Область техники, к которой относится изобретение
Изобретение относится к фармацевтическому использованию веществ, которыми являются агонисты интерлейкина 10 (IL-10). В частности, настоящее изобретение относится к использованию вещества настоящего изобретения для получения фармацевтической композиции, предназначенной для снижения продуцирования TNFα, и/или для профилактики или лечения панкреатита, уратного артрита (подагры), аллергий кожи, аллергических реакций на коже, повреждений ткани в результате гипоксии/ишемии (инфаркта, реперфузии), воспалительных реакций, вызываемых вирусными инфекциями; и/или для изготовления противозачаточного средства.
Известный уровень техники
Фармацевтические композиции, содержащие человеческий IL-10 (hIL-10) или вирусный IL-10 (vIL-10), а также использование hIL-10 или vIL-10 для изготовления фармацевтической композиции для лечения различных состояний были описаны, например, в WO 93/02693 и WO 94/04180, а некоторые агонисты IL-10 были описаны в WO 96/01318.
Краткое описание сущности изобретения
Настоящее изобретение относится к использованию вещества или полипептида формулы
X1-X2-X3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID No: 22)
где X1 представляет Ala или Gly,
Х2 представляет Тyr или Phe,
Х3, X4 и X5 независимо выбирают из группы, состоящей из Met, Ile, Leu, и Val; и
Х6 выбирают из группы, состоящей из Asn, Asp, Gin и Glu; при этом необязательно по крайней мере, один из X1, Х2, Х3, Х4, X5 и Х6 независимо замещен не встречающимися в природе или редкими аминокислотами; и/или пептид является циклизованным; и/или пептид является стабилизированным, и/или амино-концевой аминокислотный остаток является ацилированным, и/или карбокси-концевой аминокислотный остаток является амидированным; а псевдопептиды смоделированы на основе вышеуказанной формулы в целях получения фармацевтической композиции, предназначенной для снижения уровня продуцирования TNFα, и/или для профилактики или лечения панкреатита.
Кроме того, настоящее изобретение относится к веществу или полипептиду формулы
X1-X2-X3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID No: 22)
где X1 представляет Ala или Gly,
Х2 представляет Тyr или Phe,
Х3, X4 и X5 независимо выбирают из группы, состоящей из Met, Ile, Leu, и Val; и
Х6 выбирают из группы, состоящей из Asn, Asp, Gin и Glu;
где, по крайней мере, один из X1, Х2, Х3, Х4, X5 и Х6 независимо замещен не встречающимися в природе или редкими аминокислотами; и/или пептид является циклизованным; и/или пептид является стабилизированным, и/или амино-концевой аминокислотный остаток является ацилированным, и/или карбокси-концевой аминокислотный остаток является амидированным; а псевдопептиды смоделированы на основе вышеуказанной формулы; причем указанное вещество или полипептид обладает, по крайней мере, одним из нижеследующих свойств:
a) индуцирует ингибирование спонтанного продуцирования IL-8 моноцитами человека;
b) индуцирует ингибирование IL-lβ-индуцированного продуцирования IL-8 мононуклеарными клетками периферической крови человека (РВМС);
c) индуцирует продуцирование белка, являющегося антагонистом рецептора интерлейкина-1(IRAP), моноцитами человека;
d) индуцирует хемотаксическую миграцию СD8+-Т-лимфоцитов человека in vitro,
e) десенсибилизирует CD8+-Т-клетки человека, что приводит к их невосприимчивости по отношению к rhIL-10,
f) супрессирует хемотаксический ответ CD4+-T-лимфoцитoв человека по отношению к IL-8;
д) супрессирует хемотаксический ответ моноцитов человека по отношению к MCAF/MCP-1;
h) ингибирует экспрессию молекулы МНС (главного комплекса гистосовместимости) класса II на моноцитах человека, стимулированных IFN-γ;
i) индуцирует продуцирование IL-4 культивированными нормальными CD4+-T-клeткaми человека;
j) снижает продуцирование TNFα в реакции смешанной культуры лейкоцитов человека;
k) осуществляет негативную регуляцию продуцирования TNFα и IL-8 в кроличьей модели острого панкреатита, индуцированного желчной кислотой, а также способствует снижению нейтрофильной инфильтрации в легких обработанных кроликов.
Считается (как подробно изложено в нижеследующем описании иммунологических механизмов), что механизм действия указанного пептида опосредован блокированием действия медиаторов иммунной системы, а в частности цитокинов, таких как монокины, лимфокины, хемокины и антагонисты рецептора монокинов, то есть предполагается, что вещество настоящего изобретения блокирует/супрессирует продуцирование и/или действие некоторых цитокинов и тем самым ингибирует патологические процессы, приводящие к повреждению тканей, и что вещество настоящего изобретения индуцирует продуцирование природных антагонистов рецептора монокинов, что приводит к блокированию/супрессии действия некоторых цитокинов, таких как, TNFα или IL-1 и тем самым к ингибированию патологических процессов, вызывающих повреждение ткани.
Таким образом, в важном варианте своего осуществления, настоящее изобретение относится к фармацевтической композиции, содержащей в качестве активного ингредиента вещество настоящего изобретения.
В другом своем аспекте настоящее изобретение относится к использованию вещества настоящего изобретения для изготовления фармацевтической композиции, предназначенной в основном для ингибирования цитокин-ассоциированного биологического эффекта, т.е. к использованию вещества настоящего изобретения в качестве белка/пептида, лимфокина, монокина, интерлейкина, интерферона, хемокина или колониестимулирующего фактора, являющихся антагонистами рецептора IL-1. В другом своем аспекте настоящее изобретение относится к использованию вещества настоящего изобретения для изготовления фармацевтической композиции, предназначенной для профилактики или лечения состояния, связанного с нарушением системы цитокинов, т.е. системы белков/пептидов, лимфокинов, монокинов, интерлейкинов, интерферонов, хемокинов или колониестимулирующих факторов, которые являются антагонистами IL-1. В другом своем аспекте настоящее изобретение также относится к способу лечения состояний у человека, связанных с нарушением в системе цитокинов, где указанный способ предусматривает введение индивидууму эффективного количества вещества настоящего изобретения.
Клеточная иммунная система принимает участие в развитии таких нарушений, как инфекции, воспаления и опухолевые заболевания. Иммунокомпетентные клетки и их продукты могут играть важную роль в инициации, прогрессировании и, возможно, в природе развития хронических воспалительных состояний. Эти нарушения имеют часто неясную этиологию и представляют собой широко распространенные заболевания, такие как сахарный диабет, ревматоидный артрит, воспалительные заболевания желудочно-кишечного тракта и кожи. Однако помимо указанных примеров клеточно-опосредованный иммунитет или провоспалительные медиаторы вносят свой вклад во многие другие воспалительные и пролиферативные заболевания (см. табл.1).
