Изобретение относится к биотехнологии, белковой инженерии и медицине, конкретно - к получению корового антигена вируса гепатита B(HBcAg), позволяющего экспонировать на поверхности чужеродные эпитопы.
Возможность экспонирования чужеродных эпитопов на поверхности HBcAg [1] открывает перспективы для конструирования широкого спектра химерных капсидов с заданными иммунологическими свойствами. Такое экспонирование достигается в результате внедрения фрагментов ДНК, кодирующих аминокислотные последовательности эпитопа, по специально выбранным участкам гена HBcAg.
Химерные капсиды на основе HBcAg могут быть использованы в первую очередь как компоненты диагностикумов и вакцин. Последнее тем более перспективно, что HBcAg при иммунизации не только обеспечивает высокий уровень образования опосредованных B-клеточным иммунитетом антител, но и обладает ярко выраженным свойством стимулировать Т-клеточный иммунный ответ как на собственные последовательности, так и на сопутствующие эпитопы [2].
Для этих целей сконструированы так называемые "вектора" экспонирования, предназначенные для внедрения по определенным точкам HBcAg природных и синтетических ДНК-копий пептидов, которые представляют структурный или функциональный интерес, т. е. эпитопов вирусных белков, пептидных гормонов и т.п. [1-5]. При этом предполагается выполнение следующих условий:
1. внедренный пептид сохраняет свою нативную конформацию в составе химеры;
2. не нарушает капсидообразующих свойств последней;
3. экспонируется на наружной поверхности.
Таким образом, критическим моментом для HBcAg, как белка-носителя, является выбор района, подходящего для встройки чужеродного эпитопа.
В гене HBcAg существуют природные рестрикционные сайты, которые позволяют осуществлять встройку в N- и C- концы этого гена.
Известны генноинженерные конструкции, кодирующие HBcAg и экспонированными на его поверхности фрагментами pre S- участка вируса гепатита B [4] и эпитопом вируса лейкоза крупного рогатого скота (BLVgp51) [5]. В том и другом случае вставка чужеродного эпитопа произведена по 144-ой C-концевой аминокислоте гена HBcAg. При этом не нарушается самоорганизация корового белка в капсид. Химерные капсиды обладают антигенными свойствами внедренных последовательностей. Однако иммунный ответ на встроенные детерминанты оказался гораздо ниже, чем на сам носитель: во всех опытах титр анти-HBc (10-6) был значительно выше, чем титр антител против внедренных preS-последовательностей (10-3, 10-4) или BLVgp51 (1-3)[4,5].
Таким образом, эпитопы, встроенные в С-конец HBcAg, проявляют "неожиданно" низкую иммуногенность, что признают сами авторы.
Этот факт объясняется недостаточной экспонированностью таких эпитопов на поверхности HBcAg, поскольку относительно С-конца корового антигена известно, что он содержит поли-аргининовую последовательность, которая служит для упаковки вирусной ДНК и ориентирована внутрь коровой частицы.
Более перспективным местом для встроек, по-видимому, является область центральной иммунодоминантной петли HBcAg e1, соответствующая 71-89 аминокислотным остатком корового белка. Авторами работы [6] было показано, что этот район кора поверхностно экспонирован и является основной антигенной детерминантной HBcAg. В работах [7,8] были получены обнадеживающие результаты при встройке чужеродных эпитопов в этот район кора.
Наиболее близким аналогом, выбранным в качестве прототипа, является рекомбинантная плазмида pFS14 [8, прототип], несущая под контролем tac промотора полноразмерный ген HBcAg, содержащий уникальный Xho I сайт в области, соответствующей 83 аминокислотному остатку корового белка. Xho I сайт был введен в район центральной иммунодоминантной петли HBcAg при клонировании олигонуклеотида, кодирующего последовательность pre-S2 HBcAg, которая не удаляется при гидролизе XhoI.
Таким образом, полученная авторами [8] плазмидная конструкция не является универсальной, поскольку требует предварительного удаления ранее клонированного эпитопа pre-S2. К недостаткам прототипа следует отнести низкий уровень экспрессии (1% суммарного клеточного белка), а также необходимость использования индуктора tac промотора ИПТГ (изопропилтио-b-D-галактопиранозид).
