Изобретение относится к материалам и методологиям для устранения распространения инфекции, вызываемой вирусом гепатита C (HCV). Более конкретно оно относится к полипептидам, применяемым в качестве иммунологических реагентов при определении, предупреждении и лечении инфекций, вызванных HCV.
Предпосылки изобретения
HCV был впервые идентифицирован и охарактеризован в качестве причины гепатита ни A, ни B (NANBH) Houghton et al. Это привело к открытию ряда основных и специфических полипептидов, применимых в качестве иммунологических реагентов. Смотрите, например, Houghton et al., N 318,216; Houghton et al., EPO Pub. N 388,232; Choo et al., Science (1989) 244: 359-362; Kuo et al., Science (1989) 244: 362-364; Houghton et al., Hepatology (1991) 14: 381-388. Эти публикации представляют информацию с обширным кругом источников в основном по HCV, так же как по производству и использованию полипептидных иммунологических реагентов из HCV. Для краткости поэтому описание этих публикаций, в частности, включены здесь в виде ссылок.
Другие с готовностью применили и расширили разработки Houghton et al. Смотрите, например, Highfield et al., UK Pat.App.2.239245 (The Welcome Foundation Ltd); Wahg, EPO Pub. N 442394 (United Biomedical Inc.); Leung et al. , EPO Pub. N 445423 (Abbott Laboratories); Habits et al., EPO Pub. N 451.891 (Akzo N. V.); Reyes et al., PCT Pub. N WO 91/15516 (Genelabs Inc.); Maki et al. , EPO Pub. N 468.657 (Tonen Corp.); и Kamada et al., EPO Pub. N 469.348 (Shionogi Seiyaku K.K.).
Чувствительные специфичные методы для проверки на и определения носителей HCV и зараженной HCV крови и продуктов из крови являются важным достижением медицины. Гепатит после переливания крови (PTH) встречается примерно у 10% больных, которым переливали кровь, и HCV составляли до 90% этих случаев. Главной проблемой при этом заболевании является частое развитие болезни до хронического поражения печени (25-55%). Лечение больных, так же как и предупреждение передачи HCV с кровью и продуктами крови или при тесном личном контакте, требует надежных диагностических и прогностических средств, таких как полипептиды HCV, для определения антител, связанных с HCV. Такие полипептиды также применяются в качестве вакцин и иммунотерапевтических и терапевтических средств для предупреждения и/или лечения заболевания.
Так как HCV является относительно новым возбудителем, существует постоянная потребность в выявлении дополнительных иммунологических реагентов, которые дадут возможность дальнейшего изучения клинического течения болезни и эпидемиологии HCV среди населения.
Описание изобретения
Изобретение относится к определению характеристик новых эпитопов HCV. Определение характеристик этих эпитопов дает возможность производства полипептидных продуктов, которые иммунологически реагируют с антителами к HCV и/или вызывают образование антител против HCV in vivo. Эти полипептидные продукты применимы в качестве стандартов или реагентов в диагностических тестах и/или в качестве компонентов вакцин. Антитела, включая, например, как поликлональные, так и моноклональные, направленные против HCV эпитопов, которые содержатся в этих полипептидных последовательностях, также применимы как реагенты, например, для диагностических тестов, в качестве терапевтических средств, при отборе противовирусных средств и для выделения/очистки полипептидов HCV или частиц.
В наиболее широком смысле настоящее изобретение направлено на получение полипептидов, содержащих вновь охарактеризованные эпитопы HCV, описанные здесь, разработку способов получения таких полипептидов (например, химического синтеза), разработку способов применения таких полипептидов (например, диагностика, вакцины и терапия). Иммунологическое исследование для определения антител против HCV, включающее инкубацию образца, подозреваемого на содержание антител против HCV, с полипептидом, описанным выше, в условиях, которые дают возможность образования комплекса антитело-антиген: и определение комплекса антитело-антиген, содержащего полипептид, также входит в объем притязаний изобретения. Еще одним аспектом изобретения является способ индукции иммунного ответа у субъекта против HCV, включающий введение индивидууму выделенного иммуногенного полипептида, содержащего эпитоп HCV, описанный здесь, в количестве, достаточном, чтобы вызвать иммунный ответ.
Вышеназванные объекты настоящего изобретения могут быть осуществлены с применением эпитопов HCV с формулой
aax -aay,
где aa обозначает аминокислоту;
x и y являются целыми числами, такими, что y-x ≥ 14;
aax-aay показывает часть аминокислотной последовательности фигуры 1;
и x выбирают из группы: 66, 413, 540, 1218, 1940, 2244, 2281, причем указанный полипептид может иметь необязательную последовательность N-концевых аминокислот, соответствующую последовательности полипептида HCV, прилегающей к N-концу аминокислоты aax, к которой она прикреплена, или необязательную последовательность C-концевых аминокислот, соответствующую последовательности полипептида HCV, прилегающую к C-концу аминокислоты aay, к которой она прикреплена, причем общее количество дополнительных аминокислот не превышает 15 для aa413-aa427, 4 для aa465-aa480, 85 для aa540-aa554, 10 для aa2244-aa2258, и 30 для aa2281-aa2300.
Краткое описание чертежей
Фиг.1 показывает полипротеин прототипного изолята HCV-HCV1.
Фиг.2 показывает составную последовательность кДНК HCV1
Фиг. 3 показывает нуклеотидную консенсусную последовательность человеческого изолята 23, варианты последовательности показаны ниже линии последовательности. Показаны также аминокислоты, кодируемые консенсусной последовательностью.
Фиг. 4 показывает нуклеотидную консенсусную последовательность человеческого изолята 27, вариантные последовательности показаны ниже линии последовательности.
Фиг.5 показывает вытянутые в ряд нуклеотидные последовательности человеческих изолятов 23 и 27 и HCV1. Гомологичные последовательности указаны символом (*). Негомологичные последовательности представлены маленькими буквами.
Фиг. 6 показывает вытянутые в ряд аминокислотные последовательности человеческих изолятов 23 и 27 и HCV1. Гомологичные последовательности указаны символом (*). Негомологичные последовательности представлены маленькими буквами.
Фиг. 7 показывает сравнение выстроенных в ряд составных нуклеотидных последовательностей изолятов Thorn, EC1, HCT ≠ 18 и HCV1.
Фиг. 8 показывает сравнение нуклеотидных последовательностей ЕС10 и композит последовательности HCV1; последовательность ЕС10 находится на линии над точками, и последовательность HCV1 находится на линии ниже точек.
Фиг. 9 показывает сравнение аминокислотных последовательностей 117-308 (относительно HCV1), кодируемых в "EnvL" областях консенсусных последовательностей человеческих изолятов НСТ#18, JH, 2J, JH27, Thorne, EC1 и HCV1.
Фиг. 10 показывает сравнение аминокислотных последовательностей 330-360 (относительно HCV1), кодируемых в областях "EnvL" консенсусных последовательностей человеческих изолятов НСТ ≠ 18, JA23, JH27, Thorne, ECT, HCV1.
Способы осуществления изобретения
Полное перечисление публикаций, на которые ссылаются здесь, можно найти в разделах "Предпосылки создания изобретения" или "Библиография".
1. Определения
"Вирус гепатита C" или "HCV" относится к научнопризнанному вирусному виду, патогенные штаммы которого вызывают NANBH и из них получены аттенуированные штаммы или дефектные интерферирующие частицы. Смотрите в основном публикации, процитированные в разделе, озаглавленном "Предпосылки создания изобретения". Геном HCV содержит PHK. Известно, что содержание PHK вирусы имеют относительно высокие степени спонтанных мутаций, т.е. по сообщениям, порядка от 10-3 до 10-4 на включенный нуклеотид (Fields and Knipl (1986)). Поэтому из-за гетерогенности и текучести генотипа, присущей РНК вирусам, существует множество штаммов/изолятов, которые могут быть вирулентными или эвирулентными, внутри вида HCV. Распространение, идентификация, обнаружение и выделение различных штаммов HCV или изолятов хорошо представлены в литературе. Кроме того, раскрытое здесь позволяет получить диагностические препараты и вакцины для различных штаммов/изолятов, так же как и композиции, а также разработать способы, которые применимы при процедуре отбора на противовирусные средства для фармакологических целей, таких как средства, которые подавляют репликацию HCV.
Здесь раскрывается информация по нескольким разным штаммам/изолятам HCV, в частности по штамму или изоляту СДС/HCV/ (также называемого). Информация, полученная по одному штамму или изоляту, такая как по части геномной или аминокислотной последовательности, достаточна, чтобы дать возможность опытным специалистам, используя стандартные методики, выделить новые штаммы/ изоляты и определить, являются ли такие новые штаммы/изоляты HCV. Для примера несколько различных штаммов/изолятов описаны ниже. Эти штаммы, которые были получены из ряда человеческих сывороток (и из разных географических областей), были выделены с использованием информации по геномной последовательности HCV1.
Представленная здесь информация показывает, что HCV может быть отдаленно родственен флавивирусам. Семейство Flavivirus содержит большое число вирусов, которые являются мелкими, оболоченными возбудителями человека. Морфология и структура флавивирусных частиц известны и рассмотрены у Brinton (1986). В основном, что касается морфологии, флавивирусы содержат центральный нуклеокапсид, окруженный двойными липидными слоями. Вирионы являются сферическими и имеют диаметр около 40-50 нм. Их нуклеоиды имеют примерно 25-30 нм в диаметре. По наружной поверхности оболочки вириона расположены выросты, которые имеют длину около 5-10 нм с концевыми вздутиями диаметром примерно 2 нм. Типичными примерами представителей этого семейства являются вирус желтой лихорадки, вирус западного Нила, вирус лихорадки Денге. Они обладают положительно-цепочечными РНК-геномами (~ 11000 нуклеотидов), которые немного больше, чем таковые HCV, и кодируют полипротеиновый предшественник примерно в 3500 аминокислот. Отдельные вирусные белки отделяются от этого предшественника-полипептида.
Была выведена структура генома и нуклеотидная последовательность геномной РНК HCV. Геном, по-видимому, является одноцепочечной РНК, содержащей ~ 10000 нуклеотидов. Геном является положительно-цепочечным и обладает постоянной трансляционной открытой считывающей рамкой (ORF), которая кодирует полипротеин размером примерно 3000 аминокислот. В ORF структурные протеин(ы), по-видимому, кодируются примерно в первой четверти N-концевой области, причем большая часть полипротеина ответственна за неструктурные белки. При сравнении со всеми известными вирусными последовательностями наблюдаются небольшие, но важные ко-линейные гемологии с неструктурными белками семейства флавивирусов и с пестивирусами (которые, как теперь считается, являются частью семейства Flavivirus).
На основе предполагаемых аминокислот, кодируемых в нуклеотидной последовательности HCV1 и других данных возможные протеиновые домены кодируемого полипротеина HCV, так же как и приблизительные границы, являются следующими:
Предполагаемый домен - Приблизительные границы (номера аминокислот)
C (нуклеокапсидный белок) - 1-131
E1 (белок оболочки вируса) - 192-383
E2 /NS1/ (оболочка?) - 384-800
(неизвестная функция) - 800-1050
(протеаза?) - 1050-1650
(неизвестная функция) - 1651-2100
(полимераза) - 2100-3011 (конец)
Эти домены являются, однако, предварительными. Например, граница EI-NS2 находится в области 750-810, и NS3-NS4 граница находится около 1640-1650. Существует также доказательство того, что вариант C - 191 не является предшественником, который далее обрабатывается (например, до длины примерно 170 аа), и что белки NS2, NS4 и NS5 каждый, далее перерабатываются в два зрелых белка.
Как предполагается, различные штаммы, изоляты или подтипы HCV содержат варианты аминокислот и нуклеиновых кислот по сравнению с HCV1. Предполагают, что многие изоляты покажут большую степень (т.е. более, что примерно 40%) гомологии в общей аминокислотной последовательности в сравнении с HCV1. Однако, возможно, также обнаружится, что существуют другие менее гомологичные изоляты HCV. Эти изоляты могут быть определены как HCV по разным критериям, таким как, например, ORF в пределах от примерно 9000 нуклеотидов до примерно 12000 нуклеотидов, кодирующая полипротеин, сходный по размеру с таковым HCV1, кодируемый полипротеин с гидрофобными и/или антигенными характеристиками, сходными с таковыми HCV1, и наличие колинейных пептидных последовательностей, которые сохраняются из HCV1. Кроме того, был бы положительно-цепочечной РНК.
