Изобретение относится к белкам для новых АТР-(АТФ)-чувствительных калиевых каналов, huКАТР-1 и ruКАТР-1, которые экспрессируются в различных тканях человеческого и крысиного происхождения, и к генам, кодирующим их. Упомянутые белки и гены могут использоваться в качестве диагностических и лечебных средств при заболеваниях, связанных с калиевыми каналами, таких как диабет, гипертензия и эндокринная недостаточность.
Этиология диабета известна, главным образом, вследствие нарушений секреции инсулина в панкреатических β-клетках. Следовательно, ожидается, что выяснение молекулярного механизма секреции инсулина играет важную роль при выяснении причин диабета и разработке лекарственных препаратов против диабета, но детали такого молекулярного механизма еще не известны.
Уже выяснено, что АТР-чувствительный калиевый канал (КАТР), присутствуя на клеточной мембране, играет ведущую роль в клеточных функциях, таких как секреция и мышечные сокращения, посредством связывания состояния метаболизма в клетках с мембранным потенциалом.
В 1983 КАТР-канал впервые обнаружен в сердечной мышце [Noma, A., Nature 305: 147 (1983)], и затем подтверждено его присутствие в таких тканях, как панкреатические β-клетки [Cook, D.L. et al., Nature 311: 271 (1984), Misler, S. et. al. , Proc. Natl. Acad. Sci. U. S.A. 83: 7119 (1986)], гипофиз [Bernardi, H. et al., Proc, Natl. Acad. Sci. U.S.A., 90: 1340 (1993)], скелетные мышцы [Spruce, A.E. et al., Nature, 316: 736 (1985)] и головной мозг.
Кроме того, предположено, что в таких КАТР-каналах существует молекулярная неоднородность [Ashcroft, F.M., Annu. Rev. Neurosci, 11: 97 (1988)].
В частности, в панкреатических β-клетках АТР, образовавшийся при метаболизме глюкозы, является причиной притока ионов кальция из кальциевого канала благодаря запиранию КАТР-канале, что вызывает деполяризацию, приводящую в результате к секреции инсулина. Из этого очевидно, что КАТР-канал играет ведущую роль в регуляции секреции инсулина.
КАТР-канал принадлежит к семейству калиевых каналов, проявляющих электрофизиологически внутреннюю ректификацию, на основании чего семейство калиевых каналов, проявляющих внутреннюю ректификацию подразделяют на четыре подсемейства - ROMK1, IRK1 GIRK1 и с КАТР-1, основываясь на степени идентичности аминокислотной последовательности.
Несмотря на это, молекулярное строение КАТР-канала в панкреатических β-клетках не выяснено. Кроме того, отсутствуют сведения о детальной белковой структуре новых АТР-чувствительных калиевых каналов (huКАТР-1 и ruКАТР-1) настоящего изобретения и об образовании комплексов с другими белками, например с белком, связывающимся сульфонилмочевиной.
Чтобы осуществить выделение, идентификацию и функциональный анализ нового мембранного канала, требуются весьма сложные методы, такие как методы молекулярной биологии, клеточной биологии и электрофизиологические методы.
Поскольку это так, авторы настоящего изобретения широко и всесторонне использовали такие методы, чтобы выделить геномы человека и крысы, и кДНК, кодирующие новый КАТР-канал (uКАТР-1), экспрессированный в различных тканях млекопитающих, и идентифицировать их аминокислотные последовательности (см. фиг. 1, 2, 3 и 4). Идентифицированный uКАТР-1-канал экспрессируют в ооцитной системе Xenopus и в клеточных линиях млекопитающих.
Электрофизиологический анализ показал, что uКАТР-1-канал представляет собой АТР-чувствительный калиевый канал, проявляющий внутреннюю ректификацию. uКАТР-1-канал, будучи убиквитарно экспрессированным в тканях млекопитающих, включая человека и крыс, включается в поддержание мембранного потенциала через основной энергетический метаболизм.
