Область изобретения
Настоящее изобретение относится, в общем, к диагностике вирусов. В частности, изобретение относится к определению комплекса антиген/антитело для точного диагностирования инфекции вируса гепатита С.
Предпосылки изобретения
Вирус гепатита С (HCV) - основная причина парентерального гепатита не-А, не-В (NANBH), который в основном передается при переливании крови и половых контактах. Вирус присутствует у 0,4-2,0% доноров крови. Хронический гепатит развивается примерно у 50% инфицированных, и примерно у 20% из них развивается цирроз печени, который иногда приводит к гепатоцеллюлярной карциноме. Поэтому изучение и контроль этого заболевания являются важными для медицины.
HCV был впервые определен и характеризован как причина NANBH Хофтеном с соавторами (Houghten et al.). См. также Международную Публикацию № WO 89/04699, WO 90/11089 и WO 90/14436. HCV имеет геном, представленный 9,5 т.п.н. смысловой, одно-цепочечной РНК и является членом семейства вирусов Flaviridae. По меньшей мере шесть отдельных, хотя и родственных генотипов HCV было идентифицировано на основе филогенетических анализов (Simmonds et al., J.Gen.Virol (1993) 74: 2391-2399). Вирус кодирует единственный полипротеин, имеющий более 3000 аминокислотных остатков (Choo et al., Science (1989) 244:359-362; Choo et al., Proc. Natl. Acad. Sci. USA (1991) 88:2451-2455; Han et al., Proc. Natl. Acad. Sci USA (1991) 88: 1711-1715). Во время и после трансляции этот протеин преобразуется как в структурные, так в неструктурные (NS) белки.
В частности, как показано на фиг.1, геном HCV кодирует несколько белков. Порядок и номенклатура продуктов расщепления полипротеина HCV следующая: NH2-C-E1-E2-p7-NS2-NS3-NS4a-NS4b-NS5a-NS5b-COOH. Начальное расщепление полипротеина катализируется протеазами хозяина, которые высвобождают три структурных белка, N-концевой нуклеокапсидный белок (обозначаемый «кор») и два оболочечных гликопротеина, «Е1» (также известный как «Е») и «Е2» (также известный, как «E2/NS1»), также как неструктурные (NS) белки, содержащие вирусные ферменты. NS области обозначены NS2, NS3, NS4 и NS5. NS2 - интегральный мембранный протеин с протеолитической активностью. NS2, как один, так и вместе с N33, расщепляет NS2-NS3 слабую связь, которая, в свою очередь, создает N-конец NS3 и высвобождает большой полипротеин, который обладает активностью как сериновой протеазы, так и РНК-хеликазы. NS3-протеаза служит для преобразования оставшегося полипротеина. Окончание созревания полипротеина инициируется автокаталитическим расщеплением соединения NS3-NS4a, катализируемым NS3 сериновой протеазой. Последующие расщепления полипротеина HCV, опосредованные NS3, скорее всего, включают в себя распознавание расщепляющихся связей полипротеина молекулой N33 другого полипептида. В этих реакциях NS3 высвобождает NS3 кофактор - (NS4a), два белка (NS4b и NS5a) и РНК зависимую РНК-полимеразу (NS5b).
Описан ряд общих и специфических полипептидов, полученных из полипротеина HCV, пригодных в качестве иммунологических и диагностических реагентов. Например, Houghton et al., Европейская публикация №318,216 и 388,232; Choo et al., Science (1989) 244:359-362; Kuo et al., Science (1989) 244:362-364; Houghton et al., Hepatology (1991) 14:381-388; Chien et al., Proc. Natl. Acad. Sci. USA (1992) 89:10011-10015; Chien et al., J. Gastroent. Hepatol. (1993) 8-S33-39; Chien et al., Международная публикация № WO 93/00365, Chinen D. Y. Международная публикация № WO 94/01778. Эти публикации создают значительные предпосылки как для исследований HCV в целом, так и для производства и применения иммунологических реагентов для полипептида HCV.
Чувствительные, специфические способы отбора и идентификации носителей HCV и крови и ее продуктов, зараженных HCV, создадут значительный прогресс в медицине. Пост-трансфузионный гепатит встречается примерно у 10% пациентов, перенесших переливание крови, и HCV отмечен вплоть до 90% в этих случаях. Лечение больных, а также профилактика и передача HCV с кровью и продуктами крови или при близком личном контакте требует надежных диагностических и прогностических средств. Таким образом, было разработано несколько анализов для серодиагностики HCV инфекции. Например, см. Choo et al., Science (1989) 244:359-362; Kuo et al.. Science (1989) 244,: 362-364; Choo et al., Br. Med. Bull. (1990) 46:423-441; Ebeling et al., Lancet (1990) 335:982-983; van der Poel et al.. Lancet (1990) 335:558-560; van der Poel et al., Lancet. (1991) 337:317-319; Chien, D.Y., Международная публикация № WO 97/01178; Valenzuela et al., Международная публикация № WO 97/44469; и Kashiwakuma et al., U.S. Patent No. 5,871,904.
В некоторых анализах на базе сыворотки возникла серьезная проблема из-за значительного интервала между заражением и обнаружением вируса, который часто превышает 80 дней. Этот интервал может создать большой риск для реципиентов крови. Для преодоления этой проблемы были разработаны тесты на основе нуклеиновой кислоты (NAT), которые непосредственно определяют вирусную РНК, и тесты на коровый HCV антиген, которые являются анализом на вирусный антиген, вместо ответной реакции антитела. См., например, Kashiwakuma et al., Патент США №5,871,904; Beld et al.. Transfusion (2000) 40:575-579.
Тем не менее сохранилась необходимость в чувствительных и точных диагностических и прогностических средствах, позволяющих как организовать соответствующее лечение пациентов, так и предотвратить передачу HCV с кровью и продуктами крови или при близком личном контакте.
Краткое описание изобретения
Настоящее изобретение частично основано на открытии того, что отвечающие на HCV сывороточные антитела являются типично антикбровыми и анти-NS3 (хеликаза). Соответственно, изобретение связано с определением комплекса коревого антигена HCV и NS3-антитела, которое дает возможность обнаруживать как HCV-антигены, так и антитела, находящиеся в образце, с использованием отдельного твердого матрикса.
Таким образом, в одном из вариантов осуществления, предметом изобретения является твердая подложка для иммуноанализа, включающая в себя по меньшей мере одно HCV-антикоровое антитело и по меньшей мере один выделенный NS3/4a-эпитоп HCV, связанные с ней. Антитело и NS3/4а-эпитоп могут быть любыми из описанных здесь молекул. Кроме того, твердая подложка может включать в себя любой из описанных здесь слитых антигенов со множественными эпитопами, таких как слитый антиген со множественными эпитопами, включающий в себя аминокислотную последовательность, изображенную на фиг.7A-7F.
В определенных вариантах осуществления, твердая подложка включает в себя по меньшей мере два связанных с ней HCV-антикоровых антитела. Кроме того, антикоровое антитело может быть моноклональным антителом. К тому же NS3/4а-эпитоп может быть конформационным эпитопом, таким как конформационный NS3/4a-эпитоп, содержащий аминокислотную последовательность, изображенную на фиг.4A-4D.
В другом варианте осуществления, изобретение относится к твердой подложке для иммуноанализа, содержащей по меньшей мере два HCV-антикоровых моноклональных антитела и по меньшей мере один конформационный эпитоп NS3/4a HCV, содержащий аминокислотную последовательность, изображенную на фиг.4A-4D, связанные с ней.
В следующем варианте осуществления, изобретение относится к способу определения инфекции HCV в биологическом образце. Способ включает в себя в себя: (а) получение твердой подложки для иммуноанализа, как описано выше; (b) комбинирование биологического образца с твердой подложкой в условиях, которые позволяют HCV-антигенам и антителам, когда они присутствуют в биологическом образце, связываться по меньшей мере с одним антикоровым антителом и NS3/4a-эпитопом, соответственно; (с) добавление к твердой подложке со стадии (b) при комплексообразующих условиях (i) первого меченого антитела, где первое обнаруживаемое меченое антитело является антикоровым антителом HCV с обнаруживаемой меткой, где меченое антикоровое антитело направлено против другого коревого эпитопа HCV, чем по меньшей мере одно антикоровое антитело, связанное с твердой подложкой; (ii) антигена, который взаимодействует с HCV-антителом из биологического образца, реагирующим с NS3/4a-эпитопом и (iii) второго меченого антитела, где второе антитело с определяемой меткой может реагировать с антигеном (ii); и (d) определение комплексов, образованных между антителами и антигенами, если они присутствуют, в качестве указания на наличие инфекции HCV в биологическом образце. NS3/4a-эпитоп может быть конформационным эпитопом, таким как конформационный эпитоп, представленный последовательностью NS3/4a, изображенной на фигурах 4A-4D.
Еще в одном варианте осуществления, изобретение относится к,способу определения HCV инфекции в биологическом образце. Данный способ включает в себя в себя: (а) получение твердой подложки иммуноанализа как минимум с двумя связанными с ней антикоровыми антителами HCV, как было описано выше; (b) добавление биологического образца к твердой подложке в условиях, позволяющих антигенам и антителам HCV, в случае их присутствия в биологическом образце, связываться по меньшей мере с двумя антикоровыми антителами и NS3/4a-эпитопом, соответственно; (с) добавление к твердой подложке со стадии (b) в комплексообразующих условиях (i), первого антитела с обнаруживаемой меткой, где это первое антитело с обнаруживаемой меткой мечено обнаруживаемым образом антикоровым антителом HCV, где меченое антикоровое антитело направлено против иного коревого эпитопа HCV, чем антикоровые антитела, связанные с твердой подложкой; (ii) эпитопа из области с33с полипротеина HCV, соединенного с аминокислотной последовательностью hSOD; и (iii) второе антитело с обнаруживаемой меткой, где это второе антитело с обнаруживаемой меткой реагирует с аминокислотной последовательностью hSOD; и (d) определение комплексов, образованных между антителами и антигенами, если они присутствуют, в качестве указания на HCV-инфекцию в биологическом образце. NS3/4a-эпитоп может быть конформационным эпитопом, таким как конформационный эпитоп, представленный последовательностью NS3/4a, изображенной на фигурах 4A-4D.
В любом из вышеупомянутых вариантов осуществления, антикоровое антитело может быть направлено против N-концевой области HCV коревого антигена, такой как аминокислоты 10-53 HCV, пронумерованные относительно HCV1 полипротеиновой последовательности, и/или определяемо меченое HCV антикоровое антитело может быть направлено против С-концевой области HCV корового антигена, такого как аминокислоты 120-130 HCV, пронумерованные относительно HCV1 полипротеиновой последовательности. К тому же антиген, который реагирует с HCV-ангителом из биологического образца, может быть из NS3 области, например, эпитоп из с33с области полипротеина HCV, и может быть слит с аминокислотной последовательностью супероксиддисмутазы человека (hSOD). В этом варианте осуществления, второе антитело с обнаруживаемой меткой реагирует с аминокислотной последовательностью hSOD.
Еще в одном варианте осуществления изобретение направлено способ определения инфекции HCV в биологическом образце. Способ включает в себя в себя: (а) получение твердой подложки для иммунологического анализа, включающей в себя два HCV антикоровых моноклональных антитела и конформационный эпитоп, включающий в себя аминокислотную последовательность, изображенную на фигурах" 4A-4D; (b) добавление биологического образца к твердой подложке, в условиях, позволяющих антигенам и антителам HCV, когда они присутствуют в биологическом образце, связываться по меньшей мере с двумя антикбровыми антителами и NS3/4а конформационням эпитопом, соответственно; (с) добавление к твердой подложке со стадии (b) в комплексообразующих условиях (i) первого антитела с обнаруживаемой меткой, где это антитело с обнаруживаемой меткой обнаруживаемо помечено HCV антикоровым антителом, и где это антикоровое антитело с обнаруживаемой меткой направлено против другого коревого эпитопа HCV, чем как минимум два антикоровых антитела, связанных с твердой подложкой; (ii) эпитопа из области с33с полипротеина HCV, соединенного с аминокислотной последовательностью hSOD; и (iii) второго антитела с обнаруживаемой меткой, где это второе антитело с обнаруживаемой меткой реагирует с упомянутой аминокислотной последовательностью hSOD; (d) определение образующихся комплексов между антителами и антигенами, если они присутствуют, в качестве указания на присутствие HCV-инфекции в биологическом образце.
В некоторых вариантах осуществления, как минимум два антикоровых антитела направлены против N-концевой области коревого антигена HCV, например, против аминокислот 10-53 HCV, пронумерованных относительно полипротеина HCV1, и HCV-антикбровое антитело с обнаруживаемой меткой направлено против С-концевой области HCV коревого антигена, как, например, против аминокислот 120-130 HCV, пронумерованных относительно последовательности HCV1 полипротеина.
В следующем варианте осуществления, изобретение относится к способу определения HCV-инфекции в биологическом образце. Способ включает в себя: (а) получение твердой подложки для иммунологического анализа, которая включает в себя слитый антиген со множественными эпитопами; (b) комбинирование биологического образца с твердой подложкой в условиях, которые позволяют антигенам и антителам HCV, когда они присутствуют в биологическом образце, связываться по меньшей мере с одним антикоровым антителом, NS3/4a-эпитопом и слитым антигеном с множественными эпитопами; (с) добавление к твердой подложке со стадии (b) в комплексообразующих условиях (i) первого антитела с обнаруживаемой меткой, где это первое антитело с обнаруживаемой меткой является антикоровым антителом HCV с обнаруживаемой меткой, где это меченое антикоровое антитело направлено против другого HCV коревого эпитопа, чем по меньшей мере одно антикоровое антитело, связанное с твердой подложкой; (ii) первого и второго антигенов, которые реагируют с HCV-антителом из биологического образца, реагирующего с NS3/4a-эпитопом, и слитым антигеном со множественными эпитопами, соответственно; (iii) второго антитела с обнаруживаемой меткой, способного реагировать с антигенами из (ii); (d) определение комплексов, образовавшихся между антителами и антигенами, если они присутствуют, в качестве указания на присутствие инфекции HCV в биологическом образце.
Антикоровое антитело может быть направлено против N-концевой области HCV-корового антигена, и упомянутое первое HCV-антикоровое антитело с обнаруживаемой меткой может быть направлено против С-концевой области HCV-корового антигена, как описано выше. Кроме того, первый антиген, реагирующий с HCV-антителом из биологического образца, может включать в себя эпитоп из области с33с полипротеина HCV и может быть слит с аминокислотной последовательностью hSOD. Таким образом, второе определяемо меченое антитело реагирует с аминокислотной последовательностью hSOD. К тому же, второй антиген, реагирующий с HCV-антителом из биологического образца, может включать в себя эпитоп из с22 области полипротеина HCV, такой как эпитоп, включающий в себя аминокислоты Lys10-Ser99 долипротеина HCV, с делецией Аrg47 и заменой Trp на Leu в положении 44, пронумерованной относительно последовательности полипротеина HCV1. Эпитоп может быть слит с аминокислотной последовательностью hSOD. В этом случае второе антитело с обнаруживаемой меткой реагирует с аминокислотной последовательностью hSOD. Слитый антиген с множественными эпитопами может включать в себя аминокислотную последовательность, изображенную на фигурах 7A-7F.
Еще в одном варианте осуществления, изобретение относится к способу определения HCV инфекции в биологическом образце, причем указанный способ включает в себя: (а) получение твердой подложки для иммуноанализа, которая включает в себя два антикоровых моноклональных антитела HCV, NS3/4a-конформационный эпитоп HCV, включающий в себя аминокислотную последовательность, изображенную на фигурах 4A-4D, и слитый антиген с множественными эпитопами, содержащий аминокислотную последовательность, изображенную на фигурах 7A-7F, связанную с ним; (b) комбинирование биологического образца с твердой подложкой в условиях, которые позволяют антигенам и антителам HCV, когда они присутствуют в биологическом образце, связываться по меньшей мере с двумя антикоровыми антителами, NS3/4а конформационным эпитопом и слитым антигеном со множественными эпитопами, соответственно; (с) добавление к твердой подложке из стадии (b) при комплексообразующих условиях (i) первого антитела с обнаруживаемой меткой, где первое антитело с обнаруживаемой меткой является HCV антикоровым антителом с обнаруживаемой меткой, где меченое антикоровое антитело направлено против иного HCV-корового эпитопа, чем как минимум два антикоровых антитела, связанных с твердой подложкой; (ii) эпитопа из области с33с полипротеина HCV, слитого с аминокислотной последовательностью hSOD, и эпитопа из области с22 полипротеина HCV, слитого с аминокислотной последовательностью hSOD; и (iii) второго антитела с обнаруживаемой меткой, где указанное второе антитело с обнаруживаемой меткой способно реагировать с упомянутыми аминокислотными последовательностями hSOD; (d) определение комплексов, образующихся между антителами и антигенами, если они присутствуют, в качестве указания на присутствие HCV инфекции в биологическом образце.
В этом варианте осуществления, по меньшей мере два антикоровых антитела могут быть направлены против N-концевой области HCV-корового антигена, такой как аминокислоты 10-53 HCV, нумерованные относительно HCVl-полипротеина, и HCV-антикоровое антитело с обнаруживаемой меткой направлено против С-концевой области HCV-корового антигена, такой как аминокислоты 120-130 HCV, пронумерованные в соответствии с последовательностью полипротеина HVC1. К тому же, эпитоп из области с22 может включать в себя аминокислоты Lys10-Ser99, полипротеина HCV с делецией Аrg47 и заменой Trp на Leu в положении 44, пронумерованные соответственно с последовательностью полипротеина HVC1.
В других вариантах осуществления, изобретение относится к получению иммунодиагностического тестового набора, включающего в себя твердую подложку для иммуноанализа, описанную выше, и инструкции для проведения иммунодиагностического теста.
