Предпосылки создания изобретения
Изобретение относится к новой бактерии, продуцирующей L-глутаминовую кислоту, и к способу получения L-глутаминовой кислоты путем ферментации с использованием указанной бактерии. L-глутаминовая кислота является важной аминокислотой, используемой в пище, лекарственных средствах и т.п.
L-глутаминовая кислота обычно продуцируется способами ферментации с использованием так называемых коринеформных бактерий, продуцирующих L-глутаминовую кислоту, которые в основном принадлежат к роду Brevibacterium, Corynebacterium и Microbacterium, или их вариантов ("Amino Acid Fermentation", Gakkai Shuppan Center, pp.195-215, 1986). Известными методами продуцирования L-глутаминовой кислоты с использованием других бактериальных штаммов являются способы ферментации, в которых используются микроорганизмы рода Bacillus, Streptomyces, Penicillium и т.п. (патент США 3220929); методы, в которых используются микроорганизмы рода Pseudomonas, Arthrobacter, Serratia, Candida и т.п. (патент США 3563857); методы, в которых используются вариантные штаммы Esherichia coli (выложенная заявка на патент Японии (КОКА1) 5-244970 (1993)), и т.п.
Хотя продуцируемость L-глутаминовой кислоты была в основном улучшена путем размножения указанных микроорганизмов вышеупомянутыми способами или путем усовершенствования способов продуцирования, необходимость в разработке менее дорогостоящих и более эффективных способов получения L-глутаминовой кислоты, которые удовлетворяли бы ожидаемому в будущем заметному повышению спроса на аминокислоты, пока еще остается актуальной.
Краткое описание изобретения
Целью настоящего изобретения является обнаружение новой бактерии, продуцирующей L-глутаминовую кислоту и обладающей высокой способностью к продуцированию указанной L-глутаминовой кислоты, что позволяет разработать менее дорогостоящий и более эффективный способ получения L-глутаминовой кислоты.
Для достижения вышеуказанной цели авторы настоящего изобретения провели интенсивные исследования и изучение микроорганизмов, которые обладают способностью продуцировать L-глутаминовую кислоту и которые отличаются от уже известных микроорганизмов. В результате этих исследований было обнаружено, что некоторые штаммы, происходящие от микроорганизмов, принадлежащих к роду Klebsiella или Erwinia, обладают высокой степенью способности к продуцированию L-глутаминовой кислоты, и этот факт был положен в основу настоящего изобретения.
Таким образом, настоящее изобретение относится:
(1) к микроорганизму, принадлежащему к роду Klebsiella, роду Erwinia или роду Pantoea и обладающему способностью продуцировать L-глутаминовую кислоту;
(2) к микроорганизму, определенному выше в (1), которым явлется Klebsiella planticola или Pantoea agglomerans;
(3) к микроорганизму, определенному выше в (1) или (2), который обладает повышенной активностью фермента, катализирующего реакцию биосинтеза L-глутаминовой кислоты;
(4) к микроорганизму, определенному выше в п.(3), где указанным ферментом, катализирующим реакцию биосинтеза L-глутаминовой кислоты, является, по крайней мере, один фермент, выбранный из группы, включающей цитрат-синтазу (далее сокращенно обозначаемую "CS"), фосфоенолпируват-карбоксилазу (далее сокращенно обозначаемую "РЕРС"), и глутамат-дегидрогеназу (далее сокращенно обозначаемую "GDH");
(5) к микроорганизму, определенному выше в п.(4), где указанным ферментом, катализирующим реакцию биосинтеза L-глутаминовой кислоты, являются все ферменты CS, РЕРС и GDH;
(6) к микроорганизму, определенному в любом из вышеуказанных п.п.(1)-(5), который обладает пониженной или вообще не обладает активностью фермента, катализирующего реакцию, ответвляющуюся от пути биосинтеза L-глутаминовой кислоты и продуцирующую соединение, не являющееся L-глутаминовой кислотой:
(7) к микроорганизму, определенному выше в п.(6), где указанным ферментом, катализирующим реакцию, ответвляющуюся от пути биосинтеза L-глутаминовой кислоты и продуцирующую соединение, не относящееся к L-глутаминовой кислоте, является α-кетоглутарат-дегидрогеназа (обозначаемая далее "αKGDH"); и
(8) к способу продуцирования L-глутаминовой кислоты, в котором микроорганизм, определенный в любом из вышеуказанных п.п.(1)-(7), культивируют в жидкой культуральной среде, в результате чего происходит продуцирование и акумуляция L-глутаминовой кислоты в культуральной среде, а затем из этой культуральной среды собирают L-глутаминовую кислоту.
Поскольку микроорганизм настоящего изобретения обладает высокой продуктивностью (получения) L-глутаминовой кислоты, то эту способность микроорганизма можно еще больше увеличить путем использования методов селекции ранее известных для коринеформных бактерий, продуцирующих L-глутаминовую кислоту, и т.п., что позволит разработать более дешевый и более эффективный способ получения L-глутаминовой кислоты, путем соответствующего выбора условий культивирования и т.п. Краткое описание чертежей.
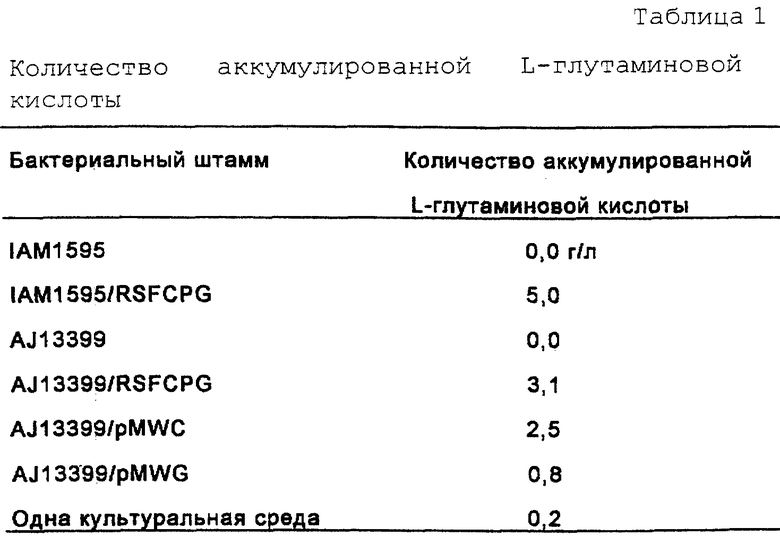
На фиг.1 показано конструирование плазмиды pMWCPG, имеющей ген gltA, ген ррс и ген gdhA.
