Изобретение относится к молекулам ДНК, в частности из Spinacia oleracea, которые содержат кодирующую область транспортера 2-оксоглутарата/малата и интродукция которых в геном растения изменяет производство и транспорт углеродных скелетов для фиксации азота и транспорт ассимилированного азота в трансгенных растениях. Кроме того, изобретение относится к плазмидам, дрожжам и бактериям, содержащим указанные молекулы ДНК, а также к трансгенным растениям, в которых изменения активности транспортера 2-оксоглутарата/малата и, следовательно, модификации метаболизма азота и углерода осуществляют путем введения указанных молекул ДНК. Кроме того, изобретение относится к трансгенным растениям, на фотореспираторную способность которых воздействуют путем изменения активности транспортера 2-оксоглутарата/малата. Изобретение также относится к применению описанных молекул ДНК, которые кодируют транслокатор 2-оксоглутарат/малат для идентификации родственных транслокаторов из Spinacia oleracea и других растений с использованием методов гибридизации в условиях пониженной жесткости или методов ПЦР, а также к применению транслокатора 2-оксоглутарата/малата в качестве мишени для гербицидов.
Только растения, бактерии и дрожжи обладают способностью к крупномасштабному превращению неорганического азота (нитратного азота) в органически фиксированный азот (обычно в виде аминокислот) путем восстановительного аминирования органических углеродсодержащих соединений. Другие представители царства животных, в частности полезные животные и люди, зависят от растений как первичных поставщиков органических азотсодержащих соединений. Ассимиляция растениями неорганического азота путем связывания с органическим углеродом зависит от доступности азота, углеродных скелетов и энергии.
На поставку азота растению могут оказывать влияние удобрения. Источником энергии для ассимиляции азота является световая реакция фотосинтеза, соответственно в корнях или в других незеленых тканях энергия образуется в результате диссимиляции, а в нормальных полевых условиях она не является лимитирующим фактором.
2-Оксоглутарат (α-кетоглутатрат) в соответствии с современными данными представляет собой первичный акцептор восстановленного азота в реакции глутамин- синтаза/глутамин - 2 - оксоглутарат-аминотрансфераза (GOGAT). В этой реакции азот (азот аммония), восстановленный глутамин -синтазой, сначала переносится на глутамат с поглощением энергии. Образуется глутамин. В следующей реакции глутаматоксоглутарат-аминотрансфераза (GOGAT; глутамат-синтаза) катализирует перенос аминогруппы глутамина на 2-оксоглутарат (трансаминирование), поглощая при этом восстанавливающие эквиваленты. Образуются две молекулы глутамата.
Все последовательные стадии реакции глутамин-синтаза/GOGAT происходят в строме пластид растений. Эти органеллы окружены двумя двухслойными липидными мембранами, причем наружная, имеющая характер молекулярного сита, является приницаемой для соединений размером приблизительно до 10 кДа (Flügge и Benz, 1984, FEBS Lett. 169: 85-89). Внутренняя мембрана проницаема для некоторых более мелких соединений, таких, как вода, двуокись углерода, кислород и нитрит, однако не для молекул с более высоким зарядом, таких, как 2-оксоглутарат.
Ключевое соединение в реакции глутамин-синтаза/GOGAT, т.е. 2-оксоглутарат, должно быть доставлено из цитозоля растительной клетки с помощью специфичного транслокатора через внутреннюю мембрану оболочки хлоропласта в строму пластиды. Транспорт 2-оксоглутарата в пластиды происходит при обмене с малатом из пластид с помощью транслокатора 2-оксоглутарата/малата. Малат, экспортированный во время этого процесса в цитазоль, транспортируется обратно в обмен на глутамат с помощью второго транслокатора, дикарбоксилатного транслокатора, который близок к транслокатору 2-оксоглутарата/малата по своей субстратной специфичности. В результате 2-оксоглутарат импортируется в хлоропласт, а конечный продукт реакции глутамин-синтаза/GOGAT, глутамат, экспортруется без чистого транспорта малата, который циркулирует с помощью обеих систем транслокации ("двойной транслокатор", Woo и др., 1987, Plant Physiol. 84: 624-632; Flügge и др., 1988, Planta 174: 534-541). Глутамат представляет собой предпочтительный для растений донор аминогруппы в сериях реакций трансаминирования, например, при биосинтезе аминокислот аланина или фенилаланина, и т. д. Кроме того, глутамат является важной транспортной формой для органически связанного азота внутри растения. Большинство азотсодержащих соединений в растении, таких, как аминокислоты, нуклеиновые кислоты или алкалоиды, нуждаются в глутамате как в первичном доноре для процесса биосинтеза аминокислот.
2-Оксоглутарат, необходимый для ассимиляции азота, синтезируется в основном путем превращения цитрата в цитоплазме клеток.
В более современных публикациях (Riesmeier и др., 1993, Proc. Natl. Acad. Sci. USA 90: 6160-6164; Heineke и др., 1994, Planta 193: 174-180) говорится о том, что эффективность реакции фотосинтеза углерода помимо прочего в значительной степени ограничена экспортом восстановленного, органически связанного углерода (триозофосфата), образование которого катализируется транслокатором, расположенным во внутренней мембране оболочки хлоропласт. Таким образом, этот протеин-транслокатор представляет собой "узкое место" в метаболизме углерода. Пластидный транслокатор 2-оксоглутарат/малат играет аналогичную роль в метаболизме азота.
Пластидный транслокатор 2-оксоглутарат/малат, таким образом, играет ключевую роль в метаболизме азота в растениях, поскольку он ответственен за доставку достаточных количеств субстрата для ассимиляции азота путем реакции глутамин-синтаза/GOGAT. Путем манипулирования активностью этого транслокатора можно было бы оказывать влияние на эффективность ассимиляции азота в растениях.
