Изобретение относится к биотехнологии, в частности к генетической инженерии, и представляет собой сконструированную "in vitro" рекомбинантную плазмидную ДНК, содержащую фрагмент гена U11 герпесвируса человека 6 типа (HHV-6), кодирующий иммунодоминантную часть (3 антигенных эпитопа) белка тегумента р100 с 494 по 842 а.к. Конструкция обеспечивает эффективный биосинтез полипептида в клетках E. coli в виде фрагмента p100 HHV-6, слитого с фрагментом β-галактозидазы с гистидиновым трактом (6*His) на С-конце. Использование очищенного с помощью аффинной хроматографии рекомбинантного белка в иммуноферментном анализе для выявления специфичных к HHV-6 иммуноглобулинов в крови пациентов показывает высокий уровень чувствительности и специфичности. Таким образом, этот рекомбинантный белок может быть использован в качестве антигена HHV-6 для научно-исследовательских работ и серологического тестирования герпесвируса человека 6 типа в клинической практике.
Структурный белок p100 герпесвируса человека 6 типа является белком, наиболее активно реагирующим с сыворотками крови человека при иммунологическом анализе. Был идентифицирован ген U11, кодирующий данный белок, и определена его первичная последовательность для А и В серотипов HHV-6 [1, 2]. Рамка считывания U11 кодирует полипептид 870 а.к., его N-концевая часть гомологична иммуногенному матриксному фосфопротеину рр150 цитомегаловируса человека (HCMV). Антигенные эпитопы р100 локализованы в С-концевой части белка, корорая не имеет существенной гомологии с рр150 HCMV.
Наиболее близким к заявляемому техническому решению (прототипом) является конструкция, описанная в работе [2]. Рекомбинантная плазмида содержит ДНК-последовательность гена U11. Описанная плазмидная конструкция позволяет экспрессировать рекомбинантный белок в виде слитого с полноразмерной β-галактозидазой белка. Недостатками прототипа является наличие полноразмерной β-галактозидазы как неспецифического антигена в последующем ИФА, а также невозможность эффективной аффинной очистки экспрессируемого белка.
Технической задачей изобретения является получение рекомбинантной плазмидной ДНК, обеспечивающей экспрессию фрагмента гена U11 герпесвируса человека 6 типа, кодирующего всю иммунодоминантную часть р100 с 494 по 842 а. к., в составе бактериального плазмидного вектора, кодирующего 6*His-мишень для аффинной очистки белка. Конструкция должна обеспечивать более высокий уровень биосинтеза рекомбинантного p100 HHV-6 в клетках Е. coli и уровень аффинной очистки с использованием 6*Нis-мишени не менее 95-98% рекомбинантного белка.
Поставленная задача решается путем конструирования рекомбинантной плазмидной ДНК pU11HHV6, кодирующей IPTG-индуцируемый биосинтез полипептида, обладающего антигенными свойствами герпесвируса человека 6 типа, в клетках Е. coli в виде фрагмента p100 (с 494 по 842 а. к.), слитого с фрагментом β-галактозидазы и 6*His на С-конце для аффинной очистки.
