Область техники, к которой относится изобретение
Изобретение относится к генной инженерии растений, в частности к трансформации пластид высших растений. Изобретение относится к новым промоторным последовательностям, обеспечивающим экспрессию представляющих интерес чужеродных генов в различных видах растений.
Предпосылки создания изобретения
Гены хлоропласта транскрибируются с участием РНК-полимеразы, содержащей кодируемые пластидной ДНК субъединицы, гомологичные α-, β- и β′ -субъединицам РНК-полимеразы Е. соli. Промоторы, используемые для экспрессии этого фермента, аналогичны σ70-промоторам Е. coli, состоящим из консенсусных элементов -35 и -10 (G.L. Igloi и Н. Kossel, Crit. Rev. Plant Sci., 10: 525, 1992; W. Gruissem и J.C. Tonkyn, Crit. Rev. Plant ScL, 12, -19, 1993). Выбор промотора для РНК-полимеразы, кодируемой пластидной ДНК, зависит от кодируемых ядерной ДНК сигмаподобных факторов (Link и др., 1994, Plant promoters and transcription factors, изд-во Springer Verlag, Heidelberg, стр. 63-83). Кроме того, транскрипционная активность некоторых промоторов модулируется кодируемыми ядерной ДНК факторами транскрипции, взаимодействующими с элементами, расположенными против хода транскрипции ядерного промотора (LA. Allison и Р. Maliga, EMBO J., 14: 3721-3730; R. Iratni, L. Baeza, A. Andreeva, R. Mache, S. Lerbs-Mache, Genes Dev., 8, 2928, 1994). Эти факторы обусловливают ядерный контроль экспрессии пластидных генов в ответ на сигналы, связанные с процессом развития и с окружающей средой.
Умозрительно было сделано предположение о существовании в пластидах второй системы транскрипции. Однако к настоящему времени не получено прямого доказательства, подтверждающего это предположение. Выявление в пластидах новой второй системы транскрипции представляет собой важное достижение в области генной инженерии растений. Такая система позволяет увеличить пластичность и расширить диапазон видов растений, у которых можно трансформировать пластиды, и облегчает тканеспецифическую экспрессию чужеродных протеинов и РНК посредством конструкций, которыми можно манипулировать с помощью методов рекомбинантной ДНК.
Краткое изложение сущности изобретения
Настоящее изобретение относится к конструкциям ДНК и к способам стабильной трансформации пластид многоклеточных растений. Конструкции ДНК по изобретению расширяют диапазон видов растений, которые могут быть трансформированы, и облегчают тканеспецифическую экспрессию представляющих интерес чужеродных генов.
В соответствии с первым вариантом осуществления изобретения предложены конструкции ДНК, содержащие новые промоторные последовательности, которые распознаются кодируемой ядерной ДНК пластидной [NEP = nuclear encoded plastid] РНК-полимеразой. Конструкция ДНК содержит трансформирующую ДНК, которая включает ориентирующий сегмент, по крайней мере один сайт клонирования, адаптированный для встраивания (инсерции) по крайней мере одного представляющего интерес чужеродного гена, причем экспрессия представляющего интерес чужеродного гена регулируется промотором, который распознается NEP-полимеразой, и пластидный селектируемый ген-маркер.
Согласно второму варианту осуществления настоящего изобретения предлагается применение элементов промотора, распознаваемых кодируемой пластидной ДНК пластидной [PEP = plastid encoded plastid] PHK-полимеразой, для усиления экспрессии представляющего интерес чужеродного гена. Подобно описанным выше конструкциям эти конструкции также содержат ориентирующий сегмент и сайт клонирования для экспрессии представляющего интерес чужеродного гена.
Промоторы, распознаваемые кодируемой пластидной ДНК пластидной РНК-полимеразой, ранее были хорошо охарактеризованы в фотосинтезирующих тканях, таких, как лист. В отличие от этого система транскрипции по настоящему изобретению, включающая кодируемую ядерную полимеразу, позволяет осуществлять экспрессию пластидных генов также в корнях, семенах и в меристематической ткани. У большинства растений, включая кукурузу, хлопчатник и пшеницу, регенерация растения осуществляется путем соматического эмбриогенеза (т.е. включая меристематическую ткань). В предпочтительном варианте осуществления изобретения эффективная трансформация пластид этих культурных растений может быть в значительной степени облегчена благодаря применению NEP-системы транскрипции в пластиде, промоторов и полимераз по настоящему изобретению.
NEP-промоторы по изобретению встраивают в доступные в настоящее время векторы для трансформации пластид и в протоколы по их применению, такие, как представленные в патенте США 5451513 и в находящейся на рассмотрении заявке на патент США 08/189256, а также описанные у Svab и Maliga, Proc. Natl. Acad. Sci. USA, 90, 913 (1993), при этом все эти публикации в полном объеме включены в настоящее описание в качестве ссылки. Для получения трансгенных растений пластиды из нефотосинтезирующих тканей трансформируют селектируемыми генами-маркерами, экспрессируемыми с помощью NEP-промоторов, и транскрибируют полимеразой, кодируемой ядерной ДНК. Для экспрессии представляющих интерес протеинов также конструируют кассеты экспрессии, обеспечивающие высокий уровень экспрессии в нефотосинтезирующей ткани с использованием NEP-промотора, получаемого в результате транскрипции полимеразой, кодируемой ядерной ДНК. В другом варианте РЕР-промоторы по изобретению включают в доступные в настоящее время векторы для трансформации пластид и в протоколы по их применению.
Согласно еще одному варианту осуществления изобретения NEP-система транскрипции также может быть объединена с системой σ70-типа путем использования двойственных NEP/PEP-промоторов.
Краткое описание чертежей
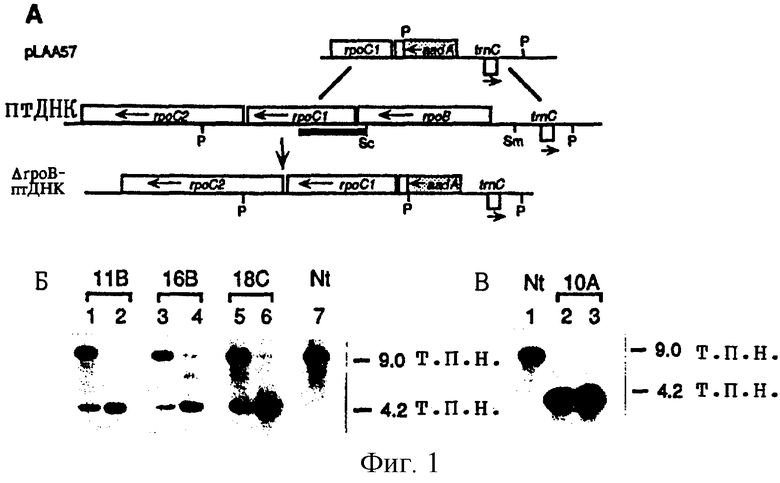
Фиг. 1: Деления гена rроВ из пластидного генома табака путем направленного замещения гена. (А) Гомологичная рекомбинация (диагональные линии) с использованием последовательностей пластидной ДНК, расположенных рядом с геном aadA в плазмиде pLAA57, приводит к замещению rроВ (SacI-SmaI-фрагмент) в пластидном геноме дикого типа (птДНК) на последовательности aadA, что позволяет получить пластидный геном ΔrроВ (ΔrроВ-птДНК). Аббревиатуры: rроВ, rpoC1, rроС2 представляют собой пластидные гены, кодирующие β,β′ и β″ -субъединицы РНК-полимеразы, подобной РНК-полимеразе Е. coil; aadA представляет собой химерный устойчивый к спектиномицину ген. Сайты, распознаваемые рестрикционным ферментом: Р, PstI; Sm, SmaI; Sc, SacI. (Б) Дефицит пигмента связан с делецией гена rроВ. Общую клеточную ДНК выделяли из зеленой (полосы 1, 3, 5) и белой (полосы 2, 4, 6) листовой ткани трех независимо друг от друга трансформированных линий растений табака (линия Nt-pLAA57-11B, полосы 1 и 2; линия Nt-pLAA57-16В, полосы 3 и 4; линия Nt-pLAA57-18C, полосы 5 и 6) и из зеленой листовой ткани растений дикого типа (Nt, полоса 7). ДНК расщепляли с помощью PstI и путем гель-блоттинга гибридизовали с фрагментом ДНК [положения нуклеотидов 22883-24486 птДНК, пронумерованные согласно К. Shinozaki и др. (ЕМВО J., 5, 2043, 1986)], содержащим часть rpoC1 (жирная черная линия на фиг. 1А). Зонд гибридизуется с фрагментом длиной 9,0 т.п.н. из генома дикого типа и с фрагментом длиной 4,2 т. п. н. из ΔrроВ-птДНК. (В) Анализ ДНК методом гель-блоттинга подтверждает отсутствие копий птДНК дикого типа в побегах белого цвета линии Nt-pLAA57-10A (полоса 2) и в потомстве белого цвета, полученном из семян трансплантированного химерного растения той же линии (полоса 3). ДНК из зеленой листовой ткани дикого типа вносили в полосу 1. Заметно отсутствие фрагмента птДНК длиной 9,0 т.п.н. в растениях линии ΔrроВ. Блоттинг проводили аналогично тому, как это описано для фиг. 1Б.
Фиг. 2: Деления гена rроВ приводит к получению фенотипа с дефицитом пигмента. (А) Показаны зеленые растения дикого типа (слева), растения с дефицитом ΔrроВ (справа) и химерные растения (в центре). (Б) Цветущее химерное растение в теплице. Видны белые края листьев, что свидетельствует о присутствии ΔrроВ-пластид во втором листовом слое, который формирует зародышевую линию клеток.
Фиг. 3: (А) Пластиды (Р) в мезофильных клетках листа растений линии ΔrроВ лишены организованных соответствующим образом фотосинтезирующих мембран. Аббревиатуры: N - ядро; V - везикулы; М - митохондрия. (Б) Для сравнения приведена электронная микрофотография хлоропласта (Ср) листа дикого типа с тилакоидными мембранами (Т). Увеличение на фиг. (А) и на фиг. (Б) составляет 7800 раз (7800Х).
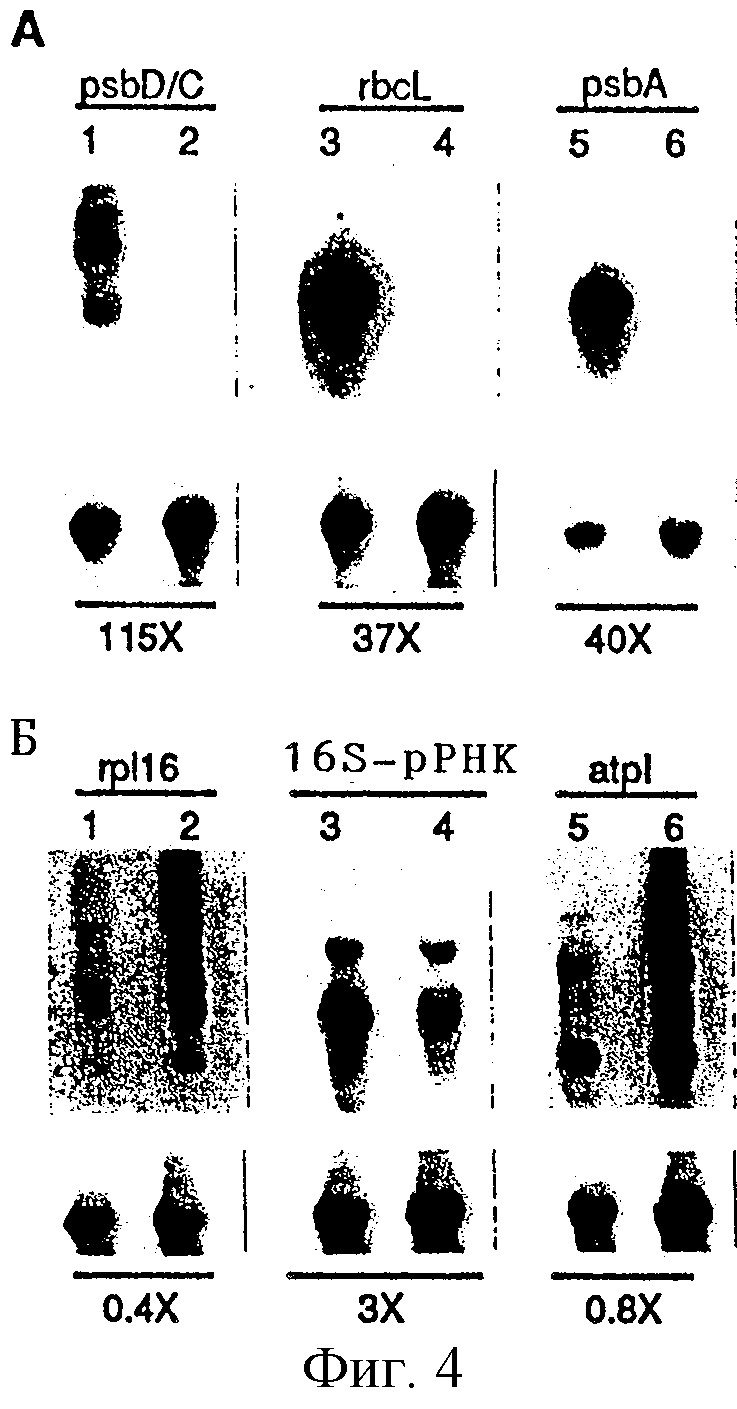
Фиг. 4. (Вверху) Накопление пластидных мРНК (А) фотосинтезирующих генов и (Б) генов генетической системы в растениях линии ΔrроВ. Для гель-блоттинга использовали общую клеточную РНК (А - 3 мкг/полоса; Б - 5 мкг/полоса) из листовой ткани растений дикого типа (полосы 1, 3, 5) и ΔrроВ-типа (полосы 2, 4, 6) и гибридизовали с указанными последовательностями пластидных генов. (Внизу) Указанные выше блотты подвергали повторному зондированию с последовательностями 25S-pДHK. Сигналы гибридизации количественно оценивали с помощью прибора Molecular Dynamics PhosphorImager и стандартизировали по отношению к сигналу 25S-pPHK. Относительное превышение интенсивности сигналов дикого типа над ΔrроВ-типом приведено для каждого зонда под полосами.