Поскольку большинство людей на земном шаре зависит от вегетарианской пищи, что приводит к постоянному неадекватному снабжению протеинами этого слоя общества, то имеется насущная потребность в растениях, имеющих повышенное содержание органических азотсодержащих соединений, в частности протеинов и аминокислот. В промышленно развитых странах в корм животных, предназначенных на убой, во все возрастающих масштабах добавляют животную и рыбную пищу с целью улучшить снабжение протеинами племенных животных. Кормовые растения с высоким содержанием протеинов несомненно были бы лучшей альтернативой, прежде всего с учетом проблем, таких, как BSE, возникающих при питании животной пищей.
На активность пластидного транслокатора 2-оксоглутарата/малата было бы возможно влиять с помощью методов генной инженерии, если бы последовательности ДНК, кодирующие этот транслокатор, были доступными. До сих пор это было не так. Получение последовательностей ДНК, кодирующих транслокатор 2-оксоглутарат/малат, кроме того, должно дать возможность идентифицировать вещества, которые специфично ингибируют этот транслокатор и которые, следовательно, могут применяться в качестве гербицидов.
В настоящее время последовательности протеинов-транслокаторов, субстратная специфичность которых описана выше, известны только в митохондриях сердца быков и в митохондриях человека (Runswick и др., 1990, Biochemistry 29: 11033-11040; Iacobazzi и др., 1992, LNA Seq. 3(2): 79-88). Эти транспортеры играют важную роль в дикарбоксилатном метаболизме в митохондриях (среди прочего в челночном переносе малат/аспартат и челночном переносе оксиглутарат/изоцитрат) и принадлежат к семейству близкородственных транспортеров митохондриальных метаболитов. Например, митохондриальные переносчики (фосфат/ОН-, АДФ/АТФ, оксоглутарат/малат и т.д.) характеризуются взаимосвязью последовательности и наличием внутренних повторов. Кроме того, можно подтвердить, что большинство митохондриальных носителей, равно как и носитель 2-оксоглутарат, включаются в митохондриальную мембрану без препоследовательности (направляющей последовательности), направляющей их к органеллам (Runswick и др., 1990, Biochemistry 29: 11033-11040). Следовательно, можно предположить, что информация о направленности содержится в зрелом протеине-переносчике. Сверхэкспрессия транспортера митохондриального дикарбоксилата в растениях, таким образом, не должна привести к увеличению транспорта оксоглутарата через мембрану оболочки пластиды, а только к увеличению митохондриального дикарбоксилатного транспорта, что является нежелательным эффектом.
В противоположность вышесказанному протеины внутренней мембраны оболочки пластиды нуждаются в препоследовательности, специфично направляющей их к пластидам (см. обзорные статьи Keegstra и др., 1989, Annu. Rev. Plant. Physiol. Plant Mol. Biol. 40: 471-501; Lubben и др., 1988, Photosynth. Res. 17: 173-194; Flügge, 1990, J. Cell Sci. 96: 351-354). В дополнение к препоследовательности, необходимой для "направленности к пластиде", дополнительная информация заложена в зрелой части (части протеина, сохраняющейся после расщепления препоследовательности с помощью специфичной протеазы) протеинов мембраны оболочки пластиды, которая препятствует транспорту протеинов мембраны оболочки через мембрану оболочки в строму пластиды или тилакоидную мембрану (согласно собственным неопубликованным наблюдениям заявителя; Li и др. , 1992, J. Biol. Chem. 267: 18999-19004). До сих пор не представлялось возможным точно установить местонахождение этой информации в зрелой части известных к настоящему времени протеинов внутренней мембраны оболочки (протеина 37 кДа: Dreses-Werringloer и др. 1991, Eur. J. Biochem. 195: 361-368; транслокатора триозофосфат/фосфат: Flügge и др., 1989, EMBO J. 8: 39-46; Willey и др., 1991, Planta 183: 451-461; Fisher и др., 1994, Plant Jour. 5(2): 215-226; Са2+ - АТФазы: Huang и др., 1993, Proc. Natl. Acad. Sci. USA 90: 10066-10070). Собственные исследования заявителя, проведенные с митохондриальным носителем, транспортером АДФ/АТФ, показали, что этот протеин не может или только очень малоэффективно может быть направлен к хлоропластам и включаться в этом месте в мембрану оболочки. Даже гибридный протеин, состоящий из препоследовательности хлоропласта (содержащей информацию для направленности к хлоропласту) и указанного митохондриального носителя, проявил лишь небольшое увеличение с точки зрения включения в мембрану оболочки хлоропласта по сравнению с аутентичным протеином (неопубликованные наблюдения). Поскольку протеин/липидное взаимодействие является важным для правильной инсерции, а мембрана оболочки хлоропласта принципиально отличается по составу липидов от таковой митохондрии (Joyard и др. 1991, Eur. J. Biochem. 199: 489-509), маловероятно, что встраивание протеина митохондрии, хотя и минорного, может быть функциональным, т.е. что транспортер из других органелл может быть включен полностью в мембрану оболочки хлоропласта в конформации и ориентации, соответствующим его функции.
Таким образом, в соответствии с современным состоянием в данной области техники не представляется возможным функционально интегрировать, например, митохондриальный или прокариотический дикарбоксилатные транспортеры во внутреннюю мембрану оболочки хлоропласта, используя известные, направляющие к пластиде последовательности. В соответствии с современным состоянием в данной области техники более перспективным представляется получение последовательности ДНК, необходимой для конструирования описанных выше растений, путем клонирования аутентичного пластидного транслокатора 2-оксоглутарата/малата.
