Настоящее изобретение относится к области химии нуклеиновых кислот. В частности оно относится к способам амплификации последовательностей нуклеиновых кислот и к способам уменьшения неспецифической амплификации.
Метод с использованием полимеразной реакции синтеза цепи (называемой также полимеразной цепной реакцией или ПЦР) для амплификации последовательностей нуклеиновых кислот широко известен в данной области техники и описан в патентах США 4683202; 4683195 и 4965188. Торговые фирмы, такие, как Perkin Elmer, Norwalk, CT, продают реагенты для ПЦР и публикуют протоколы ПЦР.
В каждом цикле ПЦР-амплификации денатурируют двухцепочечную последовательность-мишень, для каждой цепи денатурированной мишени отжигают праймеры и праймеры удлиняют под воздействием ДНК-полимеразы. Специфичность амплификации зависит от специфичности гибридизации праймера. Праймеры выбирают так, чтобы они были комплементарными или практически комплементарны последовательностям, находящимся на 3'-конце каждой цепи полинуклеотидной последовательности-мишени. При повышенных температурах, применяемых при обычной ПЦР, праймеры гибридизируются только с нужной последовательностью-мишенью. Однако реакционные смеси для амплификации обычно готовят при комнатной температуре, значительно более низкой, чем температура, необходимая для обеспечения специфичности гибридизации праймера. При таких менее жестких условиях праймеры могут неспецифично связываться с другими только частично комплементарными полинуклеотидными последовательностями (или даже с другими праймерами) и инициировать синтез нежелательных продуктов удлинения, которые могут быть амплифицированы наряду с последовательностью-мишенью. Амплификация неспецифических продуктов удлинения праймера может конкурировать с амплификацией нужных последовательностей-мишеней и может в значительной степени снижать эффективность амплификации нужной последовательности. Проблемы, вызываемые неспецифической амплификацией, описаны, кроме того, у Chou и др., 1992, Nucleic Acids Research, 20(7): 1717-1723.
Неспецифическая амплификация может быть снижена путем уменьшения образования продуктов удлинения от праймеров, связанных до начала реакции с последовательностями, не являющимися мишенями. В одном из методов, названном протоколом "hot-start" (протоколом "горячей инициации"), один или несколько важных реагентов не добавляют в реакционную смесь до тех пор, пока температура не поднимется до уровня, достаточного для обеспечения необходимой специфичности гибридизации. Таким образом, в реакционной смеси не может осуществляться удлинение праймера в то время, когда условия реакции не гарантируют специфическую гибридизацию праймера.
"Hot-start" - методы могут быть осуществлены вручную открытием реакционной пробирки после начальной стадии высокотемпературной инкубации и добавления недостающих реагентов. Однако осуществляемые вручную "hot start" - методы являются трудоемкими и увеличивают риск загрязнения реакционной смеси. "Hot-start" - методы, в которых применяют легкоплавкий материал, такой, как воск, для разделения или секвенирования реакционных компонентов, описаны в патенте США 5411876 и у Chou и др., 1992, см. выше. В этих методах высокая температура инкубации до начала реакции плавит легкоплавкий материал, позволяя, таким образом, реагентам смешиваться.
Другой способ уменьшения образования продуктов удлинения от праймеров, связанных до начала реакции с последовательностями, не являющимися мишенями, основан на ингибировании ДНК-полимеразы за счет использования соединения, которое нековалентно связывается с ДНК-полимеразой термообратимым образом. В патенте США 5338672 описано применение антител, специфичных в отношении термостабильной ДНК-полимеразы, для ингибирования ДНК-полимеразной активности. Антитела необходимо инкубировать с помощью ДНК-полимеразы в буфере при комнатной температуре до составления реакционной смеси, чтобы обеспечить образование комплекса антитело-ДНК-полимераза. Ингибирование антителом ДНК-полимеразной активности инактивируют путем высокотемпературной инкубации до начала реакции. Недостатком этого метода является то, что получение антител, специфичных в отношении ДНК-полимеразы, особенно в больших количествах, является дорогостоящим и требует значительных затрат времени. Кроме того, добавление антител в реакционную смесь может потребовать повторного планирования реакции амплификации.
Образование продуктов удлинения также можно ингибировать путем добавления соединения, которое нековалентно связывается с праймерами термообратимым образом, тем самым препятствуя гибридизации праймеров с любой последовательностью, как с мишенью, так и с какой-либо другой. Например, одноцепочечный связывающий протеин, добавленный в реакционную смесь, будет связывать праймеры, препятствуя, таким образом, гибридизации праймера и ингибируя удлинение праймера. Улучшение выхода продуктов ПЦР при использовании гена белка 32 описаны у Schwartz и др., 1990, Nucleic Acids Research 18(4): 10.
Неспецифическую амплификацию можно уменьшить путем разложения продуктов удлинения, образованных от праймеров, связанных до начала реакции с последовательностями, не являющимися мишенями, например, с применением способов, описанных в патенте США 5418149 и в Международной заявке WO 92/01814. Разложение вновь синтезированных продуктов удлинения достигают путем включения в реакционную смесь дУТФ и урацил-ДНК-гликозилазы (UNG) и инкубации реакционной смеси при 45-60oC до осуществления реакции амплификации. Недостатком этого метода является то, что разложение продукта удлинения конкурирует с образованием продукта удлинения, и удаление неспецифического продукта удлинения праймера, вероятно, будет менее полным.
Обычные методы молекулярной биологии, химии белка и химии нуклеиновых кислот, известные в данной области техники, достаточно полно описаны в литературе. См., например, Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York (Sambrook и др., 1989); Oligonucleotide Synthesis (M.J. Gait, ред., 1984); Nucleic Acid Hybridization (B.D. Hames и S.J. Higgins, ред-ы, 1984); Chemical Reagents for Protein Modification (CRC Press); и в сериях Methods in Enzymology (Academic Press, Inc. ). Все упомянутые в заявке патенты, заявки на патент и публикации, указанные как выше, так и ниже, включены в настоящее описание в качестве ссылки.
Настоящее изобретение относится к способам и реагентам для амплификации нуклеиновой кислоты с использованием реакции амплификации, основанной на применении праймера, как это более конкретно указано в прилагаемой формуле изобретения. Эти способы и реагенты обеспечивают простое и экономичное решение проблемы неспецифической амплификации. В способах используют обратимо инактивированный термостабильный фермент, который может быть реактивирован путем инкубации в реакционной смеси для амплификации при повышенной температуре. Неспецифическая амплификация заметно уменьшается вследствие того, что реакционная смесь не поддерживает удлинение праймера до тех пор, пока температура реакционной смеси не поднимется до уровня, который гарантирует специфичность гибридизации праймера.
Одним предметом настоящего изобретения являются обратимо инактивированные термостабильные ферменты, которые получают в результате реакции между термостабильным ферментом, который катализирует реакцию удлинения праймера, и модифицирующим реагентом. Реакция приводит к значительному, предпочтительно практически полному, снижению ферментативной активности. Инкубация модифицированного фермента в водном буфере при щелочном значении pH при температуре ниже приблизительно 25oC практически не приводит к увеличению ферментативной активности в течение промежутка времени менее приблизительно 20 минут. Инкубация модифицированного фермента в водном буфере, значение pH которого доведено до 8-9 при 25oC, при температуре выше приблизительно 50oC приводит по крайней мере к двукратному увеличению активности удлинения праймера в течение промежутка времени менее приблизительно 20 минут. Обратимо инактивированные термостабильные ферменты по изобретению в их активном состоянии либо катализируют удлинение праймера, либо необходимы для того, чтобы происходило удлинение праймера. Предпочтительные ферменты включают термостабильные ДНК-полимеразы и лигазы.
Предпочтительными модифицирующими реагентами являются ангидриды дикарбоновых кислот общей формулы: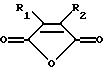
где R1 и R2 обозначают водород или органические радикалы, которые могут быть связаны, или общей формулы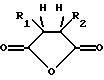
где R1 и R2 обозначают органические радикалы, которые могут быть связаны, и атомы водорода находятся в цис-положении. Органические радикалы могут быть непосредственно присоединены к кольцу через связь углерод-углерод или через связь углерод-гетероатом, такую, как связь углерод-кислород, углерод-азот или углерод-сера. Органические радикалы также могут быть связаны друг с другом с образованием циклической структуры, как, например, в случае 3,4,5,6-тетрагидрофталевого ангидрида.
Предпочтительные реагенты включают малеиновый ангидрид; замещенные малеиновые ангидриды, такие, как цитраконовый ангидрид, цис-аконитовый ангидрид и 2,3-диметилмалеиновый ангидрид; экзо-цис-3,6-эндоксо-Δ4 4-тетрагидрофталевый ангидрид и 3,4,5,6-тетрагидрофталевый ангидрид. В частности цитраконовый ангидрид и цис-аконитовый ангидрид предпочтительны для получения обратимо инактивированных ДНК-полимераз для использования в ПЦР-амплификациях.
Другим предметом настоящего изобретения являются способы проведения реакции амплификации нуклеиновой кислоты с использованием обратимо инактивированного термостабильного фермента по настоящему изобретению. В настоящем изобретении предложен способ амплификации содержащейся в образце нуклеиновой кислоты-мишени, включающий следующие стадии:
(а) введение образца в контакт с реакционной смесью для амплификации, содержащей праймер, комплементарный нуклеиновой кислоте-мишени, и модифицированный термостабильный фермент, где модифицированный термостабильный фермент получают путем реакции смеси термостабильного фермента, который катализирует реакцию удлинения праймера, с модифицирующим реагентом, причем реакцию проводят при щелочном значении pH и температуре менее приблизительно 25oC, и указанная реакция приводит к химической модификации фермента, являющейся причиной практически полной инактивации ферментативной активности, и причем инкубация модифицированного фермента в водном буфере, значение pH которого доведено до 8-9 при 25oC, при температуре выше приблизительно 50oC приводит по крайней мере к двукратному увеличению ферментативной активности в течение промежутка времени менее приблизительно 20 минут; и
(б) инкубацию образовавшейся на стадии (а) смеси при температуре выше приблизительно 50oC в течение промежутка времени, достаточного для реактивации фермента и позволяющего образовать продукты удлинения праймера.