Технической задачей изобретения является создание универсальной плазмидной конструкции, обеспечивающей конститутивный синтез аутентичного HBcAg и позволяющей экспонировать чужеродные эпитопы, представленные оптимальным образом на поверхности капсида HBcAg, т.е. служить базой для создания новых конструкций.
Поставленная задача решается путем введения с помощью олигонуклеотиднаправленного мутагенеза XhoI-сайта рестрикции в ген HBcAg в район, соответствующий 81-му аминокислотному остатку кора, и конструированием рекомбинантной плазмиды pUHBc, кодирующей синтез HBcAg, в котором первые три аминокислотных остатка заменены семнадцатью аминокислотными остатками - галактозидазы, и состоящей из: фрагмента генома вируса гепатита B, кодирующего HBcAg и содержащего уникальный рестрикционный XhoI сайт; плазмидного вектора с молекулярной массой 2,36 МДа и размером 3576 п.о., обеспечивающего экспрессию гена, кодирующего HBcAg.
В качестве фрагмента генома вируса гепатита B (ayw серотипа) используют BamHI фрагмент плазмиды p7,5C [9] длинной 890 п.о., содержащий C-ген вируса гепатита B, в котором делетированы первые 3 аминокислотных остатка.
В качестве фагового вектора, обеспечивающего встройку C-гена и необходимого для введения мутации, используют бактериофаг M13mp10 [10].
В качестве олигонуклеотида-мутагена для введения мутации по методу [11], обеспечивающего появление уникального XhoI сайта, используют синтетический фрагмент ДНК:
5'-ctaggtccctcgagattggatc-3'.
В качестве плазмидного вектора, обеспечивающего встройку C-гена и его экспрессию под контролем "сильного" lac промотора, используют плазмиду pUC19 [12].
Место встройки выбрано не случайно, а на основании компьютерного анализа структуры белка HBcAg и ряда соображений: позиция 81 а.к. расположена в центре петли e1 и находится рядом с пролином, который, как известно, обеспечивает поворот в белковой цепи, и поэтому встроенный эпитоп должен оказаться ориентирован наружу. Организация уникального XhoI-сайта была произведена с учетом вырожденности кода таким образом, чтобы аминокислотная последовательность белкового продукта мутантного гена в области e1 была аутентична последовательности природного HBcAg.
Сущность изобретения заключается в том, что для получения C-гена вируса гепатита B, содержащего уникальный рестрикционный сайт XhoI, выделяют фрагмент ДНК, содержащий C-ген, и клонируют его в ДНК фага M13mp10 по сайтам BamHI. Полученную одноцепочечную ДНК M13mp10-HBcAg используют для олигонуклеотиднонаправленного мутагенеза по методу Золлера и Смита [11]. В результате получают C-ген, в котором произведена замена 242-го нуклеотида A из C и 244-го C на G. В результате этого в данной области (соответствующей 81-ему аминокислотному остатку HBcAg) возникает уникальный XhoI сайт рестрикции без изменения аминокислотной последовательности кора. Для экспрессии мутантного C-гена BamHI фрагмент клонируют в полилинкерную область плазмиды pUC19. В результате в целевой плазмиде pUHBc первые три аминокислотные остатка HBcAg заменены семнадцатью аминокислотными остатками β-галактозидазы.
Для получения штамма-продуцента HBcAg компетентные клетки E.coli JM103 трансформируют сконструированной плазмидой pUHBc. Полученный таким образом штамм E.coli JM103/pUHBc характеризуется следующими признаками.
Морфологические признаки. Клетки мелкие утолщенной палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. При росте на агаре "Дифко" - колонии круглые, гладкие, прижатые, мутные, блестящие, серые, край ровный. При росте в жидких средах (на минимальной среде с глюкозой или LB бульоне) образуют интенсивную ровную муть. Клетки растут при температуре 39oC при оптимуме pH от 6,8 до 7,0.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к ампициллину (300 мгк/мл), обусловленную наличием плазмиды.
Существенными отличиями штамма E.coli JM103/pUHBc является то, что он обусловливает конститутивный синтез HBcAg с уровнем экспрессии около 16% суммарного клеточного белка, что в 16 раз превосходит прототип.
Полученный штамм депонирован во Всесоюзной коллекции промышленных микроорганизмов во ВНИИ генетики под номером ВКПМ-В-6956.