Однако это клонирование не может быть осуществлено путем скрининга в условиях пониженной жесткости библиотеки кДНК растения с использованием зонда, имеющего происхождение из митохондриальных дикарбоксилатных транспортеров. В соответствии с современными данными транслокаторы хлоропласта имеют первичную последовательность, совершенно отличную от всех транслокаторов из других систем (бактерии, митохондрии), даже если протеин имеет близкую функцию. Следовательно, должны быть применены процессы биохимической характеризации, очистки и выделения транслокатора 2-оксоглутарата/малата, которые особенно сложны для мембранных протеинов. В настоящее время возможна идентификация протеина-транслокатора как компонента внутренней мембраны оболочки хлоропласта, имеющего присоединенную молекулярную массу 45000 Да (Menzlaff и Flügge, 1993, Biochim. Biophys. Acta 1147: 13-18). Однако описанная методика очистки непригодна для получения достаточных количеств протеина, необходимых для секвенирования протеина, так как она не учитывает то, что N-концы протеина были блокированы и не пригодны для N-концевого секвенирования протеина путем автоматического разложения Эдмана. Следовательно, до настоящего времени было невозможно выделить, начиная с аминокислотной последовательности, молекулы ДНК, кодирующие пластидный транслокатор 2-оксоглутарат/малат.
Таким образом, задачей, лежащей в основе настоящего изобретения, является создание способов и молекул ДНК, применение которых дает возможность модифицировать растения таким образом, чтобы они приобрели способность синтезировать повышенное количество органических азотсодержащих соединений. В частности задачей, лежащей в основе изобретения, является получение молекул ДНК, кодирующих пластидный транслокатор 2-оксоглутарат/малат.
Эта задача решена с помощью вариантов осуществления изобретения, представленных в формуле изобретения.
Таким образом, изобретение относится к молекулам ДНК, кодирующим пластидный транслокатор 2-оксоглутарат/малат, причем информация, содержащаяся в нуклеотидной последовательности, приводит к формированию рибонуклеиновой кислоты после интродукции и экспрессии в растительных клетках, что позволяет интродуцировать активность транслокатора 2-оксоглутарата/малата в клетки с помощью этой рибонуклеиновой кислоты или путем подавления эндогенной активности транслокатора 2-оксоглутарата/малата.
В частности настоящее изобретение относится к молекулам ДНК, кодирующим протеин с аминокислотной последовательностью, представленной в SEQ ID N 2, а также к молекулам ДНК, содержащим кодирующую область, приведенную в SEQ ID N 1. Кроме того, оно относится к молекулам ДНК, которые гибридизируются с вышеуказанными молекулами ДНК по изобретению, и к молекулам ДНК, последовательность которых вырождена вследствие генетического кода, причем эти последние молекулы кодируют протеин, обладающий активностью транслокатора 2-оксоглутарата/малата. Еще одним предметом изобретения являются молекулы ДНК, которые комплементарны таковым, указанным выше, а также фрагменты и производные вышеуказанных молекул ДНК по изобретению, которые могут быть получены в результате инсерций, замещений или делеций и которые кодируют транслокатор 2-оксоглутарат/малат.
Термин "гибридизация" в контексте настоящего изобретения означает гибридизацию в обычных условиях, таких, как описаны, например, у Sambrook и др. (1989, Molecular Cloning, A Laboratory Manual, 2-е изд.. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY), предпочтительно гибридизацию в строгих условиях (как это описано у Sambrook и др., см. выше).
Молекулы ДНК (или их производные либо части этих молекул) по настоящему изобретению могут быть интродуцированы в плазмиды, что дает возможность осуществить мутагенез или модификацию последовательности путем рекомбинации последовательностей ДНК в системах прокариот или эукариот, тем самым позволяя модифицировать специфичность транслокатора 2-оксоглутарата/малата. Путем модификации специфичности транслокатора в направлении усиления специфичности по отношению к глутамату и одновременного снижения специфичности к малату описанная выше система "двойного транслокатора" может быть модифицирована так, чтобы достигалось дальнейшее улучшение скоростей транспорта 2-оксоглутарата при направленном обмене на глутамат. Также может быть достигнута нечувствительность к гербицидам, специфичным для транслокатора 2-оксоглутарата/малата. Стандартные методы Sambrook и др. (1989, Molecular Cloning, A Laboratory Manual, 2-е изд.. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY), могут быть использованы для осуществления обмена основаниями и/или делении оснований и/или добавления синтетических или натуральных последовательноcтей. Для внутреннего сцепления фрагментов ДНК могут быть использованы адаптеры или линкеры. Кроме того, могут применяться операции, не требующие обеспечения ДНК пригодными сайтами рестрикции или делениями. Когда осуществимы инсерции, делеции или замещения, такие, как транзиции или трансверсии, то могут быть использованы мутагенез, праймерная репарация, рестрикция или лигирование in vitro. Методы, которые применяют для анализа, обычно включают анализ последовательности, рестрикционный анализ и другие биохимические методы молекулярной биологии, такие, как экспрессия модифицированного протеина в почкующихся дрожжах и оценка измененных транспортных способностей на искусственных липосомах (см. пример 4 и Loddenkötter и др., 1993, Proc. Natl. Acad. Sci. USA 90: 2155-2159; а также у Fischer и др., 1994, Plant Journal 5: 215-226) или измерение измененных транспортных свойств протеина, экспрессированного в трансгенных растениях с использованием метода, только в последнее время адаптированного для этих целей заявителями (Flügge и Weber, 1994, Planta, 194: 181-185).
Молекулы ДНК по изобретению (или части либо производные этих молекул) могут быть использованы для выделения с помощью стандартных методик из генома растений сходных молекул ДНК, которые также кодируют транслокатор 2-оксоглутарат/малат или аналогичный дикарбоксилатный транслокатор. Наиболее пригодными методами являются скрининг библиотек кДНК путем гибридизации в условиях пониженной жесткости с использованием молекулы ДНК по изобретению или ее части в качестве зондов или конструкции таких зондов для стратегий скрининга в жестких условиях и в условиях пониженной жесткости с помощью получения вырожденных и/или невырожденных праймеров из последовательностей молекул ДНК по изобретению для ПЦР-экспериментов с ДНК или с кДНК шпината или других растений. Молекулы ДНК также могут быть использованы для идентификации и выделения молекул ДНК, которые кодируют близкородственный транслокатор глутамат-малат (дикарбоксилатный транслокатор) из мембраны оболочки пластиды (субтрактивные методы скрининга библиотеки кДНК из шпината или других растений в условиях различной жесткости и с использованием различных областей молекул ДНК по изобретению в качестве зонда).