Еще в одном варианте осуществления, изобретение относится к способам получения твердой подложки для иммуноанализа, включающим в себя в себя: (а) получение твердой подложки; и (b) связывание с ней по меньшей мере одного HCV-антикорового антитела, например одного, двух или больше, и по меньшей мере одного выделенного NS3/4a-эпитопа HCV, и, необязательно, слитого антигена с множественными эпитопами. Антикоровые антитела, NS3/4a-эпитопы и слитые антигены с множественными эпитопами описаны выше.
В дополнительных вариантах осуществления, изобретение относится к слитому антигену со множественными эпитопами, включающему в себя в себя аминокислотную последовательность, изображенную на фигурах 7A-7F, или аминокислотную последовательность, идентичную ей по меньшей мере на 80%, например, идентичную на 90% или более и, кроме того, имеющего специфическую реакцию на анти-HVC антитела, находящиеся в биологическом образце от HCV-инфицированного субъекта. В определенных вариантах осуществления, слитый антиген с множественными эпитопами состоит из аминокислотной последовательности, изображенной на фигурах 5A-5F.
В последующих вариантах осуществления, изобретение относится к полинуклеотиду, включающему в себя кодирующую последовательность для вышеупомянутого слитого антигена со множественными эпитопами, рекомбинантным векторам, включающим в себя полинуклеотиды, клеткам-хозяевам, трансформированным рекомбинантными векторами, и способам получения рекомбинантного слитого антигена со множественными эпитопами, включающим в себя: (а) получение популяции вышеупомянутых клеток-хозяев; и (b) культурирование популяции клеток в условиях, при которых экспрессируется слитый антиген с множественными эпитопами, кодируемый кодирующей последовательностью, находящейся в рекомбинантном векторе.
Эти и другие аспекты настоящего изобретения станут очевидными при ознакомлении с нижеследующим детальным описанием и прилагаемыми чертежами.
Краткое описание чертежей.
Фигура 1 представляет собой графическое изображение генома HCV, изображающее различные области полипротеина, из которого получают реагенты для настоящего анализа (белки и антитела).
Фигура 2 представляет собой схематический рисунок иллюстративного комплекса антиген/антитело согласно изобретению.
На фигуре 3 изображена аминокислотная последовательность иллюстративного NS3/4a конформационного антигена, для использования в настоящих анализах. Выделенный жирным аланин в положении 182 заменяется природным серином, в норме находящимся в этом положении.
На фигурах 4A-4D изображают ДНК и соответствующую аминокислотную последовательность другого иллюстративного NS3/4a конформационного эпитопа для использования в настоящих анализах. Аминокислоты в положениях 403-404 фигур 4A-4D представляют собой замены Thr на Pro и Ser на Ile в природной аминокислотной последовательности HCV-1.
На фигуре 5 - диаграмма строения pd.HCV1a.ns3ns4aPI
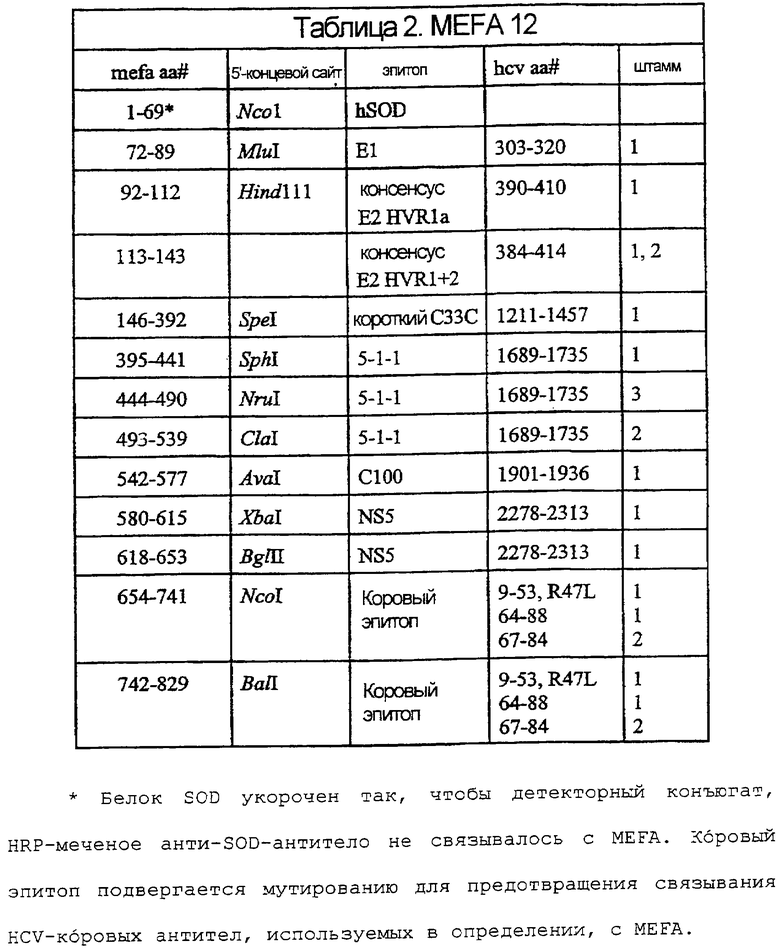
На фигуре 6 - графическое представление MEFA 12.
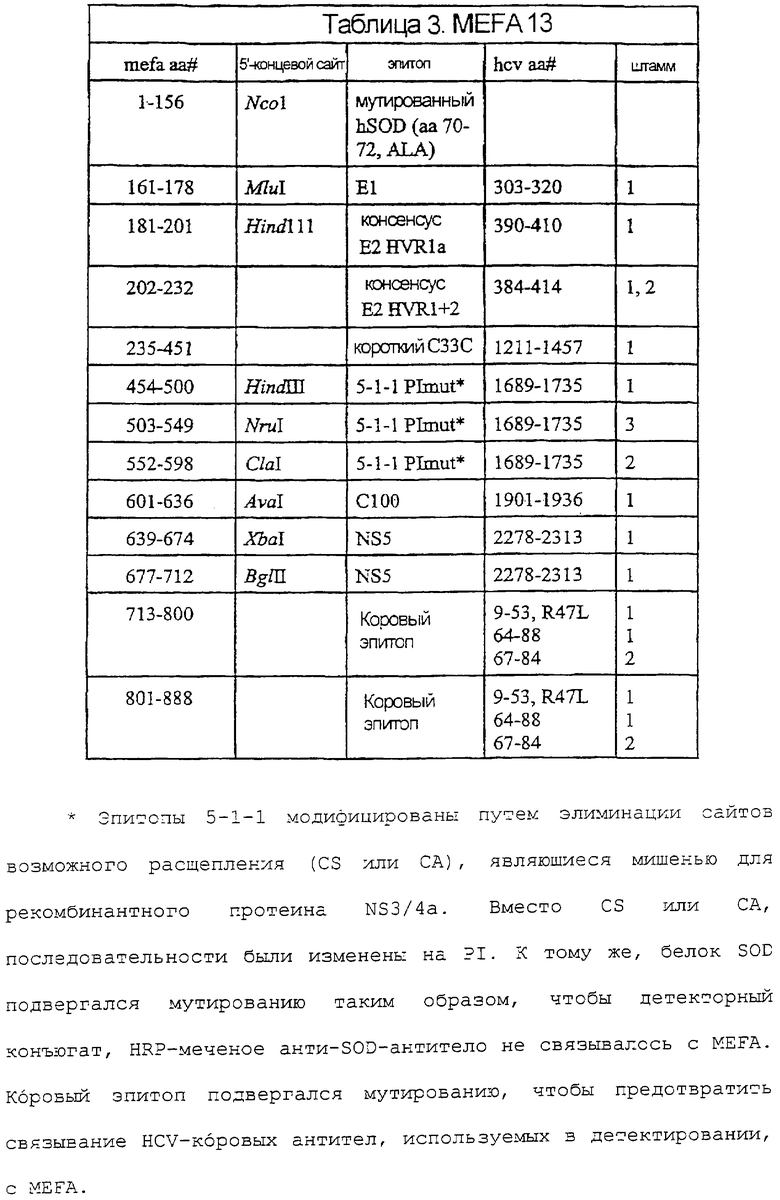
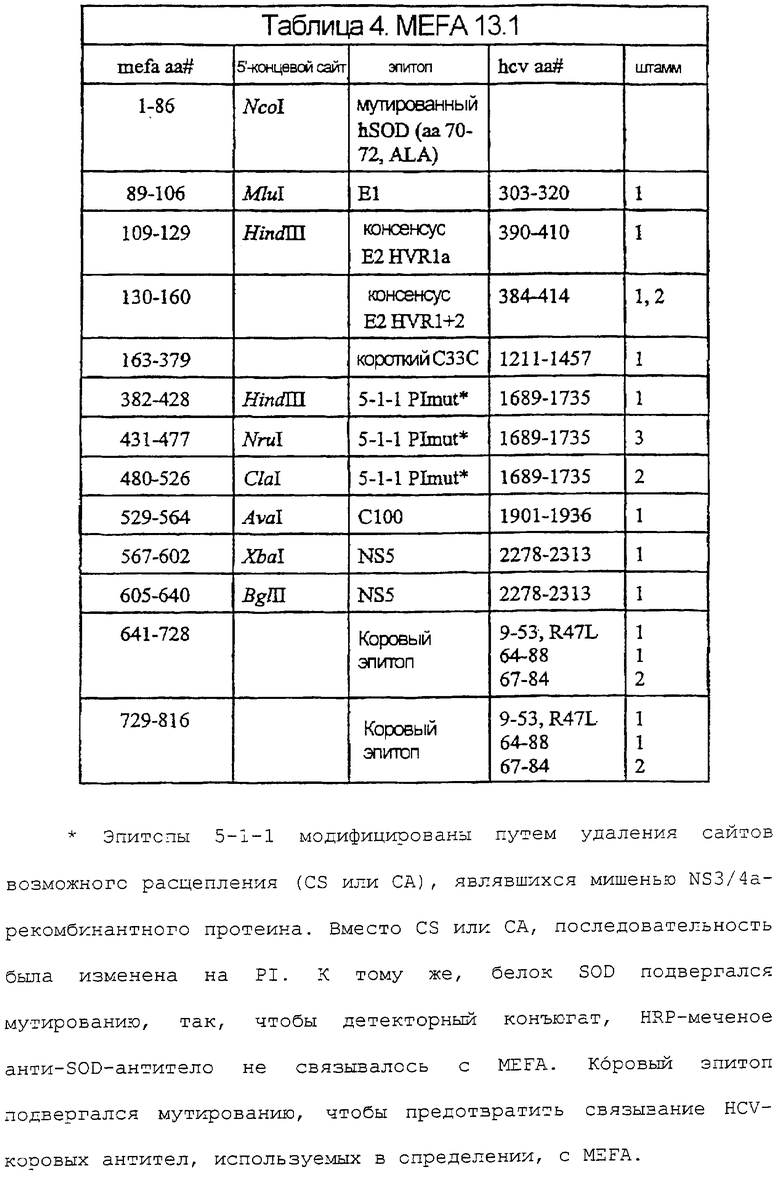
На фигурах 7A-7F изображены последовательность ДНК и соответствующая аминокислотная последовательность MEFA 12.
Фигура 8 является схематическим рисуноком иллюстративного иммунологического анализа согласно изобретению с использованием MEFA 12.
Подробное описание изобретения
В практическом применении настоящего изобретения будут использованы, если не указано иначе, обычные методы химии, биохимии, методики рекомбинации ДНК и иммунологии, в -пределах квалификации специалистов данной области. Такие методики полно представлены в литературе. См., например. Fundamental Virology, 2nd Edition, vol. I & II (B.N.Fields and D.M.Knipe, eds.); Handbook of Experimental Immunology, Vols. I-IV (D.M.Weir and C.C.Blackwell eds., Blackwell Scientific Publications); Т.Е.Creighton, Proteins: Structures and Molecular Properties (W.H.Freeman and Company, 1993); A.L.Lehninger, Biochemistry (Worth Publishers, Inc., current addition); Sambrook, et al., Molecular Cloning: A Laboratory Manual (2nd Edition, 1989); Methods In Enzymology (S.Colowick and N.Kaplan eds.. Academic Press, Inc.).
Необходимо отметить, что в данном описании и прилагаемой формуле изобретения формы единственного числа следует понимать во множественном числе, если содержание явно не диктует обратного. Так, например, «антиген» подразумевает смесь двух или более антигенов, и тому подобное.
В тексте использованы следующие сокращения названий аминокислот:
Определения
Ниже приводятся термины, используемые в описании настоящего изобретения, и их определения.
Термины «полипептид» и «белок» относятся к полимерам аминокислотных остатков и не ограничены минимальной длиной продукта. Таким образом, пептиды, олигопептиды, димеры, мультимеры и им подобное включены в это определение. Как полноразмерные белки, так и их фрагменты также включены в это определение.
Термины также включают в себя постэкспрессионные модификации полипептида, например гликозилрование, ацетилирование, фосфорилирование и им подобные. Кроме того, для целей данного изобретения, термин «полипептид» относится к белку, который содержит модификации в природной последовательности, такие как делеции, добавки или замены (как правило, консервативные по природе), до тех пор, пока этот белок сохраняет необходимую активность. Эти модификации могут быть созданы намеренно, например, при сайт-специфическом мутагенезе, или случайно, например, при мутациях у хозяина, продуцирующего белки, или ошибках ПЦР-амплификации.
Полипептид HCV является полипептидом, как определено выше, получаемым из полипротеина HCV. Нет необходимости в том, чтобы этот полипептид физически происходил из HCV, он может быть получен с помощью синтеза или рекомбинации. К тому же полипептид может быть получен из различных штаммов HCV, таких как штаммы HCV 1, 2, 3 или 4. У этих штаммов известен ряд сохраняющихся и вариабельных областей, и в целом аминокислотные последовательности эпитопов, происходящих из этих областей, будут иметь высокую степень гомологии последовательности, более 30%, предпочтительно более 40%, когда обе последовательности выровнены. Так, например, термин полипептид «NS3/4a» относится к природному NS3/4a любого из различных штаммов HCV, также как и аналоги NS3/4a, мутеины и иммуногенные фрагменты, как будет определено ниже. Известны полные генотипы многих из этих штаммов. См., например. Патент США №6150087 и GenBank поступление № AJ 238800 и AJ 238799.
Термины «аналог» и «мутеин» относятся к биологически активных производным базовой молекулы или фрагментов таких производных, которые сохраняют желаемую активность, такую как иммунореактивность в описанных здесь анализах. Обычно термин «аналог» относится к соединениям, имеющим природную полипептидную последовательность и структуру с одной или более аминокислотными вставками, заменами (обычно консервативными по природе) и/или делениями, по сравнению с природной молекулой, до тех пор, пока эти модификации не нарушают иммуногенной активности. Термин «мутеин» относится к пептидам, имеющим одну или более пептидных имитаций (пептоидов), таких, как описанные в Международной публикации № WO 091/04282. Предпочтительно, аналог или мутеин обладает по меньшей мере такой же иммуноактивностью, как и природная молекула. Способы создания аналогов и мутеинов полипептидов известны в данной области и описаны ниже.
Наиболее предпочтительные аналоги включают в себя замены, которые консервативны по природе, т. е. такие замены, которые происходят в пределах семейства аминокислот, родственного своими боковыми цепями. В частности, аминокислоты в целом разделены на четыре семейства: (1) кислотные - аспартат и глутамат; (2) основные - лизин, аргинин, гистидин; (3) неполярные - аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженные полярные глицин, аспарагин, глутамин, цистеин, серии, треонин, тирозин. Фенилаланин, триптофан и тирозин иногда классифицируют как ароматические аминокислоты. Например, вполне предсказуемо, что изолированная замена лейцина на изолейцин или валин, аспартата на глутамат, треонина на серии или подобная консервативная замена аминокислоты на структурно близкую аминокислоту не будет иметь большого эффекта на биологическую активность. Например, полипептид, представляющий интерес, может нести примерно до 5-10 консервативных и неконсервативных аминокислотных замен, или даже примерно до 15-25 консервативных или неконсервативных замен, или любое целое число между 5-25, при условии, что желаемая функция молекулы остается невредимой. Специалист в данной области может легко определить области в интересующей молекуле, допускающие замены, обратившись к схемам Hopp/Woods и Kyte-Doolittle, хорошо известным в данной области.
Термином «фрагмент» обозначен полипептид, состоящий только из части последовательности и структуры целого, полноразмерного полипептида. Этот фрагмент может включать в себя С-концевую делецию и/или N-концевую делецию природного полипептида. «Иммуногенный фрагмент» HCV - белка, в частности, обычно включает в себя не менее примерно 5-10 смежных аминокислотных остатков из полноразмерной молекулы, предпочтительно не менее 15-20 смежных аминокислотных остатков из полноразмерной молекулы, и наиболее предпочтительно - не менее примерно 20-50 или более смежных аминокислотных остатков из полноразмерной молекулы, определяющих эпитоп, или любое целое число от 5 аминокислотных остатков до полноразмерной последовательности, при условии, что рассматриваемый фрагмент сохраняет иммунореактивность в описанных здесь анализах. Например, предпочтительный иммуногенный фрагмент включает в себя, хотя и не ограничен ими, коровые фрагменты HCV, которые содержат, например, аминокислоты 10-45, 10-53, 67-88 и 120-130 полипротеина, эпитоп 5-1-1 (в NS3-области вирусного генома), также как и определенные эпитопы, полученные из областей Е1, Е2, с33с (NS3), c100 (NS4), NS3/4a и NS5 полипротеина HCV, a также любой из различных эпитопов, определенных в HCV-полипротеине. Например, см. Chien et al., Proc. Natl. Acad. Sci. USA (1992) 89:10011-10015; Chien et al., J. Gastroent. Hepatol. (1993) 8:S33-39; Chien et al., Международная публикация № WO 93/00365; Chien D. Y.,. Международная публикация № WO 94/01778; Патент США №6150087 и 6121020.