Как упоминается выше, настоящее изобретение относится к АТР-чувствительному калиевому каналу (uКАТР-1), который убиквитарно присутствует у млекопитающих, и охватывает белки для АТР-чувствительного калиевого канала, идентифицированные аминокислотные последовательности, кодирующие их, плазмиду, имеющую такие включенные последовательности, и, кроме того, рекомбинантные клетки (трансформанты), имеющие такую трасфектированную в них плазмиду. Кроме того, настоящее изобретение включает изолированные uКАТР-1-белки и рекомбинантные белки, родственные им вещества, такие как агонисты и антагонисты, и лекарственные композиции, включающие диагностические и лекарственные средства для генотерапии.
huКАТР-1 человеческого происхождения состоит из 324 аминокислотных остатков (см. фиг. 1) с молекулярной массой 47965, в то время как ruКАТР-1, происходящий от крысы, состоит, подобным образом, из 424 аминокислотных остатков (см. фиг.4) с молекулярной массой 47960. Эти два калиевых канала обнаруживают 98% идентичность аминокислотной последовательности, и такая заметная гомология приводит нас к предположению, что uКАТР-1 выполняет обычные, структурные и функциональные основные действия во всех клетках млекопитающих. Между прочим, uКАТР-1 участвует в мембранном потенциале и энергетическом метаболизме, что вызывает предположение, что он может найти применение в качестве лекарственного вещества, действующего как предупреждающее нарушения при необычных, исключительных условиях метаболизма, включая эндокринные заболевания, например диабет, голодание и ишемию.
Например, приток и отток ионов кальция, вызываемый открытием и запиранием uКАТР-1 при начале ишемии, тесно связаны с ишемическими нарушениями. Иными словами, существует вероятность того, что агонисты и антагонисты открытия и запирания uКАТР-1 могут составить средство, подавляющее ишемические нарушения.
При сравнительных исследованиях аминокислотной последовательности huKАТР-1 и ruКАТР-1 и других калиевых каналов подтвердилось, что ruКАТР-1 настоящего изобретения принадлежит к новому семейству калиевых каналов внутренней ректификации; центральная область белка uКАТР-1 показывает увеличенную гомологию с другими калиевыми каналами внутренней ректификации. Гидросхема (hydropathy plot) указывает на присутствие двух гидрофобных областей, которые состоят из двух трансмембранных областей, характерных для калиевых каналов внутренней ректификации, и одной области отверстия [Nicholas, C.G., Trends Pharmacol. Sci., 14: 320 (1993), Jan, L.Y. and Jan, Y.N., Nature, 371, 119 (1994).
В отношении ruКАТР-1 [Inagaki, N. et al., J.B.C., 270: 5691 (1995)] сообщается, что во второй внутриклеточной области существует два потенциальных сайта фосфорилирования протеинкиназы, зависящего от сАМР (Thr-234 и Ser-385), и семь потенциальных сайтов фосфорилирования, зависящего от протеинкиназы С (Ser-224, Thr-345, Ser-354, Ser-379, Ser-385, Ser-391 и Ser-397), в то время как имеются один (Thr-63) и четыре потенциальных сайта фосфорилирования, зависящего от казеинкиназы-II (Thr-234, Ser-281, Thr-329 и Ser-354) в первой и второй внутриклеточных областях, соответственно, при отсутствии N-связанного сайта гликозилирования во внутриклеточных областях. Такие же результаты получены с huKАТР-1 [Inagaki, N. et al., в печати (1995)].