В другом варианте осуществления настоящее изобретение относится к применению молекул ДНК по изобретению или их частей либо производных, полученных из этих молекул ДНК с помощью инсерции, делеции или замещения, для трансформации клеток про- и эукариот. Чтобы гарантировать экспрессию транслокатора 2-оксоглутарата/малата в трансформированных клетках, молекулы ДНК по изобретению могут быть интродуцированы в векторы, предпочтительно в плазмиды, и могут быть объединены с контролирующими элементами для экспрессии в клетках прокариот или эукариот (см. примеры 3 и 5). Такие контролирующие элементы представляют собой промоторы транскрипции или терминаторы транскрипции. Векторы могут применяться для трансформации клеток эукариот с целью экспрессии транслируемой матричной (информационной) рибонуклеиновой кислоты (мРНК), которая дает возможность синтезировать пластидный транслокатор 2-оксоглутарат/малат в трансформированных клетках, или с целью экспрессии нетранслируемой, обратно ориентированной (антисмысловая ориентация) матричной рибонуклеиновой кислоты, которая препятствует синтезу эндогенных транслокаторов 2-оксоглутарата/малата. Для этой цели также могут применяться более короткие фрагменты молекул ДНК по изобретению или молекулы ДНК, последовательности которых имеют относительно высокую степень гомологии (гомологию более чем на 65%) с последовательностями молекул ДНК по изобретению. Аналогично этому экспрессия эндогенных дикарбоксилатных транслокаторов может быть подавлена с помощью экспрессии сконструированных для этой цели рибозимов с использованием молекул ДНК по изобретению.
Таким образом, согласно еще одному варианту осуществления настоящее изобретение относится к рекомбинантным молекулам ДНК, например, к плазмидам, содержащим одну из молекул ДНК по изобретению, например, к плазмидам pBinAR-211 (DSM 9239), pEVP11-211 (DSM 9237) и pBSC-211 (DSM 9238).
Предметом изобретения являются, в частности, рекомбинантные молекулы ДНК, в которых молекулу ДНК по изобретению сцепляют с последовательностями ДНК, позволяющими осуществлять экспрессию в клетках про- и эукариот.
Согласно другому варианту осуществления настоящее изобретение относится к бактериям, содержащим молекулы ДНК по изобретению или рекомбинантные молекулы ДНК по изобретению.
Кроме того, настоящее изобретение относится к обладающим активностью транслокатора 2-оксоглутарата/малата протеинам, которые кодируются последовательностью ДНК одной из молекул ДНК по изобретению. Предпочтительными протеинами являются таковые из двудольных или однодольных растений, предпочтительно из растений семейства Chenopodiaceae (маревых) и особенно предпочтительно из Spinacia oleracea.
Производство таких протеинов может быть достигнуто в том случае, когда, например, молекулы ДНК по изобретению, которые кодируют транслокатор 2-оксоглутарат/малат, сцепляют с последовательностями ДНК, которые обеспечивают транскрипцию в клетках про- или эукариот. Полученные рекомбинантные молекулы затем могут быть интродуцированы и экспрессированы в пригодных клетках-хозяевах про- или эукариот. Полученный протеин может быть выделен в соответствии с известными методами. Можно также экспрессировать укороченный протеин, который не содержит сигнальных последовательностей, дающих возможность расположить протеин на внутренней мембране оболочки пластид.
Согласно еще одному, предпочтительному варианту осуществления настоящее изобретение относится к применению молекулы ДНК по изобретению для экспрессии транслокатора 2-оксоглутарата/малата в растительных клетках.
Путем экспрессии РНК, соответствующей молекулам ДНК по изобретению, кодирующим в растении транслокатор 2-оксоглутарата/малата, можно модифицировать метаболизм азота в растении. Экономическая важность этого факта состоит в том, что улучшение транспорта 2-оксоглутарата из цитозоля в пластиду приводит к изменению отношения углеводов (сахар, крахмал, органические кислоты) и жиров в пользу азотсодержащих соединений (аминокислот, протеинов, возможно алкалоидов). Таким путем могут быть получены растения, которые более богаты ценным протеином, но имеют пониженное содержание углеводов и жиров. Это изменение повышает питательную ценность растений, а также их экономическую ценность.
Методы генетической модификации двудольных и однодольных растений известны (Gasser и Fraley, 1989, Science 244: 1293-1299; Potrykus, 1991, Ann. Rev. Plant Mol. Biol. Plant Physiol. 42: 205-225). Для экспрессии кодирующих последовательностей в растении они должны быть сцеплены с элементами, регулирующими транскрипцию. Такие элементы, так называемые промоторы, известны (см. , в частности, у Köster-Töpfer и др., 1989, Mol. Gen. Genet. 219: 390-396). Кроме того, кодирующие области должны быть обеспечены сигналом окончания транскрипции, чтобы они могли быть правильно транскрибированы. Такие элементы также уже описаны (Gielen и др., 1989, EMBO J. 8, 23-29). Область начала транскрипции может быть как нативной или гомологичной, так и чужеродной или гетерологичной для растения-хозяина. Области терминации являются полностью взаимозаменяемыми. Последовательность ДНК областей начала транскрипции и окончания транскрипции может быть получена искусственно или получена из естетственных источников либо может содержать смесь синтетических и натуральных компонентов ДНК.
