Изобретение относится к борьбе с насекомыми-вредителями. В частности, изобретение относится к защите растений от нематод-паразитов.
Большинство растительных нематод-паразитов имеет длину менее 2 мм и развиваются из яиц, проходя три или четыре ювенильные стадии стадии (J1-J3 или J4) прежде чем достигнуть имагинальной стадии; при этом, продолжительность жизненного цикла нематод составляет от нескольких недель до нескольких месяцев. В природе существуют эктопаразиты и эндопаразиты, причем, большинство видов поражает корни растения. За очень небольшими исключениями, нематоды, для проникновения через клеточные стенки растения, используют полный стилет, с помощью которого они отсасывают содержимое клетки. Миграция нематод в растения происходит путем проникновения вовнутрь клетки через перфорированные клеточные стенки, либо путем продвижения между клетками. Некоторые эндопаразиты проникают в растения на небольшие расстояния до стадии питания, тогда как другие продвигаются постепенно вглубь растения, либо оседают на месте в расчете на последующий рост растения-хозяина, обеспечивая тем самым свое распространение внутри растения.
Некоторые виды нематод, включая такие экономические важные виды, как корневая и яванская галловая нематоды, вызывают значительные изменения в клетках растений, способствуя тем самым поддержанию сердиментарных (т.е. прикрепленных) самок. Эти особи растут примерно в течение нескольких недель, достигая высокой степени плодовитости (1000x).
Галловые нематоды (в основном, Heterodera и Globodera spp) являются главными вредителями многих сельскохозяйственных культур. Heterodera glycines являются главным патогеном сои культурной, возделываемой в США с экономическим эффектом, достигающим примерно US $ 500-1000 М в год. Heterodera shachti (свекловичная нематода) является главным врагом фермеров, возделывающих сахарную свеклу в европейских странах и в некоторых регионах США, а Heterodera avenae (злаковая нематода) поражает почти все ценные зерновые культуры, произрастающие по большей части на почвах аридной зоны, например, в некоторых регионах Австралии. Картофельные нематоды Globodera rostochiensis и G. pallida распространены на многих площадях, отведенных под возделывание картофеля. Причиняемые этими вредителями убытки сельскому хозяйству Великобритании как непосредственным разрушающим воздействием, так и опосредованным влиянием на урожай составляет Z 10 - 50 М в год.
Яванская галловая нематода (Meloidagyne spp) обитает в тропических и субтропических почвах, но имеется несколько видов патогенов, принадлежащих к этому классу нематод, которые распространены во всем мире. Таких видов достаточно много, однако, за ущерб, наносимый культурным растениям, ответственны лишь пять видов нематод; причем убыток, наносимый сельскому хозяйству паразитом вида M.incognita, составляет примерно 66% от всех экономических потерь, вызываемых этими патогенами. Для широкого ряда культурных растений в главных географических регионах тропиков, общие потери в урожаях культурных растений составляют 11 - 25%.
В настоящее время для борьбы с вредителями используются сельскохозяйственные меры борьбы с вредителями, химическая борьба, а также борьба с вредителями путем замены поражаемых растений резистентными, причем часто эти меры используются в сочетании друг с другом. Однако, проблема разработки усовершенствованного способа борьбы с нематодами в настоящее время особенно актуальна, поскольку большинство имеющихся нематоцидов, в основном, приемлемы для широкого использования.
Например, карбамат (алдикарб) и продукты его распада являются высоко токсичными для млекопитающих, загрязняют грунтовые воды в США, и вероятно, в других регионах, где широко используется этот пестицид. Сельскохозяйственные меры борьбы с вредителями несут в себе скрытые потери, что неприемлемо для фермеров, специализирующихся на выращивании определенных культур, либо высокорентабельных культур. Резистентные сорта также являются не всегда приемлемыми, и часто наиболее восприимчивые к поражениям патогенами сорта превосходят по своей урожайности резистентные сорта, а поэтому использование резистентных сортов опять же несет в себе скрытые потери, даже если растения не поражаются нематодами. Неадекватность вышеуказанных способов борьбы с вредителями вынуждает исследователей продолжать поиск более эффективных средств борьбы с нематодами.
Галловые нематоды с обычной плотностью заселения вызывают замедление роста растений с корневой системой, занимающей небольшой объем почвы, делая их чахлыми и низкорослыми. Пораженные растения обнаруживают симптомы минеральной недостаточности в листьях и легко увядают. Потери в урожаях зависят от тяжести поражения, т.е. степени превышения его пределов толерантности, и могут составлять для некоторых видов свыше 50%.
Галловая яванская нематода, помимо основных симптомов, описанных для галловых нематод, вызывает часто значительное галлообразование, сопровождающееся увеличением восприимчивости растения к вторичным патогенам.
Некоторые нематоды являются переносчиками небольшого ряда растительных вирусов (например, вирусы NEPO, переносимые Xiphinema, Longidorus, и некоторые вирусы TOAPA, переносимые Trichodorus). Например, Xiphinema spp переносит вирус GFLV на виноград. Кроме того, нематоды вместе с грибками переносятся определенными насекомыми-переносчиками и вызывают некоторые серьезные болезни растений, такие, как вертициллез сосны и нематодоз кокосовой пальмы. В некоторых районах Австралии нематода переносит Corynebаcterium на семенную шапку плевела, в результате чего он становится в высокой степени токсичным для пасущихся овец. Такая связь с бактериями и грибками, в частности, с Fusarium spp. сообщает Meloidogyne spp особый экономический статус. Благоприятное клубнеобразование бобовых культур Rhizobium spp может также подавляться соевой и гороховой нематодами.
Совершенно очевидно, что получение культурных растений, резистентных к нематодам, является важной задачей для исследователей. В отношении нематод, резистентность определяется успехом или неудачей в репродуцировании на данном виде растений-хозяев. За доминантный, полудоминантный и рецессивный механизм исследования ответственны от 1 до 3 растительных гена. Гипотеза "ген для гена" применима для некоторых случаев, когда типично доминантному R-гену резистентности противодействует рецессивный V-ген вирулентности нематод. Два примера резистентности, вводимой селекционерами, заключается в следующем.
Что касается вида Globodera, то эту популяцию картофельных нематод, распространенных в Европе, можно разделить на 5 форм G.rostochensis (RO 1-5) и три формы G. pallida (Ra 1-3). Эти типы патогенов определяются как формы одного вида, которые отличаются своей репродуктивной способностью на растениях-хозяевах, которые, как известно, экспрессируют гены резистентности. HI - ген, несущий резистентность к Globodera rostochiensis Ro 1 и Ro 4, является достаточно качественным и находит широкое коммерческое использование. В Великобритании, cv. Mаris Piper экспрессирует HI и является высококачественным резистентным сортом. К сожалению, широкое использование в Англии этого сорта коррелирует с возрастающим преобладанием по всей стране G.pallida, к которой этот сорт является полностью восприимчивым.
Что касается Meloidogyne spp., то этот вид имеет морфологически аналогичные формы с различной способностью размножаться на хозяйских растениях. Для дифференциации 4 видов M.incognita использовали стандартные растения табака (cv NC95) и хлопчатника (cv Deltaphne), два вида M.arenaria дифференцировали с использованием арахиса (cv Florrunner). Один единственный доминантный ген в табаке cv NC95 сообщал резистентность к видам 1 и 3 M.incognita, но возделывание этого сорта в США повышало превалирование других галловых нематод, в частности M.arenaria. Большинство источников резистентности были эффективными лишь для одного какого-либо вида галловых нематод, за одним лишь исключением, а именно: ген LMi от Lycopersicum peruvanium, сообщал резистентность ко многим видам нематод, кроме M.halpa. Другим ограничением генов резистентности, идентифицированных в томатах, бобах и сладком картофеле, является их температурная зависимость, которая делает их неэффективными в тех местах, где температура почвы превышает 28oC.
В соответствии с вышеуказанным, необходимость получения более резистентных к нематодам коммерчески ценных культур становится более очевидной. Новая резистентность, сообщаемая растению, должна быть более продолжительной, поскольку нематоды не имеют большого потомства за сезон культивирования, и важные изменения патотипов происходят в результате селекции существующих до этого на полях форм, а не в результате мутации после внедрения сорта. Этот последний процесс не так легко осуществляется для нематод. Например, в противоположность широко распространенной устойчивости к инсектицидам у тлей и других насекомых, полевой устойчивости к нематодам не наблюдается.
Было обнаружено, что ингибиторы протеиназы, включая ингибиторы трипсина, являются сильными противонематодными агентами, а поэтому, обработка нематод и/или очагов их обитания ингибиторами протеиназы может служить защитой растений от поражения нематодами; например, растения, трансформированные геном или другой ДНК, кодирующей ингибитор трипсина, являются резистентными к нематодам.
В первом своем варианте, настоящее изобретение относится к способу борьбы с паразитами растений - нематодами, который заключается в обработке нематод или мест их обитания ингибитором протеиназы. Для того чтобы нематоды могли поглощать ингибитор, он должен соответствовать данной протеиназе.
Во втором своем варианте, настоящее изобретение относится к использованию ингибитора протеиназы в качестве средства против растительных нематод.
Для генерирования ингибитора протеиназы в местах обитания нематод, предпочтительно использовать технику рекомбинантных ДНК. Подходящим способом достижения вышеуказанной цели является генетическая трансформация растения, при этом, предпочтительно, чтобы растение, которое является восприимчивым к поражению нематодами, само экспрессировало ингибитор протеиназы.
В третьем своем варианте, настоящее изобретение относится к способу сообщения растению резистентности к нематодам, который заключается в модифицировании или трансформации растения в целях сообщения этому растению способности экспрессировать ингибитор протеиназы. При этом, следует отметить, что сообщаемая растению резистентность не обязательно должна быть полной, однако, она должна быть достаточной для получения нужного агрономического или экономического эффекта. Борьба с нематодами может быть ограничена борьбой с самками нематод, как будет показано ниже.
Подробное описание жизненного цикла и патогенности картофельных нематод приводится в специальной литературе (Agrios I.N. (1988) "Plant Pathology", 3rd. Ed. Academic Press San Diego, стр. 803 и Jones F.I.W. и Jones M. (1974) "Pests of Field Crops", 3rd. Ed. Arnold, London). На второй личиночной стадии J2, картофельная нематода Globodera pallida мигрирует с поверхности корня вовнутрь клетки через клеточную стенку, протыкая ее своим стоматостилетом. После того, как организм достигнет эндодермы, он выбирает исходную клетку для питания, внедряется в нее, после чего клетка увеличивается в объеме, вовлекая другие клетки в синцитий, которой постепенно принимает характер системы переноса клеток примерно через 7 дней после инвазии. Эти организмы питаются из синцития, который поддерживается на протяжении всего периода питания нематоды. Животные, которые становятся самцами, питаются в течение двух стадий (J2 и J3), а животные - самки, кроме того, питаются в стадии (J4) и во взрослой стадии. Размер тела самки в 100 раз превышает размеры тела самцов, и от питания самок в значительной степени зависит патология галловых нематод. Сорт Maris Piper обладает резистентностью к Globodera rostochiensis (лишь патогены типа Ro 1 и Ro 4), и эта резистентность выражается неспособностью синцития развиваться после первых нескольких дней взаимодействия нематоды с клеткой. Самцы могут развиваться на таких растениях, а самки не способны развиваться на этих растениях, вследствие чего патогенность нематод значительно снижается. Поэтому целью сообщаемой новой резистентности является либо предупреждение, либо ограничение развития самок нематод.
В своем четвертом варианте, настоящее изобретение относится к использованию гена или другой ДНК, кодирующей ингибитор протеиназы, для получения трансгенного растения, обладающего резистентностью к нематодам.
Субстратная специфичность протеиназ, происходящих от различных источников, может варьироваться, критериев ее оценки не существует. Эти ферменты действуют посредством четырех различных каталитических механизмов, что позволяет их отнести к четырем различным классам, а именно: сериновые, цестеиновые, аспарагиновые и металлопротеиназы. Сериновые протеиназы представляют широко распространенный класс, отличающийся большим разнообразием; цистеиновые протеиназы также являются широко распространенным классом и обнаруживаются в бактериях, эукариотических микроорганизмах, растениях и животных. Металлопротеиназы также широко распространены, однако, класс аспарагиновых протеиназ является более ограниченным и обнаруживается, в основном, в эукариотах.
Существует широкий ряд ингибиторов протеиназ, многие из которых являются специфичными к одному классу протеиназ. Например, 1, 10 - фенантролин является хелатообразующим агентом, действующим как обычный ингибитор металлопротеиназы, не удаляя при этом кальций, который необходим для активности многих ферментов, включая некоторые другие протеиназы. Особый интерес представляют также ингибиторы протеиназы, которые будучи протеинами, должны быть субстратами, а не ингибиторами этих ферментов. Многие из этих ингибиторов являются запасными белками семян, которые аккумулируются в процессе развития семян, и могут представлять один из наиболее обильных белков в зрелых семенах. В настоящее время известными семействами ингибиторов протеиназы (PI) являются: семейства Bowman-Birk Kunitz, картофеля 1, картофеля 2, тыквы, подсемейство зерновых, семейства Ragi 1 - 2, кукурузы 22 кДа и семейство цистатина. Предпочтительным ингибитором протеиназы, используемым в настоящем изобретении, является семейство Bowman-Birk; другие подходящие ингибиторы принадлежат к семействам кукурузы 22 кДа и цистатину.
Семейство цистатина включает в себя орцицистатин I и II, которые происходят из риса. Другой член этого семейства происходит из кукурузы и отличается от ингибитора кукурузы 22 кДа. Ингибитор, происходящий из кукурузы 22 кДа, является бифункциональным ингибитором трипсина и α - амилазы, а также обладает противогрибковой активностью.
Семейство Bowman-Birk включает в себя ингибиторы протеиназы, выделенные из Vigna unguiculata (коровий горох). Ингибиторы трипсина коровьего гороха (Cp Tis) представляют собой небольшие полипептиды из около 80 аминокислот и являются двухконцевыми ингибиторами сериновой протеиназы. Были получены последовательности белка и кДНК CpTis (EP-A-0272144, Hilder и др.) (1989) "Protein and kDNA seguences of Bowman-Birk protease inhibitor from cowpea (Vigna unguiculata walp) "Plant Molecular Biology 13 701-710 and Hilder и др. (1987) Nature 300, 160-163). В этих работах сообщается, что изменение CpTi уровней в семенах коровьего гороха может коррелировать с полевой резистентностью к жукам Callosorbuch maculatus. Кроме того, введение Cp Ti в искусственную среду ингибирует развитие Heliothis, spodoptera, Diabrotica и Fribolium. При этом, также сообщается, что его употребление человеком вместе с пищей не приносило какого-либо вреда, а пища, приготовленная из этого коровьего гороха и скормленная крысам, не оказывала влияния на их рост. Кассета, включающая в себя промотор 35 CaMV и последовательность, кодирующую зрелый ингибитор CpTi, была введена в растение табака посредством Agrobacterium facien - опосредованной трансформации. При этом, наблюдалась коррекция между уровнем экспрессии CpTi и выживанием Heliothis vriescens, а также уровнем поражения растений личинками этих насекомых (Hilder и др. (1987) Nature, 300, 160-163; EP-A-0272144).