Экспрессию HBcAg осуществляют в клетках E.coli JM103 без использования индуктора ИПТГ. Индикацию экспрессии осуществляют с помощью белкового электрофореза и иммуноблоттинга. Рекомбинантный белок узнается антисывороткой, используемой в качестве положительного контроля в тест-системе "Векто-HBcAg-антитела" (АО "Вектор Бест" Россия). Уровень экспрессии определяют с помощью денситометрии полиакриламидного геля, окрашенного Кумасси. Уровень экспрессии составляет 16% от суммарного клеточного белка.
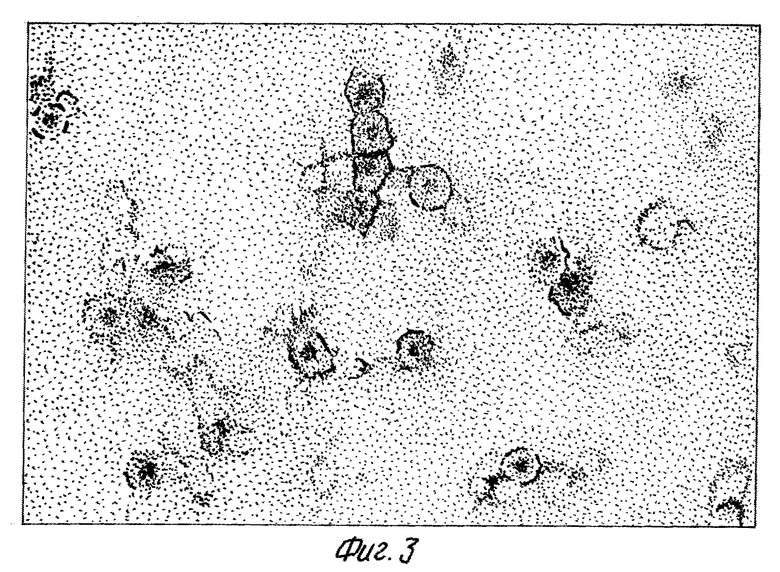
С помощью электронной микрофотографии показано, что продуцируемый белок собирается в частицы, по форме и морфологии подобные природному кору.
В качестве модельного пептида (для иллюстрации возможности экспонирования чужеродных эпитопов на поверхности HBcAg) была выбрана детерминанта белка E2 (233-240) вируса Венесуэльского энцефаломиелита лошадей (ВЭЛ). Фрагмент ДНК, колирующий пептид, имеет следующую нуклеотидную последовательность:
5'-tcgatcaacgataaatgggtgtacaacagc
agttgctatttacccacatgttgtcgagct-5'
Указанный синтетический фрагмент ДНК был клонирован по уникальному XhoI сайту плазмиды pUHBc. Полученная в результате клонирования плазмида обеспечивала синтез клетками E. coli гибридного HBcAg, содержащего эпитопы ВЭЛ. Такие гибридные капсиды хорошо узнавались антителам к ВЭЛ. Это говорит о том, что встроенная детерминанта находится в природной конформации и располагается на поверхности кора.
В качестве подтверждения универсальности заявляемой конструкции был проведен эксперимент по экспонированию основного нейтрализующего эпитопа HBsAg ( 137-147) на поверхности HBcAg. Фрагмент ДНК, кодирующий указанный пептид, имеет следующую последовательность:
5'-tcgatctgcaccaaaccgaccgatggcaactgc
agacgtggtttggctggctaccgttgacgagct -5'
Указанный синтетический фрагмент ДНК был клонирован по уникальному XhoI сайту плазмиды pUHBc. Полученные гибридные коровые частицы анализируют с помощью иммуноэлектронной микроскопии с использованием моноклональных антител (МАТ) к HBsAg, меченных иммунным золотом. На электронной микрофотографии отчетливо видно, что частицы золота, связанные с МАТ к HBsAg, располагаются на поверхности гибридных частиц HBcAg-HBsAg и отсутствуют на нативных частицах без встройки (фиг. 5). Эти данные свидетельствуют о том, что встроенный в область 81-а.к. пептид экспонирован на поверхности химерных частиц HBsAg-HBcAg и находится в своей нативной конформации.
