.
[88] Таблица 2: Селективность Связывания Aт с Комплексом ANGPTL3/8, ANGPTL8 и ANGPTL3 в Анализе ИФАОТК.
Значения представляют собой оптическую плотность (OD) при 560 нм, а фоновый сигнал, характерный при отсутствии связывания, по всем исследуемым планшетам в среднем отражается в строке «Фоновый сигнал». Этот отрицательный контроль демонстрирует фоновый сигнал в отсутствии Aт.
[89] В анализе ИФАОТК Aт, имеющее LC с последовательностью SEQ ID NO:5 и HC последовательностью SEQ ID NO:6, демонстрирует положительное связывание с комплексом ANGPTL3/8 человека и отрицательное связывание с ANGPTL8 человека и ANGPTL3 человека. Ат с вариантами LC, имеющими мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22), мутацию D33T (SEQ ID NO:24), мутацию M56T (SEQ ID NO:24) или мутацию E99Q (SEQ ID NO:25) и HC с последовательностью SEQ ID NO:6, аналогичным образом демонстрируют связывание с комплексом ANGPTL3/8 человека и отрицательное связывание с ANGPTL8 человека и ANGPTL3 человека.
[90] Таблица 3: Анализ ИФА Связывания Ат при Различных Концентрациях ANGPTL3, ANGPTL8 и Комплекса ANGPTL3/8.
Контроль представляет собой буфер без Aт.
[91] Ат, имеющее LC с последовательность SEQ ID NO:5 и HC с последовательностью SEQ ID NO:6, демонстрирует положительное связывание с комплексом ANGPTL3/8 человека в зависимости от его концентрации и отсутствие детектируемого связывания с ANGPTL8 человека и ANGPTL3 человека вплоть до концентрации антигена 100 нМ в данном анализе (Таблица 3).
[92] В дополнение к вышеупомянутому Aт против комплекса ANGPTL3/8, имеющего LC с последовательностью SEQ ID NO:5 и HC с последовательностью SEQ ID NO:6, Aт с вариантами LC, имеющими мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22) или мутацию E99Q (SEQ ID NO:25) и HC с последовательностью SEQ ID NO:6, исследуются и аналогичным образом демонстрируют положительное связывание с комплексом ANGPTL3/8 человека в зависимости от концентрации и не демонстрируют связывание с ANGPTL8 человека и ANGPTL3 человека вплоть до концентрации антигена 100 нМ в данном анализе (Таблицы 4-6).
[93] Таблица 4: Анализ ИФА Связывания варианта Ат D31S при Различных Концентрациях ANGPTL3, ANGPTL8 и Комплекса ANGPTL3/8.
[94] Таблица 5: Анализ ИФА Связывания варианта Ат D33A при Различных Концентрациях ANGPTL3, ANGPTL8 и Комплекса ANGPTL3/8.
[95] Таблица 6: Анализ ИФА Связывания варианта Ат E99Q при Различных Концентрациях ANGPTL3, ANGPTL8 и Комплекса ANGPTL3/8.
[96] Пример 3: Аффинность Рецептора In Vitro
[97] Кинетику связывания можно определить с использованием прибора Biacore® T200 (GE Healthcare Bio-Sciences Corp.; Пискатауэй, Нью-Джерси). Поверхность сенсорного чипа CM4 может быть подготовлена ковалентным связыванием человеческого Fab Binder (GE Healthcare Bio-Sciences Corp.). Кинетические эксперименты можно проводить при температуре около 25°C в рабочем буфере HBSEP+, 0,01% БСА при pH 7,4. Aт могут быть захвачены на поверхности чипа, и серии концентраций комплекса ANGPTL3/8 мыши, яванской макаки или человека могут быть введены на поверхность чипа со скоростью около 50 мкл/мин в течение около 240 секунд с временем диссоциации около 800 секунд. Для определения кинетических параметров (например, ka, kd, KD) данные были подвергнуты двойной нормализации и приведены в соответствие модели связывания 1:1 с использованием оценочного программного обеспечения Biacore T200 (GE Healthcare Bio-Sciences Corp.). В Таблице 7 ниже представлено связывание Aт против комплекса ANGPTL3/8, имеющего LC с последовательностью SEQ ID NO:5 и HC с последовательностью SEQ ID NO:6, с различными комплексами ANGPTL3/8 из разных видов при pH 7,4 и температуре 25°С.
[98] Таблица 7: Связывание Aт с Кросс-Видовыми Комплексами ANGPTL3/8 при pH 7,4 и Температуре 25°С.
[99] В дополнение к вышеупомянутому Aт против комплекса ANGPTL3/8, имеющего LC с последовательностью SEQ ID NO:5 и HC с последовательностью SEQ ID NO:6, Ат с вариантами LC, имеющими мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22) или мутацию E99Q (SEQ ID NO: 25) и HC с последовательностью SEQ ID NO:6, исследовали на связывание с различными комплексами ANGPTL3/8 из разных видов при pH 7,4 и температуре 25°C (Таблицы 8-10).
[100] Таблица 8: Связывание Варианта Aт D31S с Кросс-Видовыми Комплексами ANGPTL3/8 при pH 7,4 и Температуре 25°С.
[101] Таблица 9: Связывание Варианта Aт D33A с Кросс-Видовыми Комплексами ANGPTL3/8 при pH 7,4 и Температуре 25°С.
[102] Таблица 10: Связывание Варианта Aт E99Q с Кросс-Видовыми Комплексами ANGPTL3/8 при pH 7,4 и Температуре 25°С.
[103] Пример 4: Анализ Функциональной Активности ЛПЛ In Vitro
[104] Биотест на основе клеток используется для оценки способности Ат против комплекса ANGPTL3/8, имеющего LC с последовательностью SEQ ID NO:5 и HC с последовательностью SEQ ID NO:6, подавлять ингибирование ЛПЛ активности очищенным белком ANGPTL3/8. Ингибирующая активность Aт против ANGPTL3/8 определялась с использованием липазного субстрата EnzChek™ (ThermoFisher). Использовались клеточные линии HEK293, экспрессирующие ЛПЛ человека, яванской макаки, мыши или крысы, и очищенный белок ANGPTL3/8 человека, яванской макаки, мыши или крысы. Генерация HEK293-ЛПЛ включает линию эмбриональных клеток человека, стабильно экспрессирующую ЛПЛ человека, которую получают (HEK293-huLPL) с использованием стандартных способов. Вкратце, ЛПЛ человека клонируют в плазмиду лентивируса с промотором ЦМВ и устойчивостью к бластицидину. Плазмиду используют для создания лентивируса ЛПЛ человека с использованием смеси ViraPower Packaging Mix (Invitrogen). Клетки HEK293 инкубируют с лентивирусом ЛПЛ и отбирают клоны, устойчивые к бластицидину. Клон выбирают после подтверждения экспрессии мРНК ЛПЛ человека с помощью количественной ПЦР и активности ЛПЛ с помощью использования субстрата EnzChek™. Этот процесс повторяется для ЛПЛ яванской макаки, мыши и крысы.
[105] Способы модифицированы по сравнению со способами, описанными в Basu et al. (Basu et al. (2011) J. Lipid Res. 52:826-832): (a) Клетки HEK293-ЛПЛ добавляют на половину площади 96-луночного планшета, покрытого Поли-D-Лизином A (человека), B (яванской макаки), C (мыши) или D (крысы) при плотности 25000 клеток/лунку и инкубируют в течение ночи при 37°С, 5% СО2. Aт серийно разводят девять раз от исходной концентрации стокового раствора для получения десятиточечной CRC, а затем добавляют к очищенным белкам ANGPTL3/8 (при концентрации IC80 0,42 нМ для человека, 0,38 нМ для яванской макаки, 0,13 нМ для мыши или 0,81 нМ для крысы) в 96-луночных планшетах E (человек), F (яванская макака), G (мышь) или H (крыса).
[106] Среды для клеток HEK293-ЛПЛ в планшетах A, B, C и D заменяют смесями ANGPTL3/8 и Ат из планшета E, F, G и H, соответственно, и инкубируют 1 час при 37°С, 5% СО2. 10 мкл субстрата липазы EnzChek™ (приготовленного в концентрации 5 мкМ в 0,05% Zwittergent (3-(N, N-диметилоктадециламмонио)пропансульфонат) (Sigma)) добавляют к клеткам, смеси ANGPTL3/8 и Ат в планшетах A, B, C и D. Планшетный анализатор используется для измерения флуоресценции при длине волны Возбуждения 482 нм и Испускания 515 нм с ограничивающим фильтром 495 нм. Между временными точками планшеты инкубируют при 37°C, 5% CO2. Относительная флуоресценция (прямо пропорциональная активности ЛПЛ) рассчитывается путем вычитания сигнала на 1 мин из сигнала на 31 мин. Эффективная концентрация, при которой Ат восстанавливает активность ЛПЛ на 50% (EC50), рассчитывается с использованием Excel Fit. Концентрации ЕС50 представлены в Таблице 11.
[107] Процент дерепрессии рассчитывается следующим образом:
= (RFU - RFU(MIN))/(RFU(MAX) - RFU(MIN)) x 100,
где MAX=только клетки (ЛПЛ) и MIN=клетки (ЛПЛ) + ANGPTL3/8 КС (кондиционированная среда).
[108] Ат обычно имеет низкий уровень EC50 и, таким образом, способно подавлять дерепрессировать ЛПЛ. Aт также имеет подходящий % максимальной дерепрессии ЛПЛ.
[109] Таблица 11: EC50 Ат и % Максимальной Дерепрессии ЛПЛ для ЛПЛ Человека, Яванской Макаки, Мыши и Крысы, а также Комплекса ANGPTL3/8 Человека, Яванской Макаки, Мыши и Крысы.