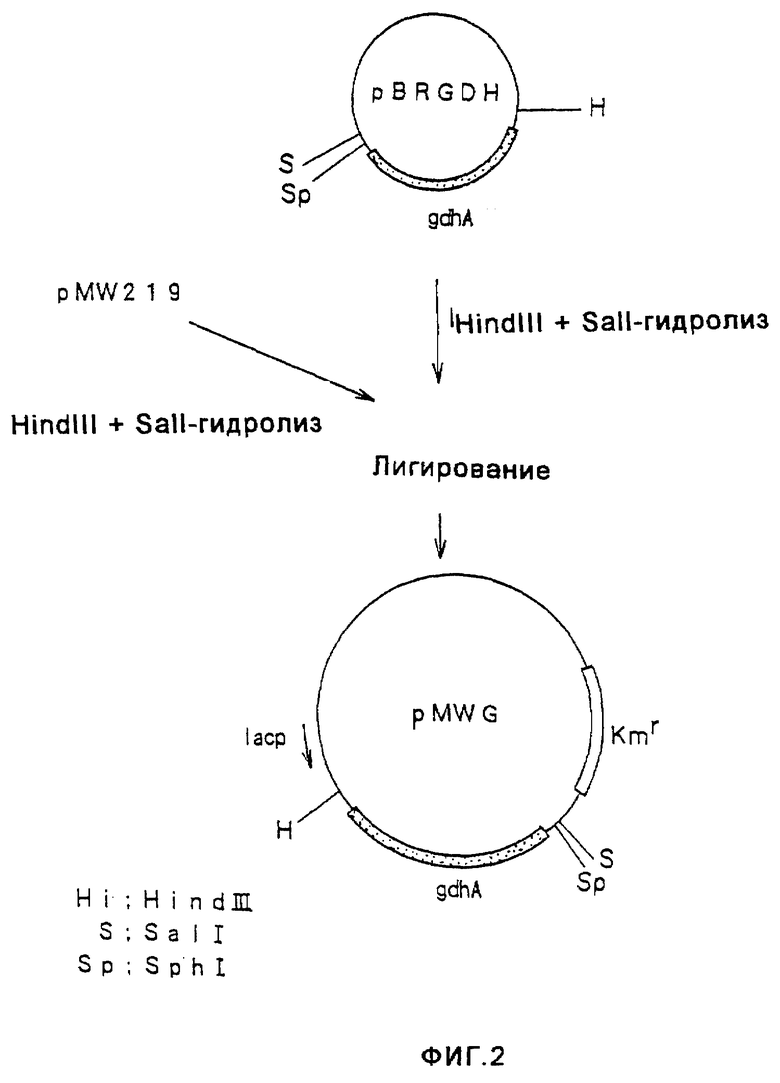
На фиг.2 показано конструирование плазмиды pMWG, имеющей ген gdhA.
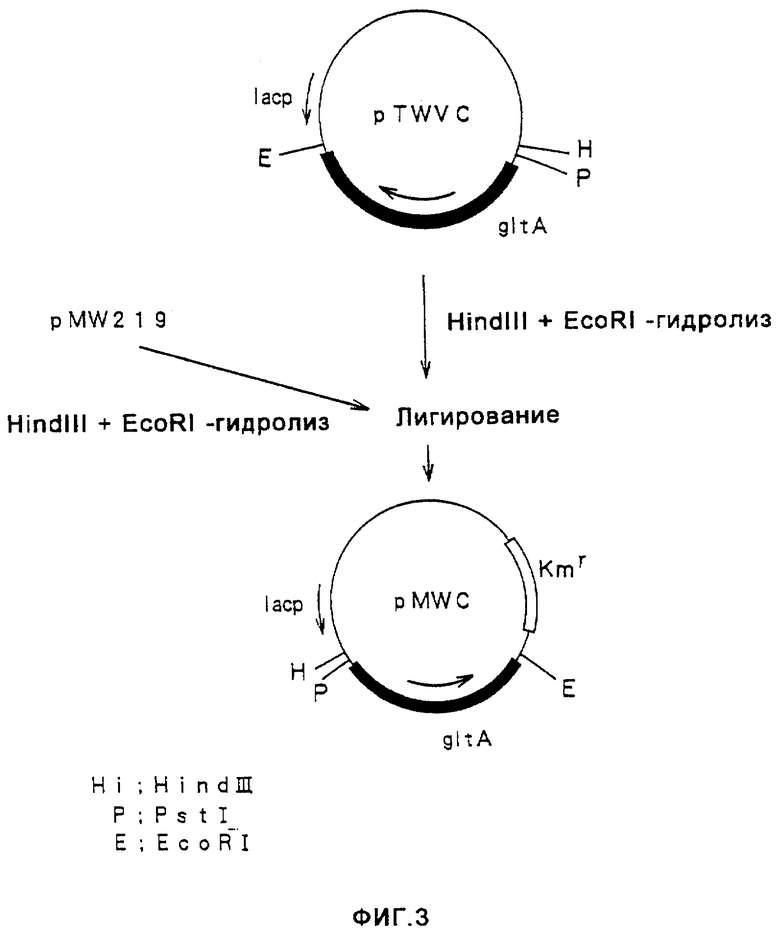
На фиг.3 показано конструирование плазмиды pMWC, имеющей ген gltA.
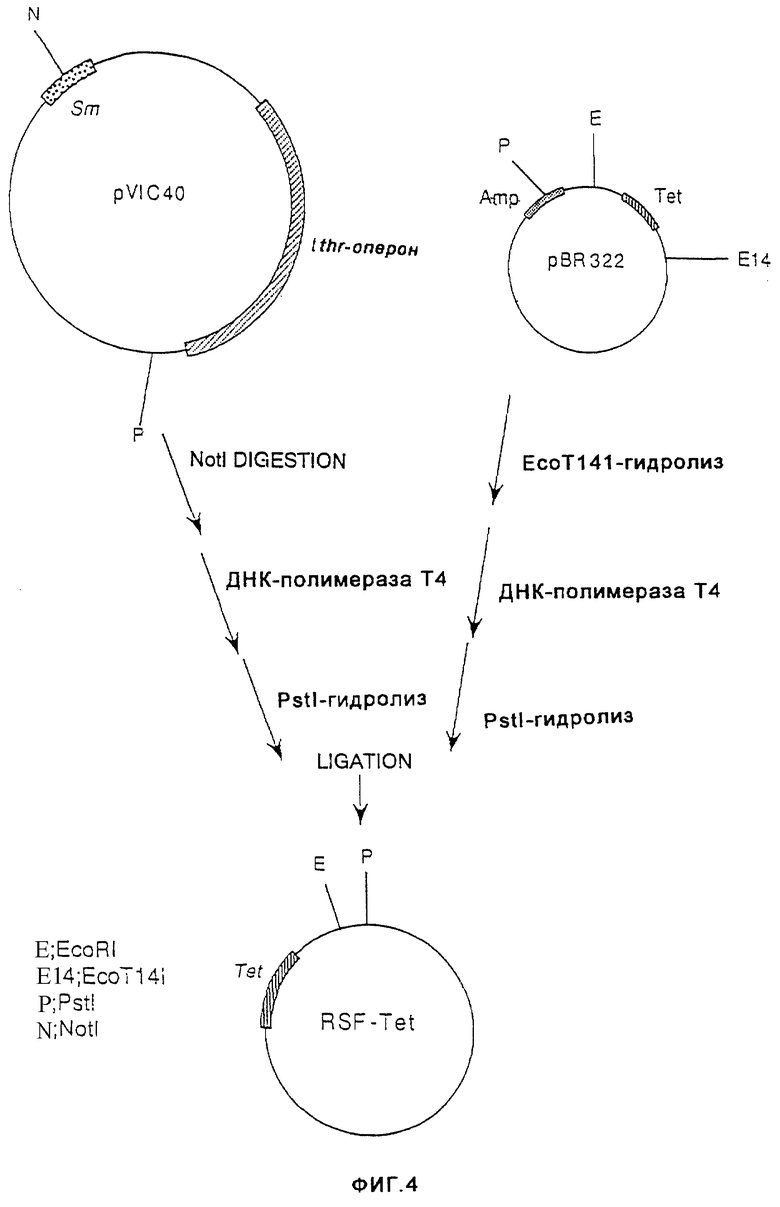
На фиг.4 показано конструирование плазмиды RSF-Tet, имеющей сайт инициации репликации плазмиды RSF1010 широкого круга хозяев дикого типа и ген резистентности к тетрациклину.