Для осуществления интродукции чужеродных генов в высшие растения приемлемо большое число векторов клонирования, которые содержат сигнал репликации для Е. соli и маркер, позволяющий осуществлять селекцию трансформированных клеток. Примерами таких векторов являются pBR322, серии pUC, серии M13mp, pACYC184 и т.д. В зависимости от метода введения требуемых генов в растения могут оказаться необходимыми другие последовательности ДНК. Если, например, для трансформации растения применяют Ti- или Ri-плазмиду, то по крайней мере правая пограничная область Т-ДНК Ti- и Ri-плазмиды, хотя часто как правая, так и левая пограничные области, должна быть присоединена в качестве фланкирующей области к интродуцируемым генам. Применение Т-ДНК для трансформации растительных клеток было тщательно исследовано и достаточно полно описано (см. Hoekema в The Binary Plant Vector System, Offsetdrukkerij Kanters B-V. Ablasserdam, 1985, раздел 5; Fraley и др., Critic. Rev. Plant Sci. 4:1-46; An и др., 1985, EMBO J. 4: 277-287). Если интродуцированная ДНК интегрирована в геном, то как правило эта интеграция стабильна и сохраняется даже в следующих поколениях первоначально трансформированных клеток. Она обычно содержит маркер селекции, который придает трансформированным клеткам растения устойчивость к биоцидам или антибиотикам, таким, как канамицин, блеомицин или гигромицин. Таким образом, индивидуально подобранный маркер должен позволять отличать трансформированные клетки от клеток, которые лишены интродуцированной ДНК.
Для введения ДНК в растительную клетку-хозяина существует много других методик, отличных от трансформации с использованием агробактерий. Эти методики включают трансформацию протопластов, микроинъекции ДНК, электропорацию, а также баллистические методы. Далее целые растения могут быть выращены из трансформированного растительного материала в пригодной для этой цели селективной среде. Растения, полученные таким образом, могут быть исследованы с помощью традиционных методов молекулярной биологии на наличие интродуцированной ДНК. Эти растения могут быть выращены обычным образом и скрещены с растениями, имеющими такой трансформированный генетический код или другие генетические коды. Образовавшиеся в результате гибридные особи обладают соответствующими фенотипическими свойствами.
Таким образом, настоящее изобретение также относится к трансгенным растительным клеткам, содержащим рекомбинантные молекулы ДНК по изобретению, а также к трансгенным растениям, которые могут быть размножены из этих трансгенных растительных клеток, и к трансгенным растениям, содержащим растительные клетки по изобретению. Эти растительные клетки и растения характеризуются тем, что они содержат рекомбинантную молекулу ДНК, интегрированную в геном, что дает возможность экспрессировать протеин, обладающий активностью транслокатора 2-оксоглутарата/малата, или экспрессировать нетранслируемую молекулу РНК, подавляющую синтез транслокаторов 2-оксоглутарат/малат. Кроме того, настоящее изобретение относится к семенам трансгенных растений по изобретению.
Согласно другому варианту осуществления настоящее изобретение относится к гетерологичной экспрессии молекул ДНК по изобретению в грибах, например, в почкующихся дрожжах (см. пример 3, 4 и Loddenkötter и др., 1993, Рrос. Natl. Acad. Sci. USA 90: 2155-2159), в частности для исследований зависимости структура-функция транслокатора 2-оксоглутарата/малата, что может привести к разработке специфичного ингибитора для этого протеина. В этом контексте также возможна разработка гербицидов, поскольку ингибирование протеина, играющего ключевую роль в метаболизме, неизбежно должно быть летальным для растения.
Следовательно, молекулы ДНК по изобретению (или части этих молекул либо производные этих молекул) могут быть введены в векторы и могут быть снабжены контролирующими элементами для экспрессии в клетках грибов, в частности в почкующихся дрожжах (см. пример 3). Введение транслокатора 2-оксоглутарата/малата приводит к существенному увеличению активности транслокатора 2-оксоглутарата/малата в рекомбинантных дрожжевых клетках, активность в которых может быть измерена с помощью восстановления в искусственных липосомах. В этом контексте следует отметить, что митохондрии несут транслокатор 2-оксоглутарат/малат, который близок по своей субстратной специфичности к таковому пластид, но который, однако, не близок к пластидному транслокатору в отношении своей последовательности ДНК и аминокислотной последовательности (Runswick и др., 1990, Biochemistry 29: 11033-11040). В рекомбинантных клетках дрожжей обнаружена более высокая транспортная активность малата, достигающая 100-кратного уровня. Следовательно, представляется возможным, используя молекулы ДНК по изобретению (или части либо производные этих молекул), изменить клетки дрожжей таким образом, чтобы они имели измененное содержание протеина. Для этой цели может быть использовано преимущество пластидного транслокатора из растений, а именно, такого, который не является объектом для эндогенной регуляции и механизмов направленности к компартаметам у почкующихся дрожжей. Высока вероятность того, что описанное увеличение транспортной активности малата в клетках дрожжей окажется невозможным с помощью экспрессии митохондриального транслокатора 2-оксоглутарата. Эти штаммы могли бы иметь особенное важное значение для индустрии продуктов питания.
Таким образом, изобретение также относится к клеткам грибов, в частности к дрожжевым клеткам, содержащим молекулу ДНК по изобретению или рекомбинантную молекулу ДНК.
Молекула ДНК по изобретению содержит области в кодирующих областях, которые способны специфично направлять синтезируемый на рибосомах в цитоплазме протеин к пластидам и препятствовать нахождению протеина в других мембранных системах клетки. Область протеина, которая направляет кодируемый молекулами ДНК по изобретению протеин к пластидам, расположена внутри первой сотни аминокислот протеина, она не требуется для транспортной функции протеина, и ее удаляют после успешной инсерции протеина в мембрану оболочки хлоропласта. Путем замены этой направленной по отношению к пластиде последовательности на одну из известных направляющих последовательностей, например, на митохондриальную, протеин-транслокатор может быть ориентирован к другой мембранной системе клеток эукариот и может иметь возможность изменять в этом месте транспортные свойства через соответствующую мембрану. Аналогично этому направляющая к пластиде последовательность транслокатора 2-оксоглутарата/малата, в частности первые 100 аминокислот, или эндогенные области зрелого протеина, могут применяться для ориентации чужеродных протеинов (например, протеинов, катализирующих активный или пассивный транспорт метаболитов через мембраны, ферментов, бактериальных транспортных протеинов или транспортеров из дрожжей) к пластидам, в частности к мембране оболочки пластиды, к строме пластиды или к тилакоидам растительных клеток.