HCV кодируется по крайней мере один эпитоп, который иммунологически сравним с эпитопом в полипротеине HCV1. Эпитоп является уникальным для HCV по сравнению с ранее известными флавивирусами. Уникальность эпитопа может быть определена по его иммунологической реактивности с антителами к известным видам флавивирусов. Методы для определения иммунологической реактивности известны в данной области науки, например, здесь представлены радиоиммуноисследование, определение по ELISA путем гемагглютинации и еще несколько примеров подходящих методик для исследования.
В дополнение к вышеописанному могут применяться для идентификации штамма/изолята как HCV следующие параметры гомологии нуклеиновых и аминокислот отдельно или в комбинации. Так как штаммы HCV и изоляты являются эволюционно родственными, ожидается, что общая гомология геномов на нуклеотидном уровне может быть на уровне 10% или больше, возможно, будет около 40% или больше, возможно, примерно 60% или больше и даже более вероятно около 80% или больше; и кроме того, будут существовать соответствующие прилегающие последовательности из по крайней мере около 13 нуклеотидов. Необходимо отметить, что вариабельные и гипервариабельные области присутствуют в геноме HCV; поэтому ожидается, что гомология в этих областях будет значительно меньше, чем гомология в целом по геному. Соответствие между предполагаемой геномной последовательностью штамма HCV и, например, последовательностью кДНК СДС/HCV1 можно определить по методикам, известным в данной области науки. Например, оно может быть определено путем прямого сравнения данных по последовательности полинуклеотида из предполагаемого HCV, и последовательностями кДНК, описанными здесь. Например, оно может быть также определено путем гибридизации полинуклеотидов в условиях, при которых образуются стабильные дуплексы между гомологичными областями (например, теми, которые использовались бы до расщепления S1), за которым следует расщепление с помощью специфичных нуклеза одноцепочечных нуклеиновых кислот с последующим определением размера фрагментов расщепления.
Благодаря эволюционному родству штаммов или изолятов HCV, предполагаемые штаммы или изоляты HCV идентифицируются по их гомологии на полипептидном уровне. В основном ожидается, что штаммы и изоляты HCV будут гомологичны по крайней мере на 10%, более чем примерно на 40%, возможно более чем примерно на 70%, и даже более вероятно более чем примерно на 80%, и некоторые могут быть гомологичны более чем примерно на 90% на полипептидном уровне. Методики для определения гомологии аминокислотных последовательностей известны специалистам. Например, можно прямо определить аминокислотную последовательность и сравнить с представленными здесь последовательностями. Или же иначе может быть определена нуклеотидная последовательность материала генома предполагаемого HCV (обычно через промежуточную кДНК), может быть определена кодируемая в ней аминокислотная последовательность и проведено сравнение соответствующих областей.
Как использовано здесь, полинуклеотид, "полученный из" указанной последовательности, относится к полинуклеотидной последовательности, которая содержит последовательность, примерно, по крайней мере, около 6 нуклеотидов, предпочтительно, по крайней мере, около 8 нуклеотидов, более предпочтительно, по крайней мере, около 10-12 нуклеотидов, и даже более предпочтительно, по крайней мере, около 15-20 нуклеотидов, соответствующих области указанной нуклеотидной последовательности. "Соответствующих" означает гомологичных или комплементарных указанной последовательности. Предпочтительно последовательность области, из которой получен полинуклеотид, гомологична или комплементарна последовательности, которая уникальна для генома HCV. Уникальна или нет последовательность для HCV генома можно определить методами, известными специалистам. Например, последовательность можно сравнить с последовательностями в банках данных (по данным на дату приоритета), например, в Genebank, чтобы определить, присутствует ли она у неинфицированных хозяев или других микроорганизмов. Последовательность можно также сравнить с известными (по данным на дату приоритета) последовательностями других вирусных агентов, включая те, которые, как известно, вызывают гепатит, например НАУ, НВУ и HDV, и представителями семейства Flaviviridae. Соответствие или несоответствие полученной последовательности другим последовательностям можно также определить путем гибридизации в соответствующих строгих условиях. Методы гибридизации для определения комплементарности последовательностей нуклеиновых кислот известны в данной отрасли. Смотрите, например, Maniatis et al. (1982). Кроме того, неподходящие пары дуплексных полинуклеотидов, образованные при гибридизации, могут быть определены известными методами, включая, например, расщепление нуклеазой, такой как SI, которая специфически расщепляет одноцепочечные области в дуплексных полинуклеотидах. Области, из которых могут быть "получены" типичные последовательности ДНК, включают (но не ограничиваются), например, области, кодирующие специфические эпитопы, так же как и несчитываемые и/или нетранслируемые области.
Полученный полинуклеотид необязательно физически получен из показанной нуклеотидной последовательности, но может производиться другим путем, включая, например, химический синтез или репликацию, или обратную транскрипцию или транскрипцию ДНК. Кроме того, комбинации областей, соответствующих этой указанной последовательности, могут быть модифицированы известными специалистам способами, чтобы соответствовать намеченному применению.
Подобным же образом полипептид или аминокислотная последовательность, "полученная из" указанной аминокислотной или последовательности нуклеиновой кислоты, соответствует полипептиду, имеющему аминокислотную последовательность, идентичную последовательности полипептида, кодируемого в этой последовательности, или его части, причем эта часть состоит из, по крайней мере, 3-5 аминокислот и более предпочтительно, по крайней мере, мере на 8-10 аминокислот, и даже более предпочтительно, по крайней мере, 10-15 аминокислот, или которая способна иммунологически идентифицироваться с помощью полипептида, кодируемого в этой последовательности. Эта терминология также включает полипептид, экспрессируемый указанной последовательностью нуклеиновой кислоты.
Рекомбинантный или производный полипептид необязательно транслируется с указанной последовательности нуклеиновой кислоты; он может производиться другим путем, включая, например, химический синтез или экспрессию рекомбинантной системы экспрессии или выделение из HCV, включая мутированные HCV. Рекомбинантный или производимый полипептид может включать один или более аналогов аминокислот или неприродных аминокислот в своей последовательности. Методы встройки аналогов аминокислот в последовательность известны в этой области науки. Он может также включать одну или более меток, которые известны опытным специалистам.
Термин "рекомбинантный полинуклеотид", так как использован здесь, подразумевает полинуклеотид геномной нДНК, полусинтетического или синтетического происхождения, который благодаря своему происхождению или обработке: (1) не связывается со всем или с частью полинуклеотида, с которым он связывается в природе, (2) соединяется с полинуклеотидом, отличным от того, с которым он связывается в природе, кили (3) не встречается в природе.
Термин "полинуклеотид", который использован здесь, относится к полимерной форме нуклеотидов любой длины, или рибонуклеотидов, или дезоксирибонуклеотидов. Этот термин относится только к первичной структуре молекулы. Таким образом этот термин охватывает удвоенные и одноцепочечные ДНК и РНК. Он также заключает в себе известные типы модификаций, например метки, которые известны специалистам, метилирование, "шапочки", замещение одного или более естественно присутствующих нуклеотидов аналогом, межнуклеотидными модификациями, такими как, например, нуклеотиды с незаряженными связями (например, метилфосфонаты, фосфотриэфиры, фосфорамидаты, карбанаты и т.д.) и с заряженными связями (например, фосфоротиосаты, фосфородитисаты и т.д.), нуклеотиды, содержащие подвешенные заместители, такие как, например, белки (включая, например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и т.д.), нуклеотиды с включениями (например, акридином, псораленом и т.д.), нуклеотиды, содержащие хелаторы (например, металлы, радиоактивные металлы, бор, окислительные металлы и т.д.), нуклеотиды, содержащие алкилаторы, нуклеотиды с модифицированными связями (например, альфааномерными нуклеиновыми кислотами и т.д.), так же как и немодифицированные формы полинуклеотидов.
"Очищенный" полипептид относится к полипептиду, находящемуся в состоянии, в котором по существу свободен от других полипептидов, т.е. в составе, который содержит минимум около 50% по весу (желаемый полипептид/общее количество полипептидов в композиции), предпочтительно, минимум, примерно 70%, и даже более предпочтительно, минимум, примерно 90% желаемого полипептида, не учитывая непротеиновые материалы в композиции. Методики очистки вирусных полипептидов известны в данной области науки. Очищенным антителам дается подобное же определение.
"Рекомбинантные клетки-хозяева", "клетки-хозяева", "клетка", "клеточные линии", "культуры клеток" и другие такие термины, обозначающие микроорганизмы или линии высших аукариотических клеток, культивируемых как одноклеточные существа, относятся к клеткам, которые могут быть использованы или использовались в качестве реципиентов для рекомбинантного вектора или другого перекоса ДНК, и включают потомство исходной клетки, которая была трансфицирована. Понятно, что потомство единственной родительской клетки может быть необязательно полностью идентично по морфологии или по геномной или общей ДНК хромосомного набора исходной родительской, из-за естественных, случайных или преднамеренных мутаций.
"Репликон" является любым генетическим элементом, например плазмидой, хромосомой, вирусом, космидой и т. д., который ведет себя как автономная единица полинуклеотидной репликации внутри клетки; т.е. способна к репликации под своим собственным контролем.
"Вектор" является репликоном, к которому присоединен другой полинуклеотидный сегмент, так чтобы вызвать репликацию и/или экспрессию присоединенного сегмента.
"Управляющая последовательность" относится к полинуклеотидным последовательностям, которые необходимы для воздействия на экспрессию кодирующих последовательностей, с которыми они связаны. Природа таких управляющих последовательностей отличается в зависимости от организма-хозяина; в прокариотах также управляющие последовательности в основном включают промотор, сайт связывания с рибосомой и терминаторы; в эукериотах в основном также управляющие последовательности включают промоторы, терминаторы и в некоторых случаях анхансеры. Термин "управляющие последовательности" предназначен для того, чтобы включить в себя, как минимум, все компоненты, присутствие которых необходимо для экспрессий, и может также включать дополнительные компоненты, присутствие которых целесообразно, например лидерные последовательности.
"Оперативно связанные" относятся к близкому расположению, при котором компоненты, так названные, находятся в связи, позволяющей им функционировать предназначенным им способом, управляющая последовательность связана таким образом, что экспрессия кодирующей последовательности достигается при условиях, совместимых с управляющими последовательностями.
"Открытая считывающая рамка" (ORF) является областью полинуклеотидной последовательности, которая кодирует полипептид; эта область может представлять часть кодирующей последовательности или общей кодирующей последовательности.
"Кодирующая последовательность" является полинуклеотидной последовательностью, которая считывается на мРНК и/или транслируется в полипептид, когда помещается под управление соответствующих регуляторных последовательностей. Границы кодирующей последовательности определяются путем трансляции стартового кодона на 5' - конце и трансляции стоп-кодона на 3'-конце. Кодирующая последовательность может включать, но не ограничивается мРНК, кДНК и рекомбинантными полинуклеотидными последовательностями.
"Иммунологически идентифицируемый с помощью/как" относится к наличию эпитопа(ов) и полипептида(ов), которые также присутствуют в указанном(ых) полипептиде(ах), обычно белках HCV. Иммунологическая идентичность может быть определена путем связывания с антителами и/или конкуренции при связывании; эти методики известны опытным специалистам.
Употребленный здесь термин "эпитоп" относится к антигенной детерминанте полипептида. Эпитоп мог включать 3 или более аминокислот, которые определяют сайт связывания антитела. В основном эпитоп состоит из по крайней мере 5 аминокислот и иногда состоит из по крайней мере 8 аминокислот. Методы картирования эпитопа известны в этой области науки.