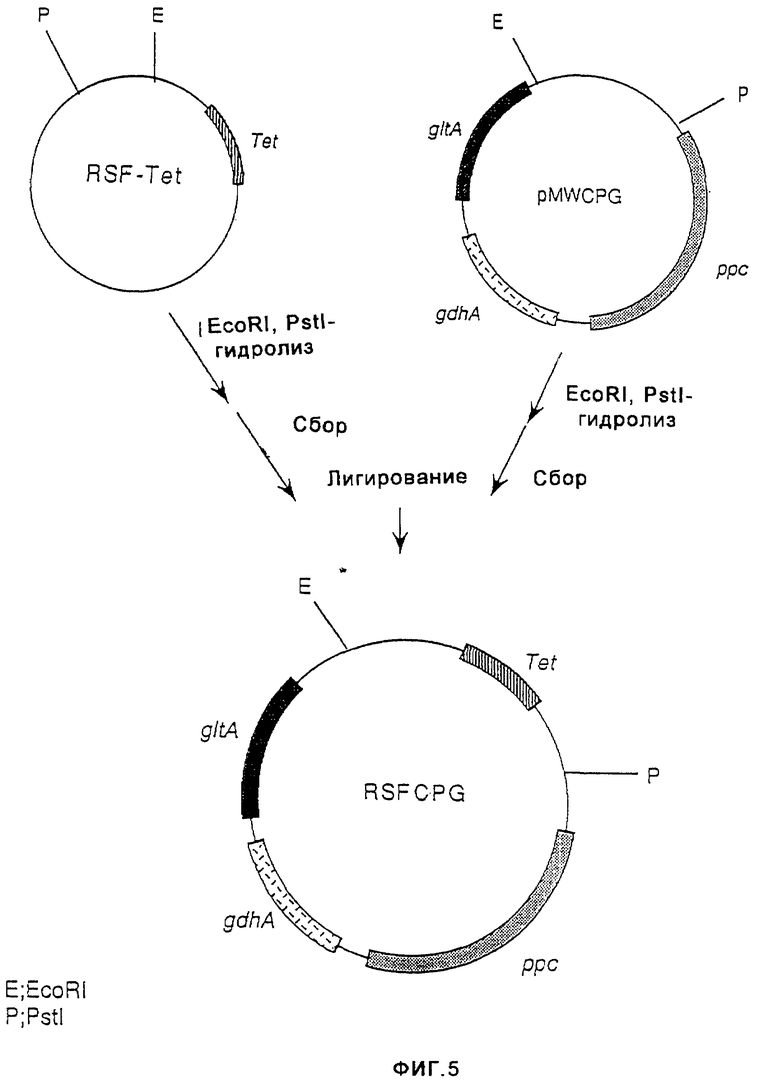
На фиг. 5 показано конструирование плазмиды RSFCPG, имеющей сайт инициации репликации плазмиды RSF1010 широкого круга хозяев дикого типа, ген резистентности к тетрациклину, ген gltA, ген ррс и ген gdhA.
Подробное описание изобретения
Ниже приводится более подробное описание изобретения. Примерами микроорганизма, принадлежащего к роду Klebsiella, Erwinia или Pantoea, которые могут быть использованы в целях настоящего изобретения, являются нижеследующие микрооганиз-мы:
Klebsiella planticola
Klebsiella terrigena
Erwinia herbicola (классифицированная в данной работе как
Pantoea agglomerans)
Erwinia ananas
Erwinia cacticida
Erwinia chrysanthemi
Erwinia mallotivora
Erwinia persicinus
Erwinia psidii
Erwinia quercina
Erwinia rhapontici
Erwinia rubrifaciens
Erwinia salicis
Erwinia uredovora
Pantoea agglomerans
Pantoea dispersa
Из вышеуказанных микроорганизмов более предпочтительными являются нижеследующие бактериальные штаммы:
Klebsiella planticola AJ13399 Erwinia herbicola IAM1595 (Pantoea agglomerans AJ12666) Штамм Klebsiella planticola AJ13399 был депонирован Национальным институтом биологических наук и Технологии человека, Агенство промышленных наук и техники, Министерство Международной торговли и промышленности 19 февраля 1998, под номером допуска FERM P-16646, а затем в соответствии с Будапештским договором он был переведен на международное депонирование, 11 января 1999 г. под номером допуска FERM ВР-6616. Микроорганизм, который был классифицирован как Erwinia herbicola, в настоящей работе классифицирован как Pantoea agglomerans. Поэтому микроорганизм Erwinia herbicola IAM1595 был обозначен Pantoea agglomerans AJ2666, и в соответствии с Будапештским договором был положен на международное депонирование Национальным институтом биологических наук и технологии человека, Агенство промышленных наук и техники, Министерство Международной торговли и промышленности, 25 февраля 1999 под номером допуска FERM BP-6660.
Klebsiella planticola AJ13399 представляет собой штамм, выделенный из почвы в Sapporo-shi Хоккайдо Япония.
Бактерия AJ13399 имеет следующие физиологические свойства:
(1) Морфология клетки палочка
(2) Подвижность: отсутствует
(3) Образование спор: отсутствует
(4) Морфология колоний на питательном агаре с LabM: кольцевая, гладкая поверхность, кремового цвета, ровная, выпуклая и блестящая
(5) Тест на глюкозу: положительна на сбраживаемость
(6) Окрашивание по Граму: отрицательна
(7) Реакция на кислород: факультативный анаэроб
(8) Каталаза: положительна
(9) Оксидаза: отрицательна
(10) Уреаза: положительна
(11) Цитохромоксидаза: отрицательна
(12) β-галактозидаза: положительна
(13) Аргинин-дегидратаза: отрицательна
(14) Орнитин-декарбоксилаза: отрицательна
(15) Лизин-декарбоксилаза: положительна
(16) Триптофан-деаминаза: отрицательна
(17) Реакция Фогеса-Проскауэра: положительна
(18) Продуцирование индола: положительна
(19) Продуцирование гидросульфида в культуральной среде TSI: отрицательна
(20) Усвоение лимонной кислоты: положительна
(21) Усвоение м-гидроксибензойной кислоты: отрицательна
(22) Разжижение желатина: отрицательна
(23) Продуцирование кислоты из сахара:
Глюкоза: положительна
Маннит: положительна
Рамноза: положительна
Арабиноза: положительна
Сахароза: положительна
Сорбит: положительна
Инозит: положительна
Мелибиоза: положительна
Амигдалин: положительна
Адонит-пептон-вода: положительна
Целлобиоза-пептон-вода: положительна
Дульцит-пептон-вода: отрицательна
Раффиноза-пептон-вода: положительна
(23) Температура роста: хороший рост при 37oС, рост отсутствует при 45oС.
Исходя из указанных бактериологических свойств, микроорганизм AJ13399 определен как Klebsiella planticola.
В "Определителе бактерий Берджи, девятое издание" бактерия Erwinia herbicola не описана, и микроорганизм, который был классифицирован как Erwinia herbicola, классифицирован как Pantoea agglomerans. Эти микроорганизмы принадлежат к роду Erwinia, а микроорганизмы, принадлежащие к роду Pantoea, являются близкородственными по отношению друг к другу. Поэтому любой из микроорганизмов, принадлежащих к роду Erwinia и к роду Pantoea, могут быть использованы в настоящем изобретении.
Метаболизм сахара бактериями, принадлежащими к роду Klebsiella, Erwinia или Pantoea, такими как бактерии, упомянутые выше, происходит по пути Эмбдена-Мейерхофа, и продуцированнный в результате этого пируват окисляется в цикле трикарбоновых кислот в аэробных условиях. Биосинтез L-глутаминовой кислоты происходит из α-кетоглутаровой кислоты, являющейся промежуточным соединением цикла трикарбоновых кислот, под действием фермента GDH или глутаминсинтетазы/глутаматсинтазы. Таким образом, эти микроорганизмы имеют один и тот же путь биосинтеза L-глутаминовой кислоты и микроорганизм, принадлежащий к роду Klebsiella, Erwinia или Pantoea, охватываются единой концепцией настоящего изобретения. Поэтому микроорганизмы, принадлежащие к роду Klebsiella, Erwinia или Pantoea и отличающиеся от видов и штаммов, конкретно упомянутых выше, также входят в объем настоящего изобретения.