Термин «эпитоп», как он использован в данном описании, относится к последовательности, состоящей по меньшей мере примерно из 3-5 аминокислот, предпочтительно, примерно из 5-10 или 15, и не более, чем примерно из 1000 (или любое целое число в данном диапазоне), которые определяют последовательность, которая сама по себе или как часть большей последовательности связывается с антителом, которое генерируется в ответ на эту последовательность. Нет критического верхнего предела длины фрагмента, который может включать в себя почти полноразмерную белковую последовательность или даже слитый белок, включающий в себя два или больше эпитопов из полипротеина HCV. Эпитоп для использования в объекте изобретения не ограничен полипептидом, обладающим точной последовательностью той части родительского белка, из которой он был получен. Действительно, вирусные геномы находятся в состоянии постоянного изменения и содержат несколько вариабельных доменов, которые демонстрируют высокие уровни изменчивости между изолятами. Таким образом, термин «эпитоп» включает в себя как последовательности, идентичные природной последовательности, так и модификации природной последовательности, такие как делеции, вставки и замены (обычно консервативной природы).
Области данного полипептида, содержащие эпитоп, могут быть идентифицированы с использованием любого числа методик картирования эпитопов, хорошо известных в данной области. Например, см. Epitope Mapping Protocols in Methods in Molecular Bioligy, Vol. 66 (Glenn E. Morris, Ed., 1966) Humana Press, Totowa, New Jersey. В частности, линейные эпитопы можно определять с помощью, например, одновременного синтеза большого числа пептидов на твердых подложках, сопоставления этих пептидов с частями молекулы белка и проведения реакции этих пептидов с антителами, когда пептиды еще связаны с подложками. Подобные методики известны в данной области и описаны, в частности, Патент США 4,708,871; Geysen et al. (1984) Proc. Natl. Acad. Sci. USA 81:3998-4002; Geysen et al. (1985) Proc. Natl. Acad. Sci. USA 82:178-182; Geysen etal (1986) Molec. Immunol. 23:709-715. С использованием этой методики было определено число эпитопов HCV. Например, см. Chien et al., Viral Hepatitis and Liver Disease (1994) pp. 320-324, и далее ниже. Также конформационные эпитопы легко определяются при определении пространственной конформации аминокислот с помощью, например, рентгеновской кристаллографии и 2-мерного ядерного магнитного резонанса. Например, см. Epitope mapping protocol, supra. Антигенные области белка можно также определить, используя стандартные графики антигенности и гидропатии, такие как вычисляемые программой Omiga, версия 1.0, доступной в Oxford Molecular Group. В этой компьютерной программе используется метод Hopp/Woods, Hopp et al, Proc. Natl. Acad. Sci USA (1981) 78:3824-3828 для определения профилей антигенности, и методика Kyte-Doolittle, Kyte et al., J. Mol. Biol. (1982) 152:105-132 для графиков гидропатии.
Термин «конформационный эпитоп», используемый здесь, означает часть полноразмерного белка, или аналог, или мутеин, обладающий' природными структурными свойствами аминокислотной последовательности, кодирующей эпитоп в полноразмерном природном белке. Природные структурные свойства включают в себя, не ограничиваясь ими, гликозилирование и трехмерную структуру. Длина последовательности, определяющей эпитоп, может широко варьировать, так как считается, что эпитопы формируются трехмерной формой антигена (например, складчатостью). Таким образом, аминокислоты, определяющие эпитоп, могут быть сравнительно малочисленны, но широко рассеяны по длине молекулы (или даже разных молекул, в случае димеров и т.п.) и собираться в правильную конформацию эпитопа за счет складчатости. Части антигена между остатками, определяющими эпитоп, могут быть несущественными для конформационной структуры эпитопа. Например, делеция или замена этих промежуточных последовательностей может не влиять на последовательности, создающие конформационный эпитоп, необходимые для сохранения конформации эпитопа (например, цистеин, участвующий в дисульфидной связи; участки гликолизирования и т.д.).
Конформационный эпитоп, находящийся в - области NS3/4a, надежно определяется с использованием способов, обсуждаемых выше. Кроме того, присутствие или отсутствие конформационного эпитопа в данном полипептиде можно легко определить с помощью скрининга интересующего антигена с антителом (поликлональным сывороточным или моноклональным к конформационному эпитопу), и сравнения его реакционной способности с денатурированной версией антигена, который сохраняет только линейные - эпигоны (если есть). В подобном скрининге с использованием поликлональных антител можно вначале абсорбировать поликлональную сыворотку денатурированным антигеном и посмотреть, остались ли антитела к интересующему антигену. Кроме того, в случае NS3/4a, молекула, которая сохраняет природную конформацию, будет также обладать ферментативной активностью протеазы и, необязательно, хеликазы. Эти активности можно определить с помощью ферментативных анализов, как будет описано ниже.
Предпочтительно, конформационный эпитоп получают путем рекомбинации, и он выделяется клеткой, из которой он может быть извлечен в условиях, которые сохраняют его желаемые структурные особенности, т.е. без денатурации эпитопа. Эти клетки включают в себя клетки бактерий, дрожжей, насекомых и млекопитающих. Выделение и изоляция рекомбинантных конформационных эпитопов из полипротеина HCV описана, в частности, в Международных публикациях №№ WO 96/04301, WO 94/01778, WO 95/33053, WO 92/08734. В качестве альтернативы, можно выделять антигены и затем ренатурировать белок после выделения. Также понятно, что химический синтез также позволяет получать мимитопы конформационного антигена, которые перекрестно реагируют с конформационным эпитопом «природного» антигена.
Термин «слитый антиген с множественными эпитопами», или "MEFA", используемый здесь, означает полипептид, в котором множественные антигены HCV являются частью единой, непрерывной цепи аминокислот, которая не встречается в природе. HCV-антигены могут быть соединены непосредственно друг с другом пептидными связями или могут быть разделены промежуточными аминокислотными последовательностями. Слитые антигены могут также содержать последовательности, экзогенные по отношению к полипротеину HCV. Более того, присутствующие последовательности HCV могут происходить из разнообразных генотипов и/или изолятов HCV. Примеры конкретных МЕРА для использования в настоящих иммунологических анализах детализированы, например, в Международной публикации № WO 97/44469 и описаны ниже.
«Антитело» означает молекулу, которая по химическим или физическим причинам специфически связывается с представляющим интерес полипептидом. Таким образом, HCV-коровое антитело является молекулой, которая специфически связывается с HCV коровым белком. Термин «антитело», как он использован здесь, включает в себя антитела, полученные как из поликлональных, так и моноклональных препаратов, так же, как следующее: гибридные (химерные) молекулы антител (см., например. Winter et al. (1991) Nature 349: 293-299; Патент США №4816567); F(ab')2- и F(ab)-фрагменты, Fv-молекулы (не-ковалентные гетеродимеры, см., например, Inbar et ai. (1972) Proc Nati Acad Scl USA 69:2659-2662; and Ehrlich et al. (1980) Biochem 19:4091-4096); одноцепочечные Fv молекулы (sFv) (см.,например, Huston et ai. (1988) Proc Nati Acad Sci USA 85:5879-5883); димерные и гримерные конструкции фрагментов антител, минитела (см., например. Pack et al. (1992) Blochem 31:1579-1584; Cumber et al. (1992) J Immunology 1498:120-126); гумманизированные молекулы антител (см., например, Riechmann et al. (1988) Nature 332: 323-327; Verhoeyan et al. (1988) Science 239:1534-1536; и Патентная публикация Англии № GB 2276169, опубликованная 21 сентября 1994 г.); и любые функциональные фрагменты, полученные из таких молекул, которые сохраняют иммунологические связывающие свойства родительской молекулы антитела.
Термин «моноклональное антитело», как он используется здесь, относится к композиции антител, имеющих гомогенную популяцию антител. Этот термин не ограничен ни видом или источником антител, ни способом их получения. Так, этот термин включает в себя антитела, полученные из мышиных гибридом, так же, как и моноклональные антитела, полученные при использовании скорее человеческих, чем мышиных гибридом. Например, см., Cote, et al. Monclonal Antibodies and Cancer Therapy, Alan R. Liss, 1985, p. 77.
«Рекомбинантный» белок - это белок, сохраняющий желательную активность и полученный с помощью методики рекомбинации ДНК, как описано здесь. В целом, представляющий интерес ген клонируют и затем экспрессируют в трансформированном организме, как описано ниже. Организм хозяина экспрессирует чужеродный ген для продуцировакия белка в условиях экспрессии.
Когда имеют в виду полипептид, под «выделенным» подразумевают, что указанная молекула выделена и отделена от целого организма, в котором эта молекула обнаружена в природе, или она находится в значительной степени в отсутствие других биологических макромолекул того же типа. Термин «выделенный» в отношении полинуклеотида означает молекулу нуклеиновый кислоты, лишенную, полностью или частично, последовательностей, обычно связанных с ней в природе; или последовательность, такую, как она существует в природе, но включающую в себя гетерологичную последовательность, с которой она связана; либо молекулу, диссоциированную с хромосомой.
Под «эквивалентной антигенной детерминантой» следует понимать антигенную детерминанту из другого подвида или штамма HCV, такого как HCV-штаммы 1, 2 или 3. Более конкретно, известны эпитопы, такие как 5-1-1, и эти эпитопы варьируют у штаммов 1, 2 и 3. Таким образом, эпитопы 5-1-1 из трех разных штаммов являются эквивалентными антигенными детерминантами и, таким образом, являются «копиями», даже если их последовательности и неидентичны. В целом, аминокислотные последовательности эквивалентных антигенных детерминант будут иметь высокую степень гомологии последовательности, например гомология аминокислотной последовательности более 30%, предпочтительно более 40%, когда две последовательности выровнены.
«Гомология» означает процент сходства между двумя полинуклеотидными или двумя полипептидными последовательностями. Две ДНК или две - полипептидные последовательности являются «существенно гомологичными» по отношению друг к другу, когда последовательности демонстрируют по меньшей мере около 50%, предпочтительно, по меньшей мере около 75%, более предпочтительно - по меньшей мере около 80-85%, и наиболее предпочтительно - по меньшей мере около 95-98% сходства последовательностей на протяжении определенной длины молекул. «Существенно гомологичный» здесь также относится к последовательностям, демонстрирующим полную идентичность с определенными ДНК или полипептидными последовательностями.
Обычно «идентичность» относится в точном нуклеотид-нуклеотид или аминокислота-аминокислота соответствии двух полинуклеотидных или полипептидных последовательностей, соответственно. Процент идентичности можно определить путем прямого сравнения информации о последовательности между двумя молекулами при помощи выравнивания последовательностей, подсчета точного числа соответствий между двумя выровненными последовательностями, деления на длину более короткой последовательности и умножения на 100.
Для помощи в анализе сходства и идентичности можно использовать легко доступные компьютерные программы, такие как ALIGN, Dayhoff, M.O. in Atlas of Protein Sequence and. Structure M.O. Dayhoff ed., 5 Suppi. 3:353-358, National biomedical Research Foundation, Washington, DC, которая адаптирует алгоритм локальной гомологии Смита и Ватермана (Smith & Waterman Advances in Appl. Math. 2: 482-489, 1981) для анализа пептидов. Программы для определения сходства и идентичности, нуклеотидных последовательностей доступны в Wisconsin Sequence Analysis Package, версия 8 (доступна от Genetics Computer Group, Madison, WI), например, программы BESTFIT, FASTA и GAP, в которых также использован алгоритм Смита и Ватермана. Эти программы легко применяются с параметрами по умолчанию, рекомендованными производителем и описаны в Wisconsin Sequence Analysis Package, упомянутом выше. Например, процентное сходство конкретной нуклеотидной последовательности с контрольной последовательностью можно определить, используя алгоритм гомологии Смита и Ватермана с установленными по умолчанию оценочной таблицей и отрицательным счетом за промежуток в шесть нуклеотидных положений.
Еще одним способом установления процентного сходства в контексте настоящего изобретения является использование пакета программ MPSRCH, с авторскими правами Университета Эдинбурга, разработанного John F.Collins and Shane S.Sturrok и распространяемого IntelliGenetics, Inc. (Mountain View, CA). У этого набора программ алгоритм Смита и Ватермана используется при параметрах по умолчанию, установленных для оценочной таблицы (например, отрицательный счет -12 при открытом промежутке, отрицательный счет - 1 при расширенном промежутке и промежуток - 6 положений). В полученных данных значение "Match" отражает «сходство последовательностей». Другие подходящие программы для расчета процентной идентичности или сходства между последовательностями, в целом, известны в данной области, например, еще одной программой выравнивания является программа BLAST, используемая с параметрами по умолчанию. Например, BLASTN и BLASTP можно использовать со следующими параметрами по умолчанию: genetic code = standard; filter = none; strand = both; cutoff = 60; expect = 10; Matrix = BLOSUM62; Descriptions = 50 sequences; sort by = HIGH SCORE; Databases = non-redundant, GenBank + EMBL + DDBJ + PDB + GenBank CDS translations + Swiss protein + Spupdate + PIR. Подробности этих программ можно найти по следующему Интернет-адресу: http://www.ncbi.nlm.gov/cgi-bin/BLAST.
В качестве альтернативы, гомологию можно определять путем гибридизации полинуклеотидов в условиях, когда образуются стабильные дуплексы между гомологичными регионами, с последующим перевариванием одноцепочечно-специфической нуклеазой (нуклеазами) и определением размеров переваренных фрагментов. Существенно гомологичные ДНК-последовательности можно определять при помощи эксперимента по Саузерн-блот-гибридизации, например, в жестких условиях, как определено в этой конкретной системе. Определение соответствующих условий гибридизации находится в пределах квалификации специалистов данной области. Например, см. Sambrook et al., supra; DNA Cloning, supra; Nucleic Acid Hybridization, supra.
«Кодирующая последовательность» или последовательность, которая «кодирует» избранный полипептид, - это молекула нуклеиновой кислоты, которая транскрибируется (в случае ДНК) или транслируется (в случае мРНК) в полипептид in vivo или in vitro, будучи помещенной под контроль соответствующих регуляторных последовательностей. Границы кодирующей последовательности определены стартовым кодоном на 5'-(амино) конце и кодоном остановки трансляции на 3'-(карбокси) конце. Последовательность, останавливающая транскрипцию, может находиться в интервале от 3' конца до кодирующей последовательности.
«Оперативно связанный» относится к расположению элементов, при котором компоненты, охарактеризованные таким образом, скомпонованы так, чтобы выполнять требуемую функцию. Так, данный промотор, оперативно связанный с кодирующей последовательностью, способен воздействовать на экспрессирование кодирующей последовательности, когда присутствуют надлежащие трансляционные факторы и т.п. Промотор необязательно должен граничить с кодирующей последовательностью, поскольку его функция состоит в управлении ее экспрессией. Так, например, промежуточные нетранслируемые, хотя и транскрибируемые, последовательности могут находиться между промоторной последовательностью и кодирующей последовательностью, как, например, транскрибируемые интроны, и промоторная последовательность может тем не менее считаться «оперативно связанной» с кодирующей последовательностью.
Термин «контролирующий элемент» относится к полинуклеотидной последовательности, которая способствует экспрессии кодирующей последовательности, с которой она связана. Термин включает в себя промоторы, последовательности, терминирующие транскрипцию, вышерасположенные регуляторные домены, сигналы полиаденилирования, нетранслируемые области, включая 5'-UTR и 3'-UTR, и, при соответствующих условиях, лидерные последовательности и энхансеры, которые коллективно обусловливают транскрипцию и трансляцию кодирующих последовательностей в клетке-хозяине.
Термин «промотор», используемый здесь, относится к регуляторной области ДНК, способной связываться с РНК-полимеразой в клетке-хозяине и инициировать транскрипцию нижележащей (в направлении 3') кодирующей последовательности, оперативно с ней связанной. Для целей настоящего изобретения промоторная последовательность включает в себя минимальное число оснований или элементов, необходимых для инициации транскрипции гена, представляющего интерес, на уровне, обнаруживаемом выше фона. Промоторная последовательность содержит сайт, инициирующий транскрипцию также, как и белок-связывающие домены (консенсусные последовательности), ответственные за присоединение РНК-полимеразы. Промоторы эукариот часто, но не всегда, содержат «ТАТА»-боксы и «САТ»-боксы.
Управляющая последовательность «руководит транскрипцией» кодирующей последовательности в клетке, когда РНК-полимераза будет присоединять промоторную последовательность и транскрибировать кодирующую последовательность в мРНК; которая затем транслируется в полипептид, кодируемый кодирующей последовательностью.
"Кассета экспрессии или "экспрессирующая конструкция" относится к совокупности последовательностей, которая способна управлять экспрессией представляющей интерес последовательности (последовательностей) или гена (генов). Кассета экспрессии включает в себя регуляторные элементы, как было описано выше, такие как промотор, который оперативно связан (для того чтобы управлять их транскрипцией) с представляющей интерес последовательностью (последовательностями) или геном (генами), и часто также включает в себя полиаденилированные последовательности. В некоторых вариантах осуществления изобретения описанная здесь кассета экспрессии может находиться внутри плазмидной конструкции. В добавление к компонентам кассеты экспрессии плазмидная конструкция может также содержать один или более выбираемых маркеров, сигнал, который позволяет плазмидной конструкции существовать в виде одноцепочечной ДНК (например, точка начала репликации М13), по меньшей мере один многократно клонирующий сайт, и точка начала репликации «млекопитающего» (например, SV40 или точка начала репликации аденовируса).
Термин «трансформация», как он использован здесь, относится к вставке экзогенного полинуклеотида в клетку-хозяина, безотносительно к способу, использованному для вставки: например, трансформация путем прямого поглощения, трансфицирования, инфицирования и т.п. Об отдельных методах трансфекции, см. ниже. Экзогенные полинуклеотиды могут содержаться в виде неинтегрированного вектора, например, как эписома, или - как альтернатива - может быть интегрирован в геном хозяина.
«Клетка-хозяин» - это клетка, которая трансформирована или способна к трансформации экзогенной последовательностью ДНК.
«Обычная твердая подложка» означает отдельную твердую основу с которой HCV-полипептиды, использующиеся в целях иммунологических анализов, связаны ковалентной связью или нековалентным способом, таким как гидрофобная адсорбция.
«Иммунологически реактивный» означает рассматриваемый антиген, который специфически реагирует с анти-НСУ-антителами, находящимися в биологическом образце, полученном у HVC-инфицированного субъекта.
