Целью настоящего изобретения является способ переустройства активности сериновой протеазы, связанной с NS3 белком HCУ, что позволяет использовать способность NS4A белка HCУ, или содержащихся в нем последовательностей в виде кофактора активности сериновой протеазы, или, в более широком смысле, энзимных активностей, связанных с NS3.
Хорошо известно, что вирус гепатита C (HCУ) является основным этиологическим агентом гепатита ниА, ниВ (NANB). Установлено, что HCУ вызывает, по крайней мере, 90% вирусного гепатита NANB, после переливания крови и 50% спорадического NANB гепатита. Хотя в выборе доноров крови и иммунологическом анализе крови, используемой для переливаний, достигнут большой прогресс, все еще наблюдается высокий уровень инфицирования острым HCУ среди пациентов, перенесших переливание крови (в мире регистрируется миллион или более таких инфекций ежегодно). Примерно 50% инфицированных HCУ пациентов демонстрируют развитие цирроза печени в течение периода времени от 5 до 40 лет. Кроме этого, последние клинические исследования подтвердили тот факт, что имеется корреляция между хроническим инфицированием HCУ и развитием гепатоклеточной карциномы.
HCУ представляет собой оболочечный вирус, содержащий РНК-положительный геном размером примерно 9,4 kb. Такой вирус является членом семейства Flaviridal, другими членами которого являются флавивирусы и пестивирусы. РНК-геном HCУ недавно был картирован. Сравнение последовательностей HCУ-геномов, выделенных в различных районах мира показало, что такие последовательности могут быть чрезвычайно гетерогенными. Большая часть НCУ-генома имеет открытую рамку считывания (ORF), которая может содержать от 9030 до 9099 нуклеотидов. Такая ORF кодирует один вирусный полипротеин, длина которого может изменяться от 3010 до 3033 аминокислот. В ходе цикла вирусного заражения такой полипротеин протеолитически процессируется в индивидуальные генные продукты, необходимые для репликации вируса. Гены, кодирующие структурные белки HCУ, находятся на 5'- конце ORF, тогда как участок, кодирующий неструктурные белки, занимает всю оставшуюся часть ORF.
Структурные белки состоят из C (сердцевины, 21 кДа), E1 (оболочка gp37) и E2 (NS1, gp61). С представляет собой негликолизированный белок 21 кДа, который, вероятно, образует вирусный нуклеокапсид. Белок E1 предоставляет собой гликопротеин, примерно 37 кДа, и предполагается, что он является структурным белком внешней оболочки вируса. E2, другой мембранный гликопротеин, 61 кДа, по-видимому, является вторым структурным белком во внешней оболочке вируса.
Неструктурная область начинается с NS3 (p24), гидрофобного белка 24 кДа, функция которого неизвестна. NS3, белок 68 кДа, следующий за NS2 в полипротеине, в соответствии с предсказаниями, имеет два функциональных домена: домен сериновой протеазы в первых 200 амино-концевых аминокислотах и домен РНК-зависимой АТФ-азы на С-конце. Генный участок, соответствующий NS4, кодирует NS4А (p6) и NS4B (p26), два гидрофобных белка, 6 и 24 кДа соответственно, функции которых до настоящего времени не выяснены. Ген, соответствующий NS5, также кодирует два белка; 5A (p56)NS и NS5B (p65), 56 и 65 кДА соответственно. Аминокислотная последовательность, присутствующая во всех РНК-зависимых РНК-полимеразах, может распознаваться в области NS5. Этот факт подтверждает то, что участок NS5 содержит части аппарата репликации вируса.
Различные молекулярно-биологические исследования показали, что сигнальная пептидаза, протеаза, связанная с эндоплазмическим рет кулумом клетки-хозяина, ответственна за протеолитический процессинг в неструктурной области, т. е. на сайтах C/E1, E1/E2 и E2/NS2 сериновая протеаза, содержащаяся в NS3, ответственна за расщепление в местах соединения между NS3 и NS4A, между NS4A и NS4B, между NS4B и NS5A, а также между NS5A и NS5B. Было обнаружено, что в результате расщепления под действием сериновой протеазы остаток цистеина или трионина остается со стороны амино-конца (положение Р1), а остаток аланина или серина остается со стороны с-конца (положение Р1') расщепляемой связи. Вторая протеазная активность HCУ, по-видимому, ответственна за расщепление между NS2 и NS3. Такая протеазная активность содержится в области, включающей как часть NS2, так и часть NS3, содержащей домен сериновой протеазы, однако, используются различные каталитические механизмы.