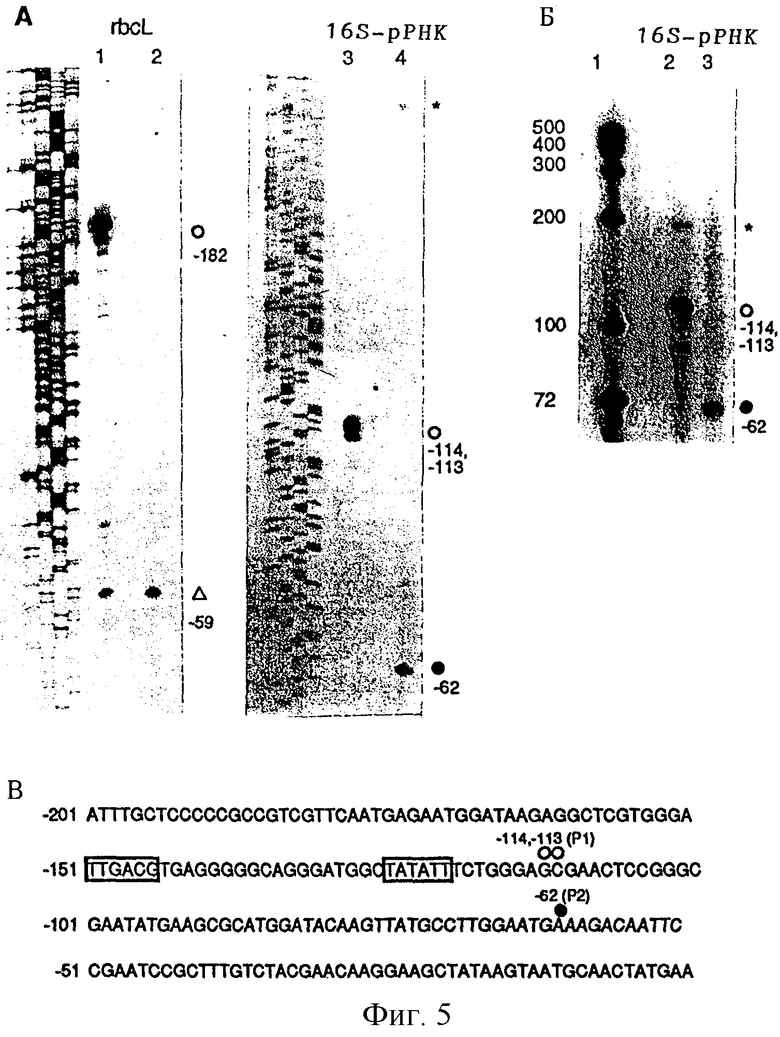
Фиг. 5: Транскрипция в растениях линии ΔrроВ инициируется неканоническим промотором. (А) Для картирования 5'-концов транскриптов rbcL и 16S-рДНК в растениях дикого типа (полосы 1, 3) и ΔrроВ-типа (полосы 2, 4) применяли анализ удлинения праймера. Первичные транскрипты обозначены кружками (не закрашенными для дикого типа, закрашенными для ΔrроВ), а образовавшиеся при процессинге транскрипты обозначены треугольниками. Транскрипты неизвестного происхождения обозначены звездочками. Соответствующие ступени последовательности (наличие загрузки GATC) получали с использованием таких же праймеров, которые применяли в реакциях удлинения праймера. Номера, указанные сбоку от каждого продукта удлинения, обозначают расстояние от первого нуклеотида кодирующей последовательности гена rbcL и от первого нуклеотида зрелой 16S-pPHK. (Б) Картирование первичных транскриптов 16S-pPHK в растениях дикого типа (полоса 2) и ΔrроВ-типа (полоса 3). Общую РНК листа (20 мкг) кэппировали in vitro, и кэппированные виды 16S-pPHK выявляли методом защиты от РНКазы после гибридизации с комплементарным РНК-зондом. Кэппированные защищенные продукты имеют такое же обозначение, что и на фиг. 5(А). Полоса 1 содержит стандарты РНК известного размера. (В) Последовательность ДНК 16S-рДНК, расположенная против хода транскрипции, с транскриптами, начинающимися от промоторов кодируемой пластидной ДНК (σ70-тип = Р1) и кодируемой ядерной ДНК (Р2) полимераз (обозначения Р1 и Р2 соответствуют таковым, принятым у A. Vera и М. Sigiura, Curr. Genet. 27, 280, 1995). Консенсусные элементы σ70-промотора (-35 и -10) обозначены прямоугольниками. Сайты инициации обозначены кружками, как на фиг. 5 (А) и 5 (Б). Нумерация начинается с первого нуклеотида против хода транскрипции области, кодирующей 16S-рДНК (-1 соответствует нуклеотиду 102757 в пластидном геноме табака).
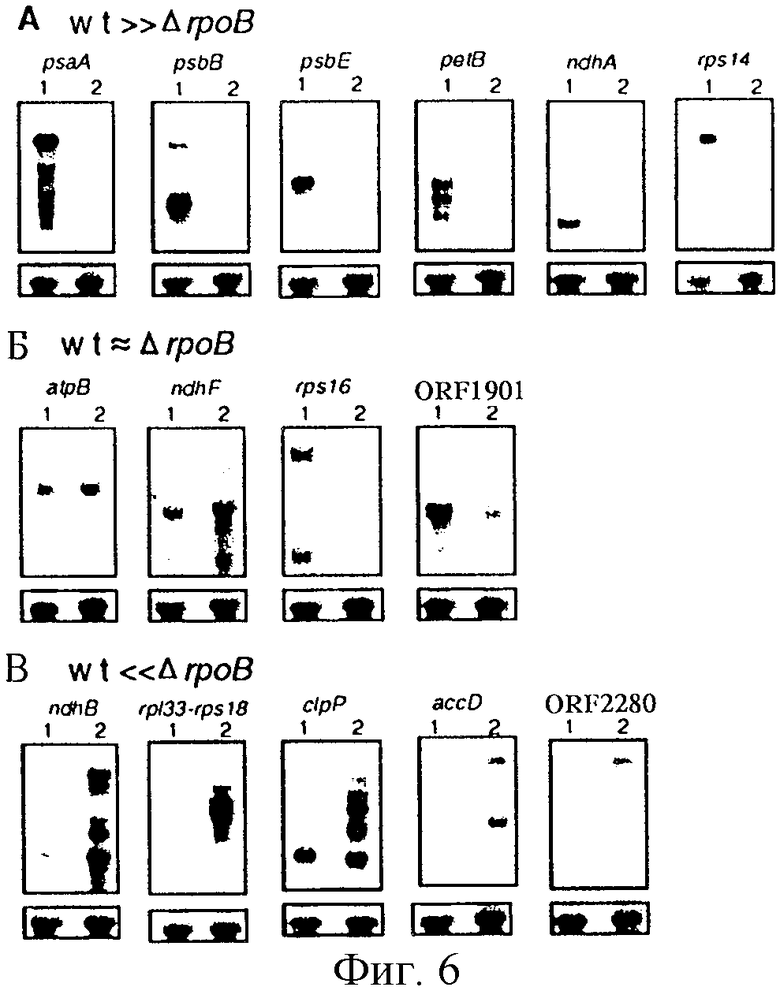
Фиг. 6: Накопление пластидных мРНК в листьях табака дикого типа (дикий тип обозначен через wt) и ΔrроВ-типа. Блотты, несущие пластидные гены (см. пример I), группируют следующим образом. (А) мРНК присутствует в существенно большем количестве в листьях дикого типа по сравнению с ΔrроВ-типом. (Б) Уровни мРНК сопоставимы в листьях дикого типа и ΔrроВ-типа или (В) мРНК больше в листьях ΔrроВ-типа. Гель-блоттинг проводили с использованием общей клеточной РНК (3 мкг/полоса) из листовой ткани дикого типа (полосы 1) и ΔrроВ-типа (полосы 2) и гибридизовали для выявления последовательностей пластидных генов. (Нижняя часть) Для контроля загрузки приведенные выше блотты повторно зондировали последовательностями 25S-рДНК.
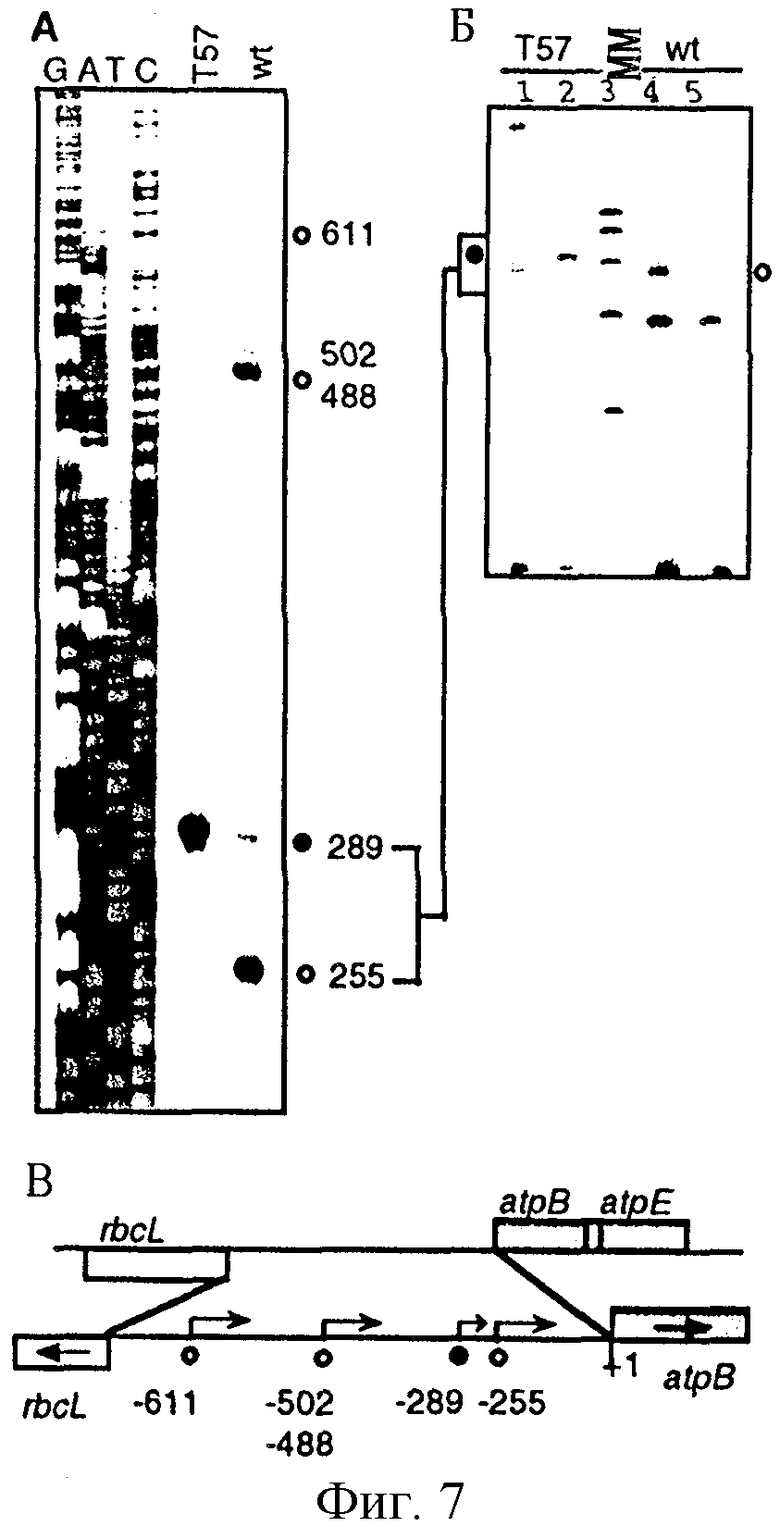
Фиг. 7: Картирование сайтов инициации транскрипции оперона atpB в листьях табака дикого типа и ΔrроВ-типа. (А) Анализ удлинения праймера. Меченные по концам продукты удлинения праймера из образцов дикого типа (wt) и ΔrроВ-типа (Т57) подвергали анализу параллельно с гомологичными последовательностями, полученными при использовании этого же праймера. Номера, проставленные сбоку от последовательности, обозначают расстояние от кодона инициации трансляции ATG. Первичные транскрипты, полученные от NEP- и РЕР-промоторов, обозначены закрашенными и незакрашенными кружками соответственно. (Б) Применение кэппирования in vitro и метода защиты от РНКазы для идентификации 5'-концов первичных транскриптов. В полосы вносили образцы РНК ΔrроВ-типа (Т57 - 1, 2) и дикого типа (wt - 4, 5) с (2, 4) и без (1, 5) защищающей комплементарной антисмысловой РНК. В полосу 3 вносили маркеры для определения молекулярной массы (ММ) (100, 200, 300, 400 и 500 нуклеотидов). 5'-конец транскрипта на фиг. 7(А) соответствует указанному в скобках размеру защищенного фрагмента: -254 (277 нуклеотидов, н.т.), -289 (311). Следует отметить артефакт, имеющий место несколько ниже маркера в 200 н.т., который присутствует в образцах незащищенной РНК. (В) Физическая карта межгенной области atpB-rbcL. Положение на карте 5'-концов первичных транскриптов для NEP- и РЕР-промоторов atpB обозначено так же, как и на фиг. 7 (А).