Существует много примеров, когда меры, предпринимаемые для защиты растений от насекомых-вредителей, не имели практического значения для защиты растения от нематод. Не имеется также данных относительно общей корреляции между природными генами резистентности к насекомым и нематодам. Никогда не наблюдалось случаев, где гены резистентности против картофельной нематоды (например, HI в Maris Piper) сообщали резистентность против жесткокрылых, например, колорадских жуков, или ген томата Mi, сообщающий резистентность против некоторых форм Meloidogyne spp., имел какое-либо значение для резистентности против насекомых. Кроме того, многие широко используемые инсектициды, такие, как хлороорганические соединения, органофосфаты и синтетические пиретроиды, не являются эффективными для нематод. Исключение составляет оксимкарбамат (алдикарб), но он имеет очень широкий спектр активности против ацетилхолинэстеразы и является также очень токсичным для млекопитающих.
Насекомые обнаруживают значительное различие в отношении своих протеиназ. Например, трипсин и химотрипсин- подобные ферменты обнаруживаются у саранчи и пчел, тогда как тиоловые протеиназы преобладают в пищеварительных процессах долгоносика Callosobrinchus maculatis. Кроме того, протеиназы насекомых требуют оптимальных pH, и ингибиторы, например, такие, как ингибитор трипсина сои, будут ингибировать протеиназы одних насекомых и не будут ингибировать протеиназы других насекомых. Поэтому эффективность CpTi против некоторых насекомых не может служить надежным показателем его эффективности против других насекомых. При этом, нет каких-либо оснований ожидать apriori, что нематоды имеют протеиназы, аналогичные протеиназам насекомых. В этом отношении нематоды отличаются от насекомых.
Можно ожидать корреляцию между проглатываемой пищей и пищеварительными ферментами. Существует большое различие в способах питания жалящих жесткокрылах насекомых, таких как амбрамный долгоносик Callosobruchus maculatis, и питающиеся флоэмой полужесткокрылые насекомые, такие как тля. Эти насекомые также вносят изменения в растения, однако, они не образуют синцитиллярных клеток, как это делают нематоды. По сравнению с травоядными жуками, галловые нематоды отличаются от них по своему механизму питания даже больше, чем тли. Из этого следует, что изучение насекомых, к сожалению, не дает надежной информации о питании нематод.
Имеющаяся информация о протеиназах нематод также слишком недостаточна, чтобы моно было с уверенностью утверждать, что CpTi или другой трипсин или даже ингибитор протеиназы оказывает влияние на пищеварительные процессы. Многие работы исследователей, посвященные организмам-паразитам, были направлены на ферменты, секретируемые инфицирующими личинками, и поэтому они скорее рассматривают инвазию хозяйских тканей, а не питание этих насекомых. В случае Ascaris suum было установлено, что инфицирующая личинка содержит сериновые протеиназы, но при этом предполагается, что в пищеварении взрослых особей участвуют тиоловые и карбоксильные протеиназы. Присутствующие ферменты являются стадие-специфическими и тесно связаны со специфическими требованиями паразитизма. Отсюда можно заключить, что любой ингибитор протеиназы, такой, как CpTi, должен оказывать воздействие на растительные нематоды, на что указывают исследования, проведенные на различных видах паразитов.
Аналогично, свободноживущие нематоды являются бактериядными, и ферменты, необходимые для переваривания их пищи, могут сильно отличаться от ферментов растительных паразитов. Действительно, в некоторых ранних работах, посвященных растительным паразитам, делается предположение, что они имеют протеиназы с более низкой или значительно более низкой активностью, чем свободно живущие нематоды, исследованные одновременно с растительными нематодами.
Из указанных выше исследований, показавших, что CpTi является эффективным ингибитором против насекомых и что нематоды обычно уничтожаются совсем другими механизмами, очевидно, что антинематодная активность ингибиторов протеиназы не может быть предсказана на основании их инсектицидной активности трипсиновых ингибиторов, описанных, например, в EP-A-0272144. Причина этого вероятно заключается в том, что указывается в EP-A-0272144 на стр. 3 (строка 58) - стр. 4 (строка 2), именно резистентность к насекомым трансгенного растения значительно увеличивается, если растение было генетически трансформировано таким образом, что оно получило способность экспрессировать ингибитор трипсина по всему растению.
Для использования в настоящем изобретении могут оказываться предпочтительными ингибиторы цистеиновых протеиназ, поскольку имеются свидетельства (представленные в Примерах), что по крайней мере некоторые из мажорных протеиназ нематод являются цистеиновыми протеиназами.
Предпочтительными ингибиторами протеиназы настоящего изобретения является ингибитор трипсина, в частности ингибитор трипсина (CpTi) коровьего гороха, однако, в некоторых случаях могут оказаться более предпочтительными ингибиторы трипсина или других протеиназ, происходящих, например, от других бобовых культур. Cp Ti может быть предпочтительно идентичным натуральному Cp Ti в одной из своих аллельных форм, но это необязательно. Например, модификации в аминокислотной последовательности могут не оказывать неблагоприятного действия на активность фермента, или даже наоборот, могут улучшить эту активность. В основном, можно сказать, что ингибитор протеиназы, используемый в настоящем изобретении, будет обладать свойствами, аналогичными свойствам натурального ингибитора протеиназы, и гомологией, достаточной для полноправного члена протеиназного семейства, такого, как Cp Ti - семейства, в соответствии с имеющимися научными критериями. Например, нуклеиновая кислота, кодирующая Cp Ti, может обладать достаточной гомологией, для гибридизации с нуклеиновой кислотой, кодирующей натуральный Cp Ti в жестких условиях гибридизации. Примерам таких жестких условий может служить использование солевого раствора приблизительно 0,9 М при температуре от 35oC до 65oC. Нуклеиновая кислота, используемая в настоящем изобретении, может, кроме того, удовлетворять вышеуказанным критериям, но для вырождения генетического кода. При необходимости, гликозилирование может присутствовать или отсутствовать.
Ингибиторы трипсина типа Bowman-Birk и коровьего гороха кодируются семейством умеренно повторяющихся генов, которые экспрессируются в семядолях коровьего гороха в виде четырех ингибиторов, которые могут быть разделены с помощью ионообменной хроматографии. Все эти четыре изо-ингибитора (fI, fII, fIII и fIV) имеют аналогичную трипсин-ингибирующую активность, измеряемую in vitro - анализом активации фермента. Главным изо-ингибитором коровьего гороха является fIV. Гены, кодирующие указанные изо-ингибиторы, обладают гомологией, достаточной для перекрестной гибридизации с кДНК - зондами полного Cp Ti в 0,45 М NaCl, 0,045 M Na-цитрата (3 xssC) при 68oC. Примеры первичной последовательности изо-ингибитора fIV (определенной с помощью секвенирования белка (Sammour (1985), PhD Thesis, University of Durham) или предсказанной в результате секвенирования кДНК Cp Ti представлены на фиг. 1 EP-A-0272144.
Трансгенные растения, обладающие резистентностью к нематодам, сообщаемой в результате экспрессии трансгена ингибитора трипсина или другого фермента, могут быть получены в соответствии с описанием, представленным в EP-A-0272144 (которое вводится в настоящее описание посредством ссылки в той степени, насколько это допускается законом). Вкратце, этот способ получения трансгенных растений включает в себя следующие стадии:
1. Выделение полиаденилированой РНК;
2. Получение кДНК для клонирования;
3. Синтез кДНК;
4. Синтез второй цепи;
5. Обработка концов-дц-ДНК для клонирования;
4. Клонирование в хозяина (например, E.Coli) с использованием вектора, такого, как плазмида (например, pBR 322 или pU C19);
7. Скренинг рекомбинантов на Cp Ti (или дугой ингибитор) - кодирующую последовательность;
8. Перенос клонированных Cp Ti - кодирующих последовательностей в вектор экспрессии (например, Agrobacterium BIN);
9. Перенос в организм хозяина для трансгенеза (например, Agrobacterium tumefaciens) и
10. Инфицирование растения хозяином.
Следует отметить, что необязательно точно следовать указанным стадиям. Например, для синтеза ингибитор-кодирующей ДНК может быть использована химическая техника синтеза ДНК. Модификации могут быть введены либо на стадии de-novo - синтеза, либо путем сайт-направленного мутагенеза, осуществляемого на уже имеющейся последовательности.
Аналогично, может быть использован соответствующий вектор экспрессии. Выбор подходящего вектора экспрессии может зависеть от присутствия в нем промотора, под контролем которого будет происходить экспрессия ингибитора. При этом может быть желательной нерегулируемая или индуцибельная экспрессия. Другой фактор, влияющий на выбор, заключается в том, что может быть предпочтительным, чтобы экспрессия происходила по всему растению, либо локализовалась в какой-либо его части, причем на практике, предпочтительными будут, вероятно, корне- или другие тканеспецифические промоторы. Для трансгенеза могут быть использованы и другие методы, например, конструирование последовательности, кодирующей соответствующий промотор, а в некоторых случаях может быть предпочтительным метод "микровыстреливания".
Настоящее изобретение может быть использовано для борьбы с некоторыми видами нематод, включая яванскую галловую нематоду и другие галловые нематоды. Примерами галловых нематод являются нематоды рода Heterodera и Globodera. В этот род входят такие виды, как H. glycines, H. shachtii (свекловичная нематода). H., аvenae (галловая нематода зерновых) и картофельные нематоды G. rostochiensis и G. pallida. Яванские галловые нематоды принадлежат к роду Mеloidogyne, в частности к видам M. javanica, M. halpa, M. arenaria и M. incоgnita. Другими важными в промышленном отношении нематодами являются нематоды рода Xiphinema, в частности, Xindex и X. Italiae (которая переносит на виноград вирус короткоузлия виноградной лозы (GFLV). X. americanum, которая имеет большое экономическое значение в США и в других регионах мира, и X. diversicaudatum (которая переносит на растения малины и другие растения вирус арибис-мозаики, на зерновые растения вирус мозаики костера, и на малину, розу, смородину черную и другие растения вирус кольцевой пятнистости земляники, и нематоды-паразиты рода Pratylenchus, в частности P. penetrans, P. brаnchurus и p. geae (которая связана с поражениями кукурузы), P. , coffeae, P. grachyurus и другие (кофе), P. coffeae и P. goodeyi (бананы), P. branchyurus (ананас посевной), P. geae и P. branchyurus (рис), P. branchyurus и P. geae eae (овощные культуры) и P. thornei (пшеница).
Следует отметить, что Pratylenchus spp. имеет широкий круг хозяев и не ассоциируется с каким-либо одним конкретным культурным растением. Padopholus similis аналогичны членам рода Pratylenchus и является даже более важным вредителем для бананов.
Нематоды-паразиты, такие, как нематоды рода Pratylenchus, проникают в корни и питаются, продвигаясь по корням и нанося сильные разрушения окружающим клеткам. Периодически, эти организмы могут покидать корни и внедряться в другие ткани, продолжая пожирать содержимое клеток и размножаться. Термин "свободноживущие" иногда используют в отношении нематод, таких, как Xiphinema, которые никогда не проникают в растение, но питаются из почвы путем протыкания растительных клеток своими стилетами. То есть, в растение проникают только их стилеты, а поэтому, фактически, эти вредители являются эктопаразитами, однако, тем не менее, настоящее изобретение также является эффективным в борьбе против этих вредителей.
Ингибитор может быть использован для защиты против более чем одной имеющей экономическое значение нематоды, поражающей культурное растение. Например, Meloidogyne и Heterodera glycines являются главными вредителями оси культурной, Meloidogyne и Pratylenchus являются главными вредителями для кукурузы, а Heterodera shachtii, Longidorus, Trichodorus и Paratrichodorus для сахарной свеклы.
Галловая нематода (такая, как Globodera или Heterodera) проникает с поверхности корня вовнутрь клетки, и разрушая клетку, обеспечивает себе исходное питание. Нематода Pratylenchus ведет себя аналогичным образом, поскольку она также проникает в клетку. Разница заключается только в том, что Pratylenchus питается от каждой клетки до того, как она проникает в нее. Если протеиназы участвуют в процессе миграции, то следует ожидать, что эти ферменты являются сходными. Аналогично, галловые нематоды при их начальном питании, перед тем, как модифицировать клетку, вытягивают из нее содержимое для своего питания. Если при таком экстракорпоральном пищеварении используются протеиназы, то они должны быть аналогичными протеиназам Pratylenchus.
Белки являются главным питанием для указанных паразитов, поскольку сами они неспособны синтезировать нужные аминокислоты. Протеиназы используются этими организмами для расщепления белков на более мелкие молекулы (пептиды), которые усваиваются ими как часть их пищи. Поэтому нематоды, питающиеся на растениях, должны обладать протеиназами, способными расщеплять белки, имеющиеся в растениях. Иначе говоря, природа этих протеиназ должна зависеть от конкретных аминокислот, имеющихся в растительных клетках и отвечающих потребностям пожирающих их организмов. Очевидно, нет таких нематод, которые бы не могли использовать имеющийся белок с помощью своих протеиназ. Поскольку многие нематоды могут поражать одинаковые растения, например, картофель является хозяином для Globodera нескольких видов Meloidogyne и Pratylenchus, то имеются все основания предполагать, что эти виды имеют одинаковые протеиназы. Поэтому, ингибиторы должны быть подходящими для ингибирования переваривания белков протеиназами способом, описанным в настоящем изобретении.
Для борьбы против конкретных видов нематод или против ряда различных нематод могут быть использованы комбинации различных ингибиторов. Этот принцип наилучшим образом может быть проиллюстрирован при рассмотрении картофельной нематоды (PCN). На второй ювенильной стадии PCN, а также для взрослой самки PCN, важную роль играет активность цистеиновой протеиназы, как в условиях кислотного pH, так и в условиях pH, близкого к нейтральному значению. Вероятно, что металлопротеиназы играют важную роль на второй ювенильной стадии при нейтральном pH, а для самок при кислотных значениях pH. Поэтому комбинация ингибиторов для этих обеих групп должна быть эффективной против начального развития на второй ювенильной стадии, а также против последующего развития самки. Ингибиторы карбоксипептидазы, происходящие из клубней картофеля и его листьев, являются характерными примерами ингибиторов металлопротеиназы.
Кроме того, как было показано выше, настоящее изобретение предусматривает использование двух (или более) ингибиторов для борьбы против двух различных нематод, которые могут поражать одни и те же культурные растения, но которые обладают до некоторой степени различными протеиназами. Например, Meloidogyne spp. и H. glycine обе поражают сою культурную, а Globodera и Meloidogyne spp. обе поражают картофель.