Действие IL-10 на продуцирование цитокинов
hIL-10 ингибирует продуцирование ряда цитокинов, включая интерферон-γ (IFM-γ), фактор некроза опухоли-α. (TNF-α), колониестимулирующий фактор гранулоцитов и макрофагов (GM-CSF), колониестимулирующий фактор гранулоцитов (G-CSF), IL-lα, IL-1β, IL-2, IL-6, IL-8 и моноцитарный хемотаксический полипептид-1 (MCP-1/MCAF), моноцитами/макрофагами и/или Т-лимфоцитами [4, 5]. IL-10 также ингибирует способность моноцитов мигрировать в ответ на хемокин MCP-1/MCAF [75] . Кроме того, hIL-10 индуцирует продуцирование эндогенного природного антагониста рецептора интерлейкина-1 (IRAP) [6], который ингибирует IL-lα и IL-lβ, путем конкурирования за связывание с рецептором. Поскольку IL-8 является в высокой степени IL-lα, IL-lβ, индуцибельным, то часть действия IL-10, ингибирующего продуцирование IL-8, обусловлено стимуляцией продуцирования антагониста рецептора IL-1, IRAP. Этот механизм имеет большое значение для настоящего изобретения, описанного и проиллюстрированого ниже. IRAP обладает противовоспалительной активностью [9], и предполагается что он оказывает терапевтическое действие при ревматоидном артрите [10]. В своих исследованиях Fischer et а1. наблюдали успешное выживание через 28 дней после обработки IRAP (р=0,015) [11], что свидетельствовало о том, что IRAP оказывает эффективное и дозозависимое действие при лечении септического синдрома. Часть противовоспалительного действия IRAP может быть обусловлена ингибированием продуцирования хемокинов, таких как IL-8.
Экспрессия IL-10 и антигена
IL-10 ингибирует экспрессию МНС класса II на моноцитах человека [8]. hIL-10 ингибирует конститутивно индуцированную и IL-4- или IFN-γ-индуцированную экспрессию HLA-DR/DP и DQ [12] . Кроме того, моноциты, предварительно инкубированные с IL-10, не реагируют на последующее индуцирование экспрессии МНС класса II цитокинами IL-4 или IFM-γ. IL-10 ингибирует экспрессию МНС класса II моноцитами человека после LPS-активации [12, 76]. Введение мышам BALB/c 1-10 мг IL-10 одновременно с летальной дозой LPS обеспечивало защиту этих мышей от гибели [6].
IL-10 ингибирует азотсодержащие промежуточные соединения и супероксид-анионы. IL-10 также ингибирует реакционноспособное азот-содержащее промежуточное соединение (NO), а также реакционноспособные кислородсодержащие промежуточные соединения (H2O2), продуцируемые макрофагами после IFNγ-активации [13].
IL-10 и Т-клеточная активность
IL-10 обладает также модулирующим действием на Т-клеточные функции/активность. Так, например, hIL-10 представляет собой хемотаксический фактор для СD8+-Т-лимфоцитов, тогда как по отношению к CD4+-T-клeткaм он не обнаруживает хемотаксического действия [14] . Кроме того, IL-10 подавляет способность CD4+-T-клeтoк реагировать на хемотаксические сигналы β-хемокина RANTES, а также α-хемокина IL-8. hIL-10 также непосредственно ингибирует пролиферацию Т-клеток периферической крови человека и клоны CD4+-T-клeтoк [14].
Обсуждение терапевтических свойств
Эти in vivo-результаты/данные и другие данные, систематизированные, например, в WO 96/01318, дают серьезные основания предположить о гомеостатической роли IL-10 в регуляции клеточно опосредованного и амплифицированного монокинами иммуновоспалительного ответа и указывают на возможность широкого терапевтического применения IL-10 или лекарственного средства с IL-10-подобной активностью для лечения заболеваний, обусловленных повышенным/недостаточным продуцированием и/или активностью IL-10.
В табл. 1 и 2 перечислены некоторые заболевания, где иммуномодулятор, аналогичный IL-10, или иммуномодулятор с IL-10-подобной активностью имеет, как предполагается, важное терапевтическое значение:
Подробное описание изобретения
Получение IL-10-гомологичного нонапептида с IL-10-подобной активностью
Неполные последовательности hIL-10, имеющие длину 9 аминокислот, были выбраны исходя из тех соображений, что последовательности vIL-10 и hIL-10 должны обладать высокой гомологией, но, по возможности, низкой гомологией по отношению к mlL-10. Было обнаружено, что синтетический нонапептид IT9302 обладает некоторой иммуносупрессорной активностью, которая имитирует активность hIL-10, как будет более подробно описано в нижеследующих примерах. IT9302 соответствует нонапептидной последовательности от С-конца hIL-10, имеющего следующую аминокислотную последовательность
NH2-Ala-Tyr-Met-Thr-Met-Lys-Ile-Arg-Asn-COOH (SEQ ID No: 1)
Нонапептид IT9302 обладает сильным действием, ингибирующим различные функции, и является очень стабильным, и, кроме того, предполагается, что он не способен неспецифически связываться с рецепторами. Нонапептид был выбран потому, что в основном 9-аминокислотная полипептидная последовательность является уникальной для белка. Однако следует отметить, что наиболее важными являются, очевидно, 6 аминокислот у самого конца hIL-10. Таким образом, в объем настоящего изобретения входит вещество или полипептид, содержащий подпоследовательность аминокислотной последовательности Ala-Tyr-Met-Thr-Met-Lys-Ile-Arg-Asn (SEQ ID No: l).
Предполагается также, что некоторые аминокислотные замены не будут оказывать неблагоприятного действия на агонистическую активность hIL-10, как показано в настоящем описании, при условии, что будут присутствовать треонин, лизин и аргинин и что между ними будет находиться одна аминокислота.
Настоящее изобретение, в частности, относится к использованию вещества или полипептида формулы
X1-X2-X3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID No: 22)
где X1 представляет Ala или Gly,
X2 представляет Тyr или Phe,
Х3, Х4 и X5 независимо выбирают из группы, состоящей из Met, Ilе, Leu, и Val; и
Х6 выбирают из группы, состоящей из Asn, Asp, Gin и Glu;
при этом необязательно по крайней мере, один из X1, X2, Х3, Х4, X5 и Х6 независимо замещен не встречающимися в природе или редкими аминокислотами; и/или пептид является циклизованным; и/или пептид является стабилизированным; и/или амино-концевой аминокислотный остаток является ацилированным; и/или карбокси-концевой аминокислотный остаток является амидированным; а псевдопептиды смоделированы на основании вышеуказанной формулы в целях получения фармацевтической композиции, предназначенной для снижения IL-1-индуцированного продуцирования TNFα, вероятно, посредством IRAP-блокирования рецептора IL-1, и/или для профилактики или лечения панкреатита.