[110] В дополнение к вышеупомянутому Aт против комплекса ANGPTL3/8, имеющего LC с последовательностью SEQ ID NO:5 и HC с последовательностью SEQ ID NO:6, Ат с вариантами LC, имеющими мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22) мутацию D33T (SEQ ID NO:23) или мутацию E99Q (SEQ ID NO: 25) и HC с последовательностью SEQ ID NO:6, исследовали на способность подавлять ингибирование ЛПЛ активности очищенным белком ANGPTL3/8 (Таблица 12).
[111] Таблица 12: EC50 Варианта Aт и % Макс. Дерепрессии ЛПЛ для ЛПЛ Человека и Мыши и Комплекса ANGPTL3/8 Человека и Мыши.
EC50 Ат
(нМ),
% Макс. Дерепрессии ЛПЛ
EC50 Ат
(нМ),
% Макс. Дерепрессии ЛПЛ
EC50 Ат
(нМ),
% Макс. Дерепрессии ЛПЛ
EC50 Ат
(нМ),
% Макс. Дерепрессии ЛПЛ
[112] Пример 5: Ответ Триглицеридов In Vivo
[113] Воздействие Ат против комплекса ANGPTL3/8, имеющего LC с последовательностью SEQ ID NO:5 и HC с последовательностью SEQ ID NO:6, на уровень ТГ в сыворотке оценивается на мышах, трансгенных по huCETP и huApolipoprotein A1 (a). У мышей берут кровь и выделяют сыворотку перед началом эксперимента. ТГ измеряется в образцах сыворотки с помощью химического анализатора Cobas® (Roche). Животные были разделены на 5 групп по 20 особей в каждой, чтобы получить группы с аналогичными средними значениями ТГ в сыворотке. Затем каждую группу из 20 особей подразделяют на 4 группы по 5 особей с аналогичными средними значениями триглицеридов в сыворотке. Aт вводят мышам путем однократной подкожной инъекции в дозе 1 мг/кг (n=20), 3 мг/кг (n=20), 10 мг/кг (n=20) или 30 мг/кг (n=20) 4 отдельным группам животных. Контрольное антигенсвязывающее мАт, соответствующее нуль-изотипу, вводят путем однократной подкожной инъекции в дозе 30 мг/кг (n=20) пятой группе животных.
[114] Забор крови у животных делают через 1 час (n=5), 8 часов (n=5), 1 день (n=5), 2 дня (n=5), 3 дня (n=5), 7 дней (n=5), 14 дней (n=5) и 21 день (n=5) после введения Ат. Кровь отбирают у животных подгруппы А через 1 час и 3 дня. Кровь отбирают у животных подгруппы B через 8 часов и 7 дней. Кровь отбирают у животных подгруппы C через 1 день и 14 дней. Кровь отбирают у животных подгруппы D через 2 дня и 21 день. Сыворотку готовили из крови, и уровни ТГ в сыворотке измеряли с помощью клинического химического анализатора Cobas® (Roche). Процентное изменение ТГ от согласованного по времени изотипического контроля рассчитывают для каждой дозы Ат в каждый момент времени. Расчет процентного изменения составляет [(Rx триглицерид сыворотки - триглицерид сыворотки изотипического контроля согласованного по времени)/(триглицерид сыворотки изотипического контроля согласованного по времени)] x 100. Данные представлены в Таблице 13.
[115] Таблица 13: Процент Снижения Уровня ТГ с Помощью Ат По Сравнению с Контролем IgG При Различных Дозах Ат в Разные Моменты Времени.
Тест Даннета использовался для каждого набора данных для сравнения каждой экспериментальной группы с согласованным по времени контролем, и значение p < 0,05 считалось статистически значимым. Группы, статистически значимые по сравнению с соответствующими по времени контролями, обозначены в таблице знаком (*).
[116] Эффективность Ат, имеющего LC с последовательностью SEQ ID NO:5 и HC с последовательностью SEQ ID NO:6, при снижении ТГ является подходящей, начиная с Дня 1, и благоприятное влияние сохраняется до Дня 21.
[117] В дополнение к вышеупомянутому Ат против комплекса ANGPTL3/8, имеющего LC с последовательностью SEQ ID NO: 5 и HC с последовательностью SEQ ID NO:6, Ат с вариантами LC, имеющими мутацию D31S (SEQ ID NO:21), мутацию D33A (SEQ ID NO:22) или мутации E99Q (SEQ ID NO:25) и HC с последовательностью SEQ ID NO:6, также исследовались для оценки изменения уровня ТГ по сравнению с контролем IgG у мышей (Таблица 14).
[118] Таблица 14: Процент Снижения Уровня ТГ По Сравнению с Контролем IgG При Различных Дозах Вариантов Ат в Разные Моменты Времени.
Тест Даннета использовался для каждого набора данных для сравнения каждой экспериментальной группы с согласованным по времени контролем, и значение p < 0,05 считалось статистически значимым. Группы, статистически значимые по сравнению с соответствующими по времени контролями, обозначены в таблице знаком (*).
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[119] Нижеследующие нуклеотидные и аминокислотные последовательности упоминаются в раскрытии выше и приводятся ниже для справки.
[120] SEQ ID NO:1
atatatagagttaagaagtctaggtctgcttccagaagaaaacagttccacgttgcttgaaattgaaaatcaagataaaaatgttcacaattaagctccttctttttattgttcctctagttatttcctccagaattgatcaagacaattcatcatttgattctctatctccagagccaaaatcaagatttgctatgttagacgatgtaaaaattttagccaatggcctccttcagttgggacatggtcttaaagactttgtccataagacgaagggccaaattaatgacatatttcaaaaactcaacatatttgatcagtctttttatgatctatcgctgcaaaccagtgaaatcaaagaagaagaaaaggaactgagaagaactacatataaactacaagtcaaaaatgaagaggtaaagaatatgtcacttgaactcaactcaaaacttgaaagcctcctagaagaaaaaattctacttcaacaaaaagtgaaatatttagaagagcaactaactaacttaattcaaaatcaacctgaaactccagaacacccagaagtaacttcacttaaaacttttgtagaaaaacaagataatagcatcaaagaccttctccagaccgtggaagaccaatataaacaattaaaccaacagcatagtcaaataaaagaaatagaaaatcagctcagaaggactagtattcaagaacccacagaaatttctctatcttccaagccaagagcaccaagaactactccctttcttcagttgaatgaaataagaaatgtaaaacatgatggcattcctgctgaatgtaccaccatttataacagaggtgaacatacaagtggcatgtatgccatcagacccagcaactctcaagtttttcatgtctactgtgatgttatatcaggtagtccatggacattaattcaacatcgaatagatggatcacaaaacttcaatgaaacgtgggagaactacaaatatggttttgggaggcttgatggagaattttggttgggcctagagaagatatactccatagtgaagcaatctaattatgttttacgaattgagttggaagactggaaagacaacaaacattatattgaatattctttttacttgggaaatcacgaaaccaactatacgctacatctagttgcgattactggcaatgtccccaatgcaatcccggaaaacaaagatttggtgttttctacttgggatcacaaagcaaaaggacacttcaactgtccagagggttattcaggaggctggtggtggcatgatgagtgtggagaaaacaacctaaatggtaaatataacaaaccaagagcaaaatctaagccagagaggagaagaggattatcttggaagtctcaaaatggaaggttatactctataaaatcaaccaaaatgttgatccatccaacagattcagaaagctttgaatgaactgaggcaaatttaaaaggcaataatttaaacattaacctcattccaagttaatgtggtctaataatctggtattaaatccttaagagaaagcttgagaaatagattttttttatcttaaagtcactgtctatttaagattaaacatacaatcacataaccttaaagaataccgtttacatttctcaatcaaaattcttataatactatttgttttaaattttgtgatgtgggaatcaattttagatggtcacaatctagattataatcaataggtgaacttattaaataacttttctaaataaaaaatttagagacttttattttaaaaggcatcatatgagctaatatcacaactttcccagtttaaaaaactagtactcttgttaaaactctaaacttgactaaatacagaggactggtaattgtacagttcttaaatgttgtagtattaatttcaaaactaaaaatcgtcagcacagagtatgtgtaaaaatctgtaatacaaatttttaaactgatgcttcattttgctacaaaataatttggagtaaatgtttgatatgatttatttatgaaacctaatgaagcagaattaaatactgtattaaaataagttcgctgtctttaaacaaatggagatgactactaagtcacattgactttaacatgaggtatcactataccttatttgttaaaatatatactgtatacattttatatattttaacacttaatactatgaaaacaaataattgtaaaggaatcttgtcagattacagtaagaatgaacatatttgtggcatcgagttaaagtttatatttcccctaaatatgctgtgattctaatacattcgtgtaggttttcaagtagaaataaacctcgtaacaagttactgaacgtttaaacagcctgacaagcatgtatatatgtttaaaattcaataaacaaagacccagtccctaaattatagaaatttaaattattcttgcatgtttatcgacatcacaacagatccctaaatccctaaatccctaaagattagatacaaattttttaccacagtatcacttgtcagaatttatttttaaatatgattttttaaaactgccagtaagaaattttaaattaaacccatttgttaaaggatatagtgcccaagttatatggtgacctacctttgtcaatacttagcattatgtatttcaaattatccaatatacatgtcatatatatttttatatgtcacatatataaaagatatgtatgatctatgtgaatcctaagtaaatattttgttccagaaaagtacaaaataataaaggtaaaaataatctataattttcaggaccacagactaagctgtcgaaattaacgctgatttttttagggccagaataccaaaatggctcctctcttcccccaaaattggacaatttcaaatgcaaaataattcattatttaatatatgagttgcttcctctatt