Полипептид является "иммунологически реактивным" с антителом, когда он связывается с антителом благодаря распознаванию антителом специфического эпитопа, содержащегося в полипептиде. Иммунологическая реактивность может быть определена путем связывания антител, более конкретно, при помощи кинетики связывания антител и/или путем конкуренции в связывании с использованием конкурентного(ых) компонента(ов) известного полипептида(ов), содержащих эпитоп, против которого направлено антитело. Методики определения, является ли полипептид иммунологически реактивным с антителом, известны в этой области науки.
Использованный здесь термин "антитело" относится к полипептиду или группе полипептидов, которые содержат по крайней мере один соединяющий антитела сайт. "Соединяющий сайт антитела" или "связывающий домен" формируется в результате складывания различных доменов молекул(ы) антител(а) для образования трехмерных пространственных формирований с внутренней формой поверхности и распределением зарядов, комплементарным этим характерным чертам эпитопа антитела, который дает возможность иммунологической реакции с антигеном. Сайт связывания антитела может формироваться доменом тяжелой и/или легкой цепи (Vн и V1 соответственно), которые формируют гипервариабельные петли, обеспечивающие связывание антигена. Термин "антитело" включает, например, позвонковые антитела, гибридные антитела, химерные антитела, измененные антитела, неполные антитела, белки Fab и антитела с единственным доменом.
Использованный здесь термин "однодоменовое антитело" (d:b) это антитело, которое включает Vн домен, иммунологически реагирующий с указанным антигеном, dAb не содержит домена V1, но может содержать другие связывающие антиген домены, которые, как известно, существуют в антителах, например, каппа и лямбда домены. Методы получения dAb известны в этой области. Смотрите, например Ward et. al. (1989).
Антитела могут также содержат Vн и V1 домены, а также другие известные связывающие антиген домены. Пример этих типов антител и методы их получения известны в этой области науки (смотрите, например, патент США N 4816467, который приведен здесь в виде ссылки) и включают следующее. Например, "позвонковые антитела" относятся к антителам, которые являются тетрамерами или их комплексами, включающими легкие и тяжелые цепи, которые обычно собраны в "у" конфигурацию и которые могут иметь или не иметь ковалентные связи между цепями. В позвонковых антителах аминокислотные последовательности всех цепей конкретного антитела являются гомологичными цепями, обнаруженными в одном антителе, произведенным лимфоцитом, который продуцирует что антитела in situ или in vitro (например, в гибридомах). Позвонковые антитела обычно включают естественные антитела, например очищенные поликлональные антитела и моноклональные антитела. Примеры методов получения этих антител описаны ниже.
"Гибридные антитела" являются антителами, в которых одна пара тяжелых и легких цепей является гомологичной таковым в первом антителе, тогда как другая пара тяжелых и легких цепей гомологична таковым другого второго антитела. Обычно каждая из этих двух пар будет связываться различными эпитопами, в частности, на различных антигенах. Это дает в результате свойство "дивалентности", т.е. способности связывать два антигена одновременно. Такие гибриды могут также образовываться при использовании химерных цепей, как изложено ниже.
"Химерные антитела" - это антитела, в которых тяжелые и/или легкие цепи являются слитыми белками. Обычно константный домен цепей происходит из одного конкретного вида и/или класса, и вареабельные домены происходят из другого вида и/или класса. Сюда также включаются любые антитела, в которых одна из или обе из тяжелых или легких цепей составлены из комбинаций последовательностей, имитирующих последовательности антител из различных источников, являются ли эти источники представителями разных классов, или видов разного происхождения, и так или иначе точка слияния находится на границе вариабельной/константной областей. Таким образом возможно получить антитела, в которых ни константная, ни вариабельная область не имитируют известные последовательности антител. Затем становится возможным, например, построить антитела, вариабельная область которых имеет высокое специфическое сродство к конкретному антигену, или константная область которых может вызывать повышенную фиксацию комплемента, или получать другие улучшения свойств, которыми обладает константная область.
Другим примером являются "измененные антитела", которые относятся к антителам, в которых естественно встречающаяся аминокислотная последовательность в позвонковых антителах изменена. Используя методики рекомбинантной ДНК, антитела могут быть перестроены для получения желаемых характеристик. Возможных изменений много и они варьируют от изменения одной или более аминокислот до полной перестройки области, например константной области. Изменения в константной области в основном предназначены для того, чтобы достичь желаемых характеристик клеточной обработки, т.е. изменений в фиксации комплемента, взаимодействии с мембранами и других эффекторных функций. Могут быть произведены изменения в вариабельной области, чтобы изменить характеристики связывания антигена. Антитела могут быть также сконструированы так, чтобы помочь специфической доставке молекулы или вещества к специфическому клеточному или тканевому сайту. Желаемые изменения могут быть произведены известными в молекулярной биологии методами, например рекомбинантные методики, сайтнаправленный мутагенез и т.д.
Еще один пример, что "неполные антитела", которые являются агрегатами, включающими димер тяжелой цепи/легкой цепи, граничащий с Fc (т.е. константной) областью второй тяжелой цепи. Этот тип антитела исключает антигенную модуляцию. Смотрите, например Glennie et.al. (1982).
В определение антител включаются также "Fab" фрагменты антител. Область "Fab" относится к тем частям тяжелых и легких цепей, которые приблизительно эквивалентны или аналогичны последовательностям, которые содержатся в ответвленной части тяжелых и легких цепей, и которые, как было показано, демонстрируют иммунологическое связывание со специфическим антигеном, но у которых отсутствует эффекторная часть Fc. "Fab" включает агрегаты одной тяжелой и одной легкой цепи (обычно известные как "Fab"), так же как и тетрамеры, содержащие 2H и 2L цепи (называемые F (ab)2), которые способны селективно реагировать с указанным антигеном или антигенным семейством. "Fab" антитело можно разделить на подсерии, биологические таковым, описанным выше, т. е. "позвонковые Fab", гибридные Fab", "химерные Fab" и "измененные Fab". Методы получения "Fab" фрагментов антител известны в этой области науки и включают, например, протеолиз и синтез с помощью рекомбинантных методик.
В термин "антитела" также включаются одноцепочечные связывающие антиген (SCA) белки, такого типа, который описан в статье в соавторстве с Schlom J. в выпуске Cancer Research от 15 июня 1992 г. (так же как и в статьях, процитированных здесь).
Так, как он использован здесь, термин "иммуногенный полипептид" обозначает полипептид, который вызывает клеточный и/или гуморальный иммунный ответ, один или связанный с носителем в присутствии или отсутствии адъюванта.
Термин "полипептид" относится к полимеру из аминокислот безотносительно к конкретной длине продукта; таким образом, пептиды, олигопептиды и белки включены в определение полипептида. Этот термин также не относится к или не допускается модификации полипептида после экспрессии, например, гликоэклирования, ацетилирования, фосфорилирования и тому подобное. В это определение включаются, например, полипептиды, содержащие один или более аналогов аминокислот (включая, например, неприродные аминокислоты, и т.д.), полипептиды с замещенными связями, так же как и другие модификации, известные в этой области, как встречающиеся в природе, так и в природе несуществующие.
"Трансформация" так, как использована здесь, относится к встройке экзогенного полинуклеотида в клетку-хозяина, независимо от метода, использованного для встройки, например, прямого поглощения, транслукции, f-спаривания или электропорации. Экзогенный полинуклеотид может быть сохранен в качестве неинтегрированного вектора, например плазмиды, или, напротив, может интегрироваться в геном хозяина.
"Лечение" так, как употребляется здесь, относится к профилактике и/или терапии.
"Индивидуум" так, как используется здесь, относится к позвоночным, конкретно к представителям видов млекопитающих, и включает, но не ограничивается животными (например, собака, кошка, крупный рогатый скот, свиньи, овцы, козы, кролики, мыши, крысы, морские свинки и т.д.) и приматами, включая обезьян, шимпанзе, бабуинов и людей.
Использованный здесь термин "смысловая цепь" нуклеиновой кислоты содержит последовательность, которая имеет токологию с последовательностью мРНК. "Антиосмысловая цепь" содержит последовательность, которая комплементарна последовательности "смысловой цепи".
Так, как использован здесь "геном с положительной цепью" вируса - это такой геном, в котором или РНК или ДНК является одноцепочечной и который кодирует вирусный(е) полипептид(ы). Примеры РНК-вирусов с положительной цепью включают Jogaviridal, Сoronaviridal, Retroviridal, Picornaviridal и Caliciviridal.
Сюда также включаются Flaviridal, которые первоначально были классифицированы как Jogaviridal. Смотрите Fields and Knipl (1986).
Так, использован здесь "компонент организма, содержащий антитела" относится к компоненту организма индивидуума, который является источником антител, представляющих интерес. Компоненты организма, содержащие антитела, известны в данной области знаний и включают (но не ограничиваются), например, плазму, сыворотку, спинномозговую жидкость, лимфу, наружное отделяемое дыхательного кишечного и мочеполового трактов, слезы, слюну, молоко, лейкоциты и миеломы.
Так, как использована здесь, "биологическая проба" относится к образцу ткани или жидкости, выделенному от индивидуума, включая (но не ограничиваясь), например, плазму, сыворотку, спинномозговую жидкость, лимфу, наружное отделяемое кожи, дыхательного, кишечного и мочеполового трактов, слезы, слюну, молоко, клетки крови, опухоли, органы, а также образцы, составляющих клеточную культуру in vitro (включая, но не ограничиваясь, средой, отвечающей стандарту, получаемой в результате роста клеток в среде для культивирования клеток, предположительно инфицированных вирусом клеток, рекомбинантных клеток и компонентов клеток).
II. Описание изобретения
В практических действиях настоящего изобретения будут применяться, если нет других указаний, общепринятые методики молекулярной биологии, микробиологии, рекомбинантной ДНК, и иммунологии, которые составляют методологическую основу этой области. Объяснение таких методик полностью дано в литературе. Смотрите, например, Maniatis, Fitsch & Sambrook, "Molecular Cloning; A Laboratory Manual" (1982); "DNA Cloning, Volumes I and II (D.N Glover ed. 1985); "Oligonucleotide Synthesis" (M. J. Gait ed, 1984); "Nucleic Acid Hybridization" (B. D. Hames & S.J. Higgins eds. 1984); "Transcription and Translation" (B. D. Hames & S.J. Higgins eds. 1984); "Animal Cell Culture" (R.I. Freshney ed. 1986); "Immobilized Cells And Enzymes" (IRL Press, 1986); B. Perbal, "A Practical Guide To Molecular Cloning" (1984); the series, "Methods in Enzymology" (Academic Press, Inc.); "Gene Transfer Vectors For Mammalian Cells" (J.H. Miller and M.P. Calos eds. 1987, Cold Spring Harbor Laboratory), Meth Enzymol Vol. 154 and Vol. 155 (Wu and Grossman, and Wu, eds. , respectively), Mayer and Walker, eds. (1987), "Immunochemical Methods In Cell And Molecular Biology" (Academic Press, London); Scopes (1987) "Protein Purification: Principles and Practice", Second 10 Edition (Springer-Verlag, N.Y.); and "Handbook of Experimental Immunology", Volumes I-IV (D.M. Weir and C. C. Blackwell eds 1986).
Все патенты, заявки на патенты и публикации, упомянутые здесь как выше, так и ниже, таким образом включены здесь в виде ссылки.
II. A. Усеченные полипептиды HCV
Применяемые полипептиды и процессы настоящего изобретения становятся возможными путем идентификации, приведенной ниже, новых эпитопов HCV. Знание этих эпитопов (или антигенных областей) дает возможность построения полипептидов, содержащих усеченные последовательности HCV, которые могут использоваться в качестве иммунологических реагентов.