Примерами конкретных полипептидов, которые, как предполагается, могут быть использованы для снижения уровня продуцирования TNFα и/или для профилактики или лечения панкреатита, являются следующие полипептиды: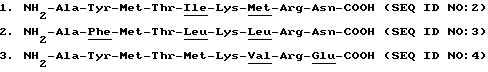
4. NH2-Gly-Tyr-Met-Thr-Met-Lys-Ile-Arg-Asp-COOH (SEQ ID NO:5)
5. NH2-Ala-Phe-Met-Thr-Met-Lys-Ile-Arg-Asp-COOH (SEQ ID NO:6)
6. NH2-Ala-Tyr-Ile-Thr-Met-Lys-Ile-Arg-Asp-COOH (SEQ ID NO:7)
7. NH2-Ala-Tyr-Leu-Thr-Met-Lys-Ile-Arg-Asp-COOH (SEQ ID NO:8)
8. NH2-Ala-Tyr-Val-Thr-Met-Lys-Ile-Arg-Asp-COOH (SEQ ID NO:9)
9. NH2-Ala-Tyr-Met-Thr-Ile-Lys-Ile-Arg-Asp-COOH (SEQ ID NO:10)
10. NH2-Ala-Tyr-Met-Thr-Leu-Lys-Ile-Arg-Asp-COOH (SEQ ID NO: 11)
11. NH2-Ala-Tyr-Met-Thr-Val-Lys-Ile-Arg-Asp-COOH (SEQ ID NO: 12)
12. NH2-Ala-Tyr-Met-Thr-Met-Lys-Ile-Arg-Asp-COOH (SEQ ID NO: 13)
13. NH2-Ala-Tyr-Met-Thr-Met-Lys-Met-Arg-Asp-COOH (SEQ ID NO: 14)
14. NH2-Ala-Tyr-Met-Thr-Met-Lys-Val-Arg-Asp-COOH (SEQ ID NO: 15)
15. NH2-Ala-Tyr-Met-Thr-Met-Lys-Ile-Arg-Gln-COOH (SEQ ID NO: 16)
16. NH2-Ala-Tyr-Met-Thr-Met-Lys-Ile-Arg-Glu-COOH (SEQ ID NO: 17)
Настоящее изобретение, в частности, относится к полипептиду, имеющему формулу
Thr-X4-Lys-X5-Arg-X6 (SEQ ID No: 19)
полипептиду, имеющему формулу
X3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID No: 20)
полипептиду, имеющему формулу
X2-X3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID No: 21)
и к полипептиду, имеющему формулу
X1-X2-X3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID No: 22),
где X1 представляет Ala или Gly,
Х2 представляет Тyr или Phe,
Х3, Х4 и X5 независимо выбирают из группы, состоящей из Met, Ile, Leu, и Val; и
Х6 выбирают из группы, состоящей из Asn, Asp, Gin и Glu;
где одна или несколько аминокислот замещены не встречающимися в природе или редкими аминокислотами; и/или пептид является циклизованным; и/или амино-концевой аминокислотный остаток является ацилированным, и/или карбокси-концевой аминокислотный остаток является амидированным; а псевдопептиды смоделированы исходя из вышеуказанной формулы; причем указанные аналоги обладают, по крайней мере, одним из нижеследующих свойств:
a) индуцируют ингибирование спонтанного продуцирования IL-8 моноцитами человека;
b) индуцируют ингибирование IL-lβ-индуцированного продуцирования IL-8 мононуклеарными клетками периферической крови человека (РВМС);
c) индуцируют продуцирование белка, являющегося антагонистом рецептора интерлейкина-1(IRAP), моноцитами человека;
d) индуцируют хемотаксическую миграцию СD8+-Т-лимфоцитов человека in vitro,
е) десенсибилизируют CD8+-T-клетки человека, что приводит к их невосприимчивости по отношению к rhIL-10;
f) супрессируют хемотаксический ответ CD4+-T-лимфoцитoв человека по отношению IL-8;
g) супрессируют хемотаксический ответ моноцитов человека по отношению MCAF/MCP-1;
h) ингибируют экспрессию молекулы МНС (главного комплекса гистосовместимости) класса II на моноцитах человека, стимулированных IFN-γ;
i) индуцируют продуцирование IL-4 культивированными нормальными СD4+-Т-клетками,
j) снижают продуцирование TNFα в реакции смешанной культуры лейкоцитов человека;
k) осуществляют негативную регуляцию продуцирования IL-8 и TNFα в кроличьей модели острого панкреатита, индуцированного желчной кислотой, а также способствуют снижению уровня нейтрофильной инфильтрации в легких обработанных кроликов.
Используемый в настоящем описании термин "по крайней мере, одна биологическая активность IT9302" означает, что IT9302 имеет, по крайней мере, одно из вышеуказанный свойств.
Любой из пептидов, рассматриваемых в настоящем изобретении, может иметь амино-концевой аминокислотный остаток, который является ацилированным, такой как ацетилированный или бензоилированный аминокислотный остаток. Любой из рассматриваемых пептидов может также иметь карбокси-концевой аминокислотный остаток, который является амидированным.
Кроме того, в настоящем изобретении рассматриваются аналоги пептидов, образованные другими консервативными аминокислотными заменами, отличающимися от тех, которые были указаны выше; заменами редкими или не встречающимися в природе аминокислотами; стабилизацией пептидов; циклизацией пептидов; и псевдопептидами, смоделированными на основе идентифицированных пептидов, являющихся агонистами IL-10.
Принцип "консервативных аминокислотных замен" основан на том, что некоторые пары аминокислот имеют совместимые боковые цепи, такие, что при замене одной аминокислоты другой аминокислотой третичная структура белка и его аффинность связывания имеют лишь минимальные изменения. Правила, которым следует руководствоваться при осуществлении консервативных замен, описаны в [78].
Используемый в настоящем описании термин "консервативный" означает, что (i) изменения являются, по возможности, конформационно нейтральными, то есть влекут за собой минимальные изменения третичной структуры мутантных полипептидов по сравнению с нативным белком: и (ii) изменения являются, по возможности, антигенно нейтральными, то есть влекут за собой минимальные изменения в антигенных детерминантах мутантных полипептидов по сравнению с нативным белком. Конформационная нейтральность желательна для сохранения биологической активности, а антигенная нейтральность желательна для предупреждения стимуляции иммуногенного ответа у пациентов или животных, которым были введены вещества настоящего изобретения. Хотя достаточно трудно с абсолютной уверенностью выбрать, который из альтернативных вариантов будет конформационно и антигенно нейтральным, однако существуют правила, которыми каждый специалист может руководствоваться для осуществления замен, которые с высокой степенью вероятности можно считать конформационно и антигенно нейтральными, см., например, [77 и 78]. Некоторые из наиболее важных правил основаны на том, что: (1) замена гидрофобных остатков с меньшей долей вероятности будет приводить к изменению антигенности, поскольку они, очевидно, расположены внутри белка, см., например, Berzovsky (цитированный выше) и Bowie et al. (цитированные выше); (2) замена подобных по своим физико-химическим свойствам, то есть синонимичных остатков с меньшей долей вероятности будет приводить к конформационным изменениям, поскольку заменяющая кислота может играть ту же самую структурную роль, что и заменяемая кислота; и (3) замена эволюционно консервативных последовательностей будет, очевидно, приводить к неблагоприятным конформационным эффектам, поскольку предполагается, что эволюционная консервативность последовательностей может играть важную функциональную роль. Помимо этих основных правил, которыми следует руководствоваться при выборе последовательностей мутеина, имеются анализы, способные подтвердить биологическую активность и конформацию сконструированных молекул. Изменения в конформации могут быть протестированы, по крайней мере, с использованием двух хорошо известных анализов: метод фиксации микрокомплемента, например, [79 и 80], широко используемый в эволюционных исследованиях третичных структур белков; и анализ на аффинность для установления конформационно-специфических моноклональных антител, например, [81]. Более полно биологические анализы для веществ настоящего изобретения описаны в нижеследующих примерах.
