Изобретение относится к селекции растений и может быть использовано для создания растений подсолнечника, устойчивых к биотическим и абиотическим стрессам, а также с измененным составом жирных кислот или запасных белков.
Подсолнечник принадлежит к четырем главным масличным культурам мира. Для России подсолнечник является главной масличной культурой, обеспечивающей в питании населения до 30% потребляемых энергокалорий в виде масла, маргаринов и жиров, произведенных на основе подсолнечного масла.
Высокоурожайные и высокомасличные сорта и линии подсолнечника в настоящее создаются методами традиционной селекции. Однако проблемы потери урожая от неблагоприятных климатических факторов, поражения фитопатогенными микроорганизмами и насекомыми весьма актуальны. Поэтому имеется настоятельная необходимость в радикальном улучшении созданных сортов подсолнечника, увеличении продуктивности в условиях разнообразных биотических и абиотических стрессов.
Для многих важных сельскохозяйственных культур задача получения растений, устойчивых к стрессовым факторам или с измененным качеством продукта, успешно решается путем использования генно-инженерных технологий. В настоящее время трансгенные сорта картофеля, хлопка, риса, кукурузы, устойчивые к насекомым и гербицидам, а также сорта рапса и сои с измененным составом жирных кислот занимают около 40 миллионов гектаров в США, Аргентине, Канаде и Китае. Трансгенные сорта кукурузы, устойчивые к насекомым, начинают распространяться в Европе. Первые работы по генетической трансформации клеток растений были осуществлены на подсолнечнике, но развитие генно-инженерных работ с подсолнечником отстает от других культур. Так, на сегодняшний день в США и странах Европы зарегистрировано около 3000 полевых испытаний трансгенной кукурузы, а подсолнечника - порядка 30. Поскольку для России подсолнечник является наиболее рентабельной и основной масличной культурой, создание растений подсолнечника, устойчивых к разнообразным неблагоприятным условиям среды и с измененным качеством масла, представляется важной экономической задачей.
Впервые методика получения трансгенного подсолнечника была опубликована в 1990 г. (В.Schrammeijer et al., Plant Cell Report, 9: 55-60, 1990), в котором использовалась методика агробактериального переноса генов в апикальную меристему проростков. Стабильно воспроизводимая агробактериальная технология трансформации подсолнечника была получена позже, чем для других сельскохозяйственных культур. Полевые испытания трансгенного подсолнечника были начаты в 1991 г. В дальнейшем методика, основанная на агробактериальной инфекции, была улучшена (D.Bidney et al., Plant Mol. Biol., 18: 301-303, 1992) за счет того, что перед сокультивацией с агробактерией эксплант подвергался поранению бомбардированием микроскопическими частицами металла. Эффективность трансформации была доведена до 2-4%.
Существует альтернативный способ генетической трансформации растений. К 1988 году был разработан способ прямого ввода генов (Т.Klein, J.Sanford et al. , Proc. Nat. Acad. Sci. USA, 85: 4305-4309, 1988). Авторы назвали свой способ "биологической баллистикой" (biolistic), его также называют "методом бомбардмента" или "методом генной пушки". Баллистический метод имеет ряд преимуществ над агробактериальным способом - он позволяет вводить одновременно несколько генов в одно растение. Например, описан случай одновременного введения 9, 10 и даже 13 генов в рис (Lili Chen et al., Nature Biotechnology, 16: 1060-1063, 1998). Такой результат невозможно получить при использовании агробактерии. Кроме того, баллистический метод позволяет трансформировать виды растений, устойчивые к агробактериальной инфекции.
Способ заключается в следующем: на золотые или вольфрамовые частицы размером в десятые доли микрона прецепитируются ДНК, содержащие генные конструкции. В специальном устройстве (существуют электрические, пневматические, гелиевые и пороховые) частицы с нанесенной на них ДНК разгоняются в вакууме до скоростей, достаточных для проникновения в клетки-мишени клеточных культур и многоклеточных эксплантатов, компетентных к регенерации растений in vitro, которые помещают в вакуумной камере под источником частиц. После такого "обстрела" необходимо выделить на селективных средах трансформированные клетки и получить из них растения-регенеранты.