Микроорганизмом настоящего изобретения является микроорганизм, принадлежащий к роду Klebsiella, Erwinia или Pantoea и обладающий способностью продуцировать L-глутаминовую кислоту. Выражение "обладающий способностью продуцировать L-глутаминовую кислоту", используемое в настоящем описании, означает, что этот микроорганизм способен аккумулировать L-глутаминовую кислоту в культуральной среде в процессе культивирования. Способностью продуцировать L-глутаминовую кислоту может обладать либо штамм дикого типа как присущим ему свойством, либо штамм, наделенный этим свойством, или штамм, у которого это свойство было усилено путем культивирования. Примерами микроорганизма, принадлежащего к роду Klebsiella, Erwinia или Pantoea и обладающего способностью продуцировать L-глутаминовую кислоту, являются, например, такие микроорганизмы, которые обладают повышенной активностью фермента, катализирующего реакцию биосинтеза L-глутаминовой кислоты, и такие микроорганизмы, которые обладают пониженной или вообще не обладают активностью фермента, катализирующего реакцию, ответвляющуюся от пути биосинтеза L-глутаминовой кислоты и продуцирующую соединение, не являющееся L-глутаминовой кислотой. Другим примером микроорганизма является микроорганизм, который обладает повышенной активностью фермента, катализирующего реакцию биосинтеза L-глутаминовой кислоты, и который обладает пониженной или вообще не обладает активностью фермента, катализирующего реакцию, ответвляющуюся от пути биосинтеза L-глутаминовой кислоты и продуцирующую соединение, не являющееся L-глутаминовой кислотой.
Примерами фермента, катализирующего реакцию биосинтеза L-глутаминовой кислоты, могут быть вышеупомянутый фермент GDH, глутаминсинтетаза, глутаматсинтаза, изоцитратдегидрогеназа, аконитатгидратаза, CS, РЕРС, пируватдегидрогеназа, пируваткиназа, энолаза, фосфоглицеромутаза, фосфоглицераткиназа, глицеральдегид-3-фосфат-дегидрогеназа, триозофосфат-изомераза, фруктозобифосфатальдолаза, фосфофруктокиназа, глюкозофосфатизомераза и т.п. Из этих ферментов предпочтительными являются один, два или три фермента из CS, РЕРС и GDH. В качестве микроорганизма настоящего изобретения более предпочтительным является микроорганизм, который имеет повышенные активности всех видов ферментов CS, РЕРС и GDH. Установление того факта, обладает ли микроорганизм повышенной активностью целевого фермента, и определение степени повышения этой активности может быть осуществлено путем измерения ферментативной активности бактериального клеточного экстракта или очищенной фракции, и сравнения этой активности с активностью штамма дикого типа или родительского штамма.
Микроорганизм настоящего изобретения, который принадлежит к роду Klebsiella, Erwinia или Pantoea и имеет повышенную активность фермента, катализирующего реакцию биосинтеза L-глутаминовой кислоты, может быть получен, например, в виде варианта, в который была введена мутация в ген, кодирующий данный фермент, или в виде генетически рекомбинантного штамма, с использованием в качестве исходного родительского штамма любого из микроорганизмов, упомянутых выше.
Так, например, для усиления активности CS, РЕРС и GDH ген, кодирующий CS, РЕРС и GDH, может быть клонирован в подходящую плазмиду, а вышеуказанный исходный родительский штамм в качестве хозяина может быть трансформирован полученной плазмидой. Это может приводить к увеличению числа копий каждого из генов, кодирующих CS, РЕРС и GDH (обозначаемых ниже "геном gltA", "геном ррс" и "геном gdhA" соответственно), в результате чего активности CS, РЕРС и GDH могут быть увеличены.
Для этого в исходный родительский штамм, упомянутый выше, вводят один, два или три вида генов, выбранных из клонированных гена gltA, гена ррс и гена gdhA в любой комбинации. В случае, когда вводят два или три вида генов, то два или три вида этих генов клонируют в одном виде плазмиды и вводят в клетку-хозяина, либо эти гены отдельно клонируют в двух или трех видах плазмид, которые могут существовать в одном и том же хозяине, и вводят в клетку-хозяина.
Тип используемой плазмиды не имеет конкретных ограничений, при условии, что эта плазмида способна автономно реплици-роваться в микроорганизме, принадлежащем к роду Klebsiella, Erwinia или Pantoea. Примерами такой плазмиды являются pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219, pMW218 и т.п. Могут быть также использованы другие плазмиды и фаговые ДНК-векторы.
Трансформация может быть осуществлена, например, способом D.M. Morrison (Methods in Enzymology 68, 326 (1979)); способом повышения проницаемости клеток-реципиентов для ДНК с использованием хлорида кальция (Mandel М. & Higa A., J. Mol. Biol., 53, 159 (1970)) и т.п.
Активности ферментов CS, РЕРС и GDH могут быть также увеличены с использованием множества копий гена gltA, гена ррс и/или гена gdhA, присутствующих на хромосомной ДНК исходного родительского штамма-хозяина. Для введения множества копий гена gltA, гена ррс и/или гена gdhA в хромосомную ДНК микроорганизма, принадлежащего к роду Klebsiella, Erwinia или Pantoea, могут быть использованы последовательности, присутствующие на хромосомной ДНК в множестве копий, такие как повторяющаяся ДНК, и инвертированные повторы, присутствующие на конце транспозиционных факторов. Альтернативно, множественные копии этих генов могут быть также введены в хромосомную ДНК с использованием транспозиции транспозонов, несущих ген gltA, ген ррс или ген gdhA. Эти методы позволяют увеличить число копий гена gltA, гена pрс и гена gdhA в клетках-трансформантах, в результате чего увеличивается активность CS, РЕРС и GDH.
В качестве источника гена gltA, гена ррс и гена gdhA, используемых для увеличения числа копий, могут быть использованы любые микроорганизмы, при условии, что они обладают CS-, РЕРС- и GDH-активностями. Из указанных микроорганизмов предпочтительными являются бактерии, то есть прокариоты, такие как бактерии, принадлежащие к роду Enterobacter, Klebsiella, Erwinia, Pantoea, Serratia, Escherichia, Corynebacterium, Brevibacterium и Bacillus. В качестве конкретного примера может быть упомянута бактерия Escherichia coli. Ген gltA, ген ррс и ген gdhA могут быть получены из хромосомной ДНК вышеупомянутых микроорганизмов.
Каждый из генов qltA, ррс и gdhA может быть получен из хромосомной ДНК любого из вышеупомянутых микроорганизмов путем выделения ДНК-фрагмента, комплементирующего ауксотрофию штамма-варианта, у которого отсутствует CS-, РЕРС- и GDH-активность. Альтернативно, поскольку нуклеотидные последовательности этих генов в бактериях рода Escherichia или Corynebacterium были уже выявлены (Biochemistry, Vol. 22, pp. 5243-5249, 1983; J. Biochem. Vol. 95, pp. 909-916, 1984; Gene, Vol. 27, pp. 193-199. 1984; Microbiology, Vol. 140, pp. 1817-1828, 1994; Mol. Gen. Genet Vol. 218, pp. 330-339, 1989; и Molecular Microbiology, Vol. 6, pp. 317-326, 1992), то эти гены могут быть получены посредством PCR (полимеразной цепной реакции) с использованием праймеров, синтезированных на основе каждой из выявленных нуклеотидных последовательностей, и хромосомной ДНК в качестве матрицы.
Активности CS, РЕРС и GDH могут быть также увеличены способами усиления экспрессии гена gltA, гена ррс или гена gdhA, не относящимися к вышеупомянутым методам амплификации генов. Так, например, экспрессия этих генов может быть усилена путем замены промотора гена gltA, гена ррс или гена gdhA другим более сильным промотором. Примерами таких сильных промоторов являются, например, промотор Iac, trp-промотор, промотор trc, промотор tac, промотор PR, и промотор PL лямбда-фага и т.п. Ген gltA, ген ррс или ген gdhA, промотор которых был заменен, клонируют в плазмиду и встраивают в микроорганизм-хозяина, или встраивают в хромосомную ДНК микроорганизма-хозяина с использованием повторяющейся ДНК, инвертированного повтора, транспозона или т.п.