9 июня 1994 г. в Немецкой коллекции микроорганизмов (German Collection of Microorganisms) (DSM) в Брунсвике, Федеративная Республика Германия, признанной в качестве международного депозитария в соответствии с требованиями Будапештского договора, были депонированы следующие плазмиды в штаммах Е. coli:
плазмида pBinAR-211 (DSM 9239);
плазмида pEVP11-211 (DSM 9237);
плазмида pBSC-211 (DSM 9238).
Описание чертежей
Фиг. 1: Восстановление транспортной активности рекомбинантного транслокатора 2-оксоглутарата/малата, экспрессированного в дрожжах. На чертеже показано поглощение [14С] малата в восстановленных липосомах, содержащих клеточные мембраны S. pombe.
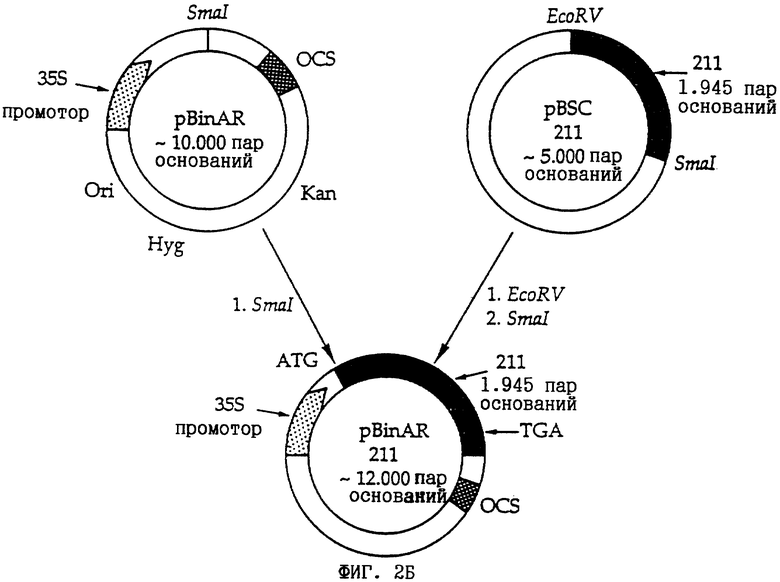
Фиг. 2: Схематическое изображение клонирующей плазмиды pEVP11-211 (фиг. 2А) и плазмиды pBinAR-211 (фиг.2Б).
Фиг. 3: Направленность предшественника протеина транслокатора 2-оксоглутарата/малата к хлоропластам и зависящая от энергии инсерция зрелого протеина во внутреннюю мембрану оболочки.
Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (ДСН) осуществляли по описанной в примере 6 методике:
Полоса 1: транслированный in vitro предшественник протеина.
Полосы 2-4: в условиях темноты.
Полосы 5-9: в условиях освещенности.
Полоса 2: без АТФ.
Полоса 3: плюс 2 мМ АТФ.
Полоса 4: плюс 2 мМ АТФ, последующая обработка протеазой (100 мкг/мл).
Полоса 5: без АТФ.
Полоса 6: плюс 2 мМ АТФ.
Полоса 7: плюс 2 мМ АТФ, последующая обработка протеазой (100 мкг/мл).
Полоса 8: плюс агент десопряжения (10 мкМ карбонилцианид-м-хлорфенилгидразон (КЦХФ)).
Полоса 9: плюс 2 мМ АТФ; предварительно обработанные протеазой хлоропласты (30 мкг/л).
С целью пояснения иллюстрирующих настоящее изобретение примеров некоторые из использованных методов объяснены ниже более подробно.
1. Способы клонирования
Для клонирования использовали фаг лямбда gt10, а также вектор pBluescript II KS (pBSC) (Short и др., 1988, Nucl. Acids Res. 16: 7583-7600).
Для трансформации дрожжей применяли вектор pEVP11 (Russel и Nurse, 1986, Cell 45: 145-153).
Для трансформации растений генные конструкции клонировали в бинарном векторе pBinAR (Höfgen и Willmitzer, 1990, Plant Sci. 66: 221-230).
2. Штаммы бактерий и дрожжей
Для вектора pBluescriptKS (pBSC), а также для конструкций pEVP11 и pBinAR применяли штаммы E.coli DH5α (Hanahan и др., 1983. J. Mol. Biol. 166: 557-580) и TG1 (Gibson, 1984, Ph.D. Thesis, Cambridge University, Англия).
Трансформацию конструкций pBinAR в растениях табака осуществляли с помощью штамма Agrobacterium tumefaciens LBA4404 (Bevan, 1984, Nucl. Acids Res. 12: 8711-8720).
3. Трансформация с использованием Agrobacterium tumefaciens
Перенос ДНК в Agrobacteria осуществляли с помощью направленной трансформации в соответствии с методом Höfgen и Willmitzer (1988, Nucl. Acids Res. 16: 9877). ДНК плазмиды трансформированной Agrobacteria выделяли в соответствии с методом Birnboim и Doly (1979, Nucl. Acids Res. 7: 1513-1523) и анализировали с помощью гель-электрофореза на правильность и ориентацию после соответствующего рестрикционного разложения.
4. Трасформация растений
Для трансформации 15 маленьких листьев стерильной культуры табака, которые были повреждены наждачной бумагой и скальпелем, помещали в 10 мл MS-среды с добавлением 2% сахарозы, содержащей 100 мкл выращенной в строго селективных условиях, трансформированной в течение ночи культуры Agrobacterium tumefaciens. После осторожного встряхивания в миксере в течение 15 минут листья помещали в MS-среду, содержащую 1,6%-ную глюкозу, 2 мг/л зеатин-рибозы, 0,02 мг/л нафтилуксусной кислоты, 0,02 мг гибберелловой кислоты, 500 мг/л Betabactyl®, 15 мг/л гигромицина и 0,8%-ный бактоагар. После инкубации при 25oС при освещенности 3000 люкс в течение одной недели концентрацию Betabactyl в среде снижали в два раза.