В качестве предпочтительного способа в настоящем изобретении предложен способ амплификации содержащейся в образце нуклеиновой кислоты-мишени, включающий:
(а) введение образца в контакт с реакционной смесью для амплификации, содержащей праймер, комплементарный нуклеиновой кислоте-мишени, и модифицированный термостабильный фермент, где модифицированный термостабильный фермент получают путем реакции смеси термостабильного фермента с ангидридом дикарбоновой кислоты общей формулы: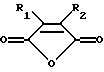
где R1 и R2 обозначают водород или органические радикалы, которые могут быть связаны, или общей формулы: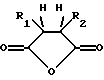
где R1 и R2 обозначают органические радикалы, которые могут быть связаны, и атомы водорода находятся в цис-положении, причем реакция приводит к практически полной инактивации ферментативной активности; и
(б) инкубацию образовавшейся на стадии (а) смеси при температуре выше приблизительно 50oC в течение промежутка времени, достаточного для реактивации фермента и позволяющего образовать продукты удлинения праймера.
В предпочтительных примерах осуществления способов по изобретению применяют обратимо модифицированные ферменты, модифицированные с использованием предпочтительных модифицирующих реагентов. В некоторых примерах осуществления изобретения стадию инкубации, т. е. стадию (б), проводят до начала реакции амплификации. В других примерах осуществления изобретения инкубация, которая приводит к реактивации фермента, является составной частью процесса амплификации. Например, стадия денатурации, которую осуществляют в каждом цикле ПЦР, может одновременно выполнять функцию реактивации модифицированной ДНК-полимеразы.
В предпочтительном примере осуществления изобретения реакция амплификации представляет собой полимеразную реакцию синтеза цепи (ПЦР) и в ней используют обратимо инактивированную термостабильную ДНК-полимеразу. Реакционную смесь инкубируют до проведения реакции амплификации при температуре выше, чем температура отжига в реакции амплификации. Таким образом, ДНК-полимеразу инактивируют до тех пор, пока температура остается выше той температуры, которая гарантирует специфичность реакции амплификации, тем самым снижая неспецифическую амплификацию.
Следующим предметом изобретения являются реакционные смеси для амплификации, которые содержат обратимо инактивированный термостабильный фермент по настоящему изобретению наряду с реагентами, необходимыми для проведения реакции амплификации. В предпочтительном примере осуществления реакционная смесь для амплификации содержит олигонуклеотидные праймеры для проведения ПЦР.
Еще одним предметом изобретения являются наборы, содержащие обратимо инактивированный термостабильный фермент по изобретению и один или несколько реагентов для амплификации.
На фиг. 1 показана структура цитраконового ангидрида, цис-аконитового ангидрида и 2,3-диметилмалеинового ангидрида и реакция между цитраконовым ангидридом и лизином.
На фиг. 2 показаны результаты амплификаций, проведенных с использованием цитраконилированной ДНК-полимеразы Taq, как описано в примере 4.
На фиг. 3 показаны результаты амплификаций, проведенных с использованием цитраконилированных ДНК-полимераз, как описано в примере 6.
На фиг. 4 показаны результаты изменения времени предреакционной инкубации при амплификациях, проведенных с использованием цитраконилированных и цис-аконитилированных ДНК-полимераз, как описано в примере 9.
На фиг. 5 показаны результаты изменения количества циклов амплификации при амплификациях, проведенных с использованием цитраконилированных и цис-аконитилированных ДНК-полимераз, как описано в примере 10.
Для лучшего понимания сущности изобретения ниже даются определения некоторых терминов.
Термины "нуклеиновая кислота" и "олигонуклеотид" относятся к праймерам, зондам и олигомерным фрагментам, подлежащим обнаружению, и должны включать полидезоксирибонуклеотиды (содержащие 2-дезокси-D-рибозу), полирибонуклеотиды (содержащие D-рибозу) и любой другой тип полинуклеотида, который представляет собой N-гликозид пуринового или пиримидинового основания или модифицированного пуринового или пиримидинового основания. Термины "нуклеиновая кислота" и "олигонуклеотид" не подразумевают различия в длине этих нуклеотидов и могут использоваться взаимозаменяемо. Эти термины относятся только к первичной структуре молекулы. Таким образом, эти термины включают двух- и одноцепочечную ДНК, а также двух- и одноцепочечную РНК. Олигонуклеотиды могут быть получены любым приемлемым способом. Обзор методов синтеза приведен у Goodchild, 1990, Bioconjugate Chemistry 1(3): 165-187.
Термин "гибридизация" относится к образованию двойной структуры в результате спаривания комплементарных оснований двух одноцепочечных нуклеиновых кислот. Гибридизация может происходить между полностью комплементарными цепями нуклеиновых кислот или между "практически комплементарными" цепями нуклеиновых кислот, которые содержат минорные области ошибочного спаривания. Условия, при которых будет происходить гибридизация только полностью комплементарных цепей нуклеиновых кислот, называются "жесткими условиями гибридизации" или "специфичными в отношении последовательности условиями гибридизации". Стабильные дуплексы практически комплементарных последовательностей могут быть получены в менее жестких условиях гибридизации. Специалисты в области технологии нуклеиновых кислот могут определить стабильность дуплекса эмпирически, анализируя различные параметры, в том числе, например, длину и концентрацию пар оснований олигонуклеотидов, ионную силу и частоту ошибочно спаренных пар оснований, следуя руководству в этой области (см., например, Sambrook и др., 1989, выше).
Обычно условия жесткой гибридизации выбирают так, чтобы они были приблизительно на 5oC ниже, чем термическая точка плавления (Тпл) для конкретной последовательности при определенных ионной силе и значении pH. Тпл представляет собой температуру (при определенных ионной силе и pH), при которой происходит разъединение 50% пар оснований. Снижение жесткости условий гибридизации будет допускать появление в некоторых пределах ошибочных спариваний последовательности; степень допустимых ошибочных спариваний может контролироваться путем соответствующей регулировки условий гибридизации.
Термин "праймер" относится к олигонуклеотиду естественного происхождения или синтетическому, способному действовать в качестве точки инициации синтеза ДНК в таких условиях, в которых индуцируется синтез продукта удлинения праймера, комплементарного цепи нуклеиновой кислоты, т.е. в присутствии четырех различных нуклеозидтрифосфатов и агента полимеризации (например, ДНК-полимеразы или обратной транскриптазы) в соответствующем буфере и при приемлемой температуре. Аналоги олигонуклеотидов, такие, как "пептидные нуклеиновые кислоты", могут действовать в качестве праймеров и подпадают под область значений термина "праймер", как он используется в контексте настоящего описания. Предпочтительно праймер представляет собой одноцепочечный олигодезоксирибонуклеотид. Приемлемая длина праймера зависит от предполагаемого применения праймера, но обычно составляет от 6 до 50 нуклеотидов. Для праймеров с короткими молекулами обычно требуются более низкие температуры для образования достаточно стабильных гибридных комплексов с матрицей. Нет необходимости в том, чтобы праймер отражал точную последовательность матричной нуклеиновой кислоты, но он должен быть в достаточной степени комплементарным, чтобы гибридизироваться с матрицей.
Термин "удлинение праймера", как он используется в контексте настоящего описания, относится как к синтезу ДНК, происходящему вследствие полимеризации отдельных нуклеозидтрифосфатов с использованием праймера в качестве точки инициации, так и к присоединению к праймеру дополнительных олигонуклеотидов с целью удлинения праймера. Термин "удлинение примера", как он используется в настоящем описании, включает лигирование (сшивание) двух олигонуклеотидов для образования более длинного продукта, который далее может служить в качестве мишени в последующих циклах амплификации. Термин "праймер", как он используется в настоящем описании, включает олигонуклеотиды, используемые в опосредуемых лигированием процессах амплификации, которые удлиняются с помощью лигирования второго олигонуклеотида, гибридизирующегося в соседнем положении.
Праймеры могут включать дополнительные элементы, способствующие обнаружению или иммобилизации праймера, но не изменяющие основное свойство праймера, а именно, его действие в качестве точки инициации синтеза ДНК. Например, праймеры могут содержать дополнительную полинуклеотидную последовательность на 5'-конце, которая не гибридизируется с нуклеиновой кислотой-мишенью, но которая облегчает клонирование амплифицированного продукта. Область праймера, комплементарная матрице в достаточной для гибридизации степени, обозначают в данном описании областью гибридизации.
Термины "область-мишень" и "нуклеиновая кислота-мишень" относятся к области или последовательности нуклеиновой кислоты, которая должна быть амплифицирована. Сайт праймерной гибридизации может рассматриваться как область-мишень для гибридизации праймера.
Олигонуклеотидный праймер, как это используется в настоящем описании, является "специфичным" в отношении последовательности-мишени, если число ошибочных спариваний, присутствующих между олигонуклеотидом и последовательностью-мишенью, меньше, чем число ошибочных спариваний, присутствующих между олигонуклеотидом и последовательностями, не являющимися мишенями, которые могут присутствовать в образце. Могут быть выбраны такие условия гибридизации, при которых стабильные дуплексы образуются только в том случае, если число имеющихся ошибочных спариваний не превосходит числа ошибочных спариваний, присутствующих между олигонуклеотидом и последовательностью-мишенью. При таких условиях олигонуклеотид может образовывать стабильный дуплекс только с последовательностью-мишенью. Таким образом, использование специфичных в отношении мишени праймеров в приемлемых жестких условиях амплификации дает возможность осуществлять специфическую амплификацию тех последовательностей-мишеней, которые содержат сайты, связывающие праймеры-мишени. Использование специфичных в отношении последовательности условий амплификации дает возможность осуществлять специфическую амплификацию тех последовательностей-мишеней, которые содержат сайты, связывающие строго комплементарные праймеры.
Термин "неспецифическая амплификация" относится к амплификации полинуклеотидных последовательностей, отличных от последовательности-мишени, которые образуются вследствие гибридизации праймеров с последовательностями, отличными от последовательности-мишени, и затем служат субстратом для удлинения праймера. Гибридизация праймера с последовательностью, не являющейся мишенью, рассматривается как "неспецифическая гибридизация" и может осуществляться при менее жестких предреакционных условиях, характеризующихся более низкой температурой.
Термин "термостабильный фермент" относится к ферменту, относительно стабильному при нагревании. Термостабильные ферменты могут выдерживать без необратимой потери активности инкубацию при высокой температуре, обычно превышающей 50oC, применяемую для удаления модифицирующих групп. Модифицированные термостабильные ферменты, применяемые в способах по настоящему изобретению, включают термостабильные ДНК-полимеразы и термостабильные лигазы.