Многие культурные растения могут быть защищены способом настоящего изобретения, а поэтому они являются кандидатами для их генетического трансформирования в целях сообщения им способности экспрессировать один или несколько трансгенных генов, кодирующих ингибиторы протеиназы. В принципе, любое культурное или какое-либо другое растение, подверженное поражению нематодами, независимо от того являются ли эти нематоды корневыми или галловыми, могут быть защищены способами настоящего изобретения. Иллюстративными, но не ограничивающими примерами таких растений являются виноград, табак, томат, хлопчатник, масличный рапс и другие культуры, такие, как соя культурная, сахарная свекла, зерновые и картофель. Средствами настоящего изобретения могут быть также защищены декоративные растения. Настоящее изобретение относится к трансгенным растениям per se, которые в своей естественной форме подвергаются поражению нематодами, но не подвергаются в значительной степени поражению насекомыми. Предпочтительные отличительные особенности трансгенных растений настоящего изобретения приводятся в настоящем описании для разных вариантов изобретения.
Приведенные ниже примеры иллюстрируют, но не ограничивают настоящее изобретение. Эти примеры сопровождаются чертежами, где:
На фиг. 1 представлена схематически структура гена Т-ДНК от p ROK-Cp Ti + 5.
На фиг. 2 показан рост Globodera pallida на трансгенных картофельных растениях, который представлен как временная зависимость объема тела от времени после введения в корни растения паразитов на ювенильных стадиях. Растения либо экспрессировали (заштрихованные кружочки), либо не экспрессировали (незаштрихованные кружочки) ген лектина гороха. Объем нематод из растений, экспрессирующих лектин гороха, составлял 70% и 85% по сравнению с контрольными через 12 и 16 дней, соответственно. Величины объемов выражены в натуральных логарифмах и показывают экспоненциальный суточный рост на 23,9% в день в течение 2-12 дней для нематод на контрольных растениях.
На фиг. 3 показан рост Globodera pallida на трансгенных картофельных растениях, который представлен как изменение объема тела со времени после введения в корни растения инфективных ювенильных особей. Растения либо экспрессировали (заштрихованные кружочки), либо не экспрессировали (незаштрихованные кружочки) ингибитор трипсина коровьего гороха (Cp Ti). Объем нематод из растений, экспрессируюих Cp Ti, был значительно меньше, чем для контрольных растений через 12 дней. Величины объемов даны в натуральных логарифмах и показывают экспоненциальный суточный рост на 21,3% в день через 2-12 дней для нематод на контрольных растениях.
На фиг. 4 показан объем тела на стадиях развития Globodera pallida, присутствующей на картофельных растениях 12 дней после введения в корни растения инфективных ювенильных особей. Растения либо экспрессировали (+), либо не экспрессировали (-) ингибитор трипсина коровьего гороха (Cp Ti). Величины объемов даны в натуральных логарифмах, а пол незрелых особей определяли на предзрелой стадии (J4). Объемы незрелых особей на предзрелой стадии и самок на Cp Ti - растениях составляла 39% и 54% для самок и самцов соответственно.
На фиг. 5 показано соотношение каждого пола Globorera pallida на картофельных растениях 12-24 дней после введения инфективных ювенильных особей в корни растения. Растения либо экспрессировали (заштрихованные символы), либо не экспрессировали (незаштрихованные символы) ген лектина гороха; (самцы представлены квадратами, а самки - кружочками).
На фиг. 6 представлена круговая диаграмма, где показаны процентные содержания каждого пола Globodera Pallida на трансгенных растениях картофеля 12 дней после введения в корни растения. Растения либо экспрессировали (правая диаграмма), либо не экспрессировали (левая диаграмма) ингибитор трипсина коровьего гороха. Результаты были получены на основе наблюдений свыше 120 нематод для каждой обработки. Особи, которые были слишком незрелыми для определения пола, обозначены (J).
На фиг. 7, которая относится к примеру 4, показано, что уровень экспрессии Cp Ti - экспрессирующих растений составляет около 1% от полного белка.
На фиг. 8, которая относится к примеру 5, проиллюстрирован ингибирующий эффект различных концентраций ингибитора протеиназы, происходящего от кукурузы.
Примеры
Пример 1
Получение трансгенных растений картофеля
Трансгенный картофель получали следующим образом:
Конструирование гена Ca MV-35S-CpTi;
Ген Cp Ti был получен из плазмиды pUSSR c 3/2 (Hilder и др. (1987), Nature 33, 160-163 и EP-A-0272144). Cp Ti - кодирующая последовательность (240 п.о.) содержалась внутри 550 п.о. Alul-Scal - рестрикционного фрагмента. Этот фрагмент, также содержащий более длинную лидерную последовательность и 3'-нетранслированную последовательность (96 п.о.), клонировали в Smal - сайт- Ti - плазмидного вектора (происходящего от Agrobacterium tumefaciens) и p ROK2 (Bevan 1984), Nucleic Acids Researsh 12, 8711-8721 и Bevan и др. (1985), EMBO J, 4, 1921 - 1926), pBOK Cp. Ti + 5 (Hilder и др., 1987, см. выше).
Плазмида pBOK 2 является производной бинарного A. tumefaciens - вектора pBIN 19 (Bevan 1984, см. выше). Кроме того, она содержит ген резистентности к канамицину, активный в растительных клетках, сайт множественного клонирования, расположенный между высокоэкспрессирующими Ca MV-35S-промотором и последовательностью терминации транскрипции гена нопалин-синтазы (Guilley и др. (1982), Ce 11 30, 763-773 и Bevan и др. (1983) Nucleis Acids Research 11(2) - 369-385). Плазмиду pPOK-Cp Ti+5 переносили в штамм LBA 4404 бак. A tumefaciens, содержащий обезвреженную Ti - плазмиду (Bevan, 1984 см. выше). Более подробно структура гена Т-ДНК из p-POK Cp. Ti + 5 показана на фиг. 1.
Получение и генерация трансгенных растений
Трансгенные побеги картофеля получали с использованием агробактерии штамма LBA4404/pAL 5505, содержащего p ROK Cp Ti + 5, и регенерировали в зрелые растения в соответствии со следующей процедурой:
Аксенические культуры побегов Solanumtuberosum (сорт Desiree) выдерживали на стандартной поддерживающей среде для культур побегов (MS соли и витамины) (Murashige и Skoog (1962) Physiologia Plantarum 15, 473-497), 2% (мас. /об. ), сахарозы и 0,8% (мас./об.) DIFCO очищенный агар, pH 5,7, в регулируемых комнатных условиях с фотопериодом 16 ч (облучение при 40 мкЕ м-2с-1) при 24oC. (DIFCO означает торговую марку). Затем верхушки побегов субкультивировали на свежей среде каждые 14 дней. Штамм Agrobacterium tumefaciens, содержащий pROK Cp Ti, культивировали в dYT-среде (16 г/л триптона, 10 г/л дрожжевого экстракта, 5 г/л NaCl), содержащей 20 мг/л канамицина в течение 24 ч при 27oC. Затем бактерии осаждали, промывали три раза в 2 мМ MgSO4, ресуспендировали в PCM-среде (MS-соли и витамины, 2% (мас./об.) сахарозы, 2 мг/л 2,4-дихлорофеноксиуксусной кислоты, 0,5 мг/л зеатина, pH 5,7 при плотности приблизительно 109 кл/мл).
Вырезанные квадраты (прибл. 7х7 мм) из мезофилла картофеля от стабилизированной культуры побегов инкубировали 30 мин в бактериальной суспензии, слегка размешивая, затем быстро блотировали и переносили (нижней поверхностью вверх) на PCM-среду, содержащую 0,8% очищенного агара DIF CO. После 2-дневного культивирования при 24oC (фотопериод 16 ч, 40 мк Ем-2с-1 PAR), эксплантаты промывали в течение ночи в PCM, содержащей 100 мг/л аугментина, блотировали и переносили на отвержденную PCM, содержащую 100 мг/л аугментила и 100 мг/л канамицина кислый сульфат. После культивирования в течение 4 дней на PCM, содержащий 100 мл/л канамицина, 100 мг/л аугментина, эксплантаты переносили на среду для индуцирования побегов (MS - соли + витамины, 2% (мас. /об. ) сахарозы, 0,5 мг/л зеатина, 2 мг/л гибберелловой кислоты, 0,8% очищенного агара DIFCO, pH 5,7, содержащей 100 мг/л аугментина и 100 мг/л канамицина. После 6-недельного культивирования, трансгенные побеги вырезали и переносили на стандартную поддерживающую среду для укоренения побегов. Проростки субкультивировали каждые 14 дней до тех пор, пока они не достигнут стадии, приемлемой для их трансплантации в почву (приблизительно 3 субкультуры). Затем растения выращивали до полной зрелости.
Вестерн-блоттинг и ДСН-ПААГ-выделенные экстракты
В основе используемого способа лежал метод, описанный Laemmli (1970) Science 227, 680 - 685. Растворимые белки экстрагировали в фосфатно-солевом буфере (PB 8 г/л NaCl, 0,02 г/л KCl, 1,15 г/л Na2HPO4, 0,02 г/л KH2PO4, pH 7,2), и концентрацию доводили до 20 мг/мл, затем 50 мкл каждого выпаренного экстракта загружали в 17% полиакриламидный гель и подвергали электрофорезу в течение ночи при 7 мА. После этого выделенные белки переносили на нитроцеллюлозу, используя систему полусухого электроблоттинга (LKB Pharmacia). Раствор свободнотекущего порошка (5% мас./об. сухого молока в PBS), как описано Jonson и др. (1984), Gene Analysis Technigues 1,3-8.
Иммунологическое обнаружение связанного Cp Ti осуществляли с использованием кроличьих анти- Cp Ti первичных антител (поставляемых Durham University), а затем вторичных 125I-конъюгированных ослиных антикроличьих антител (Amersham International plc., Bucks). Фильтр инкубировали при 42oC в течение 2 часов с первичными антителами, а затем промывали 15 минут блокирующим раствором. После этой промывки, фильтр инкубировали со вторичными антителами также при 42oC и также 2 часа. После этого фильтр промывали как и в первый раз, а затем осушали и обрабатывали. Полученные фильтры подвергали авторадиографии традиционными методами. Вестерн-анализ подтвердил, что все трансгенные клоны экспрессировали зрелый Cp Ti-белок в молодых листьях и корнях.
Экстракция ДНК и Саузерн-анализ
Приблизительно 3 г ткани листьев растений замораживали в жидком азоте и измельчали вместе с промытым кислотой песком в ступке с помощью пестика. Затем к этому измельченному материалу добавляли 5 мл 100 мМ Трис-HCl (pH 8,5), 100 мМ NaCl, 50 мМ ЭДТК, 2% ДНС, 0,1 мг/мл протеиназы K. После 1-часового легкого размешивания при комнатной температуре, к суспензии добавляли 5 мл фенольной смеси (Stlarer (1984), The Extraction of Total RNA by the Detergent and Phenol method в "Methods in Molecular Biology", т. 2, изд. Walker J. M. , Humana Press, стр. 101-108), уравновешенной 100 мМ Трис HCl, pH 8,5, 100 мМ NaCl, 50 мМ ЭДТК. После 30-минутного легкого размешивания, смесь центрифугировали, и водную фазу снова три раза экстрагировали фенольной смесью: хлорформа (1:1, по объему), и один раз хлороформом. Затем к водной фазе добавляли 0,5 мл 3М ацетата натрия, pH 5,2 и 10 мл 100% этанола при комнатной температуре. Осажденную ДНК распрямляли, используя стерильную пластиковую петлю, и ресуспендировали в 0,4 мл ТЕ-буфера, содержащего 50 мкг/мл PHKазы, в течение 1-2 часов при 37oC.
Затем ДНК снова экстрагировали, как описано выше, и осаждали из водной фазы путем добавления 0,1 объемов 3 М ацетата натрия, pH 5,2, и 3 объемов 100% этанола, и инкубировали при -20oC в течение ночи. Затем ДНК промывали два раза в 70% этаноле, ресуспендировали в 50 мкл два раза дистиллированной стерильной воды и хранили при -80oC. 10 мкл - образцы переваривали реструктирующим ферментом Hind III и подвергали в течение ночи электрофорезу на 0,8% агарозном геле при 25 В. При этом использовали стандартную технику, описанную Маниатисом и др. (1982) в "Molecular Cloning: a Laboratory Manual, Cold Spriug Harbor Laboratory".
Затем гель депуринизировали в 0,25 М HCl в течение 10 мин при комнатной температуре, промывали в дистиллированной воде и денатурировали в 1,5 М NaCl, 0,5 М NaOH в течение 20 минут при комнатной температуре. ДНК переносили на найлоновые мембраны (Gene Screen Plus, Du Pont Co, NEN Products, Boston, Mass USA), используя LKB VACUGENE - систему вакуумного блотирования (Pharmacia Ltd., Milton Keynes, Bucks, UK). (Gene Screen Plus и VACUGENE означают торговые марки). После переноса, который осуществляли в течение 60 минут при комнатной температуре в вакууме при 40 см H2O в 0,4 М NaOH, фильтр нейтрализовали в 1,5 М NaCl, 0,5 М Триса, pH 7,5, в течение 15 минут при комнатной температуре, а затем тщательно осушали воздухом. До гибридизации, фильтры хранили при 4oC.
Полученные таким образом фильтры обрабатывали одним из двух зондов (фиг. 1). Они представляли собой Alul/SсaI-фрагмент (564 п.с.) от pUSSR с 3/2, который содержал CpTi-кодирующую последовательность, и Pst1/BamHI-фрагмент (800 п.о.) от pNEO, который содержал последовательность, кодирующую ген резистентности к канамицину NPT II, pNEO является производным pUC8, содержащим химерный ген резистентности к канамицину, обнаруженный в pBIN6 (Bevan, 1984, см. выше), и полученным из Отделения биологических наук, Даремский университет. Оба зонда были меченными с высокоспецифической активностью (> 9·108 число распадов в минуту на мкг) с использованием набора для рандомизированного примирования (Boehringer mannheim Ltd., Sussex). Для гибридизации и промывки использовали стандартную технику (Маниатис и др., 1982, см. выше). Фильтры подвергали авторадиографии в течение 4 дней при -70oC.
На фиг. 1 показана структура Т-ДНК, содержащая химерный ген Cp Ti, который был введен в растения картофеля. Единичный HindIII-рестрикционный сайт находился между Cp Ti и NPT II-генами. Таким образом, HindIII-перевар генома растения, содержащего одну копию Т-ДНК, будет генерировать два сочлененных фрагмента. Поскольку сайт интеграции, а следовательно, и расстояние до следующей смежной последовательности узнавания Hind III в растительной ДНК, будет варьироваться для отдельных трансформантов, то размер соединенных фрагментов также может варьироваться. Аналогично, множественные копии Т-ДНК, интегрированной в различные сайты внутри одного и того же генома, будут продуцировать соответствующее число полос, если используются зонды с последовательностью, специфичной к одному из участков соединения. Это позволяет совершенно точно определить число копий гена.