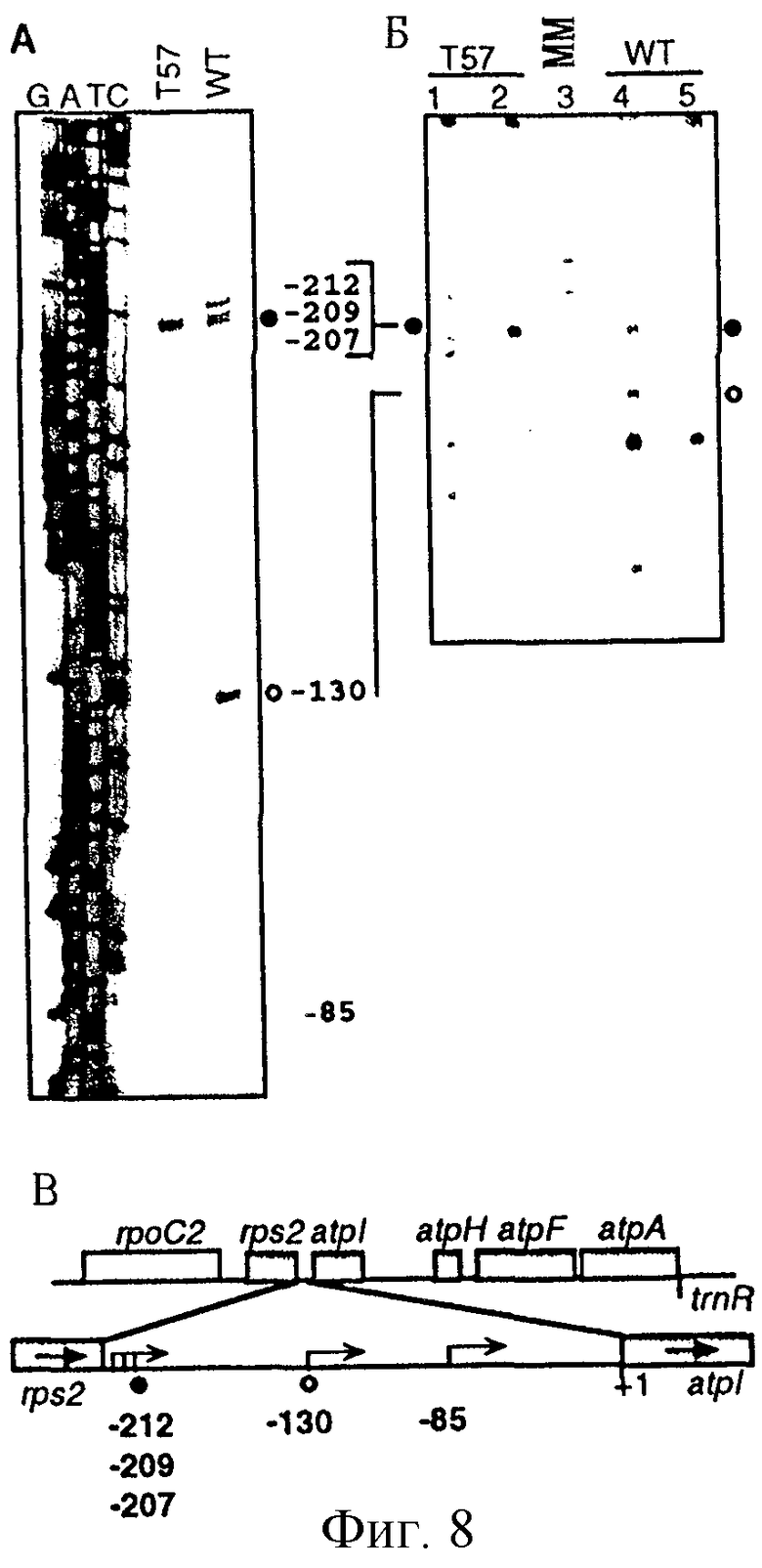
Фиг. 8: Картирование сайтов инициации транскрипции оперона atpI в листьях табака дикого типа и ΔrроВ-типа. (А) Анализ удлинения праймера. Меченные по концам продукты удлинения праймера из образцов дикого типа (wt) и ΔrроВ-типа (Т57) подвергали анализу параллельно с гомологичными последовательностями, полученными при использовании этого же праймера. Номера, проставленные сбоку от последовательности, обозначают расстояние от кодона инициации трансляции ATG. Первичные транскрипты, полученные с помощью NEP- и РЕР-промоторов, обозначены закрашенными и незакрашенными кружками соответственно. (Б) Применение кэппирования in vitro и метода защиты от РНКазы для идентификации 5'-концов первичных транскриптов. В полосы вносили образцы РНК ΔrроВ-типа (Т57 - 1, 2) и дикого типа (wt - 4, 5) с (2, 4) и без (1, 5) защищающей комплементарной антисмысловой РНК. В полосу 3 вносили маркеры для определения молекулярной массы (ММ) (100, 200, 300, 400 и 500 нуклеотидов). 5'-конец транскрипта на фиг. 8 (А) соответствует указанному в скобках размеру защищенного фрагмента: -130 (235 н.т.), -207, 209, 212 (303, 305, 309; не выявлены). Следует отметить артефакт, имеющий место несколько ниже маркера в 200 н. т., который присутствует в образцах незащищенной РНК. (В) Физическая карта межгенной области rps2-atpI. Положение на карте 5'-концов первичных транскриптов для NEP- и РЕР-промоторов atpI оперона обозначено так же, как и на фиг. 8(А).
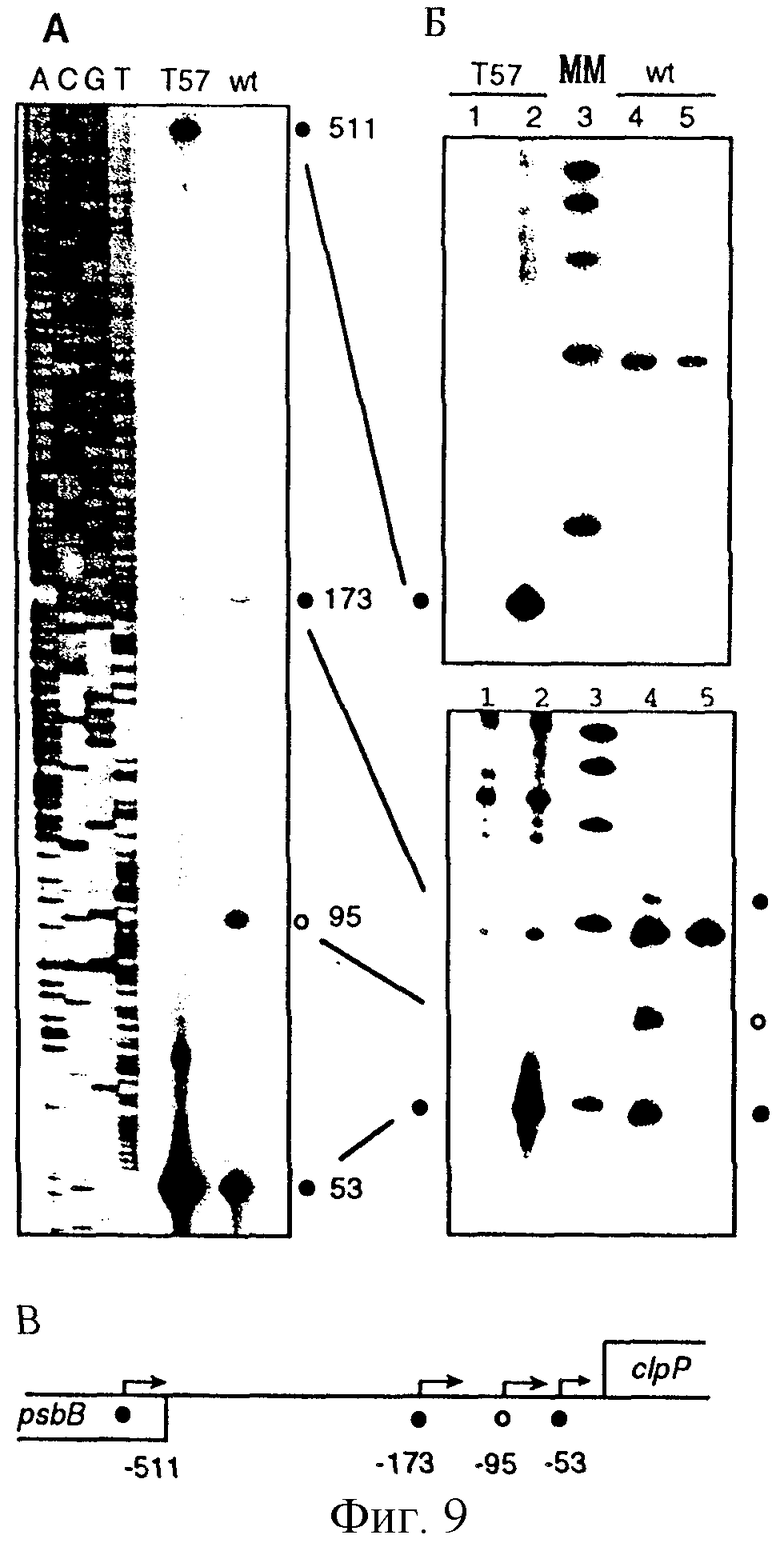
Фиг. 9: Картирование сайтов инициации транскрипции с1рР в листьях табака дикого типа и ΔrроВ-типа. (А) Анализ удлинения праймера. Меченные по концам продукты удлинения праймера из образцов дикого типа (wt) и ΔrроВ-типа (Т57) подвергали анализу параллельно с гомологичными последовательностями, полученными при использовании этого же праймера. Номера, проставленные сбоку от последовательности, обозначают расстояние от кодона инициации трансляции ATG. Первичные транскрипты, полученные с помощью NEP- и РЕР-промоторов, обозначены закрашенными и незакрашенными кружками, соответственно. (Б) Применение кэппирования in vitro и метода защиты от РНКазы для идентификации 5'-концов первичных транскриптов. В полосы вносили образцы РНК ΔrроВ-типа (Т57 - 1, 2) и дикого типа (wt - 4, 5) с (2, 4) и без (1, 5) защищающей комплементарной антисмысловой РНК. В полосу 3 вносили маркеры для определения молекулярной массы (ММ) (100, 200, 300, 400 и 500 нуклеотидов). 5'-конец транскрипта на фиг. 9 (А) соответствует указанному в скобках размеру защищенного фрагмента: -53 (96 н.т.), -95 (138 н.т.), -173 (216 н.т.) и -511 (69 н.т.). Следует отметить артефакт, имеющий место несколько ниже маркера в 200 н. т., который присутствует в образцах незащищенной РНК. (В) Физическая карта межгенной области clpP-psbB. Положение на карте 5'-концов первичных транскриптов для NEP- и РЕР-промоторов с1рР обозначено так же, как и на фиг. 9 (А).