Ниже изобретение проиллюстрировано на примерах.
Пример 1
Выделение пептидных фрагментов транслокатора 2-оксоглутарата/малата и приготовление зондов для скрининга путем гибридизации библиотек кДНК
Очищенный протеин транслокатора 2-оксоглутарата/малата (Menzlaff и Flügge, 1993, Biochem. Biophys. Acta 1147: 13-18) отделяли от оставшихся примесей с использованием препаративного электрофореза в ДСН-полиакриаламидном геле (Laemmli, 1970, Nature 227: 680-685), вырезали из геля после обнаружения протеина с помощью окрашивания сульфатом меди(II) (Lee и др., 1987, Anal. Biochem. 166: 308-312) и разлагали в гелевом матриксе эндопротеиназой LysC (Eckerskorn и Lottspeich, 1989, Chromatographia 28: 92-94). Полученные пептиды элюировали из геля и разделяли с использованием ЖХВД. Аминокислотную последовательность очищенных пептидных фракций определяли в газовой фазе с помощью автоматического разложения Эдмана (Eckerskorn и др., 1988, Eur. J. Biochem. 176: 509-519). Вырожденные олигонуклеотидные последовательности, кодирующие эти аминокислоты, имели происхождение из аминокислоты трех пептидов, а соответствующие олигонуклеотиды получали с помощью синтеза ДНК in vitro. Для применения в качестве зонда олигонуклеотиды радиоактивно метили путем присоединения 32Р-фосфатной группы к 5'-концу с помощью олигонуклеотид-киназы.
Пример 2
Клонирование транслокатора 2-оксоглутарата/малата из шпината
Из молодых листьев ростков шпината, выращенных на гидрокультуре, выделяли поли-А+-РНК и на ее основе конструировали библиотеку кДНК в векторе лямбда gt10 (Flügge и др. , 1989, ЕМВО J. 8: 39-46). Приблизительно 300000 клонов этой библиотеки подвергали скринингу с помощью синтетических олигонуклеотидов, которые создавали по образцу очищенных пептидных фрагментов транслокатора 2-оксоглутарата/малата, полученных с использованием эндопротеиназы LysC (см. пример 1). Положительно реагирующие клоны очищали с помощью стандартных методик и после получения амплифицированной ДНК фага из очищенных бляшек получали путем рестрикционного разложения с помощью EcoRI инсерцию (вставку), кодирующую транслокатор 2-оксоглутарат/малат, и проверяли его с помощью анализа с использованием саузерн-блоттинга, применяя вышеуказанные олигонуклеотиды в качестве зонда. После повторного клонирования ДНК-вставок фага в векторе pBluescript (pBSC) клоны анализировали, определяя последовательность ДНК (дидезокси-метод: Sanger и др., 1977, Proc. Natl. Acad. Sci. USA 74: 5463-5467), и из этой последовательности ДНК получали первичную структуру транслокатора 2-оксоглутарата/малата. Последовательность олигонуклеотидов или пептидов, использованная для скрининга библиотеки кДНК, может быть восстановлена.
Пример 3
Экспрессия транслокатора 2-оксоглутарата/малата из шпината в почкующихся дрожжах Schizosaccharomyces pombe
Вышеуказанную плазмиду pBluescript (pBSC), содержащую вставку, кодирующую транслокатор 2-оксоглутарат/малат, линеаризировали с помощью эндонуклеазы SaiI и полученные "липкие концы" заполняли ферментом ДНК-полимеразой фага Т4. Далее вставку отрезали от вектора путем другого рестрикционного разложения с помощью BamHI и выделяли путем электрофореза. Полученный таким образом фрагмент встраивали в определенной ориентации в применяемый для дрожжей вектор экспрессии pEVP11, который предварительно линеаризировали с помощью SacI, концы заполняли Т4-ДНК-полимеразой и затем разлагали вновь BamHI (см. также фиг.2А) и после амплификации конструкции в E.coli трансформировали в клетках S. pombe с дефицитом синтеза лейцина, которые делали компетентными с помощью LiCl/ПЭГ (Ito и др.. 1983, J. Bact. 153: 163-168). Трансформантов отбирали без лейцина путем селекции на минимальной основной среде, так как конструкция pEVP11-211 дает клеткам дрожжей способность расти на среде, лишенной лейцина.
Пример 4
Измерение активности транспорта малата в рекомбинантных линиях дрожжей
Клетки дрожжей, трансформированные с использованием плазмиды pEVP11-211 (см. фиг. 2А, клетки SP-DC3), выращивали в минимальной основной среде до оптической плотности 1,0 при 600 нм и собирали путем центрифугирования при 3000xg в течение 5 минут. Клетки разрушали путем интенсивного встряхивания смеси со стеклянными шариками, объем которых составлял 1/2 объема (по отношению к клеткам), и разделяли стеклянные шарики и клеточный дебрис путем центрифугирования (600xg в течение 1 мин). Супернатант доводили до концентрации 0,5% (масса/объем) с помощью Тритона Х-100, к смеси добавляли равный объем липосом и полученные протеолипосомы немедленно замораживали в жидком азоте. Липосомы предварительно получали путем обработки звуком фосфолипидов бобов сои (20 мг/мл в течение 10 минут при 4oС в присутствии 200 мМ трицин-NaOH (рН 7,6), 40 мМ малата и 60 мМ глюконата калия). После оттаивания протеолипосом и обработки суспензии звуком (10 импульсов в 1 с) протеолипосомы отделяли от окружающей среды с помощью гель-фильтрации на Сефадексе G-25, который был уравновешен с помощью 10 мМ трицин-NaOH (рН 7,6), 100 мМ глюконата натрия и 50 мМ глюконата калия. Элюированные протеолипосомы применяли для измерения активности транспорта малата. Измерение осуществляли в соответствии с методом "стоп-ингибитора", описанного у Menzlaff и Flügge (1993, Biochem. Biophys. Acta 1147: 13-18).