Активности CS, РЕРС и GDH могут быть также увеличены путем замены промотора гена gltA, гена ррс или гена gdhA на хромосоме другим более сильным промотором (см. , W087/03006 и выложенную заявку на патент Японии (KOKAI) 61-268183 (1986)) или путем вставки сильного промотора в положение, находящееся выше каждой кодирующей последовательности (см. Gene, 29, pp. 231-241, 1984). В частности, это может быть достигнуто путем гомологичной рекомбинации между геном gltA, геном ррс или геном gdhA, промотор которых был заменен более сильным промотором, или их ДНК-содержашей частью, и соответствующим геном на хромосоме.
Конкретными примерами микроорганизма, принадлежащего к роду Klebsiella, Erwinia или Pantoea и имеющего повышенные CS-, РЕРС- и GDH-активности, являются, например, Klebsiella planticola ATCC13399/RSFCPG и Erwinia herbicola IAM1595/RSFCPG.
Примерами фермента, катализирующего реакцию, ответвляющуюся от пути биосинтеза L-глутаминовой кислоты и продуцирующую соединение, не являющееся L-глутаминовой кислотой, могут служить, например, αKGDH, изоцитратлиаза, фосфатацетилтрансфераза, ацетаткиназа, ацетогидроксикислота-синтаза, ацето-лактатсинтаза, формиатацетилтрансфераза, лактатдегидрогеназа, L-глутамат-декарбоксилаза, 1-пирролин-дегидрогеназа и т.п. Из этих ферментов предпочтительным является αKGDH.
Для достижения уменьшения или отсутствия вышеупомянутой ферментативной активности в микроорганизме, принадлежащем к роду Klebsiella, Erwinia или Pantoea, в ген, кодирующий данный фермент, с помощью стандартной техники мутагенеза или техники генной инженерии может быть введена мутация, приводящая к снижению или к отсутствию ферментативной активности,
Примерами техники мутагенеза могут служить, например, метод с использованием облучения рентгеновским или ультрафиолетовым излучением; метод с использованием обработки мутагенным агентом, таким как М-метил-N'-нитро-N-нитрозогуанидин, и т. п Область гена, в которую вводят мутацию, может быть кодирующей областью белка-фермента или областью, регулирующей экспрессию, такой как промотор.
Примерами техники генной инженерии могут служить, например, генетическая рекомбинация, генетическая трансдукция, слияние клеток, и т.п. Так, например, для продуцирования функционально инактивированного гена (дефектного гена) в нужный ген встраивают ген резистентности к лекарственному средству. Затем этот дефектный ген вводят в клетку микроорганизма, принадлежащего к роду Klebsiella, Erwinia или Pantoea, и целевой ген на хромосоме заменяется дефектным геном посредством гомологичной рекомбинации (разрушение гена).
Для того чтобы определить, имеет ли данный микроорганизм пониженную активность целевого фермента или эта активность отсутствует, может быть измерена ферментативная активность экстракта бактериальных клеток или очищенной фракции штамма-кандидата с последующим сравнением этой активности с активностью штамма дикого типа или родительского штамма. Активность фермента αKGDH может быть измерена, например, методом Reed et al, (L.J Reed & ВВ. Mukherjee, Methods in Enzymology 1969, 13, р.55-61).
В зависимости от целевого фермента нужный вариант может быть выбран исходя из фенотипа этого варианта. Так, например, вариант, в котором отсутствует αKGDH-активность или эта активность является пониженной, не может расти на минимальной среде, содержащей глюкозу, или на минимальной среде, содержащей уксусную кислоту или L-глутаминовую кислоту в качестве единственного источника углерода, либо этот вариант обнаруживает заметно более медленную скорость роста в аэробных условиях. Однако даже при тех же самых условиях может наблюдаться нормальный рост, если в минимальную среду, содержащую глюкозу, добавить янтарную кислоту или лизин, метионин и диаминопимелат. Исходя из этого факта, может быть выбран вариант, в котором отсутствует αKGDH-активность либо эта активность является пониженной.
Способ продуцирования штамма Brevibacterium lactofermentum, не содержащего гена αKGDH, основанный на гомогенной рекомбинации, подробно описан в W095/34672, и аналогичный способ может быть использован для продуцирования микроорганизмов, принадлежащих к роду Klebsiella, Erwinia или Pantoea.
Кроме того, методы генетического клонирования, расщепления и лигирования ДНК, трансформации и т.п., подробно описаны в Molecular Cloning, 2nd edition. Cold Spring Harbor Press (1989) и т.п.
Примером вариантного штамма, не содержащего αKGDH-активности или имеющего пониженную активность, как описано выше, является штамм Klebsiella planticola AJ 13410. Штамм Klebsiella planticola AJ13410 был депонирован Национальным институтом биологических наук и технологии человека, Агенство промышленных наук и техники,. Министерство Международной торговли и промышленности 19 февраля 1998 г., под номером допуска FERM Р-16647, а затем в соответствии с Будапештским договором он был переведен на международное депонирование 11 января 1999 г. под номером допуска FERM ВР-6617.
Бактериальные штаммы, принадлежащие к роду Klebsiella, Erwinia или Pantoea, которые обладают пониженной αKGDH-активностью или вообще не обладают этой активностью, или бактериальные штаммы, которые обладают повышенной CS-, РЕРС- и GDH-активностью и которые были получены, как описано выше, должны обладать способностью продуцировать L-глутаминовую кислоту, как это будет показано в нижеприведенных примерах.
Микроорганизмы, принадлежащие к роду Escherichia, которые были классифицированы как кишечные бактерии аналогично бактериям рода Klebsiella, Erwinia или Pantoea и которые известны как штаммы, обладающие пониженной αKGDH-активностью или вообще не обладающие этой активностью, могут продуцировать L-глутаминовую кислоту (выложенная заявка на патент Японии (KOKAI) 5-244970 (1993)); причем указанные штаммы, которые обладают пониженной αKGDH-активностью или вообще не обладают этой активностью и которые обладают повышенными CS-, РЕРС- и GDH-активностями, могут продуцировать еще более высокие количества L-глутаминовой кислоты (выложенная заявка на патент Японии (KOKAI) 7-203980 (1995)), и, кроме того, штаммы, обладающие восприимчивостью к валину и повышенными CS- и GDH-активностями, могут продуцировать L-глутаминовую кислоту (W097/08294).
Что касается микроорганизмов, принадлежащих к роду Enterobacter, которые аналогично классифицируются как кишечные бактерии, то авторы настоящего изобретения обнаружили, что те штаммы, которые обладают пониженной αKGDH-активностью или вообще не обладают этой активностью, или штаммы, которые обладают повышенными CS-, РЕРС- и GDH-активностями, могут продуцировать L-глутаминовую кислоту (выложенная заявка на патент Японии (KOKAI) 10-69068 (1998)).
Кроме того, что касается микроорганизмов, принадлежащих к роду Serratia, то авторами настоящего изобретения было также обнаружено, что штаммы, обладающие повышенными CS-, РЕРС- и GDH-активностями, могут продуцировать L-глутаминовую кислоту (выложенная заявка на патент Японии (KOKAI) 10-69068 (1998)).
Исходя из этих фактов, легко предсказать, что в случае бактерий, принадлежащих к роду Klebsiella, Erwinia, Pantoea, Escherichia, Enterobacter или Serratia и не относящихся к штаммам, описанным в примерах, те штаммы, которые обладают пониженной αKGDH-активностью или вообще не обладают этой активностью, или те штаммы, которые обладают повышенными CS-, РЕРС- и GDH-активностями, могут продуцировать L-глутаминовую кислоту. Как было продемонстрировано в примерах, имеются все основания утверждать, что способность к продуцированию L-глутаминовой кислоты, которая может быть сообщена штамму Klebsiella planticola AJ13399 путем делеции αKGDH, может быть сообщена и другим бактериям рода Klebsiella, Erwinia или Pantoea.