В свете сказанного выше, NS3 протеаза рассматривается как потенциальная мишень для разработки анти-HCУ терапевтических агентов. Однако поиск таких агентов затруднен тем, что активность сериновой протеазы, показываемая NS3 in vitro, слишком мала для осуществления скрининга ингибиторов.
Авторами изобретения было неожиданно обнаружено, что такое важное ограничение может быть снято в результате применения способа согласно настоящему изобретению, который также обеспечивает дополнительные преимущества, как это следует из дальнейшего описания.
В соответствии с настоящим изобретением, способ воспроизводства in vitro протеолитической активности протеазы NS3 HCУ характеризуется применением в реакционной смеси как последовательностей, содержащихся в NS3, так и последовательностей, содержащихся в NS4A.
Оптимальная активность сериновой протеазы достигается в том случае, когда NS4A присутствует в соотношении 1:1 по отношению к NS3.
NS3 и NS4A также могут вводиться в реакционную смесь в виде NS3 - NS4A предшественника, если такой предшественник в аутопротеолитических условиях образует эквимолярные количества NS3 и NS4A.
Существует также возможность мутации сайта расщепления между NS3 и NS4A в предшественнике так, что NS4A остается ковалентно связанным с NS3. Последовательности, не оказывающие влияния на протеолитическую активность NS3, могут быть далее удалены из такого не способного к протеолизу предшественника.
Настоящее изобретение также относится к новой композиции вещества, характеризующейся тем, что она включает белки, последовательности которых описаны последовательностями SEQID N 1 и SEQID N 2 или содержащимися в них последовательностями или их производными. Следует иметь в виду, что такие последовательности могут меняться в различных изолятах HСУ, поскольку все РНК-вирусы обладают высокой степенью изменчивости. Такая новая композиция обладает протеолитической активностью, необходимой для протеолитического созревания нескольких из неструктурных HCУ белков.
Другая цель настоящего изобретения состоит в применении таких композиций для проведения энзимных анализов, способных в терапевтических целях идентифицировать соединения, ингибирующие энзимную активность, связанную с NS3, включая ингибиторы взаимодействия между NS3 и NS4A.
Выше было приведено общее описание настоящего изобретения. Теперь с помощью следующих ниже примеров будет представлено подробное описание конкретных технических решений изобретения, имеющее своей целью лучшее понимание его целей, отличий, преимуществ и способа его осуществления.
На рисунке проиллюстрированы плазмидные векторы, используемые в способе изобретения для активации HCУ NS3 протеазы в культивируемых клетках и in vitro (пример 1 и пример 2).
Пример 1
Способ активации NS3 сериновой протеазы HCУ в культивируемых клетках.
Конструировали плазмидные векторы для экспрессии NS3, 4A и других неструктурных белков HCУ в клетках HEla. Сконструированные плазмиды схематически показаны на чертеже. Отобранные фрагменты кДНК, соответствующие геному HCУ BK (HCУ-BK), клонировали ниже промотора бактериофага Т7 в плазмидном векторе pCite-1R (Novagen). Такой вектор экспрессии содержит сайт внутренний входа рибосомы вируса энцефаломиокардита с тем, чтобы гарантировать эффективную трансляцию информационной РНК, транскрибирующейся с промотором Т7 даже в отсутствии структуры CAP.