Известны многочисленные случаи применения способа баллистической трансформации на пшенице (V.Vasil et al. (1992) Bio/Technology 10(6): 667-674), сое (P. Christou et al. (1990) Trends Biotech. 8: 145-151), рисе (P. Christou et al. (1994) Bio/Technology 9: 957-962), кукурузе (T.Klein et al. (1989) Plant Pysiol. 91: 440-444), хлопчатнике (Finer et al. (1990) Plant Cell Reports 8: 586-589) и других видах. Попытки применения баллистического способа для генетической трансформации подсолнечника проводились в лаборатории Gunter Hanner (Reiner Hunold et al. (1995) Plant Science 105: 95-109). Эти работы не увенчались успехом, и авторы не сумели получить трансгенных растений. Подводя итог многолетним исследованиям, другие разработчики в данной области (напр. , Pioneer Hi-Bred International - см. пат. US 6229078) также констатируют, что подсолнечник почти не поддается баллистической трансформации.
Поэтому нами была поставлена задача разработать воспроизводимую технологию баллистической трансформации подсолнечника, которая оказалась бы достаточно эффективной.
Сущность изобретения заключается в том, что в способе получения трансгенных растений подсолнечника, включающем трансформацию клеток ткани подсолнечника введением в них экзогенной ДНК, селекцию трансформированных клеток из ткани и регенерацию из них побегов, были сделаны следующие улучшения, которые позволили придать ткани повышенную способность к трансформации (т.е. увеличить частоту трансформации ее клеток):
1. Использовав метод культивирования, регенерации и вторичной индукции эмбриоидов подсолнечника in vitro, по модифицированной нами методике Freyssnet, мы впервые в мире успешно применили баллистический способ доставки генов в компонентные к регенерации клетки и получению из них трансгенной ткани подсолнечника и разработали новый способ селекции этой ткани в жидкой фазе с последующей регенерацией и клонированном трансгенных растений.
2. Создан и использован оригинальный метод обработки незрелых зародышей пониженными температурами в течение суток (при to +4Сo), повышающий частоту транзиентной экспрессии гетерологичных генов и генетической трансформации.
3. Экспериментально отработаны оптимальные условия доставки генных конструкций в незрелые зародыши методом бомбардмента при помощи баллистической установки PIG (Particle Inflow Gun (Finer et al., Plant Cell Report, 1992, 11: 323-328)).
4. Найден эффективный промотор (RTL2) для регуляции экспрессии селективного гена Hgh, определяющего устойчивость к антибиотику гигромицину, для селекции in vitro трансформированных клеток.
5. Создана оригинальная система селекции трансформированных тканей в жидкой среде на модифицированной нами культуральной среде Chraibi [Chraibi et al., Plant Cell Report (1992), 10: 617-620].
6. Создана система выделения и микроклонирования трансгенной ткани.
7. Укоренение трансгенных растений проводили методом прививок на 7-дневные проростки подсолнечника.
Далее изобретение раскрывается более подробно со ссылками на чертежи.
На фиг.1 показана конструкция плазмиды RTL2.
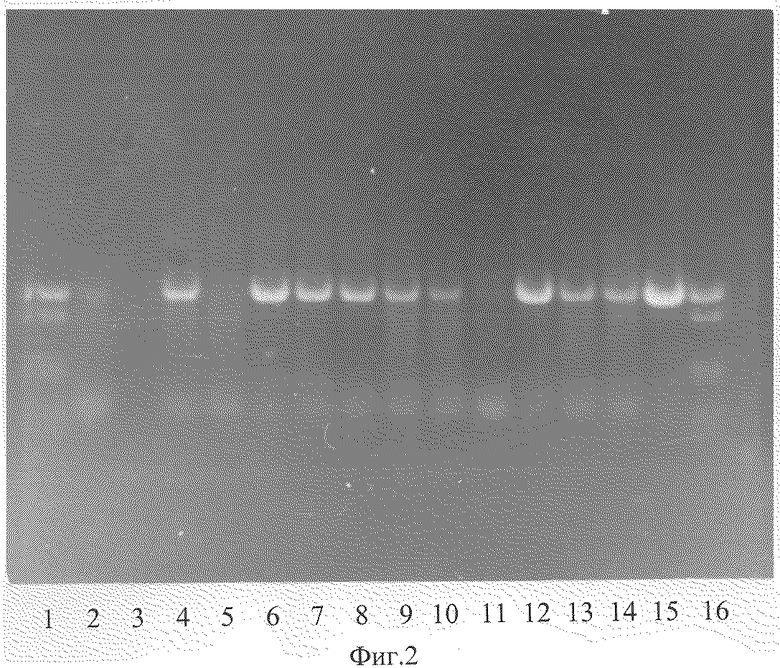
На фиг.2 показан результат ПЦР анализа наличия введенного гена гигромицин фосфотрансферазы (hgh) в ДНК семян трансгенного подсолнечника сорта Передовик улучшенный.
Далее в примерах раскрыт оптимальный вариант изобретения.