Для Саузерн-анализа, геномную ДНК экстрагировали из одной линии с относительно высокой экспрессией CP Ti и двух линий с низкой экспрессией, а затем переваривали Hind III. После выделения с помощью гель-электрофореза и последующего блотирования, полученные фильтры зондировали либо с Cp Ti - кодирующей последовательностью, либо с последовательностью, кодирующей резистентность к канамицину.
Каждая из двух низкоэкспрессирующих линий давала одну полосу, что свидетельствовало о присутствии одной копии химерного Cp Ti-гена, интегрированного в ДНК растения. Как и ожидалось, эти линии были разных размеров, что указывало на то, что интеграция имела место в различных сайтах внутри генома обоих трансформантов. Относительно высокоэкспрессирующая линия имела приблизительно семь копий Cp Ti.
Жесткая гибридизация с зондом, происходящим от NPT II-кодирующей последовательности, наблюдалась в геномной ДНК, экстрагированной из всех трех CpTi - положительных линий.
Альтернативно, трансгенный картофель может быть получен способами, описанными в EP-A-0272144.
Пример 2
Выведение и развитие G. pallida в трансгенном картофеле - культивирование трансгенных растений
Трансгенный картофель, полученный как описано выше, сначала отбирали как обычные растения, выращенные монаксенически на 50 мл питательного агара в 250-мл стеклянных сосудах с крышкой. Эти растения выращивали в вегетационной камере (Conviron SIOH) с циклом день:ночь = 16 ч : 18 ч (при 20oC и интенсивностью фотонного излучения 80 мгЭ/м2). Затем растения субкультивировали каждые 14 дней, снимая боковые стебли и сажая новые растения в дополнительные стерильные горшки. Таким способом культивировали 4 линии растений. Одна из них, как было показано выше, экспрессировала высокий уровень ингибитора трипсина коровьего гороха (Cp Ti), вторая была трансформирована аналогичной конструкцией, в которой отсутствовал Cp Ti-ген. Третий растительный экспрессировал лектин гороха, и наконец, четвертый клон имел аналогичную конструкцию, но в которой отсутствовал лектин гороха.
В первом эксперименте, растения с лектином гороха и их контроль переносили из агаровой культуры в небольшие горшки диаметром 5 см, содержащие песок с размером зерен 50-400 мкм для создания благоприятных условий для инвазии нематод. Затем в каждый горшок добавляли 500 свежевыведенных Globodera pallida. Через 4 дня, песок с нематодами, которые не были инвазированы в растения, промывали от растений, а растения пересаживали в песчаную суглинистую почву. Число особей, присутствующих на растениях, оценивали через 4 дня и через 21 день после инвазии.
Во всех последующих экспериментах, партии эксплантатов переносили не в песок или почву, а в вегетационные мешочки (Northrup king, minneapol and Vaugan Seed Co., Иллинойс). Эти растения выращивали в мешочках, содержащих 10 мл среды с 25% конц. солью Hoag lands, и в этих условиях растения культивировали 10 дней. После этого растения были готовы к инфицированию Globodera pallida.
Популяцию Globodera pallida, которая представляла собой стандартную популяцию фенотипа Pa2, выращивали на сорте Maris Peper на открытом воздухе, и инфицированную почву осушали и сохраняли. По необходимости, для этих экспериментов галлы экстрагировали из сухой почвы путем флотации (Soutkey j.F. (1986) "Laborаtory methods for Work c Plant и Soil Nematodes" Her Majеsty Stationery Office, London). Эти галлы отделяли от другого плавающего дебриса путем их "скатывания" с наклонной металлической пластины. Серии галлов (несколько сотен) погружали на 7 дней при 15oC, а затем переносили на 40 мкм найлоновое сито, которое частично погружали в диффузат из корней картофеля. Этот раствор изготавливали стандартным способом (Southey см. выше) путем пропускания воды через клубни молодого картофеля, после чего диффузат хранили в пластиковых сосудах при 4oC. Перед использованием диффузат фильтровали.
Молодые особи, появлявшиеся из галлов, использовали через два дня после их появления. Эксперименты хронировали так, чтобы имелось достаточное количество молодых особей для одновременного заражения экспериментальных партий и соответствующих контрольных растений.
Растения инфицировали в 1 см от кончика корня (4-6 корней) после того, как эти корни высаживали в бумажные пакетики, помещенные на ленточный фильтр из стекловолокна (Whatman l A). 50 особей на стадии 2 вводили примерно в 10 мкл диффузата корней картофеля в водопроводной воде на поверхность корня на расстоянии примерно 1-2 см от его верхушки, а вторую часть стекловолокна помещали над корнем. Фильтры вынимали через 24 часа и корневые системы промывали водой для удаления нематод, не внедрившихся в корни к этому времени. Затем растения возвращали в вегетационную камеру в их мешочках и держали их там до завершения эксперимента.
Инфицированные корни отделяли от растения и сразу погружали на 1 минуту в кипящий 1% кислый Fuschin (44). Затем корни обесцвечивали примерно в течение 24 часов в подкисленном глицерине при 50oC, в результате чего окрашенные нематоды были хорошо видны в корнях. После этого регистрировали число нематод в каждом кончике корня, их стадии развития и положение относительно васкулярной ткани. В некоторых экспериментах, объем каждой нематоды оценивали следующим образом. Контур особи обводили с использованием 40-х - объектива и камеры - люсида на ацетатной пластине. Микрометрическая шкала была также преобразована соответствующим образом. Затем эту пластину прикрепляли к экрану микрокомпьютера MD-DOS (AMSTRAD 1512) и контур особи копировали с помощью компьютерного манипулятора типа "мышь" (MSDOS и AMSTRAD означают торговые знаки). Для оценки объема по контуру нематоды использовали соответствующую компьютерную программу.
Результаты и обсуждение
Растения, инфицированные в песке, дали аналогичные степени инфицирования на 4-й день после инфицирования как для клонов, экспрессирующих лектин гороха, так и для их контроля. Через 21 день, большинство самцов покинуло растение, но процентное содержание особей, добавившихся к песку и уже сформировавшихся, и ставших самками, было аналогично для обоих клонов (табл. 1).
Растения были перенесены из песка в почву на 4-й день.
Величины представляют значения ± ср. кв. ош.; 100% = 500 нематод на растение.
Растения в мешочках были инфицированы обычным способом с использованием 20-30% особей, добавленных на поверхность корней с периодом инвазии 24 часа (точно). Такой ограниченный период инфицирования дает процент инвазии, аналогичный тому, который достигается в 4-дневном эксперименте с растениями, выращиваемыми на песке, но с тем преимуществом, что это инфицирование является в высокой степени синхронизированным. Тому факту, что животные не внедрялись в корни до 15 часа, свидетельствовала полоса, полученная в течение примерно 9 ч между первой и последней ивазированной особью. Это говорит об адекватной степени синхронности для данных экспериментов.
Экспрессия лектина гороха в корнях картофеля не препятствовала адаптации нематод на 2, 4 или 8 день после инвазии (табл. 2).
Все величины представляют собой процент нематод/кончик корня ± ср.кв.ош. (100% = 50 нематод на кончик корня).
Для растений с Cp Ti наблюдалась слегка более низкая инвазия, чем для их соответствующего контроля, однако этот эффект не имеет статистической значимости.
Рост Globodera Pallida после инвазии прослеживали по логарифмической кривой, которая представляет собой аппроксимацию экспоненциальной кривой, на ранних стадиях инфицирования (фиг. 1 и 2). После инвазии, оценивали объем особей, который составлял 126 ± 1,9 пл. В случае экспериментов с растениями, экспрессирующими лектин гороха, этот объем возрастал на 23,9% в течение первых 12 дней. Объем особей на растениях с лектином гороха составлял 70% от объема контрольных через 12 дней и 85% через 16 дней. Отсюда видно, что экспоненциальная фаза роста не сохранялась после 12 дня, а поэтому последующие эксперименты с Cp Ti-растениями завершали через 12 дней. Степень роста для нематод на контрольных растениях в этих экспериментах была эквивалентна 21,3%-ному возрастанию объема (в день) в течение 0-12 дней. Такая же скорость роста наблюдалась в течение 0-8 дней для растений, экспрессирующих Cp Ti, но в промежутке между 8-м и 12-м днем значительного увеличения объема не наблюдалось (фиг. 3).
Подавление скорости роста может наблюдаться для обоих полов, или оно может происходить в результате предотвращения развития самок. Последний случай имеет место для Globodera rostoсhiensis RoI на сортах, несущих ген HI, сообщающий резистентность к этому типу патогенов. Поэтому каждую особь на Cp Ti-растениях и на соответствующих контрольных растениях распределяли по их стадиям развития. Инвазивная стадия (J2) и последующая стадия очень малы, и поэтому для этих анализов их совмещали. Пол пред-взрослой стадии легко определяется и поэтому особи на этой стадии распределяли по полу. Взрослые особи прекращали развитие к 12-му дню.
Полученные результаты показали, что особи пред-взрослой стадии обоих полов были меньше, чем особи на контрольных растениях на 12-й день (фиг. 4). Самцы на пред-взрослой стадии развивались в кутикуле третьей стадии, и полный размер этих особей составлял лишь 39% от контрольных. В случае самок, пред-взрослые особи на Cp Ti-растениях составляли лишь 54% от размера этих особей на контрольных растениях.
Развитие самцов и самок более полно прослеживалось в эксперименте с лектином гороха. В этом эксперименте определяли соотношение полов на 12, 16, 18 и 24-й день после добавления особей к корням. Это соотношение составляло около 1 самец на 1 самку через 16 дней (фиг. 5). По истечении этого времени, самцы вылезали из ювенильной кутикулы, в которой они находились, и мигрировали из корней в поисках самок. Это явление давало сдвиг в соотношении полов в пользу самок, которые оставались в корнях в течение нескольких недель. Растения, экспрессирующие лектин гороха, показывали небольшое, но значимое снижение такого сдвига со значимой разницей в этих соотношениях на 18-й день. Это находится в соответствии с небольшим снижением в скорости роста особей, которое может приводить к небольшому замедлению достижения самцами взрослой стадии.
Эксперименты с Cp Ti-растениями завершали через 12 дней, поскольку небольшая скорость роста к этому времени может привести к дифференцированной гибели особей одного пола, что будет искажать данные о соотношении полов. К 12-му дню большинство животных может быть точно распределено по полу, но среди них были такие, которые не достигали взрослой стадии, и они регистрировались как молодые особи. Оба пола питаются на третьей молодой стадии, но самцы в пред-взрослой или взрослой стадии не питаются от растения. Самки же интенсивно питаются в течение более чем 25 дней, и именно на этой стадии они являются наиболее патогенными для растения. Оценивали свыше 120 особей для Cp Ti-растений и для контрольных растений. В случае контроля, наблюдалось следующее соотношение: 65 ± 55% составляли самки, 17±3% составляли самцы и 18±3% составляли особи молодой стадии (фиг. 6). Эти результаты аналогичны результатам для самок, развившихся из особей, которые рассматривались в более ранних экспериментах как стабилизированные в растениях с лектином гороха и в их контролях (табл. 1b. В противоположность этому, процентные содержания самок на Cp Ti - растениях составляли 30% (самцы 50±6%, молодые 30±5%), (фиг. 6). Такое сильное снижение числа самок по сравнению с контрольным числом было статистически значимым (X 2 < 0,005). Возможно, что это обусловлено снижением в развитии самок по отношению к самцам, что влияет на их соотношение к 12-му дню. Даже если все неидентифицированные по полу особи на 12-й день являются самками, то модифицированное соотношение 50:50 самок к самцам дает значительно меньшую частоту встречаемости самок по сравнению с контролем. Поэтому необходимо сделать дубль, чтобы исключить статистическую значимость неподтвержденного допущения, что на Cp Ti -растениях развитие самок является замедленным, а развитие самцов возрастает. Если указанное необоснованное допущение принимается для дубликата, то соотношение для Cp Ti составляет 50: 50, а для контроля 65 самок : 35 самцов. Очевидно, особенно необоснованным является допущение, что Cp Ti-растения, которые стимулируют развитие самцов до объема 54% от контрольных, должны также способствовать увеличению скорости их развития до взрослой стадии. Другое объяснение может заключаться в том, что если пол для этих видов является необратимым, то меньшее количество молодых самок будет проникать именно в Cp Ti-растения, а не в контрольные. Более интересным объяснением может служить тот факт, что определение пола происходит экзогенно, что приводит к относительному снижению числа особей, способных развиваться в самки на Cp Ti - растениях.
Полученные результаты выявили два эффекта воздействия Cp Ti - растений на Globodera pallida. Скорость роста животных снижается, а также снижается вероятность развития животных в особи - самки. Такое снижение скорости роста приводит к снижению конечного размера самки или к увеличению периода времени, необходимого для достижения ими полного размера. В обоих случаях, репродуцирование животных на культурном растении будет в целом снижаться либо путем снижения их полного размера и продуцирования яиц самками, либо путем замедления их жизненного цикла и соответственного снижения числа последующих генераций, которые могут затем развиваться на этом растении. Полный эффект снижения числа самок на растении или снижения их продуктивности, либо увеличения препатентного периода является значительным. Например, снижение числа самок в этом эксперименте составляет от 0,65 до 0,2 от полного числа, что в целом составляет снижение частоты встречаемости на 70%. Если рост самок после 12-го дня прекращается, то они будут слишком малы, чтобы содержать более чем 10 яиц, что составляет приблизительно 5-10% от их нормального размера. Кажется, что этот эффект влияния на число самок не так велик, но реально, продуцирование яиц снижается на 75%, что в целом, приводит к общему результату от двух указанных эффектов, который может быть оценен как (1-0,75)·(1-0,65) = 0,0875. Этот результат эквивалентен снижению репродуктивности более чем на 90%.
Регулирование популяций галловых нематод может быть достигнуто с помощью одной технологии по борьбе с вредителями или с помощью комбинированных методов. Экономические пороги для PCN варьируются в зависимости от агрономических условий, но обычно, они составляют 10 жизнеспособных яиц на 1 г почвы при посадке. Цели, которые ставятся при борьбе с вредителями, направлены на то, чтобы обеспечить гибель достаточного количества особей до их размножения, либо добиться снижения количества яиц, которое было бы достаточным, чтобы после снятия урожая и при посадке следующего, популяция особей этого вида не достигла своего прежнего уровня. Интенсивность размножения является параметром, зависящим от плотности посадки и от условий окружающей среды, но обычно, экономический порог величины популяции при посадке составляет 10х. Поэтому, для достижения полного экономического эффекта, необходимо, чтобы технология по борьбе с вредителями, применяемая отдельно, давала по крайней мере 90% смертности. Если эта мера недостаточно эффективна, чтобы дать такой результат (т. е. 90% гибели), то необходимо использовать комбинированные технологии. Например, сорт Maris Piper способен дать вышеуказанный результат 90% гибели Globodera rostochiensis, однако, другие сорта, обладающие резистентностью против G. pallida, часто не дают такого уровня подавления репродуктивности особей. Результаты, полученные с Cp Ti - растениями, очень обнадеживают. Очевидно, что сочетание таких факторов, как снижение скорости роста, уменьшение размеров самок и их продуктивности, и сдвиг в соотношении полов в пользу самцов, позволило бы достичь общего процента уничтожения вредителей свыше 90%.