Затем авторы настоящего изобретения идентифицировали нуклеотидные последовательности и полные аминокислотные последовательности huKАТР-1 и ruКАТР-1, создавая, таким образом, возможность синтеза в больших количествах не только самих белков huKАТР-1 и ruКАТР-1, но также их мутантов, путем экспрессии ДНК, кодирующих huKАТР-1 и ruКАТР-1 и их мутантов в бактериях или клетках животных с применением известных методов генной инженерии. Также следует добавить, что huKАТР-1 и его фрагменты являются пригодными для гибридизационной диагностики ДНК элиминированного huKАТР-1, причем мутанты huKАТР-1 используются при исследованиях обмена сахара в клетках, в частности, при инсулинзависимом и инсулиннезависимом диабете.
ДНК новых huKАТР-1 и ruКАТР-1 по настоящему изобретению идентифицируют на основе библиотеки кДНК и геномной библиотеки. ДНК, кодирующая huKАТР-1, показывает длину в 9,7 т.п.о., состоит из трех экзонов и присутствует на хромосоме 12р11.23. Хромосомная ДНК может быть получена путем зондирования геномной библиотеки с применением кДНК для uKАТР-1 и также его фрагментов. Выделенная ДНК uKАТР-1 для получения мутантов может быть легко подвергнута нуклеотидной элиминации, инсерции и замещению известными методами.
Используя известные технические приемы, к uKАТР-1 или его вариантам по 5'- или 3'-концам легко присоединить нуклеотидные последовательности, кодирующие другие белки или синтетические полипептиды, и получить посредством этого слитые белки или их производные.
Например, слитый белок получают в форме белка-предшественника и подвергают расщеплению in vivo или in vitro, чтобы посредством этого обнаружить его функции; в дополнение к своей собственной функции такой слитый белок обеспечивает направленность на ткань или мембранную ориентацию. В таком случае слитые белки содержат связывающие сахара аминокислоты и могут быть модифицированы до производных, обладающих свойствами тканевой ориентации или физиологической активностью, активированной путем добавления новых цепей сахаров.
Чтобы получить uKАТР-1, его мутанты или их производные, соответствующие кодирующие ДНК вводят в воспроизводимую плазмиду и инкубируют клетки-хозяева, трансформированные такой плазмидой. Клетки-хозяева включают бактерии, дрожжи и клетки животных.
Для клонирования дезоксирибонуклеотидов подходят прокариоты, такие как бактерии. Например, плазмида рВR 322, произведенная от Е. соli, содержит ген, резистентный к ампициллину или тетрациклину, и может обеспечить практические средства идентификации трансформированных клеток. Кроме того, микробные плазмиды содержат промотор, который может быть использован для экспрессии их собственных белков. Кроме прокариотов могут хорошо работать эукариоты, такие как дрожжи, причем можно использовать плазмиду YRр7, особенно когда допускается экспрессия в дрожжах вида Saccharomyces [Steinchomb et al. , Nature, 282: 39 (1979)].
Клетки животных также используют в качестве клеток-хозяев, и, в частности, проста для применения и является обычным способом инкубации клеток позвонков [Krause and Paterson, Tissue Culture, Academic Preess (1973)]. В качестве клеточных линий упоминаются AlT-20, клетки HeLa, яичника китайского хомячка (СНО), СОМSM6, COS-7 и т.п. Для регуляции функции экспрессионной плазмиды в таких клеточных линиях используют промоторы полиомавируса, аденовируса 2, цитомегаловируса и ОВ-40, при этом рсМV представляет собой плазмиду, которая находит широкое применение в системах экспрессии клеток животных [Thomsen et al., PNAS, 81: 659 (1984)].
Последовательности ДНК для канального белка и huKАТР-1 и ruКАТР-1 в соответствии с настоящим изобретением начинаются с инициирующего кодона "АТG". В случаях, когда для синтеза такого белка используют рекомбинантные клетки, нет необходимости добавлять АТG к нужной ДНК, что делает манипуляцию легкой. В результате, когда uKАТР-1 экспрессируется в прокариоте, трансформированной Е. соli, как правило, синтезируется белок с аминокислотной последовательностью, начинающейся с Меt. N-Концевой Меt полученного в результате белка может быть элиминирован в зависимости от цели применения.