Пример 1. Получение культуры незрелых зародышей подсолнечника in vitro
1.1 Получение растительного материала
Донорные растения могут быть выращены в поле или в оранжерее при освещении ртутными и натриевыми лампами, обеспечивающими освещенность не менее 40000 люкс, при фотопериоде 15/9 часов, при температуре 26oС и 70% влажности. Еженедельно растения подкармливают добавочно KNO3 и микроэлементами. В условиях оранжереи еженедельно ведется опрыскивание ядохимикатами для предотвращения размножения белокрылки. Донорные растения опыляются ручным способом щеткой или корзинкой об корзинку.
1.2. Выделение и культивирование незрелых зародышей
Корзинки донорных растений, срезанных днем на 7-12 день после опыления, хранят до следующего утра при температуре +4oС. Семена проверяются на оптимальный размер зародышей разрезанием корзинки на две половины и исследуется размер зародышей в каждом ряду. Цветение начинается с наружного края корзинки к центру и длится около недели, поэтому только один-два ряда имеют размер зародышей 1-1,5 мм, которые используют для трансформации. Семена подходящего размера стерилизуют в 25%-м растворе Clorox (коммерческий отбеливатель), добавляя 3-4 капли Twen-80, затем промывают 3 раза стерильной дистиллированной водой. Все операции выполняют в условиях ламинарного бокса, обеспечивающего асептические условия. Незрелые зародыши размером 1-1,5 мм выделяют, отрезая верхнюю 1/3 семени и осторожно выдавливая зародыш на поверхность чашки Петри тупой стороной скальпеля. Зародыши помещают на модифицированную добавкой 500 мг/л гидролизата казеина среду SFRI для индукции эмбриоидов (Freyssinet et al., патент US 5017491, 1991), содержащую 0,5-1 мг/л ВАП, рН 5,7. Зародыши культивируют в темноте при температуре 27oС. В результате ткань эмбриоидов существенно увеличивает способность к трансформации экзогенной ДНК.
Пример 2. Условия бомбардмента эксплантов
Зародыши размещают в виде кольца на той же среде в чашках Петри, наружный диаметр которого составляет 5 см, внутренний - 1 см. Зародыши раскладывают на среде, помещая одну из семядолей в среду, обеспечивая перпендикулярность поверхности гипокотиля направлению потока частиц с нанесенной на них ДНК (см. ниже пример 3), которыми она будет бомбардироваться.
Для бомбардировки могут быть использованы частицы любого подходящего твердого материала, например вольфрама, золота, платины и др. В приведенном примере осуществления изобретения были использованы микрочастицы вольфрама.
После 2-4 дней прекультивирования на среде SFRI незрелые зародыши подвергали бомбардированию частицами вольфрама размера 0,7-2,5 микрон, используя PIG [Particle Inflow Gun (Finer et al., Plant Cell Reports, 11: 323-328, 1992)]. Перед бомбардированием чашки с зародышами оставляли открытыми на один - два часа для легкого подсыхания зародышей, обеспечивающего плазмолиз в клетках-мишенях, который также обусловлен высоким осмотическим давлением среды, содержащей 9% сахарозы и высокую концентрацию аминокислот. Расстояние от источника частиц составляло 12 см, давление Гелия - 6 атм, вакуум в камере - 0,3 атм. Между источником частиц и эксплантами помещалась предохранительная сетка с размером ячейки 0,5 мм. Каждый зародыш после первого бомбардмента переворачивался на другую сторону и подвергался вторичному бомбардаменту.
Пример 3. Генные конструкции для баллистической трансформации подсолнечника
Баллистическая трансформация может осуществляться одной плазмидой, несущей необходимые для трансформации гены - селективные: ген nptII устойчивости к антибиотику канамицину, или ген hph устойчивости к антибиотику гигромицину, или ген bar устойчивости к гербициду Basta, или ген EPSP synthase устойчивости к гербициду Glyphosate; репортерные: ген β-glucuronidase [uidA или (GUS)] или GFP (green fluorescent protein); и гены, представляющие интерес для улучшения агрономических свойств растения.
Возможно использовать и смесь нескольких плазмид, каждая из которых несет один из указанных видов генов. При использовании смеси двух плазмид в соотношении 1:1 вероятность их коинтеграции составляет от 30 до 75%.
В данном случае использовалась плазмида pRT-TL-Hyg, несущая ген устойчивости к гигромицину hph, в котором кодирующая часть гена находится под управлением промотора RTL2, содержащего двойной промотор 35S из вируса мозаики цветной капусты и TL энхансер из вируса табака. В качестве терминатора транскрипции использован терминатор вируса мозаики цветной капусты.