Термин "термостабильная ДНК-полимераза" относится к ферменту, относительно стабильному при нагревании и катализирующему полимеризацию нуклеозидтрифосфатов для образования продуктов удлинения праймера, которые являются комплементарными одной из полинуклеотидных цепочек последовательности-мишени. Фермент инициирует синтез на 3'-конце праймера и продолжает его в направлении к 5'-концу матрицы до тех пор, пока не закончится синтез. Очищенные термостабильные ДНК-полимеразы описаны в патенте США 4889818; патенте США 5352600; патенте США 5079352; в заявках WO 91/09950; WO 92/03556; WO 92/06200; WO 92/06202; патенте США 5491086; в заявке WO 92/09689 и патенте США 5210036.
Фермент, "происходящий" из организма, в контексте настоящего описания относится к ферменту, который выделяют и очищают из организма, или к рекомбинантной форме фермента, который выделяют и очищают из организма, и этот термин включает ферменты, в которых аминокислотная последовательность была модифицирована с использованием методов молекулярной биологии.
Термин "обратимо инактивированный", как это используется в настоящем описании, относится к ферменту, который был инактивирован путем реакции с соединением, приводящей к ковалентной модификации (также включающей и химическую модификацию) фермента, при которой модифицирующее соединение может быть удалено при соответствующих условиях. Реакция, которая приводит к удалению модифицирующего соединения, не должна быть обратной к реакции модификации. В том случае, если существует реакция, которая приводит к удалению модифицирующего соединения и к восстановлению функции фермента, фермент рассматривают как обратимо инактивированный.
Термин "реакционная смесь" относится к раствору, содержащему реагенты, необходимые для проведения данной реакции. Термин "реакционная смесь для амплификации" относится к раствору, содержащему реагенты, необходимые для проведения реакции амплификации, и эта смесь обычно содержит олигонуклеотидные праймеры и ДНК-полимеразу или лигазу в приемлемом буфере. "Реакционная смесь для ПЦР" обычно содержит олигонуклеотидные праймеры, термостабильную ДНК-полимеразу, дНТФ и двухвалентный катион металла в приемлемом буфере. Реакционную смесь рассматривают как полную, если она содержит все реагенты, необходимые для проведения реакции, и как неполную, если она содержит только часть набора необходимых реагентов. Для специалиста в данной области техники очевидно, что для удобства, стабильности при хранении и для обеспечения независимого регулирования концентрации компонентов в зависимости от применения компоненты реакции обычно хранят в виде отдельных растворов, каждый из которых содержит часть полного набора компонентов, а затем реакционные компоненты объединяют перед реакцией для получения полной реакционной смеси.
Способы по настоящему изобретению включают проведение реакции амплификации с использованием активированного нагреванием термостабильного фермента, где активный фермент необходим для удлинения праймера. Перед высокотемпературной инкубацией, которая активирует фермент, реакционная смесь для амплификации не поддерживает удлинение праймера и в ней не образуются неспецифические или какие-либо другие продукты удлинения. После высокотемпературной инкубации, которая реактивирует фермент, реакцию амплификации поддерживают при повышенных температурах, которые обеспечивают специфичность реакции. Таким образом, продукты удлинения праймера образуются только в условиях, которые обеспечивают специфичность амплификации.
В способах по настоящему изобретению активированный нагреванием фермент в своем активном состоянии катализирует реакцию удлинения праймера. Для применения в обычной реакции амплификации, например, в ПЦР, активированный нагреванием термостабильный фермент в своем активном состоянии обладает ДНК-полимеразной активностью. Для применения в системах амплификации, опосредуемых лигазой, активированный нагреванием термостабильный фермент в своем активном состоянии обладает ДНК-лигазной активностью.
В системах амплификации, опосредуемых лигазой, "продукт удлинения" образуется путем лигирования первого олигонуклеотида (в настоящем описании подпадающего под термин "праймер") со вторым олигонуклеотидом, который гибридизируется вблизи 3'-конца первого олигонуклеотида. Второй олигонуклеотид может быть гибридизирован непосредственно вблизи праймера, в этом случае необходимо только лигирование, или может быть гибридизирован с одним или несколькими основаниями вдали от праймера, в этом случае необходима полимеразная активность для удлинения праймера перед лигированием. В любом случае соединение двух олигонуклеотидов, которые гибридизируются с соседними областями ДНК-мишени, рассматривается в настоящем описании как подпадающее под термин "удлинение праймера".
Обратимо инактивированные термостабильные ферменты по изобретению получают путем реакции между ферментом и модифицирующим реагентом, которая приводит к обратимой химической модификации фермента и в результате которой наблюдается полная или практически полная потеря активности фермента. Модификация заключается в ковалентном связывании модифицирующей группы с протеином. Модифицирующее соединение выбирают так, чтобы инкубация при повышенной температуре в буфере для реакции амплификации делала модификацию обратимой. Пригодные для этой цели ферменты и модифицирующие группы описаны ниже.
Обратимо инактивированные ферменты, обладающие в своем активном состоянии ДНК-полимеразной активностью, получают из термостабильных ДНК-полимераз. Применяемые в реакциях амплификации термостабильные ДНК-полимеразы хорошо известны в данной области техники и могут иметь происхождение из различных источников, таких, как некоторые виды термофильных эубактерий или архебактерий, относящиеся к родам Thermus, Thermogota, Thermococcus, Pyrodictium, Pyrococcus и Thermosipho. Характерные виды, из которых могут быть выделены термостабильные ДНК-полимеразы, пригодные для ПЦР-амплификации, включают Thermus aquaticus, Thermus thermophilus, Thermotoga maritima, Pyrodictium occultum, Pyrodictium abyssi и Thermosipho africanus. Термостабильные ДНК-полимеразы описаны в патенте США 4889818; патенте США 5352600; патенте США 5079352; Международных заявках WO 91/09950; WO 92/03556; WO 92/06200; WO 92/06202; патенте США 5491086; в заявке WO 92/09689 и патенте США 5210036. Термостабильные ДНК-полимеразы поставляются на рынок фирмой Perkin Elmer, Norwalk, CT.
Обратимо инактивированные термостабильные ферменты, пригодные для применения в других процессах амплификации, таких, как опосредуемые лигазой амплификации, получают из термостабильных ферментов, описанных в приведенных ниже ссылках, в которых представлены различные способы амплификации.
Способы по настоящему изобретению не ограничены применением ферментов, приведенных в качестве примеров. Например, любая описанная в литературе для применения в реакциях амплификации термостабильная ДНК-полимераза может быть модифицирована в соответствии с описанием настоящего изобретения для получения обратимо инактивированного фермента, пригодного для использования в настоящих способах. В целом любой фермент, который катализирует удлинение праймера или необходим для того, чтобы произошло удлинение праймера, и который является достаточно термостабильным, чтобы противостоять реактивации при высокотемпературной инкубации, не становясь при этом необратимо инактивированным, и который может быть модифицирован, как описано в настоящем изобретении, для получения обратимо инактивированного фермента, может быть применен в способах по настоящему изобретению. Специалист в данной области техники может оптимальным образом подобрать условия реакции модификации и реакции амплификации для любого конкретного фермента, руководствуясь настоящим описанием.
В предпочтительных примерах осуществления изобретения обратимую инактивацию термостабильного фермента выполняют с помощью обратимой блокады остатков лизина путем химической модификации ε--аминогруппы в остатках лизина. Модификация лизина в активной области протеина приводит к инактивации протеина. Кроме того, модификация лизина вне активной области может оказать влияние на инактивацию протеина посредством пространственного взаимодействия или конформационных изменений. В литературе описано большое количество соединений, которые взаимодействуют с аминогруппами обратимым образом. Например, аминогруппы обратимо модифицировали путем трифторацетилирования (см. Goldberger и Anfinsen, 1962, Biochemistry 1: 410), амидирования (см. Hunter и Ludwig, 1962, J. Amer. Chem. Soc. 84: 3491), малеилирования (см. Butler и др. , 1967, Biochem. J. 103: 78), ацетоацетилирования (см. Marzotto и др., 1967, Biochem. Biophys. Res. Commun. 26: 517; и Marzotto и др., 1968, Biochem. Biophys. Acta 154: 450), тетрафторсукцинилирования (см. Braunitzer и др. , 1968, Hoppe-Seyler's Z. Physiol. Chem. 349:265) и цитраконилирования (см. Dixon и Perham, 1968, Biochem. J. 109: 312-314; и Habeeb и Atassi, 1970, Biochemistry 9 (25): 4939-4944).
Предпочтительными реагентами для химической модификации ε--аминогруппы в остатках лизина являются ангидриды дикарбоновых кислот общей формулы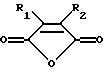
где R1 и R2 обозначают водород или органические радикалы, которые могут быть связаны, или общей формулы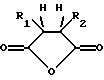
где R1 и R2 обозначают органические радикалы, которые могут быть связаны, и атомы водорода находятся в цис-положении. Органические радикалы могут быть непосредственно присоединены к кольцу через связь углерод-углерод или через связь углерод-гетероатом, такую, как связь углерод-кислород, углерод-азот или углерод-сера. Органические радикалы также могут быть связаны друг с другом с образованием циклической структуры, как, например, в случае 3,4,5,6-тетрагидрофталевого ангидрида.
Ангидриды дикарбоновых кислот взаимодействуют с аминогруппами протеинов, давая соответствующие ацилированные продукты, как показано на фиг. 1 для цитраконового ангидрида. Обратимость вышеуказанных ангидридов дикарбоновых кислот, как полагают, должна усиливаться в присутствии либо двойной связи цис-углерод-углерод, либо атомов водорода в цис-положении, что поддерживает концевую карбоксильную группу ацилированных остатков в пространственной ориентации, пригодной для взаимодействия с амидной группой и последующего деацилирования. Описание возможных механизмов реакций как ацилирования, так и деацилирования см. у Palacian и др., 1990, Mol. Cell. Biochem. 97: 101-111. Другие заместители могут аналогичным образом ограничивать вращение вокруг 2,3-связи ацильного остатка в ацилированном продукте, и можно ожидать, что такие соединения будут иметь функциональное значение в способах по настоящему изобретению.