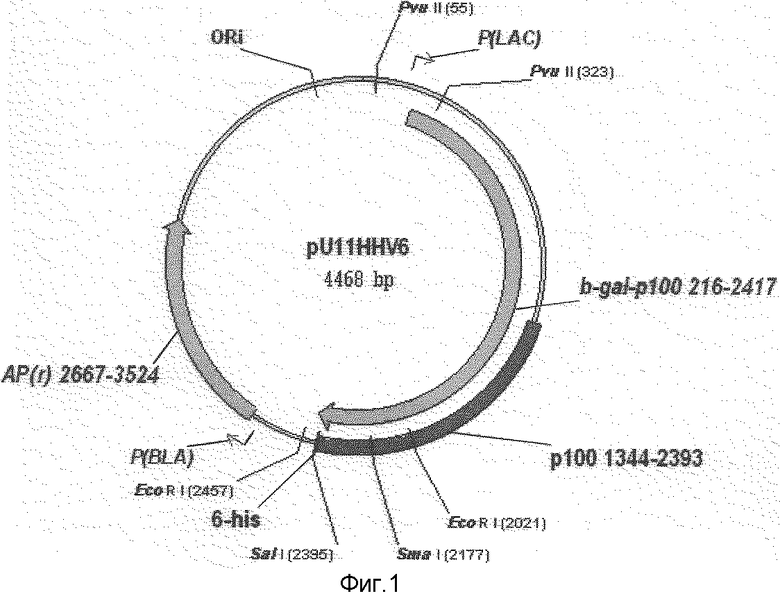
Рекомбинантная плазмидная ДНК pU11HHV6, кодирующая иммунодоминантную часть белка тегумента р100 герпесвируса человека, характеризуется следующими признаками (фиг.1):
- имеет молекулярную массу 2,95 мегадальтон (4,468 т. п. о.);
- кодирует аминокислотную последовательность иммунодоминантной части белка p100 HHV-6 (с 494 по 842 а. к.);
- состоит из:
- HindIII/EcoRV - фрагмента ДНК плазмидного вектора pUR290 (3396 т. п. о.)[5];
- PCR-фрагмента гена U11 (1050 п.о.), кодирующего всю иммунодоминантную часть белка p100 HHV-6 (с 494 по 842 а. к.);
- содержит:
- в качестве генетического маркера ген blа β-лактамазы, определяющий устойчивость трансформированных плазмидой pU11HHV6 клеток к ампициллину;
- нуклеотидную последовательность, которая находится в рамке считывания гена U11 и кодирует полигистидиновый тракт L44, состоящий из 6 молекул гистидина;
- уникальные сайты узнавания рестрикционными эндонуклеазами, имеющие следующие координаты:
Bgl 11 (AGATCT) - 1500, 2199; EcoR I (GAATTC) - 2021; Sma I (CCCGGG) - 2177.
Существенными преимуществами предложенной плазмидной конструкции по сравнению с прототипом являются наличие экспрессируемого фрагмента гена U11, кодирующего только иммунодоминантную часть белка p100 HHV-6, наличие в С-концевой области полигистидинового тракта, что в совокупности обеспечивает уровень синтеза целевого антигена 40-50% с выходом очищенного продукта после аффинной хроматографии до 98-99% от суммарных клеточных белков.
Перечень графических материалов.
Фиг.1. Физическая карта рекомбинантной плазмиды pU11HHV-6.
Фиг. 2. Нуклеотидная последовательность гена U 11, кодирующая иммунодоминантную часть белка р100.
Фиг. 3. Аминокислотная последовательность иммунодоминантного фрагмента белка p100 HVV-6, кодируемого рекомбинантной плазмидой pU11HVV-6.
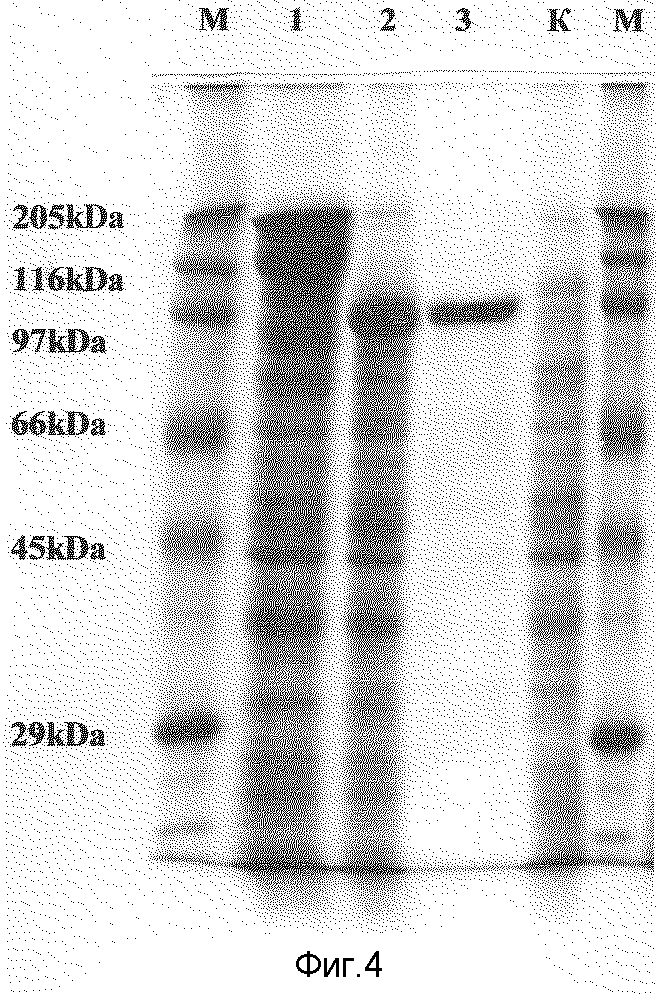
Фиг.4. Электрофореграмма лизатов клеток Е.coli (штамм TG-1)
Дорожки М: маркер белковых весов (Sigma).