Различные фрагменты HCУ-ВК кДНК клонировали в плазмиду pCite-1R с использованием методов, известных в практике молекулярной биологии. pCite /NS3/ включает часть HCУ-ВК генома, содержащуюся между нуклеотидами 3351 и 5175 (аминокислоты 1007-1615 полипротеина). pCite/NS4B/5A включает часть HCE-ВК генома, содержащуюся между нуклеотидами 5652 и 7467 (аминокислоты 1774-2380). pCite/NS3/4A включает часть HCУ-ВК генома, содержащуюся между нуклеотидами 3711 и 5465 (аминокислоты 991-1711). pCite/NS4A/ включает часть HCУ-ВК генома, содержащуюся между нуклеотидами 5281 и 5465 (аминокислоты 1649-1771). pCite/NS5AB/ включает часть HCУ-ВК генома, содержащуюся между нуклеотидами 6224 и 9400 (аминокислоты 1965-3010). Приведенная выше нумерация согласуется с последовательностями генома и полипротеина, приведенными для HCУ-ВК в статье Jakamizava с сотр. "Структура и организация генома вируса гепатита C, выделенного из человеческих носителей, (1991), J.Virol. 65, 1105-1113.
С целью достижения эффективной экспрессии различных частей HCУ полипротеина клетки Hela заражали VTF7-3, рекомбинантным вирусом осповакцины, который позволяет осуществлять синтез РНК-полимеразы бактериофага Т7 в цитоплазме зараженных клеток. Такие клетки после заражения далее трансфицировали плазмидными векторами, выбранными среди тех, что показаны на чертеже. Зараженные и трансфицированные таким образом клетки Hela далее метаболически метили (35S) метионином, и рекомбинантные белки, закодированные различными плазмидами, могли быть идентифицированны методом иммуноосаждения с помощью поликлональных кроличьих антител, распознающих NS3, NS4 или NS5A. Способ, описанный в настоящем примере, предназначенный для анализа рекомбинантных HCУ белков, уже был опубликован Z.Jomei, с сотр. в статье "NS3-сериновая протеаза, требуемая для обработки полипротеина вируса гепатита C", J.Virol (1993) 67,1017-1026 и в указанной там библиографии.
В результате трансфекции плазмиды pCite (NS3) в клетки Hela, зараженные yTF7-3, можно наблюдать синтез белка, содержащего каталитический домен HCУ NS3 протеазы. pCite (NS4B5A) кодирует часть HCУ полипротеина, содержащего пептидную связь в месте соединения между NS4B и NS5B, которое, как можно ожидать, гидролизуется под действием активности сериновой протеазы, связанной с NS3. Однако при осуществлении контрансфекции pCite NS3 с pCite NS4B5A не наблюдалось признаков протеолитического расщепления. В отличие от этого при экспрессии NS3 серинового протеазного домена в комбинации с NS4A протеолитическое расщепление предшественника, закодированного pCite (NS4B5A), может протекать обычным образом. Коэкспрессия NS3 серинового протеазного домена и 4A может достигаться, например путем трансфекции эквимолярными количествами плазмид pCite (NS3) и pCite (NS4A), трансфекцией плазмиды, кодирующей предшественник, содержащий как NS3, так и NS4A (pCite(NS34A), или путем трансфекции производного последней из указанных плазмид, у которой вся последовательность, не имеющая отношения к протеолизу, была подвергнута делеции [pCite(NS34A)] , или путем трансфекции производного последней плазмиды, у которой вся последовательность, не имеющая отношения к протеолизу, была подвергнута делеции [pCite(NS3 Δ int 12371635)]. Таким образом, NS4A, кратковременно экспрессированный в клетках Hela, может активировать протеолитическую активность, связанную с NS3, чего не наблюдается в других случаях,
Пример 2.
Способ активации сериновой NS3 протеазы HCУ в in vitro трансляционном анализе.
Плазмиды, показанные на чертеже, также могут использоваться для in vitro синтеза мРНК, кодирующей соответствующие HCУ белки с использованием очищенного РНК-полимеразного фермента фага Т7 (Promega).
Обычно плазмиды, являющиеся производным pCite-1R, линеаризуют с использованием подходящих рестриктаз и транскрибируют с использованием предписаний производителя (Promega). Такая синтетическая мРНК может быть далее использована для синтеза соответствующих белков в экстрактах ретикулоцитов кроликов в присутствии микросомальных мембран поджелудочной железы собаки. Ретикулоцитные экстракты, микросомальные мембраны собачьей поджелудочной железы, как и другие требуемые материалы, были приобретены от Promega, причем производитель также снабжает инструкциями для описанного выше способа синтеза белка in vitro.