L-глутаминовая кислота может быть продуцирована путем культивирования микроорганизма, принадлежащего к роду Klebsiella, Erwinia или Pantoea и обладающего способностью продуцировать L-глутаминовую кислоту в жидкой культуральной среде, с продуцированием и аккумуляцией L-глутаминовой кислоты в этой среде и с последующим ее выделением из этой культуральной среды.
Культуральной средой может быть обычная питательная среда, содержащая источник углерода, источник азота и неорганические соли, а также, если это необходимо, органические питательные микроэлементы, такие как аминокислоты, витамины и т. п. Этой средой может быть синтетическая среда или природная среда. Для культуральной среды может быть использован любой из углеродных и азотных источников, при условии, что они могут быть утилизованы культивируемым микроорганизмом.
Источником углерода может являться сахарид, такой как глюкоза, глицерин, фруктоза, сахароза, мальтоза, манноза, галактоза, гидролизаты крахмала, мелассы и т.п. Кроме того, может быть также использована органическая кислота, такая как уксусная кислота и лимонная кислота, отдельно или в комбинации с другими углеродными источниками.
Источником азота может являться аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония и ацетат аммония, а также нитраты, и т.п.
В качестве органических микроэлементов используются аминокислоты, витамины, жирные кислоты, нуклеиновые кислоты, материалы, содержащие эти вещества, такие как пептон, казамино-кислота, дрожжевой экстракт, и продукты разложения соевого белка, и т.п., а в случае ауксотрофного варианта, которому для его роста требуется аминокислота или т.п., эти микроэлементы необходимы для восполнения требуемых питательных веществ.
В качестве неорганической соли используются фосфаты, соли магния, соли кальция, соли железа, соли марганца и т.п.
Что касается условий культивирования, то оно может быть осуществлено в аэробных условиях при температуре 20-42oС и при рН 4-8. Для аккумуляции значительного количества L-глутаминовой кислоты в жидкой культуральной среде культивирование может продолжаться в течение периода времени от 10 часов до 4 дней.
После завершения культивирования L-глутаминовая кислота, аккумулированная в культуральной среде, может быть собрана известным способом. Так, например, она может быть выделена способом, предусматривающим концентрирование среды после удаления клеток с кристаллизацией продукта; методом ионообменной хроматографии или т.п.
Примеры
Более подробно, настоящее изобретение описано ниже со ссылками на следующие примеры.
(1) Конструирование плазмиды, имеющей ген gltA, ген ррс и ген gdhA.
Процедура конструирования плазмиды, имеющей ген gltA, ген ррс и ген gdhA, описана со ссылками на фиг. 1, 2 и 3.
Плазмиду pBRGDH, имеющую ген gdhA, происходящий от Esherichia coli (выложенная заявка на патент Японии (KOKAI) 7-203980 (1995)), гидролизовали ферментами HindIII и SphI и оба конца затупляли путем обработки ДНК-полимеразой Т4. Затем ДНК-фрагмент, содержащий ген gdhA, очищали и выделяли. С другой стороны, плазмиду pMWCP, имеющую ген gltA и ген ррс, происходящие от Escherichia coli (W097/08294), гидролизовали ферментом Xbal и оба конца затупляли путем обработки ДНК-полимеразой Т4. Этот продукт смешивали с ДНК-фрагментом, содержащим ген gdhA, очищенный, как описано выше, и лигировали с помощью лигазы Т4, в результате чего получали плазмиду pMWCPG, которая соответствовала плазмиде pMWCP, кроме того, несущей ген gdhA (фиг.1).
ДНК-фрагмент, имеющий ген gdhA и полученный путем гидролиза pBRGDH ферментами HindIII и Sail, очищали и выделяли, а затем вводили в HindIII-SalI-сайт плазмиды pMW219 (поставляемой от Nippon Gene), в результате чего получали плазмиду pMWG (фиг. 2) Кроме того, плазмиду pTWVC, имеющую ген gltA, происходящую от Escherichia coli (W097/08294), гидролизовали ферментами HindIII и EcoRI и полученный ДНК-фрагмент, имеющий ген gltA, очищали и выделяли, а затем вводили в HindIII-EcoRI-сайт плазмиды pMW219, в результате чего получали плазмиду pMWC (фиг. 3).
В то же время, продукт, полученный путем гидролиза плазмиды pVIC40, содержащей сайт инициации репликации плазмиды pSF1010 широкого круга хозяев (выложенная заявка на патент Японии (КОКАI) 8-047397 (1996)), ферментом NotI, с последующей обработкой ДНК-полимеразой Т4 и гидролизом ферментом PstI, и продукт, полученный путем гидролиза плазмиды p8R322 ферментом ЕсоТ141 с последующей обработкой ДНК-полимеразой Т4 и гидролизом ферментом PstI, смешивали и лигировали с помощью лигазы Т4, в результате чего получали плазмиду RSF-Tet, содержащую сайт инициации репликации RSF1010, и ген резистентности к тетрациклину (фиг.4).
Затем плазмиду pMWCPG гидролизовали ферментами EcoRI и PstI и ДНК-фрагмент, содержащий ген gltA, ген ррс или ген gdhA, очищали и выделяли. Аналогично, RSF-Tet гидролизовали ферментами EcoRI и PstI и ДНК-фрагмент, содержащий сайт инициации репликации RSF1010, очищали и выделяли. Эти ДНК-фрагменты смешивали и лигировали с помощью лигазы Т4, в результате чего получали плазмиду RSFCPG, состоящую из RSF-Tet, несущую ген gltA, ген ррс и ген gdhA (фиг. 5). Экспрессию гена gltA, гена ррс и гена gdhA в полученной плазмиде RSFCPG подтверждали исходя из комплементации ауксотрофии штаммов Escherichia coli, не содержащих гена gltA, гена ррс или гена gdhA, и путем измерения активности каждого фермента. Аналогично, экспрессию гена gltA или гена gdhA в плазмиде pMWG или в плазмиде pMWC подтверждали исходя из комплементации ауксотрофии штаммов Escherichia coli, не содержащих гена gltA или гена gdhA, и путем измерения активности каждого фермента.
(2) Введение RSFCPG, pMWC и pMWG в бактерию рода Klebsiella и бактерию рода Erwinia (Pantoea) и оценка продуцируемости L-глутаминовой кислоты.
Erwinia herbicola IAM1595 (Pantoea agglomerans AJ2666) и Klebsiella planticola AJ13399 трансформировали плазмидами RSFCPG, pMWC и pMWG путем электропорации (Miller J.H. "A Short Course in Bacterial Genetics; Handbook" Cold Spring Harbor Laboratory Press, USA, 1992), в результате чего получали трансформанты, обладающие резистентностью к тетрациклину.
Каждый из полученных трансформантов и родительские штаммы инокулировали в лабораторном сосуде объемом 50 мл, содержащем 5 мл культуральной среды, включающей 40 г/л глюкозы, 20 г/л сульфата аммония, 0,5 г/л гептагидрата сульфата магния, 2 г/л дигидрофосфата калия, 0,5 г/л хлорида натрия, 0,25 г/л гептагидрата хлорида кальция, 0,02 г/л гептагидрата сульфата железа, 0,02 г/л тетрагидрата сульфата марганца, 0,72 мг/л дигидрата сульфата цинка, 0,64 мг/л пентагидрата сульфата меди, 0,72 мг/л гексагидрата хлорида кобальта, 0,4 мг/л борной кислоты, 1,2 мг/л дигидрата молибдата натрия, 2 г/л дрожжевого экстракта и 30 г/л карбоната кальция, и культивировали, встряхивая, при 37oС до тех пор, пока глюкоза, содержащаяся в культуральной среде, не была израсходована. В культуральную среду трансформантов добавляли 25 мг/л тетрациклина. После завершения культивирования измеряли количество L-глутаминовой кислоты, аккумулированной в культуральной среде. Результаты представлены в таблице 1.