В случаях, когда uKАТР-1 синтезируют в рекомбинантных клетках животных, аналогично биосинтезируются белки, содержащие Меt или элиминированные на N-конце, и оба белка пригодны для отдельно обозначенных целей применения.
uKАТР-1 и его фрагменты могут быть введены животным с целью их иммунизации, чтобы посредством этого получить антитела. Иммунизация животных также позволяет продуцировать антитело из клеток, секретирующих нужное антитело.
Стало легко получать uKАТР-1 в больших количествах, что обеспечивает, таким образом, лучшее понимание того же процесса на молекулярном уровне. Соответственно, получение uKАТР-1 и его мутантов или аналогов увеличивает возможность разработки диагностических или лечебных средств для заболеваний, связанных с калиевыми каналами.
В частности, такие белки могут использоваться в методах исследования в веществе, предназначенном для диагностики и лечения, или в веществе, которое вызывает агонистическое или антагонистическое действие uKАТР-1. Например, тестирование с клетками животных может быть проведено путем инъекции в клетки кДНК uKАТР-1 для проведения экспрессии с последующим добавлением сульфонилмочевины для изучения их взаимодействия [Kayano, T. et al., J. Biol. Chem., 265: 13 276 (1990), Example 4].
Кроме того, получена подходящая информация о ДНК-последовательностях uKАТР-1, облегчающая получение ДНК или РНК, кодирующих их неполные последовательности. Такие относительно короткие ДНК-последовательности обладают способностью к гибридизации с геном, который выбирается, и могут найти применение в качестве зондов, которые являются эффективными для детекции кДНК в различных тканях.
Зонды, полученные с использованием uKАТР-1, могут использоваться для продуцирования нуклеиновых кислот, способных к гибридизации с различными организмами и их тканями. Полученные в результате нуклеиновые кислоты могут быть того же типа, как uKАТР-1 или его изоформа, и включают нуклеиновые кислоты, кодирующие новые белки.
Полученные зонды применимы при генной диагностике заболеваний, связанных с калиевыми каналами; исследования могут проводиться с нуклеотидными последовательностями пациентов, гибридизованными с зондом, способным детектировать генные заболевания.
Прежде в качестве лечебных средств против диабета и гипертензии применяли блокаторы и агенты для открывания калиевого канала. Переработанные в фармацевтические препараты, uKАТР-1 и его мутанты, их производные и моноклональные тела к ним могут быть введены пациентам, чтобы посредством этого, через нейтрализацию, частично снять вредное действие, оказываемое избытком таких блокаторов или открывателей, введенных при лечении. Когда сам uKАТР-1 показывает функциональную недостаточность, такие фармацевтические препараты могут вводиться для восполнения таких дефицитных функций uKАТР-1.
Настоящее изобретение включает получение лекарственных средств для генотерапии, применяемой при поддерживающем способе лечения. Нуклеотидные последовательности для uKАТР-1 или его мутантов и их производных могут быть введены в плазмиду или в стволовые клетки, которые затем даются пациентам, и открывают возможность применения в качестве лекарственного средства для генотерапии.
Ниже приводятся примеры для более детальной иллюстрации настоящего изобретения со ссылками на прилагаемые чертежи.
На прилагаемых чертежах изображается следующее.