«Иммунный комплекс» означает комплекс, образовавшийся, когда антитело связывается с эпитопом антигена.
Термин «биологический образец», как он используется здесь, относится к образцу ткани или жидкости, полученному у субъекта, включающему в себя, но не ограниченного ими, например, кровь, плазму, сыворотку, фекальные массы, мочу, костный мозг, желчь, спинномозговую жидкость, лимфу, образцы кожи, наружные выделения кожи, дыхательной системы, кишечного и мочеполового трактов, слезы, слюну, молоко, клетки крови, органы, биопсии и также образцы компонентов культуры клеток in vitro, включающие в себя, но не ограниченные ими, кондиционированную среду, полученную в результате роста клеток и тканей в культуральной среде, например, рекомбинантные клетки и клеточные компоненты.
Термины «метка» и «обнаруживаемая метка» относятся к молекуле, которую возможно обнаружить, включая в себя, но не ограничиваясь ими: радиоактивные изотопы, люминофоры, хемилюминесцентные вещества, хромофоры, ферменты, субстраты ферментов, кофакторы ферментов, ингибиторы ферментов, хромофоры, красители, ионы металлов, золи металлов, лиганды (например, биотин, стрепавидин или гаптены) и им подобные. Термин «люминофор» относится к веществу или его дозе, которая способна флуоресцировать а регистрируемом диапазоне. Частные примеры меток, которые можно использовать в данном изобретении, включают в себя, не ограничиваясь ими: пероксидазу хрена (HRP), флуоресцеин, FITC, родамин, дансил, умбеллиферон, диметилакридиновый эфир (DMAE), Техасский красный, люминол, НАДФН и α-β-галактозидазу.
II. Варианты осуществления изобретения
Прежде чем детально описывать настоящее изобретение, необходимо понять, что данное изобретение не ограничивается частными рецептурами или параметрами процесса, поскольку они, естественно, могут варьировать. Также должно быть ясно, что терминология, используется здесь только для описания конкретных вариантов осуществления данного изобретения, и не должна трактоваться как ограничительная.
Хотя в практической реализации данного изобретения можно использовать различные композиции и способы, подобные или эквивалентные описанным здесь, в тексте описаны предпочтительные материалы и способы.
Как отмечалось выше, настоящее изобретение основано на открытии новых диагностических способов точного определения ранней инфекции HCV. Способы основаны на идентификации и использовании высокоиммуногенных антител и антигенов HCV, которые присутствуют на ранних стадиях HCV-сероконверсии, таким образом увеличивая точность определения и уменьшая долю ошибочных результатов. Данные способы могут быть легко применены в виде единого анализа.
Конкретнее, анализ проводится на твердой подожке, с которой сшиты одно или более HCV-антикоровых антител (направленных против либо одного и того же, либо различных HCV-коровых эпитопов), и эпитоп, полученный из NS3/4a-области полипротеина HCV. Примеры конкретных антикоровых антител, применимых в настоящем изобретении, включают в себя, не ограничиваясь ими, молекулы антител, такие как моноклональные антитела, направленные против эпитопов в коровой области, найденной между аминокислотами 10-53, аминокислотами 10-45, аминокислотами 67-88, аминокислотами 120-130, или антитела, направленные против любых из коровых эпитопов, установленных, например, в: Houghton et ai., Патент США №5350671; Chien et al., Proc. Natl. Acad. Sci. USA (1992) -89:10011-10015; Chien et al., J. Gastroent. Hepatol. (1993) 8:S33-39; Chien et al., Международная публикация № WO 93/00365; Chien, D. Y., Международная публикация № WO 94/01778; Патентная заявка США с одним правопреемником. Сер. №08/403590 и 08/444818.
Описана NS3/4a-область полипротеина HCV, и аминокислотная последовательность и полная структура белка описана, например, в Yao et al.. Structure (November 1999) 7:1353-1363; Sali et al., Blochezn. (1998) 37:3392-3401; and Bartenschlager, R., J.Viral Hepat. (1999) 6:165-181. Также см. Dasmahapatra et al., Патент США №5843752. Заявленью иммунологические анализы используют по меньшей мере один конформационный эпитоп, полученный из NS3/4a-области, найденной в природной частице HCV или ее инфицирующем продукте, что подтверждается сохранением протеазной или - необязательно - хеликазной активности, в норме демонстрируемой генным продуктом NS3/4a и/или иммунореактивностью антигена с антителами в образце, полученном у HCV-инфицированного субъекта, и потерей иммунореактивности эпитопа при денатурации антигена. Например, конформационный эпитоп можно разрушить путем нагревания, изменения рН в сторону очень кислых или щелочных значений или добавления известных органических денатураторов, таких как дитиотреитол (DTT), или подходящего детергента. Например, см. Protein Purification Methods, a practical approach (E.L.V.Harris and S.Angal eds., IRL Press), и денатурированный продукт сравнивают с продуктом, который не обрабатывали, как описано выше.
Протеазную и хеликазную активности можно определить, используя стандартные ферментные анализы, хорошо известные в данной области. Например, протеазную активность можно определить, используя анализы, хорошо известные в данной области. Например, Takeshita et al. Anal. Biochem. (1997) 241:242-246: Kakiuchi et al., J. Biochem. (1997) 122:749-755; Sali et al., Biochemistry (1998) 37:3392-3401; Cho et al., J. Virol. Meth. (1998) 72:109-115; Cerretani et al., Anal. Biochem. (1999) 266:192-197; Zhang et al., Anal. Biochem. (1999) 270:268-275; Kakiuchi et al., J. Virol. Meth. (1999) 80:77-84; Fowler et al., J. Blomol. Screen. (2000) 5:153-158; and Kim et al., Anal. Biochem. (2000) 284:42-48. Конкретный подходящий анализ для тестирования протеазной активности описан далее в примерах.
Точно так же, анализы хеликазной активности хорошо известны в данной области и хеликазную активность NS3/4a-эпитопа можно определять, используя, например, анализ ELISA, как описано, например, в Hsu et al., Biochem. Blophys. Res. Common. (1998) 253:594-599; систему анализа сцинтилляционного сходства, как описано у Kyono et al., Anal. Biochem. (1998) 257;120-126; анализы на основе скрининга высокой пропускной способности, как писано например, в Hicham et al., Antiviral Res. (2000) 466:181-193 and Kwong et al., Methods Mol. Med. (2000) 24:97-116; также как и с помощью других аналитических методов, известных в данной области. Например, Khu et al., J. Virol. (2001) 75:205-214; Utama et al.. Virology (2000) 273:316-324; Paolini et al., J. Gen. Virol. (2000) 81;1335-1345; Preugschat et al., Biochemistry (2000) 39:5174-5183; Preugschat et al., Methods Mol. Med. (1998) 19:353-364; and Hesson et al., Biochemistry (2000) 39:2619-2625.
Длина данного антигена достаточна для того, чтобы содержать иммунореактивный конформационный эпитоп. Часто полипептид, содержащий используемый антиген, может быть полноразмерным, однако полипептид может быть и укороченным, например, для увеличения растворимости или для улучшения секреции. Как правило, конформационный эпитоп, найденный в NS3/4a, экспрессируется в клетке как рекомбинантный полипептид, и этот полипептид образует эпитоп в требуемой форме, как будет описано в деталях ниже.
Иллюстративные аминокислотные последовательности NS3/4a полипептидов показаны, в фигуре 3 и фигурах 4A-4D. Выделенный жирным аланин, занимающий положение 182 на фигуре 3, заменяет природный серин, найденный в этом положении, для того, чтобы предотвратить автокатализ молекулы, который в противном случае может произойти. Аминокислотная последовательность, показанная в положении 2-686 в фигурах 4A-4D, соответствует аминокислотным положениям 1027-1711 HCV-1. Инициирующий кодон (ATG), кодирующий Met, показан в позиции 1. Кроме того, Thr, в норме встречающийся в положении 1428 HCV-1 (аминокислотное положение 403 на фигуре 4) изменяется на Pro, и Ser, в норме встречающийся в положении 1429 HCV-1 (аминокислотное положение 404 на фигуре 4) изменяется на Ile. Однако либо природная последовательность, имеющая или не имеющая N-концевой Met, изображенный аналог, имеющий или не имеющий N-концевой Met, либо другие аналоги или фрагменты могут быть использованы в заявленных анализах, при условии, что эпитоп получают с использованием метода, который сохраняет или восстанавливает его природную конформацию, такую как протеазную активность, и - необязательно - сохраняется хеликазная активность. В обоих патентах: Dasmahapatra et al., Патент США №5843752 и Zhang et al. Патент США №5990276 - описываются аналоги NS3/4a.
NS3-протеаза из NS3/4a найдена приблизительно в положениях 1027-1207, пронумерованных соответственно HCV-1, положениях 2-182 на фиг.4. Структура NS3-протеазы и ее активного центра известны. Например, см. De Francesco et al., Antivir. Ther. (1998) 3:99-109; Koch et al., Biochemistry (2001) 40:631-640. В норме допускаются изменения в природной последовательности, которые находятся вне активного центра молекулы. В частности, желательно сохранять аминокислоты с 1-й или 2-й по 155-ю (фиг.4), с небольшим числом замен или только консервативными заменами. Аминокислоты после 155-й допускают бóльшие изменения. Кроме того, если используют фрагменты последовательности NS3/4a, изображенной на фиг.4, эти фрагменты должны, как правило, включать в себя по меньшей мере аминокислоты 1- или 2-155, предпочтительно аминокислоты 1- или 2-175, и наиболее предпочтительно аминокислоты 1- или 2-182, с N-концевым Met или без него. Домен хеликазы обнаружен в области положений 1193-1657 HCV-1 (положения 207-632, фигура 4). Таким образом, если необходима хеликазная активность, эта часть молекулы должна быть сохранена с небольшими или только консервативными изменениями. На основе известной структуры NS3/4a специалист в данной области легко определит другие области, в которых изменения допустимы.
Твердая подложка может также включать в себя другие антигены. Например, слитые антигены с множественными эпитопами (обозначенные «MEFA»), как это описано в Международной публикации WO/44469, которые могут быть пришиты к твердой подложке для использования в заявленных анализах. Подобные MEFA содержат множественные эпитопы, полученные из двух или более разных вирусных областей, показанных на фигуре 1 и в таблице 1. В частности, как показано в фиг.1 и таблице 1, при расщеплении полипротеин HCV образует не менее 10 отдельных продуктов в порядке NH2-Kop-E1-E2-p7-NS2-NS3-NS4a-NS4b-NS5a-NS5b-COOH. Коровый полипептид находится в положениях 1-191, пронумерованных относительно HCV-1 (см. Choo et al. (1991) Proc. Natl. Acad. Sci. USA 88:2451-2455, для HCV-1 генома). Этот полипептид в дальнейшем обрабатывается для получения HCV приблизительно с аминокислотами 1-173. Полипептиды оболочки Е1 и Е2 находятся примерно в положениях 192-383 и 384-746, соответственно. Домен Р7 найден примерно в положениях 747-809. NS2 - интегральный мембранный протеин с протеолитической активностью обнаружен примерно в положениях полипротеина 810-1026. NS2, как один, так и в комбинации с NS3 (находящийся примерно в положениях 1027-1657) расщепляет слабую связь NS2-NS3, которая, в свою очередь, образует N-конец NS3 и высвобождает большой полипротеин, включающий в себя активности как сериновой протеазы, так и РНК-хеликазы. Протеаза NS3, обнаруженная примерно в положениях 1027-1207, служит для обработки остающегося полипротеина. Хеликазная активность обнаруживается в районе положений 1193-1657. Завершение созревания полипротеина инициируется автокаталитическим расщеплением связи NS3-NS4a, катализируемое сериновой протеазой NS3. Обнаружено, что NS3-опосредственные расщепления полипротеина HCV включают в себя распознавание расщепляемых связей полипротеина молекулой NS3 другого полипептида. В этой реакции NS3 высвобождает кофактор NS3 (NS4a, 1658-1711), два белка (NS4b, отмеченный примерно в положениях 1712-1972, и NS5a, находящийся примерно в положениях 1973-2420) и РНК - зависимую РНК-полимеразу (NS5b, находящуюся примерно в положениях 2421-3011).
Множественные HCV-антигены являются частью единой, непрерывной цепи аминокислот, которая не встречается в природе. Таким образом, линейный порядок эпитопов отличается от их линейного порядка в геноме, в котором они встречаются. Предпочтительно, чтобы линейный порядок последовательностей MEFA, используемых здесь, был аранжирован для получения оптимальной антигенности. Предпочтительно, чтобы эпитопы были более чем из одного штамма HCV, обеспечивая таким образом дополнительную возможность обнаруживать разные штаммы HCV в одном анализе. Таким образом MEFA, используемые здесь, могут содержать различные иммуногенные области, полученные из описанного выше полипротеина. К тому же, белок, полученный при сдвиге рамки считывания в коровой области полипротеина, такой, как описанный в Международной публикации № WO 99/63941, может быть использован для MEFA. При желании по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более одного или более эпитопов, полученных из HCV-полипротеина, могут находиться в слитом белке.
Например, эпитопы, полученные, в частности, из сверхвариабельной области Е2, такой как область, охватывающая аминокислоты 384-410 или 390-410, могут быть включены в MEFA-антиген. Особенно эффективный эпитоп Е2 является одним из тех, которые включают в себя консенсусную последовательность, полученную из этой области, такую, как консенсусная последовательность Gly-Ser-Ala-Ala-Arg-Thr-Thr-Ser-Gly-Phe-Val-Ser-Leu-Phe-Ala-Pro-Gly-Ala-Lys-Gln-Asn, которая представляет собой консенсусную последовательность аминокислот 390-410 генома HCV типа 1. Иллюстративный Е2-эпитоп, представленный в MEFA согласно изобретению, может содержать гибридный эпитоп, охватывающий аминокислоты 390-444. Подобный гибридный Е2-эпитоп может включать в себя консенсусную последовательность, представляющую аминокислоты 390-410, слитые с природной аминокислотной последовательностью 411-444 из Е2 HCV.
Кроме того, антигены можно получать из различных штаммов HCV. Известно много вирусных штаммов HCV, и в слитом белке можно использовать эпитопы, полученные из этих штаммов. Хорошо известно, что любой вид организмов варьирует от одной особи к другой и, более того, что каждый организм, включая вирус, может иметь ряд различных пород (штаммов). Например, как показано выше, HCV включает в себя по меньшей мере 6 генотипов. Каждый из этих генотипов включает в себя эквивалентные антигенные детерминанты. Более конкретно, каждый штамм содержит ряд антигенных детерминант, которые находятся во всех штаммах вируса, но слегка варьируют от одного вирусного штамма к другому. Например, HCV содержит антигенную детерминанту, известную как 5-1-1 (см. фиг.1). Эта конкретная антигенная детерминанта встречается в трех различных формах в трех разных вирусных штаммах HCV. Соответственно, в предпочтительном варианте осуществления данного изобретения все три формы 5-1-1 присутствуют в слитом антигене со множественными эпитопами, используемом в заявленных иммунологических анализах. Точно так же, могут присутствовать эквивалентные антигенные детерминанты из коровых областей различных штаммов HCV. Обычно эквивалентные антигенные детерминанты обладают высоким уровнем гомологии в отношении аминокислотной последовательности, их уровень гомологии, когда они выровнены, обычно составляет 30% или более, предпочтительно 40% или более. Мультикопийный эпитоп согласно изобретению может также включать в себя множественные копии, являющиеся точной копией одного и того же эпитопа.
Иллюстративные MEFA, для применения в анализах согласно изобретению, описаны в международной публикации № WO 97/44469. Дополнительные иллюстративные MEFA, используемые здесь, включают в себя MEFA 12, MEFA 13 и MEFA 13.1. Надо понимать, что эти MEFA показаны только в качестве образца, и другие эпитопы, полученные из генома HCV, также могут быть использованы в данных анализах и могут быть встроены в эти или другие MEFA.
Последовательность ДНК и соответствующая аминокислотная последовательность MEFA 12 показаны на фигурах 7A-7F. Общая структурная формула MEFA 12, показанная на фиг.6, выглядит следующим образом: hSOD-E1(тип1)-Е2 HVR консенсус(тип 1а)-Е2 HVR консенсус(типы 1 и 2)-с33с короткий(тип 1)-5-1-1(тип 1)-5-1-1(тип 3)-5-1-1(тип 2)-с100(тип 1)-NS5(тип l)-NS5(тип 1)-кор(типы 1 и 2) - кор(типы 1 и 2). Этот мультикопийный эпитоп включает в себя следующую аминокислотную последовательность, пронумерованную соответственно HCV-1 (нумерация аминокислот, данная ниже, следует нумерации, приведенной Choo, et al. (1991) Proc. Natl. Acad. Sci. USA 88:2451-2455, где аминокислота №1 - это 1-й метионин, - кодируемый кодирующей последовательностью коровой области): аминокислоты 1-69 супероксиддисмутазы (SOD, используемая для усиления рекомбинантной экспрессии белка); аминокислоты 303-320 полипротеина из области Е1; аминокислоты 390-410 полипротеина, представляющие собой консенсусную последовательность гипервариабельной области из Е2 HCV-1a, аминокислоты 384-414 полипротеина из области Е2, представляющие собой консенсусную последовательность гипервариабельных областей Е2 из HCV-1 и HCV-2; аминокислоты 1211-1457 из HCV-1-полипротеина, определяющие хеликазу; три копии эпитопа из 5-1-1, аминокислоты 1689-1735, один из HCV-1, один из HCV-3 и один из HCV-2, чьи копии являются эквивалентными антигенными детерминантами из трех разных вирусных штаммов HCV; HCV-полипептид С100 из HCV-1, аминокислоты 1901-1936 полипротеина; две точные копии эпитопа из NS5-области HCV-1, каждая с аминокислотами 2278-2313 HCV-полипротеина, и две копии трех эпитопов из коровой области, двух из HCV-1 и одного из HCV-2, чьи копии являются эквивалентными антигенными детерминантами, представленными аминокислотами 9-53 и 64-88 HCV-1 и 67-84 HCV-2.