Хотя Erwinia herbicola IAM1595 и Klebsiella planticola АЛ 3399 не аккумулировали L-глутаминовую кислоту, однако штаммы, у которых CS-, РЕРС- и GDH-активности были амплифицированы путем введения RSFCPG, аккумулировали 5,0 г/л и 3,1 г/л L-глутаминовой кислоты соответственно. Штамм АJ1 3399, у которого была амплифицирована лишь одна CS-активность, аккумулировал 2,5 г/л L-глутаминовой кислоты, а штамм, у которого была амплифицирована лишь одна GDH-активность, также аккумулировал 0,8 г/л L-глутаминовой кислоты.
(3) Клонирование фрагмента, содержащего часть гена αKGDH штамма Klebsiella planticola AJ 13399
Клонирование фрагмента, содержащего часть гена αKGDH Klebsiella planticola AJ13399, осуществляли с помощью PCR с использованием олигонуклеотидов, каждый из которых имел нуклеотидную последовательность гомологичной области гена αKGDH организмов с известными нуклеотидными последовательностями, то есть Azotobacter vinelandii, Bacillus subtilis, Escherichia coli, Corynebacterium glutamicum, Haemophilus influenzae, человека, и Saccharomyces cerevisiae (Eur. J. Biochem. Vol. 187, pp. 235-239, 1990; Mol. Gen. Genet. Vol. 234, pp. 285-296, 1992; Eur. J. Biochem. Vol. 141, pp. 351-359, 1984; Microbiology, Vol. 142, pp.3347-3354, 1996; Science, Vol. 269, pp. 496-512, 1995; Proc. Natl. Acad. Sci. USA Vol. 89, pp.1963-1967, 1992; and Mol. Cel. Biol. Vol. 9, pp. 2695-2705, 1989), и с использованием EcoRI-сайта в качестве праймеров.
В частности, были использованы нижеследующие праймеры:
(Праймер 1)
5'-CCGGGAATTCGGTGACGTNAARTAYCA 3' SEQ ID No:1
(Праймер 2)
5'-GGCGAATTCGGGAACGGGTASAGYTGYTC 3' SEQ ID No:2.
Хромосомную ДНК штамма Klebsiella planticola AJ13399, используемую в качестве матрицы для PCR, выделяли тем же самым методом, который обычно используется для экстракции хромосомной ДНК из Escherichia coli (Seibutsu Kogaku Jikkensho (Textbook of Bioengineering Experiments), Ed. by the Society of Fermentation and Bioengineering, Japan, p.97-98, Baifukan, 1992).
PCR осуществляли в 30 циклов, каждый из которых проводили в течение 1 минуты при 94oС, в течение 1 минуты при 50oC и в течение 3 минут при 73oС, и полученный ДНК-фрагмент гидролизовали ферментом EcoRI и встраивали в плазмидный вектор pT7Blue, гидролизованный ферментом EcoRI, в результате чего получали рекомбинантную плазмиду рТ7КР. Используемый плазмидный вектор pT7Blue (резистентный к ампициллину) является коммерческим продуктом, поставляемым фирмой Novagen.
Нуклеотидная ДНК-последовательность клонированного фрагмента и аминокислотная последовательность, кодируемая этой последовательностью, представлены в SEQ ID No:3. Та же самая последовательность отдельно представлена в SEQ ID No: 4. Эта последовательность обнаруживает 82,3%-ную гомологию с геном субъединицы αKGDH Escherichia coli (обозначаемым далее "геном sucA") и она четко идентифицируется как часть гена sucA штамма Klebsiella ptanticola AJ13399. В нуклеотидной последовательности, представленной в SEQ ID No:3, нуклеотиды 18-1659 происходят от гена sucA, а нуклеотиды 0-17 и 1660-16679 происходят от праймеров.
(4) Получение штамма, дефицитного по αKGDH и происходящего от Klebsiella planticola AJ13399
Штамм Klebsiella planticola, дефицитный по αKGDH, был получен, как описано выше, путем гомологичной рекомбинации с использованием фрагмента, содержащего часть гена sucA Klebsiella planticola.
Для этого сначала рТ7КР гидролизовали ферментом BstEII и в этот рестриктированный сайт был встроен фрагмент гена резистентности к канамицину, который был клонирован посредством PCR из плазмиды pNEO (поставляемой фирмой Pharmacia), и в оба конца которого были введены BstEII-сайты, в результате чего была получена плазмида рТ7КРКm, в которой ген резистентности к канамицину был встроен в центральную часть гена sucA Klebsiella planticola.
Для клонирования гена резистентности к канамицину были использованы следующие праймеры:
(Праймер 3)
S'-TACTGGGTCACCTGACAGCTTATCATCGAT 3' (SEQ ID No:5)
(Праймер 4)
S'-CGTTCGGTGACCACCAAAGCGGCCATCGTG 3' (SEQ ID No:6)
Затем плазмиду рТ7КРКт гидролизовали ферментом KpnI, и в этот рестриктированный сайт был встроен фрагмент гена резистентности к тетрациклицину, который был клонирован посредством PCR из плазмиды pBR322 (поставляемой фирмой Takara Shuzo) и в оба конца которого были введены KpnI-сайты, в результате чего была получена плазмида рТ7КРКmТс, в которой ген резистентности к тетрациклицину был встроен в центральную часть гена sucA Klebsiella planticola вместе со встроенным фланкирующим геном резистентности к тетрациклину.
Для клонирования гена резистентности к тетрациклину были использованы следующие праймеры:
(Праймер 5)
S'-GGGGTACCCAAATAGGCGTATCACGAG 3' (SEQ ID No:7)
(Праймер 6)
S'-GGGGTACCCGCGATGGATATGTTCTG 3' (SEQ ID No:8).
Затем плазмиду pT7KPKmTc гидролизовали ферментами SacI и XbaI для вырезания ДНК-фрагмента, содержащего ген резистентности к канамицину, встроенный в центральную часть гена sucA Klebsiella planticola вместе с встроенным фланкирующим геном резистентности к тетрациклину, и этот фрагмент вводили в плазмидный вектор pGP704, который был встроен в хромосому грам-отрицательной бактерии (Marta Herrero et al. , Journal of Bacteriology, 1990, 172, p. 6557-6567) и который был гидролизован ферментами Sad и Xbal, в результате чего получали плазмиду pUTONOTK.
С использованием плазмиды pUTONOTK, полученной, как описано выше, штамм Klebsiella planticola AJ13399 трансформировали путем электропорации, а штамм, в хромосому которого встраивали плазмиду pUTONOTK путем гомогенной рекомбинации фрагмента гена sucA, отбирали по резистентности к тетрациклину и резистентности к канамицину. Из этого штамма, исходя из восприимчивости к тетрациклину и резистентности к канамицину, был, кроме того, получен штамм Klebsiella planticola AJ13410, не содержащий гена sucA, где ген sucA на хромосоме был заменен геном sucA, в центральную часть которого был встроен ген резистентности к канамицину. Для подтверждения того, что полученный, как описано выше, штамм AJ13410 является дефицитным по αKGDH-активности, его ферментативную активность определяли методом Reed (L.J. Reed & В.В. Mukherjee, Methods in Enzymology 1969, 13, р. 55-61). В результате этого αKGDH-активность не была обнаружена в
штамме AJ13410, как показано в таблице 2, что свидетельствует о том, что этот штамм, как и требовалось, не содержит гена sucA.
(5) Оценка продуцируемости L-глутаминовой кислоты штаммом Klebsiella planticola, который является дефицитным по αKGDH.