Пример 3
Иллюстрация того факта, что можорные протеиназы нематод относятся к цистеиновым протеиназам
О роли протеиназ в инвазии, прикреплении и питании галловых нематод мало что известно, хотя было установлено, что до некоторой степени, эти протеиназы отвечают самым различным требованиям в процессе развития паразитических форм вредителей. Экстракорпоральная секреция протеиназ может иметь место во время миграции, индуцирования синцития и последующего питания, а поэтому, эти ферменты, очевидно, играют значительную интракорпоральную роль в пищеварении. Результат, полученный при эксперименте на G. pallida с использованием Cp Ti - растений является прямым указанием на то, что в данном случае, имеет место прямое воздействие на протеиназы нематод, а не опосредованное действие на растительные протеиназы в синцитии или в других участках растения. Поэтому для подтверждения присутствия таких ферментов при питании особей паразитов проводили эксперименты по изучению протеиназной активности у самок G. pallida.
(1) pH - зависимость протеолитической активности G. pallida
Самки G. pallida были удалены из корней картофеля и гомогенизированы в дистиллированной воде, забуференной ЭДТК 1 мм - NaOH (pH 6,0). Нематодный гомогенат осветляли с помощью центрифугирования при 13000 g в течение 2 мин и пропускали через неабсорбирующую хлопковую вату. После этого фильтрат использовали как источник неочищенной протеиназы. pH контролировали с использованием 0,1 М цитрат-фосфатного, натрий-фосфатного и фосфатно-карбонатного буферов для доведения pH до 3,0 - 6,0, pH 6,5 - 7,5 и 8,0 - 8,5, соответственно. Эти буферы смешивали с равными объемами нематодного гомогената и RUBISCO (150 мкг) в 0,04% ДСН, в результате чего получали конечный реакционный объем 75 мкмл. Смесь инкубировали при 25oC в течение 60 мин, и ферментную активность гасили путем добавления 25 мкл 15% (мас./об.) ТСА. Затем пробирки помещали на лед в течение 30 мин для полного осаждения. Супернатант осветляли путем центрифугирования (13000 g, 2 мин). Высвобождение ТСА - растворимых аминогрупп из переваленного RUВISCO использовали для количественной оценки протеолитической активности нематодных гомогенатов с помощью 2,4,6-тринитробензсульфоновой кислоты (TNBS). TNBS - реагент получали путем смешивания одного объема 0,3% (мас./об.) TNBS в воде с четырьмя объемами 4% (мас./об.) тетрабората натрия в 0,15 М NaOH. 250 мкл реагента смешивали с 75 мкл образца, и проводили реакцию 60 мин при 30oC, после чего реакцию прекращали путем добавления 1,25 мкмл 0,5 М HCl. Затем считывали оптическую плотность при 340 нМ для гомогенатов и RUBISCO - контролей. Результаты выражали в нМ TCA - растворимых аминогрупп, высвобожденных за 1 минуту при 25oC, при этом, использовали стандартную кривую на основе L-лейцина в CP - буфере.
Затем проводили эксперимент при pH 5,7 с использованием CP-буфера. Самок G. pallida промывали в 50 мМ Cp-буфера + 1 мМ ЭДТК и гомогенизировали в охлажденном льдом CP-буфере (50 мМ), центрифугировали и фильтровали, как описано выше. В экспериментах с использованием ингибиторов или активаторов (см. ниже) стадию промывки ЭДТК не проводили.
(2) Протеиназный анализ, проводимый для исследования ингибиторов и активаторов
25 мкл RUBISCO (из маточного раствора в CP-буфере, содержащего 1 мг/мл 0,04% ДСН) добавляли к 20 мкл образца гомогената и к либо 5 мкл CP-буфера либо 5 мкл ингибитора/активатора. После 60-минутной инкубации, реакцию прекращали путем добавления 30 мкл денатурированного буфера и кипячения в течение 2,5 мин.
(3) Ингибиторы и активаторы
Маточные растворы ингибитора и активатора (табл. 3) разводили в буфере и добавляли к нематодному экстракту. Перед добавлением к белковому субстрату, ингибитор/активатор предварительно инкубировали с ферментными растворами при 25oC в течение 30 мин.
(4) Электрофорез
ДНС - денатурированную смесь подвергали ДСН - электрофорезу в ПААГ с использованием 12,5% разрешающего геля и тока 30 мМ примерно в течение 4 часов. Гель окрашивали с использованием кумасси бриллиантовым голубым.
(5) Исследование с использованием желатина
Самок G. pallida гамогенизировали в CP-буфере, как описано выше, центрифугировали при 13000 g в течение 2 мин и пропускали через неабсорбирующую хлопковую вату. Прозрачный фильтрат использовали вместе с 0,6 М сахарозой и 0,001% бромофенолового синего для электрофореза в неденатурирующем (нативном) полиакриламидном геле (NPA GE).
(а) Электрофорез с использованием желатина
9% полиакриламидные гели, содержащие 0,1% желатин, получали путем растворения желатина в разделяющей гелевой смеси при ок. 50oC. Нематодные образцы загружали в 2,5% концентрирующий гель, не содержащий желатина, и проводили электрофорез при 30 мА в течение 4 часов, охлаждая водопроводной водой. Высокомолекулярный желатиновый субстрат, но не являющийся целевым белком иммобилизовали в 9% полиакриламидном геле.
Гели быстро промывали в дистиллированной воде, помещали в 75 мМ CP-буфер, содержащий 0,02% ДСН, и инкубировали при 25oC в течение ночи. Протеолитическую активность визуализировали как зоны пониженной окраски после окрашивания кумасси синим. Во втором эксперименте, образцы помещали в 9% гели, не содержащие желатин. Из 10 см геля вырезали две серии срезов (10х1 см). Одну серию сразу денатурировали путем кипячения с буферным образцом в течение 2,5 мин, а другую серию инкубировали в течение ночи с RUBISCO (25oC), а затем денатурировали. Не обработанные RUBISCO сегменты подвергали ДСН - элетрофорезу (12,5% разделяющий гель, 4 часа, фиксировали в смеси метанол: уксусная кислота:вода (40:10:50, по объему), и полученные фракционированные белки визуализировали путем окрашивания серебром. Сегменты, инкубированные с RUBISCO, кипятили в буферном образце, и образцы подвергали электрофорезу на 12% ДСН-ПААГ. Гель окрашивали кумасси голубым.
(в) Желатин-аффинная хроматография
Протеиназы, связывающиеся с желатином, экстрагировали желатин-агарозой (Sigma). Осветленный гомогенат самок (0,5 мл) загружали на 0,75 - мл слой желатина-агарозы (12,0х0,8 мм), которому предварительно уравновешивали СР-ДТТ. Колонку промывали СО-ДТТ, а конечные фракции буфер-ДТТ - элюирования собирали (0,5 мл). Затем загружали элюирующий буфер (содержащий 0,2 М глицина, 0,25 М цистеина и 0,5 М NaCl), и собирали шесть 0,5 мл - фракций. И наконец, СР - ДТТ пропускали через колонку и собирали три 0,5 - мл фракции. Отдельные фракции диализовали против 25 мМ CP-буфера при 4oC для удаления NaCl и аминокислот. Аликвоты брали либо для инкубирования с RUBISCO с последующим ДСН-ПААГ-анализом, либо сразу денатурировали. Образцы, обработанные RUBISCO, инкубировали в течение ночи при 25oC, а затем реакцию прекращали путем кипячения с буфером для образца в течение 2,5 мин.
(6) Использование колонки с Меркурилом для выделения протеиназ G. pallida
Предполагаемые цистеиновые протеиназы G. pallida должны иметь активные тиольные группы, которые могут способствовать их выделению с помощью органомеркуриловых реагентов, таких, как п-аминомеркурибензоат (РАМВА), который является сильным ингибитором G. pallida-протеиназ (табл. 3). Осажденный объем 0,75 мл п-гидрокси амино меркурибензоат-агарозы (sigma) (Sigma) выливали в 2-миллиметровый пластиковый шприц и промывали шестью колоночными объемами 50 мМ натрийфосфатного буфера (pH 8,0). Затем подготавливали агарозную колонку с 2,5 мМ меркаптоэтанола, 15 мМ сульфита натрия в натрийфосфатном буфере (pH 8,0). Этот тиоловый реагент максимизирует связывание тиола с белком и благоприятствует обратимому взаимодействию между белком и ингибитором. Затем колонку промывали пятью колоночными объемами 50 мМ PC - буфера (pH 5,7).
Незрелых самок гомогенизировали в PC - буфере (pH 5,6), содержащем 10% (об. /об.) диметилсульфоксида, 0,5% (об./об.) субанола, 0,1 M KCl, 1 мМ ЭДТК и 10 сульфата натрия. 0,5 мл гомогената добавляли в колонку, и этот объем элютанта рециркулировали в колонку (3х), после чего конечный элютант 0,5 мл сохраняли (1 фракция).
Колонку промывали для удаления не связанных с тиолом белков 0,5 мл х 5 объемами PC - буфера (pH 5,7), и все фракции собирали. Нужные белки выделяли из колонки либо с помощью 2 мМ HgCl2, либо с помощью 10 мМ меркаптоэтанола в CP-буфере pH 5,7. И опять, 0,5 мл х 5-фракции собирали, а колонку промывали 0,5 мл х 3 CP - буфера. Каждую фракцию концентрировали путем диализа против 20% (мас. /об. ) полиэтиленгликоля (мол. масса 3500), а HgCl2 - фракции обрабатывали денатурирующим буфером и подготавливали для ДСН-ПААГ. Фракции, собранные в результате элюирования меркаптоэтанолом, экстенсивно диализовали против CP-буфера (pH 5,7) при 4oC для удаления меркаптоэтанола и концентрировали путем диализа против полиэтиленгликоля. Образцы разделяли на две части, и первую аликвоту добавляли к 0,6 М сахарозе, 0,001% бромофенолового синего, и загружали в 9% - желатин -NPAG, после чего обрабатывали трис-веронал-буферной системой (pH 8,0). Гели в течение 5 часов вымачивали в PC -буфере (pH 5,7) + 0,02% ДСН. Гель окрашивали кумасси голубым и обесцвечивали в соответствии со стандартными процедурами.
Вторую аликвоту инкубировали в течение ночи с RUBISCO, затем реакцию прекращали путем кипячения в денатурирующем буфере в течение 2,5 мин. Образцы подвергали электрофорезу на ДСН-ПААГ, а гель окрашивали кумасси голубым.
pH -зависимость протеолитической активности G. pallida.
Влияние pH на гидролиз RUBISCO с помощью гомогената развивающихся самок было проиллюстрировано для pH 3,0 - 8,5 и pH 5,0 - 6,5. Оптимальное значение pH для RUBISCO - переваривающих протеиназ составляло 5,7 с возможным вторичным пиком при 7,5. Это свидетельствует о том, что либо (I) две группы протеиназ экспресссируют максимальную активность при их соответствующих оптимальных pH; либо (II) одна мажорная протеиназа демонстирирует свою активность как в кислотных, так и в щелочных условиях. Первый случай, очевидно, относится к разряду случаев, когда активность наблюдается в диапазоне pH 5,0-6,5. Это позволяет предположить наличие более чем одной кислотной протеиназы. Внезапное падение ферментной активности при нейтральном pH, а затем ее повышение при pH 7,5 указывает на наличие щелочной протеиназы.
Более подробное исследование было направлено на кислотные протеиназы, которые обнаруживали максимум протеолитической активности при pH 5,7. Кроме того, стабильно высокая активность в кислотном диапазоне pH 5,4-6,5 не сбрасывает со счетов возможность присутствия более чем одной протеиназы в неочищенных экстрактах самок-нематод.
Фосфат включали во все буферы, так, чтобы стимулировать негативное торможение по типу обратной связи протеиновых киназ, защищая протеиназы от фосфорилирования, которое может способствовать их инактивации.
Действие активаторов и ингибиторов
Для выявления влияния активаторов и ингибиторов на протеолитическую активность нематодных протеиназ, RUBISCO обрабатывали экстрактом при 25oC и при соотношении субстратного белка к нематодному белку (масс./масс.), равном 8. После 30-минутного инкубирования, реакцию завершали и анализировали с помощью ДСН-ПААГ. Полосу RUBISCO - субстрата в геле оценивали количественно с помощью денситометрии (Joyce Loble Chzomscan 3), причем, степень активности протеиназы оценивали по количеству оставшегося белка.
Присутствие сульфидриловых реагентов ДТТ и цистеина стимулировало эндопротеиназную активность (табл. 3). Напротив, когда экстракт инкубировали со всеми тремя меркуриловыми соединениями (РАМВА, РМА и PAMSA), наблюдалось почти полное ингибирование. Наблюдалось слабое ингибирование протеолитической активности цистеиновым ингибитором E-64 (21%, табл. 3). [E - 64 (L-транс-эпоксисукцинил-лейциламид -(4-гуанидино) - бутан является сильным нереверсивным ингибитором цистеиновых протеиназ, который не воздействует на цистеиновые остатки в других ферментах, и который не реагирует с низкомолекулярным тиолами, такими, как 2-меркаптоэтанол]. Это позволяет предположить, что главная протеиназа, ответственная за деградацию RUBISCO при pH 5,7, является натуральной цистеиновой протеиназой. Подтверждение этому было получено благодаря повышению активности при инкубировании ферментов с ДТТ и цистеином. Однако, остается неясным, способствуют ли эти агенты непосредственному увеличению активности протеиназ, либо они способствуют увеличению восприимчивости субстрата к ферментному расщеплению.
Протеолитическая активность в желатин-полиакриламидном геле
Протеолитическая активность в верхних областях геля свидетельствует о присутствии высокомолекулярных протеиназ. Имеются две или три зоны гидролиза, которые представлены Rf-величинами 0,28; 0,33 и 0,38. Они могут свидетельствовать о наличии различных ферментов или различных изоформ одного фермента. Оценка сегментов геля в эксперименте с отсутствием желатина, выявила лишь один мажорный полипептид с Mr около 62K. Кроме того, инкубирование участков, экспрессирующих деградацию желатина, с RUBISCO приводило к гидролизу субстратного белка. Еще одна область RUBISCO - гидролиза была обнаружена в последнем сантиметре сегмента геля, но полипептидная полоса была обнаружена после ДСН-ЭФ-ПААГ. Очевидно, что в нематодных экстрактах присутствует более чем одна мажорная желатин /RUBISCO - переваривающая протеиназа.