Дорожки. К: контроль бактериальных белков - клеточный лизат E.coli.
Дорожки 1-2: клеточные лизаты бактериальных штаммов-продуцентов рекомбинантных антигенов.
1: штамм, содержащий плазмиду pUR-Р100.
2: штамм, содержащий плазмиду рР100.
Дорожки 3: хроматографически очищенный рекомбинантный белок p100.
Изобретение иллюстрируется следующими примерами.
Пример 1. Конструирование промежуточной плазмидной ДНК pUR290-6*his, кодирующей последовательность из 6 гистидинов после сайта узнавания энлонуклеазой рестрикции Sail.
10 мкг плазмидной ДНК pUR290 [5] обрабатывают последовательно эндонуклеазой рестрикции HindIII, фрагментом Кленова и SalI, и сшивают с синтетическим адаптером длинной 50 пар нуклеотидов, обработанным последовательно эндонуклеазой рестрикции KpnI, фрагментом Кленова и SalI:
в лигазной реакции в 20 мкл лигазного буфера. 10 мкл лигазной смеси используют для трансформации компетентных клеток E. coli TG-1 [4]. Трансформанты высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Из выросших за 1 сутки колоний клонов выделяют плазмидную ДНК. ДНК каждого клона проверяют на присутствие вставки 116 п.н., обрабатывая эндонуклеазой рестрикции EcoRI, и на отсутствие сайта эндонуклеазы рестрикции PstI. Отбирают клон, плазмидная ДНК которого содержит последовательность синтетического адаптера.
Пример 2. Конструирование рекомбинантной плазмидной ДНК рU11HHV6.
10 мкг плазмидной ДНК pUR290-6*his обрабатывают рестриктазами BamHI и SalI в соответствии с методикой, описанной в работе [4], и из полученного гидролизата выделяют в 4% полиакриламидном геле векторный фрагмент длиной 5,34 т. п. о.
Проводят полимеразную цепную реакцию (ПЦР) с суммарной ДНК, выделенной из культуры клеток МТ-4, которая была заражена герпесвирусом человека 6 типа, штамм Z29, в соответствии с методикой, описанной в работе [3]. 10 мкг ДНК из реакционной смеси после проведения ПЦР обрабатывают рестриктазами BamHI и SalI и из полученного гидролизата выделяют в 4% полиакриламидном геле фрагмент длиной 1,05 т. п. о.
Полученный фрагмент и векторную часть плазмиды pUR290-6*his сшивают при помощи лигазной реакции в 30 мкл буфера для лигирования. 10-20 мкл реакционной смеси используют для трансформации компетентных клеток Е. coli TG-1 [4] . Трансформанты высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК и анализируют ее путем обработки набором эндонуклеаз рестрикции BglIII, EcoRI, PspALI, BamHI и SalI с последующим электрофоретическим анализом длин рестрикционных фрагментов в 4% полиакриламидном геле. Из 12 проанализированных клонов 2 показали нужный набор рестрикционных фрагментов.
Далее полученную плазмидную ДНК последовательно обрабатывают рестриктазами EcoRV и BamHI. Из полученного гидролизата выделяют в 4% полиакриламидном геле векторный фрагмент длиной 3,42 т. п. о. Достраивают концы выделенного векторного фрагмента с помощью фрагмента Кленова ДНК-полимеразы I и сшивают при помощи лигазной реакции в 30 мкл буфера для лигирования. 10-20 мкл реакционной смеси используют для трансформации компетентных клеток Е. coli TG-1. Трансформанты высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК рU100ННV6 и анализируют ее путем обработки эндонуклеазой рестрикции EcoRI с последующим электрофоретическим анализом длин рестрикционных фрагментов в 4% полиакриламидном геле.
Целевая плазмида pU11HHV6 (фиг.1) содержит уникальные сайты узнавания рестрикционными эндонуклеазами, имеющие следующие координаты:
BgIII - 1500, 2199; EcoRI - 2021, 2457; SmaI - 2177; SalI - 2395.