Пример 4. Технология нанесения ДНК на частицы вольфрама
50 мг частиц стерилизовались в 500 мкл 95% этанола в течение 20 минут. Частицы осаждались центрифугированием, затем ресуспендировались в 500 мкл стерильной воды. Эта операция повторялась трижды. К 25 мкл ресуспендированных в пробирке частиц добавлялись 5 мкл ДHК, смешивались пипеткой. Далее добавлялось 25 мкл проавтоклавированного раствора 2,5 М CaCl2, все смешивалось и к смеси быстро добавлялось 10 мкл 100 mМ спермидина. После смешивания раствор-суспензия и помещалась на 5 минут на лед. После этого 50 мкл супернатанта удалялось, а оставшаяся суспензия частиц с прецепитированной ДНК использовалась в течение 15 минут. На каждый выстрел расходовалось по 2-3 мкл взвеси микрочастиц.
Пример 5. Селекция на среде с гигромицином
Обработанные зародыши культивировали в течение 5 дней в темноте на модифицированной среде SFRI. После этого для селекции трансформированной ткани экспланты переносили в 250-миллилитровые колбы Эрленмейера емкостью 250 мл, содержащие 35 мл среды LBM (Chraibi et al., 1992). Среда содержала 3% сахарозы, 1 мг/л БАП, 1 мг/л НУК и 15 мг/л гигромицина. Колбы помещали на ротационный шейкер, частота колебаний составляла около 137 мин-1. Селекцию проводили при освещении и температуре 27oС. Среду меняли каждые 5-7 дней. Селекцию осуществляли в течение 3-4 недель.
Пример 6. Регенерация трансгенных побегов
После 2-3 недель селекции основная ткань эксплантов некротизировалась, приобретая коричневый и черный цвет. Экспланты переносились в чашки Петри и проверялись на наличие растущей зеленой ткани. При наличии таковой экспланты переносились в жидкую среду без селективного агента на два-три дня, после чего зеленую ткань, растущую в области гипокотиля, вырезали и помещали на среду SFRI и культивировали на свету, меняя среду каждые 7 дней. Ткани с регенерационными зонами переносили на среду SFRII, содержащую 3% сахарозы и БАП в концентрации 0,5-1 мг/л. Для удлинения развивающихся побегов последние переносили на среду с концентрацией БАП менее 0,2 мг/л или на среду, не содержащую БАП. Полученные клоны могут быть размножены при культивировании на среде SFRI и SFRII в течение двух месяцев.
Пример 7. Укоренение на среде для укоренения или методом прививки на побеги подсолнечника
Для получения взрослых фертильных растений использовали среды для укоренения и метод прививания побегов длиной 3-5 мм на 7-дневные проростки подсолнечника в культуре in vitro. Использовали побеги, достигшие 3-5 мм длины и имеющие пазушную почку. У выращенных в асептических условиях проростков подсолнечника удаляли семядоли и апекс побега. Скальпелем в гипокотиле, на расстоянии 5-7 мм от верхней части, проводили сквозной вертикальный разрез, и в образовавшуюся щель вставляли побег. Привитые проростки культивировали в пробирках при температуре 27oС при фотопериоде 16/8 часов. Через две-три недели растения из пробирок переносили в горшки с почвой и обеспечивали создание "влажной камеры". Далее растения пересаживали в большие горшки и выращивали в условиях теплицы. Частота выживания прививок, достигших цветения, составляла до 70%.
Пример 8. Анализ природы полученных растений методом ПЦР
Доказательство встраивания вводимого гена в геном трансгенных растений осуществляли по стандартной методике полимеразой цепной реакции (Mullis К.В. & Faloona F. A. Meth. Enzymol, 155, 335, 1987). На фиг.2 показан результат ПЦР анализ наличия введенного гена гигромицин фосфотрансферазы (hgh) в ДНК семян трансгенного подсолнечника сорта Передовик улучшенный. Дорожки 1 и 16 - молекулярный маркер - фагλ; дорожки 2 и 3 - ДНК нетрансформированного подсолнечника, негативный контроль; дорожка 15 - плазмида с геном hgh; дорожки - с 4 по 14 ДНК из растений - трансформантов.