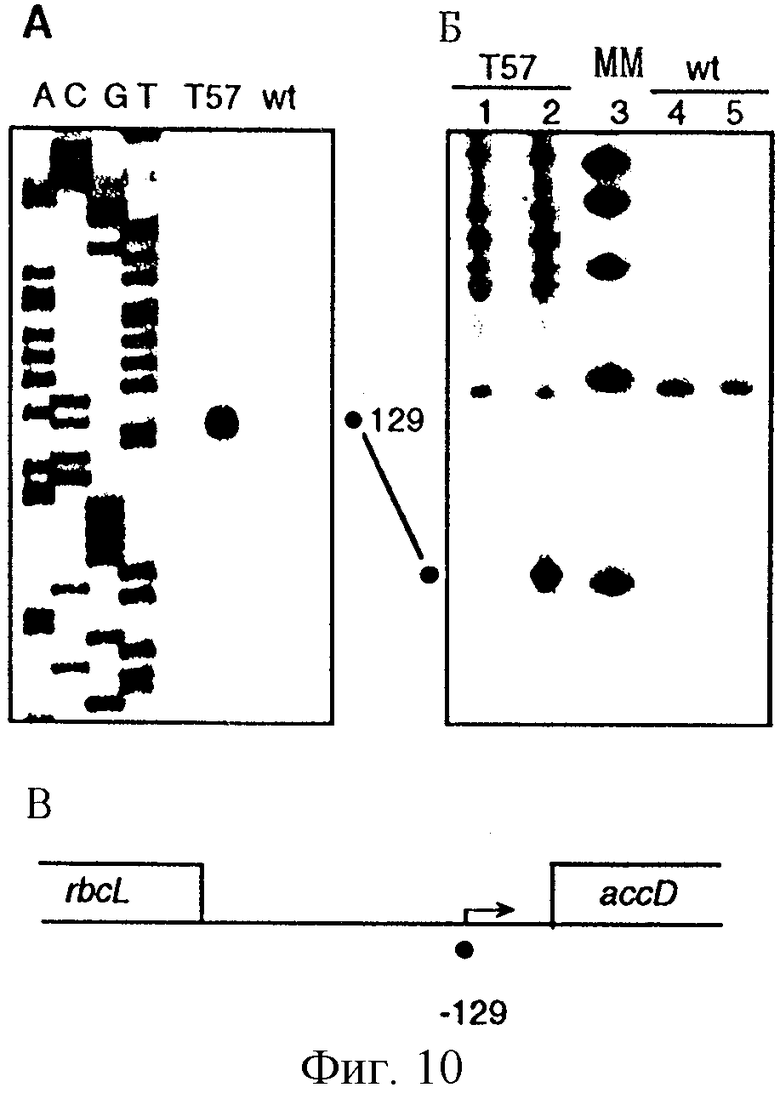
Фиг. 10. Картирование сайтов инициации транскрипции accD в листьях табака дикого типа и ΔrроВ-типа. (А) Анализ удлинения праймера. Меченные по концам продукты удлинения праймера из образцов дикого типа (wt) и ΔrроВ-типа (Т57) подвергали анализу параллельно с гомологичными последовательностями, полученными при использовании этого же праймера. Номера, проставленные сбоку от последовательности, обозначают расстояние от кодона инициации трансляции ATG. Первичный транскрипт, полученный от NEP-промотора PaccD-129, обозначен закрашенным кружком. (Б) Применение кэппирования in vitro и метода защиты от РНКазы для идентификации 5'-концов первичных транскриптов. В полосы вносили образцы РНК ΔrроВ-типа (Т57 - 1, 2) и дикого типа (wt - 4, 5) с (2, 4) и без (1, 5) защищающей комплементарной антисмысловой РНК. В полосу 3 вносили маркеры для определения молекулярной массы (ММ) (100, 200, 300, 400 и 500 нуклеотидов). 5'-конец транскрипта -57 на фиг. 10 (А) соответствует защищенному фрагменту размером 103 н.т. Следует отметить артефакт, имеющий место несколько ниже маркера в 200 н.т., который присутствует в образцах незащищенной РНК. (В) Физическая карта межгенной области accD--rbcL. Обозначено положение на карте 5'-конца первичного транскрипта для NEP-промотора PaccD-129.
Фиг. 11: Сранительный анализ первичной структуры последовательностей ДНК, соседних с сайтами инициации транскрипции NEP-промотора. Прямоугольниками обозначены нуклеотиды, имеющие более 6 совпадений. Консенсусная последовательность, соседняя с сайтом инициации транскрипции, приведена ниже. Положение 5'-концов обозначено закрашенными кружками. Следует отметить, что 5'-концы Prpsl2-153 и Prpsl6-107 не кэппировались и могут не относиться к первичным транскриптам.
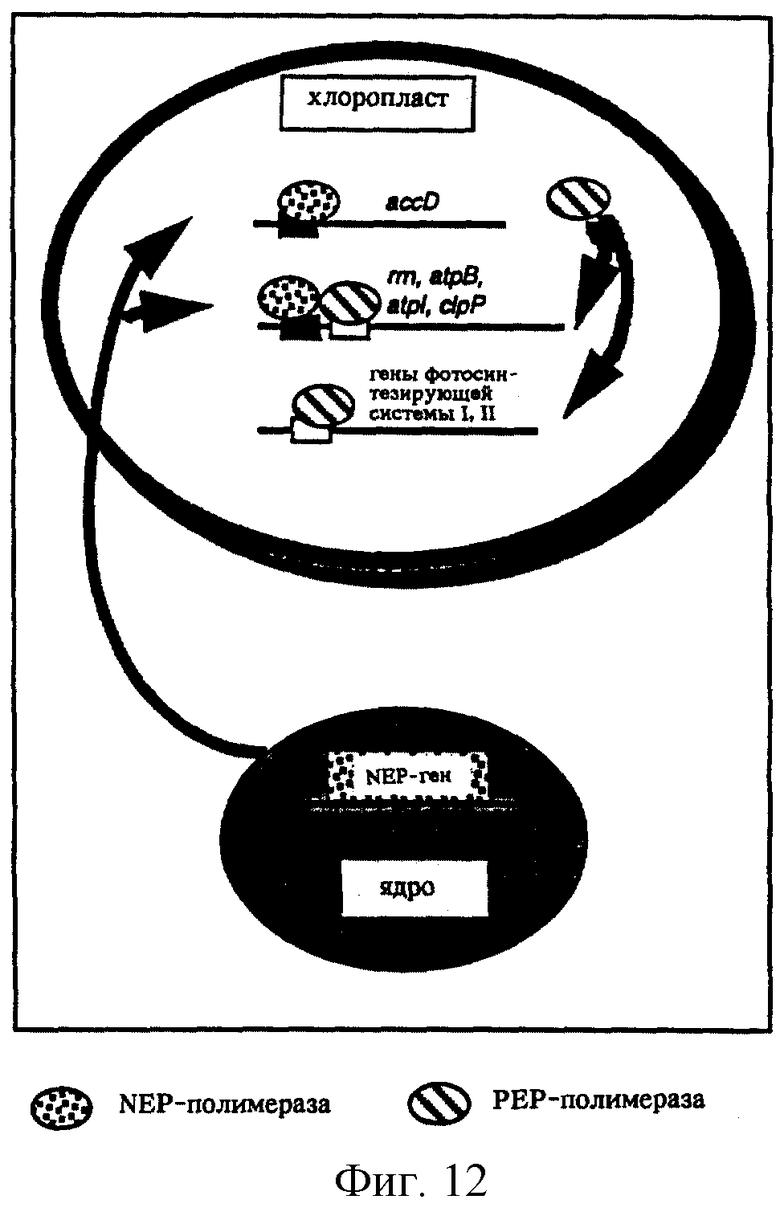
Фиг. 12: NEP- и РЕР-полимеразы благодаря способности распознавать различные промоторы обеспечивают механизм избирательной транскрипции пластидных генов. Следует отметить, что некоторые гены имеют только РЕР-промоторы (фотосинтезирующая система I и фотосинтезирующая система II), а другие имеют как PEP-, так и NEP-промоторы (большинство генов "домашнего хозяйства", или только NEP-промоторы (accD)).
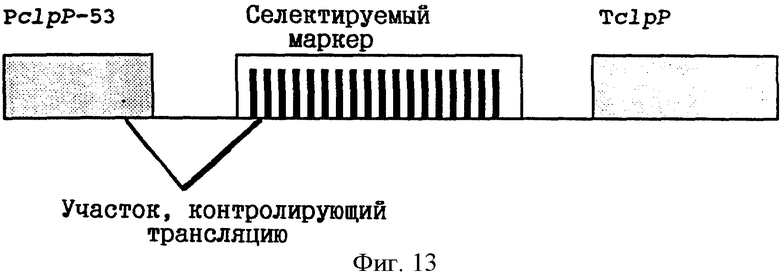
Фиг. 13: Схематическая диаграмма химерного пластидного гена, экспрессируемого с помощью NEP-промотора.
Описание изобретения
В некоторых исследованиях было высказано предположение о существовании дополнительной локализованной в пластиде РНК-полимеразы, кодируемой ядерной ДНК (эти данные обобщены у Gruissem и Tonkyn, 1993; Igloi и Kossel, 1992; Mullet, 1993; Link, 1994).
Путем удаления гена rроВ, кодирующего основную β-субъединицу РНК-полимеразы табака, подобной РНК-полимеразе Е. coli, доказано существование в пластиде второй системы транскрипции, которая кодируется ядерной ДНК (Allison и др. , 1996, EMBO J. 15: 2802-2809). Деления гена rроВ приводила к получению растений, дефектных с точки зрения фотосинтеза с дефицитом пигмента. Изучение ультраструктуры пластид в клетках мезофилла листа растений с делецией rроВ (ΔrроВ) позволило выявить пропластидоподобные органеллы, у которых отсутствовало упорядочнное строение тилакоидных мембран, характерное для активных в отношении фотосинтеза хлоропластов. Уровни транскриптов фотосинтезирующих генов rbcL, psbA и psbD были очень невысокими, тогда как мРНК генов rрl16, atpI и 16S-рДНК накапливались до уровней, близких к таковым в диком типе или до более высоких уровней, чем у дикого типа. Отсутствие накопления транскриптов для фотосинтезирующих генов являлось следствием отсутствия промоторной активности σ70-типа. В то время как в листьях табака дикого типа оперон рибосомной РНК в норме транскрибируется с помощью промотора σ70-типа, в растениях ΔrроВ-типа оперон рРНК транскрибировался с помощью промотора, не относящегося к σ70-типу. Оперон рРНК является первой единицей транскрипции, для которой выявлена и кодируемая пластидной ДНК, и кодируемая ядерной ДНК пластидная РНК-полимераза (PEP- и NEP-полимераза соответственно).
Анализ промоторных областей других генов показал, что оперон рРНК не явлется единственным. Он является представителем большого класса пластидных генов, каждый из которых имеет по крайней мере один промотор для PEP- и NEP-полимеразы, и обладает способностью экспрессировать любую из двух пластидных РНК-полимераз. Кроме того, были выявлены пластидные гены, которые транскрибируются исключительно при участии NEP-РНК-полимеразы. Таким образом, эти результаты позволяют предположить, что дополнительные генспецифические механизмы регулируют уровни NEP-транскриптов в пластидах различных типов.
Сайт инициации транскрипции NEP был выявлен на расстоянии приблизительно 62 основания против хода транскрипции от 5'-конца зрелой 16S-pPHK. Последовательность, окружающая сайт инициации, является высококонсервативной у многочисленных изученных видов растений и не имеет сходства с консенсусной последовательностью РЕР-промотора. Консенсусные последовательности NEP-промотора, важные для распознавания и связывания полимеразы, кодируемой ядерной ДНК (аналогично последовательностям -10 и -35 сайта инициации транскрипции E. coli-типа), предпочтительно локализованы внутри области длиной приблизительно 50 нуклеотидов в любом направлении от сайта инициации транскрипции NEP. Как описано более подробно в примере I, существуют несколько различных NEP-промоторов, и иногда обнаруживали NEP-промоторы, конъюгированные с РЕР-промоторами.