Ингибирование спонтанного продуцирования IL-8 моноцитами человека тестировали, как показано в Примере 1, с использованием синтезированного вещества или пептида вместо IT9302. Если продуцирование IL-8 супрессируется не более чем на 50% при использовании 1 нг/мл вещества или пептида, то это вещество или пептид входит в объем настоящего изобретения.
Ингибирование IL-lβ-индуцированнго продуцирования IL-8 мононуклеарными клетками периферической крови человека (РВМС) тестировали, как показано в Примере 2, с использованием синтезированного вещества или пептида вместо IT9302. Если процент ингибирования продуцирования IL-8 составляет, по крайней мере, 50% при использовании 1 нг/мл вещества или пептида, то это вещество или пептид входит в объем настоящего изобретения.
Продуцирование белка, обладающего антагонистической активностью по отношению к рецептору интерлейкина-1 (IRAP), моноцитами человека, тестировали, как показано в Примере 3, с использованием синтезированного вещества или пептида вместо IT9302. Если индуцирование IRAP составляет, по крайней мере, 30 нг/мл при использовании 10 нг/мл вещества или пептида, то это вещество или пептид входит в объем настоящего изобретения.
Индуцирование хемотаксической миграции СD8+-Т-лимфоцитов человека in vitro тестировали, как показано в Примере 4, с использованием синтезированного вещества или пептида вместо IT9302. Если при используемой концентрации 10 нг/мл данное вещество или пептид является эффективным, то есть дает значение 2 или более, то это вещество или пептид входит в объем настоящего изобретения.
Десенсибилизацию СD8+-Т-клеток человека, приводящую к их невосприимчивости в отношении к rhIL-10, тестировали, как показано в Примере 5, с использованием синтезированного вещества или пептида вместо IT9302. Если предварительное инкубирование клеток с данным веществом или пептидом приводит, в основном, к полной супрессии восприимчивости СD8+-клеток по отношению к rhIL-10, то есть дает значение около 1, такое как 0,8-1,2, при концентрации вещества или пептида 10 нг/мл, то это вещество или пептид входит в объем настоящего изобретения.
Супрессию хемотаксического ответа CD4+-T-лимфoцитoв человека по отношению к IL-8 тестировали, как показано в Примере 6, с использованием синтезированного вещества или пептида вместо IT9302. Если добавление вещества или пептида к суспензии СD4+-Т-лимфоцитов человека приводит, в основном, к полному ингибированию ответа СD4+-клеток по отношению к IL-8, то есть дает величину около 1, такую как 0,8-1,2 при концентрации вещества или пептида 10 нг/мл, то это вещество или пептид входит в объем настоящего изобретения.
Супрессию хемотаксического ответа моноцитов человека по отношению к MCAF/MCP-1 тестировали, как показано в Примере 1, с использованием синтезированного вещества или пептида вместо IT9302. Если добавление вещества или пептида к суспензии моноцитов человека приводит, в основном, к полному ингибированию хемотаксического ответа моноцитов по отношению к MCAF/MCP-1, то есть дает величину около 1, такую как 0,8-1,2 при концентрации вещества или пептида 10 нг/мл, то это вещество или пептид входит в объем настоящего изобретения.
Ингибирование экспрессии молекул МНС класса II на IFNγ-стимулированных моноцитах человека тестировали, как показано в Примере 8, с использованием вещества или пептида вместо IT9302. IFNγ осуществляет позитивную регуляцию экспресии антигена МНС класса II в клеточной популяции на уровне от 36,8 до 58,4%, и эта стимуляция блокирует или осуществляет негативную регуляцию до уровня 25,2% при концентрации 10 нг/мл rhIL-10 и до уровня 31,2% при концентрации 1 нг/мл IT9302. Если данное вещество или пептид блокирует или осуществляет негативную регуляцию экспрессии МНС класса II на моноцитах до нестимулируемого уровня при добавлении этого вещества в количестве 1-10 нг/мл, то указанное вещество или пептид входит в объем настоящего изобретения.
Если добавление данного вещества или пептида приводит к блокированию эффекта стимуляции IFNγ при концентрации этого вещества или пептида 10 нг/мл, то указанное вещество или пептид входит в объем настоящего изобретения.
Индуцирование продуцирования IL-4 культивированными нормальными CD4+-T-клeткaми человека тестировали, как показано в Примере 9, с использованием синтезированного вещества или пептида вместо IT9302. Если добавление данного вещества или пептида индуцирует продуцирование IL-4 в CD4+-T-лимфoцитax при концентрации вещества или пептида 10 нг/мл, то это вещество или пептид входит в объем настоящего изобретения.
Снижение продуцировния TNFα в реакции смешанной культуры лейкоцитов тестировали, как описано в Примере 10, с использованием синтезированного вещества или пептида вместо IT9302. Если добавление данного вещества или пептида приводит к значительному снижению продуцировния TNFα в реакции смешанной культуры лейкоцитов в течение 24 часов при концентрации вещества или пептида 10 нг/мл, то это вещество или пептид входит в объем настоящего изобретения.
Негативную регуляцию продуцирования TNFα и IL-8 в кроличьей модели острого панкреатита, индуцированного желчной кислотой, и снижение уровня нейтрофильной инфильтрации в легких обработанных кроликов тестировали, как описано в Примере 14, с использованием синтезированного вещества или пептида вместо IT9302. Если добавление данного вещества или пептида при концентрации 100 мкг/кг приводит к значительному снижению смертности испытуемых животных, то это вещество или пептид входит в объем настоящего изобретения.
Таким образом, в одном из важных вариантов своего осуществления настоящее изобретение относится к полипептиду, в котором, по крайней мере, один аминокислотный остаток заменен другим аминокислотным остатком, и/или в котором, по крайней мере, один аминокислотный остаток был делегирован или добавлен так, чтобы полученный в результате этого полипептид содержал аминокислотную последовательность или подпоследовательность, включающую указанную аминокислотную последовательность, определенную ниже, и при этом чтобы он в основном обладал hIL-10-агонистической активностью, определенной выше.