Усеченные аминокислотные последовательности HCV, кодирующие, по крайней мере, один вирусный эпитоп, применимы как иммунологические реагенты. Например, полипептиды, включающие такие усеченные последовательности, могут использоваться в качестве реагентов в иммунологических исследованиях. Эти полипептиды также являются кандидатами в субъединицы антигенов в композициях для получения антисывороток или вакцин. Тогда как эти усеченные последовательности могут быть получены путем различных известных обработок природного вирусного белка, в основном предпочтительно производить синтетические или рекомбинантные полипептиды, включающие последовательность HCV. Полипептиды, включающие эти усеченные последовательности HCV, могут быть составлены полностью из HCV последовательностей (одного или более эпитопов, примыкающих или непримыкающих) или из последовательностей и гетерологичных последовательностей в едином белке. Применимые гетерологичные последовательности включают последовательности, которые можно выделить из рекомбинантного хозяина, которые усиливают иммунологическую реактивность эпитопа(ов) или облегчают соединение полипептида с подложкой при иммуноанализе или с венцинным носителем. Смотрите, например, EPO Pub N 116 201; US Pat, N 4722840; EPO Pub N 259149; US. Pat. N 4629783, описания которых включены здесь в виде ссылки.
Размер полипептида, включающего усеченные HCV последовательности, может широко варьировать, причем минимальный размер определяется последовательностью достаточного размера, чтобы создать HCV эпитоп, тогда как максимальный размер не имеет решающего значения. Для удобства максимальный размер обычно существенно не превышает того, который необходим, чтобы обеспечить желаемые HCV эпитопы и функцию(и) гетерологичной последовательности, если такая присутствует. Обычно усеченная аминокислотная последовательность HCV будет варьировать в пределах от примерно 5 (или 8) до примерно 100 аминокислот в длину. Более типично, однако, что HCV последовательность будет длиной максимум в примерно 50 (или 40) аминокислот, и иногда максимум в примерно 20, 25 или 30 аминокислот. Обычно желательно выбирать HCV последовательности по крайней мере примерно в 8, 10, 12 или 15 аминокислот.
Примеры усеченных HCV аминокислотных последовательностей (октамеров), которые являются пригодными, как описано здесь, представлены ниже в примерах. Необходимо понять, что эти пептиды не обязательно точно картируют один эпитоп. Иммуногенные части последовательности могут быть определены при помощи общепринятых методик и устранены из описанных последовательностей. Кроме того, здесь могут быть идентифицированы и описаны дополнительные усеченные аминокислотные последовательности HCV, которые включают эпитоп или являются иммуногенными.
Полипептидные продукты, содержащие усеченные HCV аминокислотные последовательности, раскрываемые ниже, могут быть получены в виде дискретных пептидов или включенными в полипептид большего размеры и могут найти применение, здесь описанное. При предпочтительном применении усеченные последовательности из доменов E1 и/или E2 употребляются в вакцинах в терапевтических продуктах. Хотя в общем любой из доменов может иметь какое-то применение для диагностики, особенно предпочтительны C, N S3, N S4, и N S5, причем комбинации C эпитопов с эпитопами из одного или более из N S3, N S4 или N S5 наболее предпочтительны.
П.В. Получение полипептидов
Доступность последовательностей ДНК, кодирующих аминокислотные последовательности HCV, дает возможность построения и экспрессии векторов, кодирующих антигенно активные области полипептида (Смотрите, например, фиг. 2). Эти антигенно активные области могут происходить из антигенов оболочки или из антигенов ядра или из антигенов, которые не являются структурно включенными, например, из связанных с полинуклеотидом белков, полинуклеотидной(ых) полимераз)(ы) и других вирусных белков, необходимых для репликации и/или сборки вирусной частицы. Фрагменты, кодирующие желаемые полипептиды, получаются, например, из клонов вирусной кДНК при использовании общепринятого рестрикционного расщепления или методами синтеза и лигируются в векторы, которые могут, например, содержать части соединяемых последовательностей, такие как бетагалактозидазы или супероксидисмутазы (SOD), предпочтительно. Методы и векторы, которые применимы для продукции полипептидов, содержащих соединяемые последовательности SOD, описаны в European Patent Office Publication N 0196056, опубликованном 1 октября 1986 г. Векторы, кодирующие соединяемые пептиды и полипептиды HCV, т.е. NANB5-1-1, NANB81 и C100-3, которые кодируются в составе кДНК HCV, описываются в разделах IV.B.I., IV.B.2 в IV.B.4 соответственно. Любая желаемая часть кДНК HCV, содержащая скрытую считывающую рамку (или ее синтетический вариант), может использоваться для экспрессии рекомбинантного полипептида, такого, как зрелый или (слитый) объединенный протеин.
Или иначе, полипептид, содержащий эпитопы HCV, может быть получен путем химического синтеза с использованием стандартных методик на основе аминокислотной последовательности, представленной на фигурах и в примерах.
ДНК, кодирующая желаемый полипептид, или в объединенной или не в объединенной форме, содержащая или не содержащая сигнальную последовательность, дающую возможность выделения, может быть лигирована в вектор экспрессии, приемлемый для какого-либо подходящего хозяина. В настоящее время для выработки рекомбинантных пептидов используются системы клеток-хозяев как эукариотов, так и прокериотов, и линии клеток-хозяев представлены в EPO Pub. N 318216. Затем полипептид выделяется из лизированных клеток или ее культуральной среды и очищается до степени, необходимой для предназначенного ему применения. Очистка может быть выполнена известными в этой области методами, например, дифференциальной экстракцией, солевым фракционированием, хроматографией на ионообменных смолах, эффинной хроматографией, центрифугированием и т. п. Смотрите, например, Methods in Enzimology на различные методы очистки белков. Такие полипептиды могут использоваться в качестве диагностикумов, или те, которые дают повышение нейтрализующих антител, могут оформляться в виде вакцин. Антитела, выработанные против этих полипептидов, могут также использоваться в качестве диагностикумов или для пассивной иммунотерапии. Кроме того, как обсуждается ниже, антител к этим полипептидам применимы, например, для выделения и идентификации частиц HCV.
Полипептиды HCV могут быть также выделены из вирионов HCV и обрезаны (усечены) (если это еще не сделано). Вирионы могут выращиваться в инфицированных HCV клетках в культуре ткани или в инфицированном хозяине.
П.С. Получение антигенных полипептидов и конъюгация с носителем
Антигенная область полипептидов в основном небольшая - обычно от 8 до 10 аминокислот или менее в длину. Фрагменты из такого небольшого числа, как 5 аминокислот, могут определять антигенную область. Эти сегменты могут соответствовать областям антигена HCV. В соответствии с этим, используя кДНК HCV в качестве основы, ДНК, кодирующая короткие сегменты полипептидов HCV, может экспрессироваться рекомбинантно или в виде объединенных белков, или в виде отдельных полипептидов. Кроме того, короткие аминокислотные последовательности можно удобно получить с помощью химического синтеза. В случаях, когда синтезированный полипептид правильно сформирован, так что представляет правильный эпитоп, но слишком мал, чтобы быть иммуногенным, полипептид может быть связан с подходящим носителем.
Для получения такого связывания в этой области науки известен ряд методик, включая образование дисульфидных связей при использовании N-сукцинимидил-3-(2-пиридилтио)пропионата (SpDp) и сунцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилата (SM CC), получаемых от Pirce Company, Rockford, Jllinois (если у пептида нет сульфгидрильной группы, таковая может быть получена добавлением цистеинового остатка). Эти реагенты создают дисульфидную связь между собой и пептидными цистеиновыми остатками на одном протеине и амидной связью через эпсилон-аминогруппу на лизине, или другие свободные аминогруппы других аминокислот. Известен ряд таких дисульфид/амидобразующих средств. Смотрите, например, Jmmun, Rev (1982), 62:185. Другие бифункциональные соединенные агенты образуют тиоэфирную, а не дисульфидную связь. Многие из этих тиоэфир-образующих агентов коммерчески доступны и включают реакционноспособные эфиры 6-малеимидокапроновой кислоты, 2-бромуксусной кислоты, 2-иодуксусной кислоты, 4-(N-малеимидометил)циклогексан-1-карбоновой кислоты и тому подобное. Карбоксильные группы могут быть активированы путем соединения их с сукцинимидом или 1-гидроксил-2-нитро-4-сульфоновой кислоты натриевой соли. В дополнительных методах связи антигенов применяется система ротавирус /"связывающий пептид", описанной в EPO Pub. N 259149, раскрытие которой включается здесь в виде ссылки. Вышеприведенный список не рассматривается как исчерпывающий, и ясно, что могут быть использованы модификации вышеназванных соединений.
Может использоваться любой носитель, который сам не вызывает образования антител, опасных для хозяина. Удобными носителями является обычно большие, медленно метаболизируемые молекулы, такие как протеины; полисахариды, такие, как латекс, функционализированный Сефарозой®, агароза, целлюлоза, целлюлозные гранулы и тому подобное; полимерные аминокилсоты, такие как полиглютаминовая кислота, полилизин и тому подобное; аминокислотные сополимеры; неактивные вирусные частицы, смотрите, например, раздел
II. D. Особенно пригодными протеиновыми субстратами являются сывороточные альбумины, гемоцианин моллюска "блюдечко" молекулы иммуноглобулинов, тироглобулин, овальбумин, столбнячный токсоид и другие белки, известные опытным специалистам.
II. E. Получение частичек гибридных иммуногенов, содержащих эпитопы HCV
Иммуногенность эпитопов HCV может быть также увеличена путем получения их в системах клеток млекопитающих или дрожжей, слитых или соединенных с протеинами, образующими частицы, такими как, например, те, которые связаны с поверхностным антигеном гепатита B. Смотрите, например, патент США 4622840. Конструкции, в которых эпитоп HCV связан непосредственно с последовательностями, кодирующими белок, образующий частицы, продуцируют гибриды, которые являются иммуногенными в отношении эпитопа HCV. Кроме того, все полученные векторы включают эпитопы, специфичные и имеющие различную степень иммуногенности, такие как, например, пре-S-пептид. Таким образом, частицы, построенные из белка, образующего частицы, которые включают HCV последовательности, являются иммуногенными в отношении HCV и HBV.
Поверхностный антиген гепатита (HBSAg), как было показано, формируется и собирается в частицы в S.ceverisial P.Valenjuela et al. (1982)), так же как, например, и в клетках млекопитающих (P.Vallnguela et.al. (1984)). Образование таких частиц, как было показано, усиливает иммуногенность мономерной субъединицы. Конструкции могут также включать иммунодоминантный эпитоп HBSAg содержащий 55 аминокислот предповерхностной (pre-S) области. Neurath et.al. (1984). Конструкции частицы Pre-S-HBSAg, экспрессируемой в дрожжах, раскрываются в EPO 174444, опубликованном 19 марта 1966 г.; гибриды, включающие гетерологичные вирусные последовательности для экспрессии в дрожжах, раскрываются в EPO 175261, опубликованном 26 марта 1966. Эти конструкции могут также экспрессироваться в клетках млекопитающих, таких как яйцеклетки китайского хомяка (CHO) при использовании SV40-дигидрофолатредуктазного вектора (wichelle et.al. (1984)).
Кроме того, части последовательности, кодирующей белок, образующий частицы, могут замещаться колонами, кодирующими эпитоп HCV. При этом замещении области, которые не нужны, чтобы опосредовать агрегацию единиц при образовании иммуногенных частиц в дрожжевых клетках или клетках млекопитающих, могут быть устранены, исключая таким образом дополнительные антигенные сайты HBV из конкуренции с эпитопом HCV.
II. E. Получение вакцин
Вакцины могут быть изготовлены из одного или более иммуногенных полипептидов, полученных из HCV. Эти полипептиды могут экспрессироваться в различных клетках-хозяевах (например, бактериях, дрожжах, клетках насекомых и млекопитающих), или иначе, могут выделяться из препаратов вирусов или производиться синтетически. Одно- или поливалентные вакцины против HCV могут включать один или более эпитопов из одного или более неструктурных белков. Эти вакцины могут состоять из, например, рекомбинантных белков. Эти вакцины могут состоять из, например, рекомбинантных полипептидов HCV и/или полипептидов, выделенных из вирионов. В частности, предполагается, что вакцины содержат один или более из следующих протеинов или субъединицы антигенов, полученных из них: E1, E2, C, NS2, NS3, NS4 и NS5. Особенно предпочтительны вакцины, включающие E1 и/или E2 или их субъединицы.
В дополнение к вышеприведенному, также возможно получить живые вакцины аттенуированных микроорганизмов, которые экспрессируют один или более комбинантных полипептидов HCV. Подходящие аттенуированные микроорганизмы известны в этой области и включают, например, вирусы (например, вакцинные вирусы (смотрите Brown et.al. (1986)), так же как и бактерии.