Примеры предпочтительных реагентов включают малеиновый ангидрид; замещенные малеиновые ангидриды, такие, как цитраконовый ангидрид, цис-аконитовый ангидрид и 2,3-диметилмалеиновый ангидрид; экзо-цис-3,6-эндоксо- Δ4 -тетрагидрофталевый ангидрид и 3,4,5,6-тетрагидрофталевый ангидрид. Реагенты являются коммерчески доступными и поставляются, например, фирмами Aldrich Chemical Co. (Milwaukee, WI), Sigma Chemical Co. (St. Louis, MO) или Spectrum Chemical Mfg. Corp. (Gardena, CA). Модификации термостабильных ДНК-полимераз с использованием в качестве реагентов замещенных малеиновых ангидридов, в частности цитраконового ангидрида и цис-аконитового ангидрида, описаны в примерах.
Относительная стабильность ацилированных аминогрупп снижается в следующем порядке при использовании вышеуказанных реагентов: малеиновый ангидрид; экзо-цис-3,6-эндоксо-Δ4-тетрагидрофталевый ангидрид; цитраконовый ангидрид; 3,4,5,6-тетрагидрофталевый ангидрид; цис-аконитовый ангидрид; и 2,3-диметилмалеиновый ангидрид (см. Palacian и др., выше). Оптимальные условия ингибирующей активации для ферментов, модифицированных конкретным реагентом, определяют эмпирическим путем, как описано в примерах.
В патенте США 5262525 описаны способы химической модификации протеинов, в которых применяют соединения, представляющие собой ангидриды дикарбоновых кислот, полученные путем реакции Дильса-Альдера между малеиновым ангидридом и диеном. Соединения, описанные в указанном патенте, обладающие указанной в данном описании стабильностью, могут быть пригодны для настоящего изобретения.
Способы по настоящему изобретению не ограничены указанными в качестве примеров модифицирующими соединениями или модификацией протеина путем химической модификации остатков лизина. Любые описанные в литературе соединения, взаимодействие которых с протеином вызывает обратимую потерю всей или практически всей ферментативной активности, причем модификация обратима путем инкубации при повышенной температуре в буфере для реакции амплификации, являются пригодными для получения обратимо инактивированного фермента. При появлении новых соединений, обладающих способностью обратимо модифицировать протеины, они также могут оказаться пригодными для применения в способах по настоящему изобретению. Таким образом, соединения для получения модифицированных термостабильных ферментов по настоящему изобретению включают соединения, обладающие следующими свойствами, заключающимися в способности:
(1) к взаимодействию с термостабильным ферментом, который катализирует удлинение праймера, приводящему к значительной инактивации фермента;
(2) к инкубации образовавшегося модифицированного фермента в водном буфере при значении pH приблизительно 8-9 при температуре, приблизительно равной или ниже комнатной температуры (25oC), не приводящей к существенному увеличению ферментативной активности в течение промежутка времени менее приблизительно 20 минут; и
(3) к инкубации образовавшегося модифицированного термостабильного фермента в буфере для реакции амплификации, значение pH которого доведено до приблизительно 8-9 при комнатной температуре, при повышенной температуре, превышающей приблизительно 50oC, приводящей по крайней мере к двукратному увеличению ферментативной активности в течение промежутка времени менее приблизительно 20 минут.
Пригодность конкретного модифицирующего соединения может быть определена эмпирически традиционными способами, следуя указаниям, приведенным в настоящем описании. Экспериментальные методы для оценки вышеуказанных свойств, степени снижения ферментативной активности, являющейся результатом модификации протеина, и степени восстановления ферментативной активности в результате инкубации при повышенных температурах в реакционной смеси для амплификации описаны в примерах.
Получение обратимо инактивированных термостабильных ферментов
Химическая модификация остатков лизина в протеинах основана на способности ε -аминогруппы этого остатка вступать в реакцию в качестве нуклеофила. Не имеющая протона аминогруппа представляет собой реактивную форму, оптимальную для щелочного значения pH. Реакцию модификации проводят при значении pH от 8,0 до 9,0 в водном буфере при температуре, равной или ниже комнатной температуры (25oC). Взаимодействие является практически полным после инкубации в течение 12-24 часов. Пригодные условия взаимодействия известны в данной области техники и описаны ниже в примерах.
Ангидриды дикарбоновых кислот легко взаимодействуют с водой с получением соответствующих кислот. Следовательно, большая часть реагента гидролизуется во время модификации аминогрупп протеина. Скорость гидролиза возрастает при увеличении значения pH. Усиление гидролиза, происходящее при значениях pH, выше приблизительно 9, может привести к субоптимальному ацилированию протеина.
Обычно в реакции ацилирования используют молярный избыток модифицирующего агента по отношению к протеину. Оптимальное молярное отношение модифицирующего реагента к ферменту зависит от примененного реагента и его определяют эмпирическим путем. Так, например, ДНК-полимераза Taq практически полностью инактивируется (< 5% начальной активности) при взаимодействии с 20-кратным или более молярным избытком цитраконового ангидрида. Минимальное молярное отношение модификатора, приводящее к практически полной инактивации фермента, может быть определено при проведении реакции инактивации с серийными разбавлениями модифицирующего реагента, как описано в примерах.
В способах по настоящему изобретению не является необходимым, чтобы фермент был полностью инактивированным, а необходимо лишь, чтобы фермент был существенно инактивированным. Согласно настоящему описанию фермент является существенно инактивированным, если активность фермента после взаимодействия с модификатором составляет менее приблизительно 50% начальной активности. Уменьшение неспецифической амплификации может быть достигнуто с использованием существенно инактивированного фермента. Молярное отношение модификатора к ферменту в реакции может быть выбрано эмпирическим путем, следуя указаниям, приведенным в настоящем описании, так, чтобы оно приводило либо к практически полной инактивации, либо к существенной инактивации фермента. Приемлемые молярные отношения приведены в примерах. Соответствующие условия реакции для инактивации ферментов, не приведенные в примерах, могут быть определены традиционными экспериментальными способами, следуя указаниям, приведенным в настоящем описании.
Важной характеристикой ферментов по настоящему изобретению, инактивированных путем нагревания, является их стабильность при хранении. Обычно соединения, приведенные в данном описании, являются стабильными в течение продолжительного периода времени, что исключает необходимость их получения непосредственно перед каждым применением. Например, было установлено, что цитраконилированная ДНК-полимераза Taq оставалась инактивированной при хранении по крайней мере в течение четырех недель при 25oC. Рекомендуемые условия хранения варьируются в зависимости от того, какой модификатор применяют, но обычно препарат инактивированного фермента должен храниться при температуре, равной или ниже комнатной (25oC), предпочтительно замороженным. В частности менее стабильные модифицированные ферменты, такие, которые были модифицированы 2,3-диметилмалеиновым ангидридом, должны храниться замороженными.
Способы по настоящему изобретению включают применение реакционной смеси, содержащей обратимо инактивированный термостабильный фермент, и высокотемпературную инкубацию реакционной смеси до реакции амплификации или в ее процессе. Высокотемпературная инкубация приводит к деацилированию аминогрупп и восстановлению ферментативной активности.
Деацилирование модифицированных аминогрупп является результатом как увеличения температуры, так и сопутствующего понижения значения pH. Реакции амплификации проводят обычно в трис-HCl-буфере, значение pH которого доведено до 8,0-9,0 при комнатной температуре. При комнатной температуре для ацилированной формы аминогруппы благоприятна щелочная среда реакционного буфера. Хотя значение pH реакционного буфера доводят до 8,0-9,0 при комнатной температуре, pH реакционного трис-HCl-буфера понижается при увеличении температуры. Таким образом, значение pH реакционного буфера понижают при повышенных температурах, при которых проводят амплификацию и, в частности, при которых проводят активирующую инкубацию. Понижение значения pH реакционного буфера является благоприятным для деацилирования аминогрупп.
Изменение значения pH, происходящее вследствие высокотемпературных условий реакции, зависит от используемого буфера. Зависимость pH от температуры для различных буферов, применяемых в биологических реакциях, приведена у Good и др., 1966, Biochemistry 5(2): 467-477. Для трис-буфера изменение рКа, т.е. значения pH в средней точке диапазона буферного действия, связано с температурой следующим образом: Δ рКа/oC=-0,031. Например, у трис-HCl-буфера, приготовленного при 25oC, происходит падение рКа на 2,17 при увеличении температуры до 95oC для активирующей инкубации.
Хотя реакции амплификации обычно проводят в трис-HCl-буфере, реакции амплификации могут быть проведены в буферах, у которых происходит меньшее или большее изменение значения pH при изменении температуры. В зависимости от используемого буфера может потребоваться более или менее стабильный модифицированный фермент. Например, применение модифицирующего реагента, приводящего к получению менее стабильного модифицированного фермента, позволяет в достаточной мере восстановить ферментативную активность при меньших изменениях значений pH буфера. Эмпирическое сравнение относительной стабильности ферментов, модифицированных различными реагентами, как показано выше, определяет выбор модифицированного фермента, пригодного для применения в определенных буферах.
В способах по настоящему изобретению активацию модифицированного фермента достигают путем инкубации, проводимой при температуре, равной или более высокой, чем температура гибридизации (отжига) праймера, которая используется в реакции амплификации для обеспечения специфичности амплификации. Продолжительность инкубации, необходимая для восстановления ферментативной активности, зависит от температуры и pH реакционной смеси и от стабильности ацилированных аминогрупп фермента, что зависит от модифицирующего реагента, используемого для получения модифицированного фермента. Приемлем широкий диапазон условий инкубации; оптимальные условия для каждой реакции определяют эмпирическим путем. Обычно инкубацию проводят в буфере для реакции амплификации при температуре выше приблизительно 50oC в течение промежутка времени от 10 с до 20 мин. Не приведенная в примерах оптимизация условий инкубации для реактивации ферментов или для реакционных смесей может быть установлена традиционными экспериментальными способами, следуя указаниям, приведенным в настоящем описании.
В предпочтительном примере осуществления изобретения ПЦР-амплификацию проводят, применяя обратимо инактивированную термостабильную ДНК-полимеразу. Температура отжига, применяемая при ПЦР-амплификации, обычно составляет приблизительно 55-75oC, а предреакционную инкубацию проводят при температуре, равной или превышающей температуру отжига, предпочтительно при температуре выше 90oC. Предпочтительно реакционную смесь для амплификации инкубируют при приблизительно 90-100oC в течение промежутка времени, не превышающего приблизительно 12 мин, чтобы реактивировать ДНК-полимеразу до проведения температурных циклов. Пригодные предреакционные условия для обычных ПЦР-амплификаций описаны в примерах наряду с данными о воздействии на амплификацию различных условий предреакционной инкубации.