Полимеразы по изобретению могут быть очищены с помощью хроматографии с использованием стандартных методов. NEP-полимеразная активность в колоночных фракциях может быть проанализирована в реакциях транскрипции in vitro с использованием сегментов ДНК, содержащих область NEP-промотора в качестве матриц. В альтернативном варианте последовательности NEP-промотора могут быть присоединены к матриксу, который может быть разделен с помощью ряда способов (например, с помощью магнитных гранул). Связанную с матриксом ДНК инкубируют с растительным экстрактом в условиях, при которых ожидается, что полимераза, кодируемая ядерной ДНК, будет связываться с ДНК. Комплекс матрикс/ДНК/полимераза затем отделяют от растительного экстракта и связанный протеин далее может быть выделен и охарактеризован. Очищенный любым из вышеуказанных способов протеин может использоваться для получения антител для зондирования библиотек экспрессии, для цели выделения ядерных генов или кДНК, кодирующих полимеразу, кодируемую ядерной ДНК.
В качестве альтернативного подхода к выделению NEP-полимеразы может быть использовано выделение протеинов, обладающих специфической аффинностью по отношению к фрагменту промотора, а N-концевая аминокислотная последовательность может быть определена путем микросеквенирования. Затем аминокислотная последовательность может быть использована для конструирования соответствующих ПЦР-праймеров, предназначенных для выделения генов.
Активность ранее известной кодируемой пластидной ДНК системы транскрипции σ70-типа была в достаточной степени охарактеризована в тканях с высокой фотосинтезирующей активностью, таких, как лист. В отличие от этого предлагаемая система транскрипции с использованием полимеразы, кодируемой ядерной ДНК, управляет экспрессией пластидных генов также и в корнях, семенах и в меристематической ткани. У большинства растений, включая кукурузу, хлопчатник и пшеницу, регенерация растения осуществляется путем соматического эмбриогенеза (т.е. включая меристематическую ткань). Эффективная пластидная трансформация этих культурных растений может быть осуществлена или в значительной степени облегчена благодаря применению предлагаемой NEP-системы транскрипции в пластиде.
NEP-промоторы по изобретению могут быть включены в доступные в настоящее время векторы для трансформации пластиды согласно протоколам по их применению, таким, как описанные в патенте США 5451513 и в находящейся на рассмотрении заявке на патент США 08/189256, а также приведенные у Svab и Maliga, Proc. Natl. Acad. Sci. USA, 90, 913 (1993), при этом все эти публикации в полном объеме включены в настоящее описание в качестве ссылки. Для получения трансгенных растений пластиды из нефотосинтезирующих тканей трансформируют селектируемыми генами-маркерами, экспрессируемыми с помощью NEP-промоторов, и транскрибируют с помощью полимеразы, кодируемой ядерной ДНК. Аналогично этому для экспрессии представляющих интерес протеинов конструируют кассеты экспрессии, обеспечивающие высокий уровнь экспрессии в нефотосинтезирующей ткани с использованием NEP-промотора, транскрибируемого полимеразой, кодируемой ядерной ДНК. NEP-система транскрипции также может быть объединена с системой σ70-типа с использованием двойных NEP/PEP-промоторов. В некоторых случаях также может требоваться экспрессия трансгенов с помощью NEP-промоторов в фотосинтезирующей ткани.
В приведенном ниже в примерах I-III подробном описании изложены предпочтительные способы создания и применения конструкций ДНК по настоящему изобретению и практического воплощения способов по изобретению. Любые конкретно не описанные методики молекулярного клонирования и рекомбинантных ДНК осуществляют в соответствии со стандартными способами, как это в общих чертах изложено, например, у Ausubel (ред.), Current Protocols in Molecular Biology. John Wiley и Sons, Inc. (1994).
Ниже изобретение более подробно проиллюстрировано на примерах, не ограничивающих его объем.
Пример I
Демонстрация существования в пластидах второй, отличающейся от известной, системы транскрипции путем делеции гена rроВ
Для доказательства существования в пластидах РНК-полимеразы, не подобной таковой Е. coli, ген одной из основных субъединиц подобного Е. coli фермента удаляли путем делеции из пластидного генома табака. Затем определяли уровни мРНК в мутантных пластидах. Данные свидетельствуют о том, что в отсутствие кодируемого пластидной ДНК подобного Е. coli фермента экспрессия некоторых фотогенов резко снижается. В противоположность этому уровни транскриптов пластидных генов, кодирующих аппарат экспрессии генов, близки к уровням в растениях дикого типа. Следовательно, РНК-полимераза, не подобная таковой Е. coli, избирательно транскрибирует определенную подгруппу пластидных генов. Этот второй аппарат транскрипции не инициируется обычными для Е. coli σ70-промоторами, но обладает способностью распознавать новую промоторную последовательность.
Материалы и методы примера I
Конструирование плазмиды. Плазмида pLAA57 является производной pBSKS+ (фирма Stratagene), которая несет SacI-BamHI-фрагмент (нуклеотиды с 22658 по 29820) птДНК. Внутренний фрагмент SacI-SmaI ДНК внутри вставки между нуклеотидами 24456 и 28192 птДНК замещали химерным устойчивым к спектиномицину (aadA) геном. Ген aadA идентичен таковому, описанному у Z. Svab и Р. Maliga, Proc. Natl. Acad. Sci. USA, 90, 913 (1993), за исключением того, что 3'-область psbA короче и находится в фрагменте XbaI-DraI, как описано у J.M. Staub и Р. Maliga, Plant J. 6, 547, (1994).
Трансформация растения. Для трансформации пластиды частицы вольфрама покрывали (Z. Svab и Р. Maliga, Proc. Natl. Acad. Sci. USA, 90, 913, 1993) ДНК pLAA57 и интродуцировали в листья растений табака Nicotiana tabacum, используя инжектор для биобаллистического метода типа DuPount PDS10000He при давлении 1100 фунтов/кв. дюйм. Трансгенные побеги отбирали в асептических условиях на среде RMOP (Z. Svab, P. Hajdukiewicz, P. Maliga, Proc. Natl. Acad. Sci. USA, 87, 8526, 1990), содержащей 500 мг/мл дигидрохлорида спектиномицина. Трансгенные черенки укореняли и выращивали на RM-среде, состоящей из затвердевших в агаре MS-солей (Т. Murashige и F. Skoog, Physiol. Plant., 15, 493, 1962), содержащей 3% сахарозы.
Электронная микроскопия. С помощью электронной микроскопии исследовали полностью распустившиеся листья черенков дикого типа и линии ΔrроВ, выращенные в стерильной культуре на RM-среде с 3% сахарозы. Ткань фиксировали в течение 2 ч в 2%-ном глутаровом альдегиде, 0,2М сахарозе и 0,1М фосфатном буфере (рН 6,8) при комнатной температуре и трижды промывали в 0,2М сахарозе, 0,1М фосфатном буфере. Зафиксированные ткани подвергали постфиксации в буфере из 1%-ного тетроксида осмия с 0,2М сахарозой, дегидрировали в постепенно меняющихся концентрациях этанола, заливали в эпоксидную смолу Спурра (твердая), делали срезы и окрашивали уранилацетатом и цитратом свинца для трансмиссионной электронной микроскопии.
Гель-блоттинг. Общую ДНК листа получали по методике, описанной у I.J. Mettler, Plant Mol. Biol. Rep., 5, 346 (1987), обрабатывали рестрикционной эндонуклеазой PstI, разделяли на 0,7%-ных агарозных гелях и переносили на Hybond N (фирма Amersham), используя устройство для переноса типа Posiblot Transfer (фирма Stratagene). Гибридизацию со случайным образом примированным меченым фрагментом проводили в буфере для быстрой гибридизации (Rapid Hybridization Buffer, фирма Amersham) в течение ночи при 65oС. Общую РНК листа получали с использованием TRIzol (GIBCO BRL), следуя протоколу производителя. РНК подвергали электрофорезу на гелях 1% агарозы/формальдегид, затем переносили на найлоновую мембрану и зондировали аналогично тому, как это описано для блоттинга ДНК.
Синтез зондов. Двухцепочечные ДНК-зонды для psbA, atpI и rрl16 получали с помощью случайным образом примированных, меченных с помощью 32Р фрагментов ДНК, полученных с использованием ПЦР. Последовательность применяемых для ПЦР праймеров, а также их положения в птДНК табака (К. Shinozaki и др., EMBO J., 5, 2043, 1986) следующие: psbA, 5'-праймер = 5'-CGCTTCTGTAACTGG-3' (комплементарный нуклеотидам 1550-1536 птДНК), 3'-праймер = 5'-TGACTGTCAACTACAG-3' (нуклеотиды 667-682); atpI, 5'-праймер = 5'-GTTCCATCAATACTC-3' (комплементарный нуклеотидам 15985-15971), 3'-праймер = 5'-GССGСGGСТАААGТТ-3'(нуклеотиды 15292-15306); rрl16, 5'-праймер = 5'-TCCCACGTTCAAGGT-3'(комплементарный нуклеотидам 84244-84230), 3'-праймеp = 5'-TGAGTTCGTATAGGC-3'(нуклеотиды 83685-83699). Для создания зондов к rbcL, psbD/C и 16S-pPHK метили с помощью 32P следующие расщепленные рестриктазами фрагменты ДНК: rbcL, BamHI-фрагмент (нуклеотиды 58047-59285 в птДНК); psbD/C, SacII-HindIII-фрагмент оперона табака psbD/C (нуклеотиды 34691-36393); 16S-pPHK, EcoRI-EcoRV-фрагмент (нуклеотиды 138447-140855 в птДНК).
Зонд для 25S-рРНК табака получали из плазмиды pKDRl (D. Dempsey, K.W. Wobbe, D. F. Klessig, Mol. Plant Path., 83, 1021, 1993), содержащей EcoRI-фрагмент длиной 3,75 т. п. н. из локуса 25S/18S табака, клонированного в плазмиде pBR325. При гибридизации с помощью гель-блоттингов для 25S-pPHK меченный с помощью 32Р двухцепочечный ДНК-зонд смешивали с немеченой плазмидой pKDRl в количестве, соответствующем 2-кратному избытку относительно количества РНК, присутствующего на фильтре.
Стандартизация уровней ДНК по количеству копий пластидного генома. С целью определить, влияют ли изменения в количестве копий пластидного генома на выявленные различия в экспрессии генов, получали общую клеточную ДНК и РНК из равных количеств листовой ткани растений дикого типа и ΔrроВ-типа. Для сравнения количества копий пластидного генома на эквивалент листовой массы гель-блоттинги ДНК проводили с равным объемом каждого препарата ДНК и зондировали радиоактивномеченным EcoRI-EcoRV-фрагментом (нуклеотиды 138447-140845 птДНК) (К. Shinozaki и др. , EMBO J., 5, 2043, 1986) последовательности 16S-рДНК. Количественная оценка с помощью PhosporImage-анализа показала одинаковое количество копий пластидного генома в каждом образце. Количество 16S-pPHK, полученное из одинаковых образцов ткани и измеренное с помощью гель-блоттингов РНК при использовании равных объемов каждого препарата РНК, оказалось сниженным в 2,5 раза в растениях линии ΔrроВ. Это значение соответствует 3-кратному снижению, выявленному при стандартизации с сигналом цитоплазматической 25S-pPHK (фиг. 3Б).
Реакции удлинения праймеров. Реакции удлинения праймеров проводили с использованием 3 мкг (дикий тип) или 10 мкг (ΔrроВ-типа) общей РНК листа, как описано у L.A. Allison и Р. Maliga, EMBO J., в печати), применяя следующие праймеры: для 16S-pPHK: 5'-TTCATAGTTGCATTACTTATAGCTTC-3' (комплементарный нуклеотидам 102757-102732); для rbcL: 5'-ACTTGCTTTAGTCTCTGTTTGTGGTGACAT-3'(комплементарный нуклеотидам 57616-57587). Ступени последовательности получали с этими же праймерами, используя набор секвиназы II (набор Sequenase II, фирма USB).
Идентификация первичных транскриптов с помощью кэппинга in vitro. Общую РНК (20 мкг) листа из растений дикого типа и линии ΔrроВ подвергали кэппингу в присутствии [α-32P]-ГТФ (J.C. Kennell и D.R. Pring, Mol. Gen. Genet., 216, 16, 1989). Меченые 16S-pPHK выявляли путем защиты от рибонуклеазы (A. Vera и М. Sigiura, Plant Mol. Biol, 19, 309, 1992), используя набор RPAII (фирма Ambion). Для получения защищенной комплементарной РНК расположенную против хода транскрипции область 16S-pДHK (нуклеотиды 102526-102761 птДНК) подвергали ПЦР-амплификации с использованием следующих праймеров: 5'-праймер представлял собой праймер 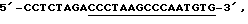 соответствующий нуклеотидам 102526-102541 птДНК (К. Shinozaki и др., EMBO J., 5, 2043, 1986), (подчеркнуты) плюс сайт XbaI; 3'-праймер представлял собой праймер
соответствующий нуклеотидам 102526-102541 птДНК (К. Shinozaki и др., EMBO J., 5, 2043, 1986), (подчеркнуты) плюс сайт XbaI; 3'-праймер представлял собой праймер 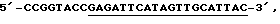 комплементраный нуклеотидам 102761-102742 птДНК (подчеркнуты) плюс сайт KpnI. Амплифицированный продукт клонировали в виде XbaI-KpnI-фрагмента в расщепленном рестриктазами ХbаI и KpnI векторе pBSKS+ (фирма Stratagene). Для создания немеченой РНК, комплементарной 5'-концу 16S-pPHK, полученную плазмиду линеаризовали с помощью XbaI и транскрибировали с использованием реакции типа Megascript (фирма Ambion) с Т3-РНК-полимеразой. Маркеры (100, 200, 300, 400 и 500 нуклеотидов) получали с помощью набора матриц РНК (RNA Centure Markers Template Set, фирма Ambion), следуя протоколу производителя. Маркер нуклеотида 72 представлял собой зрелый транскрипт, полученный в результате процессинга из пластидного гена trnV и созданный с помощью защиты от РНКазы.