Таким образом, впервые получена универсальная плазмидная конструкция pUHBc, которая может быть использована двояко: во-первых, для получения собственно аутентичного HBcAg, и, во-вторых, для создания новых гибридных частиц HBcAg, экспонирующих чужеродные эпитопы в области e1. Кроме того, полученный рекомбинантный штамм E.coli JM103/pUHBc обеспечивает продукцию рекомбинантного HBcAg с уровнем 16% от суммарного клеточного белка, что в 16 раз выше, чем в прототипе, и без использования индуктора, также в отличие от прототипа.
Изобретение иллюстрируется следующими фигурами:
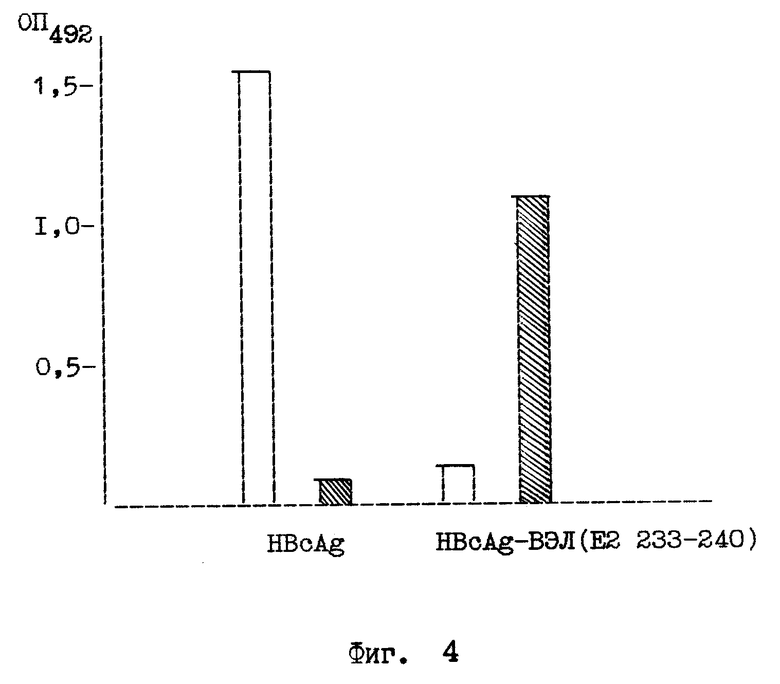
на фиг. 1 - схема плазмидной ДНК pUHBc и способа ее конструирования. BamHI, XhoI - сайты рестрикции; Klenov - фрагмент Кленова ДНК полимеразы I E. coli; ligase - ДНК-лигаза фага 14;  - олигонуклеотид-мутаген; на фиг. 2 - анализ клеточного лизата E.coli JM103, несущего плазмиду pUHBc в 12% полиакриламидном геле, и денситометрия геля. 1 - лизат клеток E.coli, трансформированных pUHBc; 2 - денситограмма геля (стрелкой указан пик, соответствующий HBcAg); на фиг. 3 - электронная фотография рекомбинантного HBcAg - продукта плазмиды pUHBc, препараты HBcAg готовят методом негативного контрастирования 2%-ным раствором фосфорно-вольфрамовой кислоты и исследуют в электронном микроскопе "JEM 100C"; на фиг. 4 - антигенные свойства химерных капсид HBcAg-ВЭЛ по данным иммуноферментного анализа. На планшеты сорбируют лизаты E. coli, содержащие либо продукт плазмиды pUHBc, либо химерный HBcAg-ВЭЛ, затем проводят связывание с антителами к HBcAg (белые прямоугольники) или с антителами к вирусу ВЭЛ (заштрихованные прямоугольники), выявляют связавшиеся антитела антивидовыми конъюгатами, меченными пероксидазой; на фиг. 5 - иммуноэлектронная фотография гибридных частиц HBcAg-HBsAg и рекомбинантного HBcAg без встроек, частицы предварительно инкубируют с моноклональными антителами к HBsAg, а затем с анти-IgG, меченным коллоидным золотом, затем препараты готовят методом негативного контрастирования 2%-ным раствором фосфорно-вольфрамовой кислоты и исследуют в электронном микроскопе "JEM 100C".