В результате программирования in vitro трансляционной смеси мРНК, транскрибированной из pCite (NS3), можно наблюдать синтез белка с ожидаемым молекулярным весом (68 кДа), содержащего полный NS3 сериновый протеазный домен. мРНК, транскрибированная из pCite (NS5AB), регулирует синтез предшественника с молекулярным весом 115 кДа, который содержит NS5A и NS5B и, таким образом, служит субстратом для протеолитической активности, связанной с NS3.
Однако в том случае, когда два таких белка, содержащих NS3 сериновый протеазный домен и субстрат с сайтом, соответствующим месту соединения между NS5A и NS5B, синтезируют в одной и той же реакционной смеси, явное свидетельство протеолитической активности NS3 отсутствует.
С другой стороны, мРНК, транскрибированная из pCite (NS34A), транслируется в белок-предшественник весом примерно 76 кДа, который самопроцессируется протеолитически in vitro с образованием эквимолярных количеств двух белков весом 70 кДа и 6 кДа, содержащих NS3 и NS4A соответственно.
Если, помимо мРНК, транскрибированной из pCite (NS3A), мРНК, транскрибированная из pCite (NS5AB), вводится в in vitro трансляционную смесь, то помимо самопротеолиза на сайте между NS3 и NS4A, можно наблюдать образование двух новых белков весом 55 кДа и 65 кДа, которые содержат NS5A и NS5B соответственно. Такие белки представляют собой продукт протеолиза предшественника, содержащего NS5A и NS5B, под действием NS3. Аналогичным образом белковые продукты весом в 56 кДа и 65 кДа, протеолитически образовавшиеся из NS5AB предшественника, получаются в том случае, если мРНК, транскрибированная из pCite (NS3 Δ int 1237-1635), контранслируется с мРНК, транслированной из pCite (NS5AB).
Резюмируя такой результат, можно заключить, что в условиях in vitro протеазный домен NS3 сам по себе не способен проявлять протеазную активность на субстрате, содержащем NS5A и NS5B. Однако серин-протеазная активность NS3 становится очевидной, если другая белковая последовательность, содержащая NS4A, присутствует помимо NS3 протеазного домена.
Пример 3
Способ активации NS3 протеазы HCУ с использованием синтетического пептида, содержащего NS4A последовательности.
Синтетический пептид, содержащий последовательность SEQ ID N 3, синтезировали на твердой фазе. Такая последовательность является производной C-терминальной части последовательности ID N 2. Синтез пептида проводили на твердой фазе в соответствии со способами, известными специалистам в данной области. В этом пептиде карбокситерминальный цистеин заменяли на альфа-аминомасляную кислоту (Abu).
Такой пептид добавляли в in vitro трансляционную среду, одновременно программируемую мРНК, транскрибируемыми из плазмид pCite (NS3) и pCite (NS5AB).
Таким образом, возникала возможность наблюдать протеолитеческую активность, связанную с сериновым протеазным доменом NS3, в результате чего происходит протеолитическое расщепление субстрата на два продукта, содержащих белки NS5A и NS5B. Такая активность зависит от одновременного присутствия NS3 серинового протеазного домена и синтетического пептида с последовательностью SEQ ID N 3.
Пример 4
Способ анализа рекомбинантной NS3 сериновой протеазы HCУ на пептидном субстрате.
Плазмиду рТ7-7 NS3 (1027-1206), показанную на чертеже и описанную в примере 4, конструировали с целью осуществления экспрессии в E.coli белкового фрагмента, находящегося между аминокислотой 1 и аминокислотой 180 последовательности SEQ ID N 1. Такой фрагмент содержит сериновый протеазный домен NS3, что установлено экспериментально. Фрагмент кДНК HCУ, кодирующий описанный NS3 фрагмент, клонировали в плазмиду рТ7-7, вектор экспрессии, содержащий Т7 РНК-полимеразный промотор ⊘ 10 и сайт инициации трансляции для гена белка 10 фага Т7 (Studier и Moffat. Применение РНК-полимеразы бактериофага Т7 для обеспечения селективной экспрессии клонированных генов (1986), J. Мol. Biol, 189, стр. 113-130). Фрагмент кДНК, кодирующий NS3 сериновый протеазный домен, как это описано выше, клонировали после промотора бактериофага Т7 и в рамке с первым ATG кодоном гена 10 белка Т7 с использованием способов, известных в молекулярно-биологической практике. Плазмида рТ7-7 также содержит ген β-лактамазного энзима, который может использоваться в качестве маркера селекции клеток E.coli, трансформированных плазмидами, производными рТ7-7.