Пример 9. Частота трансформации
Частота трансформации, полученная при использовании разработанного метода, вычислялась как отношение количества трансгенных побегов, для которых показано встраивание гетерологичного гена в геном методом ПЦР, к количеству обстрелянных чашек с эксплантами или числу использованных для трансформации эксплантов. Данные ряда независимых опытов представлены в таблице.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ПШЕНИЦЫ С ИСПОЛЬЗОВАНИЕМ БИОБАЛЛИСТИКИ | 2016 |
|
RU2646108C1 |
| СПОСОБ СОЗДАНИЯ РАСТЕНИЙ ПШЕНИЦЫ, УСТОЙЧИВОЙ К КЛОПУ-ВРЕДНОЙ ЧЕРЕПАШКЕ (EURYGASTER INTEGRYCEPS PUTON) | 2004 |
|
RU2277586C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ РАСТЕНИЙ САХАРНОЙ СВЕКЛЫ С ИСПОЛЬЗОВАНИЕМ AGROBACTERIUM TUMEFACIENS | 2004 |
|
RU2278162C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ СОРГО | 2002 |
|
RU2229793C1 |
| СПОСОБ УЛУЧШЕННОЙ ТРАНСФОРМАЦИИ С ИСПОЛЬЗОВАНИЕМ АГРОБАКТЕРИИ | 2012 |
|
RU2620975C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОГО РАСТЕНИЯ КУКУРУЗЫ | 1991 |
|
RU2114911C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ С ПОВЫШЕННОЙ УСТОЙЧИВОСТЬЮ К ФИТОПАТОГЕНАМ | 2002 |
|
RU2261275C2 |
| Промотор pro-SmAMP-D1 из растения звездчатка средняя (Stellaria media L.) для биотехнологии растений | 2022 |
|
RU2799014C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ОДНОДОЛЬНЫХ РАСТЕНИЙ | 2007 |
|
RU2351121C1 |
| ТРАНСГЕННОЕ РАСТЕНИЕ БЕРЕЗЫ С ПОВЫШЕННОЙ ПРОДУКТИВНОСТЬЮ | 2013 |
|
RU2593721C2 |
Изобретение относится к селекции растений. Для получения трансгенных растений подсолнечника их трансформируют экзогенной ДНК, вводимой в растительные клетки на частицах твердого носителя. Затем из клеток регенерируют побеги. Экзогенная ДНК может быть представлена, например, генами устойчивости к антибиотикам или гербицидам. Использование промотора RTL2 для экспрессии гена, определяющего устойчивость к гигромицину, и селекции трансформированных клеток в жидкой фазе позволяют получить высокий процент трансформантов подсолнечника. 5 з.п.ф-лы, 2 ил., 1 табл.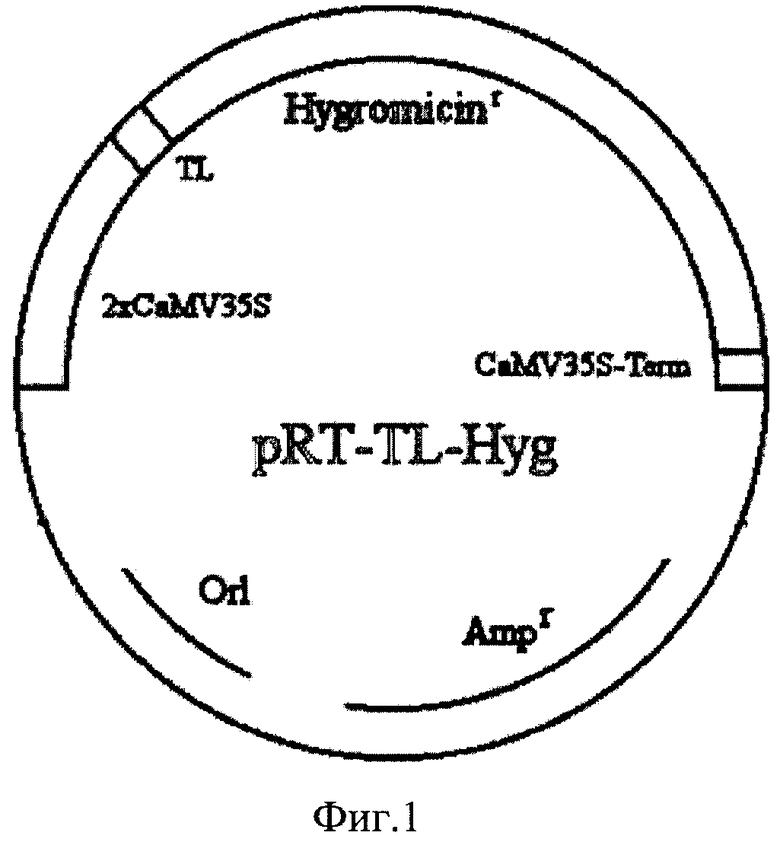
| US 6229078 A, 08.05.2001 | |||
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| US 5017491 A, 21.05.1991. | |||