- олигонуклеотид-мутаген; на фиг. 2 - анализ клеточного лизата E.coli JM103, несущего плазмиду pUHBc в 12% полиакриламидном геле, и денситометрия геля. 1 - лизат клеток E.coli, трансформированных pUHBc; 2 - денситограмма геля (стрелкой указан пик, соответствующий HBcAg); на фиг. 3 - электронная фотография рекомбинантного HBcAg - продукта плазмиды pUHBc, препараты HBcAg готовят методом негативного контрастирования 2%-ным раствором фосфорно-вольфрамовой кислоты и исследуют в электронном микроскопе "JEM 100C"; на фиг. 4 - антигенные свойства химерных капсид HBcAg-ВЭЛ по данным иммуноферментного анализа. На планшеты сорбируют лизаты E. coli, содержащие либо продукт плазмиды pUHBc, либо химерный HBcAg-ВЭЛ, затем проводят связывание с антителами к HBcAg (белые прямоугольники) или с антителами к вирусу ВЭЛ (заштрихованные прямоугольники), выявляют связавшиеся антитела антивидовыми конъюгатами, меченными пероксидазой; на фиг. 5 - иммуноэлектронная фотография гибридных частиц HBcAg-HBsAg и рекомбинантного HBcAg без встроек, частицы предварительно инкубируют с моноклональными антителами к HBsAg, а затем с анти-IgG, меченным коллоидным золотом, затем препараты готовят методом негативного контрастирования 2%-ным раствором фосфорно-вольфрамовой кислоты и исследуют в электронном микроскопе "JEM 100C".
A - частицы HBcAg,
B - частицы HBcAg-HBsAg.
Для лучшего понимания сущности предлагаемого изобретения ниже следуют примеры его осуществления.
Пример 1. Способ введения уникального XhoI сайта в область 81 аминокислотного остатка HBcAg и конструирования рекомбинантной плазмиды pUHBc.
В качестве источника C-гена используется плазмида p7,5C [9], содержащая BamHI фрагмент ДНК (890 п.о.) вируса гепатита B, субтипа ayw.
50 мкг плазмиды p7,5C расщепляют BamHI эндонуклеазой в реакционной смеси, содержащей 100 ед. BamHI, 50 мМ трис-HCl pH 7,6; 10 мМ MgCl2, 100 мМ NaCl; 1 мМ DTT. Реакцию ведут 1 час при 37oC. После этого фрагмент ДНК длиной 890 п.о. (содержащий C-ген с делетированными нуклеотидами, соответствующими первым трем аминокислотным остаткам HBcAg) выделяют методом электрофореза в агарозном геле с последующей электроэлюцией.
Отдельно расщепляют репликативную форму ДНК фага M13mp10 BamHI эндонуклеазой, инактивируют фермент прогреванием при 68oC 30 мин и лигируют с 1 мкг BamHI фрагмента плазмиды p7,5C в стандартных условиях [13]. Полученной лигазной смесью трансформируют компетентные клетки E.coli JM109(RecA-). С помощью рестрикционного анализа отбирают клоны, содержащие вставку и обозначаемые далее M13HBc. Отобранные клоны используют для наработки одноцепочечной ДНК и проведения мутагенеза по методу Золлера-Смита [11]. 1 пмоль одноцепочечной ДНК M13HBc смешивают с 20-кратным избытком олигонуклеотида, несущего запланированную мутацию в 10 мкл раствора, содержащего 20 мМ DTT. Пробирку с реакционной смесью нагревают до 75oC и постепенно охлаждают до комнатной температуры. Затем добавляют 10 мкл раствора, содержащего 20 мМ трис-HCl pH 7,5, 10 мМ MgCl2, 10 мМ DTT, 1 мМ ATP, 1 мМ dATP, 1 мM dTTP, 1 мМ dGTP, 1 мМ dCTP, 4 ед. акт. ДНК - лигазы, 2 ед. акт. фрагмента Кленова ДНК-полимеразы I и инкубируют смесь в течение ночи при 10oC. Смесью трансформируют клетки E.coli. Клоны, содержащие мутантный фаг, отбирают гибридизацией, с использованием в качестве зонда олигонуклеотида-мутагена. Отобранные фаговые ДНК, обозначенные M13HBcX, анализируют с помощью рестрикционного анализа и секвенирования по методу Сангера [14].