В таблице 2 показаны положения аминокислот различных эпитопов в MEFA 12 со ссылкой на фиг.7A-7F. Нумерация в таблице дана соответственно HCV-1. См. Choo, et al. (1991) Proc. Natl. Acad. Scl. USA 88:2451-2455. MEFA 13 и 13.1 разделяют общую формулу, приведенную выше для MEFA 12, с изменениями, как показано в таблицах 3 и 4, соответственно.
Согласно одному из анализов, образец соединяют с твердой подложкой, как описано ниже. Если образец инфицирован HCV, коровый антиген, также как и антитела к эпитопам, находящимся на твердой подложке, будут связываться с компонентами твердой подложки. Затем добавляют антикоровое антитело с обнаруживаемой меткой. Меченое антикоровое антитело направленно против эпитопа, отличного от того, которое связано с твердой подложкой. Это антикоровое антитело связывается с коровым антигеном, захваченным антикоровыми антителами на твердой подложке.
Также добавляют антиген, реагирующий с захваченным HCV-антителом из биологического образца, где захваченное антитело из образца может реагировать с эпитопом NS3/4a. Предпочтительно этот антиген является эпитопом, полученным из NS3-области полипротеина HCV. Этот антиген связывает захваченное HCV-антитело из образца. Число антигенов, включающих такие эпитопы, известно и включает в себя, не ограничиваясь ими, антигены, полученные из областей с33с и с100, также как и слитые белки, включающие в себя NS3-эпитоп, такой как с25. Эти и другие NS3-эпитопы можно использовать в настоящих анализах, они известны в данной области и описаны, например, у Houghton et al., патент США 5350671; Chien et al., Proc. Natl. Acad. Scl. USA (1992) 89:10011-10015; Chien et al., J. Gastroent. Hepatol. (1993) 8:S33-39; Chien et al., Международная публикация № WO 93/00365; Chien, D. Y. Международная публикация № WO 94/01778; и Патентная заявка США с одним правопреемником №08/403590 и 08/444818.
Добавляют второе меченое антитело, против описанного выше антигена. Это антитело может быть направлено против любого эпитопа, находящегося в этом антигене. Например, это антитело может быть направлено против NS3-области, находящейся в антигене; Напротив, если упомянутый антиген экспрессируется как составной белок, второе меченое антитело может быть направлено против партнера слияния. К анализу могут быть добавлены дополнительные антигены и антитела, в особенности если твердая подложка включает в себя MEFA. Подобные анализы описаны ниже.
Иллюстративный анализ согласно изобретению изображен в фигуре 2. Как показано на фигуре, твердая подложка содержит два антикóровых моноклональных антитела, обозначенных с11-3 и c11-7. Эти антитела направлены против эпитопа, находящегося в N-концевой области коревого протеина, на аминокислотах 10-53, пронумерованных соответственно последовательности полипротеина HCV1. Твердая подложка также включает в себя эпитоп к NS3/4a. Биологический образец добавляют к твердой подложке. HCV-коровый антиген, также как и антитела, направленные против эпитопа NS3/4a, вместе присутствующие в образце, будут связывать реагенты захватывания на твердой подложке.
Затем добавляли меченое пероксидазой хрена (HPR) антикóровое моноклональное антитело c11-14, направленное против С-концевой области кора, занимающее аминокислотные положения 120-130, пронумерованные соответственно последовательности полипротеина HCV1. В качестве второго HPR-меченого антитела, направленного против SOD-части слитого белка, добавляют слитый белок, включающий в себя последовательность из SOD человека (hSOD) и эпитоп из области с33с. Слияние SOD-с33с свяжется с анти-NS3-антителом и анти-SOD-антителом, в свою очередь, свяжет слитый белок SOD-с33с. Обнаружение метки свидетельствует о присутствии HCV-инфекции.
Другой иллюстративный анализ согласно изобретению изображен на фиг.8. Конфигурация анализа с использованием антител представляет собой анализ с захватом сэндвича антиген-антитело-антиген с использованием как NS3/4a, так и MEFA 12. Твердая подложка включает в себя два вышеописанных антикоровых моноклональных антитела, эпитоп к NS3/4a, также как и иллюстративные MEFA, MEFA12, которые включают в себя укороченную версию SOD человека. Как в случае вышеописанного анализа, к твердой подложке добавляют биологический образец. HCV-кбровый антиген, также как и антитела, направленные против NS3/4a-эпитопа и эпитопов MEFA, присутствующие в образце, будут связывать реагенты захватывания на твердой подложке. Добавляют два антигена, один - способный реагировать с антителами образца, которые связывают NS3/4a (как описано выше) и один - способный реагировать с антителами образца, которые связывают MEFA 12. На фигуре 8 антиген, реагирующий с комплексом MEFA 12/антитело образца, является слиянием между молекулой SOD и c22ks Δ47-L44W. Антиген c22ks происходит из коровой области и содержит аминокислоты полипротеина Lys10-Ser99, также как и присутствующую в норме делецию Аrg47 и замену Trp на Leu в положении 44. Детектируемый конъюгат антител является вторым HPR-меченым моноклональным анти-SOD-антителом, описанным выше.
Вышеописанные анализы по определению комплекса антиген/антитело особенно удачны, так как и HCV-коровый антиген, и антитела к NS3/4a и/или кору могут быть обнаружены с помощью той же подложки в том же анализе. К тому же, для того чтобы покрыть другие неструктурные эпитопы HCV, в комбинированном коктейле можно использовать, как описано выше, дополнительные HCV-эпитопы, такие как SOD слитый с с100, 5-1-1, NS5-антигены, также как и белок, полученный в результате сдвига рамки считывания в коровой области полипротеина, такой как описан в Международной публикации № WO 99/63941.
Для лучшего понимания данного изобретения ниже дается более подробное обсуждение получения антител используемых в заявленных иммунологических анализах, получения полипептидов, используемых в иммунологических анализах, и способов проведения иммунологических анализов.
Получение антител для применения в HCV-иммунологических анализах
Как показано выше, в данном анализе используются различные антитела, которые связаны с твердой подложкой (например, одно или более антикоровых антител), и таким образом обнаруживают комплексы антиген/антитело, образовавшиеся, если в образце присутствует инфекция HCV. Эти антитела могут быть поликлональными или моноклональными препаратами антител, моноспецифическими антисыворотками, антителами человека, или могут быть гибридными или химерными антителами такими, как гумманизированные антитела, измененные антитела, F(ab')2-фрагменты, F(ab)-фрагменты, Fv-фрагменты, однодоменные антитела, димерные или гримерные конструкции фрагментов антител, минитела или их функциональные фрагменты, которые связываются с рассматриваемым антигеном.
Антитела получают с использованием методик, хорошо известных специалистам в данной области и раскрытых, например, в Патентах США №4011308, 4722890, 4016043, 3876504, 3770380 и 4372745. Например, поликлональные антитела получают путем иммунизации антигеном, представляющим интерес, подходящих млекопитающих, таких как мышь, крыса, кролик, овца или коза. Для усиления иммуногенности антиген можно связать с носителем до иммунизации. Подобные носители хорошо известны специалистам в данной области. Иммунизация обычно выполняется посредством смешивания или эмульгации антигена в физиологическом растворе, предпочтительно в адъюванте, таком как полный адъювант Фрейнда, и парентерального введения смеси или эмульсии (обычно подкожно или внутримышечно). Животное обычно подвергают вторичной инъекции антигена через 2-6 недель, одной или более инъекциями антигена в физиологическом растворе, предпочтительно используя неполный адъювант Фрейнда. Антитела можно получать также с помощью иммунизации in vitro, при помощи методов, известных в данной области. Затем от иммунизированного животного получают поликлональную антисыворотку. Для описания получения анти-HCV-поликлональных антител см., например, Houghton et al., Патент США №5350671.
Моноклональные антитела обычно получают, используя метод Колера и Мильштейна (Kohler, Milstein; 1975), Nature 256: 495-497, или его модификацию. В типичном случае мышь или крысу иммунизируют, как описано выше. Однако вместо отбора крови для выделения сыворотки, лучше извлекать селезенку (или, необязательно, несколько крупных лимфатических узлов) и разделять на отдельные клетки. Если требуется, клетки селезенки можно отсортировать (после удаления неспецифически-адгезивных клеток) путем нанесения суспензии клеток на планшет или лунку, покрытую антигеном. В-клетки, экспрессирующие специфический для данного антигена связанный с мембраной иммуноглобулин, будут связываться с планшетом и не будут смываться вместе с остатками суспензии. Оставшиеся В-клетки, или все диссоциированные клетки селезенки, затем индуцируют для слияния с клетками миеломы для образования гибридом, а затем культивируют в селективной среде (например, гипоксантине, аминоптерине, тимидиновой среде, «HAT»). Получившиеся в результате гибридомы размещают на планшете при ограниченном разбавлении и проверяют на продуцирование антител, которые специфически связываются с иммунизирующим антигеном (и которые не связываются с посторонними антигенами). Затем селективные моноклональные, секретирующие антитела гибридомы культивируют in vitro (например, в бутылях для культуры тканей или реакторах с полыми волокнами) или in vivo (например, асциты мышей).
Получение различных анти-HCV-моноклональных антител описано, например, у Houghton et al. Патент США №5350671, Chien et al. Международная публикация № WO 93/00365, Патентная заявка США с одним правопреемником №08/403590 и 08/444818 и Kashiwakuma et al. Патент США №5871904.
Как показано выше, фрагменты антител, которые сохраняют способность распознавать представляющий интерес антиген, также найдут применение в заявленных иммунологических анализах. В данной области известен целый ряд фрагментов антител, которые содержат антиген-связывающие сайты, способные проявлять иммунологические связывающие свойства целой молекулы антитела. Например, можно получать функциональные фрагменты антител путем расщепления константных областей молекулы антитела, не ответственных за связывание с антигеном, с помощью, например, пепсина, чтобы получить F (ab')2-фрагменты. Эти фрагменты будут включать в себя два антиген-связывающих сайта, но в них будет отсутствовать часть константной области из каждой из тяжелых цепей. Также, если необходимо, Fab-фрагменты, включающие в себя единственный антиген-связывающий сайт, можно получать, например, путем переваривания поликлональных или моноклональных антител папаином. Функциональные фрагменты, содержащие только вариабельные области тяжелых и легких цепей, можно также получать с помощью стандартных методик, таких как получение рекомбинантов или избирательное. протеолитическое расщепление молекул иммуноглобулина. Эти фрагменты известны как Fv. Например, см. Inbar et al. (1972) Proc. Nat. Acad. Sci. USA 69:2659-2662; Hochman et al. (1976) Blochem 15:2706-2710; and Ehrlich et al. (1980) Biochem 19:4091-4096.
Одноцепочечный полипептид. Fv ("sFv", или "scFv") ковалентно связывается с VH-VL-гетеродимером, который экспрессируется слитым геном, включающим в себя VH- и VL-кодирующие гены, связанные пептид-кодирующим линкером. Huston et al., Proc. Nat. Acad. Scl. USA 85:5879-5883. Описано некоторое число методов различения и усовершенствования химических структур (линкеров), превращающих естественно агрегированные, но химически отдельные тяжелые и легкие цепи из области V-антитела, в sFv-молекулу, которая складывается в трехмерную структуру, весьма сходную со структурой антиген-сзязывающего сайта. Например, см. Патент США №5091513, 5132405 и 4946778. Молекулы sFv можно получать, используя методики, описанные в данной области. Например, см. Huston et al. (1988) Proc. Nat. Acad. Sci. USA 85:5879-5883; Патент США №5091513, 5132405 и 4946778. Критерии дизайна включают в себя определение соответствующей длины, занимающей расстояние между С-концом одной цепи и N-концом другой, где линкер обычно образован небольшим гидрофильным аминокислотным остатком, который не имеет тенденции к скручиванию или образованию вторичных структур. Эти методы описаны в данной области. Например, см. Патент США №5091513, 5132405 и 4946778. Подходящие линкеры обычно содержат полипептидные цепи из перемежающихся групп остатков глицина и серина и могут включать в себя вставки остатков глутаминовой кислоты и лизина для увеличения растворимости.
«Мини-антитела», или «мини-тела» также найдут применение в настоящем изобретении. Мини-тела являются цепями sFv-полипептида, которые включают в себя олигомеризованные домены на своем С-конце, отделенные от sFv областью петли. Pack et al. (1992) Biochem 31:1579-1584. Олигомеризация домена включает в себя самоассоциирующиеся α-спирали, например, лейциновые «молнии», которые в дальнейшем могут быть стабилизированы дополнительными дисульфидными связями. Олигомеризованный домен должен быть совместимым с векторной упаковкой поперек мембраны - процессом, призванным содействовать пространственной упаковке полипептида in vivo в функциональный связывающий белок. Обычно минитела получают, используя методы рекомбинации, хорошо известные в данной области. Например, Pack et al. (1992) Biochem 31:1579-1584; Cumber et al. (1992) J Immunology 149B:120-126.
Продуцирование антигенов для применения в иммунологических анализах HCV
Как показано выше, молекулы согласно изобретению обычно получают с помощью рекомбинации. Так, полинуклеотиды, кодирующие антигены HCV, для использования в данном изобретении можно получать, используя стандартные методики молекулярной биологии. Например, полинуклеотидную последовательность, кодирующую описанные выше молекулы, можно получить, используя методы рекомбинации, такие как скрининг циклической ДНК и библиотек генов из клеток, экспреозирующих данный ген, или извлечение гена из вектора, который заведомо его содержит. Более того, желаемый ген можно выделять непосредственно из молекул нуклеиновой кислоты вируса, используя методики, известные в данной области, такие как Houghton et al. Патент США №5350671. Ген, представляющий интерес, лучше синтезировать, чем клонировать. Можно построить молекулы с соответствующим кодоном для конкретной последовательности. Затем полную последовательность собирают из перекрывающихся олигонуклеотидов, приготовленных по стандартной методике и собранных в полную кодирующую последовательность. Например, см. Edge (1981) Nature 292:756; Nambair et al. (1984) Science 223:1299; and Jay et al. (1984) J. Biol. Chem. 259:6311.
Таким образом, конкретные нуклеотидные последовательности можно получать из векторов, содержащих требуемые последовательности, или синтезировать, полностью или частично, используя, когда целесообразно, различные способы синтеза олигонуклеотидов, известные в данной области, такие как сайт-специфический мутагенез и полимеразную цепную реакцию (ПЦР). Например, см. Sambrook, выше. В частности, существует один метод получения нуклеотидной последовательности, кодирующей требуемую последовательность при помощи отжига комплементарных наборов перекрывающихся синтетических олигонуклеотидов, полученных в стандартном автоматическом полинуклеотидном синтезаторе, с последующим лигированием соответствующей ДНК-лигазой и амплификацией лигированной нуклеотидной последовательности с помощью ПЦР. Например, см. Jayaraman et al. (1991) Proc. Natl. Acad. Sci. USA 88:4084-4088. Кроме того, в рамках изобретения можно использовать прямой олигонуклеотидный синтез (Jones et al. (1986) Nature 54:75-82), нуклеотид-направленный мутагенез предсуществующих нуклеотидных областей (Riechmann et al. (1988) Nature 332:323-327 and Verhoeyen et al. (1988) Science 239:1534-1536), и ферментативное заполнение брешей в олигонуклеотидах при помощи ДНК-полимеразы Т4 (Queen et al. (1989) Proc. Natl. Acad. Sci. USA 86:10029-10033) для получения молекул, обладающих измененными или усиленными антиген-связывающими свойствами и/или сниженной иммуногенностью.
Когда кодирующие последовательности получены или выделены, такие последовательности можно клонировать в подходящий вектор или репликон. Специалистам в данной области известны многочисленные клонирующие векторы, и подбор подходящего клонирующего вектора - это вопрос выбора. Подходящие векторы включают в себя, не ограничиваясь ими, плазмиды, фаги, транспозоны, космиды, вирусные хромосомы, способные к репликации, в ассоциации с подходящими контрольными элементами.
Затем кодирующая последовательность помещается под контроль подходящих контрольных элементов, в зависимости от системы, используемой для экспрессии. Так, кодирующая последовательность может быть помещена под контроль промотора, области связывания с рибосомами (для бактериальной экспрессии) и, необязательно, оператора, так, чтобы последовательность ДНК, представляющая интерес, транскрибировалась в РНК подходящим трансформантом. Кодирующая последовательность может содержать (а может и не содержать) сигнальные пептиды или лидерные последовательности, которые затем могут быть удалены хозяином в посттрансляционном процессинге. Например, см. Патенты США №4431739, 4425437 и 4338397.
В дополнение к управляющим последовательностям желательно добавить регуляторные последовательности, которые позволяют регулировать экспрессию последовательностей в соответствии с ростом клетки-хозяина. Регуляторные последовательности известны специалистам в данной области, и в числе их примеров - такие, которые вызывают включение или выключение экспрессии гена в ответ на физические или химические стимулы, включая присутствие регуляторного соединения. Другие типы регуляторных элементов также могут присутствовать в этом векторе. Например, в нем можно использовать энхансерные элементы для увеличения уровней экспрессии конструкций. Примеры включают в себя ранний ген энхансер SV40 (Dijkema et al., (1985) EMBO J. 4:761), энхансер/промотор, полученный из длинного концевого повтора (LTR) вируса саркомы Рауса (German et al. (1982) Proc. Natl. Acad. Sci. USA 79:6777), и элементы, полученные из CMV человека (Boshart et al. (1985) Cell 41:521), такие как элементы, входящие в А-последовательность CMV-интрона (Патент США №5688688). Кассета экспрессии может дополнительно содержать точку начала репликации для автономной репликации в подходящей клетке-хозяине, один или более выбираемых маркеров, один или более сайтов рестрикции, потенциал для большого числа копий и сильный промотор.