Желатин - аффинная хроматография
Фракции, которые были элюированы с колонки с желатин-агарозой, переваривали RUBISCO и содержали белок с Mr 62K. 30 мкг белка самок, загруженного в колонку с желатином-агарозой, было достаточно для обнаружения М625 - белка путем окрашивания серебром.
(6) Использование колонки с меркурилом для выделения G.pallida протеиназ
Белки, выделенные из колонки с PAMBA - агарозой, обнаруживали протеолитическую активность в желатин-акриламидном геле, но в меньшей степени, чем протеиназы, полученные непосредственно из неочищенных экстрактов (фиг.7). Окрашивание серебром выявило белок с Mr 62К, иммобилизованный на РАМВА - агарозе. Этот эксперимент подтверждает предварительное предположение о том, что мажорная протеиназа, обнаруживаемая в нематодах-самках, имеет именно это значение Mr, и что эта протеиназа является, вероятно, протеиназой цистеинового типа.
Пример 4
Действие низких уровней экспрессии Cp Ti
Продуцирование поликлональных антител против Cp Ti
Двум самкам-мышам в возрасте 5 недель (Ba I b c) путем внутрибрюшинной инъекции вводили 10 мкг Cp Ti, в смеси полного адъюванта Фрейнда и дистиллированной воды (50:50). Через три недели, обеим мышам вводили аналогичные дополнительные инъекции с использованием неполного адъюванта Фрейнда. После предварительного тестирования титра антител, одну мышь умерщвляли через три недели после последней инъекции, и ее сыворотку использовали в разведении 152500 для обнаружения Cp Ti в корнях. В этих целях использовали стандартный твердофазный иммуноферментный анализ (ELISA). Корни измельчали на льду, используя ступку и пестик, в PBS (фосфатно-солевой буфер, pH 7) + нерастворимый поливинилпирролидон. Этот материал гомогенизировали, затем 2 мин центрифугировали, а супернатант собирали. Планшеты для микротитрования (NUNC) в течение более чем 48 часов при 4oC покрывали 5 мкг белка корней на мл карбонатного буфера при pH 9,6. Растения, экспрессирующие Cp Ti и не экспрессирующие Cp Ti, обрабатывали аналогичным способом, и использовали серию внутренних стандартов путем добавления 0-60 кг дополнительного Cp Ti /5 мкг белка корней. Планшеты промывали PBS и блокировали с использованием 1% альбумина бычьей сыворотки (BSA, Sigma) в PBS-твине. После дополнительного промывания в PBS -твине, добавляли анти- Cp Ti -антитела при разведении 1:2500 в PBS - твине + 0,1% BSA в течение 2 часов, затем планшеты 8 раз промывали в PBS -твине, после чего, в концентрации, рекомендованной поставщиком (Sigma продукт A5153), добавляли вторичные конъюгированные со щелочной фосфатазой антитела против мышиных lgG в 0,1% BA + 1% козьей сыворотки, в течение 2 часов. Планшеты снова промывали, как описано выше, и добавляли субстратный п-нитрофенилфосфат при 1 мл на 1 мл буфера при pH 9,8 (10% диэтаноламин и 0,5 мМ MgCl2). Через 24 часа измеряли число отсчетов при 405 нм с использованием планшет-ридера (Biorad). Представлена серия результатов для корней растений, которые находятся в соответствии с данными, показанными на фиг. 3-6. Одна линия регрессии показана для Cp Ti - экспрессирующих растений (+Cp Ti), а другая для растений, не экспрессирующих (Cp Ti(Cp Ti)). В обоих случаях, для субвыборки полученных гомогенатов добавляли 0-60 нг Cp Ti. Разница между двумя линиями регрессии на фиг. 7 указывает на уровень экспрессии в Cp Ti-экспрессирующих растениях, который составлял около 1% полного белка.
Влияние низких уровней экспрессии Cp Ti на G. pallida.
Cp Ti - экспрессирующие растения, которые, как было показано с помощью ELISA, экспрессировали 0,1-0,5% полного растворимого белка в виде Cp Ti, использовали для подтверждения влияния Cp Ti на развитие G. pallida, и для определения являются ли уровни около 1% необходимыми для достижения значительного воздействия на нематоды. Эксперименты проводили с растениями в мешочках, как было описано ранее, с добавлением 50J2 на отобранные кончики корней.
Результаты показали, что растения, экспрессирующие низкие уровни Cp Ti (0,1-0,5%), имели значительно более низкое отношение самок к самцам по сравнению с двумя типами контрольных растений (фиг. 4). Это различие не может быть приписано замедлению развития в тот момент, когда может быть определен пол незрелой нематоды (фиг. 5).
Указанные в табл. 4 величины представляют собой число незрелых самцов и самок, наблюдаемых на корнях растений картофеля через 12-19 дней после инвазии. В качестве нетрансформированных растений использовали сорт Desiree, а Cp Ti - негативные и Cp Ti -позитивные растения были аналогичны растениям, описанным ранее, за тем лишь исключением, что Cp Ti - позитивные растения обнаруживали только 0,1 - 0,5% Cp Ti -экспрессию. Плотность особей на этих растениях составляла 4-12 нематод/инфицированный кончик корня с контролируемой инфекцией максимально до 4 кончика корня на растение.
Полученные данные были собраны от большого количества Cp Ti-растений по сравнению с контрольными растениями.
Отношение самцы : самки выше для Cp Ti-позитивных растений, чем для Cp Ti-негативных растений (X2=17,4; P<0,005).
Растения сорта Desirее и Cp Ti-растения собирали через 12 дней после инфицирования, и отношение незрелых особей делили на те, которые могут или не могут быть выделены по половому признаку к этому времени (см. табл. 5).
Для двух типов растений (X2=0,35; NS) значительной разницы в соотношении нематод, которые могут быть разделены по половому признаку, не наблюдалось.
Влияние низких уровней экспрессии Cp Ti на Meloidogyne incognita.
Cp Ti-экспрессирующие и неэкспрессирующие картофельные растения переносили из тканевой культуры в песчаную суглинистую почву на 4 недели для их укоренения в почве. Затем их переносили в почву, в которой были выращены томаты, сильно инфицированные Meloidogyne incognita. Растения выращивали 36 дней при 25oC в камере с регулируемыми условиями, имитирующими окружающую среду, а затем собирали корни. Большую часть каждой корневой системы собирали для подсчета яиц, используя 4 растения, экспрессирующих Cp Ti, и 5 растений, не экспрессирующих Cp Ti, и 5 растений, не экспрессирующих этого гена. Из каждого корневого образца удаляли яйца с использованием 1% гидрохлорида натрия в течение 2 мин, и для каждого растения подсчитывали число яиц с использованием стандартных препаратов для подсчета. Соотношение имеющихся эмбрионов и образовавшихся молодых особей оценивали для 100 яиц, выбранных наугад. Небольшую часть от каждой корневой системы брали от двух линий растений для ELISA-анализа на обнаружение Cp Ti, как описано выше.
Результаты, полученные из ELISA-анализа, позволяют предположить, что уровни экспрессии Cp Ti составляли лишь 0,1-0,2% от полного растворимого белка. Несмотря на такой низкий уровень экспрессии, Cp Ti-позитивные растения оказывали влияние на M. incognita. Число яиц, отложенных самками к 36-му дню, было значительно меньше для Cp Ti-экспрессирующих растений, чем для контрольных растений (P<0,0005, табл. 6). Процент откладывания эмбрионных яиц самками, паразитирующими на двух линиях растений были почти аналогичными (табл. 6). Если Cp Ti замедлял созревание самок, то следовало бы ожидать, что неэмбрионных яиц для этих растений будет больше, чем для контрольных. Однако, полученные результаты позволяют предположить, что основной эффект заключается в снижении уровня ежедневной кладки яиц, которое, очевидно, не влияет на процентное содержание яиц, из которых будут образовываться зародыши и на поверхности галла.
Разница в числе яиц/г корня является статистически значимой величиной. Средние значения были получены с использованием преобразованных значений (Iog10), но представлены как арифметические значения после обратного преобразования, t < 0,005, на основе анализа логарифмически преобразованных данных.
Данные, представленные в этом примере, имеют следующие значения:
a) продуцирование антител позволяет измерить уровень Cp Ti-экспрессии в корнях. Это измерение подтверждает присутствие Cp Ti в растениях;
b) для растений, обнаруживающих экспрессию в 1%, были получены данные, свидетельствующие о их очевидном влиянии на соотношение полов нематод;
c) эксперименты с низкими уровнями экспрессии Cp Ti подтверждают исходные данные о влиянии на соотношение полов, что позволяет предположить, что Cp Ti является эффективным в период после начала питания особей, но перед их половой дифференциацией. Это согласуется с результатами первоначальных экспериментов и указывает на то, что Cp Ti воздействует на стадии J2 и J3, которые являются первыми двумя стадиями питания личинок в растении. Это раннее действие Cp Ti не является результатом замедления скорости развития (табл. 7);
d) результаты, полученные для Meloidogyne, представляют особый интерес. Самка этой нематоды (яванская галловая нематода) питается из гигантских клеток, которые она образует в корнях растений, а растение реагирует на это образованием галла (типа опухоли) вокруг каждой самки или небольшой группы самок. Яйца откладываются индивидуально со скоростью несколько яиц в день на внутренней поверхности галла посредством их извержения через вульву самки. То есть, самка выделяет желатинированную массу вокруг себя, в которую и откладываются яйца (для выделения яиц из этой массы применяли стандартный способ с использованием гипохлорита). Тот факт, что эмбрионные яйца для Cp Ti+ и для Cp Ti- были одинаковыми, то можно предположить, что эти яйца были отложены в один и тот же период времени. Поэтому разница, наблюдаемая в табл. 6, обусловлена различием в яйценоскости, а не во времени развития самок до той стадии, в которой они должны откладывать яйца.
Пример 5
Ингибиторы протеиназ, происходящие от риса и кукурузы, также обнаруживают активность in vitro.
Получение ингибиторов протеиназы из риса
Процедуры экстракции осуществляли в соответствии с описанием, данным Abe K. и Arai (1987 Agric. Biol. Chem 51 2763-2768), для выделения оризацистатина (см. ниже). 50 г муки неполированного риса экстрагировали в течение 1 часа в 130 мл 25 мМ фосфатного буфера pH 7, содержащего 0,15 M NaCl. Супернатант собирали путем центрифугирования при 7000 g и 4oC в течение 20 мин. Прозрачный супернатант нагревали 10 мин при 80oC. Затем смесь снова центрифугировали, как описано выше, и в течение 1 часа, охлаждая льдом, к супернатанту добавляли 30% сульфат аммония. Осадок удаляли путем центрифугирования, а затем добавляли еще сульфата аммония до 65% насыщения. Осадок собирали центрифугированием, ресуспендировали в 10 мл 50 мМ ацетатного буфера (pH 4,9) и диализовали в течение ночи против того же самого буфера при 4oC. Раствор пропускали через колонку с Сефадексом C50 СМ 1,5х24 см, которую предварительно уравновешивали ацетатным буфером (pH 4,9). Белки элюировали в линейном градиенте 0-0,4 M NaCl, и собирали те фракции, которые обнаруживали способность ингибировать папаин в планшетах для титрования, как описано ниже. Фракции с ингибирующей активностью объединяли и получали препарат риса I (табл. 7). Аликвоту диализовали против 50 мМ фосфатного буфера (pH 6), содержащего 2 мМ ЭДТК и 2 мМ цистеина, после чего ее загружали снова в колонку с папаин-агарозой, уравновешенную тем же самым буфером, но содержащим также 0,1% В 35. Белки элюировали 50 мМ K3PO4 (pH 11,50) в 0,5 M NaCl. Элютант доводили до pH 6,5 с помощью 3М натрийформатного буфера (pH 2,9) и диализовали против dH2O, а затем использовали в качестве препарата риса II.
Получение ингибиторов из кукурузной муки
40 г кукурузной муки в течение ночи смешивали в 250 мл 25 мМ фосфатного буфера (pH 7,1), содержащего 0,1 M NaCl и при 4oC фильтровали через фильтровальную бумагу Whatman Vol, фильтрат центрифугировали при 2000 g и 4oC в течение 30 мин, и прозрачный супернатант доводили до 25 - 85% насыщения сульфата аммония. Осадок, полученный в результате центрифугирования при 20000 g и 4oC в течение 10 мин, снова растворяли в воде и диализовали в течение ночи при 4oC, а затем использовали как препарат, содержащий кукурузный ингибитор.
Протеолитическая активность G. pallida
Свежевыведенные молодые особи G. pallida инкубировали в 2,5 мМ 5 гидрокситриптамина в воде при 20oC в течение 4 часов. Эта процедура стимулирует выделение содержимого из ротового отверстия особей, которое может быть визуализировано с помощью белкового красителя Ponceau S, который диффундирует через небольшую аликвоту экспериментального образца, содержащего нематоды, и помещенного на предметном стекле микроскопа и под покровным стеклом. Нематоды удаляли из среды путем центрифугирования при 3000 g в течение 2 мин, а затем жидкость диализовали при 4oC против дистиллированной воды в течение ночи. Полученный материал лиофилизовали в аликвотах при -70oC и хранили вплоть до использования, а непосредственно перед использованием препарат восстанавливали до 2,5 мкг белка/мл dH2O при 5oC. Анализ проводили в реакционной смеси, полученной путем смешивания 50 мкл этого раствора с 20 мкл либо 100 мМ фосфатного буфера при pH 6,8, либо 100 мМ цитратного/фосфатного буфера при pH 5 + либо 10 мл испытуемого ингибитора при концентрациях, приведенных в табл. 7, либо dH2O для соответствующего контроля неингибируемого экссудата.
Анализ осуществляли путем покрытия планшетов для микротитрования (NHNC) 50 мкл 0,01 мкг/мл биотина/желатина в карбонатном буфере (pH 9,6) в течение ночи при 4oC. Эти планшеты блокировали 1% желатина и промывали PBS, а затем добавляли в течение 18 часов 75 мкл свежеприготовленной смеси. Реакцию останавливали путем 8-кратного промывания лунок PBS-твином, а затем добавляли 100 мкл авидин-конъюгированной щелочной фосфатазы с 1% козьей сывороткой в течение 1 часа и в каждую лунку. Эти лунки, как и раньше, промывали PBS-твином, а затем 2 раза в PBS, после чего добавляли 100 мкл 1 мг/мл P-нитрофенилфосфата в буфере для субстрата (10% диэтаноламина и 0,5 мМ MgCl2) при pH 9,8. Развитие окраски оценивали при 405 нм с использованием планшет-ридера (Biorad). Все образцы сравнивали со стандартной "пустышкой", в которых отсутствовали препараты нематодного экссудата в реакционной смеси.
Результаты показали, что экссудаты обнаруживают две различные картины ингибирования протеолитической активности при двух значениях pH. При pH 5 обнаруживается цистеиновая протеиназная активность, которая ингибируется E64 и стимулируется двумя известными активаторами цистеиновых протеиназ (цистеин и ЭДТК). Эту активность ингибируют Cp Ti и препарат риса I.