Репликативную форму ДНК M13HBcX разрезают эндонуклеазой BamHI. Фрагмент ДНК, содержащий мутантный C-ген (несущий уникальный XhoI сайт), выделяют из агарозного геля с помощью электроэлюции. Плазмиду pUC19 расщепляют BamHI нуклеазой, инактивируют фермент прогреванием при 68oC 30 мин и лигируют с 1 мкг BamHI фрагмента фага M13HBcX. Полученной лигазной смесью трансформируют компетентные клетки E.coli JM103. С помощью рестрикционного анализа отбирают клоны, содержащие вставку. Полученную таким образом плазмиду обозначают pUHBc. Схема плазмидной ДНК pUHBc и способ ее конструирования представлены на фиг. 1.
Пример 2. Характеризация рекомбинантного HBcAg - продукта плазмиды pUHBc.
Клетки E. coli JM103, содержащие плазмиду pUHBc, растят на среде LB в течение ночи без использования индуктора ИПТГ. Биомассу собирают центрифугированием и анализируют с помощью электрофореза по Лэммли и иммуноблоттинга с антисывороткой, используемой в качестве положительного контроля в тест-системе "Векто-HBcAg-антитела" (АО "Вектор Бест" Россия). (В качестве контроля сравнения использовали рекомбинантный HBcAg, любезно предоставленной Г. П. Борисовой [4]).
При сканировании полосы в полиакриламидном геле, соответствующей HBcAg, показан уровень продукции, составляющий 16% суммарного клеточного белка (фиг. 2).
Выделенные с помощью ультрацентрифугирования в градиенте сахарозы частицы рекомбинантного HBcAg исследуют с помощью электронной микроскопии (фиг. 3). В полученных препаратах обнаружены частицы, по форме и морфологии аналогичные природному кору.
Пример 3. Иллюстрация возможности экспонирования эпитопа Вируса Венесуэльского энцефаломиелита лошадей на поверхности HBcAg.
Плазмиду pUCHBc расщепляют XhoI нуклеазой, инактивируют фермент прогреванием при 68oC 30 мин и лигируют по методике [12] с синтетическим фрагментом ДНК, кодирующим эпитоп Вируса Венесуэльского энцефаломиелита лошадей (ВЭЛ) E2 (233 - 240), имеющим следующую нуклеотидную последовательность:
5'-tcgatcaacgataaatgggtgtacaacagc
agttgctatttacccacatgttgtcgagct-5'
Полученной лигазной смесью трансформируют компетентные клетки E.coli JM103. С помощью рестрикционного анализа отбирают клоны, содержащие вставку. Полученную таким образом плазмиду обозначают pUHBc-ВЭЛ.
Клетки E.coli JM103, содержащие плазмиду pUHBc-ВЭЛ, растят на среде LB в течение ночи без использования индуктора ИПТГ. Биомассу собирают центрифугированием и анализируют с помощью электрофореза по Леммли и иммуноблоттинга с антисывороткой к пептиду ВЭЛ E2 (233 - 240). Показано, что гибридный белок HBcAg-ВЭЛ выявляется антителами к пептиду ВЭЛ E2 (233 -240).
Выделенные с помощью ультрацентрифугирования в градиенте сахарозы частицы химерного HBcAg-ВЭЛ исследуют с помощью иммуноферментного анализа (ИФА). С помощью ИФА было показано, что химерные частицы реагируют как с антителами к пептиду ВЭЛ E2 (233 - 240), так и антителами к целому вирусу ВЭЛ, но перестают узнаваться антителами к HBcAg (поскольку встройка была проведена в иммунодоминантной области HBcAg) (фиг. 4). Эти данные говорят о том, что пептид экспонирован на поверхности химерных частиц HBcAg-ВЭЛ и находится в своей нативной конформации.
Пример 4. Иллюстрация возможности экспонирования основного нейтрализующего эпитопа HBsAg на поверхности HBcAg.