Экспрессирующий вектор сконструирован так, чтобы конкретная кодирующая последовательность была расположена в векторе вместе с соответствующими регуляторными последовательностями, и расположение и ориентация кодирующей последовательности относительно управляющих последовательностей было таким, чтобы кодирующая последовательность транскрибировалась под «контролем» управляющих последовательностей (т.е. РНК-полимераза, которая связывается с молекулой ДНК у контрольных последовательностей, транскрибирует кодирующую последовательность). Для достижения этого результата может потребоваться модификация последовательности, кодирующей молекулу, представляющую интерес. Например, в ряде случаев может быть необходимо модифицировать эту последовательность так, чтобы она могла прикрепляться к управляющей последовательности, имея соответствующую ориентациию, т.е. сохраняя рамку считывания. Управляющие последовательности и другие регуляторные последовательности могут быть лигированы с кодирующей последовательностью до вставки в вектор. Либо кодирующая последовательность может быть клонирована непосредственно в вектор экспрессии, который уже содержит управляющие последовательности и соответствующий сайт рестрикции.
Как показано выше, может быть также желательным получение мутантов или аналогов антигена, представляющего интерес. Это особенно справедливо в случае NS3/4a. Методы осуществления этого описаны, например, в Dasmahapatra et al. Патент США №5843752, и Zhang et al., Патент США №5990276. Мутанты или аналоги этого или другого белков HCV для применения в заявленных анализах могут быть получены путем делеции части последовательности, кодирующей интересующий полипептид, вставки последовательности и/или замены одного или более нуклеотидов в данной последовательности. Методики модификации нуклеотидных последовательностей, такие как сайт-направленный мутагенез, и т.п., хорошо известны специалистам в данной области. Например, см. Sambrook et al., supra; Kunkel, T. A. (1985) Proc. Natl. Acad. Sci. USA (1985) 82:448; Geisselsoder et al. (1987) BioTechnlques 5:786; Zoller and Smith (1983) Methods Enzyiaol. 100:468; Dalbie-McFarland et al. (1982) Proc. Natl. Acad. Sci USA 79:6409.
Молекулы могут быть экспрессированы в самых разнообразных системах, включая системы экспрессии насекомых, млекопитающих, бактерий, вирусов и дрожжей, также известные в данной области.
Например, системы экспрессии клетки насекомого, такие как бакуловирусные системы, известны специалистам в данной области и описаны, например, Summers and Smith, Texas Agricultural Experiment Station Bulletin No. 1555 (1987). Материалы и методы в виде набора для экспрессионной системы бакуловирус/клетка насекомого доступны для приобретения, кроме прочего, в Invitrogen, Сан-Диего, Калифорния (набор «МахВас»). Также системы экспрессии на основе бактериальных клеток и клеток млекопитающего хорошо известны в данной области и описаны, например, в Sambrook et al., выше. Дрожжевые системы экспрессии также известны в данной области и описаны, например, в Yeast Genetic Engineering (Barr et al., eds., 1989) Butterworths, London.
Число подходящих клеток-хозяев для применения в упомянутых системах также известно. Например, клеточные линии млекопитающих известны в данной области и включают в себя иммортализованные клеточные линии, доступные в Американской Коллекции Типовых Культур (АТСС), включая, но не ограничиваясь ими; клетки яичников Китайского хомяка (СНО), клетки HeLa, клетки почки детеныша хомяка (ВНК), клетки почки обезьяны (COS), клетки почки человеческого эмбриона, клетки гепатоцеллюлярной карциномы человека (например, Нер G2), клетки Медин-Дерби (Madin-Darby) бычьей почки ("MDBK"), а также и другие. Также бактериальные хозяева, такие как Е.Coli, Bacillis subtilis и Streptococcus spp., найдут применение в настоящих экспрессирующих конструкциях. Хозяева-дрожжи, которые можно использовать в настоящем изобретении, включают в себя, кроме прочих, Saccharomyces cerevisiae, Candida albicans, Candida maltosa, Hansenula polymorpha, Kluyveromyces fragilis, Kluyveromyces. lactis, Pichia guillerimondii, Pichia pastoris, Schizosaccharomyces pombe и Yarroma lipolytica. Клетки насекомых, используемые в бакуловирусных экспрессирующих векторах, включают в себя, кроме прочего, Aedes aegypti, Autographs californica, Bombyx mori, Drosophila melanoqaster, Spodoptera frugiperda, и Trichoplusiani.
Молекулы нуклеиновой кислоты, включающие в себя нуклеотидные последовательности, представляющие интерес, могут быть устойчиво интегрированы в геном клетки-хозяина или сохраняться в стабильном эписомном элементе в подходящей клетке-хозяине, с помощью разнообразных методик доставки гена, хорошо известных в данной области. Например, см. Патент США №5399346.
В зависимости от системы экспрессии и выбранного хозяина, при росте клеток, трансформированных описанным выше экспрессирующим вектором, получают молекулы в условиях, при которых экспрессируется белок. Затем экспрессированный белок выделяют из клеток-хозяев и очищают. Если системы экспрессии секретирует белок в среду для выращивания, продукт очищают прямо из этой среды. Если секреции не происходит, продукт можно выделить из клеточных лизатов. Подбор подходящих условий выращивания и способов восстановления находится в пределах квалификации специалистов в данной области.
Описано получение разнообразных HCV-антигенов при помощи рекомбинации. Например, см. Houghton et al., Патент США №5350671, Chien et al. J. Gastroent. Hepatol. (1993). 8:S33-39; Chien et al. Международная публикация № WO 93/00365, Chien, D.Y., Международная публикация № WO 94/01778.
Иммунодиагностические анализы
Сразу после получения упомянутые антикоровые антитела и NS3/4a-антигены помещают на соответствующую твердую подложку для использования в заявленных иммуноанализах. Твердая подложка для целей согласно изобретению может быть любым материалом, который может являться нерастворимой основой с жесткой или полужесткой поверхностью. Типичная твердая подложка включает в себя, не ограничиваясь ими, такие субстраты как нитроцеллюлоза (например, в форме мембраны или лунки для микротитрования), полихлорвинил (например, в форме листов или лунок для микротитрования), полистирольный латекс (например, в форме шариков или планшет для микротитрования), поливинилидинфторид, диазотированная бумага, нейлоновые мембраны, активированные шарики, шарики для магнитных мешалок и т.п. Частные случаи подложек включают в себя планшеты, гранулы, диски, капилляры, полые волокна, булавки, иглы, сплошные волокна, целлюлозные шарики, пористые стеклянные шарики, силикагели, полистироловые шарики, необязательно поперечносшитые с дивинилбензолом, шарики из привитого сополимера, полиакриламидные шарики, латексные шарики, диметилакриламидные шарики, необязательно поперечносшитые с N-N'-бис-акрилоилэтилендиамином, и стеклянные частицы, покрытые гидрофобным полимером.
При желании, молекулам, добавленным к твердой подложке, может быть легко придана функция образования стироловых или акрилатных фрагментов, таким образом создается возможность встраивания молекул в полистирол, полиакрилат и другие полимеры, такие как полиимид, полиакриламид, полиэтилен, поливинил, полидиацетилен, полифенилен-винилен, полипептид, полисахарид, полисульфон, полипиррол, полиимидазол, политиофен, полиэфир, эпоксидные смолы, кварцевое стекло, силикагель, силоксан, полифосфат, гидрогель, агароза, целлюлоза и т.п.
В этом контексте, твердая подложка вначале реагирует с HCV-антикоровыми антителами и NS3/4a-эпитопом (вместе называемые здесь «твердофазными компонентами») и, необязательно, с одним или более MEFA, при подходящих условиях сшивки, при которых молекулы в достаточной степени иммобилизированы на подложке. Иногда иммобилизация на подложке может быть усилена первой связью антигена и/или антитела с белком, обладающим лучшими свойствами твердофазного связывания. Подходящие связывающие белки включают в себя, не ограничивася ими, такие макромолекулы как сывороточные альбумины, включая бычий сывороточный альбумин (BSA), гемоцианин морского блюдечка, молекулы иммуноглобулина, тироглобулин, яичный альбумин и другие белки, хорошо известные специалистам в данной области. Другие реагенты, которые можно использовать для связывания молекул с подложкой, включают в себя полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, аминокислотные сополимеры и т.п. Такие молекулы и методы связывания этих молекул с антигенами хорошо известны специалистам в данной области. Например, см. Brinkley, М.А. (1992) Blo conjugate Chem. 3:2-13; Hashida et al. (1984) J. Appi. Biochem. 6:56-63; и Anjaneyulu and Staros (1987) International J. of Peptide and. Protein Res. 30:117-124.
После взаимодействия твердой подложки с твердофазными компонентами все неиммобилизированные твердофазные компоненты удаляют с подложки промыванием, и затем компоненты, связанные с подложкой, вступают в контакт с биологическим образцом, подозреваемым в наличии антител или антигенов HCV (вместе называемых здесь «лигандными молекулами») в подходящих для связывания условиях. После промывки для удаления всех несвязанных лигандных молекул в подходящих для связывания условиях добавляют второе антикоровое антитело, направленное против эпитопа, отличного от антикорового антитела, связанного с подложкой. Добавленное антикоровое антитело включает в себя обнаруживаемую метку, как описано выше, и связывает любой антикоровый антиген, который может присутствовать в образце, который прореагировал с антикоровым антителом, связанным с подложкой. Также добавляют один или более антигенов, которые могут реагировать с присутствующими в образце антителами, которые, в свою очередь, прореагировали с NS3/4a-эпитопом. Как объяснялось выше, этот антиген обычно получают из NS3-области HCV-полипротеина и, в частности, из с33с-области HCV. Данная область и полученные из нее эпитопы описаны у Houghton et al. Для описания этой области и полученных из нее эпитопов см. Патент США №5350671; Chien et al., Proc. Nat. Acad. Sci. (1989) 89:10011-10015; Международная публикация № WO 93/00365 и Патентная заявка США с одним правопреемником №08/403590 и 08/444818. Также добавляют меченое антитело, направленное против этого антигена. Антитело затем связывается с антигеном, который прореагировал с анти-NS3-антителами, находящимися а образце. Эпитоп с33с можно легко получить для этой цели как слияние между с33с и супероксиддисмутазой человека (hSOD), полученное с помощью рекомбинации, например, по методике, описанной у Hough-ton et al.. Патент США №5350671. Нуклеотидные и аминокислотные последовательности SOD человека известны и опубликованы в Haliewell et al., Патент США №5710033. Поэтому меченое антитело, направленное против SOD человека, может быть использовано для обнаружения присутствия комплексов, образованных между NS3/4а-эпитопом, любыми антителами в образце, которые реагируют с этим эпитопом, и HCV-полипептидами, которые, в свою очередь, связываются с этим антителом в образце.
Если MEFA присутствует на твердой подложке, в анализ можно также добавить один или более дополнительных антигенов, которые реагируют с антителами из биологического образца, связанными с антигенами, представленными в MEFA. Особенно полезен в этом контексте антиген, полученный из коровой области HCV, и, в частности, из антигена с22, который включает в себя 119 N-концевых кóровых аминокислот полипротеина HCV. Таким антигеном, полученным из с22, является c22ks Δ47-L44W, который включает в себя аминокислоты полипротеина Lys10-Ser99, также как и в норме присутствующую делецию Аrg47 и замену Trp на Leu в положении 44. Как и с описанном выше эпитопом с33с, этот антиген можно получить как слияние с hSOD, и то же самое меченое антитело, направленное против SOD человека, можно использовать для обнаружения наличия комплексов, образованных между антителами, присутствующими в образце, и NS3/4а-эпитопом и/или MEFA, чьи комплексы также связаны с HCV-антигенами (например, с33с и с22).
В частности, можно использовать метод ELISA, в котором лунки планшета для микротитрования покрыты твердофазными компонентами. Затем в покрытые лунки добавляют биологический образец, который содержит или предположительно содержит лигандные молекулы. После периода инкубации, достаточного для того, чтобы лигандные молекулы связались с иммобилизированными твердофазными компонентами, планшет (планшеты) можно промыть для удаления несвязавшихся фрагментов и добавить вторично связывющую молекулу с обнаруживаемой меткой (меченое антикоровое антитело), NS3-эпитоп-содержащую молекулу и антитело, направленное против NS3-эпитопсодержащей молекулы. Этим молекулам дается время прореагировать с любым захваченным антигеном и антителом из образца, планшет промывают, и определяют присутствие меченых антител с помощью методики, хорошо известной в данной области.
Реагенты для - анализа, описанные выше, включая твердую подложку для иммунологического анализа с антителами и антигенами, также как и антитела и антигены для реакции с захваченным образцом, могут поставляться в виде наборов, с соответствующей инструкцией и другими необходимыми реагентами, для проведения иммуноанализа, как было описано выше. В зависимости от конкретного иммуноанализа набор может содержать соответствующие метки и другие упакованные реагенты и материалы (например, промывочный буфер и т.п.). С помощью этих наборов можно проводить описанные выше стандартные иммунологические анализы.
III. Экспериментальная часть
Ниже даны примеры специфических вариантов осуществления настоящего изобретения. Данные примеры предлагаются только для иллюстративных целей и не преследуют задачу ограничить рамки настоящего изобретения каким-либо образом.
Были предприняты определенные усилия, чтобы гарантировать точность в отношении использованных цифр (например, количества, температуры и т.п.), но необходимо учесть возможность некоторых экспериментальных ошибок и отклонений.
ПРИМЕР 1
Иммунологический анализ образования комплекса HCV-антиген/антитело.
Настоящий иммунологический анализ образования комплекса HCV-антиген/антитело сравнивали с другими HCV-анализами, чтобы протестировать пределы выявления сероконверсии и сравнить эти пределы с таковыми, полученными в других доступных для приобретения анализах, как следует ниже.
А. Материалы и методы
Образцы крови: Использовали выборки образцов доступной для приобретения человеческой крови. Эти выборки образцов доступны, например, в Boston Biomedica, Inc., West Bridgewater, MA (BBI); Bioclinical Partners, Franklin, MA (BCP); и North American Biologies, Inc., BocoRatan, FL (NABI). Дни, показанные в таблицах 5 и 6, означают дни, когда кровь отбирали у доноров.
Моноклональные антитела: Моноклональные антитела с11-3, с11-7 и c11-14 были получены в Ortho Clinical Diagnostics, Raritan, New Jersey. Антитела с11-3 и с11-7 направлены против N-концевой части кора (аминокислоты 10-53, пронумерованные соответственно HCV1-полипротеина). Моноклональное антитело c11-14 направлено против С-концевой части кора (аминокислоты 120-130, пронумерованные соответственно HCV1-полипротеина). Антитело c11-14 конъюгировали с пероксидазой хрена (HPR) с использованием стандартных методов.
Моноклональное антитело 5А-3 и анти-SOD-антитело, направленное против аминокислот 1-65 SOD, получали с помощью стандартных методик. Антитело конъюгировали с (HPR), как описано выше.
В. Антигены:
Антиген с33с (266 аминокислот, аминокислоты 1192-1457 HCV1-полипротеина) экспрессировали как внутренний SOD-слитый полипептид в Е.coli, с помощью методики, описанной для синтеза 5-1-1-антигена. (Choo, et al. Science (1989) 244:359-362). Рекомбинантный антиген был очищен и описан Chien, et al., Proc. Natl. Acad. Sci. (1989) 89:10011-10015. Также см. протокол получения SOD-с33с у Houghton et al. Патент США №5350671.
Эпитоп NS3/4a, использованный в анализе, является конформационным эпитопом, обладающим последовательностью, показанной на фиг.3.
С. Форматы иммунологических анализов:
Анализ Abbot PRISM (Abbott Laboratories, Abbott Park, IL), является доступным для приобретения анализом на основе антител. Анализ проводили, используя инструкции производителя.
Тестовая система ELISA версия 3.0 ORTHO HCV (обозначенная здесь Ortho 3.0 анализ, Ortho Clinical Diagnostics, Raritan, New Jersey) является доступным для приобретения анализом на основе PCR. Анализ проводили, используя инструкции производителя.
Анализ Roche Amplicor (Roche, Pleasant, CA) является доступным для приобретения PCR-тестом. Анализ проводили, используя инструкции производителя.
Анализ Gen-Probe TMA (San Diego, Ca) является доступным для приобретения транскрипционно-опосредованным амплификационным анализом. Анализ проводили, используя инструкции производителя.
Антигеный анализ Ortho (Ortho Clinical Diagnostics, Raritan, New Jersey) является анализом на основе антител. Анализ проводили, используя инструкции производителя.
Заявленный иммуноанализ образования комплекса HCV-антиген/антитело проводили следующим образом. Очищенные моноклональные антитела (4 мг/мл каждое) СП-7 и СП-3 в 1х фосфантно-соляном растворе (PBS), рН 7,4, объединяли, хорошо перемешивая. В тот же покрывающий буфер добавляли 90 нг рекомбинантного антигена NS3/4a. Перед покрытием раствор перемешивали в течение 30 минут. Добавляли указанный раствор в лунки 96-луночного микротитровального планшета для связывающей среды Costar (Corning, Inc.), 200 мкл на лунку. Планшеты инкубировали при 15-30°С в течение 16-24 часов. Планшеты дважды промывали дистиллированной H2O, с последующим добавлением 300 мкл/лунку послепокровного буфера (1% бычий сывороточный альбумин (BSA), 1х PBS) на один час и 300 мкл/лунку стабилизирующего буфера (1х PBS, 1% BSA, маннит, полиэтилен гликоль (PEG), желатин), на 1 час. Жидкость с планшетов удаляли, и сушили их при 4°С в течение 24 часов в лиофилизаторе. Планшеты помещали в пакеты с поглотителем влаги.