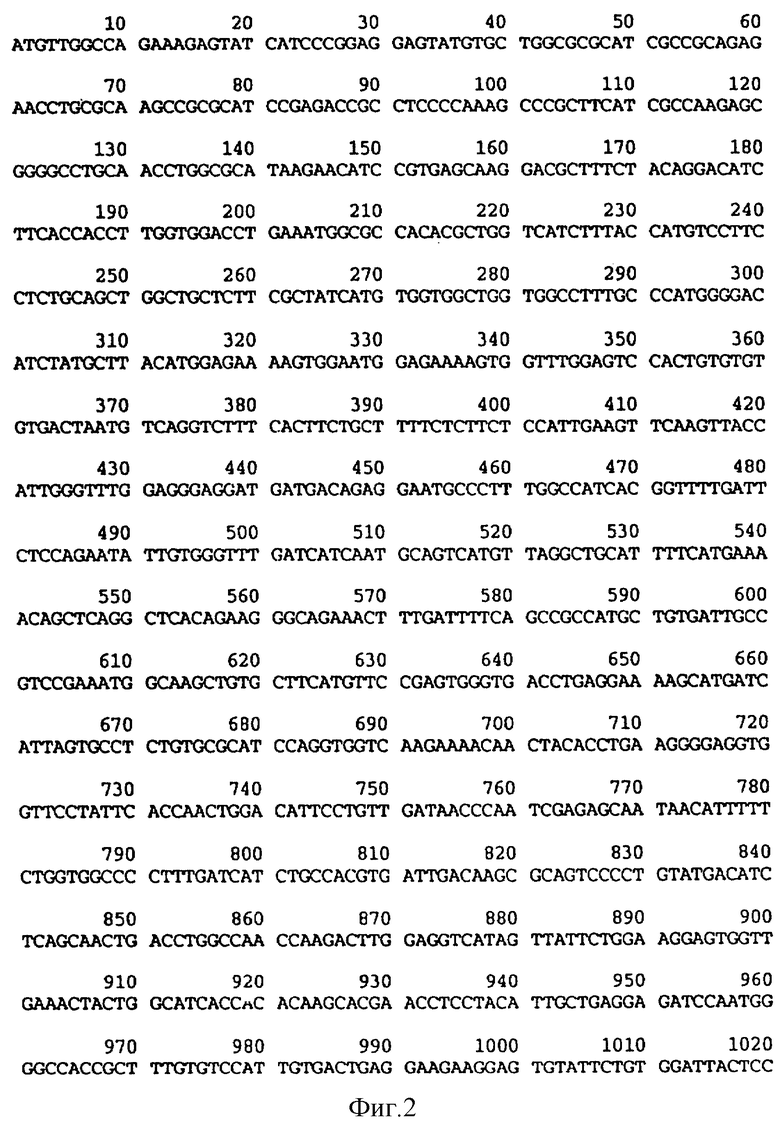
На фиг. 1 приводятся аминокислотные последовательности, соответствующие последовательностям оснований, показанным на фиг.2.
На фиг.2 и 2A (фиг.2A - продолжение фиг.2) приводится последовательность оснований uKАТР-1 человеческого происхождения, полученная в примере 5.
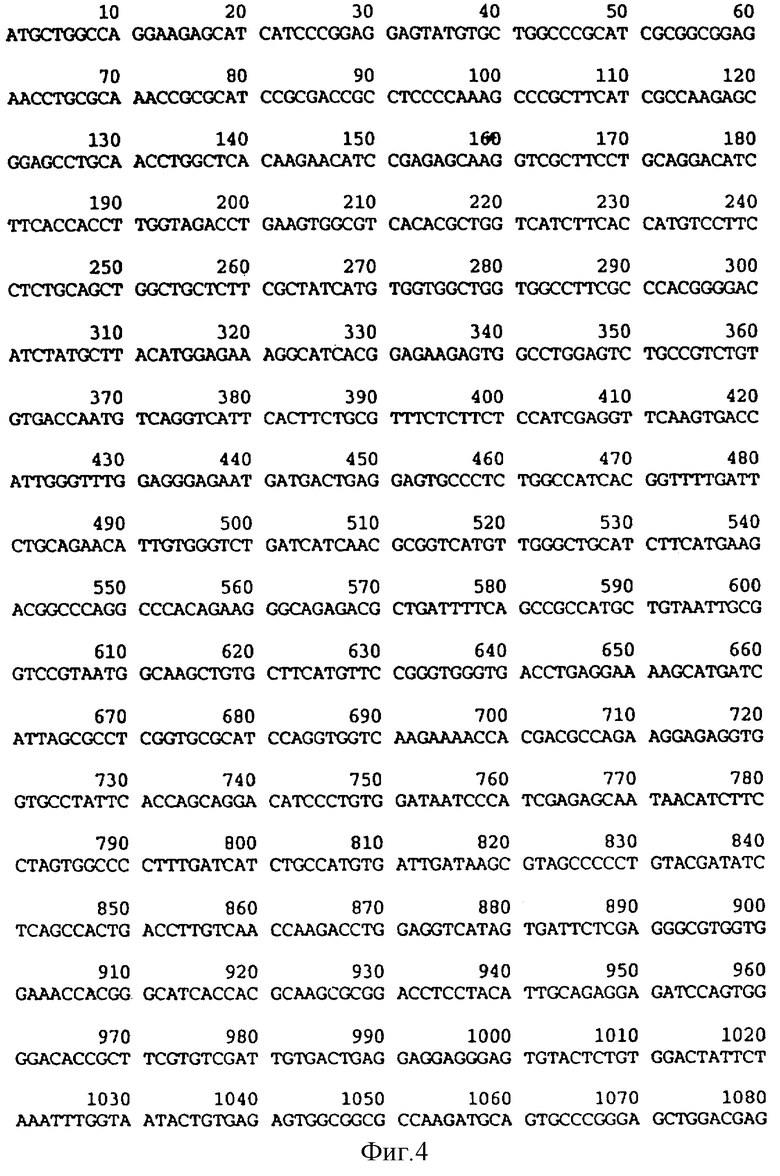
На фиг.3 дается аминокислотная последовательность, соответствующая фиг. 4.
На фиг.4 и 4A (фиг.4A - продолжение фиг.4) приводится последовательность оснований ruКАТР-1 крысиного происхождения.
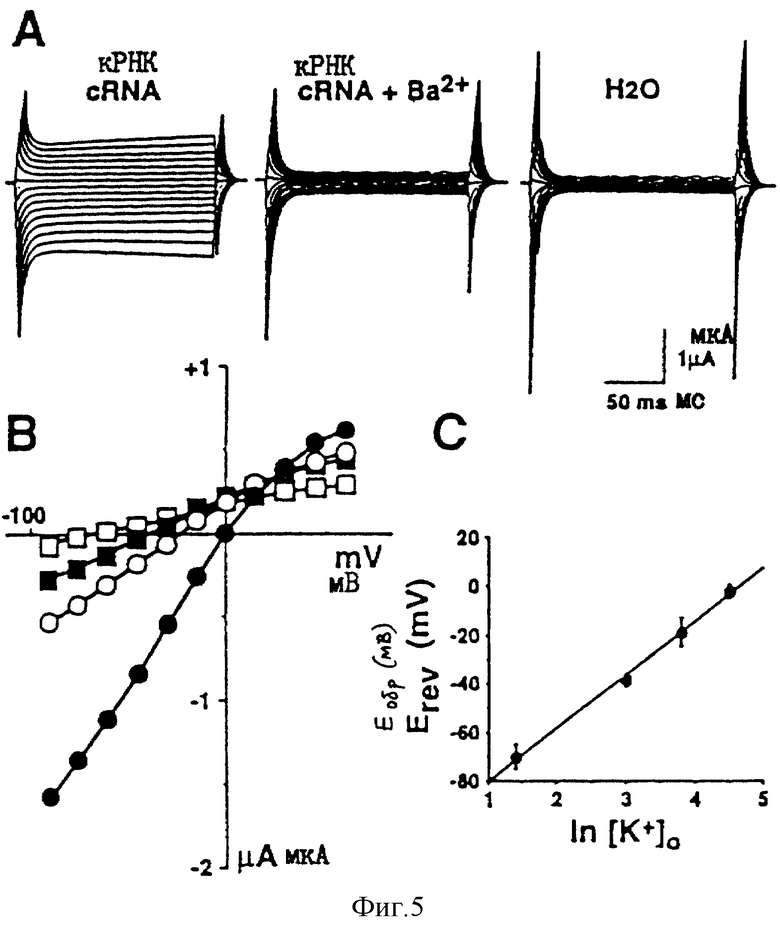
На фиг. 5: А - результаты электрофизиологического анализа ruКАТР-1 с использованием ооцитов Хеnopus. Ооциты, инъецированные кРНК ruКАТР-1, обнаруживают внутреннюю ректификацию, при