комплементраный нуклеотидам 102761-102742 птДНК (подчеркнуты) плюс сайт KpnI. Амплифицированный продукт клонировали в виде XbaI-KpnI-фрагмента в расщепленном рестриктазами ХbаI и KpnI векторе pBSKS+ (фирма Stratagene). Для создания немеченой РНК, комплементарной 5'-концу 16S-pPHK, полученную плазмиду линеаризовали с помощью XbaI и транскрибировали с использованием реакции типа Megascript (фирма Ambion) с Т3-РНК-полимеразой. Маркеры (100, 200, 300, 400 и 500 нуклеотидов) получали с помощью набора матриц РНК (RNA Centure Markers Template Set, фирма Ambion), следуя протоколу производителя. Маркер нуклеотида 72 представлял собой зрелый транскрипт, полученный в результате процессинга из пластидного гена trnV и созданный с помощью защиты от РНКазы.
Результаты
Нарушение активности РНК-полимеразы, подобной РНК-полимеразе Е. coli, в пластидах табака приводит к появлению фенотипа дефицита пигмента. Для предотвращения нарушения других функций пластидных генов в качестве мишени для делеции был выбран ген rроВ, поскольку является первым в рамке считывания оперона, кодирующего исключительно субъединицы Е. coli-подобной пластидной полимеразы (К. Shinozaki и др., EMBO J., 5, 2043, 1986). Делецию осуществляли путем замещения большей части кодирующей области rроВ (3015 из 3212 пар оснований) и 691 пары оснований некодирующей области, расположенной против хода транскрипции, на химерный устойчивый к спектиномицину (aadA) ген (Z. Svab и Р. Maliga, Proc. Natl. Acad. Sci. USA, 90, 913 (1993) в клонируемом фрагменте пластидной ДНК (птДНК). Полученную плазмиду интродуцировали путем бомбардировки частицами в хлоропласты табака, при этом ген aadA интегрировался в геном пластиды с помощью фланкирующих последовательностей пластидной ДНК, как показано в виде диаграммы на фиг. 1А. Поскольку генетическая система пластиды является высоко полиплоидной, характеризующейся тем, что в каждой клетке листа содержится до 10000 идентичных копий птДНК, проводили селективную амплификацию копий трансформированного генома, выращивая обработанную путем бомбардировки ткань на среде, содержащей спектиномицин (Р. Maliga, Trends Biotechnol., 11, 101, 1993).
После первой стадии селекции получали несколько устойчивых к спектиномицину растений, у которых были обнаружены секторы белой листовой ткани (фиг. 2А). Анализ с помощью гель-блоттинга ДНК секторов белого и зеленого цвета показал, что дефицит пигмента коррелировал с делецией rроВ в трех независимо друг от друга трансформированных линиях (фиг. 1Б). Большая часть образов ДНК из ткани с дефицитом пигмента, например, полоса 4 на фиг. 1Б, содержала смесь копий генома дикого типа и трансформированного генома. Полное отсутствие копий птДНК дикого типа имело принципиальное значение для интерпретации данных. Далее для получения растений, содержащих только трансформированные пластидные геномы, побеги регенерировали из секторов ткани белого цвета. Этот способ позволил получить только белые растения (фиг. 2А), которые не содержали птДНК дикого типа, что подтверждено с помощью гель-блоттинга ДНК (фиг. 1В). Регенерация из этих белых листьев на среде без спектиномицина привела к получению только побегов с дефицитом пигмента, что свидетельствует о полном отсутствии пластидных геномов дикого типа во всех слоях листа и во всех типах клеток.
Оказалось трудным получить семена растений табака, выращенных в стерильной культуре. Случайно в процессе регенерации растения из первичных трансформантов авторы получили периклинальную химеру (S. Poethig, Trends Genetics, 5, 273, 1989), гомоплазматическую по пластидной мутации в L2-слое листа (фиг. 2А). Эту линию трансплантировали в линию табака дикого типа и выдерживали для созревания в теплице (фиг. 2Б). Из семян, полученных из самоопыленных цветков, получили только белые проростки, в которых методом гель-блоттинга ДНК не смогли обнаружить пластидные геномы дикого типа (фиг. 1В).
В пластидах в листьях линии ΔrроВ-растений отсутствуют тилакоидные мембраны. Растения линии ΔrроВ с дефицитом пигмента не способны расти в фотоавтотрофных условиях. Однако, если их выращивать на среде, содержащей сахарозу, с целью компенсировать ее недостаток из-за отсутствия фотосинтеза, они растут нормально, но с меньшей скоростью по сравнению с растениями дикого типа и не проявляют заметных изменений в морфологическом строении органов. Более того, проростки линии ΔrроВ обладали высокой всхожестью и развивались в растения. Эти результаты свидетельствуют о том, что пластидная РНК-полимераза, подобная РНК-полимеразе Е. coli, не требуется для поддержания функций нефотосинтезирующих пластид, необходимых для роста и дифференцировки растения.
Изучение ультраструктуры пластид в мезофильных клетках листа растений линии ΔrроВ показало, что мутантные пластиды имели меньший размер и были более округлыми, чем хлоропласты дикого типа, и в среднем их длина составляла 2-5 мкм по сравнению с 5-9 мкм для хлоропластов дикого типа. Таким образом, пластиды линии ΔrроВ являются более крупными, чем недифференцированные пропластиды, средний размер которых составляет 1 мкм (M.R. Thomas и RJ. Rose, Planta, 158, 329, 1983). Кроме того, пластиды линии ΔrроВ обычно содержали многочисленные везикулы неодинакового размера и формы и у них отсутствовало упорядоченное строение тилакоидных мембран, характерное для активно фотосинтезирующих хлоропластов (фиг.3).
Транскрипция пластидных генов поддерживается в ΔrроВ-пластидах. Можно ожидать, что при отсутствии β-субъединицы не будет происходить транскрипция с помощью пластидных промоторов σ70-типа. С целью выяснить, поддерживалась ли какая-либо транскрипционная активность в ΔrроВ-пластидах, методом гель-блоттинга РНК оценивали накопление РНК. Анализировали транскрипты для двух различных классов пластидных генов (К. Shinozaki и др., EMBO J., 5, 2043, 1986). Первая группа включала гены, кодирующие субъединицы фотосинтезирующего аппарата: оперон psbD/C, кодирующий субъединицы D2 и СР43 фотосинтезирующей системы II; rbcL, кодирующий большую субъединицу рибулозо-1,5-бифосфаткарбоксилазы; и psbA, кодирующий субъединицу D1 реакционного центра фотосинтезирующей системы II. Вторая группа содержала гены для компонентов аппарата экспрессии генов: rрl16, кодирующий субъединицу рибосомного протеина, и ген 16S-рДНК. Количества всех пластидных РНК были стандартизированы по отношению к уровням цитоплазматической 25S-рибосомной РНК.
Неожиданно было обнаружено накопление мРНК для всех изученных генов. Однако роль rроВ-делеции в накоплении транскриптов очень сильно различалась для двух классов генов. Постоянные уровни мРНК, участвующих в фотосинтезе генов psbD/C, rbcL и psbA, снизились в 40-100 раз по сравнению с соответствующими уровнями у растений дикого типа (фиг. 4А), и при более длительном экспонировании были обнаружены сигналы во всех полосах ΔrроВ. В отличие от этого уровни транскриптов ядерных генов, кодирующих полимеразу, оказались подверженными изменению в существенно меньшей степени. Было выявлено 3-кратное снижение 16S-рРНК, а также было обнаружено фактическое увеличение количества транскриптов, полученных с помощью полицистронного оперона, содержащего ген rрl16 (фиг. 4Б). Эти данные свидетельствуют о том, что в то время как экспрессия генов, кодирующих фотосинтезирующий аппарат у растений линии ΔrроВ, является дефектной, РНК генов "домашнего хозяйства" (конститутивных генов) накапливаются приблизительно до уровней дикого типа или даже до более высоких уровней.
Ген 16S-рДНК в растениях линии ΔrроВ транскрибируется с помощью нового промотора. Накопление пластидных РНК подтверждает, что в пластидах, лишенных β-субъединицы, существует РНК-полимеразная активность Е. coli-подобного фермента. Однако из литературы известно, что существует миграция пластидных генов в ядро (S. L. Baldauf и J.D. Palmer, Nature, 334, 262, 1990; J.S. Gantt, S. L. Baldauf, P. J. Calie, N.F. Weeden, J.D. Palmer, EMBO J., 10, 3073, 1991; M.W. Gray, Curr. Op. Genet. Dev., 3, 884, 1993). Следовательно, можно предположить, что транскрипция в пластидах ΔrроВ-типа еще может инициироваться промоторами σ70-типа, если имеется ядерная копия гена rроВ, продукт которого может импортироваться в пластиды и объединяться в функциональный Е. coli-подобный фермент. С целью выяснить, являлись ли пластидные транскрипты, выявленные в ΔrроВ-растениях, продуктами транскрипции промотора σ70-типа, картировали 5'-концы транскриптов четырех генов: rbcL (К. Shinozaki и М. Sugiura, Gene, 20, 91, 1982), 16S-рДНК (A. Vera и М. Sugiura, Curr. Genet. 27, 280, 1995), psbA (M. Sugita и М. Sugiura, Mol. Gen. Genet., 195, 308, 1984) и psbD (W.B. Yao, B.Y. Meng, M. Tanaka, M. Sugiura, Nucl. Acids Res., 17, 9583, 1989), для которых ранее были выявлены сайты инициации транскрипции. Никакие из картированных 5'-концов не соответствовали сайтам инициации промоторов σ70-типа (на фиг. 5А показаны
карты для rbcL и 16S-рДНК). Таким образом, было сделано заключение, что остаточная РНК-полимеразная активность в пластидах ΔrроВ-типа не соответствовала Е. coli-подобному ферменту, а являлась второй уникальной системой транскрипции в пластиде. Эта вторая, отличная от первой РНК-полимеразная активность обозначена в контексте данного описания как кодируемая ядерной ДНК пластидная РНК-полимераза (Nuclear Encoded Plastid RNA polimerase = NEP) с той целью, чтобы ее отличать от Е. coli-подобного фермента, который обозначен как кодируемая пластидной ДНК пластидная РНК-полимераза (Plastid Encoded Plastid RNA polymerase = PEP). Поскольку пластидный геном табака был полностью секвенирован, а последовательности нескольких неидентифицированных рамок считывания не имеют сходства с последовательностями известных субьединиц РНК-полимеразы (К. Shinozaki и др., ЕМВО J., 5, 2043, 1986), транскрипция при участии кодируемой ядерной ДНК РНК-полимеразы основана на продуктах ядерного гена.
При отстутствии в растениях линии ΔrроВ транскрипции от промоторов σ70-типа остается вопрос: какие промоторы были источником пластидных РНК. По данным генетического картирования 5'-конец 16S-pPHK, выявленный в растениях линии ΔrроВ, содержал 62 нуклеотида, расположенных против хода транскрипции от 5'-конца зрелой 16S-pPHK (фиг. 5А). Кэппированием in vitro установлено, что этот 5'-конец принадлежит первичному транскрипту (фиг. 5Б). Известный первичный транскрипт с аналогичным 5'-концом недавно был описан в пропластидах культивируемых в гетеротрофных условиях клеток табака и был обозначен как Р2 (A. Vera и М. Sugiura, Curr. Genet. 27, 280, 1995). Этот транскрипт также присутствует в очень незначительных количествах в клетках листа дикого типа (А. Vera и М. Sugiura, Curr. Genet. 27, 280, 1995; фиг. 5А, более длительная экспозиция, не показано). Последовательность, окружающая сайт инициации, является наиболее высоко консервативной среди всех изученных видов растений и не имеет сходства с консенсусной последовательностью σ70-типа (А. Vera и М. Sugiura, Curr. Genet. 27, 280, 1995). Основываясь на ее широком использовании в растениях линии ΔrроВ, было сделано заключение, что этот уникальный промотор используется аппаратом транскрипции NEP.