Активность транспорта малата в pEVP11-211-трансформантах сравнивали с активностью транспорта малата в трансформантах, которых трансформировали только с использованием вектора pEVP11 без 211-вставки. Было установлено, что активность транспорта малата в pEVP11-211-трансформантах (измеренная в пмолях транспортированного 14С-малата/мг протеина в час) была в 100 раз выше, чем в контрольных pEVP11-трансформантах (фиг.1). Кроме того, можно подтвердить, что рекомбинантный протеин-транслокатор проявляет идентичные транспортные характеристики по сравнению с аутентичым протеином мембраны хлоропласта. Эти данные приведены в табл.1.
Пример 5
Трасформация растений с помощью конструкции для сверхэкспрессии кодирующей области транслокатора 2-оксоглутарата/малата
Из вектора pBluescript-211 (pBSC-211), который содержал в качестве вставки кДНК транслокатора 2-оксоглутарата/малата из шпината (см. пример 2, фиг. 2Б), вставку выделяли путем рестрикционного разложения с помощью EcoRV и SmaI и клонировали в векторе pBinAR (Höfgen и Willmitzer, 1990, Plant Sci. 66: 221-230), который разлагали ферментом SmaI. После амплификации полученной конструкции pBinAR-211 в Е. coli конструкцию трансформировали в агробактерии и последнюю применяли для заражения сегментов листьев табака и картофеля.
Полученные трансформанты подвергали скринингу с использованием анализов саузерн-блоттинга в присутствии интактного, не перегруппированного химерного гена. Используя "метод восстановления целого листа" (Flügge и Weber, 1994, Planta, 194: 181-185), оценивали транспортную активность малата, сравнивая с таковой у контрольных трансформантов (трансформированных вектором pBinAR без вставки), а также отношение C/N, скорость фотосинтеза, транспирацию и рост.
Пример 6
Направленность протеина-предшественника транслокатора 2-оксоглутарата/малата к хлоропластам и зависящая от энергии инсерция зрелого протеина во внутреннюю мембрану оболочки
Транскрипцию in vitro плазмиды pBSC-211, которая была линеаризована с помощью SmaI, осуществляли с использованием Т3-РНК-полимеразы в соответствии с инструкциями производителя (фирма Pharmacia). Последующую трансляцию in vitro осуществляли в лизате ретикулоцита (Boehringer-Mannheim). Для транспортов протеинов в интактные хлоропласты шпината применяли пост-рибосомный супернатант. Эксперименты проводили как в темноте, так и на свету; препарат содержал буфер для импорта (Flügge и др.. 1989, EMBO J. 8: 39-46), интактные хлоропласты шпината (соответвующие 200 мг хлорофилла) и различные аддитивы, указанные в описании к фигуре 3 (полный объем 300 мл). После выдерживания в течение 15 минут при 25oС хлоропласты промывали и выделяли мембраны оболочки (Flügge и др. , 1989, EMBO J. 8: 39-46). Их анализировали с использованием электрофореза в ДСН-полиакриламидном геле (Laemmli, 1970, Nature 227: 680-685) и последующей флуорографии (Bonner и Laskey, 1974, Eur. J. Biochem. 46: 84-88) (фиг.3). Полоса 1 соответствует транслированному in vitro протеину-предшественнику (р). Полосы 2-4 соответствуют опытам, проведенным в условиях темноты, полосы 5-9 - опытам, проведенным в условиях освещенности. В любых условиях препоследовательность транслокатора 2-оксоглутарата/малата направляет связанный зрелый протеин точно к его мембране-мишени, т.е. внутренней мембране оболочки хлоропластов; его отщепляют во время процесса импорта с помощью специфичной протеазы; получают зрелый протеин (m). В темноте встраивание транслокатора осуществляют с помощью дополнительной энергии, поставляемой АТФ (полосы 3 и 4). При отсутствии АТФ (полоса 1) импорт не обнаружен. На свету энергия, необходимая для импорта протеина, может быть получена в виде АТФ путем фотосинтетического фосфорилирования; импорт в этих условиях не зависит от экзогенного АТФ (полоса 5), но может быть усилен при добавлении АТФ извне (полоса 6). Если фотосинтетическому фосфорилированию и получению в результате этого АТФ препятствовать путем добавления агента десопряжения, такого, как карбонилцианид-м-хлорфенилгидразон (КЦХФ), то импорт протеина будет блокирован (полоса 8). Зрелый протеин присутствует включенным во внутреннюю мембрану: добавление протеаз (например, термолизина), которые не обладают способностью проникать через внешнюю мембрану оболочки, не может оказать влияния на зрелый протеин, включенный во внутреннюю мембрану (линии 4 и 7). Предварительная обработка хлоропластов протеазой (например, термолизином) приводит к полной потере связывания и импорта транслокатора (полоса 9). Это свидетельствует о том, что препоследовательность (направляющая последовательность) транслокатора на первой стадии нуждается в специфичном связывании с рецепторами, локализованными на наружной мембране. Только после этого могут быть осуществлены следующие стадии встраивания протеина.










Изобретение относится к генной инженерии и может быть использовано в селекции бактерий, дрожжей и растений. Введение молекул ДНК, кодирующих пластидный транслокатор 2-оксоглутарат/малат, в генном про- или эукариот изменяет в них метаболизм азота и углерода. В частности, молекулы ДНК из Spinacia oleracia способны модифицировать продуцирование и транспорт углеродных скелетов для фиксации азота и транспорт ассимилированного азота в растениях, а также влияет на их фотореспираторную способность. 5 с. и 7 з.п. ф-лы, 4 ил., 1 табл.
| Экономайзер | 0 |
|
SU94A1 |
| Экономайзер | 0 |
|
SU94A1 |
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| АВТОМАТИЧЕСКИЙ ФИКСАТОР ШТОКА ТОРМОЗНОГО ЦИЛИНДРА В ПОЛОЖЕНИИ ТОРМОЖЕНИЯ | 0 |
|
SU255340A1 |