Затем плазмидой рТ7-7 NS3 (1027-1206) трансформировали штамм E.coli (DE53), который обычно используют для высоко-уровневой экспрессии генов, клонированных в векторы экспрессии, содержащие Т7 промотор. В таком штамме E. coli полимераза гена Т7 находится на бактериофаге λ DE53, который интегрирован в хромосому B21. Экспрессия изучаемого гена индуцируется добавлением изопропилтиогалактозида (IPTG) в растительную среду в соответствии с описанной выше методикой (Studier и Moffat). Применение РНК-полимеразы бактериофага Т7 для направления селективной высоко-уровневой экспрессии клонированных генов, (1986), J. Mol. Biol. - 189, стр. 113-130).
Фрагмент рекомбинантного NS3, содержащий сериновый протеазный домен, может быть очищен из E.coli BL21 (DE53), трансформированной плазмидой рТ7-7 NS3 (1027-1206), в соответствии с описанными ниже способами.
Вкратце, клетки E.coli B121 (DE53), содержащие рТ7-7 NS3(1027-1206) плазмиду, выращивают при 37oC до оптической плотности, при длине волны 600 нм, порядка 0,8 единиц поглощения. После этого среду охлаждают до 22oC и продуцирование желаемого белка индуцируют добавлением IPTG до конечной концентрации 0,4 мМ. Через 4-6 часов при 22oC в присутствии IPTG клетки собирали и подвергали лизису с помощью пресовального устройства (French-pressure cell) в буффере, содержащем 20 мМ фосфата натрия, pH 6,5, 0,5% (вес./объем) (3-/(3-холамидопропил)-диметиламмоний/1-пропансульфоната (CHAPS), 50% (об./об.) глицерина, 10 мМ дитиотреитола и 1 мМ ЕДТА (лизисный буфер). Клеточные обломки удаляли центрифугированием (1 час при скорости 120000 xg) и полученный в результате осадок ресуспендировали в лизисном буфере, расщепляли ДМКазой 1, регомогенизировали и повторно центрифугировали в описанных выше условиях. Ионо-обменную смолу S-Sepharose Fast Flow (Pharmacia), предварительно уравновешенную в лизисном буфере, добавляли к слитым супернатантам (30% об. /об. ) и полученную суспензию перемешивали в течение 1 часа при 4oC. Смолу седиментировали и тщательно промывали лизисным буфером и после этого загружали в хроматографическую колонку. NS3 протеазу элюировали со смолы, применяя O-1M градиент NaCl. Фракции, содержащие протеазу, уравновешивали 50 мМ натрийфосфатного буфера pH 7,5, 10% (об./об.) глицерина, 0,5% (вес./об.) CHAРS и 2 мМ дитиотреитола. После осуществления такой стадии белок имел чистоту 90-95%. Очистка до степени больше 98% достигалась последующей хроматографией на Гепарин Сефарозе, уравновешенной 50 мМ Трис pH 7,5 10% (об./об.) глицерина, 0,5% (вес./об.) CHAPS и 2 мМ дитиотреитола. Элюирование NS3 протеазы с такой колонки осуществляли с применением линейного O-1M градиента NaCl.
Концентрацию очищенного белка определяли с помощью Био-Рад (Bio-Rad) анализа на белок (Био-Рад категория 500-0006).
Рекомбинантная NS3 сериновая протеаза, полученная согласно описанному выше способу в E.coli, должна быть подвергнута анализу на активность расщепления субстрата, обеспечивающему детектируемые количества продуктов дробления. Детекцию сигнала предварительно осуществляют методами колориметрии или флуорометрии. Можно применять такие способы, как HPLC и т.п.