[121] SEQ ID NO:2
MFTIKLLLFIVPLVISSRIDQDNSSFDSLSPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTSEIKEEEKELRRTTYKLQVKNEEVKNMSLELNSKLESLLEEKILLQQKVKYLEEQLTNLIQNQPETPEHPEVTSLKTFVEKQDNSIKDLLQTVEDQYKQLNQQHSQIKEIENQLRRTSIQEPTEISLSSKPRAPRTTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTDSESFE
[122] SEQ ID NO:3
ataccttagaccctcagtcatgccagtgcctgctctgtgcctgctctgggccctggcaatggtgacccggcctgcctcagcggcccccatgggcggcccagaactggcacagcatgaggagctgaccctgctcttccatgggaccctgcagctgggccaggccctcaacggtgtgtacaggaccacggagggacggctgacaaaggccaggaacagcctgggtctctatggccgcacaatagaactcctggggcaggaggtcagccggggccgggatgcagcccaggaacttcgggcaagcctgttggagactcagatggaggaggatattctgcagctgcaggcagaggccacagctgaggtgctgggggaggtggcccaggcacagaaggtgctacgggacagcgtgcagcggctagaagtccagctgaggagcgcctggctgggccctgcctaccgagaatttgaggtcttaaaggctcacgctgacaagcagagccacatcctatgggccctcacaggccacgtgcagcggcagaggcgggagatggtggcacagcagcatcggctgcgacagatccaggagagactccacacagcggcgctcccagcctgaatctgcctggatggaactgaggaccaatcatgctgcaaggaacacttccacgccccgtgaggcccctgtgcagggaggagctgcctgttcactgggatcagccagggcgccgggccccacttctgagcacagagcagagacagacgcaggcggggacaaaggcagaggatgtagccccattggggaggggtggaggaaggacatgtaccctttcatgcctacacacccctcattaaagcagagtcgtggcatctcaaaaaaaaaaaaaaaaa
[123] SEQ ID NO:4
MPVPALCLLWALAMVTRPASAAPMGGPELAQHEELTLLFHGTLQLGQALNGVYRTTEGRLTKARNSLGLYGRTIELLGQEVSRGRDAAQELRASLLETQMEEDILQLQAEATAEVLGEVAQAQKVLRDSVQRLEVQLRSAWLGPAYREFEVLKAHADKQSHILWALTGHVQRQRREMVAQQHRLRQIQERLHTAALPA
[124] SEQ ID NO:5
DIVMTQTPLSLPVTPGEPASISCRSSQSLLDSDDGNTYLDWYLQKPGQSPQLLIYMLSYRASGVPDRFSGSGSGTDFTLKISRVEAEDVGIYYCMQRIEFPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
[125] SEQ ID NO:6
QVTLKESGPTLVKPTQTLTLTCTFSGFSLSISGVGVGWIRQPPGKALEWLALIYRNDDKRYSPSLKSRLTITKDTSKNQVVLTLTNMDPVDTATYYCARTYSSGWYGNWFDPWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLG
[126] SEQ ID NO:7
Gatattgtgatgacccagacccccctgtctctgcctgtcactccgggggaaccggcctcgatctcatgccggtcgagccagtccctgctggactccgatgacgggaacacttatttggattggtacctccaaaagcctggacagagcccgcagctcctgatctacatgctgtcctaccgggcctccggagtgccagaccgcttctcgggaagcggctccggtaccgacttcacactgaagatctcccgcgtggaagctgaggacgtgggcatctactactgtatgcaaagaatcgagttccccctcaccttcggcggcgggactaaggtcgagattaagagaaccgtggccgcaccatccgtgttcatttttcccccgtccgatgaacagctgaagtccggaaccgcctccgtcgtgtgcctgctcaacaacttctacccgagggaagcgaaagtgcagtggaaagtggacaatgcgctgcagtccggaaactcccaagagtccgtgaccgaacaggactccaaggactcaacctactcgctgagctcaacgctgaccctgagcaaggccgactacgagaagcacaaggtctacgcctgcgaagtgacccatcagggtttgagctcgcccgtgaccaagtccttcaaccggggagagtgc
[127] SEQ ID NO:8
caagtcacattgaaggagagcggtccgaccctggtcaagccgactcagaccctgaccctgacgtgcactttctcgggcttctcattgtccatttctggagtgggcgtgggatggatcagacagcccccggggaaggccctcgagtggctcgcgctgatctaccgcaacgacgacaaacgctactccccctcactgaaatcccggctgaccatcactaaggatacgtccaagaaccaggtcgtgttgaccctcaccaacatggatcccgtggatactgccacctactattgtgcacggacctatagcagcggttggtacggaaactggttcgacccgtggggccagggaactcttgtgacggtgtcctccgcaagcaccaagggtccttctgtgttccccctggcgccgtgctcgcggagcacctcagagtccaccgccgccctcggctgccttgtgaaggactacttcccggagccagtcaccgtgtcctggaacagcggggccctgacttccggcgtgcacaccttccctgcggtgctgcagagctcaggcctctattcgctgtcatccgtcgtgaccgtgccttcctcgtccctgggcactaagacctacacttgcaacgtggaccataagcccagcaacaccaaagtggacaagagagtggaatccaaatacggaccgccatgtccgccctgccccgccccggaagctgccgggggacccagcgtgttcctgttcccacctaagccgaaggacactctgatgatctcaaggactcccgaagtcacttgcgtggtcgtggacgtgtcccaggaggaccccgaagtccagtttaattggtacgtggatggtgtcgaggtccacaacgccaagaccaagcctcgcgaggaacagttcaattccacctaccgggtcgtgtccgtcctgaccgtgctgcatcaggactggctgaacggaaaggagtacaagtgcaaagtgtccaacaagggactcccttcctccatcgaaaagaccatcagcaaggccaagggccagcctcgcgaaccacaagtctacaccctgcccccatcgcaagaggaaatgaccaagaaccaagtgtcgctgacatgcctcgtcaagggattctacccgtcggatattgcggtggaatgggagtccaacggacagcccgagaacaactacaagaccaccccgccggtgttggactccgacggctcctttttcctgtactcccggctcactgtggacaagtcgcggtggcaggaggggaacgtgttctcctgttccgtgatgcacgaagctctgcacaaccactacacccagaagtcgctgagcctctcactggga
[128] SEQ ID NO:9
DIVMTQTPLSLPVTPGEPASISCRSSQSLLDSDDGNTYLDWYLQKPGQSPQLLIYMLSYRASGVPDRFSGSGSGTDFTLKISRVEAEDVGIYYCMQRIEFPLTFGGGTKVEIK
[129] SEQ ID NO:10
QVTLKESGPTLVKPTQTLTLTCTFSGFSLSISGVGVGWIRQPPGKALEWLALIYRNDDKRYSPSLKSRLTITKDTSKNQVVLTLTNMDPVDTATYYCARTYSSGWYGNWFDPWGQGTLVTVSS
[130] SEQ ID NO:11
RSSQSLLDSDDGNTYLD
[131] SEQ ID NO:12
YMLSYRAS
[132] SEQ ID NO:13
MQRIEFPLT
[133] SEQ ID NO:14
TFSGFSLSISGVGVG
[134] SEQ ID NO:15
LIYRNDDKRYSPSLKS
[135] SEQ ID NO:16
ARTYSSGWYGNWFDP
[136] SEQ ID NO:17
MFTIKLLLFIVPLVISSRIDQDNSSFDSLSPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTSEIKEEEKELRRTTYKLQVKNEEVKNMSLELNSKLESLLEEKILLQQKVKYLEEQLTNLIQNQPETPEHPEVTSLKTFVEKQDNSIKDLLQTVEDQYKQLNQQHSQIKEIENQLRRTSIQEPTEISLSSKPRAPRTTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTDSESFEAAADYKDDDDK
[137] SEQ ID NO:18
METDTLLLWVLLLWVPGSTGDHHHHHHDAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQSPFEDHVKLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKRYKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKERQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQAALGLAPAPAPAPAPAPAPAPAPAPLEVLFQGPGRAAPMGGPELAQHEELTLLFHGTLQLGQALNGVYRTTEGRLTKARNSLGLYGRTIELLGQEVSRGRDAAQELRASLLETQMEEDILQLQAEATAEVLGEVAQAQKVLRDSVQRLEVQLRSAWLGPAYREFEVLKAHADKQSHILWALTGHVQRQRREMVAQQHRLRQIQERLHTAALPA
[138] SEQ ID NO:19
SRIDQDNSSFDSLSPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTSEIKEEEKELRRTTYKLQVKNEEVKNMSLELNSKLESLLEEKILLQQKVKYLEEQLTNLIQNQPETPEHPEVTSLKTFVEKQDNSIKDLLQTVEDQYKQLNQQHSQIKEIENQLRRTSIQEPTEISLSSKPRAPRTTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTDSESFEAAADYKDDDDK
[139] SEQ ID NO:20
GPGRAAPMGGPELAQHEELTLLFHGTLQLGQALNGVYRTTEGRLTKARNSLGLYGRTIELLGQEVSRGRDAAQELRASLLETQMEEDILQLQAEATAEVLGEVAQAQKVLRDSVQRLEVQLRSAWLGPAYREFEVLKAHADKQSHILWALTGHVQRQRREMVAQQHRLRQIQERLHTAALPA
[140] SEQ ID NO:21
DIVMTQTPLSLPVTPGEPASISCRSSQSLLSSDDGNTYLDWYLQKPGQSPQLLIYMLSYRASGVPDRFSGSGSGTDFTLKISRVEAEDVGIYYCMQRIEFPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
[141] SEQ ID NO:22
DIVMTQTPLSLPVTPGEPASISCRSSQSLLDSADGNTYLDWYLQKPGQSPQLLIYMLSYRASGVPDRFSGSGSGTDFTLKISRVEAEDVGIYYCMQRIEFPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
[142] SEQ ID NO:23