Плазмиду pUHBc расщепляют XhoI нуклеазой, инактивируют фермент прогреванием при 68oC 30 мин и лигируют по методике [12] с синтетическим фрагментом ДНК, кодирующим эпитоп основного нейтрализующего эпитопа HBsAg (137 - 147), имеющим следующую нуклеотидную последовательность:
5'-tcgatctgcaccaaaccgaccgatggcaactgc
agacgtggtttggctggctaccgttgacgagct-5'
Полученной лигазной смесью трансформируют компетентные клетки E.coli JM103. Полученную таким образом плазмиду обозначают pUHBc-HBsAg, растят на среде LB в течение ночи без использования индукторов ИПТГ. Биомассу собирают центрифугированием, проводят выделение гибритных коровых частиц с помощью ультрацентрифугирования в градиенте сахарозы и анализируют полученные частицы с помощью иммуноэлектронной микроскопии с использованием моноклональных антител (МАТ) к HBsAg, меченных иммунным золотом.
На электронной микрофотографии отчетливо видно, что частицы золота, связанные с МАТ к HBsAg, располагаются на поверхности гибритных частиц HBcAg-HBsAg и отсутствуют на нативных частицах без встройки (фиг. 5). Эти данные свидетельствуют о том, что встроенный в область 81-а.к. пептид экспонирован на поверхности химерных частиц HBcAg-HBsAg и находится в своей нативной конформации.
Таким образом, впереди получена универсальная плазмидная конструкция для создания новых гибридных частиц HBcAg, экспонирующих чужеродных эпитопы в области e1 кора, а также штамм-продуцент, обеспечивающий высокую экспрессию аутентичного HBcAg без использования индукторов.
Литература
1. Грен Э. Я., Пумпен П.П. Рекомбинантные вирусные капсиды - новое поколение иммуногенных белков.// ЖВХО.-1988.-Т.33.-531-536.
2. Clarke B. E., Newton S.E., Carrol A.R., Francis M.J., Appleyard G., Syred A. D., Highfild P.E., Rowlands D.J., Brown F. Improved immunogenicity of a peptide epitope after fusion to hepatitis В core protein.// Nature-1987.-U.330.-P.381-384.
3. Francis M.J., Hastings G.Z., Brown A.L., Grace K.G., Rowlands D.J., Brown F. , Clarke B.E. Immunological properties of hepatitis В core antigen fusion proteins.// Proc.Natl. Acad.Sci.USA.-1990.-U. 87.- P. 2545-2549.
4. Борисова Г.П., Берзинь И.Г., Цибиногин В.В., Лосева В.Я., Пушко П.М., Осе В.П., Дрейлиня Д.Э.. Грен Э.Я. Кор-антиген вируса гепатита В в качестве носителя функционально активных эпитопов: экспонирование preS-участков на капсидах.// Доклады АН СССР.-1990.- Т.312.N3.-C-751-754.
5. А.с. СССР N 1742331, МКИ C 12 N 12/48, опубл. БИ N 23 от 23.06.92.
6. Argos P., Fuller S.D. A model for the hepatitis В virus core protein: prediction of antigenic sites and relationship to RNA virus capsid.// EMBO J.-1988.-V.7.-P.819-824.
7. Schodel F. , Milich D. R. , Will H. Hepatitis В virus nucleocapsid/pre-S2 fusion proteins expressed in attenuated Salmonella for oral vaccination.// J.lmmunol.-1990.-V. 145, N12.-P.4317-4321.
8. Schodel F., Weimer Т., Will H., Milich D. Recombinant HBV particles carrying immunodominant B-cell epitopes of the HBV pre-S2 region.// Vaccines'90. Cold Spring Harbor (N.Y.), 1990.-P.193-198.
9. Патент РФ N 1651555, МКИ C 12 N 15/51, опубл. БИ N 10 от 30.05.94.
10. Messing J., Vieira J. A new pair of M13 vectors for selecting either DNA strand of double-digest restriction fragments.//Gene.-1982.-V. 19, N 3. -P. 269-276.
11. Zoller M. J., Smith M. Oligonucleotide-directed mutagenesis of DNA fragments cloned into M13 vectors. In:Meth.Enzymol./Wu R., Grossman L., Moldave K., eds.N.Y., London: Acad.Press.-1983 .-U .100 .-P. 468-500.
12. Vierra J., Messing J. The pUC plasmids, an M13mp7-derived system for insertion mutagenesis and sequencing with systhetic universal primers. //Gene. -1982. -V. 19. -P. 259-265.