Так, например, мы использовали в качестве субстрата синтетические пептиды, соответствующие NS4A/4B соединению HCУ полипротеина и содержащие аминокислотную последовательность SEQ ID N 4 или ее часть.
С другой стороны, пептидные эфиры, имеющие общую структуру, показанную последовательностью SEQ ID N 5, также могут использоваться в такой операции.
Анализ на активность осуществляли путем инкубации 5-1000 мкМ субстрата и 0,05-2 мкМ протеазы в буфере, содержащем 25 мМ Трис/HCl pH 7,5 3 мМ дитиотреитола, 0,5% (вес. /об.) CHAРS и 10% (об./об.) глицерина, в течение 1-3 часов при 22oC. Реакцию останавливали добавлением трифторуксусной кислоты с достижением конечной концентрации порядка 0,1% (вес./об.).
Затем продукты реакции разделяли методом HPLC на C18 обратно-фазовой колонке и проводили их количественное определение по их поглощению в дальней УФ-области.
Протеолитическая активность, показываемая рекомбинантной NS3 серийной протеазой, очищенной из E.coli, оказывалась очень низкой в том случае, когда анализ на активность проводили описанным выше способом. Однако было обнаружено, что увеличение количеств синтетического пептида, изображенного последовательностью SEQ ID N 3, стимулирует протеолитическую активность рекомбинантной NS3 сериновой протеазы, увеличивая ее в 20 раз. Максимальная активность достигается в том случае, когда рекомбинантная NS3 сериновая протеаза и синтетический пептид присутствуют в эквимолярных количествах.
Описанный выше анализ может использоваться для поиска ингибиторов протеазы. Поскольку активность NS3 протеазы в таком анализе зависит от взаимодействия NS3 серинового протеазного домена с аминокислотными последовательностями - производными NS4A, также появляется возможность с использованием описанного выше анализа проводить поиск антагонистов такого взаимодействия между NS3 и NS4A, которые в конечном счете ингибируют протеолитическую активность, связанную с NS3.
Пример 5
Подробная конструкция плазмид на единственном рисунке
pCite (NS3) содержит часть HCУ-BK генома, находящуюся между нуклеотидами 3351 и 5175 (аминокислоты 1007-1615 полипротеина). Конструкция такой плазмиды описана в статье I.Tomei с сотр., "NS3-сериновая протеаза, требующаяся для процессирования полипротеина вируса гепатита C", J. Virol (1993) 67, 1017-1026.
pCite (NS4B/5A) получали клонированием ScaI-BamHI фрагмента, произведенного из плазмиды pCite (NS4-5), описанной в статье Tomei с сотр., в pCite (NS3), которая предварительно расщеплена MSCl и BamH1. pCite (NS4B/5A) содержит часть HCУ генома, находящуюся между нуклеотидами 5652 и 7467 (аминокислоты 1774-2380 полипротеина).
pCite (NS5AB) кодирует белок, включающий последовательность от аминокислоты 1965 до аминокислоты 3010 полипротеина HCУ-BK. Для конструирования такой плазмиды плазмиду pCite (SX), описанную в статье Tomei с сотр., (1993), см. выше, вначале переваривали Asel и обрабатывали фрагментом Кленова ДНК-полимеразы. После инактивации энзима Кленова плазмиду переваривали XbaI. Полученный в результате фрагмент кДНК, содержащий область между нуклеотидами 6224 и 9400, подвергали очистке и вставляли в сайты BstXI и XbaI вектора pCite -1R, после затупления конца генерированного с помощью BstXI путем воздействия Т4 ДНК-полимеразы.
pCite (NS3/4A) получали следующим образом. Фрагмент кДНК, соответствующий области между нуклеотидами 3711 и 5465 НСУ-ВК генома, синтезировали с помощью полимеразной цепной реакции (PCR), используя в качестве затравок олигонуклеотиды, специфичные к последовательности. В антисмысловой олигонуклеотид подходящим образом включали UAG стоп-кодон. После РСР амплификации полученную в результате кДНК расщепляли на 5'-конце с помощью SAL1 и продукт, состоящий из 750 пар оснований, направленно клонировали в сайты SAL2 и NheI плазмиды pCite (SX), после затупления Nhel конца с помощью фрагмента Кленова ДНК-полимеразы. Полученная в результате плазмида кодирует часть НСУ-ВК полипротеина, находящуюся между аминокислотами 991 и 1711.