Аналоги синтетических пептидов могут быть также получены путем замены отдельных остатков синтетическими или редкими аминокислотами. Последовательности биологически активных пептидов происходят от белков, которые состоят из двадцати природных L-аминокислотных остатков. Однако способ химического синтеза, используемый для конструирования синтетических пептидов, предусматривает замену альтернативными остатками, включая D-аминокислоты, β-аминокислоты, N-замещенные аминокислоты, редко встречающиеся природные аминокислоты, или не встречающиеся в природе синтетические аналоги аминокислот [93] . Неограничивающими примерами аминокислот, которые могут быть использованы в настоящем изобретении, являются следующие аминокислоты:
Aad 2-Аминоадипиновая кислота
bAad 3-Аминоадипиновая кислота
bAla бета-Аланин, бета-аминопропионовая кислота
Abu 2-Аминомасляная кислота
4Abu 4-Аминомасляная кислота, пиперидиновая кислота
Аср 6-Аминокапроновая кислота
Ahe 2-Аминогептановая кислота
Aib 2-Аминоизомасляная кислота
bAib 2-Аминоизомасляная кислота
Аpm 2-Аминопимелиновая кислота
Dbu 2,4-Диаминомасляная кислота
Des Десмозин
Dpm 2,2'-Диаминопимелиновая кислота
Dpr 2,3-Диаминопропионовая кислота
EtGly N-Этилглицин
EtAsn N-Этиласпарагин
Hyl Гидроксилизин
aHyl алло-Гидроксилизин
3Нyр 3-Гидроксипролин
4Hyp 4-Гидроксипролин
Ide Изодесмозин
alle алло-Изолейцин
MeGly N-Метилглицин, саркозин
Melle N-Метилизолейцин
MeLys 6-N-Mетиллизин
MeVal N-Метилвалин
Nva Норвалин
Nle Норлейцин
Orn Орнитин
Другие и неограничивающие примеры редко встречающихся и не встречающихся в природе аминокислот или сконструированных блоков перечислены в следующих работах: Novabiochem 1994/95 Catalog (Calbiochem-Novabiochem AG, Weidenmattweg 4, CH-4448  /Switzerland), pp. 65-125; Bachem Feinkemikalien AG 1995 Catalog (Bachem Feinkemikalien AG, Haupstrasse 144, CH-4416 Budendorf/Switzerland), pp. 753-831; Neosystem Laboratoire Catalogue 1997/98 (Neosystem Laboratoire, 7 rue de Boulogne, 67100 Strasbourg, France) pp. 131-176.
/Switzerland), pp. 65-125; Bachem Feinkemikalien AG 1995 Catalog (Bachem Feinkemikalien AG, Haupstrasse 144, CH-4416 Budendorf/Switzerland), pp. 753-831; Neosystem Laboratoire Catalogue 1997/98 (Neosystem Laboratoire, 7 rue de Boulogne, 67100 Strasbourg, France) pp. 131-176.
Вышеописанные альтернативные остатки могут быть использованы (а) для замены химически реакционноспособных остатков и повышения стабильности синтетического пептида в отношении, например, ферментативной и протеолитической деградации; (b) для включения аналитической метки, используемой при детекции синтетического пептида; и (с) для модуляции биологической активности синтетического пептида путем повышения или снижения аффиности связывания пептида с рецептором IL-10, например, путем введения конформационных затруднений, которые ограничивают свободу вращения для специфических химических связей.
В объем настоящего изобретения также входят вещества, в которых один или несколько остатков Thr, Lys или Arg в вышеуказанной формуле заменены не встречающимися в природе или редкими аминокислотами, перечисленными выше. Если это не оговорено особо, то все полипептидные последовательности, упомянутые в настоящем описании и в формуле изобретения, считываются от N-конца к С-концу в стандартном формате.
Для синтеза пептидов и их аналогов был выбран метод твердофазного пептидного синтеза (ТФПС). Этот метод был предложен Merrifield (100) в 1963 г., и с использованием этой техники было синтезировано множество пептидов. Прекрасный обзор современных методов химического синтеза пептидов и белков представлен в работе S.B.H. Kent [101].
На практике синтез пептидов проводят постадийно методом ТФПС. С-концевую аминокислоту в N-альфа-защищенной форме (если необходимо получить защищенное по боковой цепи реакционноспособное производное) ковалентно связывают либо непосредственно, либо посредством подходящего линкера с твердым носителем, например полимерной смолой, которая набухает в органическом растворителе. Затем N-альфа-защитную группу удаляют и постадийно присоединяют последующие защищенные аминокислоты в соответствии с нужной последовательностью.
После синтеза полной пептидной цепи защитные группы для боковой цепи удаляют и пептид отщепляют от смолы, что может быть сделано одновременно или отдельными стадиями.
Из нескольких различных стратегий синтеза, разрабатывавшихся в течение многих лет, в настоящее время в основном применяются два способа, которые основаны на использовании различных N-альфа-защитных групп и соответствующих защитных групп для боковых цепей. Merrifield использовал в качестве N-альфа-защитной группы трет-бутилоксикарбонил (Воc), a Caprino и Han [102] вводили 9-флуоренилметоксикарбонил (Fmoc). Практическое применение этих двух методик, включая выбор твердых носителей, защитных групп для боковых цепей, процедуры активации, процедуры отщепления, оборудование, а также аналитическую технику и мониторинг, описано в нескольких монографиях, из которых могут быть упомянуты следующие: Stewart & Young [103], Atherton & Sheppard [104], и Pennington & Dunn [105]. Пептиды настоящего изобретения и их аналоги с редкими или не встречающимися в природе аминокислотами были в основном синтезированы в соответствии с вышеуказанными схемами синтеза.
Аналоги синтетических линейных пептидов могут быть получены путем химической трансформации структур в циклическую форму. Циклизация линейных пептидов может способствовать модуляции биологической активности путем усиления или снижения аффинности связывания пептида с белком-мишенью [94]. Линейные пептиды очень пластичны и способны принимать множество различных конформаций в растворе. Циклизация приводит к ограничению ряда имеющихся конформаций и тем самым благоприятствует более активным или неактивным структурам пептида. Иммуногенность синтетических пептидов коррелирует с экспериментально наблюдаемыми конформационными предпочтениями в растворе [95]. Различия в иммуногенности могут указывать на различия в аффинности связывания специфических антител для циклических пептидов.
Циклизацию линейных пептидов осуществляют либо путем образования пептидной связи между свободными N- и С-концами (гомодетные циклопептиды) или путем образования новой ковалентной связи между аминокислотным остовом и/или боковыми группами (гетеродетные циклопептиды) [93]. Последний способ циклизации предусматривает использование альтернативных химических стратегий для образования ковалентных связей, например дисульфидов, лактонов (оба они также присутствуют в природных пептидах), эфиров или тиоэфиров. Линейные пептиды, состоящие из более чем пяти остатков, могут быть циклизованы относительно легко. Способность пептида к образованию бета-конформации в центральных четырех остатках облегчает образование как гомо-, так и гетеродетных циклопептидов. Присутствие пролиновых или глициновых остатков у N- или С-концов также облегчает образование циклопептидов, особенно из линейных пептидов, которые имеют длину менее чем 6 остатков.