В отличие от 16S-pPHK основные транскрипты участвующих в фотосинтезе генов rbcL и psbD/C по данным картирования расположены на ранее охарактеризованных процессированных концах (данные для rbcL приведены на фиг. 5; L. Hanley-Bowdoin, E.M. Orozco, N.-H. Chua, Mol. Cell. Biol., 5, 2733, 1985; J. E. Mullet, E.M. Orozco. N.-H. Chua, Plant Mol. Biol. 4, 39, 1985; S. Reinbothe, С. Reinbothe, С. Heintzen, С. Seidenbecher, В. Parthier, EMBO J., 12, 1505, 1993). Дополнительные концы минорных транскриптов выявлены картированием против хода транскрипции процессированных концов. Таким образом, низкие уровни накопления транскриптов этих фотосинтезирующих генов являются результатом промоторной активности против хода транскрипции и последующего процессинга считываемых РНК, что приводит к образованию транскриптов соответствующих размеров.
Предполагаемая роль двух пластидных систем транскрипции. В растениях линии ΔrроВ обнаружено накопление РНК, транскрибируемых NEP-системой. Это свидетельствует о роли кодируемой ядерной ДНК РНК-полимеразы в поддержании экспрессии пластидных генов "домашнего хозяйства". Вероятно, эти уровни экспрессии достаточны для поддержания роста и дифференцировки нефотоавтотрофных растений. В отличие от этого Е. coli-подобная РЕР-РНК-полимераза необходима для получения высоких уровней транскриптов пластидных генов, необходимых для развития активных в отношении фотосинтеза хлоропластов. Предполагаемая роль кодируемой ядерной ДНК РНК-полимеразы предъявляет высокие требования к ее функции во время ранних фаз развития хлоропласта до того, как РЕР-РНК-полимераза становится активной (J.E. Mullet, Plant Physiol., 103, 309, 1993). Регуляция кодируемой ядерной ДНК РНК-полимеразы в процессе развития подтверждается тем, что промотор кодируемой ядерной ДНК полимеразы Р2 гена 16S-рДНК является более активным в пропластидах клеток культурного табака, чем в хлоропластах листа (A. Vera и М. Sigiura, Curr. Genet. 27, 280, 1995).
Пример II
Транскрипция с участием двух различных РНК-полимераз представляет собой общий регуляторный механизм экспрессии генов у высших растений
Как описано в примере I, накопление транскриптов в растениях, лишенных РЕР-полимеразы, позволяет обнаружить NEP-промотор для оперона пластидной рибосомной РНК (Allison и др., EMBO J., 14, 3721-3730, 1996). Для облегчения картирования дополнительных NEP-промоторов оценивали накопление мРНК для основных классов пластидных генов в растениях ΔrроВ-типа. Новые последовательности промоторов, представленные в данном описании, могут применяться для расширения диапазона видов, для которых возможна такая трансформация пластид, и для осуществления экспрессии представляющих интерес чужеродных генов тканеспепифическим образом.
Материалы и методы примера II
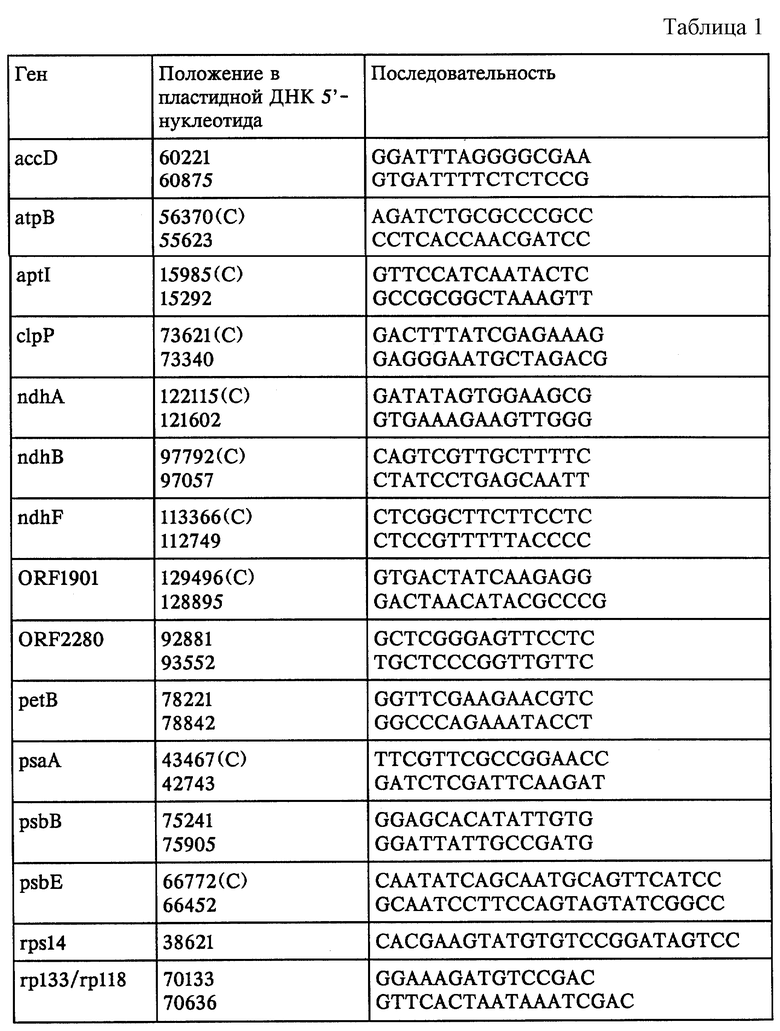
Гель-блоттинг РНК. Общую РНК листа получали с использованием TRIzol (GIBCO BRL), следуя протоколу производителя. РНК подвергали электрофорезу на гелях 1% агарозы/формальдегид, а затем переносили на Hybond N (фирма Amersham), используя устройство для переноса типа Posiblot Transfer (фирма Stratagene). Гибридизацию со случайным образом примированным меченым фрагментом проводили в буфере для быстрой гибридизации (Rapid Hybridization Buffer, фирма Amersham) в течение ночи при 65oС. Двухцепочечные ДНК-зонды получали с помощью случайным образом примированных меченных с помощью 32Р фрагментов ДНК, полученных с использованием ПЦР. Последовательность применяемых для ПЦР праймеров, а также их положения в птДНК табака (К. Shinozaki и др., 1986, см. выше) представлена в табл.1.
мРНК гена rpsl6 зондировали EcoRI-фрагментом, выделенным из плазмиды pJS40, содержащей последовательности между нуклеотидами 4938/5363 и 6149/6656 птДНК табака (Shinozaki и др., 1986, см. выше). Зонд для 25S-pPHK табака получали из плазмиды pKDRl (Dempsey и др., Mol. Plant. Path., 83, 1021, 1993), содержащей EcoRI-фрагмент длиной 3,75 т.п.н. из 25S/18S-локуса табака, клонированного в плазмиде pBR325. Для гибридизации посредством гель-блоттинга с 25S-pPHK 32Р меченый двухцепочечный ДНК-зонд смешивали с немеченой плазмидой pKDRl в количестве, соответствующем 2-кратному избытку относительно количества РНК, присутствующего на фильтре.
Реакции удлинения праймеров. Реакции удлинения праймеров проводили с использованием 10 мкг (дикий тип) или 10 мкг (ΔrроВ-тип) общей РНК листа, как описано у Allison и Maliga, EMBO J., 15, 2802-2809, 1995). Праймеры представлены в табл. 2. Подчеркнутые нуклеотиды также использовали для получения конструкций для кэппинга.
Ступени последовательностей получали с этими же праймерами, используя набор секвеназы II (набор Sequenase II, фирма USB).
Идентификация первичных транскриптов с помощью кэппинга in vitro. Общую РНК (20 мкг) листа из растений дикого типа и ΔrроВ-типа подвергали кэппингу в присутствии [α-32P] -ГTФ (Kennell и Pring, Mol. Gen. Genet., 216, 16-24, 1989). Меченые РНК выявляли путем защиты от рибонуклеазы (A. Vera и М. Sugiura, 1992, см. выше), используя набор RPAII (фирма Ambion). Для получения защищенной комплементарной РНК расположенную против хода транскрипции область 16S-рДНК (нуклеотиды 102526-102761 птДНК) подвергали ПЦР-амплификации с использованием указанных в табл.3 праймеров. 5'-праймеры конструировали таким образом, чтобы добавить сайт рестрикции XbaI (подчеркнут) против хода транскрипции амплифицируемого фрагмента. 3'-праймеры конструировали таким образом, чтобы добавить сайт рестрикции KpnI (подчеркнут) по ходу транскрипции аплифицируемой последовательности. Амплифицированный продукт клонировали в виде XbaI-KpnI-фрагмента в расщепленном рестриктазами XbaI и KpnI векторе pBSKS+ (фирма Stratagene). Для создания немеченой комплементарной к 5'-концу РНК полученную плазмиду линеаризовали с помощью XbaI и транскрибировали с использованием реакции типа Megascript (фирма Ambion) с Т3-РНК-полимеразой. Маркеры (100, 200, 300, 400 и 500 нуклеотидов) получали с помощью набора матриц РНК (RNA Centure Markers Template Set, фирма Ambion), следуя протоколу производителя.
Анализ последовательности ДНК. Анализ последовательности ДНК проводили с использованием программного обеспечения Wisconsin Sequence Analysis Package (фирма Genetics Computer Group, Inc.).
Результаты
На основе данных о накоплении мРНК в листьях дикого типа и ΔrроВ-типа пластидные гены могут быть разделены на три класса. Первый класс включает гены, мРНК которых накапливаются до высоких уровней в листьях дикого типа и присутствуют в очень низких уровнях в листьях растений линии ΔrроВ (фиг. 6А). Генами, принадлежащими к этому классу, являются psaA (ген фотосинтезирующей системы I), psbB и psbE (гены фотосинтезирующей системы II), petB (ген комплекса цитохром b6/f), ndhA (гомолог гена НАДН-дегидрогеназы дыхательной цепи; Matsubayashi и др., Mol. Gen. Genet., 210, 385-393, 1987) и rpsl4 (ген рибосомного протеина). Второй класс включает пластидные гены, мРНК которых накапливаются в приблизительно одинаковых количествах в листьях растений дикого типа и ΔrроВ-типа (фиг. 6Б). К этому классу относятся atpB (ген АТФ-синтазы), ndhF (ген, гомологичный гену НАДН-дегидрогеназы дыхательной цепи; Matsubayashi и др., 1987, см. выше), rpsl6 (ген рибосомного протеина) и ORF1901 (ген с неизвестной функцией; Wolfe и др., J. Mol. Biol., 223: 95-104, 1992). Третий класс включает гены, мРНК которых существенно больше в листьях ΔrроВ-типа, чем в листьях растений дикого типа (фиг. 6В). Типичными для этого класса являются rр133 и rр118 (гены рибосомных протеинов), accD (кодирующий субъединицу ацетил-СоА-карбоксилазы; Sasaki и др., Plant Physiol. , 108: 445-449, 1993) и ORF2280 (мнимая АТФаза с неизвестной функцией; Wolfe, Curr. Genet., 25: 379-383, 1994). Два дополнительных гена этого класса ndhB (ген, гомологичный гену НАДН-дегидрогеназы дыхательной цепи; Matsubayashi и др. , 1987, см. выше) и с1рР (кодирующий протеолитическую субъединицу АТФ-зависимой протеазы Cip; Maurizi и др., J. Biol. Chem., 265, 12546-12552, 1990; Gray и др., Plant Mol. Biol., 15, 947-950, 1990) формируют подгруппу этого класса, для которой характерны высокие уровни мРНК в листьях дикого типа.