Каждый из штаммов AJ13399 и AJ13410 инокулировали в колбе объемом 500 мл, содержащей 20 мл культуральной среды, включающей 40 г/л глюкозы, 20 г/л сульфата аммония, 0,5 г/л гептагидрата сульфата магния, 2 г/л дигидрофосфата калия, 0,5 г/л хлорида натрия, 0,25 г/л гептагидрата хлорида кальция, 0,02 г/л гептагидрата сульфата железа, 0,02 г/л тетрагидрата сульфата марганца, 0,72 мг/л дигидрата сульфата цинка, 0,64 мг/л пентагидрата сульфата меди, 0,72 мг/л гексагидрата хлорида кобальта, 0,4 мг/л борной кислоты, 1,2 мг/л дигидрата молибдата натрия, 2 г/л дрожжевого экстракта и 30 г/л карбоната кальция, 200 мг/л моногидрохлорида L-лизина, 200 мг/л L-метионина и 200 мг/л DL-α,ε-диаминопимелиновой кислоты (DAP), и культивировали, встряхивая, при 37oС до тех пор, пока глюкоза, содержащаяся в культуральной среде, не была израсходована. После завершения культивирования измеряли количество L-глутаминовой кислоты и α-кетоглутаровой кислоты (обозначаемой далее "αKG"), аккумулированных в культуральной среде. Результаты представлены в таблице 3.
Штамм AJ13410, который является дефицитным по αKGDH-активности, аккумулировал 12,8 г/л L-глутаминовой кислоты и одновременно аккумулировал 1,5 г/л αKG.
(6) Введение RSFCPG в штамм Klebsiella planticola, дефицитный по αKGDH, и оценка продуцируемости L-глутаминовой кислоты
Штамм AJ13410 трансформировали плазмидой RSFCPG, и полученный штамм с введенной плазмидой RSFCPG, то есть штамм AJ13410/RSFCPG, инокулировали в колбе объемом 500 мл, содержащей 20 мл культуральной среды, включающей 40 г/л глюкозы, 20 г/л сульфата аммония, 0,5 г/л гептагидрата сульфата магния, 2 г/л дигидрофосфата калия, 0,5 г/л хлорида натрия, 0,25 г/л гептагидрата хлорида кальция, 0,02 г/л гептагидрата сульфата железа, 0,02 г/л тетрагидрата сульфата марганца, 0,72 мг/л дигидрата сульфата цинка, 0,64 мг/л пентагидрата сульфата меди, 0,72 мг/л гексагидрата хлорида кобальта, 0,4 мг/л борной кислоты и 30 г/л карбоната кальция, 200 мг/л моногидрохлорида L-лизина, 200 мг/л L-метионина и 200 мг/л DL-α,ε-DAP, и культивировали, встряхивая, при 37 oС до тех пор, пока глюкоза, содержащаяся в культуральной среде, не была израсходована. После завершения культивирования измеряли количество L-глутаминовой кислоты и αKG, аккумулированных в культуральной среде. Результаты представлены в таблице 4.
В штамме, CS-, РЕРС- и GDH-активности которого были амплифицированы путем введения RSFCPG, наблюдалось снижение аккумулированного количества αKG, а аккумулированное количество L-глутаминовой кислоты было еще более высоким.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОРГАНИЧЕСКАЯ АЗОТСОДЕРЖАЩАЯ КОМПОЗИЦИЯ И УДОБРЕНИЕ, ВКЛЮЧАЮЩЕЕ ЕЕ | 2002 |
|
RU2291139C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2282662C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2194076C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2288271C2 |
| ШТАММ ENTEROBACTER AGGLOMERANS, СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ МИКРООРГАНИЗМА ДЛЯ ВЫРАБОТКИ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2000 |
|
RU2261272C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2003 |
|
RU2307165C2 |
| СПОСОБ ПРОИЗВОДСТВА L-АМИНОКИСЛОТ | 2007 |
|
RU2422530C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2515044C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ СЕМЕЙСТВА ГЛУТАМАТА ИЛИ L-ВАЛИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2009 |
|
RU2418064C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |











Изобретение относится к способу получения L-глутаминовой кислоты. Микроорганизм, принадлежащий к роду Klebsiella, Erwinia или Pantoea и обладающий способностью продуцировать L-глутаминовую кислоту, культивируют в жидкой культуральной среде. Из этой культуральной среды собирают продуцированную L-глутаминовую кислоту. Используемым микробным штаммом является предпочтительно штамм, который обладает пониженной активностью или вообще не обладает активностью фермента, катализирующего реакцию, ответвляющуюся от пути биосинтеза L-глутаминовой кислоты и продуцирующую соединение, не являющееся L-глутаминовой кислотой; или штамм, который обладает повышенной активностью фермента, катализирующего реакцию биосинтеза L-глутаминовой кислоты. Изобретение позволяет повысить продуцирование L-глутаминовой кислоты. 6 з.п. ф-лы, 5 ил., 4 табл.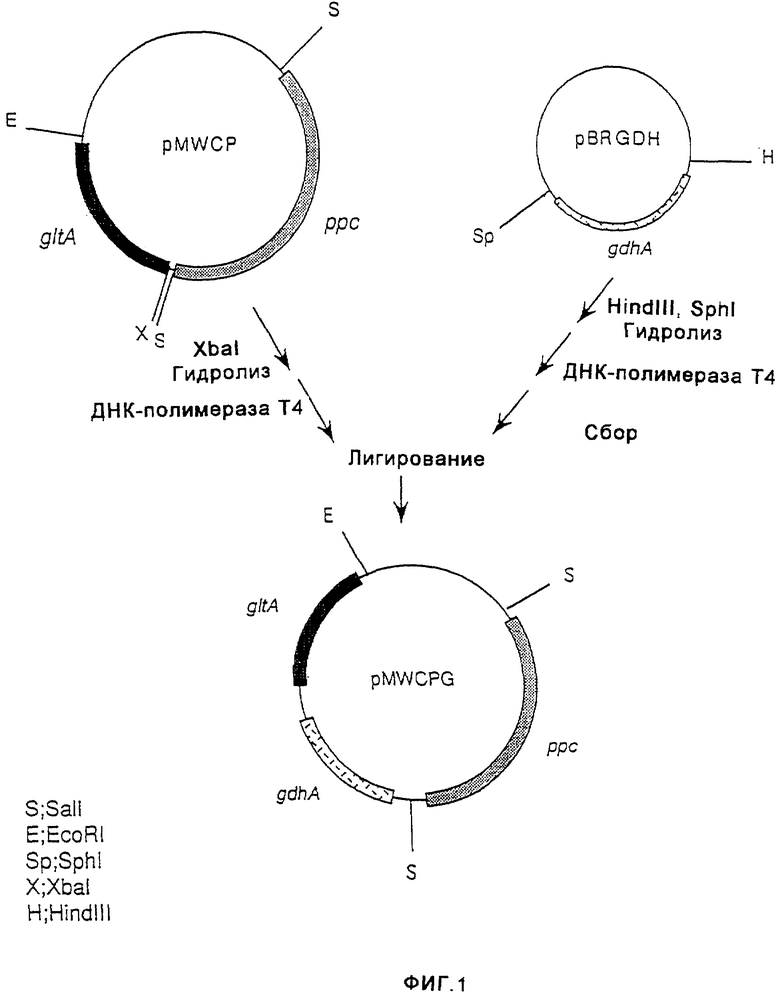
| US 4338302 А, 06.07.1982 | |||
| US 5134077 А, 28.07.1992 | |||
| EP 0780477 А4, 25.06.1997 | |||
| Способ чистовой вырезки | 1973 |
|
SU469517A1 |
| JP 62044171 А, 26.02.1987 | |||
| KR 9705914 А, 22.04.1997 | |||
| ШТАММ БАКТЕРИЙ BREVIBACTERIUM FLAVUM - ПРОДУЦЕНТ L-ГЛУТАМИНА (ВАРИАНТЫ) | 1994 |
|
RU2084520C1 |
| Способ получения -аспарагиновой кислоты | 1977 |
|
SU659611A1 |