Получение вакцин, которые содержат иммуногенные полипептиды в качестве активных ингредиентов, известно специалистам. Обычно такие вакцины получают в инъекционной форме или в виде растворов в жидкостях или в виде суспензий; могут быть также получены формы в виде плотного вещества, пригодного для получения раствора или суспензии в жидкости перед инъекцией. Препарат может также быть эмульгированным или белок может инкапсулироваться в липосомы. Активные иммуногенные ингредиенты часто смешиваются с наполнителями, которые фармацевтически приемлемы и совместимы с активным ингредиентом. Подходящими наполнителями являются, например, вода, физраствор декстроза, глицерин, этанол или тому подобное и их комбинации. Кроме того, если желательно, вакцина может содержать небольшие количества вспомогательных веществ, таких как увлажняющие или эмульгирующие средства, буферные средства поддержания pH и/или адъюванты, которые повышают эффективность вакцины. Примеры адъювантов, которые могут быть эффективны, включают (но не ограничиваются): гидроксид алюминия, N-ацетилмурамил-L-треонилизоглютамин (тр-МДФ), N-ацетил-нор-мурамил-L-аланил-D-изоглютамин (CGP 11637, называемый нор-MDP), N-ацетилмурамил-L-аланил-D-изоглютаминил-L-аланил-2-(1'-2'-дипальмитоил -Sn-глицеро-3-гидрогсифосфорилокси)-этиламин (CGP 19835A, называемый МТР-PE) и RIBI, который содержит три компонента, экстрагируемые из бактерий, монофосфорил-липид A, лимиколет трегазолы и остов клеточной станки (MP + TDM + CWS) в 2% эмульсии сквален/ твин® BO. Эффективность адъюванта может определяться путем измерения количества антител, направленных против иммуногенного полипептида, содержащего антигенную последовательность HCV, полученных в результате применения этого полипептида в вакцинах, которые также содержали различные адъюванты.
Вакцины по общепринятым правилам вводятся парентерально путем инъекции, например, или подкожно или внутримышечно. Дополнительные лекарственные формы, которые пригодны для других способов введения, включают свечи и в некоторых случаях пероральные лекарственные формы. Для свечей традиционные связующие и основы могут включать, например, полиалкиленгликоли или триглицериды; такие свечи могут формироваться из смесей, содержащих активный ингредиент в пределах от 0,5% до 10%, предпочтительно 1-2%. Пероральные лекарственные формы включают также обычно используемые наполнители, как, например, фармацевтические сорта маннитола, лактозы, крахмала, стеарата магния, натрий-сахарин, целлюлозу, карбонат магния и тому подобное. Эти композиции принимают форму растворов, суспензий, таблеток, пилюль, капсул, лекарственных форм с длительным выделением или порошков и содержат 10-95% активного ингредиента, предпочтительно 25-70%.
Белки могут включаться в вакцины как нейтральные вещества или в форме солей. Фармацевтически приемлемые соли включают соли с кислотами (образованные со свободными аминогруппами пептида) и те, которые образуются с неорганическими кислотами, такими как, например, соляная или фосфорная кислоты, или с такими органическими кислотами, как уксусная, щавелевая, виноградная, малеиновая и тому подобное. Соли, образованные со свободными карбоксильными группами, могут также получаться с неорганическими основаниями, такими как, например, натрия, калия, аммония, кальция или железа гидроксиды, и с такими органическими основаниями, как изопропиламин, триметиламин, 2-этиламиноэтанол, гистидин, прокаин и тому подобное.
II. F. Дозировка и введение вакцин
Вакцины вводятся путем, совместимым с лекарственной формой дозы, и в таком количестве, которое будет профилактически и/или терапевтически эффективно. Количество, которое необходимо ввести, и которое в основном находится в пределах от 5 мкг до 250 мкг антигена на дозу, зависит от субъекта, которого нужно лечить, способности иммунной системы субъекта синтезировать антитела и степени желаемой защиты. Точные количества активного ингредиента, необходимые для введения, могут зависеть от оценки практикующуго врача и могут быть индивидуальны для каждого субъекта.
Вакцина может даваться по схеме однократного введения или предпочтительно по схеме многократного введения доз. Схема с многократным введением доз это та, при которой первичный курс вакцинации может состоять из 1-10 отдельных доз, за которым следуют другие дозы, даваемые через последовательные интервалы времени, необходимые, чтобы поддержать и усилить иммунный ответ, например, через 1-4 месяца для второй дозы, и если необходимо последующая (ие) доза(ы) после нескольких месяцев. Режим дозировки будет также, по крайней мере частично, определяться потребностями индивидуума и будет зависеть от оценки практикующего врача.
Кроме того, вакцина, содержащая иммуногенные антигены HCV, может вводиться вместе с другими иммунорегуляторными средствами, например иммуноглобулинами.
II. G. Получение антител против эпитопов HCV
Иммуногенные полипептиды, описанные здесь, используются для продукции антител, включая поликлональные и моноклональные. Если желательны поликлональные антитела, избранное млекопитающее (например, мышь, кролик, козел, лошадь и т. д.) иммунизируется иммуногенным полипептидом, несущим эпитоп (ы) HCV. Сыворотка от иммунизированного животного собирается с обрабатывается по известной процедуре. Если сыворотка, содержащая поликлональные антитела к эпитопу HCV, содержит антитела к другим антигенам, поликлональные антитела могут быть очищены путем иммуноаффинной хроматографии. Методики получения и обработки поликлональной антисыворотки известны в этой области, смотрите, например Mayer и Walker (1987). Или иначе поликлональные антитела могут быть выделены от млекопитающего, которого предварительно инфицировали HCV. Моноклональные антитела, направленные против эпитопов HCV,могут быть также легко получены опытными специалистами. Основная методология производства моноклональных антител с помощью гибридов хорошо известна. Непрерывные линии антителопродуцирующих клеток других методик, таких как прямая трансформация B лимфоцитов с помощью онкогенной ДНК или трансфекцией вирусом Эпштейна-Барра. Смотрите, например, M. Schrier et. al. (1980); Hammerling et.al. (1981); Kennet et. al. (1980); смотрите также патенты США N 4341761, 4399121, 4427783, 4444887; 4466917; 4472500; 4491632; и 4495890. Панели моноклональных антител, выработанных против эпитопов HCV, могут быть отсортированы по различным свойствам; т.е. по изотипу, сродству к эпитопу и т.д.
Антитела, как моноклональные, так и поликлональные, которые направлены против эпитопов HCV, особенно полезны при диагностике, а те, которые являются нейтрализующими, пригодны для пассивной иммунотерапии. Моноклональные антитела, в частности, могут использоваться, чтобы индуцировать антидиоптилические антитела.
Антиидиотипические антитела являются иммуноглобулинами, которые несут "внутренний образ" антигена инфекционного агента, против которого желательна защита. Смотрите, например, Nisonoff A., et.al. (1981) и Dieesman et.al. (1985). Методики индуцирования антиидиотипических антител известны в этой области. Смотрите, например, Grzych (1985), Mac Namara et.al. (1984) Vytadehaag et. al. (1985). Эти антиидиотипические антитела могут также применяться для лечения, вакцинации и/или диагностики NANBH, так же как и для выявления иммуногенных областей антигенов HCV.
П.Н. Иммуноанализ и диагностические наборы
Как полипептиды, так и антитела настоящего изобретения применимы для иммуноанализов с целью определения присутствия антител к HCV или присутствие вируса и/или HCV полипептидов (или эпитопов), например, в биологических пробах. Разработка иммуноанализов является предметом для большого количества вариаций и в этой области известно много форматов. В иммуноанализе будет использоваться по крайней мере один вирусный эпитоп, полученный из HCV. При одном из осуществлений в иммуноанализе используется комбинация вирусных эпитопов, полученных из HCV. Эти эпитопы могут быть получены из одного и того же или из разных вирусных полипептидов, и могут быть в отдельных рекомбинантных или природных полипептидах или вместе в одних и тех же рекомбинантных полипептидах. В иммуноисследованиях могут использоваться, например, моноклональные антитела, направленные к вирусному(ми) эпитопу (ам), комбинации моноклональных антител, направленных к эпитопам одного вирусного антигена, моноклональные антитела, направленные к эпитопам различных вирусных антигенов, поликлональные антитела, направленные к одному и тому же вирусному антигену или поликлональные антитела, направленные к разным вирусным антигенам. Процедуры могут основываться, например, на конкуренции или прямой реакции, или на определениях сэндвичного типа. В процедурах могут также, например, использоваться твердые подложки или может производиться иммунопреципитация. Большинство исследований включают использование меченых антител или полипептида; метки могут быть, например, ферментными, флюоресцентными, хемилюминесцентными, радиоактивными или молекулами красителя. Исследования, в которых усиливаются сигналы от пробы, также известны; примерами которых являются исследования, в которых используется биотин и авидин, и иммуноисследования с ферментной меткой и опосредственные иммуноисследования, такие как ELISA (описанное ниже).
Обычно иммуноисследование на антитела против HCV будет включать отбор и подготовку испытуемого образца, подозреваемого на содержание антител, такого, как биологическая проба, затем инкубацию его с антигенным (т.е. содержащим эпитоп) полипептидом (ами) HCV в условиях, которые дают возможность образования комплексов антиген-антитело, и затем определения образования таких комплексов. Подходящие условия инкубации хорошо известны в этой области. Иммуноисследование может быть без ограничений в гетерогенной или гомогенной форме и стандартного или конкурентного типа.
В гетерогенной форме полипептид обычно связывается с твердой подложкой, чтобы облегчить отделение образца от полипептида после инкубации. Примерами твердых носителей (подложек), которые могут использоваться, являются нитроцеллюлоза (например, в форме мембраны или микротитровальной ячейки), поливинилхлорид (например, в виде листочков или микротитровальных ячеек), полистирольный латекс (например, в виде гранул или микротитровальных плат), поливинилидинфторид (известный как иммулон® ), диазотированная бумага, нейлоновые мембраны, активированные гранулы и гранулы протеина A. Например, при гетерогенной форме могут использоваться микротитровальные платы Dynatech Immulon® 1 или Immulon® 2, или 0,25 дюймовые полистирольные гранулы (Precisi on Plastic Bale).
Твердая подложка, содержащая антигенный полипептид, обычно отмывается после ее отделения от испытуемого образца и перед определением связанных антител. Известны как стандартные, так и конкурентные формы исследования.
При гомогенной форме испытуемый образец инкубируется с антигеном в растворе. Например, это может происходить при условиях, при которых будут осаждаться какие-либо комплексы антиген-антитело, которые образуются при этом. Известны как стандартная, так и конкурентная формы этих исследований.
При стандартной форме непосредственно определяется количество антител HCV, образующих комплекс антитело-антиген. Это может быть выполнено путем определения, будут ли связываться меченые антиксеногенные (например, античеловеческие) антитела, которые распознают эпитоп анти-HCV антител, благодаря образованию комплекса. При конкурентной форме количество антител к HCV в образце устанавливается путем контроля конкурентного действия на связывание известного количества меченых антител (или других конкурирующих лиганд) в комплекс.
Образованные комплексы, включающие анти-HCV антитела (или в случае конкурентных исследований количество конкурирующих антител), определяются любым из числа известных методов, в зависимости от формы исследования. Например, немеченые антитела в комплексе могут определяться при использовании конъюгата антиксеногенного Ig в комплексе с меткой (например, ферментной меткой).
В иммуноисследованиях, где полипептиды HCV являются анализируемыми веществами, испытуемый образец, обычно биологическая проба, инкубируется с анти-HCV антителами при условиях, которые дают возможность образования антиген-антительных комплексов. Можно использовать различные формы исследования. Например, можно использовать "сэндвичное исследование", когда антитела, связанные с твердой подложкой, инкубируются с испытуемыми образцом: отмываются; инкубируются со вторыми мечеными антителами к анализируемому веществу, и подложка основы отмывается. Анализируемое вещество регистрируется путем определения, связались ли вторые антитела с подложкой. При конкурентной форме, которая может быть или гетерогенной или гомогенной, испытуемый образец обычно инкубируется с антителами и мечеными конкурирующими антигенами или последовательно или одновременно. Эти и другие формы хорошо известны в этой области науки.