Для проведения иммунологического анализа в планшет добавляли 100 мкл усиливающего лизис буфера (1% N-лаурилсаркозин, 0,65 М NaCl, 50 мг/мл мышиного IgG, технического качества (Sigma, St. Louis, МО), 1% модифицированного сульфогидрилом BSA (Bayer), 0,1% казеин). Затем добавляли 100 мкл образца. Все это инкубировали на шейкере при 40°С в течение часа. Планшеты промывали шесть раз с 1х PBS и 0,1% Tween-20 на мойке Ortho Plate Washer. 200 мкл раствора коньюгата (c11-14-HPR в разведении 1:75 с SOD-сЗЗс 250 нг/анализ, плюс анти-SOD-HPR мыши, разведенный 1:5000 в разбавителе для образца HCV 3.0 (ORTHO HCV версия 3.0 тестовая система ELISA, Ortho Clinical Diagnostics, Raritan, New Jersey) без экстракта SOD, все приготовляется за 30 минут до добавления). Раствор инкубировали при встряхивании в течение 45 минут при 40°С. Затем промывали шесть раз, как показано выше, и добавляли 200 мкл раствора субстрата (1 таблетка ODP/10 мл). Таблетка ODP, доступная в Sigma, St.Louis, МО, содержит о-фенилендиаминдигидрохлорид и перекись водорода для окрашивания реакции пероксидазы хрена. Все это инкубировали 30 минут при 15-30°С в темноте. Реакцию останавливали добавлением 50 мл 4н H2SO4, и планшеты считывали при 492 нм, относительно поглощения при 690 нм в качестве контроля.
D. Результаты:
Результаты различных анализов показаны в таблицах 5 и 6, где изображены два отдельных эксперимента с образцами крови, инфицированными HCV, как показано. Серым показано обнаружение вируса. Как показано ниже, анализ комбинации антиген/антитело Чайрона (Chiron) выявлял сероконверсию во всех образцах, тогда как все другие анализы на основе антигенов и антител не смогли выявить сероконверсию ни в одном из образцов. В частности, ни один из анализов на основе антитела не выявил сероконверсию по меньшей мере до 18-го дня (таблица 5). Таблица 6 показывает, что ни один из анализов на основе антитела не выявил присутствие инфекции HCV на 22 день. Более того, после 85-ти дней анализ на основе Ortho-антигена не смог выявить сероконверсии.
Таким образом, на основе вышеуказанных результатов становится ясно, что новый анализ комбинации антиген/антитело уменьшает число ложных негативных результатов, полученных при использовании других стандартных анализов на основе антигенов и антител.
Сероконверсия HCV
Сероконверсия HCV
ПРИМЕР 2
Получение конформационного эпитопа NS3/4a с заменами Thr на Pro и Ser на Ile
Конформационный эпитоп NS3/4a получали следующим образом. Этот эпитоп обладает последовательностью, показанной в фигурах 4A-4D и отличается от природной последовательности в положениях 403 (аминокислота 1428 полноразмерной последовательности HCV-1) и 404 (аминокислота 1429 полноразмерной последовательности HCV-1). Более точно, Thr в норме, находящийся в положении 1428 природной последовательности, мутирован на Pro, a Ser, встречающийся в положении 1429, мутирован на Ile.
В частности, использовали описанный выше дрожжевой экспрессирующий вектор рВ324.1. Плазмиду pd.hcvla.ns3ns4aPL, которая кодировала иллюстративный эпитоп NS3/4a, использованный в заявляемых иммуноанализах, получали следующим образом. Использовали двухстадийную методику. Сначала лигировали следующие участки ДНК: (а) синтетические олигонуклеотиды, которые образуют 5' HindIII клонирующий сайт, за которым следует последовательность АСААААСААА, инициатор ATG и кодоны для HCVIa, начинающиеся с аминокислоты 1027 и продолжающиеся до сайта BglI на аминокислотах 1046; (b) фрагмент рестрикции 683 bp BglI-ClaI (кодирующий аминокислоты 1046-1274) из pAcHLTns3ns4aPI; и (с) вектор pSP72 (Promega, Madison, WI, GenBank/EMBL Accession Number X65332), который был переварен HindIII и ClaI, дефосфорилирован и очищен гель-фильтрацией. Плазмида pAcHLTns3ns4aPI была получена из pAcHLT, экспрессирующий вектор бакуловируса, доступный для приобретения в BD Pharmigen (Сан-Диего, Калифорния). В частности, был получен вектор pAcHLT EcoRI-PstI, также, как и следующие фрагменты: EcoRI-AlwnI, 935 пн, соответствующий аминокислотам 1027-1336 генома HCV-1; AlwnI-SacII, 247 пн, соответствующий аминокислотам 1336-1419 генома HCV-1; HinfI-BglI, 175 пн, соответствующий аминокислотам 1449-1509 генома HCV-1; BglI-PstI, 619 пн, соответствующий аминокислотам 1510-1711 генома HCV-1; плюс кодон терминации транскрипции. Синтезированный 91 пн фрагмент SacII-HinfI, соответствующий аминокислотам 1420-1448 генома HCV-1 и содержащий PI мутации (Thr-1428 заменен на Pro, Ser-1429 заменен на Ile), лигировали с 175 пн фрагментом HinfI-BglI, и 619 пн фрагментом BglI-PstI, описанными выше, и субклонировали в вектор pGEM-5Zf(+), переваренный SacII и PstI. pGEM-5Zf(+) является доступным для приобретения вектором Е.coli (Promega, Madison, WI, GenBank/EMBL Accession Number X65308). После трансформации компетентными клетками НВ101, минискринингового анализа индивидуальных клонов и верификации последовательности, фрагмент 885 пн Sac II-Pstl из pGEM5. PI клон 2 очищали путем гель-фильтрации. Этот фрагмент лигировали с фрагментом EcoRI-AlwnI 935 пн, фрагментом AlwnI-SacII 247 пн и вектором pAcHLT EcoRI-PstI, описанным выше. Получившаяся в результате конструкция получила название pAcHLT Tns3ns4aPI.
Вышеупомянутая лигационная смесь была трансформирована в HBlOl-компетентных клетках и помещена на агаровые планшеты Luria, содержащие 100 мкг/мл ампициллина. Miniprep анализы индивидуальных клонов приводили к определению мнимых позитивов, два из которых были амплифицированы. Плазмидную ДНК pSP72 1aHC клонов, #1 и #2, получали с помощью набора Qiagen Maxiprep и секвенировали.
Затем, следующие фрагменты были лигированы друг с другом: (а) 761 пн фрагмент HindIII-CalI из р8Р721аНС #1 (pSP72.laHC получали путем лигирования друг с другом: pSP72, который переварили HindIII и ClaI, синтетических олигонуклеотидов, которые образуют сайт клонирования 5' HindIII, за которым следует последовательность АСААААСААА, кодон инициации ATG и кодоны HCVIa, начинающиеся с аминокислоты 1027 и продолжающиеся до аминокислоты 1046 сайта BglII, и фрагмента рестрикции BglII-ClaI в 683 пн (кодирующего аминокислоты 1046-1274) из pAcHLTns3ns4aPI); (b) фрагмент BamHI-HindIII в 1353 пн гибридного промотора дрожжей ADH2/GAPDH; (с) фрагмент ClaI-SalI в 1320 пн (кодирующий аминокислоты HCVIa 1046-1711 с Thr 1428, замененным на Pro, и Ser 1429, замененным на Не) из pAcHLTns3ns4aPI; и (d) экспрессирующий вектор дрожжей pBS24.1, который был переварен BamHI и SalI, дефосфорилирован и очищен с помощью гель-фильтрации. Лигационная смесь была трансформирована в HB101-компетентных клетках и помещена на агаровые планшеты Luria, содержащие 100 мкг/мл ампициллина. Miniprep-анализы индивидуальных клонов приводили к определению отдельных клонов с ожидаемой вставкой BamHI-SalI в 3446 пн, которая состояла из промотора ADH2/GAPDH, инициирующего кодона ATG и NS3/4a из HCVIa, состоящего из аминокислот 1027-1711 (показанных как аминокислоты 1-686 на фиг.4A-4D) с Thr 1248 (аминокислотное положение 403 на фиг.4A-4D), замененным на Pro, и Ser (аминокислотное положение 404 на фиг.4A-4D), замененным на Не. Конструкция названа pd.HCVla.ns3ns4aPI (см. фиг.5).
Штамм S.cerevisiae AD3 трансформировали pd.HCVla.ns3ns4aPI и после истощения глюкозы в среде, проверяли на предмет экспрессии у отдельных трансформантов. В дрожжах был отмечен высокий уровень экспрессии рекомбинантного белка, как было показано с помощью красителя Кумасси голубого и подтверждено с помощью анализа иммуноблоттинга с использованием поликлонального антитела к хеликазному домену N33.
ПРИМЕР 3
Очистка конформационного эпитопа NS3/4a
Конформационный эпитоп NS3/4a очищали следующим образом. Упомянутые клетки S. cerevisae, экспрессирующие эпитоп NS3/4a, собирали, как было описано выше. Клетки суспендировали в лизирующем буфере (50 мМ Tris pH 8,0; 150 мМ NaCl; 1 мМ EDTA; 1 мМ PMSF; 0,1 мкМ пепстатин; 1 мкМ леупептин) и лизировали в Dyno-Mill (Wab Willy A. Bachofon, Базель, Швейцария) или аналогичном аппарате с использованием стеклянных шариков в соотношении 1:1:1 клетки:буффер:0,5 мм стеклянные шарики. Лизат центрифугировали при 30100 × g в течение 30 минут при 4°С и осадок, содержащий фракцию нерастворимого белка, добавляли к промывочному буферу (6 мл на грамм начального веса клеточного комочка) и перемешивали путем взбалтывания при комнатной температуре в течение 15 минут. Промывочный буфер состоял из 50нМ NaPO4 pH 8,0; 0,3 М NaCl; 5 мМ β-меркаптоэтанола, 10% глицерина, 0,05% октилглюкозида, 1 мМ EDTA; 1 мМ PMSF, 0,1 мкМ пепстатина; 1 мкМ леупептина. Клеточные остатки удаляли путем центрифугирования при 30100 × g в течениие 30 минут при 4°С. Супернатант выливали и оставляли осадок.
Белок из осадка получали следующим образом. Добавляли 6 мл/г экстрагирующего буфера и перемешивали путем взбалтывания при комнатной температуре в течение 15 минут. Экстрагирующий буфер состоял из 50 нМ Tris рН 8,0; 1 М Nad; 5 мМ β-меркаптоэтанола, 10% глицерина, 1 мМ EDTA; 1 мМ PMSF, 0,1 мкМ пепстатина; 1 мкМ лейпептина. Все это центрифугировали при 30100 × g в течение 30 минут при 4°С. Сохраняли супернатант и добавляли сульфат аммония до 17,5%, используя следующую формулу: объем супернатанта (мл) умноженный на х% сульфата аммония/(1 - х% сульфата аммония) = мл 4,1 М насыщенного сульфата аммония, который следует добавить в супернатант. Сульфат аммония добавляли по каплям при перемешивании на льду, и раствор перемешивали на льду в течение 10 минут. Раствор центрифугировали при 17700 × g в течение 30 минут при 4°С и осадок оставляли и сохраняли при 2-8°С вплоть до 48 часов.
Осадок ресуспендировали и пропускали через колонку Poly U (Poly U-Сефароза 4В, Amstersham Pharmacia) при 4°С следующим образом. Осадок ресуспендировали в Poly U-уравновешивающем буфере - 6 мл на грамм веса осадка. Уравновешивающий буфер состоял из 25 mm HEPES рН 8,0, 200 мМ NaCl, 5 мМ DTT (добавляют свежим), 10% глицерина, 1,2 октилглюкозида. Раствор перемешивали путем взбалтывания при 4°С в течение 15 минут и центрифугировали при 30100 × g в течениие 30 минут при 4°С.
Подготавливали Poly U-колонку (1 мл смолы на грамм начального веса осадка). Линейная скорость течения составляла 60 см/час, а скорость при набивке была 133% от 60 см/час. Колонка была уравновешена уравновешивающим буфером, и супернатант ресуспендированного аммонийно-сульфатного осадка загружали в уравновешенную колонку. Колонку промывали до исходного состояния уравновешивающим буфером и белок элюировали постадийной элюцией в следующем Poly U-элюирующем буфере: 25 мМ HEPES рН 0,8; 1М NaCl, 5 мМ DTT (добавляли свежим), 10% глицерин, 1,2 октилглюкозид. Элюат с колонки пропускали через SDS-PAGE (окрашенный Кумасси), и аликвоты замораживали и сохраняли при -80°С. Присутствие эпитопа NS3/4a подтверждали с помощью Вестерн-блот-анализа, с использованием поликлонального антитела, направленного против NS3-протеазного домена, и моноклонального антитела против 5-1-1-эпитопа (HCV 4а).
Дополнительно, в процессе очистки отслеживали протеазную ферментативную активность следующим образом. Пептид NS4A (KKGSVVIVGRIVLSGKPAIIPKK) и образец, содержащий конформационный эпитоп NS3/4a, растворяли в 90 мкл реакционного буфера (25 мМ Tris, рН 7,5; 0,15 мМ NaCl, 0,5 мМ EDTA, 10% глицерин, 0,05 н-додецил-В-D-мальтозид, 5мМ DTT) и позволили перемешиваться в течение 30 минут при комнатной температуре. В планшет для микротитрования (Costar, Inc., Corning, NY) добавляли 90 мкл смеси и 10 мкл HCV субстрата (AnaSpec, Inc., Сан-Хосе, Калифорния). Содержимое планшета перемешивали и считывали на ридер-планшетах Fluostar. Результаты выражали в единицах относительной флюоресценции (RFU) в минуту.
При использовании этих методов, продукт экстракции 1М NaCl имел активность 3,7 RFU/мин, преципитат сульфата аммония имел активность 7,5 RFU/мин, продукт Poly U очистки имел активность 18,5 RFU/мин.
Пример 4
Конкурентные исследования.
Были проведены следующие конкурентные исследования оценки того, может ли конформационный эпитоп NS3/4a распознавать другие антитела, по сравнению с иными HCV-антигенами. В частности, антиген NS3/4a сравнивали с антигеном с200 следующим образом.
0,5 мкг и 0,1 мкг NS3/4a, полученных, как описано выше, или с200 (Hepatology (1992) 15; 19-25, доступно в ORTHO HCV Version 3.0 ELISA Test System, Ortho Clinical Diagnostics, Raritan, New Jarsey) смешивали с 20 мкл образца PHV914-5 (образец ранней сероконверсии, полученный из крови инфицированного субъекта) в полном объеме 220 мкл (1 × PBS). Смесь инкубировали в течение часа в микролунке при 37°С. Планшеты промывали и проводили анализ следующим образом.
1 мкг антигена с200 добавляли к 10 мкг образца PHV914-5 в полном объеме 220 мкл. Смесь инкубировали в течение 1 часа в микролунке при 37°С, и 200 мкл переносили на планшет, покрытый NS3/4a (100 нг/анализ), и инкубировали в течение 1 часа при 37°С. Планшеты промывали пять раз с помощью 1 × PBS, 0,1 Tween-20. Добавляли 200 мкл раствора конъюгата (описан выше), планшеты инкубировали, и проводили анализ. Контрольные повторности, которые состояли из PHV914-5 и 1 × PBS (без антигена), обрабатывали таким же образом.
Результаты показаны в таблице 7. Результаты процентного ингибирования, показанные в колонке 4, вычисляли следующим образом: колонка 3 минус (колонка 2, деленная на колонку 3 и умноженная на 100). Как можно видеть, данные показывают, что NS34a нейтрализуются антителами ранней сероконверсии, а с200 - нет. Достигался сильный сигнал, когда антитела в PHV914-5 с33с из выборки образцов ранней сероконверсии реагировали с NS34a, нанесенным на микропланшет. Антиген с200 не был нейтрализован этими антителами. Это показано в верхней части таблицы 7. Когда NS34a смешивали с образцом PHV914-5, он был нейтрализован, и поэтому в образце не оказалось антител, реагирующих с NS34a, нанесенным на микропланшет. Данные показывают, что NS34a мог распознать класс антител, отличных от тех, которые распознает с200.
Конкурентные исследования, показывающие, что NS34a антиген распознает другие антитела в выборке образцов ранней с33с сероконверсии. по сравнению с с200 антигеном
ПРИМЕР 5
Исследования стабильности конформационного эпитопа NS34a.
Для оценки роли стабильности NS3/4а-эпитопа в выполнении анализа было проведено следующее исследование, определяющее зависимость иммунореактивности NS3/4a от времени при комнатой температуре. Небольшие аликвоты имеющегося в наличии NS3/4a хранили при комнатной температуре и замораживали через интервалы, показанные в таблице 8. Все пузырьки были покрыты одновременно и протестированы на двух выборках образцов ранней сероконверсии NS3.
Как видно из таблицы 8, запас NS3/4a нестабилен и иммунореактивность уменьшается со временем. К тому же для сохранения иммунореактивности необходимо поддержание конформации NS3/4a.
Дальнейшие исследования стабильности были поставлены следующим образом. Два конформационных моноклональных антитела против NS3/4a, созданных с помощью стандартных процедур, заменили на выборку образцов ранней анти-HCV сероконверсии. Пузырьки NS3/4a из основного запаса хранили при комнатной температуре, увеличивая время хранения с интервалами 3, 6 и 24 часа. NS3/4a из замороженных пузырьков (90 нг/мл) наносили, и проводили анализ, используя описанную выше методику. Результаты свидетельствуют о том, что оба моноклональных антитела действительно были конформационными и их реакционная способность чувствительна к манипулирванию NS3/4a-антигеном из основного запаса при комнатной температуре. Реакционная способность моноклонального антитела в положительном контроле не изменялась.
ПРИМЕР 6
Иммунореактивность конформационного эпитопа NS3/4a в сравнении с денатурированным NS3/4a.
Иммунореактивность конформационного эпитопа NS3/4a, полученного как описано выше, сравнивали с NS3/4a, который был денатурирован путем добавления SDS к препарату конформационного эпитопа NS3/4a до итоговой концентрации 2%. Денатурированный NS3/4a и конформационный NS3/4a наносили на планшеты для микротитрования, как было описано выше. Антиген с200 (Hepatology (1992) 15:19-25, доступный в ORTHO HCV Version 3.0 ELISA Test System, Ortho Clinical Diagnostics, Raritan, New Jarsey), также наносили на планшеты для микротитрования. Антиген с200, который использовали для сравнения, считается неконформационным из-за присутствующего в его рецептуре редуцирующего агента (DTT) и детергента (SDS).