Примеры способов образования дисульфидных связей и проведения других реакций циклизации пептидов приводятся в работе Pennington & Dunn [105], в главе 7 и 11.
Технология конструирования псевдопептидов была разработана для получения молекулярных имитаторов пептидов. Возможность успешного конструирования таких молекул зависит от знания свойств линейной пептидной последовательности и конформации, в которой она презентируется рецептору IL-10. Благодаря синтезу имитаторов можно получить вещества, обладающие лучшей биологической активностью, повышенной растворимостью и стабильностью [96].
В качестве примера могут служить нижеследующие псевдопептиды, которые были получены на основе α-спиральных матриц в имитаторах С-концевого пептида цитокинов, описанных в патенте США 5446128 [97], с учетом того известного факта, что С-конец IL-10 существует в форме x-спирали [98].
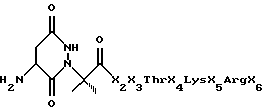
При присоединении этой небольшой молекулы у N-конца пептида α-спиральная структура синтетического пептида стабилизируется, и этот пептид становится более устойчивым к протеолитическому расщеплению. Другие псевдопептиды могут быть получены исходя из описания, приведенного в патенте США 5445128. Такие вещества, где были сделаны замены в других остатках, а не в X1 и/или были сделаны замены молекулами, отличающимися от N-концевой молекулы, показанной в вышеуказанной формуле, входят в объем настоящего изобретения.
В соответствии с настоящим изобретением, термин "аналог пептида" означает любое фармацевтически активное и фармацевтически приемлемое вещество, полученное исходя из вышеуказанной
формулы и обладающее, по крайней мере, одной биологической активностью, аналогичной активности IT9302, включая производные таких аналогов, а в частности их фармацевтически приемлемые "соли, сложные эфиры и сольваты.
Используемые в настоящем описании термины: "цитокин", "лимфокин", "интерлейкин", "монокин", "хемокин", "интерферон", "колониестимулирующий фактор" и "полипептид" определены в WO 96/01318.
В представляющем интерес варианте настоящее изобретение относится к полипептиду, в котором полное число аминокислот составляет от 6, 7, 8, 9 или 10 до 100, например, 11, 12, 13, 14, или 15 аминокислот, или даже более, например 20 или 30 аминокислот.
В предпочтительном варианте настоящего изобретения вещество или полипептид используется в основном в чистом виде. Для получения такого полипептида может потребоваться его очистка. Примерами способов, которые могут быть использованы для очистки полипептидов, являются: (i) иммунопреципитация или аффинная хроматография с использованием антител; (ii) аффинная хроматография с использованием подходящего лиганда; (iii) другие хроматографические методы, такие как гель-фильтрация, ионообменная или высокоразрешающая жидкостная хроматография или модификация любого из этих методов; (iv) электрофоретические методы, такие как, электрофорез на полиакриламидном геле, электрофорез на агарозном геле и изоэлектрическое фокусирование; (v) любые другие методы специфической солюбилизации и/или очистки.
В объем настоящего изобретения также входит фармацевтическая композиция, содержащая вещество или полипептид настоящего изобретения и фармацевтически приемлемый носитель. Эта композиция может содержать, например, очищенный синтезированный белок или очищенный рекомбинантный полипептид.
Агонист IL-10, используемый в настоящем изобретении, может быть получен в виде препарата в фармацевтически приемлемой среде, например в физиологическом растворе, забуференном фосфатом физиологическом растворе (PBS) растворе Рингера, декстрозе/физиологическом растворе, растворе Хэнкса и глюкозе. Эти композиции могут содержать фармацевтически приемлемые добавки, необходимые для создания условий, по возможности приближающихся к физиологическим, такие как забуферивающие агенты; агенты, придающие раствору тоничность; смачивающие агенты; детергенты и т. п. Такими добавками могут быть также дополнительные активные ингредиенты, например бактериальные агенты или стабилизаторы. Количество вещества, вводимого пациенту, может варьироваться в зависимости от конкретного вещества; цели его введения, такой как профилактика или лечение; от состояния пациента; способа введения и т.п.
Эти фармацевтические композиции обычно предназначены для чрескожного или парентерального введения, например внутривенного, подкожного или внутримышечного введения. Формы для перорального введения также являются желательными и могут быть получены путем модификации композиции для шунтирования окружающей среды желудка. Такая композиция может быть использована для профилактики и/или терапевтического лечения.
Фармацевтическими композициями настоящего изобретения, подходящими для местного применения, могут быть, например, кремы, мази, лосьоны, жидкие мази (линименты), гели, растворы, суспензии, пасты, косметические карандаши, аэрозоли или порошки. Такая композиция может быть использована для пропитки или нанесения, например, на подушечки, пластыри или полоски, и, таким образом, она может применяться 1-10 раз в день.
Композиции для местного применения, в основном, содержат 1-80% активного вещества по полной массе препарата, например, 0,001-25 мас.%, а в частности 0,1-10%, 0,5-5% или 2-5% активного вещества. Эта композиция может быть получена в соответствии со стандартной фармацевтической практикой с использованием фармацевтических наполнителей, обычно используемых для приготовления препаратов местного применения. Помимо воды в композициях могут быть использованы другие носители, которые могут быть твердыми или жидкими, такие как умягчители, растворители, влагоудерживающие вещества, загустители и порошки.
рН композиций может, в принципе, варьироваться в широких пределах, например от 3 до 9, хотя предпочтительно, чтобы рН составлял от около 4 до 8. Для получения нужного рН могут быть использованы стандартные забуферивающие агенты.
Композиции для чрескожного введения могут, например, содержать 1 мг вещества (IT9302), растворенного в 1 г основы для крема, такой как нейтральный крем Moistion с 0,05% салициловой кислотой (фармация Arhus Kommunehospital) и могут быть нанесены на кожу в количестве 0,4-0,5 мг в виде пластыря. Эта композиция использована в Примерах 16 и 17.
Альтернативно, фармацевтические композиции могут быть введены внутривенно. Так, например, настоящее изобретение относится к композициям, которые включают вещество, являющееся агонистом IL-10, растворенное или суспендированное в приемлемом носителе, предпочтительно в водном носителе. Эти композиции могут быть стерилизованы с использованием стандартной техники стерилизации либо они могут быть отфильтрованы в стерильных условиях. Полученные водные растворы могут быть упакованы в таком виде, в каком они были получены, либо в виде лиофилизованного препарата, который перед его введением разбавляют стерильным водным носителем. Агонист IL-10 может быть также введен вместе со вторым биологически активным агентом, таким как стандартный хемотерапевтический агент. Такими агентами являются, но не ограничиваются ими, винкристин, даунорубицин, L-аспарагиназа, митоксантрон и амсакрин.