Эффективные системы определения инфекции, вызванной HCV, могут включать использование панелей эпитопов, как описано выше. Эпитопы на панели могут быть встроены в один или множество полипептидов. Исследования на различные эпитопы могут быть последовательными или одновременными.
Твердофазный иммуноферментный анализ (ELISA) может использоваться для определения концентраций или антигена, или антитела. Этот метод основан на соединении фермента с антигеном или с антителом, и связанная ферментативная активность используется в качестве количественной метки. Чтобы определить антитела, известный антиген фиксируется на твердой фазе (например, на макроплате или пластиковой чашечке), инкубируется с испытуемыми разведениями сыворотки, отмывается, инкубируется с антииммуноглобулином, меченным ферментом, и снова отмывается. Фременты, пригодные для метки, известным в этой области науки и включают, например, пероксидазу хрена, ферментативная активность, связанная с твердой фазой, определяется путем добавления специфического субстрата и определением образования продукта или использования субстрата колориметрически. Связанная ферментативная активность является прямой функцией количества связанных антител.
Чтобы определить антиген, известное специфическое антитело фиксируется на твердой фазе, и добавляется испытуемый материал, содержащий антиген, после инкубации твердая фаза отмывается и добавляются вторые, меченные ферментом антитела. После отмывания добавляется субстрат и определяется колориметрически ферментативная активность и связывается с концентрацией антигена. Наборы, пригодные для иммунодиагностики и содержащие соответствующие меченые реагенты, создаются путем упаковки соответствующих материалов, включая полипептиды этого изобретения, содержащих эпитопы HCV или антитела, направленные против эпитопов HCV в подходящие контейнеры вместе с остальными реагентами и материалами, необходимыми для проведения этого исследования, так же как и соответствующий набор инструкций.
III. Основные методы
Основные методики, использованные на практике в настоящем изобретении, можно найти, например, в ссылках, цитированных здесь, в частности, EPO Pub, NN 318216 и 388232, так же как и в ссылках в библиографии, которые включены здесь для справки.
IV. Примеры
Описанное ниже является примерами этого изобретения, которые представлены в целях иллюстрации только и не ограничивают объем этого изобретения. В свете настоящего описания специалистам будут очевидны многочисленные примеры осуществления в объеме формулы этого изобретения.
IV. A.I. Синтез перекрывающих пептидов
Полиэтиленовые шпеньки, собранные в блок в порядке 8 х 12 (Coselco, Mimetopes, Victoria, Австралия), подготавливали путем помещения шпеньков в ванну (20% объем/объем пиперидина в диметилформамиде (DMF)) на 30 мин при комнатной температуре. Затем шпеньки удалялись, отмывались в течение 5 мин, затем отмывались в метаноле четыре раза (2 мин/на промывание). Шпенькам давали высохнуть на воздухе в течение по крайней мере 10 мин и затем отмывали последний раз в (5 мин). 1-гидроксибензотриазол (HOBt, 367 мг) растворяли в DMF (80 мл) для использования при соединении Fmос защищенных аминокислот: Fmoc-L-Ala-OPfp, Fmoc-L-Cys(Trt)-OPfp, Fmoc-L-Asp(O-tBu)-OPfp, Fmoc-L-Glu(O-tBu)-OPfp, Fmoc-L-Phe-OPfp, Fmoc-Gly-OPfp, Fmoc-L-His(Boc)-OPfp, Fmoc-L-Ile-OPfp, Fmoc-L-Lys(Boc)-OPfp, Fmoc-L-Leu-OPfp, Fmoc-L-Met-OPfp, Fmoc-L-Asu-OPfp, Fmoc-L-Pro-OPfp, Fmoc-L-Gln-OPfp, Fmoc-L-Arg(Mtr)-OPfp, Fmoc-L-Ser(t-Bu)-ODhbt, Fmoc-L-Thr(t-Bu)-ODhbt, Fmoc-L-Val-OPfp, Fmoc-L-Tyr-OPfp
Защищенные аминокислоты помещали в ячейки микротитровальной платы с HOBt и на плату помещали блок шпеньков, погружая шпеньки в ячейки. Затем комплект запечатывали в пластиковый мешочек и давали прореагировать при 25oC в течение 18 ч, чтобы первые аминокислоты присоединить к шпенькам. Затем блок вынимали и шпеньки отмывали в DMF (2 мин), чтобы очистить и убрать защиту со связанных аминокислот. Процедура повторялась для каждой дополнительной присоединяемой аминокислоты, до тех пор, пока не будут получены все октамеры.
Свободные N-концы затем апетилируют, чтобы компенсировать свободный амид, так как большинство эпитопов находится не на N-конце и таким образом не должны иметь связанный положительный заряд. Ацетилирование выполнялось путем заполнения ячеек микротитровальной платы DMF/ацетангидридом/ триэтиламином (5: 2: 1 об/об/об), давая возможность веществам на шпеньках прореагировать в ячейках в течение 90 мин при 20oC. Шпеньки затем отмывали DMF (2 мин) и MeOH (4 и 2 мин) и высушивали на воздухе в течение по крайней мере 10 мин.
Боковые цепи защищающих групп удаляли путем обработки шпеньков трифторуксусной кислотой/фенолом/дитиоэтаном (95: 2,5:2,5 об/об/об) в полипропиленовых мешочках в течение 4 ч при комнатной температуре. Шпеньки затем промывали в дихлорметане (2 х 2 мин), 5% диизопропалэтиламина/дихлорметане (2 х 5 мин), дихлорметане 5 мин) и высушивали на воздухе в течение по крайней мере 10 мин. Шпеньки затем промывали в воде (2 мин), MeOH (18 ч), высушивали под вакуумом и сохраняли в запечатанных пластиковых мешочках над силикагелем.
IV. A.2. Исследование пептидов
Несущие октамеры шпеньки, полученные как описано выше, сначала обрабатывали ультразвуком в течение 30 мин в буфере для дезинтеграции (1% додецилсульфат натрия, 0,1% 2-меркаптоэтанол, 0,1 М NaH2PO4) при 60oC. Шпеньки затем несколько раз погружали в воду (60oC), с последующим кипячением в MeOH (2 мин), и давали высохнуть на воздухе. Затем шпоньки предварительно покрывали в течение 1 ч при 25oC в микротитровальных ячейках, содержащих 200 мкл блокирующего буфера (1% овальбумина, 1% БСА, 0,1% Твин® и 0,05% NaN3 в PB9) при помешивании. Шпеньки затем погружали в микротитровальные ячейки, содержащие 175 мкл антисыворотки, полученной от больных людей, у которых диагностировано заболевание, вызванное HCV, и инкубировали при 4oC в течение ночи. Проводилось исследование со шпеньками в отношении антисывороток от трех разных больных. Образец ≠≠ PAA 3663-S ("A") продемонстрировал сильную реакцию с HCV по вестерн-блоттингу, HCV конкурентного ELISA, HCV ELISA к клону C100-3 (при разведении 1:1000) и реакции RIBA > 4 + с C100, 5-1-1 т C33с (C22 не делали). (Названия антигена/клона представлено в EPO Pub. N 318216 и 388232, так же как и описанные в литературе, относящейся к иммуноисследованиям по HCV, доступным от Ortho Diagnostics Systemes, Inc). Чистую плазму разводили 1:500 в блокирующем буфере. Образец ≠≠ PAA 33028 ("B") проявил сильную реакцию с HCV по вестерн-блоттингу, HCV конкурентному ELISA, HCV ELISA с клоном C100-3 (при разведении 1:500) и реакции RIBA > 4 + с C100, 5-1-1, C33C и C22. Поликлональную антисыворотку частично очищали путем пропускания через колонку с протеином A и использовали в разведении 1:200 и блокирующем буфере. Образец ≠ PAA S 32931 ("C") проявлял умеренную реакцию с HCV по вестерн-блоттингу (3+), HCV конкретному с клоном C100-3 (в разведении 1: 64), и реакции RIBA 3+ и 4+ с C100 и 5-1-1 соответственно (C33с и C22 не делали). Поликлональную антисыворотку частично очищали путем пропускания через колонку с протеином A и использовали в разведении 1:500 в блокирующем буфере.
Шпеньки отмывали в PBS Твин® 20 (4 х 10 мин) при комнатной температуре, затем инкубировали в микротитровальных ячейках, содержащих меченную пероксидазой хрена козлиную против человеческого Ig антисыворотку (175 мкл, разведение 1:2000 в блокирующем буфере без NaN3) в течение 1 ч при 25oC при помешивании. Античеловеческая антисыворотка специфична легким и тяжелым цепям Ig человека и реагирует с обоими классами, как IgG, так и IgM. Шпеньки опять отмывали в PBS Твин® 20 (4 х 10 мин) при комнатной температуре. Раствор субстрата готовили путем разведения NaH2PO4 (1 М, 200 мл) и лимонной кислоты (1 М, 160 мл) до 2 л дистиллированной водой, доводя pH до 4,0. Непосредственно перед использованием к 100 мл буфера добавляли азино-диэтилбенативаодинсульфонат (ABTS 50 мг) и перекись водорода (0,3 мкл/мл), чтобы завершить получение субстратного раствора. Раствор субстрата (150 мкл) добавляли в каждую ячейку микротитровальной платы и шпеньки погружали в ячейки и инкубировали при 25oC в темноте. После развития окраски реакции останавливаются путем удаления шпеньков и снимали показатели поглощения растворов при 405 нм.
Октамеры, перечисленные в конце описания, были иммунореактивны с анти-HCV антисыворотками. Пептиды, реагирующие со всеми тремя антисыворотками, перечисляются как эпитопы, тогда как пептиды, реагирующие только с одной или двумя антисыворотками, перечислены как слабые эпитопы (показаны " ~ "). Особенно сильные эпитопы помечены буквами, а не номерами (например, EpAA).
IV. В. Дифференциальное исследование
Следующее исследование выполнялось, чтобы различить ранние антигены от поздних антигенов. Антитела к ранним антигенам могут быть определены и использованы для диагноза инфекции, вызванной HCV, более быстро.
Были получены серийные образцы крови от больного человека, имевшего повышенные показатели ALT, но отрицательные на анти-C100-3 антитела. Пять образцов крови, полученные перед завершением сероконверсией (C100-3 положительной), объединяли и использовали в исследовании в разведении 1:2000. Исследование проводилось как описано выше в разделе IV, A. Однако один двойной набор шпеньков инкубировали с козлиной против человеческого IgG специфической антисывороткой, меченной пероксидазой хрена, в то время как другой набор инкубировался с козлиной против человеческого IgM специфической антисывороткой, меченной пероксидазой - хрена. Эпитопы, иммунореактивные с антителами IgM, являются ранними эпитопами.
Результаты показали, что большинство ранних эпитопов находятся в области, располагающейся от примерно аминокислоты 480 до примерно аминокислоты 650. Особенно сильными эпитопами и IgM были октамеры, начиная с аминокислот номер 506, 510, 523, 553, 562, 580 и область от 590 до 620. Исследования, при которых используются антигены, несущие эпитопы из этой области, дадут возможность диагностики инфекции, вызванной HCV, на более ранней стадии, чем исследования с использованием других антигенов.
Мы дополнительно исследовали серийные образцы плазмы, полученные от пяти больных гепатитом NANB в результате переливания крови с последующим изучением в течение 3-12 лет. Сроки получения первоначальных образцов крови были менее чем с интервалом в одну неделю. Каждый образец испытывался на IgG и IgM против ядерного антигена (G22), двух оболочечных антигенов (E1 и E2), и трех антигенов неструктурной области (С33c, C100 и N 5) с помощью иммуноферментативного анализа. Мы обнаружили, что IgM ответ на C22 и C33c превышал IgG ответ на эти антигены.