условиях, когда концентркция [К+] во внеклеточной жидкости составляет 45 мМ, однако ректификация блокируется 300 мкМ Ва2+, добавляемого во внеклеточную жидкость. При контроле, который заключается в инъекции воды, наблюдают образование только очень слабого электрического тока; В - график величины зависящего от концентрации калия тока против напряжения, подтверждающий, что uКАТР-1, очевидно, является калиевым каналом внутренней ректификации; С - график величины обратимого напряжения против логарифма внеклеточной концентрации К+ в ооците, инъецируемом кРНК для ruКАТР-1, указывающий на зависимость обратимого напряжения от внеклеточной концентрации К+.
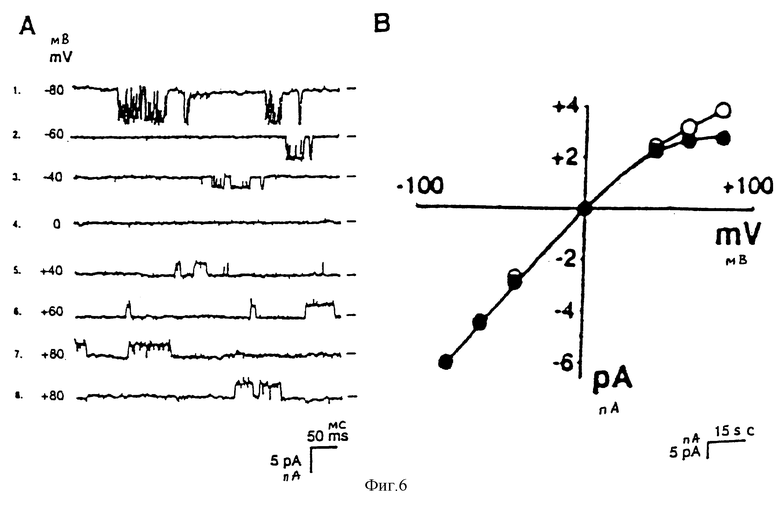
На фиг. 6 дается однокальный анализ трасформированных НЕК 239 клеток, в которых экспрессирован uKАТР-1: А - регистрация одноканального тока; В - соотношение ток - напряжение, что показывает присутствие К+-тока, показывающего внутреннюю ректификацию.
Пример 1. Клонирование кДНК нового калиевого канала внутренней ректификации (ruКАТР-1)
Фрагмент кДНК GIRK - крысиного регулирующего G-белка, калиевого канала внутренней ректификации, амплифицируют методом полимеразной цепной реакции (ПЦР). Используя в качестве зонда фрагмент кДНК крысиного GIRK, помещенного 32Р, проводят поиск в библиотеке кДНК, полученной из островков Лангерганса крысы в векторе λgt22. Выделенную кДНК ruКАТР-1 вставляют в подходящие фрагменты ДНК и после субклонирования в М13mp18 или mр19, выполняют секвенирование оснований методом обрыва цепи (см. фиг.5 или 6).