Протеолитическая активность при pH 6,8 не ингибируется Cp Ti или стимулируется цистеином, как и ожидалось. Однако, оба рисовых препарата обнаруживают ингибирующее действие на эту протеолитическую активность, которая также ингибируется ЭДТК. Кукурузный препарат является очень эффективным ингибитором этой протеолитической активности.
Последующие исследования были основаны на том же самом анализе, но с использованием целых гемогенатов самок G. pallida как источника протеиназной активности. Эти результаты подтверждают два оптимальных значения протеолитической активности при pH 5,7 и pH 6,8, которые были получены другими методами. Наиболее эффективным ингибитором в условиях эксперимента был белковый препарат, полученный из кукурузной муки. Поэтому были продолжены исследования с использованием этого кукурузного препарата с различной концентрацией белка в пределах 0-2 мкг в анализе, проводимом на планшетах для микротитрования так же, как и в вышеописанном анализе. Результаты показали, что 50% ингибирования имеет место при концентрации белка около 10 мкг в этом ингибирующем препарате, при этом, активность практически отсутствовала при 20 мкг/мл (фиг. 8). Пропускание препарата через папаин-аффинную колонку (Sigma) не препятствовало ингибированию в анализе на планшетах, что свидетельствовало о присутствии ингибитора, который не принадлежит к подгруппе цистеиновых протеиназ со специфической активностью против папаина.
Полученные данные имеют следующее значение:
a) Экссудат был, в основном (но необязательно), изготовлен из выделений, которые нематоды экскретируют в растения.
b) Использование экссудата от особи на стадии J2 является хорошим свидетельством ингибирования не столько тех протеиназ, которые находятся в нематодах, сколько тех, которые секретируются в растения.
c) Эти данные показали эффективность Cp Ti, против нематоды на стадии 2 при pH 5, но не при pH 6,8. Протеиназа при pH 5 имеет цистеиновую активность.
d) Результаты показали некоторую эффективность рисовых препаратов, содержащих оризацистатин.
e) Данные показали, что особенно высокой эффективностью обладает ингибитор, полученный из кукурузы при pH 6,8. Это также проиллюстрировано на фиг. 8.
Пример 6
Выделение гена, кодирующего другие ингибиторы цистеиновых протеиназ (Оризацистатин).
Результаты исследований, проведенных для Cp Ti, показали его воздействие на Globodera pallida и Meloidogyne incognita в трансгенных растениях, а также на протеиназы в гомогенатах самок от G. pallida и экссудатах от молодых особей этой нематоды. Сообщалось, то Cp Ti действует, в основном, как ингибитор сериновой протеиназы, тогда как данные, полученные выше (табл. 3 и 7), показали, что G. pallida также обладает цистеиновой протеиназой.
Это указывает на то, что рациональные способы получения резистентности против нематод должны включать в себя следующие стадии:
(I) Определение присутствия протеиназ в животных с использованием методов, описанных в данной заявке.
(II) Выделение ингибиторов протеиназ и идентификация их на активность против данной протеиназы.
(III) Выделение генов, кодирующих ингибиторы.
(IV) Трансформация культурного растения с использованием кассеты, содержащей нужные гены (пример был дан выше).
В качестве примера для ингибирования протеиназ нематод использовали G. pallida на стадии I и ингибиторы, такие, как Cp Ti и цистеиновые ингибиторы, полученные из кукурузы и риса. Для выделения ингибиторов протеиназы могут быть использованы процедуры, которые были описаны выше для выделения ингибиторов из кукурузы и риса. Выделенный белок или пептидные фрагменты, полученные путем переваривания протеиназами, такими как эндопротеиназа Glu-C из Staphylococcus aureus V8 (Sigma), могут быть использованы для получения информации о пептидной последовательности с использованием стандартных процедур или наборов, имеющихся в продаже. Эта информация может быть использована для построения группы повторяющихся олигонуклеотидных праймеров для экспериментов по выделению гена. Такие процедуры обычно осуществляли с помощью сконструированных олигонуклеотидных зондов для скрининга кДНК-библиотеки для риса в целях получения из этого растения цистеинового ингибитора оризацистатина (Abl, Emori, Kondo, Suzuki and Arai, 1987, J.Biol, Chem, 262, 16793-16797). В качестве примера стадии III следует указать, что были синтезированы как раз не повторяющиеся олигонуклеотиды, поскольку последовательность оризацистатина I является известной в результате секвенирования геномного клона (Kondo, Emozi, Abl, Suzuki и Arai, 1989, Gene 81, 259 - 265). Этот ген содержал интрон в середине своей кодирующей последовательности. Поэтому праймеры конструировали для ДНК-последовательности у 5'- и 3'-концов экзонов 1 и 2. Это конструирование (см. табл. 8) дало возможность ввести Smdl-сайт и Ndel-сайт в праймер 1 и 5' до старт-кодона и Eco RI-сайт у 3' до кодирующей области экзона 2 (праймер 4).
Oryza Sabiva L. japonicum выращивали до тех пор, пока побеги не достигнут примерно 10 см высоты, затем их собирали и помещали в жидкий азот приблизительно в 1 г - аликвотах. Геномную ДНК экстрагировали из этого материала с использованием стандартных методов, описанных De IIaporta, Wood и Hick (1984, A Maize DNA Miniprep стр. 36 - 37 в "Malmbеrg Messing Suss ex (изд.) Mo I ecular Biology of Pland: a Laborabory Coyrse Manual, Cold, Spring Harbor Press, Cold Spring Harbor, Нью-Йорк, США).
Оризацистатин I амплифицировали из геномной ДНК с использованием полимерно-цепной реакции (PCR) (см. Mс Pherson, Oliver и Guru, 1991, b "Mс Pherson, Anirke и Taylor изд. PCR: A Practical Approach, 1 RL, Oxford University Press. Оксфорд, Великобритания). Эти аликвоты были помещены в пробирку: 5 мкл 5хPCR - буфер (Northumbria Biological) 5 мкл 2 мМ dNTP-раствор (Pharmacia) 10 мМ каждого праймера (праймеры 1 и 4 в табл. 8), 0,1-0,5 мкг ДНК-мишени в 1 мкл и дост. кол. dH2O с получением полного объема 50 мкл после добавления 2,5 ед. Tag-полимеразы (Northumbria Biologicаl). Перед началом PCR, реакционную смесь покрывали 50 микролитрами минерального масла. Реакцию проводили при температуре термоячейки (LEP) в условиях 95oC, 5 мин, с последующими 40 циклами при 95oC, 1 мин, 55oC, 1 мин и 72oC, 2 мин. Затем образцы охлаждали до 4oC и собирали.
PCR-продукты подвергали электрофорезу с использованием низкоплавкой агарозы, и ДНК-полосу ожидаемой длины (около 650 п.м.) визуализировали с использованием этидийбромида (см. Сэмбрук, Фитч и маниатис (1989) Molecular Cloning: A Laboratory Manual, 2-е издание, 3 том, Cold Spring Harbor Press, Gold Spring Harbor, Нью-Йорк, США, техника, описанная в этой работе, использовалась в настоящих экспериментах).
Нужную ДНК выделяли из геля путем вырезания полосы и помещения ее в центрифужную пробирку spin-X (Costar). Пробирки охлаждали до -20oC в течение 20 мин, центрифугировали в микроцентрифуге в течение 5 мин, последние две стадии повторяли, после чего ДНК собирали в около 500 мкл раствора. ДНК выделяли путем осаждения этанолом (Сэмбрук и др., 1989, см. выше).
Эту ДНК - пробу использовали в PCR, как описано выше, затем лишь исключением, что в (I) использовали праймеры 1 и 2, а в (II) использовали праймеры 3 и 4 в целях амплификации экзонов 1 и 2, соответственно, причем с праймерами добавляли дополнительные основания (табл. 8). Эти продукты выделяли как описано выше.
Идентичность первоначально амплифицированной ДНК и последующих экзонов 1 и 2 была подтверждена с помощью трех рестриктирующих ферментов. Для экзона 1:1 мкг ДНК, 2 мкл одного из трех буферов (Pharmacia), 5 мкл dH2O-буфера и 5 ед. Sa II. Для экзона 2: так же, как и для экзона 1, но либо 5 ед. NaCl, либо 5 ед. Hind III. Контроль проводили для обоих экзонов, но без рестриктирующих ферментов. В обоих случаях ДНК вырезали в ожидаемых положениях в известных рестрикционных сайтах (Kondo, Emozi, Abe, Suzuki и Arai 1989, см. выше).
Экзон 1 и экзон 2 сшивали, используя PCR-метод рекомбинации с помощью перекрывающегося удлиняющего материала (Horton и Pease 1991, и в "Directed Mutagenesis: A Practical Approach; Ed Mc Pherson M.J.IkL Press at Oxford University Press Oxford и K). Праймеры 1 и 4 использовали в условиях, описанных выше, за исключением того, что температура циклов составляла: 94oC, 2 мин, 94oC, 30 с, 58oC, 2 мин и 72oC, 1 мин. Продукты PCR-реакции выделяли с помощью электрофореза с использованием 1% низкоплавкой агарозы. Мажорную линию (около 300 п.о.) вырезали из геля и выделяли, как описано выше. ДНК переваривали с использованием Eco R1 и Smal, как описано выше, после чего реакцию прекращали путем инактивации нагреванием. Продукты переваривания лигировали в полученный pBluescript IIKS + и SK + (Strategene). Смесь для лигирования использовали для трансформации E.coli DM5-альфа. Рекомбинанты отбирали путем инсерционной инактивации галактозидазы (скрининг Blne/white) и помещали в LB-среду. Последующие плазмидные препараты из этих клонов дали 300 п. о. вставок, которые переваривали Eco RI и SmAI. Секвенирование этой вставки подтвердило, что она содержит экзон 1, соединенный в своем 3'-конце с экзоном 2, которые были сшиты без добавления или потерь в кодирующей последовательности. Эти стандартные процедуры осуществляли с использованием инструкций поставщика и стандартной техники (Сэмбрук и др., 1989, см. выше). PCR в значительной степени зависит от конкретных параметров термоячейки и используемых химических реагентов. Поэтому, может оказаться необходимым корректировать условия для каждой PCR-реакции для оптимизации продуцирования реакционных продуктов.
Выделенная ДНК была походящей для использования в кассете, предназначенной для трансформации растения, как было описано ранее в данной патентной заявке.
Полученные в этом примере данные имели следующее значение:
a) Благодаря этому изобретению имеются все основания ожидать, что другие ингибиторы протеиназы могут быть, по крайней мере, такими же эффективными, как Cp Ti. По всей вероятности, существуют различия в протеиназах нематод различных видов, а также нематод одного вида, но на разных стадиях развития. Были описаны оризацистатин I и оризацистатин II.
b) Ингибиторы цистеиновых протеиназ представляют особый интерес. Благодаря настоящему изобретению стало возможным взять нематоду и определить, какие именно ингибиторы являются эффективными против нее, а также выделить ген для его использования в кассете в целях трансгенеза.