NS - 5 также вызывал IgM ответ, но этот ответ не превышал IgM ответ на этот антиген. Таким образом, можно разработать анализы, позволяющие определить очень ранние стадии инфекции, путем использования эпитопов, полученных из областей C22 и C33c и определенных по связыванию с IgM . Антитела к области C33c присутствовали в течение наибольшего периода времени и это наводит на мысль, что наиболее надежными должны быть диагностические исследования, направленные на C33c.
IV. C. Варианты последовательностей в изолятах HCY от разных индивидуумов.
У отдельных людей, часть из которых были серологически положительны на анти- C100-3 антитела (EC10 был отрицательным на антитела), были определены изоляты HCV, которые содержат последовательности с отклонениями от CDC/HСV/. Идентификация этих новых изолятов выполнялась путем клонирования и секвенирования сегментов генома HCV, которые усиливались по методу PCR с использованием последовательностей CDC/HCl. При этом методе используются праймеры и пробы на основе кДНК последовательностей HCV, описанных здесь. Первой стадией в этом методе является синтез кДНК к или геному HCY или его репликационному посреднику с использованием обратной транскриптазы. После синтеза кДНК HCV и перед амплификацией РНК в образце расщепляется известными в этой области методами. Указанный сегмент кДНК HCV затем амплифицируется путем использования соответствующих праймеров. Амплифицированные последовательности клонируются, и клоны, содержащие амплифицированные последовательности, определяются с помощью пробы, которая комплементарна последовательности, лежащей между праймерами, но которая не перекрывает праймеры.
IV. C.I. Изоляты HCV, выделенные от людей в США.
Образцы крови, которые использовались в качестве источника вирионов HCV, получали из Американского красного креста в Шарлотте, Северная Каролина, и из Общественного центра переливания крови Канзаса. Канзас сити, Миссури. Образцы отбирали на антитела к антигену C100-3 HCV, используя исследование по ELISA, и подвергали дополнительному вестерн-блот анализу с использованием поликлональных козлиных противочеловечьих HRP-антител, чтобы количественно определить анти- HCV антитела. Два образца ≠≠ 23 и ≠≠ 27 из Американского Красного Креста и из Общественного центра переливания крови Канзаса соответственно, как было определено, являлись положительными на HCV по этим исследованиям.
Вирусные частицы, присутствующие в сыворотке этих образцов, выделялись с помощью ультрацентрифугирования при условиях, описанных Bradley et.al. (1985). РНК экстрагировали из частиц путем расщепления с помощью протеиназы К и додецилсульфатом натрия при конечных концентрациях протеиназы К 10 мкг/мл и додецилсульфата натрия 0,1%; расщепление проходило в течение 1 ч при 37oC. Вирусную РНК дальше очищали путем экстракции хлороформ-фенолом.
РНК HCV в препарате РНК подвергалась обратному считыванию на кДНК. После того, как синтезировались обе нити кДНК, полученная в результате кДНК, затем амплифицировалась методом PCR. Для кДНК HCV из трех клонов, полученной из каждого изолята HCY, выполняли анализ последовательности. Анализ в основном выполнялся по методу, описанному у Chen и Seeburg (1985).
Консенсусные последовательности клонов, полученных из HCV в образцах 23 и 27, показаны на фиг. 3 и фиг. 4 соответственно. Вариабельные последовательности также показаны на этих фигурах, так же как и аминокислоты, кодируемые в консенсусных последовательностях.
Фигуры 5 и 6 показывают сравнение вытянутых в линию нуклеотидных последовательностей положительных цепей (фиг. 5) предполагаемых аминокислотных последовательностей (фиг. 6) образцов 23, 27 и HCV 1. Аминокислотная последовательность HCV1 на фиг. 6 представляет аминокислоты с номерами 129-467 полипротеина HCV, кодируемого большим ORF в геномной РНК HCV. Изучение фиг. 5 и 6 показывает, что в последовательностях трех изолированных клонов существуют вариации. Варианты последовательностей на нуклеотидном уровне и на аминокислотном уровне суммированы в таблице непосредственно ниже. В табл. 1 полипептиды, обозначенные S и NSI, представляют аминокислоты номеров с 130 до~ 380, и c 380 до ~ 470 соответственно, так как эти домены были ранее ранее известны. Отсчет ведется от предполагаемого инициатора метионина. Терминология S и NSI основывается на определении местоположения последовательностей, кодирующих полипептиды, используя модель флавивируса. Как обсуждалось выше, однако, недавние данные наводят на мысль, что не существует полной корреляции между и флавивирусами относительно вирусных полипептидных доменов, в частности в предполагаемых доменах E/NSI. Конечно, полипептиды и колирующие их домены могут демонстрировать существенные отклонения от флавивирусной модели.
Хотя существуют варианты вновь выделенных последовательностей HCV, клонированные последовательности из образцов 23 и 27 (названных HCV 23 и HCV 27) каждая содержит 1019 нуклеотидов, что указывает на отсутствие делеции и дополнительных мутаций в этой области в выбранных клонах. Последовательности на фигурах 5 и 6 также показывают, что выделенные последовательности не перегруппированы в этой области.
Сравнение консенсусных последовательностей для HCVI и для других изолятов HCV суммируется в таблице, выше. Различия в последовательностях между изолятом от шимпанзе HCV1 и HCV, выделенных от людей примерно такие же, что и наблюдаемые между HCV-ми человеческого происхождения.
Интересно, что различия в последовательностях в двух из предполагаемых доменов не одинаковы. Последовательность в предполагаемой области, по-видимому, является относительно постоянной и случайно распределена по этой области. Напротив, предполагаемая область NSI имеет более высокую ступень вариабельности, чем вся последовательность, и изменение, по-видимому, является гипервариабельным карманом из примерно 28 аминокислот, который располагается примерно на 70 аминокислот в прямом направлении от предполагаемого N-конца предполагаемого полипропилена.
Хотя можно доказать, что определяемые изменения были введены во время процесса амплификации, маловероятна, что все изменения произошли в результате этого. Было установлено, что Tag полимеризата вводит ошибки в последовательность примерно в одно основание на 10 килооснований ДНК матрицы на цикл (Saiki et.al. (1988). На основании этой оценки до 7 ошибок могло быть введено во время амплификации с помощью PCR фрагмента ДНК из 1019 пар оснований. Однако три субклона HCV-23 и HCV-27 дали 29 и 14 изменений оснований соответственно. Следующее наводит на мысль, что эти изменения встречаются в природе. Примерно 60% изменений оснований являются молчащими мутациями, которые не изменяют аминокислотной последовательности. Изменения, вводимые Ta; полимеразой во время амплификации путем PCR, как ожидалось бы, встречаются хаотично; однако результаты показывают, что отличающиеся последовательности собираются по крайней мере в одной специфической области.
IV.C. 2.Изоляты HCV от людей из Италии и США
Сегменты РНК HCV, присутствующие в различных изолятах, амплифицировали по методу HCV/cPCP. Эти сегменты соединяют область от 0,6 К до 1,6 К в прямом направлении от метионина, кодирующего стартовый кодон предположительного полипропилена HCV. Изоляты происходят из биологических образцов, полученных от индивидуумов, инфицированных HCV. Более конкретно, изолят HCT ≠≠ 18 происходит из человеческой плазмы от индивидуума в США, ECI и EC10 получены из биопсии печени итальянского больного и Th получен из периферической крови, мононуклеоцитной фракции, американского больного. Сравниваемые сегменты РНК HCV были выделены от шимпанзе.
РНК экстрагировали из образцов человеческой плазмы, используя экстракцию фенолом: CHCl3 : изоамиловым спиртом. Или 0,1 мл или 0,01 мл плазмы разводили до конечного объема 1,0 мл раствором TE NB/ протеиназы К/додецилсульфата натрия (0,05 М Трис-HCl, pH 8,0, 0,001 М ЭДТА, 0,1 М NaCl, 1 мг/мл протеиназы К и 0,5% додецилсульфата натрия), содержащим от 10 до 40 мкг/мл полиадениловой кислоты, и инкубировали при 37oC в течение 60 мин. После этого расщепления протеиназой К полученные в результате фракции плазмы освобождали от белков путем экстракции TE (50 нМ Трис-HCl, pH 8,0, 1 мМ ЭДТА) насыщенным фенолом, pH 6; 5. Фенольная фаза отделялась путем центрифугирования и повторно экстрагировалась TENB, содержащим 0,1% додецилсульфата натрия. Полученные в результате водные фазы из каждой экстракции объединялись и дважды экстрагировались равным объемом фенола/хлороформа/изоамилового спирта (1:1(99: 1)) и затем дважды объемом 99:1 смеси хлороформа/изоамилового спирта. После разделения фаз с помощью центрифугирования водную фазу доводили до конечной концентрации 0,2 М ацетатом Na, и нуклеиновые кислоты осаждали путем добавления двух объемов этанола. Осажденные нуклеиновые кислоты выделяли путем ультрацентрифугирования и SW 41 роторе при 38 К в течение 60 мин при 4oC или в микроцентрифуге в течение 10 мин при 10 К, 4oC.
РНК, извлеченная из биопсии печени, была получена доктором F. Bonino, Ospedale Maggiore di Giovanni Battista Турин, Италия. Мононуклеоцитная фракция получалась путем осаждения аликвотных проб крови от индивидуума при помощи Ficoll-Paque® (Pharmacia Corp.), при использовании указаний производителя. Всю РНК экстрагировали из фракции, используя гуанидин-тиоцианатную процедуру, описанную у Choo et. al. (1989).
Синтез кДНК NCV из образцов выполнялся с использованием обратной транскриптазы. После преципитации этанолом осажденная РНК или фракция нуклеиновых кислот высушивалась и ресуспендировалась в DEPC, обработанном дистиллированной водой. Вторичные структуры в нуклеиновых кислотах разрушали путем нагревания при 65oC в течение 10 мин, и образцы немедленно охлаждали на льду, кДНК синтезировали, используя 1 до 3 мкг общей РНК из печени или из нуклеиновых кислот (или РНК), извлеченной из 10 до 100 мкл плазмы. При синтезе использовали обратную транскриптазу и реакцию проводили в объеме 25 мкл, используя протокол, предоставляемый производителем, BRL. Все реакционные смеси для синтеза кДНК содержали 23 единицы ингибитора рНКазы, Rnasin® (Fischer Promepa).
После синтеза кДНК реакционные смеси разбавляли водой, кипятили в течение 10 мин и быстро охлаждали на льду.
Каждый выбор образцов подвергали двум циклам амплификации с помощью PCP. Праймеры для реакций выбирали, чтобы амплифицировать области, обозначенные "EnvL" и "En vR". Область "EnvL" заключает нуклеотиды 669-1243, и предполагаемые аминокислоты от 117 до 308; область "EnvR" включают нуклеотиды 1215-1629 и кодирует предполагаемые аминокислоты 300-408 (предполагаемые аминокислоты пронумерованы, начиная от предполагаемого метионинового кодона инициации).
Реакция РСР проводили по существу по указаниям производителя (Cetus-Perkin-Elmer), за исключением добавления 1 мкг РНКазы А. Реакции проводились в конечном объеме 100 мкл, выполняли в количестве 30 циклов, используя режим 94oC (1 мин), 37oC (2 мин), и 72oC (3 мин), с развитием в течение 7 мин при 72oC для последнего цикла. Образцы затем экстрагировали фенолом; CHCl2, осаждали этанолом два раза, ресуспендировали в 10 мМ Трис HCl, pH 8,0 и концентрировали, используя фильтрацию в Centricon - 30 (Amicon). Эта процедура эффективно удаляет олигонуклеотиды, меньшие по размеру, чем из 30 нуклеотидов; таким образом, праймеры из первого цикла амплификации путем PCR удаляются.