Первая стадия обычной ПЦР-амплификации состоит в тепловой денатурации двухцепочечной нуклеиновой кислоты-мишени. Точные условия, необходимые для денатурации образца нуклеиновой кислоты, зависят от длины и состава образца нуклеиновой кислоты. Обычно инкубация при 90-100oC в течение от приблизительно 10 с до приблизительно 4 мин является эффективной для полной денатурации образца нуклеиновой кислоты. Начальная стадия денатурации может служить в качестве предреакционной инкубации для реактивации ДНК-полимеразы. Однако в зависимости от продолжительности и температуры начальной стадии денатурации и от модификатора, используемого для инактивации ДНК-полимеразы, восстановление активности ДНК-полимеразы может оказаться неполным. Если требуется максимальное восстановление ферментативной активности, предреакционная инкубация может быть более продолжительной или, наоборот, может быть увеличено число циклов амплификации.
В предпочтительном примере осуществления изобретения модифицированный фермент и условия начальной денатурации выбирают так, чтобы только часть восстанавливаемой ферментативной активности восстанавливалась в течение начальной стадии инкубации. Последующие циклы ПЦР, каждый из которых включает высокотемпературную стадию денатурации, приводят к дальнейшему восстановлению ферментативной активности. Таким образом, активацию ферментативной активности приостанавливают в течение начальных циклов амплификации. Обнаружено, что это "время высвобождения" активности ДНК-полимеразы способствует дальнейшему снижению неспецифической амплификации. Известно, что избыток ДНК-полимеразы способствует неспецифической амплификации. В способах по настоящему изобретению существующий уровень ДНК-полимеразной активности низок в течение начальных стадий амплификации, когда число последовательностей-мишеней мало, что снижает количество образующихся неспецифических продуктов удлинения. Максимальная ДНК-полимеразная активность существует на более поздних стадиях амплификации, когда число последовательностей-мишеней велико, что позволяет достичь высокого выхода продукта амплификации. При необходимости число циклов амплификации может быть увеличено для компенсации более низкого уровня ДНК-полимеразной активности, имеющегося на начальных циклах. Влияние на амплификацию варьирования числа циклов амплификации показано в примерах.
Преимущество способов по настоящему изобретению состоит в том, что в них не требуется манипулирование с реакционной смесью после первоначального приготовления реакционной смеси. Таким образом, способы являются идеальными для применения в автоматизированных системах амплификации и в методах амплификации in situ, где добавление реагентов после начальной стадии денатурации или применение восковых барьеров является нежелательным или непрактичным.
Способы по настоящему изобретению особенно пригодны для снижения неспецифической амплификации в ПЦР. Однако изобретение не ограничено какой-либо конкретной системой амплификации. Обратимо инактивированные ферменты по настоящему изобретению могут быть использованы в любой основанной на применении праймера системе амплификации, в которой применяются термостабильные ферменты и в которой температура реакции является основой для достижения специфичности амплификации. Настоящие способы могут быть применимы к изотермическим системам амплификации, в которых используются термостабильные ферменты. Для восстановления ферментативной активности требуется только кратковременная инкубация при повышенной температуре. После того, как реакционную смесь подвергают высокотемпературной инкубации для восстановления ферментативной активности, реакцию проводят при соответствующей температуре реакции.
Другие, кроме ПЦР (патенты США 4683195; 4683202; и 4965188), способы амплификации включают, но не ограничены ими, следующие способы: лигазная реакция синтеза цепи (Ligase Chain Reaction = LCR, Wu и Wallace, 1989, Genomics 4:560-569 и Barany, 1991, Proc. Natl. Acad. Sci. США, 88: 189-193); полимеразно-лигазная реакция синтеза цепи (Barany, 1991, PCR Methods and Applic. 1: 5-16); "брешь"-LCR (Gap-LCR) (публикация PCT WO 90/01069); реакция репарации цепи (публикация Европейского патента 439182-А2), 3SR (Kwoh и др., 1989, Proc. Natl. Acad. Sci. США, 86: 1173-1177); Guatelly и др., 1990, Proc. Natl. Acad. Sci. США, 87: 1874-1878; публикация PCT WO 92/08800) и NASBA (патент США 5130238). Настоящее изобретение не ограничено какой-либо конкретной системой амплификации. В случае разработки других систем эти системы могут оказаться пригодными для использования на практике настоящего изобретения. Современный обзор систем амплификации был опубликован Abramson и Myers, 1993, Current Opinion in Biotechnology 4: 41-47.
Способы приготовления образцов, пригодные для каждой реакции амплификации, описаны в данной области техники (см., например, Sambrook и др., выше, и ссылки, указанные выше, где описываются способы амплификации). Простые и быстрые способы приготовления образцов для ПЦР-амплификации последовательностей-мишеней описаны у Higuchi, 1989, в PCR Technology (Erlich ред., Stockton Press, New York) и в PCR Protocols, главы 18-20 (Innis и др., ред., Academic Press, 1990). Специалист в данной области техники способен выбрать и эмпирически оптимизировать соответствующий протокол.
Способы обнаружения продуктов амплификации широко представлены в литературе. Стандартные способы включают анализ с использованием гель-электрофореза или гибридизацию с олигонуклеотидными зондами. Обнаружение гибридов, образованных между зондами и амплифицированной нуклеиновой кислотой, может быть выполнено различными методами, включающими дот-блоттинг-анализ и обратный дот-блоттинг-анализ. (См. Saiki и др., 1986, Nature 324: 163-166; Saiki и др. , 1989, Proc. Natl. Acad. Sci. США 86: 6230; публикация РСТ WO 89/11548; патенты США 5008182 и 5176775; PCR Protocols: A Guide to Methods and Applications (Innis и др., ред., Academic Press, San Diego, CA): 337-347). Способы обратного дот-блоттинга с использованием микролуночных планшетов описаны в находящейся в процессе одновременного рассмотрения заявке на патент США, N 141355 (соответствует заявке на Европейский патент ЕР-А-420260); патенте США 5232829; у Loeffelholz и др., 1992, J. Clin. Microbiol. 30(11): 2847-2851; Mulder и др., 1994, J. Clin. Microbiol. 32(2): 292-300; и Jackson и др., 1991, AIDS 5: 1463-1467.
Другой пригодный метод оценки, названный 5'-нуклеазным анализом, описан в патенте США 5210015 и у Holland и др., 1991, Proc. Natl. Acad. Sci. США 88: 7276-7280. При 5'-нуклеазном анализе меченые зонды разлагают одновременно с удлинением праймера с помощью экзонуклеазной активности в направлении 5' ---> 3' ДНК-полимеразы, например, ДНК-полимеразы Taq. Обнаружение продукта разложения зонда свидетельствует как о том, что произошла гибридизация между зондом и ДНК-мишенью, так и о том, что произошла реакция амплификации. В патенте США 5491063 (соответствует ЕР-А-699768) и ЕР-А-713921 описаны улучшенные способы обнаружения разложения зонда, которое происходит одновременно с амплификацией.
Другой способ обнаружения амплификации нуклеиновой кислоты путем контроля за увеличением общего количества двухцепочечных ДНК в реакционной смеси описан у Higuchi и др., 1992, Bio/Technology 10: 413-417; Higuchi и др., 1993, Bio/Technology 11: 1026-1030; и в опубликованных Европейских патентах 487218 и 512334. Обнаружение двухцепочечной ДНК-мишени основано на увеличении флуоресценции, которую испускают бромид этидия (EtBr) и другие ДНК-связывающие метки, когда они связаны с двухцепочечной ДНК. Увеличение двухцепочечных ДНК, происходящее в результате синтеза последовательностей-мишеней, приводит к увеличению флуоресценции, которое может быть обнаружено. Проблема этого метода состоит в том, что синтез последовательности, не являющейся мишенью, т.е. неспецифическая амплификация, приводит к увеличению флуоресценции, что мешает измерению увеличения флуоресценции, происходящей в результате синтеза последовательностей-мишеней. Таким образом, способы по настоящему изобретению являются особенно полезными, поскольку они снижают неспецифическую амплификацию, минимизируя тем самым увеличение флуоресценции, происходящее в результате амплификации последовательностей, не являющихся мишенями.
Настоящее изобретение также относится к наборам, многоконтейнерным блокам, содержащим соответствующие компоненты для практического осуществления настоящего способа. Необходимый набор содержит обратимо инактивированный термостабильный фермент и один или несколько реагентов для проведения реакции амплификации, таких, как олигонуклеотидные праймеры, нуклеозидтрифосфат в качестве субстрата, кофакторы и соответствующий буфер.
Представленные ниже примеры осуществления настоящего изобретения приведены только для иллюстрации и не ограничивают объем изобретения. Обычным специалистам в данной области техники после ознакомления с предыдущим описанием и последующими примерами должны быть очевидны многочисленные варианты осуществления изобретения, подпадающие под объем пунктов формулы изобретения, которые приведены после примеров.
Пример 1
Определение полимеразной активности
Все измерения ДНК-полимеразной активности, описанные ниже в примерах, проводили с использованием следующего метода определения ДНК-полимеразной активности. Метод практически соответствует таковому, описанному у Lawyer и др. , 1989, J. Biol. Chem. 264:6427-6437 и в инструкции к ДНК-полимеразному продукту AmpliTaq® (фирма Perkin Elmer, Norwalk, CT), оба документа включены в настоящее описание в качестве ссылки.
За единицу ферментативной активности принимают количество, равное 10 нмолям дНТФ, которое может включить нерастворимый в кислоте материал в течение 30 минут при 10-минутной инкубации при 74oC. Вследствие лабильности модифицированных ферментов активность измеряли при 50oC и приводили к стандартному раствору ДНК-полимеразы Taq, который также анализировали при 74oC. Реакции проводили в объеме 50 мкл, содержащем следующие реагенты:
25 мМ ТАПС (натриевая соль трис-(гидроксиметил)метиламинопропан-сульфоновой кислоты), pH 9,3 (при комнатной температуре);
50 мМ KCl;
2 мМ MgCl2;
1 мМ β -меркаптоэтанол;
20 мкМ каждого из дАТФ, дГТФ и дТТФ;
100 мкМ [α -32P] - дЦТФ (0,05-0,1 Ки/нмоль);
активированная ДНК спермы лосося.