При терапевтическом применении фармацевтические композиции вводят пациенту в количестве, достаточном для продуцирования нужного эффекта и определяемом как "терапевтически эффективная доза". Терапевтически эффективная доза агониста IL-10 может варьироваться в зависимости, например, от конкретной цели лечения, для которого используется данная доза, от способа введения, состояния здоровья пациента и конкретного назначения врача. Так, например, доза для непрерывного вливания будет, в основном, составлять от 500 нг/кг/день до 50 мкг/кг в день. Эта доза рассчитывается исходя из рандомизированных клинических испытаний IL-10 в контролируемых условиях с участием человека [90].
Концентрация агониста IL-10 в фармацевтических композициях может широко варьироваться, то есть от около 10 мкг до около 5 мг/мл, а предпочтительно от около 100 мкг до около 2 мг/мл. Выбор концентрации осуществляют, главным образом, исходя из объемов жидкости, вязкости, и т.п. в соответствии с конкретным способом введения. Так, например, типичная фармацевтическая композиция для внутривенного вливания может быть изготовлена так, чтобы она содержала 250 мл декстрозы/физиологического раствора и 2,5 мкг агониста IL-10.
В твердых композициях могут быть использованы стандартные нетоксичные твердые носители, которыми, например, являются фармацевтически чистые маннит, лактоза, крахмал, стеарат магния, натрийсодержащий сахарин, тальк, целлюлоза, глюкоза, сахароза, карбонат магния, и т.п. Для перорального введения фармацевтически приемлемую нетоксичную композицию приготавливают путем включения в нее обычно используемых носителей, таких как как носители, перечисленные выше, и в основном 10-95%, а предпочтительно 25-75% активного ингредиента, то есть вещества, являющегося агонистом IL-10.
Для аэрозольного введения агонист IL-10 предпочтительно используют в тонкодисперсной форме вместе с поверхностно-активным веществом и пропеллентом. Обычно процентное содержание агониста IL-10 составляет 0,01-20 мас.%, а предпочтительно 1-10%. Само собой разумеется, что это поверхностно-активное вещество должно быть нетоксичным и предпочтительно растворимым в пропелленте. Типичными представителями таких веществ являются сложные эфиры или неполные сложные эфиры жирных кислот, содержащие от 6 до 22 атомов углерода, такие как эфиры, образованные капроновой, октановой, лауриновой, пальмитиновой, стеариновой, линолевой, линоленовой, олестериновой и олеиновой кислотой с алифатическим многоатомным спиртом или с их циклическим ангидридом, таким как, например, этиленгликоль, глицерин, эритрит, арбит, маннит, сорбит, ангидриды гексита, происходящие от сорбита, и полиоксиэтиленовые и пролиоксипропиленовые производные этих сложных эфиров. Могут быть также использованы смешанные или природные глицериды.
Поверхностно-активные вещества (ПАВ) могут составлять 0,1-20% по массе всей композиции, а предпочтительно 0,25-5%. Остальную часть композиции обычно составляет пропеллент. Ожиженные пропелленты обычно представляет собой вещества, являющиеся газами в нормальных условиях, которые были конденсированы под давлением. Подходящими сжиженными пропеллетнами являются низшие алканы, содержащие до 5 атомов углерода, такие как бутан и пропан, а предпочтительно фторированные или фторхлорированные алканы. Могут быть также использованы смеси вышеперечисленных веществ. Для получения аэрозоля контейнер, снабженный подходящим клапаном, наполняют соответствующим пропеллентом, содержащим тонкоиэмельченный(ые) полипептид(ы) и ПАВ. Таким образом, эти ингредиенты содержатся под давлением до тех пор, пока они не будут высвобождены при помощи клапана.
Для увеличения периода полужизни в сыворотке агонист IL-10 может быть инкапсулирован, включен в полость липосом, изготовлен в виде коллоида или использован с применением другой техники, обеспечивающей более продолжительный период попу-жизни полипептидов. Так, например, в некоторых вариантах настоящего изобретения, агонист IL-10 может быть инкапсулирован в липосому. Для получения липосом существует несколько методов, которые описаны, например, в [83, 84, 85 и 86].
Как описано выше, было обнаружено, что IT9302, а также его аналоги и варианты могут быть использованы для предупреждения действия цитокинов, о которых известно, что они играют патогенную роль в вышеописанных патологических состояниях.
Поэтому в данном описании рассматриваются возможности терапии с использованием полипептида настоящего изобретения или его аналогов или производных, и такие возможности должны быть исследованы для всех заболеваний, при которых предполагается терапевтический эффект hIL-10 и/или IRAP (см. табл. 1 и 2).
Пояснения к чертежам
Фиг.1 представляет диаграмму, показывающую, что IT9302 ингибирует спонтанное продуцирование IL-8 очищенными культивированными моноцитами человека.
Фиг. 2 представляет диаграмму, показывающую, что IT9302 ингибирует IL-1-индуцированное (1 нг/мл) продуцирование IL-8 мононуклеарными клетками периферической крови человека.
Фиг.3 иллюстрирует продуцирование IRAP IТ9302-стимулированными моноцитами человека.
Фиг.4 иллюстрирует продуцирование IRAP IL-10-стимулированными моноцитами человека.
Фиг.5 иллюстрирует хемотаксическое действие IT9302 на СD8+-Т-клетки.
Фиг. 6 иллюстрирует десенсибилизацию СD8+-Т-клеток пептидом IT9302, что приводит к невосприимчивости СD8+-Т-клеток к IL-10 (10 нг/мл)-индуцированному хемотаксису.
Фиг. 7 иллюстрирует ингибирование пептидом IT9302 активности IL-8 по отношению к CD4+-T-клeткaм.
Фиг. 8 представляет диаграмму, показывающую, что IT9302 ингибирует MCAF/MCP-1-индуцированный хемотаксис моноцитов.
Фиг. 9 иллюстрирует продуцирование IL-4 в цитозольных фракциях СD4+-Т-клеток с помощью ЭХЛ Вестерн-блот-анализа.
Фиг. 10 иллюстрирует продуцирование TNF-α в цитозольных фракциях смешанной культуры лимфоцитов человека с помощью ЭХЛ-Вестерн-блот-анализа. Вестерн-блот-анализ для TNFα был осуществлен, как описано в "Материалах и методах" для IL-4, за исключением того, что было использовано кроличье антитело против TNFα человека (Pepro Tech. Inc., London, England) и меченное пероксидазой хрена вторичное антитело (кат. Р 217, Dako, Denmark).
Фиг. 11 иллюстрирует регуляцию Т-клеточной пролиферации цитокинами IL-10 и IT9302.
Фиг. 12 иллюстрирует экспрессию HLA-DR на моноцитах человека (проточная цитометрия).
Фиг.13 иллюстрирует, что LPS-индуцированный шок и лейкопения были модулированы IT9302, о чем свидетельствует подсчет полного числа лейкоцитов.
Фиг. 14 иллюстрирует, что инъекция кроликам мышиного IT9302 перед тем, как был индуцирован панкреатит, предупреждает лейкопению.