Окончательную структуру рекомбинантной ДНК pU11HHV6 подтверждают определением нуклеотидной последовательности в районе встроенного фрагмента гена U11HHV6 (фиг.2).
Пример 3. Экспрессия рекомбинантного антигена.
Экспрессию целевого гена U11HHV6 проверяют по наличию рекомбинантного белка 82,46 килодальтон, выделяемого с помощью аффинной хроматографии на Ni-NTA-resin, после индукции IPTG трансформированной целевой плазмидой pU11HHV6 клеток Е. coli TG-1 (фиг.4, дорожка 3).
Таким образом, заявляемое техническое решение позволяет получить экспрессирующую плазмидную ДНК pU11HHV6, кодирующую фрагмент гена U11 HHV6. Трансформированная этой плазмидой культура клеток Е. coli TG-1 при индукции IPTG осуществляет биосинтез полипептида размером 82,46 килодальтон, состоящего из фрагмента β-галактозидазы, фрагмента белка р100 (с 494 по 842 а. к.) герпесвируса человека 6 типа, содержащего 3 антигенных эпитопа, и расположенный на С-конце полигистидиновый тракт. Все это позволяет упростить процесс получения высокоочищенного (до 98-99%) рекомбинантного антигена р100 HHV6, а также увеличить синтез целевого полипептида в 40-50 раз. Этот рекомбинантный белок после аффинной хроматографии может быть использован в качестве ННV6-антигена для серологического анализа герпесвируса 6 типа в клинической практике.
Источники информации
1. Neipel F., Ellinger К., Fleckenstein В. Gene for the major antigenic structural protein (p100) of human herpesvirus 6//J. Virol, 1992, V.66, 6, p.3918-24.
2. Pellet P. et al. A Strongly Immunoreactive Virion Protein of Human Herpesvirus 6 Variant В Strain Z29: Identification and Characterization of the Gene and Mapping of a Variant-Specific Monoclonal Antibody Reactive Epitope.//Virology 1993, V.195, p.521-531.
3. Вирусология. Методы./Под редакцией Б. Мейхи. М.: Мир, 1988, с. 287-284.
4. Маниатис Т., Фрич, Сэмбук Дж. (1984) Молекулярное клонирование. Пер. с англ. М.: Мир.
5. Новое в клонировании ДНК. Методы./Под редакцией Д. Гловера. М.: Мир, 1989, с.140-141.


Изобретение относится к биотехнологии, в частности к генетической инженерии, и представляет собой сконструированную in vitro рекомбинантную плазмидную ДНК, содержащую фрагмент гена U11 герпесвируса человека 6 типа (ННV-6), кодирующий иммунодоминантную часть (3 антигенных эпитопа) белка тегумента р100 с 494 по 842 а.к. Конструкция обеспечивает эффективный биосинтез полипептида в клетках Е.coli в виде фрагмента р100 ННV-6, слитого с фрагментом β-галактозидазы с гистидиновым трактом (6 His) на С-конце. Использование очищенного с помощью аффинной хроматографии рекомбинантного белка в иммуноферментном анализе для выявления специфичных к ННV-6 иммуноглобулинов в крови пациентов показывает высокий уровень чувствительности и специфичности. Рекомбинантный белок может быть использован в качестве антигена ННV-6 для научно-исследовательских работ и серологического тестирования герпесвируса человека 6 типа в клинической практике. 4 ил.
Рекомбинантная плазмидная ДНК рU11HHV6 с молекулярной массой 2,95 мегадальтон, обеспечивающая экспрессию фрагмента гена UI1 герпесвируса человека 6 типа, кодирующего иммунодоминантную часть белка тегумента р100 (с 494 по 842 а. к. ) в клетках бактерий Escherichia coli, которая содержит участок начала репликации; HindIII/EcoRV - фрагмент ДНК плазмиды рUR290; PCR-фрагмент гена U11 (1050 п. о. ), кодирующий иммунодоминантную часть белка р100 ННV6, представленный на фиг. 3; ген-маркер bla - лактамазы, определяющий устойчивость трансформированных плазмидой рU11HHV6 клеток Е. coli к ампициллину; нуклеотидную последовательность, кодирующую полигистидиновый тракт в рамке считывания гена U11; уникальные сайты узнавания рестрикционными эндонуклеазами Bgl II, EcoR I, Sma I.
| US 6174685 С1, 16.01.2001 | |||
| US 5827519 С1, 27.10.1998. |