Гены АТФ-синтазы atpB и atpI имеют как NEP, так и РЕР-промоторы. Анализ РНК методом гель-блотгинга выявил ряд генов и оперонов, для которых в листьях ΔrроВ-типа поддерживаются высокие уровни транскриптов. Для идентификации дополнительных NEP-промоторов 5'-концы нескольких транскриптов картировали с помощью метода удлинения праймера. 5'-концы могут представлять собой 5'-концы первичных транскриптов, идентифицирующих промотор, или полученные в результате процессинга РНК. Поскольку первичные пластидные транскрипты сохраняют трифосфатные группы на своих 5'-концах, специфическое встраивание в эти молекулы РНК [32Р]-ГМФ с помощью фермента гуанилилтрансферазы позволяет точно разделить концы первичных транскриптов и процессированные концы. Для оперона табака atpB 5'-концы транскрипта были определены Orozco и др. (Current. Gen. , 17, 65-71, 1990) на нуклеотидах в положениях -611, -502, -488, -289 и -255 против хода транскрипции от кодона инициации трансляции (фиг. 7В). 5'-концы нумеруют по их положению относительно кодона инициации трансляции (ATG), при этом нуклеотид, расположенный непосредственно за А против хода транскрипции, имеет положение -1. Метод удлинения праймера позволил выявить каждый из этих 5'-концов в представляющих интерес растениях дикого типа (фиг. 7А). В образце ΔrроВ-типа присутствовали только виды -289-РНК, 5'-концы которых являлись субстратом для гуанилилтрансферазы (фиг. 7Б). Следовательно, -289-РНК траскрибируется с помощью NEP-промотора PatpB-289. Определенный интерес представляет то, что транскрипт -289 присутствует и в листьях дикого типа, хотя и в меньшем количестве, чем в растениях линии ΔrроВ. Транскрипты -255, -488 и -611 отсутствуют в растениях линии ΔrроВ (фиг. 7А). Фрагменты ДНК, содержащие эти промоторы (но не промотор PatpB-289), распознаются E. coli-подобной РНК-полимеразой (Orozco и др., 1990, см. выше) и транскрибируются в пластидах при участии РЕР-полимеразы. Оперон atpA включает гены atpI, -atpH-atpF-atpA (фиг. 8В). В листьях табака дикого типа 5'-концы мРНК были картированы в виде трех областей, расположенных против хода транскрипции от atpI: -209-область, 5'-концы которой расположены на нуклеотидах -212, -209 и -207, и областей, 5'-концы которых расположены на нуклеотидах -130 и -85. В листьях линии ΔrроВ выявлен только транскрипт -207 (фиг. 8А). Этот транскрипт мог быть кэппирован в образце РНК ΔrроВ-типа (фиг. 8Б), и, следовательно, он транскрибируется с помощью NEP-промотора. В реакции кэппирования in vitro образцов РНК дикого типа также был получен сигнал в этом положении. Транскрипты -209 и -212 могут быть образованы в результате активности перекрывающего РЕР-промотора или образованы вследствие образования множественных транскриптов от NEP-промотора в растениях дикого типа. Транскрипт -130, который присутствует только в листовой РНК дикого типа, также мог быть кэппирован (фиг. 8А, 8Б). Поскольку имеются последовательности, подобные элементам -10/-35 на правильном спейсерном участке, расположенном против хода транскрипции от этого 5'-конца, он транскрибируется при участии РЕР-полимеразы.
NEP-Промотор, обеспечивающий высокий уровнь экспрессии гена с1рР в хлоропластах. Ген с1рР субъединицы протеазы также принадлежит к классу, который имеет как NEP-, так и РЕР-промоторы. Анализ удлинения праймера в растениях дикого типа выявил 5'-концы РНК на нуклеотидах в положениях -53, -95 и -173, тогда как в растениях ΔrроВ-типа картирование выявило 5'-концы в положениях -53, -173 и -511 (фиг. 9А). Реакция кэппирования in vitro подтвердила, что каждый из них является первичным транскриптом (фиг. 9Б). Три транскрипта происходили от NEP-промоторов. Промотор PclpP-53 обеспечивает высокий уровень экспрессии как в растениях дикого типа, так и в растениях ΔrроВ-типа, представляя таким образом другой класс NEP-промоторов, обеспечивающих высокий уровень экспрессии в различных типах ткани. Промотор PclpP-53 является достаточно консервативным в шпинате (Westhoff, Mol. Gen. Genet., 1985, 201: 115-123). Дополнительными с1рР-промоторами для NEP являются PpclP-173 и PclpP-511. Поскольку транскрипт PclpP-511 накапливается только в растениях ΔrроВ-типа (фиг. 9А), то он, вероятно, может рассматриваться как регулируемый NEP-промотором. Также следует отметить, что PclpP-511 локализован на кодирующей области psbB, и на его экспрессию может влиять конвергентный РЕР-промотор области psbB (фиг. 9В).
Единственным РЕР-промотором, расположенным непосредственно против хода транскрипции гена с1рР, является PclpP-95. РНК, полученные с помощью этого промотора, накапливаются только в листьях растений дикого типа, а PclpP-95 имеет расположенные против хода транскрипции последовательности, сходные с консервативными элементами -10/-35 (не показано).
Ген ассD транскрибируется исключительно с помощью NEP-промотора. мРНК гена биосинтеза липидов accD накапливается до высоких уровней только в растениях ΔrроВ-типа. Основной транскрипт начинается на нуклеотиде в положении -129 (фиг. 10А), и он может кэппироваться in vitro (фиг. 10Б). Следовательно, эта РНК транскрибируется с помощью NEP-промотора. Поскольку PaccD-129 не проявляет заметной активности в фотосинтезирующих клетках мезофила листа, можно предположить, что он служит для регулируемой NEP-промотором другой схемы тканеспецифичной экспрессии.
NEP-промоторы имеют свободную консенсусную область, соседнюю с сайтом инициации транскрипции. Для выявления консервативных элементов NEP-промотора проводили сравнительный анализ первичной структуры последовательностей, соседних с сайтами инициации транскрипции (фиг. 11). В анализ первичной структуры последовательности были включены девять промоторов, идентифицированных в данном исследовании, и Рггn-62, представляющий собой NEP-промотор, описанный у Allison и др. (1996, см. выше). Кроме того, в этот анализ были включены последовательности для PORF2280-1577 и PORF1901-41, 5'-концы которых, как было показано с помощью кэппирования in vitro, относятся к первичным транскриптам (данные не приведены). Оба эти промотора активны в листьях растений ΔrроВ-типа, но не активны в листьях растений дикого типа. В сравнительный анализ первичной структуры также включены сомнительные NEP-промоторы генов rps2 и rpsl6, количество мРНК которых больше в листьях ΔrроВ-типа. 5'-концы этих транскриптов картировали с помощью анализа удлинения праймера. Метод кэппирования in vitro не позволил получить результаты из-за малого числа копий мРНК (данные не приведены). Множественный сравнительный анализ первичной структуры последовательности областей, непосредственно примыкающих к 5'-концам NEP-промотора, выявил свободную консенсусную последовательность, состоящую из 10 нуклеотидов, расположенную вокруг сайта инициации транскрипции (фиг. 11). Также выявлена консервативность дополнительных нуклеотидов, расположенных против и по ходу транскрипции. Неожиданно было обнаружено, что отсутствует общая консервативная последовательность в PclpP-53, который является единственным NEP-промотором, обладающим высокой активностью в хлоропластах, и в других NEP-промоторах. Принимая во внимание отсутствие подобия последовательностей, эта последовательность не была включена в сравнительный анализ первичной структуры. Последовательности, окружающие сайт инициации транскрипции промотора PclpP-53, приведы отдельно в нижней части фиг. 11.
NEP- и РЕР-полимеразы благодаря способности распознавать различные промоторы обеспечивают механизм избирательной транскрипции пластидных генов (фиг. 12). Приведенные в настоящем описании данные показывают, что некоторые гены имеют только PEP- или только NEP-промоторы, тогда как другие имеют как PEP-, так и NEP-регуляторные последовательности.
Пример III
NEP-промоторы для экспрессии селектируемых генов-маркеров
Для разнообразных и универсальных применений целесообразно обеспечить высокий уровнь экспрессии селектируемых генов-маркеров при трансформации пластид во всех типах тканей. Селектируемые гены-маркеры в применяемых в настоящее время векторах для трансформации пластид экспрессируются с помощью РЕР-промоторов, распознаваемых кодируемой пластидной ДНК РНК-полимеразой. РЕР-полимераза транскрибирует фотосинтезирующие гены и некоторых из генов "домашнего хозяйства" и, следовательно, вероятно является доминантной РНК-полимеразой в активных в отношении фотосинтеза тканях листа. Эффективная трансформация пластид в растениях табака была достигнута за счет трансформации хлоропластов в клетках листа. Однако регенерация растения из листьев большинства важных с сельскохозяйственной точки зрения зерновых культур, включая кукурузу, рис, пшеницу и хлопчатник, невозможна или не может быть осуществлена на практике. Для этих культур трансгенные растения обычно получают путем трансформации эмбриогенной ткани культуры клеток или ткани проростка. Принимая во внимание то, что эти ткани не являются фотосинтезирующими, экспрессия генов-маркеров при участии NEP-промоторов, активных в незеленых тканях, вероятно, является целесообразной с практической точки зрения и должна облегчить трансформацию пластид во всех нефотосинтезирующих типах ткани.
Промотором, особенно пригодным для осуществления экспрессии генов-маркеров, является промотор PclpP-53. Этот промотор обладает высоким уровнем экспрессии в пропластидах растений линии ΔrроВ, и, следовательно, он также может экспрессироваться с высоким уровнем в пропластидах в культурах эмбриогенных клеток, из которых получают трансгенные злаковые растения. Гены-маркеры, экспрессируемые с помощью этих промоторов, также должны быть пригодны для отбора пластидных трансформантов в бомбардируемых культурах листа, поскольку этот промотор, как было обнаружено, является активным в хлоропластах. Гены-маркеры, экспрессируемые с помощью таких промоторов, как PclpP-53, могут найти широкое применение для получения трансформированных пластид.
Селектируемые гены-маркеры могут быть сконструированы с использованием принципов, изложенных в патенте США 5451513 и в находящейся на рассмотрении заявке на патент США 08/189256, которые включены в настоящее описание в качестве ссылки. Трансформирующая конструкция ДНК показана на фиг. 13. В частности, промотор PclpP-53 может быть клонирован против хода транскрипции сегмента ДНК, кодирующего пластидный селектируемый маркер. Сигналы трансляции могут быть получены путем включения пригодных последовательностей ДНК между фрагментом промотора и областью, кодирующей селектируемый маркер. Для получения сигналов терминации транскрипции и для стабилизации химерной мРНК 3'-нетранслируемые сегменты пластидного гена должны быть клонированы по ходу транскрипции относительно селектируемого маркера. Использование 3'-нетранслируемой области генов, экспрессируемых с помощью NEP-промоторов, является предпочтительным, поскольку требования к терминации транскрипции для NEP- и РЕР-полимераз могут быть различными. PclpP-53 является особенно сильным NEP-промотором. Однако растения с трансформированными пластидами могут также быть получены и при использовании слабых промоторов. В предыдущих примерах представлено несколько примеров таких слабых промоторов, например, PclpP-173.
Экспрессия тканеспепифичных пластидных трансгенов, полученных с помощью NEP-промоторов. Тканеспецифичная экспрессия пластидных трансгенов целесообразна для разных аспектов их применения. Тканеспецифичная экспрессия протеина, который делает ткань растения репеллентной или токсичной для корневых нематод, может быть целесообразна в корнях. Однако экспрессия этого же протеина в листьях может истощать ресурсы растений и может влиять на функцию надземных частей растения. Приведенные в настоящем описании NEP-промоторы, экспрессия которых наиболее часто происходит в незеленых тканях, и промоторы, экспрессируемые с помощью NEP-полимеразы в целом, представляют собой богатый источник тканеспецифичных промоторов для трансгенной экспрессии.
Несколько NEP-промоторов, например, PclpP-511, обеспечивают высокий уровень экспрессии в пропластидах растений линии ΔrроВ. Пропластиды присутствуют в съедобной части цветной капусты. Следовательно, можно предположить, что использование этого промотора обеспечит высокий уровень экспрессии чужеродных генов в съедобных частях цветной капусты.
Пластидный ген accD кодирует субъединицу прокариотической ацетил-СоА-карбоксилазы, представляющей собой фермент, участвующий в биосинтезе липидов. Интересно отметить, что в листьях растений дикого типа количество копий мРНК гена accD мало, тогда как их много в пропластидах растений линии ΔrроВ. Эти данные позволяют предположить, что промотор PaccD-129 активен в пластидах незеленых тканей, активно участвующих в биосинтезе липидов, таких, как пластиды развивающихся семян, которые богаты маслом.





Конструкции ДНК для стабильной трансформации пластид включают трансформирующую ДНК с ориентирующим сегментом, маркерный ген, придающий селектируемый фенотип растительным клеткам, содержащим трансформированные пластиды, и сайт клонирования для инсерции дополнительной ДНК, кодирующей представляющий интерес чужеродный ген. Наличие в конструкции ДНК 5'-промоторного элемента, распознаваемого и транскрибируемого пластидной РНК-полимеразой, кодируемой ядерной ДНК, а также дополнительного 5'-промоторного элемента, который распознается и транскрибируется пластидой РНК-полимеразой, кодируемой пластидной ДНК, существенно облегчает трансформацию широкого круга растений и позволяет осуществлять тканеспецифическую экспрессию ДНК, которой трансформируют пластиды многоклеточных растений. 3 с. и 8 з.п. ф-лы, 13 ил., 3 табл.
| PLANT CELL, vol.4, 1992 | |||
| PLANT PHYSIOLOGY, vol | |||
| Клапанный регулятор для паровозов | 1919 |
|
SU103A1 |
| MOLECULAR GENERAL GENETICS, vol.241, 1993. | |||