DIVMTQTPLSLPVTPGEPASISCRSSQSLLDSTDGNTYLDWYLQKPGQSPQLLIYMLSYRASGVPDRFSGSGSGTDFTLKISRVEAEDVGIYYCMQRIEFPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
[143] SEQ ID NO:24
DIVMTQTPLSLPVTPGEPASISCRSSQSLLDSDDGNTYLDWYLQKPGQSPQLLIYTLSYRASGVPDRFSGSGSGTDFTLKISRVEAEDVGIYYCMQRIEFPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
[144] SEQ ID NO:25
DIVMTQTPLSLPVTPGEPASISCRSSQSLLDSDDGNTYLDWYLQKPGQSPQLLIYMLSYRASGVPDRFSGSGSGTDFTLKISRVEAEDVGIYYCMQRIQFPLTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЭЛИ ЛИЛЛИ ЭНД КОМПАНИ
<120> АНТИТЕЛА ПРОТИВ КОМПЛЕКСА ANGPTL3/8 И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ
<130> X21977
<150> US 62/783,260
<151> 2018-12-21
<150> US 62/783,265
<151> 2018-12-21
<160> 25
<170> PatentIn версия 3.5
<210> 1
<211> 2951
<212> ДНК
<213> Homo sapiens
<400> 1
atatatagag ttaagaagtc taggtctgct tccagaagaa aacagttcca cgttgcttga 60
aattgaaaat caagataaaa atgttcacaa ttaagctcct tctttttatt gttcctctag 120
ttatttcctc cagaattgat caagacaatt catcatttga ttctctatct ccagagccaa 180
aatcaagatt tgctatgtta gacgatgtaa aaattttagc caatggcctc cttcagttgg 240
gacatggtct taaagacttt gtccataaga cgaagggcca aattaatgac atatttcaaa 300
aactcaacat atttgatcag tctttttatg atctatcgct gcaaaccagt gaaatcaaag 360
aagaagaaaa ggaactgaga agaactacat ataaactaca agtcaaaaat gaagaggtaa 420
agaatatgtc acttgaactc aactcaaaac ttgaaagcct cctagaagaa aaaattctac 480
ttcaacaaaa agtgaaatat ttagaagagc aactaactaa cttaattcaa aatcaacctg 540
aaactccaga acacccagaa gtaacttcac ttaaaacttt tgtagaaaaa caagataata 600
gcatcaaaga ccttctccag accgtggaag accaatataa acaattaaac caacagcata 660
gtcaaataaa agaaatagaa aatcagctca gaaggactag tattcaagaa cccacagaaa 720
tttctctatc ttccaagcca agagcaccaa gaactactcc ctttcttcag ttgaatgaaa 780
taagaaatgt aaaacatgat ggcattcctg ctgaatgtac caccatttat aacagaggtg 840
aacatacaag tggcatgtat gccatcagac ccagcaactc tcaagttttt catgtctact 900
gtgatgttat atcaggtagt ccatggacat taattcaaca tcgaatagat ggatcacaaa 960
acttcaatga aacgtgggag aactacaaat atggttttgg gaggcttgat ggagaatttt 1020
ggttgggcct agagaagata tactccatag tgaagcaatc taattatgtt ttacgaattg 1080
agttggaaga ctggaaagac aacaaacatt atattgaata ttctttttac ttgggaaatc 1140
acgaaaccaa ctatacgcta catctagttg cgattactgg caatgtcccc aatgcaatcc 1200
cggaaaacaa agatttggtg ttttctactt gggatcacaa agcaaaagga cacttcaact 1260
gtccagaggg ttattcagga ggctggtggt ggcatgatga gtgtggagaa aacaacctaa 1320
atggtaaata taacaaacca agagcaaaat ctaagccaga gaggagaaga ggattatctt 1380
ggaagtctca aaatggaagg ttatactcta taaaatcaac caaaatgttg atccatccaa 1440
cagattcaga aagctttgaa tgaactgagg caaatttaaa aggcaataat ttaaacatta 1500
acctcattcc aagttaatgt ggtctaataa tctggtatta aatccttaag agaaagcttg 1560
agaaatagat tttttttatc ttaaagtcac tgtctattta agattaaaca tacaatcaca 1620
taaccttaaa gaataccgtt tacatttctc aatcaaaatt cttataatac tatttgtttt 1680
aaattttgtg atgtgggaat caattttaga tggtcacaat ctagattata atcaataggt 1740
gaacttatta aataactttt ctaaataaaa aatttagaga cttttatttt aaaaggcatc 1800
atatgagcta atatcacaac tttcccagtt taaaaaacta gtactcttgt taaaactcta 1860
aacttgacta aatacagagg actggtaatt gtacagttct taaatgttgt agtattaatt 1920
tcaaaactaa aaatcgtcag cacagagtat gtgtaaaaat ctgtaataca aatttttaaa 1980
ctgatgcttc attttgctac aaaataattt ggagtaaatg tttgatatga tttatttatg 2040
aaacctaatg aagcagaatt aaatactgta ttaaaataag ttcgctgtct ttaaacaaat 2100
ggagatgact actaagtcac attgacttta acatgaggta tcactatacc ttatttgtta 2160
aaatatatac tgtatacatt ttatatattt taacacttaa tactatgaaa acaaataatt 2220
gtaaaggaat cttgtcagat tacagtaaga atgaacatat ttgtggcatc gagttaaagt 2280
ttatatttcc cctaaatatg ctgtgattct aatacattcg tgtaggtttt caagtagaaa 2340
taaacctcgt aacaagttac tgaacgttta aacagcctga caagcatgta tatatgttta 2400
aaattcaata aacaaagacc cagtccctaa attatagaaa tttaaattat tcttgcatgt 2460
ttatcgacat cacaacagat ccctaaatcc ctaaatccct aaagattaga tacaaatttt 2520
ttaccacagt atcacttgtc agaatttatt tttaaatatg attttttaaa actgccagta 2580
agaaatttta aattaaaccc atttgttaaa ggatatagtg cccaagttat atggtgacct 2640
acctttgtca atacttagca ttatgtattt caaattatcc aatatacatg tcatatatat 2700
ttttatatgt cacatatata aaagatatgt atgatctatg tgaatcctaa gtaaatattt 2760
tgttccagaa aagtacaaaa taataaaggt aaaaataatc tataattttc aggaccacag 2820
actaagctgt cgaaattaac gctgattttt ttagggccag aataccaaaa tggctcctct 2880
cttcccccaa aattggacaa tttcaaatgc aaaataattc attatttaat atatgagttg 2940
cttcctctat t 2951
<210> 2
<211> 460
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Met Phe Thr Ile Lys Leu Leu Leu Phe Ile Val Pro Leu Val Ile Ser
5 10 15
Ser Arg Ile Asp Gln Asp Asn Ser Ser Phe Asp Ser Leu Ser Pro Glu
20 25 30
Pro Lys Ser Arg Phe Ala Met Leu Asp Asp Val Lys Ile Leu Ala Asn
35 40 45
Gly Leu Leu Gln Leu Gly His Gly Leu Lys Asp Phe Val His Lys Thr
50 55 60
Lys Gly Gln Ile Asn Asp Ile Phe Gln Lys Leu Asn Ile Phe Asp Gln
65 70 75 80
Ser Phe Tyr Asp Leu Ser Leu Gln Thr Ser Glu Ile Lys Glu Glu Glu
85 90 95
Lys Glu Leu Arg Arg Thr Thr Tyr Lys Leu Gln Val Lys Asn Glu Glu
100 105 110
Val Lys Asn Met Ser Leu Glu Leu Asn Ser Lys Leu Glu Ser Leu Leu
115 120 125
Glu Glu Lys Ile Leu Leu Gln Gln Lys Val Lys Tyr Leu Glu Glu Gln
130 135 140
Leu Thr Asn Leu Ile Gln Asn Gln Pro Glu Thr Pro Glu His Pro Glu
145 150 155 160
Val Thr Ser Leu Lys Thr Phe Val Glu Lys Gln Asp Asn Ser Ile Lys
165 170 175
Asp Leu Leu Gln Thr Val Glu Asp Gln Tyr Lys Gln Leu Asn Gln Gln
180 185 190
His Ser Gln Ile Lys Glu Ile Glu Asn Gln Leu Arg Arg Thr Ser Ile
195 200 205
Gln Glu Pro Thr Glu Ile Ser Leu Ser Ser Lys Pro Arg Ala Pro Arg
210 215 220
Thr Thr Pro Phe Leu Gln Leu Asn Glu Ile Arg Asn Val Lys His Asp
225 230 235 240
Gly Ile Pro Ala Glu Cys Thr Thr Ile Tyr Asn Arg Gly Glu His Thr
245 250 255
Ser Gly Met Tyr Ala Ile Arg Pro Ser Asn Ser Gln Val Phe His Val
260 265 270
Tyr Cys Asp Val Ile Ser Gly Ser Pro Trp Thr Leu Ile Gln His Arg
275 280 285
Ile Asp Gly Ser Gln Asn Phe Asn Glu Thr Trp Glu Asn Tyr Lys Tyr
290 295 300
Gly Phe Gly Arg Leu Asp Gly Glu Phe Trp Leu Gly Leu Glu Lys Ile
305 310 315 320
Tyr Ser Ile Val Lys Gln Ser Asn Tyr Val Leu Arg Ile Glu Leu Glu
325 330 335
Asp Trp Lys Asp Asn Lys His Tyr Ile Glu Tyr Ser Phe Tyr Leu Gly
340 345 350
Asn His Glu Thr Asn Tyr Thr Leu His Leu Val Ala Ile Thr Gly Asn
355 360 365
Val Pro Asn Ala Ile Pro Glu Asn Lys Asp Leu Val Phe Ser Thr Trp
370 375 380
Asp His Lys Ala Lys Gly His Phe Asn Cys Pro Glu Gly Tyr Ser Gly
385 390 395 400
Gly Trp Trp Trp His Asp Glu Cys Gly Glu Asn Asn Leu Asn Gly Lys
405 410 415
Tyr Asn Lys Pro Arg Ala Lys Ser Lys Pro Glu Arg Arg Arg Gly Leu
420 425 430
Ser Trp Lys Ser Gln Asn Gly Arg Leu Tyr Ser Ile Lys Ser Thr Lys
435 440 445
Met Leu Ile His Pro Thr Asp Ser Glu Ser Phe Glu
450 455 460
<210> 3
<211> 888
<212> ДНК
<213> Homo sapiens
<400> 3
ataccttaga ccctcagtca tgccagtgcc tgctctgtgc ctgctctggg ccctggcaat 60