13. Маниатис Т., Фрич Э., Сэмбрик Дж. Методы генетической инженерии. Молекулярное клонирование. -М.: Мир.-1984.
14. Sanger F., Coulson A.R., Barrel B.G. Cloning in singlestranded bacteriophage as an aid to rapid DNA sequensing. //J. Mol. Biol. -1980.-V. 143. -P. 161-178.

Рекомбинантная плазмидная ДНК р UНВс предназначена для кодирования белка корового антигена вируса гепатита В. Обладает молекулярной массой 2,36 МДа и размером 3576 п.о. Рекомбинантная плазмидная ДНК содержит плазмидный вектор рUС19, промотор Plac, уникальные сайты рестрикции Nde 1(183), Nar 1(236), Afl III (806), Cfr 10 1 (1779), Ssp 1 (2501), Bam Н1 - фрагмент, содержащий c-ген вируса гепатита В, генетические маркеры, ген ампициллин-резистентности. Изобретение позволяет экспонировать чужеродные эпитопы, представленные на поверхности капсида НВс Ag и служить базой для создания новых конструкций. 5 ил.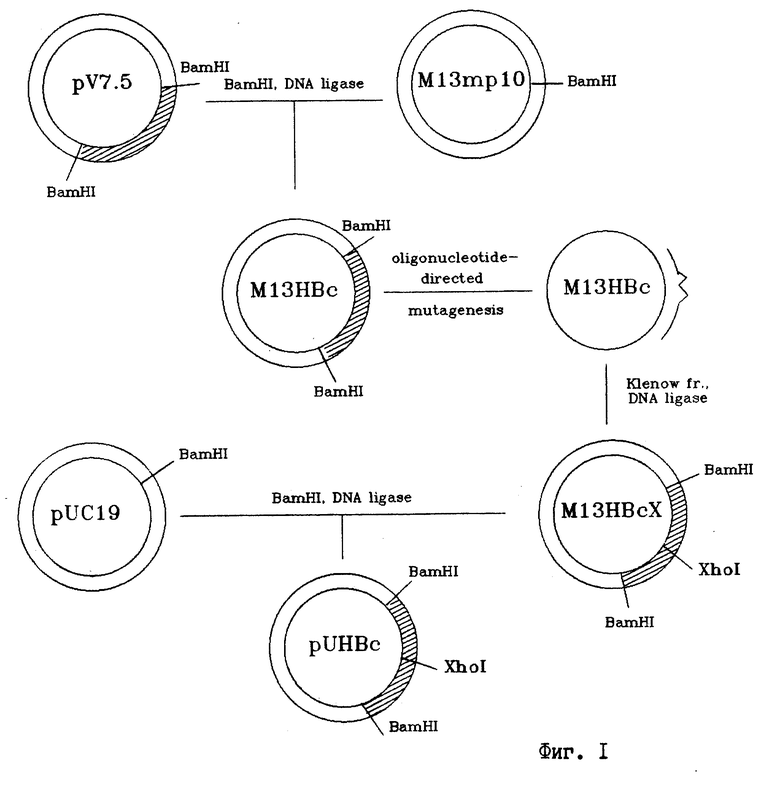
Рекомбинантная плазмидная ДНК p UHBc, кодирующая ген белка корового антигена вируса гепатита B, позволяющая экспонировать чужеродные эпитопы на поверхности кора, молекулярной массой 2,36 Md и размером 3576 п.о., содержащая плазмидный вектор p UC19 размером п.о., раскрытый по Bam HI-сайту и содержащий ген ампициллин-резистентности, сайт инициации репликации плазмиды p BR 322, часть lac-гена из M13mp 19, промотор Pl ас, полилинкерную последовательность из M 13 mp 19, уникальные сайты рестрикции: NdeI (183), Nar 1(236), Afl Ш (806), Cfr 10 1(1779), SspI (2501), - Bam HI - фрагмент ДНК плазмиды p 7,5 C размером 890 п.о., содержащий C-ген вируса гепатита B и уникальный XhoI-сайт в районе, соответствующем 81 - 82 аминокислотным остатком HB c Ag, генетические маркеры: bla -ген ампициллин (ген β-лактамазы), определяющий устойчивость к ампициллину при трансформации E.coli.