Пример 2
Цитраконилирование ДНК-полимеразы Taq
В данном примере описана модификация ДНК-полимеразы Taq с помощью цитраконового ангидрида. Измерения активности цитраконилированной ДНК-полимеразы Taq, показывающие молярное отношение модификатора к ферменту в реакции инактивации, необходимое для достижения полной инактивации ДНК-полимеразной активности, описаны ниже в примере 3.
ДНК-полимеразу Taq (AmpliTaq® Perkin Elmer, Norwalk, CT) применяли в исходной концентрации 1,3 мг/мл. В первых экспериментах ДНК-полимеразу Taq сначала подвергали диализу в 1000-кратном избытке по объему 0,1М натрий-боратного буфера при pH 8,63. Было обнаружено, что эта стадия не является решающей и в последующих экспериментах ДНК-полимеразу Taq использовали непосредственно в трис-буфере (50 мМ трис-HCl, 1 мМ ЭДТК, 65 мМ KCl, pH 7,5), не подвергая диализу в растворе бората натрия.
Цитраконовый ангидрид (11,06 М) является коммерчески доступным (Aldrich, Milwaukee, WI). Исходный раствор цитраконового ангидрида готовили путем 100-кратного разбавления 11,06М цитраконового ангидрида в ДМФ (N,N-диметилформамиде).
В одной из серий реакций модификации серийные разбавления раствора цитраконового ангидрида готовили путем повторных двукратных разбавлений в ДМФ. Для каждого раствора в сериях 4 мкл разбавленного раствора цитраконового ангидрида добавляли к 400 мкл раствора ДНК-полимеразы Taq (после диализа в борате натрия), получая растворы, содержащие молярные отношения цитраконового ангидрида к ДНК-полимеразе Taq, приблизительно равные 80/1, 40/1, 20/1 и 10/1. Растворы инкубировали в течение ночи при 4oC для того, чтобы инактивировать ДНК-полимеразу Taq. В настоящем описании фермент, который был модифицирован в реакции с N-кратным молярным избытком модификатора, обозначают как NX-фермент. Таким образом, полученные цитраконилированные ДНК-полимеразы Taq в настоящем описании обозначают как ДНК-полимеразы Taq 80Х, 40Х, 20Х и 10Х.
Дополнительные реакции модификации проводили с использованием молярных отношений цитраконового ангидрида к ДНК-полимеразе Taq (без диализа в борате натрия) приблизительно 80Х, 160Х и 240Х. Отношения 160Х и 240Х получали путем соответствующего подбора исходного разбавления 11,06М цитраконового ангидрида в ДМФ (N,N-диметилформамиде). Например, для конечного отношения 160Х цитраконовый ангидрид (11,06М) разбавляли ДМФ в отношении 1/50 и 4 мкл образовавшегося исходного раствора цитраконового ангидрида добавляли к 400 мкл раствора ДНК-полимеразы Taq. Образовавшиеся цитраконилированные ДНК-полимеразы Taq в настоящем описании обозначают как ДНК-полимеразы Taq 240Х, 160Х и 80Х.
Пример 3
Инактивация и восстановление при нагревании ДНК-полимеразной активности при использовании цитраконового ангидрида
В этом эксперименте представлены данные об измерении активности цитраконилированных ДНК-полимераз Taq из примера 2 как до, так и после реактивации цитраконилированной ДНК-полимеразы Taq путем инкубации при нагревании. Оценивали влияние pH на уровень активности, восстановленной вследствие реактивации при нагревании цитраконилированных ДНК-полимераз Taq.
Образцы цитраконилированной ДНК-полимеразы Taq разбавляли в отношении 1/200 буфером, состоящим из 10 мМ трис-HCl, 100 мМ KCl, 2 мМ MgCl2, 0,5% твина 20, 0,5% NP-40, 16% глицерина. При комнатной температуре значение pH буфера было равно 8,25. Разбавленные образцы цитраконилированной ДНК-полимеразы Taq инкубировали при 90oC в течение 20 мин или выдерживали при комнатной температуре в качестве контроля. После обработки образцы разбавляли в отношении 1/5 буфером для разбавления фермента (25 мМ трис-HCl, 50 мМ KCl, 1 мМ β -меркаптоэтанол, 0,5% твина 20 (товарный знак TweenTM 20), 0,5% NP-40, 0,1% желатина) и определяли активность, как указано в примере 1. Данные об активности ДНК-полимеразы после обработки представлены ниже. Молярное отношение обозначает примененное в реакции модификации молярное отношение цитраконового ангидрида к ДНК-полимеразе Taq. Каждое значение активности представляет собой среднее по двум повторностям, измеренным для двух одинаковых образцов (см. табл. 1).
Полную инактивацию ДНК-полимеразы Taq получали, используя более чем 20-кратный молярный избыток цитраконового ангидрида. После инкубации полностью инактивированной ДНК-полимеразы Taq при 90oC в течение 20 мин восстановилось как минимум 16% активности.
Хотя при использовании цитраконилированной ДНК-полимеразы Taq 40Х восстанавливалось больше ферментативной активности, чем при использовании цитраконилированной ДНК-полимеразы Taq 80Х, с практической точки зрения вероятно лучше использовать в коммерческом наборе цитраконилированную ДНК-полимеразу Taq 80Х (или с большим избытком модификатора) с тем, чтобы получать большую устойчивость при изготовлении.
Аналогичные эксперименты проводили с использованием буфера, значение pH которого доводили до 7,75 при комнатной температуре. Результаты представлены в табл. 2.
Количество восстановленной ДНК-полимеразной активности оказалось большим при более низком значении pH.
Активности ДНК-полимераз Taq 80Х и 160Х (без диализа в борате натрия) измеряли до и после реактивации путем инкубации при нагревании практически так же, как описано выше. Значение pH при комнатной температуре использованного для инкубации буфера составляло 8,0. Результаты приведены в табл. 3. Каждое значение активности представляет собой среднее по двум повторностям, измеренным для двух одинаковых образцов.
Влияние значения pH на реактивацию может быть оценено, кроме того, путем сравнения активности, восстановленной у ДНК-полимераз Taq 80Х, в трех процессах реактивации при разных значениях pH. Приведенные выше данные свидетельствуют о том, что ферментативная активность восстанавливается даже, когда в реакции модификации используют большой молярный избыток модификатора.
Пример 4
ПЦР-амплификация с применением цитраконилированных ДНК-полимераз Taq
В этом примере представлены данные об использовании в ПЦР-амплификации цитраконилированной термостабильной ДНК-полимеразы Taq, описанной в примере 2.
ПЦР-протокол
Амплификацию проводили, используя разбавления в отношениях 1/20, 1/40 и 1/80 модифицированной ДНК-полимеразы Taq 240Х, описанной в примере 2. Разбавления проводили с использованием буфера, состоящего из 20мМ трис-HCl, pH 8,0 (при комнатной температуре), 100 мМ KCl, 0,1 мМ ЭДТК (этилендиаминтетрауксусная кислота), 1 мМ ДТТ (дитиотреитол), 50% глицерина, 0,5% твина-20, 0,5% Nonidet Р40 (буфер для хранения AmpliTaq® , Perkin Elmer, Norwalk, CT). Для сравнения амлификации также проводили, используя разбавления в отношениях 1/10, 1/20, 1/40 и 1/80 немодифицированной ДНК-полимеразы Taq.
Клонированную геномную последовательность HTLV-I (вирус Т-клеточной лимфомы человека) амплифицировали с использованием примеров SK432 и SK111. Последовательности праймеров приведены в патенте США 5418149, включенном в настоящее описание в качестве ссылки. ПЦР проводили в реакционном объеме 100 мкл при следующих условиях реакции.
Реакционная смесь:
30 копий матрицы ДНК HTLV-I;
10 мМ трис, pH 8,3;
50 мМ KCl;
по 0,5 мкМ каждого праймера;
200 мкМ дАТФ, дЦТФ и дГТФ;
400 мкМ дУТФ;
0,5 мкл раствора цитраконилированной ДНК-полимеразы Taq;
2,5 мМ MgCl2;
1 нг вмДНК;
1 ед. урацил-ДНК-гликозилазы (UNG) (Perkin Elmer, Norwalk, CT).
Термический профиль циклов см. в табл. 4.
Амплифицированные продукты анализировали при помощи электрофореза на 4%-ной агарозе с применением подвижного буфера IX ТБЭ (0,089М трис, 0,089М борная кислота, 0,0025М двунатриевая соль ЭДТК). Электрофорез проводили при 100 В в течение приблизительно 2 часов. Для окраски любой присутствующей ДНК после электрофореза добавляли бромид этидия (0,5 мкг/мл). Гель обесцвечивали в 1X ТБЭ и окрашенные бромидом этидия зоны, соответствующие ДНК, визуализировали с использованием ультрафиолетового облучения.
Результаты
Результаты представлены на фиг. 2. Обозначена зона, соответствующая амплифицированной последовательности-мишени. Проявленные на геле зоны, отличные от зоны, соответствующей последовательности-мишени, соответствуют продуктам, образовавшимся в результате неспецифической амплификации последовательностей, не являющихся мишенями. Влияние амплификации с использованием цитраконилированной ДНК-полимеразы Taq (меченной по Taq, HS) можно увидеть при сравнении строения зон и интенсивности каждой полосы. Поскольку амплификация неспецифических продуктов конкурирует с амплификацией последовательности-мишени, увеличение амплификации последовательности-мишени, кроме того, свидетельствует о степени уменьшения неспецифической амплификации. Таким образом, изменение в относительном содержании продуктов в каждой полосе наилучшим образом свидетельствует о влиянии предреакционных обработок на неспецифическую амплификацию.
Амплификации с использованием немодифицированной ДНК-полимеразы Taq в основном привели к получению продукта неспецифической амплификации. Применение цитраконилированной ДНК-полимеразы Taq привело к существенному увеличению интенсивности зоны, соответствующей амлифицированной последовательности-мишени, и существенному снижению интенсивности зон, соответствующих продуктам неспецифической амплификации. Данные свидетельствуют о том, что ПЦР-амплификация с применением обратимо инактивированной ДНК-полимеразы существенно снижает неспецифическую амплификацию и существенно увеличивает количество нужной амплифицированной последовательности-мишени.