Образцы, сконцентрирвоанные в Centricon-30, затем подвергали второму циклу амплификации при помощи PCR. Амплификация путем PCR осуществлялась за 35 циклов, с использованием режима 94oC (1 мин), 60oC ( 1 мин) и 72oC (2 мин) с развитием в течение 7 мин при 72oC для последнего цикла. Образцы затем экстрагировали фенолом: CHCl, осаждали два раза и расщепляли ε cp RI. Продукты реакции PCR анализировали путем разделения продуктов путем электрофореза в 6% полиакриламидном геле. ДНК с примерно установленным размером ожидаемого продукта PCR электроиллюировали из геля и субклонировали или в плазмидный вектор pGEM-4 или в λ gt 11. После первого цикла амплификации ожидаются продукты с размерами для EnvL и EnvR 615 bp и 683 bp соответственно; после второго цикла амплификации ожидаются продукты с размерами для EnvL и EnvR 414 bp и 575 bp соответственно. Плазмиды, содержащие амплифицированные продукты, использовали для трансформации клеток-хозяев; плазмида pGEM-4 использовалась для трансформации DH5-альфа, и λ gt 11 использовалась для трансформации C600 дельта-HFL. Клоны трансформированных клеток, которые или гибридизуются с соответствующими пробами HCV или те, которые имели вставки правильного размера, отбирали. Вставки затем клонировали в М13 и секвенировали. Пробы для всех продуктов HCY/cPCP состояли из меченых 32P секций кДНК HCV, которые были получены путем амплификации с помощью РСР.
Информация о последовательности по вариантам в области En получены из 3 клонов из HCT ≠≠ 18, двух клонов из ТН, 3 клонов из ECI и из клонов HCV1. Сравнение составной нуклеотидной последовательности каждого изолята, полученного из этих клонов, показано на фиг. 7. На фигуре каждая последовательность показана от 5' до 3' для смысловой цепи EnvL области, и последовательности были вытянуты в линию. Вертикальные линии и заглавные буквы показывают гомологию последовательности, отсутствие линии и незаглавная буква указывает отсутствие гомологии. Последовательности, показанные в линиях, являются следующими: линии I, Thorn; линия 2, EC1; линия 3, HCT ≠≠ 18; линия 4 HCV 1.
Информация о последовательности по вариантам в EnVR области получена из двух клонов EC10 и из клонов HCV1. Два клона ЕC10 отличались только одним нуклеотидом. Сравнение нуклеотидных последовательностей EC10 (клон 2) и составных последовательностей HCV показано на фиг. 8; каждая последовательность показана от 5' до 3' для смысловой цепи EnvR области, и последовательности были вытянуты в линию. Двойные точки между последовательностями были вытянуты в линию. Двойные точки между последовательностями показывают гомологию последовательности.
Сравнение аминокислотных последовательностей, кодируемых в EnvL (аминокислоты 117-308) и EnvR области (аминокислоты ≠≠ 300-438) для каждого из изолятов, показано на фиг. 9 и фиг. 10 соответственно. В фигуры включены последовательности для изолятов JH 23 и JH 27, описанных выше. Также указаны последовательности из японского изолята; эти последовательности предоставлены доктором T. Miyamura, Япония. На фигурах аминокислотная последовательность для этой области дана полностью для HCV1, и негомологичные аминокислоты указаны в разных изолятах.
Как видно на фиг. 9, в EnvL области существует в целом примерно 93% гомологии между HCV1и другими изолятами. HCT18, Th и EC1 имеют примерно 97% гомологии с HCV1; JH 23 и JH 27 имеют примерно 96% и примерно 95% гомологии соответственно с HCV1. Фиг. 10 показывает, что гомология в EnvR области меньше, чем в EnvL области; кроме того, одна подобласть, по-видимому, является гипервариабельной (т. е. из аминокислот 383-405). Эти данные суммированы в табл. 2.
IV. Промышленная пригодность
Эпитопы, идентифицированные здесь, могут использоваться для получения полипептидных продуктов, как описано выше, для применения такого, как анализ образцов крови на инфекцию, вызванную HCV, клиническая диагностика HCV, образование антител, получение лекарственных препаратов. Другое применение описано выше и другие способы использования будут легко видны специалистам.


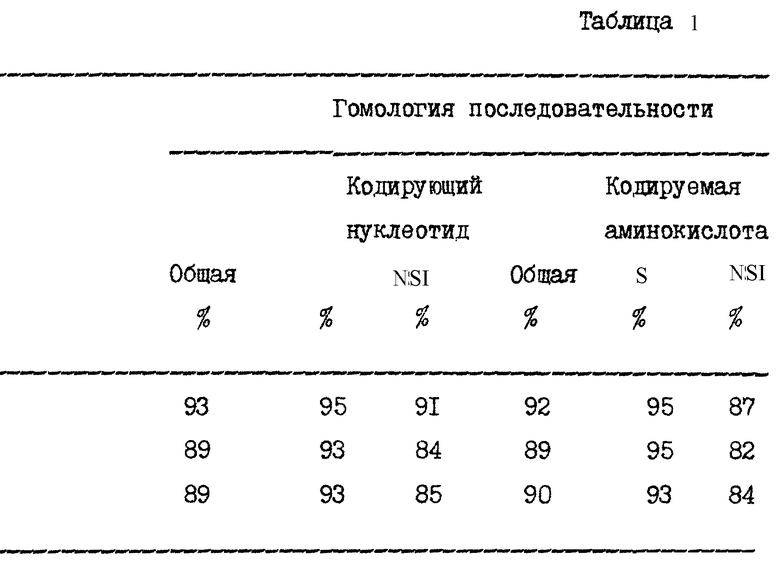




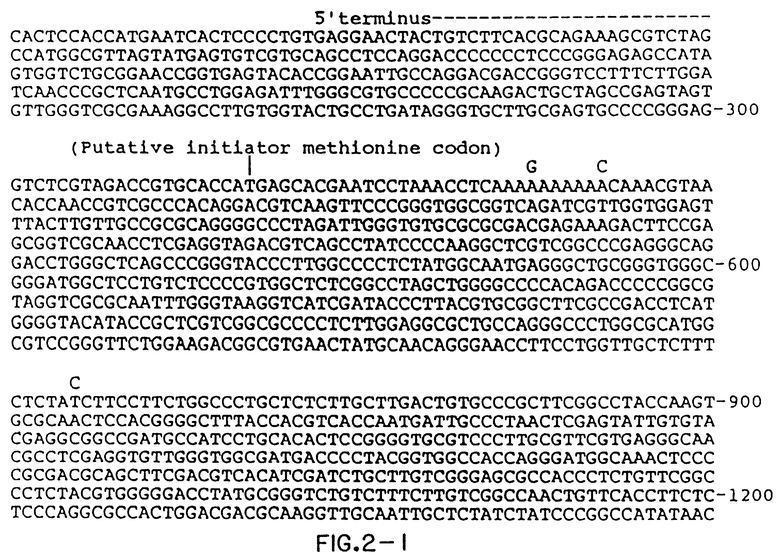














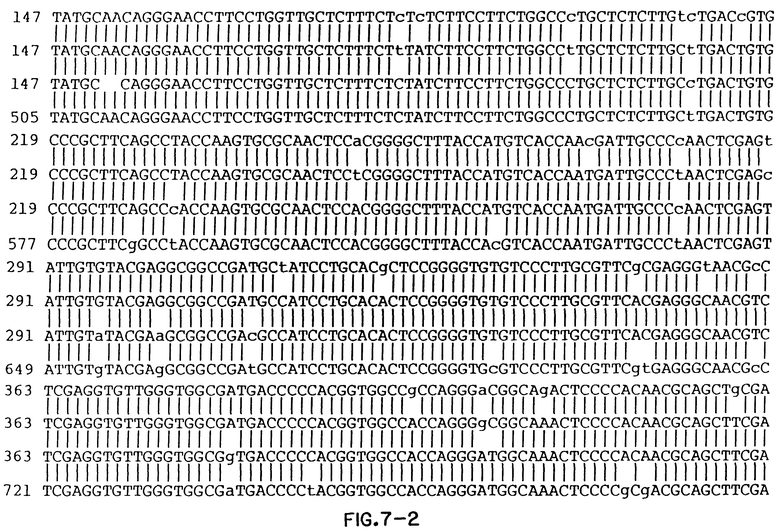
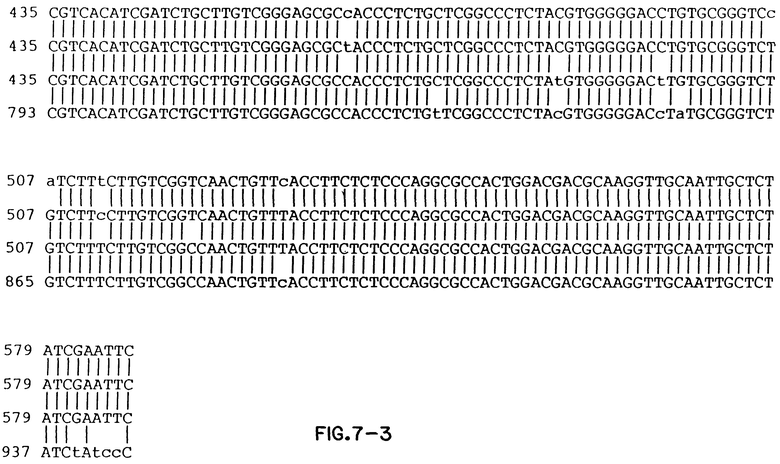



Изобретение относится к полипептидам, применяемым в качестве иммунологических реагентов при определении, предупреждении и лечении инфекций, вызванных HCV. Описывается новый полипептид, включающий усеченную последовательность HCV (вируса гепатита С), содержащую эпитоп HCV с формулой аах-аау, способный индуцировать иммунный ответ у субъекта, где аа обозначает аминокислоту, х и у являются целыми числами, такими, что у-х ≥ 14, аах-аау обозначает часть аминокислотной последовательности с фиг. 1 и х выбирают из группы: 66, 413, 465, 540, 1218, 1940, 2244, 2281, причем указанный полипептид может иметь необязательную последовательность N-концевых аминокислот, соответствующую последовательности полипептида HCV, прилегающей к N-концу аминокислоты аах, к которой она прикреплена, или необязательную последовательность С-концевых аминокислот, соответствующую последовательности полипептида HCV, прилегающую к С-концу аминокислоты аау, которой она прикреплена, причем общее количество дополнительных аминокислот не превышает 15 для аа413-аа-аа427, 4 для аа465-аа480, 85 для аа540-аа554 10 для аа2244-аа2258, и 30 для аа2281-аа2300. Описывается также способ определения присутствия антител, способ индукции иммунного ответа. 5 с. и 1 з.п. ф-лы, 2 табл., 10 ил.
aax - aay,
способный индуцировать иммунный ответ у субъекта, где aa обозначает аминокислоту, x и y являются целыми числами, такими, что y - x ≥ 14, aax - aay обозначает часть аминокислотной последовательности с рисунка 1 и x выбирают из группы: 66, 413, 465, 540, 1218, 1940, 2244, 2281, причем указанный полипептид может иметь необязательную последовательность N-концевых аминокислот, соответствующую последовательности полипептида HCV, прилегающей к N-концу аминокислоты aax, к которой она прикреплена, или необязательную последовательность C-концевых аминокислот, соответствующую последовательности полипептида HCV, прилегающую к C-концу аминокислоты aay, в которой она прикреплена, причем общее количество дополнительных аминокислот не превышает 15 для aa413 - aa427, 4 для aa465 - aa480, 85 для aa540 - aa554, 10 для aa2244 - aa2258 и 30 для aa2281 - aa2300.
| Способ твердофазного синтеза пептидов | 1975 |
|
SU539874A1 |
| Способ получения пептидов | 1988 |
|
SU1598881A3 |
| СПОСОБ СТАБИЛИЗАЦИИ ГАЛОИДСОДЕРЖАЩИХ УГЛЕВОДОРОДОВ | 0 |
|
SU318216A1 |
| ДАТЧИК ФАЗЫ | 0 |
|
SU388232A1 |
| EP 0419182 A, 27.03.1991 | |||
| CHARD T., An Introduction to radioimmunoas | |||
| say and related techniques, Elsevier North Holland Biochemical Press, 1978, p.30 | |||
| Остерманн Л.А | |||
| Исследования биологических макромолекул электрофокусированием, иммунноэлектрофорезом и радиоизотопными методами | |||
| - М.: Наука, 1983, с.99 - 105. | |||