Для конструирования pCite (NS4A) фрагмент кДНК, соответствующий области между нуклеотидами 5281 и 5465 HCУ-BK генома (аминокислоты 1649-1711), получали амплификацией полимеразной цепной реакцией (PCR) в присутствии в качестве затравки олигонуклеотидов, специфичных к последовательности. Полученную в результате PCR амплификации кДНК далее клонировали в сайты BstxI и Stul плазмиды pCite-1R, после затупления ВstxI переваренного конца с помощью ДНК-полимеразы бактериофага Т4.
pCite (NS3 Δ int 1237-1635) представляет собой производное pCite (NS3/4A), из которой все последовательности, находящиеся между нуклеотидами 4043 и 5235, подвергнуты делеции. Этот продукт получали перевариванием pCite (NS3/4A) BsteII и частично Scal. Фрагмент, содержащий интересующую делецию, далее замыкали с использованием Т4 ДНК-лигазы. Эта плазмида кодирует белок, имеющий те же амино- и карбокси-концы, что и закодированные pCite (NS3/4A), однако, как было установлено экспериментально, все аминокислотные остатки, находящиеся между аминокислотой 1237 и аминокислотой 1635, несущественны для серин-протеазной NS3 активности и они были подвергнуты делеции.
pT7-7 /NS3 (1027-1206)/ содержит НСУ последовательность от нуклеотида 3411 до нуклеотида 3951, кодирующую НСУ NS3 фрагмент между аминокислотой 1027 и аминокислотой 1206. Для получения такой плазмиды создают ДНК-фгармент амплификацией кДНК HCУ полимеразой цепной реакцией (PCR) с использованием тех олигонуклеотидов, на которые ссылались, как на последовательность SEQ ID N 6 и SEQ ID N 7. Фрагмент кДНК, полученный с помощью PCR, фосфорилировали, переваривали с Ndel и затем клонировали после промотора бактериофага Т7, сразу после первого ATG кодона гена 10 Т7 в векторе рТ7-7, предварительно переваренном Ndel и SmaI (Studier и Moffatt). Применение РНК-полимеразы бактериофага Т7 для направления селективной высоко-уровневой экспрессии клонированных генов, (1986), J. Mol. Biol. 189, стр. 113-130). Следует отметить, что амбер-кодон вставляли сразу после HCУ-производной последовательности.





Изобретение относится к биотехнологии и может быть использовано для разработки анти-НСУ терапевтических агентов. Репродукцию протеолитической активности NS3 протеазы вируса гепатита С (НСУ) осуществляют путем введения в реакционную смесь вместе с протеазой NS3 аминокислотной последовательности NS4A в соотношении 1:1. NS3 и NS4A могут быть введены в виде предшественника NS3 - NS4A. Изобретение позволяет увеличить активность сериновой NS3 протеазы in vitro. 3 з.п. ф-лы, 1 ил.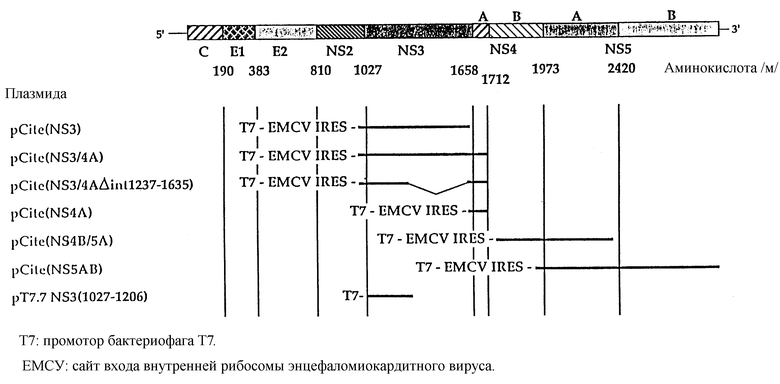
| J | |||
| of Virology, 1993, v.67, N 7, p.1017-1026 | |||
| Огнетушитель | 0 |
|
SU91A1 |