Пример 5
Другие цитраконилированные термостабильные ДНК-полимеразы
В этом примере представлены данные о цитраконилировании нескольких других термостабильных ДНК-полимераз в дополнение к ДНК-полимеразе Taq, описанной выше. Модифицировали следующие термостабильные ДНК-полимеразы:
1) термостабильную ДНК-полимеразу из Thermus thermophilus (rTth, Perkin Elmer, Norwalk, CT), как описано в заявке WO 91/09950, включенной в настоящее описание в качестве ссылки;
2) мутантную термостабильную ДНК-полимеразу из Thermatoga maritima (UlTmaTM, Perkin Elmer, Norwalk, CT), как описано в заявке WO 92/03556, включенной в настоящее описание в качестве ссылки;
3) мутантную форму термостабильной ДНК-полимеразы из Thermus aquaticus, лишенную экзонуклеазной активности в направлении 3'--->5', как описано в заявке WO 92/06200, включенной в настоящее описание в качестве ссылки. Этот фермент обозначен в настоящем описании как CS Taq или CS AmpliTaq®.
Для каждой из вышеуказанных трех ДНК-полимераз готовили исходный раствор с концентрацией приблизительно 200 ед./мкл. Для сравнения, концентрация 1,3 мг/мл ДНК-полимеразы Taq, используемой в предыдущих примерах, приблизительно эквивалентна 260 ед./мкл. Каждую ДНК-полимеразу модифицировали практически так же, как описано выше в примере 2. Десять мкл цитраконового ангидрида разбавляли 500 мкл ДМФ. Затем 10 мкл разбавленного цитраконового ангидрида объединяли с 1000 мкл раствора каждого фермента. Образовавшиеся растворы инкубировали в течение ночи при 4oC.
Пример 6
ПЦР-амплификация с применением цитраконилированных ДНК-полимераз
В этом примере представлены данные об использовании в ПЦР-амплификациях цитраконилированных термостабильных ДНК-полимераз, описанных в примерах 2 и 5.
ПЦР-протокол
Амплификации проводили, применяя разбавленнные растворы модифицированных ДНК-полимераз. Разбавления проводили с использованием буфера, состоящего из 20 мМ трис-HCl, pH 8,0 (при комнатной температуре), 100 мМ KCl, 0,1 мМ ЭДТК (этилендиаминтетрауксусная кислота), 1 мМ ДТТ (дитиотреитол), 50% глицерина, 0,5% твина (товарный знак TweenTM 20), 0,5% Nonidet P40 (буфер для хранения AmpliTaq® , Perkin Elmer, Norwalk, CT).
Геномную последовательность ВИЧ-1 амплифицировали с использованием праймеров SK145 и SK431 (Perkin Elmer, Norwalk, CT). Праймеры SK145 и SK431 описаны в патенте США 5481149 и в следующих научных публикациях: SK145 описан у Kwok и др., 1990, Nucleic Acids Res. 18: 999-1005; SK431 описан у Jackson и др., 1991, AIDS 5: 1463-1467. ПЦР осуществляли в реакционном объеме 100 мкл при следующих условиях реакции.
Реакционная смесь:
100 копий матрицы ДНК ВИЧ-1;
10 мМ трис, pH 8,3;
50 мМ KCl;
по 0,5 мкМ каждого праймера;
200 мкМ дАТФ, дЦТФ и дГТФ;
400 мкМ дУТФ;
0,5 мкл раствора ДНК-полимеразы;
2,5 мМ MgCl2;
1 мкг вмДНК;
1 ед. урацил-ДНК-гликозилазы (UNG) (Perkin Elmer, Norwalk, CT).
Термический профиль циклов см. в табл. 5.
Стадия предреакционной инкубации также служит в качестве начальной стадии денатурации. Стадию начальной денатурации обычно применяют в типичной реакции амплификации, для того чтобы гарантировать полноту денатурации двухцепочечной мишени. Каждый цикл ПЦР начинается со стадии денатурации при 94oC в течение 1 мин. Таким образом, сразу же после начальной предреакционной инкубации, проводимой при 95oC в течение 12 мин, реакционную смесь инкубируют при 94oC в течение 1 мин во время стадии денатурации в первом цикле.
Амплифицированные продукты анализировали при помощи гель-электрофореза на агарозе (100 мл 3%-ного NuSieve® и 0,5%-ного SeaChem®) с применением подвижного буфера 1X ТБЭ (0.089М трис, 0.089М борная кислота, 0.0025М двунатриевая соль ЭДТК). Электрофорез проводили при 100 В в течение приблизительно 1 часа. Для окраски любой присутствующей ДНК после электрофореза добавляли бромид этидия (0,5 мкг/мл). Гель обесцвечивали в ТБЭ и окрашенные бромидом этидия зоны, соответствующие ДНК, визуализировали с использованием ультрафиолетового облучения.
Амплификации с использованием цитраконилированных ДНК-полимераз
Цитраконилированные ДНК-полимеразы, разбавленные в отношениях 1/10, 1/20, 1/40 и 1/80, описанные в примере 5, и цитраконилированную ДНК-полимеразу Taq 240Х, описанную в примере 2, применяли при амплификациях нуклеиновой кислоты ВИЧ-1 и амплифицированные продукты анализировали при помощи электрофореза на агарозе. Для сравнения проводили амплификации с использованием разбавлений немодифицированной ДНК-полимеразы Taq.
Результаты представлены на фиг. 3. Обозначена зона, соответствующая амплифицированной последовательности-мишени. Проявленные на геле зоны, отличные от зоны, соответствующей амплифицированной последовательности-мишени, соответствуют продуктам, образовавшимся в результате неспецифической амплификации последовательностей, не являющихся мишенями. Влияние амплификации с использованием цитраконилированной ДНК-полимеразы можно увидеть при сравнении строения зон и интенсивности в пределах каждой из полос. Поскольку амплификация неспецифических продуктов конкурирует с амплификацией последовательности-мишени, увеличение амплификации последовательности-мишени, кроме того, свидетельствует об уменьшении неспецифической амплификации. Таким образом, изменение в относительном содержании продуктов в пределах каждой из полос наилучшим образом свидетельствует о влиянии предреакционных обработок на неспецифическую амплификацию.
Амплификации с использованием немодифицированной ДНК-полимеразы Taq для разбавленных ферментов в основном привели к амплификации неспецифического продукта. Применение цитраконилированной ДНК-полимеразы Taq привело к увеличению интенсивности зоны, соответствующей амплифицированной последовательности-мишени для всех разбавлений, кроме 1/80, и к существенному уменьшению интенсивности зоны, соответствующей продукту неспецифической амплификации. Данные свидетельствуют о том, что ПЦР-амплификация с применением обратимо инактивированной ДНК-полимеразы существенно уменьшает неспецифическую амплификацию и существенно увеличивает количество нужной амплифицированной последовательности-мишени.
Применение цитраконилированных ДНК-полимераз U1Tma, CS Taq и rTth также привело к увеличению образования продукта специфической амплификации по сравнению с амплификациями, в которых использовали немодифицированную ДНК-полимеразу Taq, наряду с сопутствующим уменьшением количества продукта неспецифической амплификации. Результаты показывают, что способы по настоящему изобретению применимы в целом к термостабильным ДНК-полимеразам.
Следует отметить, что хотя результаты свидетельствуют о функциональности настоящего изобретения, результаты, полученные с применением цитраконилированных ДНК-полимераз Taq, U1Tma, CS Taq и rTth, не могут непосредственно сравниваться, поскольку условия их модификации не сравнимы, они оптимизировались для каждой ДНК-полимеразы. В частности, хотя каждый исходный раствор ДНК-полимеразы содержал одно и то же количество ед./мл, молярность растворов не определяли и, следовательно, не определяли молярный избыток цитраконового ангидрида в каждой реакции модификации. Для специалиста в данной области техники очевидно, что оптимальные условия модификации могут быть определены эмпирическим путем с использованием представленных в настоящем описании протоколов.
Пример 7
Цис-аконитилированная ДНК-полимераза
В этом примере описана модификация ДНК-полимеразы Taq с использованием цис-аконитового ангидрида.
Модификацию с использованием цис-аконитового ангидрида проводили практически так же, как описано в примере 2, а основное отличие состоит в том, что цис-аконитовый ангидрид продается в виде порошка, а не жидкости. ДНК-полимеразу Taq (AmpliTaq®, Perkin Elmer, Norwalk, CT) в трис-буфере (50 мМ трис-HCl, 1 мМ ЭДТК, 68мМ HCl, pH 7,5) применяли в исходной концентрации 1,3 мг/мл. Исходный раствор цис-аконитового ангидрида (Aldrich, Milwaukee, WI) готовили, растворяя 20 мг цис-аконитового ангидрида в 1 мл 100%-ного EtOH.
Добавляли 10 или 20 мкл раствора цис-аконитового ангидрида к 1000 мкл раствора ДНК-полимеразы Taq, получая растворы, содержащие цис-аконитовый ангидрид и ДНК-полимеразу Taq в молярных отношениях приблизительно 90/1 и 180/1. Растворы инкубировали в течение ночи при 4oC для того, чтобы инактивировать ДНК-полимеразу Taq.
Сравнения амплификаций с применением цис-аконитилированной ДНК-полимеразы Taq и амплификаций с применением цитраконилированной ДНК-полимеразы Taq приведены ниже.
Пример 8
Инактивация и восстановление при нагревании ДНК-полимеразной активности при использовании цис-аконитового ангидрида
Уровни активности цис-аконитилированных ДНК-полимераз Taq 90Х и 180Х, полученных в примере 7, определяли до и после реактивации путем инкубации при нагревании, практически так же, как описано выше в примере 3. Значение pH во время инкубаций составляло 8,0. Полученные результаты приведены в табл. 6. Каждое значение активности представляет собой среднее по двум повторностям, измеренным для двух одинаковых образцов.
Сравнение результатов с таковыми, полученными в примере 3, показывает, что цис-аконитилированнный продукт более легко восстанавливается, чем цитраконилированный. Для полной инактивации ДНК-полимеразы необходим больший молярный избыток цис-аконитового ангидрида и после высокотемпературной инкубации восстанавливается более высокий уровень активности. Причина, по которой измеренный уровень активности после модификации с 90-кратным молярным избытком цис-аконитового ангидрида превышает 100%, неизвестна, и это может быть следствием неточного измерения активности или может отражать фактическую модификацию ДНК-полимеразы.