Иммунореактивность тестировали по отношению к двум выборкам образцов ранней HCV сероконверсии PHV 904 и PHV 914 (доступные для приобретения образцы крови человека из Boston Biomedica, Inc., West Bridgewater, MA). Результаты показаны в таблице 9. Данные показывают, что денатурированная или линеаризованная форма NS3/4a (так же, как и с200) не распознает выборки образцов ранней сероконверсии так же рано, как NS3/4а-конформационный эпитоп.
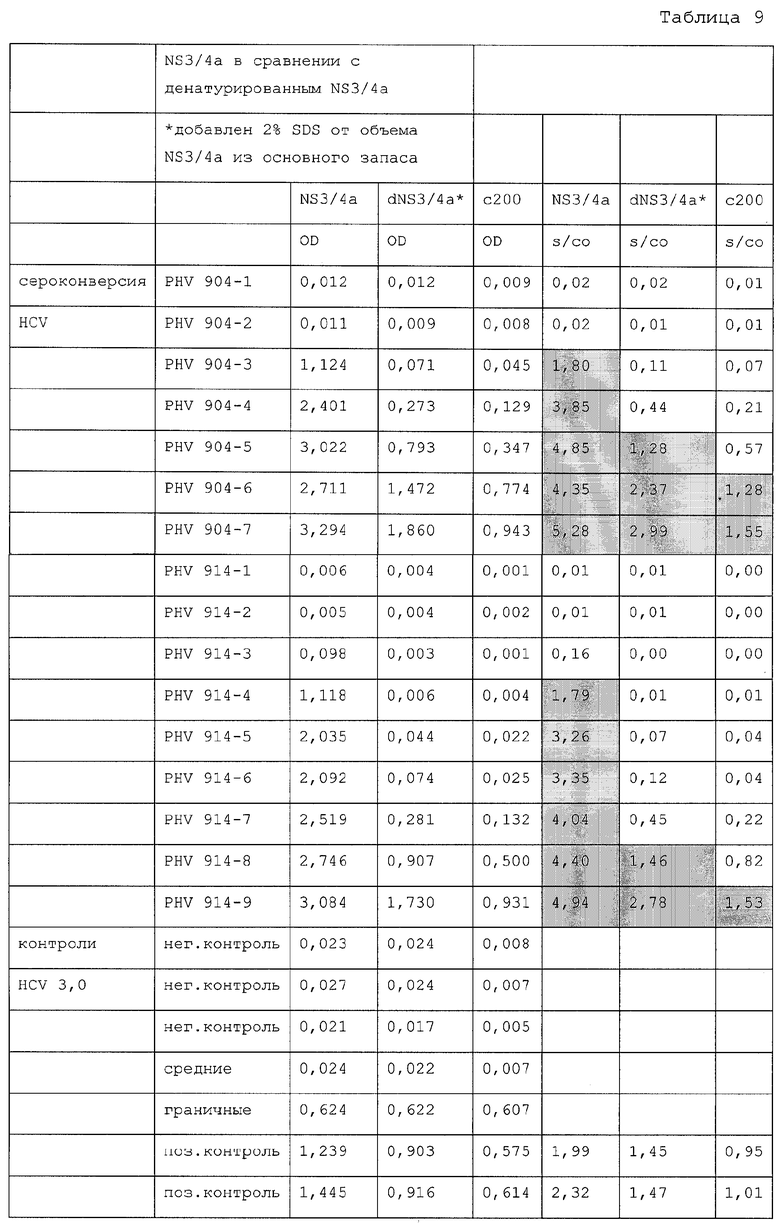
Иммунореактивность конформационного эпитопа также тестировали с использованием моноклональных антител к NS3/4a, полученных стандартными методами. Моноклональные антитела были затем тестированы в формате ELISA против NS3/4a, денатурированного NS3/4a и антигена с200. Данные показывают, что анти-NS3/4а-моноклональные антитела сходным образом реагируют с NS3/4a и денатурированным NS3/4a на сероконверсионных панелях, показанных в таблице 10. Этот результат также является дальнейшим подтверждением того, что NS3/4a является конформационным по природе, так же, как и моноклональные антитела, которые можно изготовить с близкой реакционной способностью к выборкам образцов ранней с33с сероконверсии.
Соответственно, открыты новые способы анализа для обнаружения HCV. На основании вышеописанного становится понятно, что несмотря на то, что конкретные варианты осуществления данного изобретения были описаны в иллюстративных целях, могут быть внесены разнообразные изменения без изменения сущности и рамок данного изобретения.
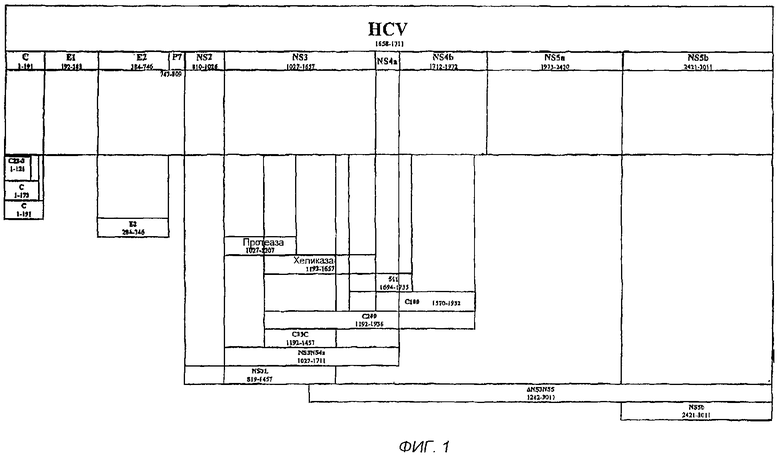




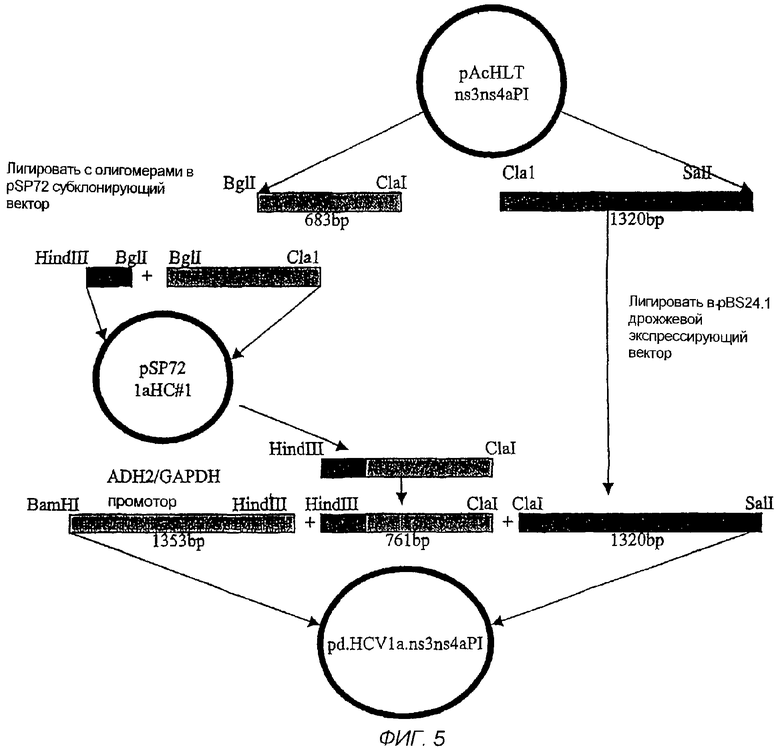
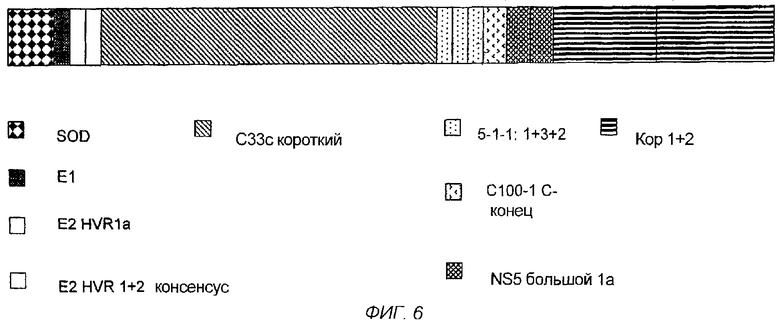





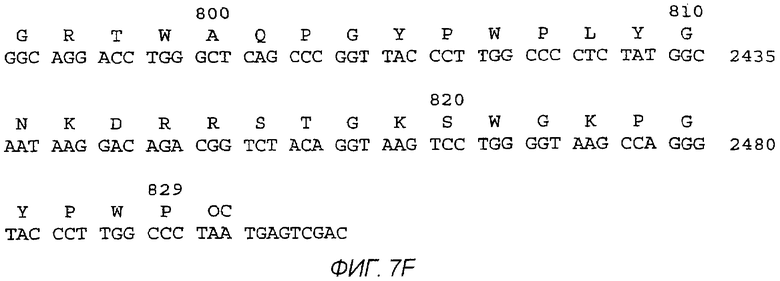

Изобретение относится к области медицины, а именно к диагностике гепатита С, и касается определения комплекса HCV-антиген/антитело и включает способ определения комплекса, образуемого между HCV-антикоровым антигеном и NS3/4a-антителом, который может распознавать как HCV антигены, так и антитела, присутствующие в образце, при использовании единой твердой основы, а также и твердые подложки для применения в иммуноанализе. Преимущество изобретения заключается в повышении чувствительности диагностики гепатита С. 16 н. и 31 з.п. ф-лы, 10 табл., 8 ил.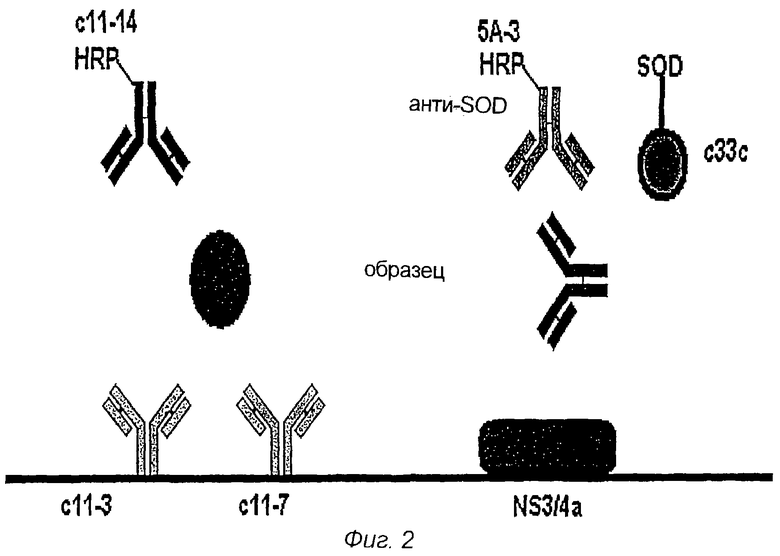
(a) получение твердой подложки для иммуноанализа по п.1;
(b) соединение биологического образца с указанной твердой подложкой в условиях, которые позволяют антигенам и антителам HCV, если они присутствуют в биологическом образце, связываться с указанным по меньшей мере одним антикоровым антителом и указанным NS3/4а-эпитопом, соответственно;
(c) добавление к твердой подложке со стадии (b) в условиях комплексообразования (i) первого антитела с обнаруживаемой меткой, где указанное первое антитело с обнаруживаемой меткой является HCV-антикоровым антителом с обнаруживаемой меткой, где указанное меченое антикоровое антитело направлено против корового эпитопа HCV, отличного по меньшей мере от одного антикорового антитела, связанного с твердой подложкой; (ii) антигена, который реагирует с HCV-антителом из биологического образца, способным реагировать с указанным конформационным NS3/4а-эпитопом; и (iii) второго антитела с обнаруживаемой меткой, где указанное второе антитело с обнаруживаемой меткой способно реагировать с антигеном (ii);
(d) обнаружение комплексов, образующихся между антителами и антигенами, если они есть, в качестве указания на HCV-инфекцию в биологическом образце.
(a) получение твердой подложки для иммуноанализа в соответствии с п.2;
(b) соединение биологического образца с твердой подложкой в условиях, которые позволяют антигенам и антителам HCV, если они присутствуют в биологическом образце, связываться с указанными по меньшей мере двумя антикоровыми антителами и указанным конформационным NS3/4a-эпитопом, соответственно;
(c) добавление к твердой подложке со стадии (b) в условиях комплексообразования (i) первого антитела с обнаруживаемой меткой, где указанное первое антитело с обнаруживаемой меткой является HCV-антикоровым антителом с обнаруживаемой меткой, где указанное меченое антикоровое антитело направлено против HCV-корового эпитопа, отличного от по меньшей мере двух антикоровых антител, связанных с твердой подложкой; (ii) эпитопа из с33с-области полипротеина HCV, слитого с аминокислотной последовательностью hSOD; и (iii) второго антитела с обнаруживаемой меткой, где указанное второе антитело с обнаруживаемой меткой способно реагировать с указанной аминокислотной последовательностью hSOD;
(d) обнаружение комплексов, образованных между антителами и антигенами, если они есть, в качестве указания на инфекцию HCV в биологическом образце.
(a) получение твердой подложки для иммуноанализа, в соответствии с п.9;
(b) соединение биологического образца с указанной твердой подложкой в условиях, которые позволяют антигенам и антителам HCV, если они присутствуют в биологическом образце, связываться с указанными по меньшей мере двумя антикоровыми антителами и указанным конформационным NS3/4a-эпитопом, соответственно;
(с) добавление к твердой подложке со стадии (b) в условиях комплексообразования (i) первого антитела с обнаруживаемой меткой, где указанное первое антитело с обнаруживаемой меткой является HCV-антикоровым антителом с обнаруживаемой меткой, где указанное меченое антикоровое антитело направлено против HCV-корового эпитопа, отличного от по меньшей мере двух антикоровых антител, связанных с твердой подложкой; (ii) эпитопа из с33с-области полипротеина HCV, слитого с аминокислотной последовательностью hSOD; и (iii) второго антитела с обнаруживаемой меткой, где указанное второе антитело с обнаруживаемой меткой способно реагировать с аминокислотной последовательностью hSOD;
(d) обнаружение комплексов, образованных между антителами и антигенами, если они есть, в качестве указания на инфекцию HCV в биологическом образце.
(a) получение твердой подложки для иммуноанализа в соответствии с п.7;
(b) соединение биологического образца с указанной твердой подложкой в условиях, которые позволяют антигенам и антителам HCV, если они присутствуют в биологическом образце, связываться с указанным по меньшей мере одним антикоровым антителом, указанным конформационным NS3/4a-эпитопом и указанным слитым антигеном со множественными эпитопами, соответственно;
(c) добавление к твердой подложке со стадии (b) в условиях комплексообразования (i) первого антитела с обнаруживаемой меткой, где указанное первое антитело с обнаруживаемой меткой является HCV-антикоровым антителом с обнаруживаемой меткой, где указанное меченое антикоровое антитело направлено против HCV-корового эпитопа, отличного по меньшей мере от одного антикорового антитела, связанного с твердой подложкой; (ii) первого и второго антигенов, которые реагируют с HCV-антителом из биологического образца, способным реагировать с указанным NS3/4а-эпитопом и указанным слитым антигеном со множественными эпитопами, соответственно; и (iii) второго антитела с обнаруживаемой меткой, где указанное второе антитело с обнаруживаемой меткой способно реагировать с антигенами (ii);
(d) обнаружение комплексов, образованных между антителами и антигенами, если они есть, в качестве указания на инфекцию HCV в биологическом образце.
(a) получение твердой подложки для иммуноанализа в соответствии с п.12;
(b) соединение биологического образца с указанной твердой подложкой в условиях, которые позволяют антигенам и антителам HCV, если они присутствуют в биологическом образце, связываться с указанными по меньшей мере двумя антикоровыми антителами, указанным конформационным NS3/4а-эпитопом и указанным слитым антигеном со множественными эпитопами, соответственно;
(c) добавление к твердой подложке со стадии (b) в условиях комплексообразования (i) первого антитела с обнаруживаемой меткой, где указанное первое антитело с обнаруживаемой меткой является HCV-антикоровым антителом с обнаруживаемой меткой, где указанное меченое антикоровое антитело направлено против HCV-корового эпитопа, отличного от по меньшей мере двух антикоровых антител, связанных с твердой подложкой; (ii) эпитопа из с33с-области полипротеина HCV, слитого с аминокислотной последовательностью hSOD, и эпитопа из с22-области полипротеина HCV, слитого с аминокислотной последовательностью hSOD; (iii) второго антитела с обнаруживаемой меткой, где указанное второе антитело с обнаруживаемой меткой способно реагировать с аминокислотной последовательностью hSOD;
(d) обнаружение комплексов, образованных между антителами и антигенами, если они есть, в качестве указания на инфекцию HCV в биологическом образце.
(a) получение твердой подложки; и
(b) связывание с ней по меньшей мере одного антикорового антитела против вируса гепатита С (HCV) и по меньшей мере одного выделенного NS3/4a-конформационного эпитопа HCV.
(a) получение твердой подложки; и
(b) связывание с ней по меньшей мере двух антикоровых антител против вируса гепатита С (HCV) и выделенного конформационного NS3/4a-эпитопа HCV.
(a) полинуклеотид в соответствии с п.44;
(b) и контрольные элементы, оперативно связанные с указанным полинуклеотидом, посредством которого кодирующая последовательность может быть транскрибирована или транслирована в клетке-хозяине.
(a) получение популяции линии клеток по п.46; и
(b) культивирование указанной популяции клеток в условиях, при которых экспрессируется слитый антиген со множественными эпитопами, кодируемый кодирующей последовательностью, присутствующей в указанном рекомбинантном векторе.
Приоритет по пунктам и признакам:
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| Двигатель внутреннего горения | 1921 |
|
SU450A1 |
| US 5683864 A, 04.11.1997. | |||