ggtgacccgg cctgcctcag cggcccccat gggcggccca gaactggcac agcatgagga 120
gctgaccctg ctcttccatg ggaccctgca gctgggccag gccctcaacg gtgtgtacag 180
gaccacggag ggacggctga caaaggccag gaacagcctg ggtctctatg gccgcacaat 240
agaactcctg gggcaggagg tcagccgggg ccgggatgca gcccaggaac ttcgggcaag 300
cctgttggag actcagatgg aggaggatat tctgcagctg caggcagagg ccacagctga 360
ggtgctgggg gaggtggccc aggcacagaa ggtgctacgg gacagcgtgc agcggctaga 420
agtccagctg aggagcgcct ggctgggccc tgcctaccga gaatttgagg tcttaaaggc 480
tcacgctgac aagcagagcc acatcctatg ggccctcaca ggccacgtgc agcggcagag 540
gcgggagatg gtggcacagc agcatcggct gcgacagatc caggagagac tccacacagc 600
ggcgctccca gcctgaatct gcctggatgg aactgaggac caatcatgct gcaaggaaca 660
cttccacgcc ccgtgaggcc cctgtgcagg gaggagctgc ctgttcactg ggatcagcca 720
gggcgccggg ccccacttct gagcacagag cagagacaga cgcaggcggg gacaaaggca 780
gaggatgtag ccccattggg gaggggtgga ggaaggacat gtaccctttc atgcctacac 840
acccctcatt aaagcagagt cgtggcatct caaaaaaaaa aaaaaaaa 888
<210> 4
<211> 198
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Met Pro Val Pro Ala Leu Cys Leu Leu Trp Ala Leu Ala Met Val Thr
5 10 15
Arg Pro Ala Ser Ala Ala Pro Met Gly Gly Pro Glu Leu Ala Gln His
20 25 30
Glu Glu Leu Thr Leu Leu Phe His Gly Thr Leu Gln Leu Gly Gln Ala
35 40 45
Leu Asn Gly Val Tyr Arg Thr Thr Glu Gly Arg Leu Thr Lys Ala Arg
50 55 60
Asn Ser Leu Gly Leu Tyr Gly Arg Thr Ile Glu Leu Leu Gly Gln Glu
65 70 75 80
Val Ser Arg Gly Arg Asp Ala Ala Gln Glu Leu Arg Ala Ser Leu Leu
85 90 95
Glu Thr Gln Met Glu Glu Asp Ile Leu Gln Leu Gln Ala Glu Ala Thr
100 105 110
Ala Glu Val Leu Gly Glu Val Ala Gln Ala Gln Lys Val Leu Arg Asp
115 120 125
Ser Val Gln Arg Leu Glu Val Gln Leu Arg Ser Ala Trp Leu Gly Pro
130 135 140
Ala Tyr Arg Glu Phe Glu Val Leu Lys Ala His Ala Asp Lys Gln Ser
145 150 155 160
His Ile Leu Trp Ala Leu Thr Gly His Val Gln Arg Gln Arg Arg Glu
165 170 175
Met Val Ala Gln Gln His Arg Leu Arg Gln Ile Gln Glu Arg Leu His
180 185 190
Thr Ala Ala Leu Pro Ala
195
<210> 5
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 5
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Asp Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Leu Leu Ile Tyr Met Leu Ser Tyr Arg Ala Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
65 70 75 80
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Met Gln
85 90 95
Arg Ile Glu Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 6
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 6
Gln Val Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Ile Ser
20 25 30
Gly Val Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Leu Ile Tyr Arg Asn Asp Asp Lys Arg Tyr Ser Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Leu Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Thr Tyr Ser Ser Gly Trp Tyr Gly Asn Trp Phe Asp Pro
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
435 440 445
Gly
<210> 7
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 7
gatattgtga tgacccagac ccccctgtct ctgcctgtca ctccggggga accggcctcg 60
atctcatgcc ggtcgagcca gtccctgctg gactccgatg acgggaacac ttatttggat 120
tggtacctcc aaaagcctgg acagagcccg cagctcctga tctacatgct gtcctaccgg 180
gcctccggag tgccagaccg cttctcggga agcggctccg gtaccgactt cacactgaag 240
atctcccgcg tggaagctga ggacgtgggc atctactact gtatgcaaag aatcgagttc 300
cccctcacct tcggcggcgg gactaaggtc gagattaaga gaaccgtggc cgcaccatcc 360
gtgttcattt ttcccccgtc cgatgaacag ctgaagtccg gaaccgcctc cgtcgtgtgc 420
ctgctcaaca acttctaccc gagggaagcg aaagtgcagt ggaaagtgga caatgcgctg 480
cagtccggaa actcccaaga gtccgtgacc gaacaggact ccaaggactc aacctactcg 540
ctgagctcaa cgctgaccct gagcaaggcc gactacgaga agcacaaggt ctacgcctgc 600
gaagtgaccc atcagggttt gagctcgccc gtgaccaagt ccttcaaccg gggagagtgc 660
<210> 8
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 8
caagtcacat tgaaggagag cggtccgacc ctggtcaagc cgactcagac cctgaccctg 60
acgtgcactt tctcgggctt ctcattgtcc atttctggag tgggcgtggg atggatcaga 120
cagcccccgg ggaaggccct cgagtggctc gcgctgatct accgcaacga cgacaaacgc 180
tactccccct cactgaaatc ccggctgacc atcactaagg atacgtccaa gaaccaggtc 240
gtgttgaccc tcaccaacat ggatcccgtg gatactgcca cctactattg tgcacggacc 300
tatagcagcg gttggtacgg aaactggttc gacccgtggg gccagggaac tcttgtgacg 360
gtgtcctccg caagcaccaa gggtccttct gtgttccccc tggcgccgtg ctcgcggagc 420
acctcagagt ccaccgccgc cctcggctgc cttgtgaagg actacttccc ggagccagtc 480
accgtgtcct ggaacagcgg ggccctgact tccggcgtgc acaccttccc tgcggtgctg 540
cagagctcag gcctctattc gctgtcatcc gtcgtgaccg tgccttcctc gtccctgggc 600
actaagacct acacttgcaa cgtggaccat aagcccagca acaccaaagt ggacaagaga 660
gtggaatcca aatacggacc gccatgtccg ccctgccccg ccccggaagc tgccggggga 720
cccagcgtgt tcctgttccc acctaagccg aaggacactc tgatgatctc aaggactccc 780
gaagtcactt gcgtggtcgt ggacgtgtcc caggaggacc ccgaagtcca gtttaattgg 840
tacgtggatg gtgtcgaggt ccacaacgcc aagaccaagc ctcgcgagga acagttcaat 900
tccacctacc gggtcgtgtc cgtcctgacc gtgctgcatc aggactggct gaacggaaag 960
gagtacaagt gcaaagtgtc caacaaggga ctcccttcct ccatcgaaaa gaccatcagc 1020
aaggccaagg gccagcctcg cgaaccacaa gtctacaccc tgcccccatc gcaagaggaa 1080
atgaccaaga accaagtgtc gctgacatgc ctcgtcaagg gattctaccc gtcggatatt 1140
gcggtggaat gggagtccaa cggacagccc gagaacaact acaagaccac cccgccggtg 1200
ttggactccg acggctcctt tttcctgtac tcccggctca ctgtggacaa gtcgcggtgg 1260
caggagggga acgtgttctc ctgttccgtg atgcacgaag ctctgcacaa ccactacacc 1320
cagaagtcgc tgagcctctc actggga 1347
<210> 9
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 9
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Asp Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Leu Leu Ile Tyr Met Leu Ser Tyr Arg Ala Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
65 70 75 80
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Met Gln
85 90 95
Arg Ile Glu Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 10
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 10
Gln Val Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Ile Ser
20 25 30
Gly Val Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Leu Ile Tyr Arg Asn Asp Asp Lys Arg Tyr Ser Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Leu Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Thr Tyr Ser Ser Gly Trp Tyr Gly Asn Trp Phe Asp Pro
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 11
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 11
Arg Ser Ser Gln Ser Leu Leu Asp Ser Asp Asp Gly Asn Thr Tyr Leu
5 10 15
Asp
<210> 12
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 12
Tyr Met Leu Ser Tyr Arg Ala Ser
5
<210> 13
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 13
Met Gln Arg Ile Glu Phe Pro Leu Thr
5
<210> 14
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 14
Thr Phe Ser Gly Phe Ser Leu Ser Ile Ser Gly Val Gly Val Gly
5 10 15
<210> 15