Пример 9
Влияние продолжительности предреакционной инкубации
В этом примере описано влияние продолжительности предреакционной инкубации на количество получаемого продукта.
Амплификации проводили с использованием цитраконилированных и цис-аконитилированных ДНК-полимераз Taq, полученных как описано выше. Для каждого набора условий амплификации применяли ДНК-полимеразы Taq, модифицированные с использованием 80- и 160-кратного молярного избытка цитраканового ангидрида и 90- и 180-кратного молярного избытка цис-аконитового ангидрида. Амплификации проводили, используя модельную систему ВИЧ-1, описанную выше в примере 6, за исключением того, что изменяли условия предреакционной инкубации. Для каждого препарата фермента амплификации проводили с предреакционной инкубацией в течение 12, 6, 3 или 0 мин.
Как отмечено выше, стадия предреакционной инкубации также служит в качестве стадии начальной денатурации. Каждый цикл ПЦР начинается со стадии денатурации. Сразу же после начальной предреакционной инкубации, проводимой при 95oC в течение 12, 6, 3 или 0 мин, каждую реакционную смесь инкубируют при 94oC в течение 1 мин на стадии инкубации первого цикла. Таким образом, даже когда применяли предреакционную инкубацию, ферментативная активность восстанавливалась в течение стадии денатурации начальных циклов.
Продукты амплификации анализировали с помощью гель-электрофореза на агарозе. Результаты представлены на фиг. 4. Обозначена зона, соответствующая амплифицированной последовательности-мишени. Проявленные на геле зоны, отличные от зоны, соответствующей последовательности-мишени, соответствуют продуктам, образовавшимся в результате неспецифической амплификации последовательностей, не являющихся мишенями.
Результаты свидетельствуют о том, что при использовании цис-аконитилированной ДНК-полимеразы при всех изученных продолжительностях предреакционной инкубации получали выраженную зону, соответствующую амплифицированному продукту. Амплификации, проведенные без предреакционной инкубации, приводили к получению приблизительно такого же количества продукта, как и амплификации, в которых применяли продолжительные предреакционные инкубации.
И, наоборот, результаты показывают, что при использовании цитраконилированной ДНК-полимеразы, была необходима предреакционная инкубация по крайней мере в течение 3 минут, для того чтобы получить максимальное количество амплифицированного продукта. Амплификации, проведенные без предреакционной инкубации, приводили к получению существенно меньшего количества амплифицированного продукта. Результаты свидетельствуют о том, что активность цис-аконитилированных ДНК-полимераз восстанавливается быстрее, чем цитраконилированных ДНК-полимераз.
Пример 10
Влияние количества циклов
Этот пример описывает роль увеличения количества циклов амплификации для компенсации короткого времени предреакционной инкубации.
Амплификации проводили, применяя ДНК-полимеразы Taq, модифицированные с использованием 80- и 160-кратного молярного избытка цитраконового ангидрида и 90- и 180-кратного молярного избытка цис-аконитового ангидрида. Амплификации проводили практически так же, как описано выше, используя модельную систему ВИЧ-1, описанную в примере 6, за исключением того, что изменяли начальные условия предреакционной инкубации и количество циклов. Для каждого препарата фермента амплификации проводили при следующих условиях:
Предреакционная инкубация - Кол-во циклов амплификации
12 минут, 80oC - 60
0 - 60
0 - 48
0 - 43
0 - 39
Продукты амплификации анализировали с помощью гель-электрофореза на агарозе. Результаты представлены на фиг. 5. Результаты показывают, что увеличение количества циклов амплификации может компенсировать уменьшение эффективности амплификации в результате неполной реактивации активности ДНК-полимеразы, когда не применяют предреакционную инкубацию.
Для каждой ДНК-полимеразы увеличение количества циклов амплификации привело к увеличению количества амплифицированного продукта. Эффект оказался наименьшим, когда использовали цис-аконитилированную ДНК-полимеразу Taq, для которой, как было показано в примере 9, требуется непродолжительная или не требуется вообще предреакционная инкубация.






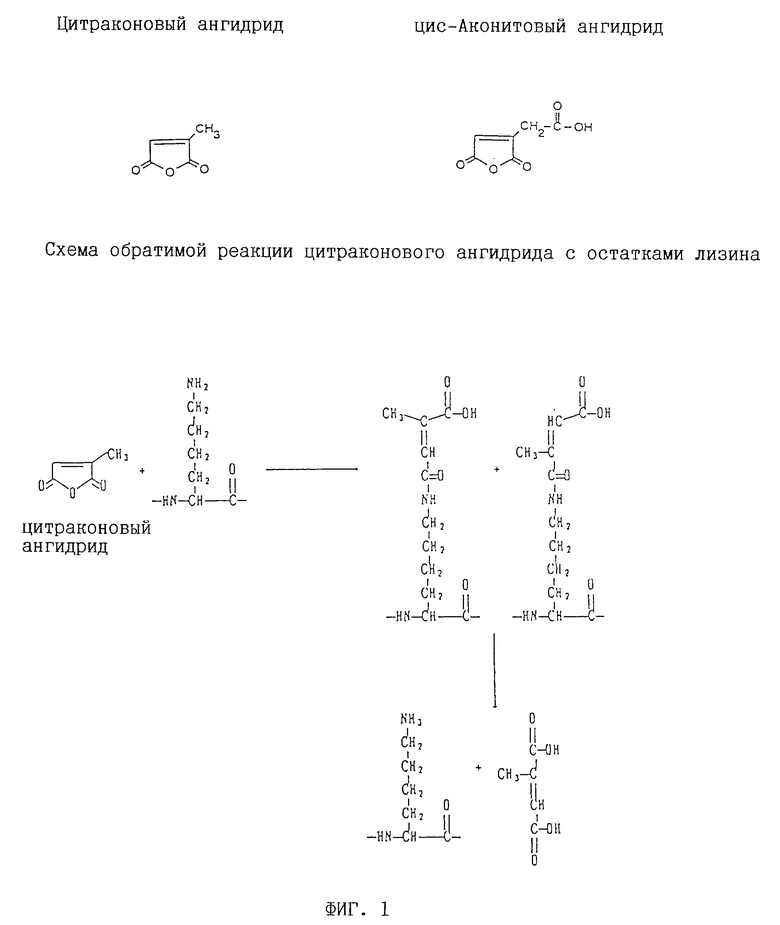

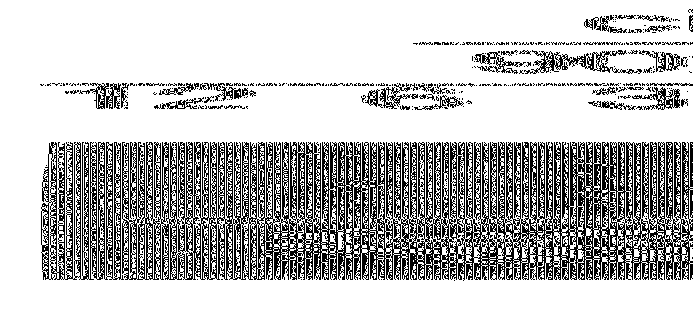
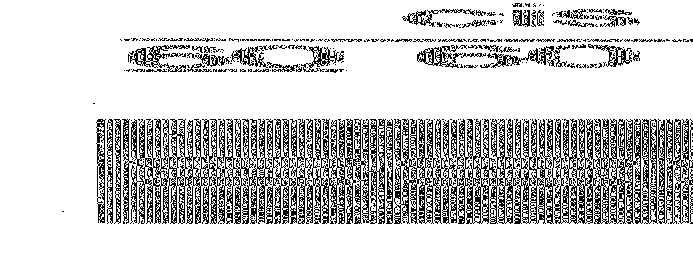
Изобретение относится к биотехнологии к области нуклеиновых кислот, касается амплификации последовательностей нуклеиновых кислот. Химически модифицированный фермент характеризуется тем, что термостабильная ДНК-полимераза или термостабильная лигаза обратимо инактивирована в водном буфере при щелочном значении рН, при температуре ниже приблизительно 25°С модифицирующим реагентом, что не приводит к существенному увеличению ферментативной активности в течение промежутка времени менее приблизительно 20 мин. Модифицирующим реагентом выбран ангидрид дикарбоновой кислоты. Инкубация химически модифицированного фермента в водном буфере, значение рН которого доведено до приблизительно 8-9 при 25°С, при температуре выше приблизительно 50°С приводит по крайней мере к двукратному увеличению ферментативной активности в течение промежутка времени менее приблизительно 20 мин. Химически модифицированную лигазу или ДНК полимеразу применяют для амплификации нуклеиновых кислот. Для чего активность инактивированного фермента восстанавливают путем инкубации реакционной смеси при повышенной температуре перед реакцией амплификации или этот процесс является составной частью реакции амплификации. Необратимо инактивированный термостабильный фермент, применяемый в ПЦР-амплификации, существенно снижает неспецифическую амплификацию и существенно увеличивает количество нужной амплифицированной последовательности-мишени. 5 с. и 10 з.п. ф-лы, 5 ил., 6 табл.
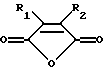
где R1 и R2 обозначают водород или органические радикалы, которые могут быть связаны, или общей формулы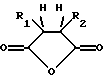
где R1 и R2 обозначают органические радикалы, которые предпочтительно связаны, и атомы водорода находятся в цис-положении.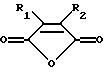
где R1 и R2 обозначают водород или органические радикалы, которые могут быть связаны, или общей формулы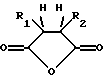
где R1 и R2 обозначают органические радикалы, которые предпочтительно связаны, и атомы водорода находятся в цис-положении и реакцию проводят до практически полной инактивации ферментативной активности.
| US 5262525, 16.11.1993 | |||
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Fransis Barany | |||
| Genetic disease detection and DNA amhlitication using cloned thermostable ligase | |||
| Proc | |||
| Natl | |||
| Acad | |||
| Sci, 1991, v.88, p.189-193 | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| US 4683202, 28.07.1987 | |||
| US 4695188, 23.10.1990. | |||