Пример 2. Экспрессия в ооцитах Xenopus laevis и электрофизиологический анализ
Инъецируют 20 нг кРНК, синтезированной in vitro из плазмиды рGEM11Z, содержащей кДНК ruКАТР-1 полной длины, с РНК-полимеразой, после перевода в линейную форму путем обработки ферментом рестрикции Not1, в ооциты Xenopus, и затем, спустя 2 или 3 дня, проводят электрофизиологический анализ (см. фиг.5). Как иллюстрируется фиг.5 А и В, наблюдают К+-электроток, показывающий слабую внутреннюю ректификацию. Путем добавления Ва2+ во внеклеточную жидкость К+-электроток подавляют.
Пример 3. Одноканальный анализ клеток НЕК 239, содержащих экспрессированный ruКАТР-1
Выращивают клетки НЕК 239 в минимальной поддерживающей среде Игла с добавлением 10% лошадиной сыворотки. Экспрессирующую плазмиду (рсМV6b), несущую кДНК, кодирующую ruКАТР-1 полной длины, трансфецируют в клетки НЕК 239 с применением липофектамина, и получают трансформированные клетки НЕК 239. Трансформированные клетки, полученные таким способом, подвергают одноканальному анализу, причем результаты приводятся на фиг.6 А и В.
Как очевидно из фиг.6 А и В, наружный электрический ток, текущий через канал, подавляется внутриклеточным Мg2+, что показывает, что uKАТР-1 является К+-каналом внутренней ректификации; uKАТР-1 демонстрирует одноканальную проводимость примерно 70 пс. Фиг.5 иллюстрирует влияние АТР на активность канала uKАТР-1 при наблюдении по методу наизнанку. Когда на внутреннюю сторону клеточной мембраны добавляют 1 мкМ АТР, канал открывается, но закрывается полностью после добавления 1 мА АТР. Результаты указывают, что uKАТР-1 является АТР-регулируемым uKАТР-каналом.
Пример 4. Блоттинг-анализ РНК
По 20 мкг РНК, экстрагированных по отдельности из разных тканей и клеточных линий, а также по 10 мкг РНК, экстрагированных из гипофиза и щитовидной железы, денатурируют формальдегидом и подвергают электрофорезу в 1%-ном агарозном геле с последующим переносом на нейлоновую мембрану. Проводят гибридизацию, используя в качестве зонда кДНК uKАТР-1, меченного 32Р, причем наблюдают экспрессию мРНК uKАТР-1 почти во всех тканях.
Пример 5. Клонирование кДНК и гена uKАТР-1 человеческого происхождения
Чтобы выделить кДНК, кодирующую uKАТР-1 человеческого происхождения, выполняют поиск в библиотеке кДНК легкого человека, используя в качестве зонда кДНК, меченного 32Р, ruКАТР-1 крысиного происхождения. Полученный в результате клон подвергают субклонированию в М13mp18, M13mp19 и pGEM3Z с последующим секвенированием оснований по методу обрыва цепи.




Изобретение относится к области биотехнологии и касается нового белка АТР-чувствительного калиевого канала и кодирующего его дезоксирибонуклеотида. Сущность изобретения включает новый белок повсеместно распространенного АТР-чувствительного калиевого канала человеческого происхождения, имеющий аминокислотную последовательность, представленную на фиг.1, а также дезоксирибонулеотид, который имеет последовательность оснований, представленную на фиг.2. Преимущество изобретения заключается в идентификации аминокислотной последовательности нового белка АТР-чувствительного калиевого канала. 2 с. и 1 з.п. ф-лы, 6 ил.
| US 5356775 A, 18.10.1994 | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Экономайзер | 0 |
|
SU94A1 |
| NOBUYA JNAGAKIT et al | |||
| Cloning and Functional characterization of a novel ATP sensitive potassium channel ubiguitously expressed in Rat tissues including pancreatic islits, pituitary, skelеtal muscle and heart, J | |||
| of Biol | |||
| Chemistry | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| ПРИБОР ДЛЯ ИСПЫТАНИЯ РАФИНАДНОГО САХАРА НА ИЗЛОМ | 1927 |
|
SU5991A1 |