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 15
Leu Ile Tyr Arg Asn Asp Asp Lys Arg Tyr Ser Pro Ser Leu Lys Ser
5 10 15
<210> 16
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 16
Ala Arg Thr Tyr Ser Ser Gly Trp Tyr Gly Asn Trp Phe Asp Pro
5 10 15
<210> 17
<211> 471
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 17
Met Phe Thr Ile Lys Leu Leu Leu Phe Ile Val Pro Leu Val Ile Ser
5 10 15
Ser Arg Ile Asp Gln Asp Asn Ser Ser Phe Asp Ser Leu Ser Pro Glu
20 25 30
Pro Lys Ser Arg Phe Ala Met Leu Asp Asp Val Lys Ile Leu Ala Asn
35 40 45
Gly Leu Leu Gln Leu Gly His Gly Leu Lys Asp Phe Val His Lys Thr
50 55 60
Lys Gly Gln Ile Asn Asp Ile Phe Gln Lys Leu Asn Ile Phe Asp Gln
65 70 75 80
Ser Phe Tyr Asp Leu Ser Leu Gln Thr Ser Glu Ile Lys Glu Glu Glu
85 90 95
Lys Glu Leu Arg Arg Thr Thr Tyr Lys Leu Gln Val Lys Asn Glu Glu
100 105 110
Val Lys Asn Met Ser Leu Glu Leu Asn Ser Lys Leu Glu Ser Leu Leu
115 120 125
Glu Glu Lys Ile Leu Leu Gln Gln Lys Val Lys Tyr Leu Glu Glu Gln
130 135 140
Leu Thr Asn Leu Ile Gln Asn Gln Pro Glu Thr Pro Glu His Pro Glu
145 150 155 160
Val Thr Ser Leu Lys Thr Phe Val Glu Lys Gln Asp Asn Ser Ile Lys
165 170 175
Asp Leu Leu Gln Thr Val Glu Asp Gln Tyr Lys Gln Leu Asn Gln Gln
180 185 190
His Ser Gln Ile Lys Glu Ile Glu Asn Gln Leu Arg Arg Thr Ser Ile
195 200 205
Gln Glu Pro Thr Glu Ile Ser Leu Ser Ser Lys Pro Arg Ala Pro Arg
210 215 220
Thr Thr Pro Phe Leu Gln Leu Asn Glu Ile Arg Asn Val Lys His Asp
225 230 235 240
Gly Ile Pro Ala Glu Cys Thr Thr Ile Tyr Asn Arg Gly Glu His Thr
245 250 255
Ser Gly Met Tyr Ala Ile Arg Pro Ser Asn Ser Gln Val Phe His Val
260 265 270
Tyr Cys Asp Val Ile Ser Gly Ser Pro Trp Thr Leu Ile Gln His Arg
275 280 285
Ile Asp Gly Ser Gln Asn Phe Asn Glu Thr Trp Glu Asn Tyr Lys Tyr
290 295 300
Gly Phe Gly Arg Leu Asp Gly Glu Phe Trp Leu Gly Leu Glu Lys Ile
305 310 315 320
Tyr Ser Ile Val Lys Gln Ser Asn Tyr Val Leu Arg Ile Glu Leu Glu
325 330 335
Asp Trp Lys Asp Asn Lys His Tyr Ile Glu Tyr Ser Phe Tyr Leu Gly
340 345 350
Asn His Glu Thr Asn Tyr Thr Leu His Leu Val Ala Ile Thr Gly Asn
355 360 365
Val Pro Asn Ala Ile Pro Glu Asn Lys Asp Leu Val Phe Ser Thr Trp
370 375 380
Asp His Lys Ala Lys Gly His Phe Asn Cys Pro Glu Gly Tyr Ser Gly
385 390 395 400
Gly Trp Trp Trp His Asp Glu Cys Gly Glu Asn Asn Leu Asn Gly Lys
405 410 415
Tyr Asn Lys Pro Arg Ala Lys Ser Lys Pro Glu Arg Arg Arg Gly Leu
420 425 430
Ser Trp Lys Ser Gln Asn Gly Arg Leu Tyr Ser Ile Lys Ser Thr Lys
435 440 445
Met Leu Ile His Pro Thr Asp Ser Glu Ser Phe Glu Ala Ala Ala Asp
450 455 460
Tyr Lys Asp Asp Asp Asp Lys
465 470
<210> 18
<211> 820
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 18
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
5 10 15
Gly Ser Thr Gly Asp His His His His His His Asp Ala His Lys Ser
20 25 30
Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala
35 40 45
Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu
50 55 60
Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys
65 70 75 80
Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu
85 90 95
Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly
100 105 110
Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys
115 120 125
Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg
130 135 140
Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr
145 150 155 160
Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe
165 170 175
Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe
180 185 190
Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys
195 200 205
Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg
210 215 220
Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala
225 230 235 240
Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala
245 250 255
Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys
260 265 270
Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala
275 280 285
Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu
290 295 300
Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val
305 310 315 320
Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe
325 330 335
Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val
340 345 350
Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr
355 360 365
Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu
370 375 380
Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val
385 390 395 400
Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys
405 410 415
Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn
420 425 430
Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro
435 440 445
Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys
450 455 460
Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu
465 470 475 480
Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val
485 490 495
Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg
500 505 510
Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu
515 520 525
Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser
530 535 540
Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val
545 550 555 560
Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp
565 570 575
Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu
580 585 590
Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala
595 600 605
Ala Leu Gly Leu Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro
610 615 620
Ala Pro Ala Pro Ala Pro Ala Pro Leu Glu Val Leu Phe Gln Gly Pro
625 630 635 640
Gly Arg Ala Ala Pro Met Gly Gly Pro Glu Leu Ala Gln His Glu Glu
645 650 655
Leu Thr Leu Leu Phe His Gly Thr Leu Gln Leu Gly Gln Ala Leu Asn
660 665 670
Gly Val Tyr Arg Thr Thr Glu Gly Arg Leu Thr Lys Ala Arg Asn Ser
675 680 685
Leu Gly Leu Tyr Gly Arg Thr Ile Glu Leu Leu Gly Gln Glu Val Ser
690 695 700
Arg Gly Arg Asp Ala Ala Gln Glu Leu Arg Ala Ser Leu Leu Glu Thr
705 710 715 720
Gln Met Glu Glu Asp Ile Leu Gln Leu Gln Ala Glu Ala Thr Ala Glu
725 730 735
Val Leu Gly Glu Val Ala Gln Ala Gln Lys Val Leu Arg Asp Ser Val
740 745 750
Gln Arg Leu Glu Val Gln Leu Arg Ser Ala Trp Leu Gly Pro Ala Tyr
755 760 765
Arg Glu Phe Glu Val Leu Lys Ala His Ala Asp Lys Gln Ser His Ile
770 775 780
Leu Trp Ala Leu Thr Gly His Val Gln Arg Gln Arg Arg Glu Met Val
785 790 795 800
Ala Gln Gln His Arg Leu Arg Gln Ile Gln Glu Arg Leu His Thr Ala
805 810 815
Ala Leu Pro Ala
820
<210> 19
<211> 455
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 19
Ser Arg Ile Asp Gln Asp Asn Ser Ser Phe Asp Ser Leu Ser Pro Glu
5 10 15
Pro Lys Ser Arg Phe Ala Met Leu Asp Asp Val Lys Ile Leu Ala Asn
20 25 30
Gly Leu Leu Gln Leu Gly His Gly Leu Lys Asp Phe Val His Lys Thr
35 40 45
Lys Gly Gln Ile Asn Asp Ile Phe Gln Lys Leu Asn Ile Phe Asp Gln
50 55 60