c) Оризацистатины I и II также доступны, как и кДНК-клоны.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЕ ИНГИБИТОРЫ ПРОТЕИНАЗЫ | 1995 |
|
RU2182598C2 |
| СПОСОБ ПОВЫШЕНИЯ УСТОЙЧИВОСТИ ИЛИ ТОЛЕРАНТНОСТИ К НЕМАТОДАМ РАСТЕНИЯ И ЕГО ПОТОМСТВА, КОНСТРУКЦИЯ ДНК, КОДИРУЕМЫЙ ЕЮ СЛИТЫЙ ПРОТЕИН, ПОВЫШАЮЩИЙ УСТОЙЧИВОСТЬ К НЕМАТОДАМ РАСТЕНИЯ И ЕГО ПОТОМСТВА | 1998 |
|
RU2230787C2 |
| СПОСОБ БОРЬБЫ С НЕМАТОДАМИ | 1991 |
|
RU2129373C1 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ФЛУОПИРАМ, ДЛЯ КОНТРОЛЯ НЕМАТОД | 2017 |
|
RU2773720C2 |
| ФРАГМЕНТ ДНК, ПОЛУЧАЕМЫЙ ИЗ Arabidopsis thaliana, ЕГО СУБФРАГМЕНТ ИЛИ КОМБИНАЦИЯ СУБФРАГМЕНТОВ, ПОСЛЕДОВАТЕЛЬНОСТЬ ХИМЕРНОЙ ДНК И ЕЕ ИСПОЛЬЗОВАНИЕ, РЕПЛИКОН (ВАРИАНТЫ) | 1996 |
|
RU2198219C2 |
| КОНСТРУКЦИЯ ДНК, КОНСТРУКЦИЯ РЕКОМБИНАНТНОЙ ДНК И СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ | 1994 |
|
RU2136757C1 |
| СИСТЕМА ДЛЯ УНИЧТОЖЕНИЯ РАСТИТЕЛЬНЫХ КЛЕТОК | 2001 |
|
RU2285045C2 |
| СИСТЕМА ДЛЯ УНИЧТОЖЕНИЯ РАСТИТЕЛЬНЫХ КЛЕТОК | 2001 |
|
RU2261277C2 |
| РАСТЕНИЕ И РАСТИТЕЛЬНАЯ КЛЕТКА, ТРАНСФОРМИРОВАННЫЕ ХИМЕРНЫМ ГЕНОМ | 2001 |
|
RU2275426C1 |
| БИО-НЕМАТОЦИДНАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ, УНИЧТОЖЕНИЯ ИЛИ БОРЬБЫ С НЕМАТОДАМИ В КУЛЬТУРАХ СЕМЕЙСТВА ПАСЛЁНОВЫХ И ЕЕ ПРИМЕНЕНИЕ | 2012 |
|
RU2636550C2 |
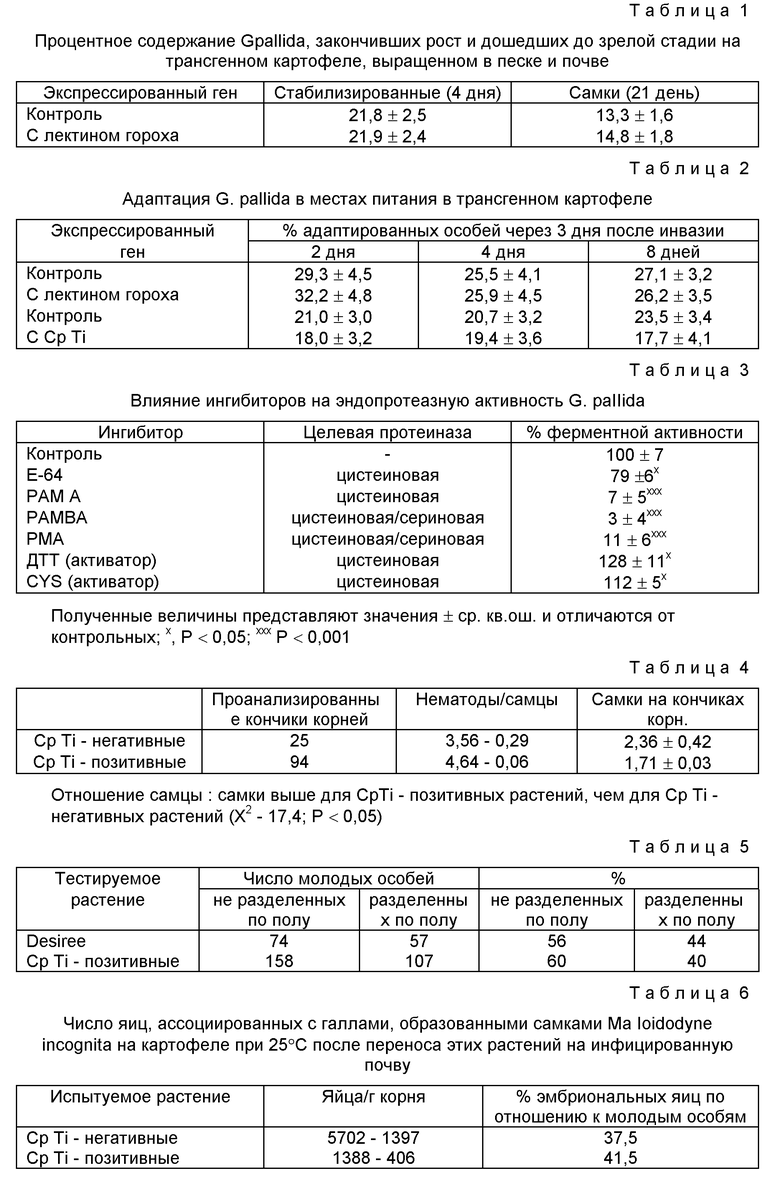
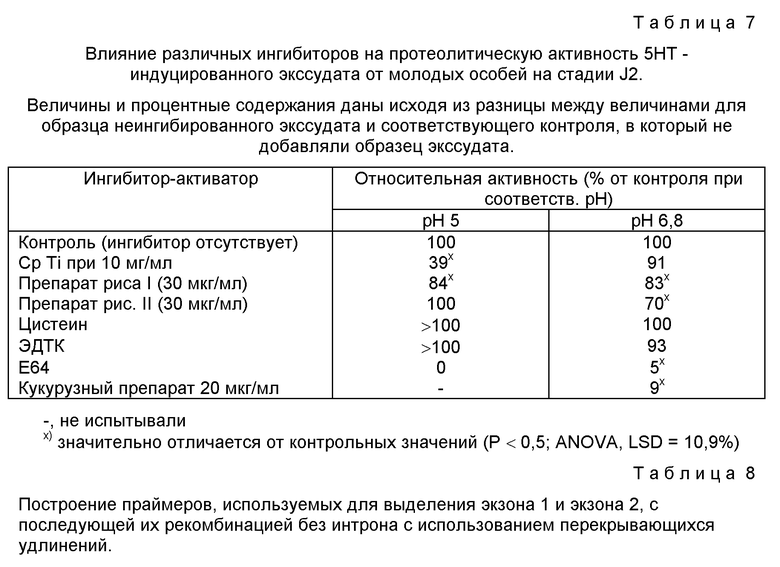

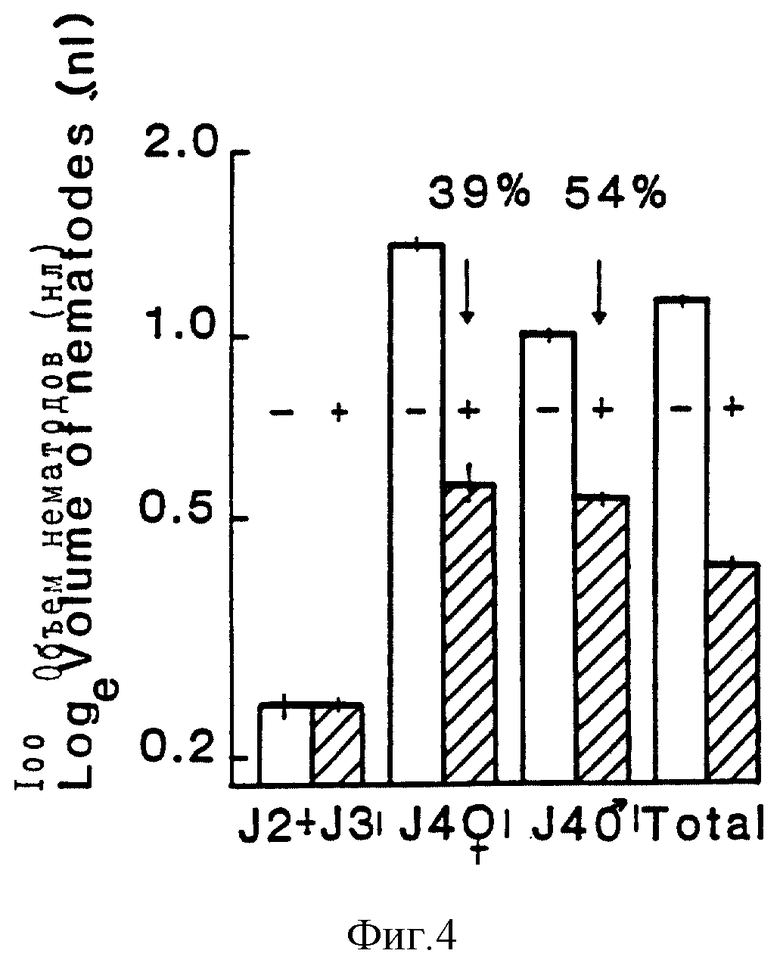
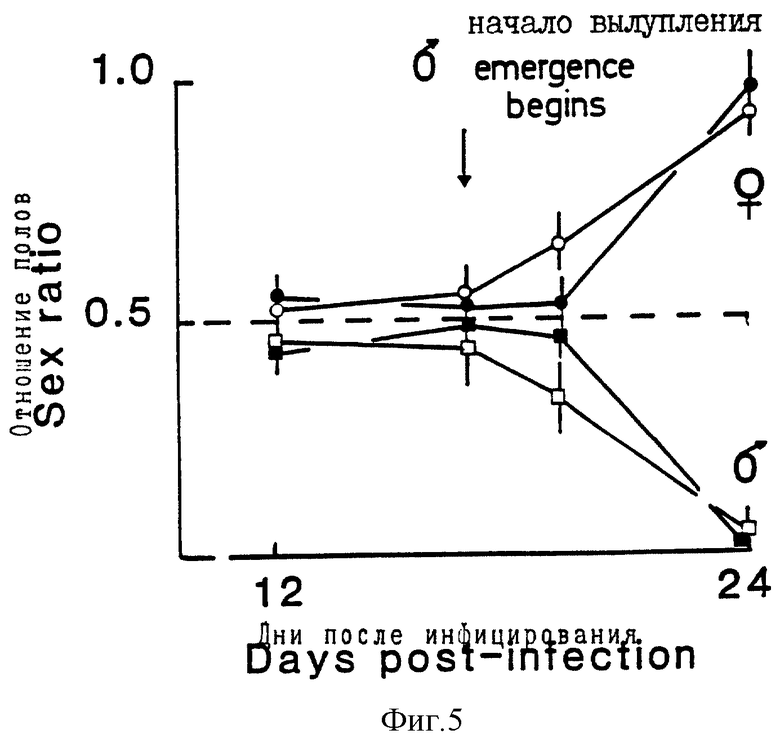
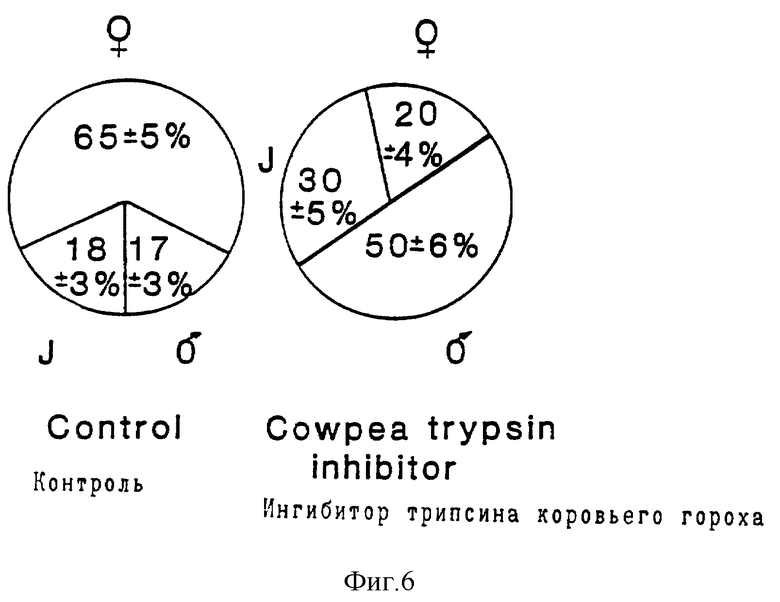
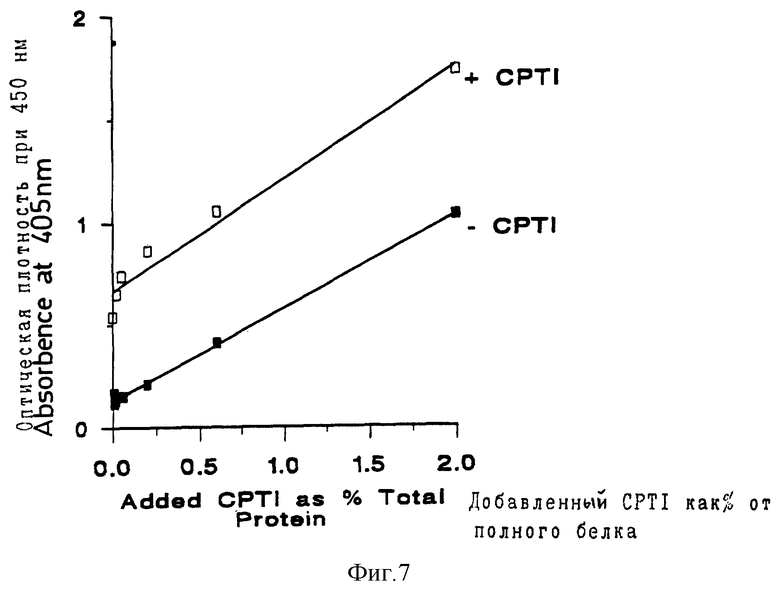
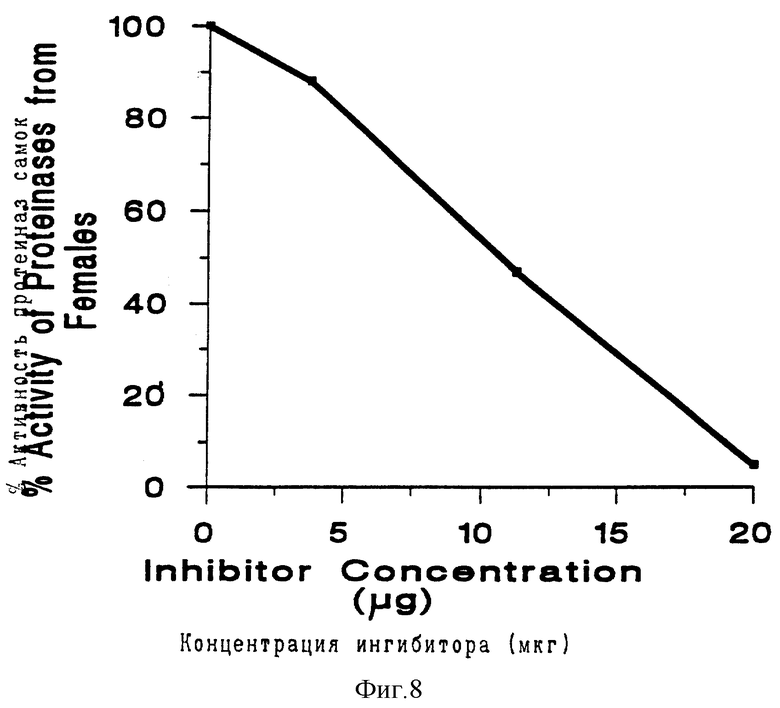
Изобретение относится к борьбе с насекомыми-вредителями, в частности с нематодами. Для снижения способности к размножению растительных паразитических нематод используют растение, в котором экспрессируется белок ингибитора трипсина коровьего гороха (СрTi) или белок ингибитора протеиназы цистеина, полученные из кукурузы или риса. Из белков ингибиторов может быть использован изоингибитор fIV или оризацистатин I или II. Способ может быть использован для защиты таких культур, как соя, сахарная свекла, картофель, зерновые культуры, табак, хлопок, масличный рапс и другие против нематод Heterodera glycines, Heterodera shachtii, Meloidоgyne javanica. 2 с. и 8 з. п. ф-лы, 8 ил., 8 табл.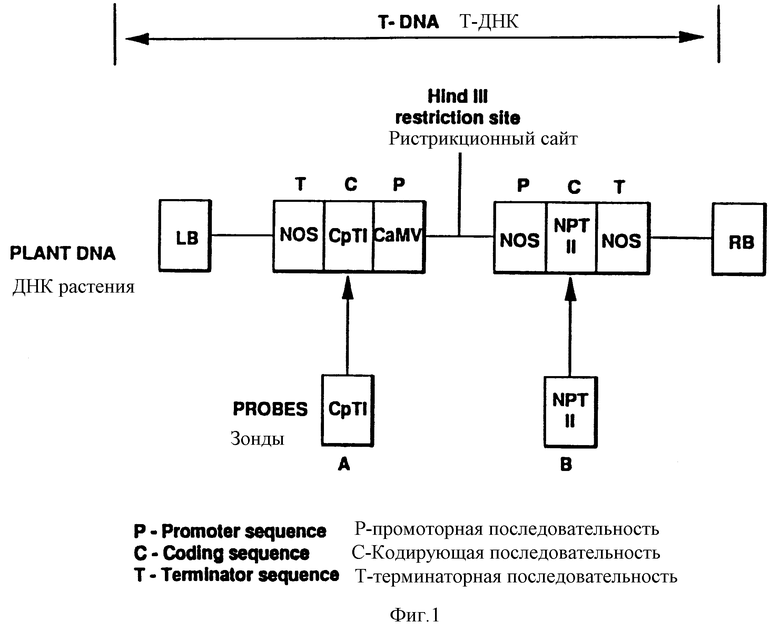
| ЗАТВОР ВОДОПРОВОДНОЙ ГАЛЕРЕИ | 0 |
|
SU272144A1 |
| ЗВУКОПОГЛОЩАЮЩИЙ КОЖУХ | 0 |
|
SU348348A1 |