Ser Phe Tyr Asp Leu Ser Leu Gln Thr Ser Glu Ile Lys Glu Glu Glu
65 70 75 80
Lys Glu Leu Arg Arg Thr Thr Tyr Lys Leu Gln Val Lys Asn Glu Glu
85 90 95
Val Lys Asn Met Ser Leu Glu Leu Asn Ser Lys Leu Glu Ser Leu Leu
100 105 110
Glu Glu Lys Ile Leu Leu Gln Gln Lys Val Lys Tyr Leu Glu Glu Gln
115 120 125
Leu Thr Asn Leu Ile Gln Asn Gln Pro Glu Thr Pro Glu His Pro Glu
130 135 140
Val Thr Ser Leu Lys Thr Phe Val Glu Lys Gln Asp Asn Ser Ile Lys
145 150 155 160
Asp Leu Leu Gln Thr Val Glu Asp Gln Tyr Lys Gln Leu Asn Gln Gln
165 170 175
His Ser Gln Ile Lys Glu Ile Glu Asn Gln Leu Arg Arg Thr Ser Ile
180 185 190
Gln Glu Pro Thr Glu Ile Ser Leu Ser Ser Lys Pro Arg Ala Pro Arg
195 200 205
Thr Thr Pro Phe Leu Gln Leu Asn Glu Ile Arg Asn Val Lys His Asp
210 215 220
Gly Ile Pro Ala Glu Cys Thr Thr Ile Tyr Asn Arg Gly Glu His Thr
225 230 235 240
Ser Gly Met Tyr Ala Ile Arg Pro Ser Asn Ser Gln Val Phe His Val
245 250 255
Tyr Cys Asp Val Ile Ser Gly Ser Pro Trp Thr Leu Ile Gln His Arg
260 265 270
Ile Asp Gly Ser Gln Asn Phe Asn Glu Thr Trp Glu Asn Tyr Lys Tyr
275 280 285
Gly Phe Gly Arg Leu Asp Gly Glu Phe Trp Leu Gly Leu Glu Lys Ile
290 295 300
Tyr Ser Ile Val Lys Gln Ser Asn Tyr Val Leu Arg Ile Glu Leu Glu
305 310 315 320
Asp Trp Lys Asp Asn Lys His Tyr Ile Glu Tyr Ser Phe Tyr Leu Gly
325 330 335
Asn His Glu Thr Asn Tyr Thr Leu His Leu Val Ala Ile Thr Gly Asn
340 345 350
Val Pro Asn Ala Ile Pro Glu Asn Lys Asp Leu Val Phe Ser Thr Trp
355 360 365
Asp His Lys Ala Lys Gly His Phe Asn Cys Pro Glu Gly Tyr Ser Gly
370 375 380
Gly Trp Trp Trp His Asp Glu Cys Gly Glu Asn Asn Leu Asn Gly Lys
385 390 395 400
Tyr Asn Lys Pro Arg Ala Lys Ser Lys Pro Glu Arg Arg Arg Gly Leu
405 410 415
Ser Trp Lys Ser Gln Asn Gly Arg Leu Tyr Ser Ile Lys Ser Thr Lys
420 425 430
Met Leu Ile His Pro Thr Asp Ser Glu Ser Phe Glu Ala Ala Ala Asp
435 440 445
Tyr Lys Asp Asp Asp Asp Lys
450 455
<210> 20
<211> 182
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 20
Gly Pro Gly Arg Ala Ala Pro Met Gly Gly Pro Glu Leu Ala Gln His
5 10 15
Glu Glu Leu Thr Leu Leu Phe His Gly Thr Leu Gln Leu Gly Gln Ala
20 25 30
Leu Asn Gly Val Tyr Arg Thr Thr Glu Gly Arg Leu Thr Lys Ala Arg
35 40 45
Asn Ser Leu Gly Leu Tyr Gly Arg Thr Ile Glu Leu Leu Gly Gln Glu
50 55 60
Val Ser Arg Gly Arg Asp Ala Ala Gln Glu Leu Arg Ala Ser Leu Leu
65 70 75 80
Glu Thr Gln Met Glu Glu Asp Ile Leu Gln Leu Gln Ala Glu Ala Thr
85 90 95
Ala Glu Val Leu Gly Glu Val Ala Gln Ala Gln Lys Val Leu Arg Asp
100 105 110
Ser Val Gln Arg Leu Glu Val Gln Leu Arg Ser Ala Trp Leu Gly Pro
115 120 125
Ala Tyr Arg Glu Phe Glu Val Leu Lys Ala His Ala Asp Lys Gln Ser
130 135 140
His Ile Leu Trp Ala Leu Thr Gly His Val Gln Arg Gln Arg Arg Glu
145 150 155 160
Met Val Ala Gln Gln His Arg Leu Arg Gln Ile Gln Glu Arg Leu His
165 170 175
Thr Ala Ala Leu Pro Ala
180
<210> 21
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 21
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Ser Ser
20 25 30
Asp Asp Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Leu Leu Ile Tyr Met Leu Ser Tyr Arg Ala Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
65 70 75 80
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Met Gln
85 90 95
Arg Ile Glu Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 22
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 22
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Ala Asp Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Leu Leu Ile Tyr Met Leu Ser Tyr Arg Ala Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
65 70 75 80
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Met Gln
85 90 95
Arg Ile Glu Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 23
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 23
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Thr Asp Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Leu Leu Ile Tyr Met Leu Ser Tyr Arg Ala Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
65 70 75 80
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Met Gln
85 90 95
Arg Ile Glu Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 24
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 24
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Asp Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Leu Leu Ile Tyr Thr Leu Ser Tyr Arg Ala Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
65 70 75 80
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Met Gln
85 90 95
Arg Ile Glu Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 25
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 25
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Asp Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Leu Leu Ile Tyr Met Leu Ser Tyr Arg Ala Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
65 70 75 80
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Met Gln
85 90 95
Arg Ile Gln Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ПОВРЕЖДЕНИЯ ХРЯЩА И АРТРИТА | 2017 |
|
RU2777504C2 |
| АНТИТЕЛА К PD-1 И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2761640C2 |
| Терапевтические антитела к CD47 | 2016 |
|
RU2748401C2 |
| СКОНСТРУИРОВАННЫЕ PH-ЗАВИСИМЫЕ АНТИТЕЛА К CD3, А ТАКЖЕ СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2832079C2 |
| НОВЫЕ КАПСИДНЫЕ БЕЛКИ АДЕНО-АССОЦИИРОВАННОГО ВИРУСА | 2017 |
|
RU2779318C2 |
| PD1/CTLA4-СВЯЗЫВАЮЩИЕ ВЕЩЕСТВА | 2016 |
|
RU2755724C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2020 |
|
RU2833880C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ИНТЕРЛЕЙКИНА-33 (IL-33) | 2015 |
|
RU2829846C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2018 |
|
RU2809244C2 |
| ПЕПТИД, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) И ЛЕКАРСТВЕННОЕ СРЕДСТВО НА ЕГО ОСНОВЕ, СПОСОБ ЛЕЧЕНИЯ И/ИЛИ ПРЕДУПРЕЖДЕНИЯ ДИАБЕТА ИЛИ ЗАБОЛЕВАНИЙ, СОПУТСТВУЮЩИХ ОЖИРЕНИЮ, СПОСОБ УМЕНЬШЕНИЯ АППЕТИТА, УМЕНЬШЕНИЯ ПОТРЕБЛЕНИЯ ПИЩИ, УМЕНЬШЕНИЯ ПОТРЕБЛЕНИЯ КАЛОРИЙ, УМЕНЬШЕНИЯ МАССЫ ТЕЛА И УВЕЛИЧЕНИЯ РАСХОДА ЭНЕРГИИ | 2007 |
|
RU2492183C2 |
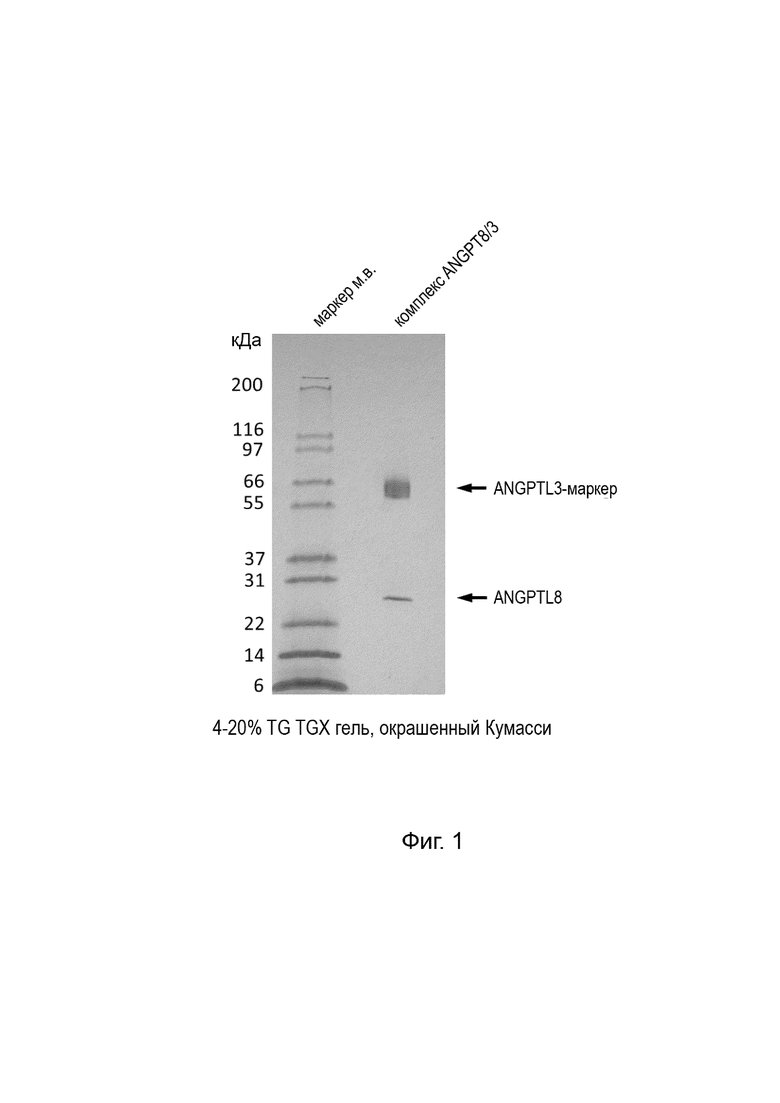
Изобретение относится к области биотехнологии, в частности к антителам, которые связываются с комплексом ANGPTL3/8 человека и тем самым нейтрализуют их; к фармацевтическим композициям, содержащим упомянутые антитела, к клеткам млекопитающих, экспрессирующим антитела к комплексу ANGPTL3/8, к способу получения антител к комплексу ANGPTL3/8, к способу снижения уровня триглицеридов, а также лечения заболеваний, связанных с метаболизмом липидов. Изобретение позволяет повышать активность липопротеинлипазы и тем самым эффективно снижать уровень триглицеридов, что в дальнейшем может быть использовано для лечения заболеваний и нарушений, связанных с метаболизмом липидов. 20 н. и 16 з.п. ф-лы, 1 ил., 14 табл., 5 пр.
1. Антитело, которое связывается с комплексом ANGPTL3/8 человека, содержащее определяющие комплементарность области легкой цепи, LCDR1, LCDR2 и LCDR3 и определяющие комплементарность области тяжелой цепи, HCDR1, HCDR2 и HCDR3, где
LCDR1 имеет аминокислотную последовательность RSSQSLLDSDDGNTYLD (SEQ ID NO: 11),
LCDR2 имеет аминокислотную последовательность YMLSYRAS (SEQ ID NO: 12),
LCDR3 имеет аминокислотную последовательность MQRIEFPLT (SEQ ID NO: 13),
HCDR1 имеет аминокислотную последовательность TFSGFSLSISGVGVG (SEQ ID NO: 14),
HCDR2 имеет аминокислотную последовательность LIYRNDDKRYSPSLKS (SEQ ID NO: 15), и
HCDR3 имеет аминокислотную последовательность ARTYSSGWYGNWFDP (SEQ ID NO: 16).
2. Антитело по п.1, содержащее вариабельную область легкой цепи (LCVR), где LCVR имеет аминокислотную последовательность SEQ ID NO: 9.
3. Антитело по п.1, содержащее вариабельную область тяжелой цепи (HCVR), где HCVR имеет аминокислотную последовательность SEQ ID NO: 10.
4. Антитело по любому из пп.1-3, где
LCVR имеет аминокислотную последовательность SEQ ID NO: 9, и
HCVR имеет аминокислотную последовательность SEQ ID NO: 10.
5. Антитело по любому из пп.1-4, где антитело представляет собой изотип IgG4.
6. Антитело, которое связывается с комплексом ANGPTL3/8 человека, содержащее легкую цепь (LC) и тяжелую цепь (HC), где
LC содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, 21, 22, 23, 24 и 25, и
HC содержит аминокислотную последовательность SEQ ID NO: 6.
7. Антитело по п.6, где
LC имеет аминокислотную последовательность SEQ ID NO: 5, и
HC имеет аминокислотную последовательность SEQ ID NO: 6.
8. Антитело по п.6, где
LC имеет аминокислотную последовательность SEQ ID NO: 21, и
HC имеет аминокислотную последовательность SEQ ID NO: 6.
9. Антитело по п.6, где
LC имеет аминокислотную последовательность SEQ ID NO: 22, и
HC имеет аминокислотную последовательность SEQ ID NO: 6.
10. Антитело по п.6, где
LC имеет аминокислотную последовательность SEQ ID NO: 23, и
HC имеет аминокислотную последовательность SEQ ID NO: 6.
11. Антитело по п.6, где
LC имеет аминокислотную последовательность SEQ ID NO: 24, и
HC имеет аминокислотную последовательность SEQ ID NO: 6.
12. Антитело по п.6, где
LC имеет аминокислотную последовательность SEQ ID NO: 25, и
HC имеет аминокислотную последовательность SEQ ID NO: 6.
13. Антитело, которое связывается с комплексом ANGPTL3/8 человека, полученное путем культивирования клетки млекопитающего, содержащей молекулу кДНК, где молекула кДНК кодирует полипептиды, содержащие аминокислотные последовательности SEQ ID NO: 5 и 6, в таких условиях, что полипептиды экспрессируются, и выделения антитела.
14. Антитело, которое связывается с комплексом ANGPTL3/8 человека, полученное путем культивирования клетки млекопитающего, содержащей молекулу кДНК, где молекула кДНК кодирует антитело, содержащее аминокислотные последовательности SEQ ID NO: 21 и 6, в таких условиях, что антитело экспрессируется, и выделения антитела.
15. Антитело, которое связывается с комплексом ANGPTL3/8 человека, полученное путем культивирования клетки млекопитающего, содержащей молекулу кДНК, где молекула кДНК кодирует антитело, содержащее аминокислотные последовательности SEQ ID NO: 22 и 6, в таких условиях, что антитело экспрессируется, и выделения антитела.
16. Антитело, которое связывается с комплексом ANGPTL3/8 человека, полученное путем культивирования клетки млекопитающего, содержащей молекулу кДНК, где молекула кДНК кодирует антитело, содержащее аминокислотные последовательности SEQ ID NO: 23 и 6, в таких условиях, что антитело экспрессируется, и выделения антитела.
17. Антитело, которое связывается с комплексом ANGPTL3/8 человека, полученное путем культивирования клетки млекопитающего, содержащей молекулу кДНК, где молекула кДНК кодирует антитело, содержащее аминокислотные последовательности SEQ ID NO: 24 и 6, в таких условиях, что антитело экспрессируется, и выделения антитела.
18. Антитело, которое связывается с комплексом ANGPTL3/8 человека, полученное путем культивирования клетки млекопитающего, содержащей молекулу кДНК, где молекула кДНК кодирует антитело, содержащее аминокислотные последовательности SEQ ID NO: 25 и 6, в таких условиях, что антитело экспрессируется, и выделения антитела.
19. Антитело, которое связывается с комплексом ANGPTL3/8 человека, полученное путем культивирования клетки млекопитающего, содержащей по меньшей мере две молекулы кДНК, где первая молекула кДНК кодирует легкую цепь антитела, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, 21, 22, 23, 24 и 25, и вторая молекула кДНК кодирует тяжелую цепь антитела, содержащую аминокислотную последовательность SEQ ID NO: 6, в таких условиях, что антитело экспрессируется, и выделения антитела.
20. Антитело по любому из пп.1-19, где указанное антитело связывает и нейтрализует комплекс ANGPTL3/8 человека в стандартном анализе активности липопротеинлипазы с EC50 3,0 нМ или менее.
21. Антитело по любому из пп.1-19, где антитело связывается с комплексом ANGPTL3/8 человека с константой диссоциации менее или равной 1×10-6 M.
22. Антитело по любому из пп.1-19, где антитело связывается с ANGPTL3 человека и ANGPTL8 человека с константой диссоциации более 1×10-6 M.
23. Антитело по любому из пп.1-19, где указанное антитело демонстрирует положительное связывание с комплексом ANGPTL3/8 человека зависимым от концентрации образом и не демонстрирует детектируемое связывание с ANGPTL3 человека и ANGPTL8 человека вплоть до концентрации антигена, равной 100 нМ.
24. Антитело по любому из пп.1-19, где антитело снижает уровень триглицеридов in vivo по меньшей мере на 50% по сравнению с контролем IgG при дозе 10 мг/кг через 14 дней после введения дозы.
25. Фармацевтическая композиция для снижения уровня триглицеридов, содержащая указанное антитело по любому из пп.1-24 и приемлемый носитель, разбавитель или эксципиент.
26. Клетка млекопитающего, содержащая молекулу ДНК, которая содержит полинуклеотидную последовательность, кодирующую полипептиды, содержащие аминокислотные последовательности SEQ ID NO: 5 и 6, где клетка способна экспрессировать антитело по п.13.
27. Клетка млекопитающего, содержащая молекулу ДНК, которая содержит полинуклеотидную последовательность, кодирующую полипептиды, содержащие аминокислотные последовательности SEQ ID NO: 21 и 6, где клетка способна экспрессировать антитело по п.14.
28. Клетка млекопитающего, содержащая молекулу ДНК, которая содержит полинуклеотидную последовательность, кодирующую полипептиды, содержащие аминокислотные последовательности SEQ ID NO: 22 и 6, где клетка способна экспрессировать антитело по п.15.
29. Клетка млекопитающего, содержащая молекулу ДНК, которая содержит полинуклеотидную последовательность, кодирующую полипептиды, содержащие аминокислотные последовательности SEQ ID NO: 23 и 6, где клетка способна экспрессировать антитело по п.16.
30. Клетка млекопитающего, содержащая молекулу ДНК, которая содержит полинуклеотидную последовательность, кодирующую полипептиды, содержащие аминокислотные последовательности SEQ ID NO: 24 и 6, где клетка способна экспрессировать антитело по п.17.
31. Клетка млекопитающего, содержащая молекулу ДНК, которая содержит полинуклеотидную последовательность, кодирующую полипептиды, содержащие аминокислотные последовательности SEQ ID NO: 25 и 6, где клетка способна экспрессировать антитело по п.18.
32. Способ получения антитела, включающий культивирование клетки млекопитающего по пп.26-31 в условиях, при которых происходит экспрессия антитела, и выделение экспрессированного антитела.
33. Способ лечения заболевания, связанного с метаболизмом липидов, выбранного из группы, состоящей из: атеросклеротического сердечно-сосудистого заболевания (АССЗ), диабета, ожирения, неалкогольного стеатогепатита (НАСГ), хронической почечной недостаточности (ХПН) или гипертриглицеридемии или их комбинации, включающий введение пациенту, нуждающемуся в этом, эффективного количества указанного антитела по любому из пп.1-24.
34. Способ снижения уровня триглицеридов, включающий введение пациенту эффективного количества указанного антитела по любому из пп.1-24.
35. Антитело по любому из пп.1-24 для применения при лечении АССЗ, диабета, ожирения, НАСГ, ХПН или гипертриглицеридемии, или их комбинации.
36. Фармацевтическая композиция для применения при лечении АССЗ, диабета, ожирения, НАСГ, ХПН или гипертриглицеридемии, или их комбинации, содержащая эффективное количество указанного антитела по любому из пп.1-24.
| WO 2017177181 A1, 12.10.2017 | |||
| CHI X | |||
| et al | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Vol | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| No | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| ЦЕПНОЙ ВЕТРЯНОЙ ДВИГАТЕЛЬ | 1924 |
|
SU1137A1 |
| ZHANG R | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Механический пресс | 1976 |
|
SU620064A1 |
| WO 2018094112 A1, 24.05.2018. | |||