ПримерЫ
Материалы и методы
Цитокины и хемоатрактанты
Рекомбинантный hIL-10 получали от фирмы Pepro Tech. Inc., NJ. (кат. 200 10). Рекомбинантный hIL-lβ и рекомбинантный hIL-8 были любезно предоставлены Dainippon Pharmaceutical Company, Osaka, Japan. В качестве культуральной среды использовали RPMI 1640 GIBCO, не содержащую LPS, в соответствии с лимулюс-тестом (проба с лизатом амебоцитов) (набор Sigma Е-ТОХАТЕ кат. 210-А1). rh-MCAF/MCP-1 был любезно предоставлен профессором Kouji Matsushima, Kanazawa, Japan.
Анализ на хемотаксис лейкоцитов
Хемотаксис Т-клеток
Субпопуляции CD4+-CD8+-T-лимфoцитoв, характеризующиеся экспрессией либо антигенов CD4, либо CD8, выделяли из гепаринизированной крови нормальных доноров. Так, например, мононуклеарные клетки периферической крови (РВМС) очищали из гепаринизированной крови путем разведения 100 мл крови сбалансированным солевым раствором Хэнкса (HBSS)(1:1), с последующим выделением путем нанесения слоя клеток на LymphopacTM (Nycomed Pharma, Oslo, Norway), и с последующим градиентным центрифугированием при 2000 об/мин в течение 20 минут. Мононуклеарные клетки промывали три раза в HBSS, и клеточный осадок разводили в 4 мл HBSS, содержащем 1% фетальную сыворотку теленка, а затем сортировали при 4oС с использованием Dynabeads, покрытых моноклональным антителом против CD4- или CD8-антигенов (Dynabeads M-450 CD4 кат. 111,16, Dynabeads М-450 CD8 кат. 111.08, DETACHaBEAD кат. 125.04). Отношение гранула: клетка составляло 10:1, а время инкубирования составляло 1 час. Гранулы отделяли путем добавления поликлонального антимышиного антитела в соответствии с инструкциями производителей.
Хемотаксический анализ представляет собой метод с использованием 48-луночной микрокамеры (Neuroprobe, Rock-ville, MD), как описано ранее (74; см. ссылку 3 и ссылку 14). Хемоатрактанты разводили в среде RPMI 1640 (GIBCO кат. 61870-010) с 1%-ной стерильно отфильтрованной околоплодной сывороткой теленка и помещали в нижнюю часть 25 мкл камеры. В случае оценки хемотаксиса Т-клеток Т-клетки (5•106/мл) суспендировали в среде и 50 мкл суспензии помещали в верхнюю часть камеры, отделенную от нижней части камеры поликарбонатным, не содержащим поливинилпирролидона фильтром, имеющим размер пор 5 мкм (Nucleopore Corp., Pleasanton, CA), и покрытым коллагеном типа IV (Sigma кат. С 0543). Клетки оставляли на 2 часа для миграции при 37oС и при 5% СО2. Затем фильтры осторожно удаляли, фиксировали в 70% метанола и окрашивали в течение 5 минут в Кумасси бриллиантовом голубом. Клетки, прикрепленные к нижней поверхности фильтра, подсчитывали путем измерения их площади с использованием видеокамеры на микроскопе, подсоединенном к компьютерной системе для цифрового анализа и поддерживаемом программным обеспечением для объективного определения хемотаксической миграции. Приблизительно 5% Т-клеток будут спонтанно мигрировать, что соответствует 12000-13000 клеткам; причем эта величина может варьироваться в различные дни эксперимента, но очень мало варьируется в день проведения эксперимента. Поэтому, как было описано ранее, для выражения результатов было выбрано отношение между числом клеток, мигрирующих в образце и в негативном контроле, который отражает спонтанную миграцию. Это отношение называется индексом хемотаксиса (CI). Все образцы анализировали в трех дубликатах, а миграцию клеток в каждой лунке измеряли в трех участках, после чего оценивали среднюю величину площади. В некоторых экспериментах мембрану для хемотаксиса не покрывали коллагеном, а поэтому в этой системе анализа мигрирующие клетки опускаются на дно нижней лунки камеры для хемотаксиса.
В первом эксперименте хемотаксическое действие IT9302 на CD8+-Т-клетки оценивали путем тестирования серийных разведений IT9302, добавленных в нижнюю часть камеры, и оценки хемотаксиса, как описано выше.
Во втором эксперименте способность IT9302 десенсибилизировать миграцию СD8+-Т-клеток в ответ на rHIL-10 (10 нг/мл), исследовали путем добавления IT9302 к клеткам-мишеням за 30 минут до хемотаксиса. IT9302 добавляли в серийных концентрациях и хемотаксический ответ rhIL-10 оценивали, как описано выше.
В третьем эксперименте способность IT9302 к супрессии хемотаксического ответа СD4+-Т-клеток на rhIL-8 (10 нг/мл) исследовали путем добавления IT9302 к клеткам-мишеням за 30 минут до осуществления хемотаксиса. IT9302 добавляли в серийных концентрациях и хемотаксический ответ rhIL-8 оценивали, как описано выше.
Хемотаксис моноцитов
Хемотаксис моноцитов измеряли с использованием той же камеры Бойдена, которая была описана выше для Т-клеток. Хемоатрактант MCAF/MCP-1 разводили в среде RPMI 1640 с 0,5% BSA и вводили в нижнюю часть камеры при концентрации 10 нг/мл. Моноциты, очищенные стандартным способом путем адгезии к пластику от нормальных человеческих РВМС, полученных, как описано выше, суспендировали в среде RPMI 1640 с 0,5% BSA, а затем инкубировали в течение 30 минут в присутствии IT9302 при различных концентрациях. Затем клетки вводили в верхнюю часть камеры для хемотаксиса при концентрации 106 клеток/мл. Верхняя и нижняя части камеры были отделены поликарбонатным фильтром, не содержащим поливинилпирролидона и имеющим размер пор 8 мкм (Nucleopore, Pleasanton, CA). Содержимое камеры инкубировали при 37oС в течение 90 минут. Мембраны, содержащие мигрирующие клетки, обрабатывали, как описано выше, а индекс хемотаксиса вычисляли способом, описанным выше.
Продуцирование IL-8 нормальными периферическими мононуклеарными клетками человека (РВМС)
РВМС выделяли путем очистки из гепаринизированной крови здоровых доноров. После градиентного центрифугирования с использованием LymphoprepTM (Nycomed Pharma, Oslo, Norway), мононуклеарные клетки разводили до плотности 2•106 клеток/мл в среде RPMI 1640 (Gibco кат. 6187-010), не содержащей LPS и включающей 1%-ную стерильно отфильтрованную инактивированную нагреванием фетальную сыворотку теленка, а также пенициллин (10000 МЕ/мл), стрептомицин (10 мг/мл) и гентамицин (2,5 мг/мл). Клетки культивировали в 24-луночных планшетах Nunc Micro Plates (Nunc, Denmark) и в присутствии различных концентраций IT9302 (0,1 мкг, 100 нг, 10 нг, 1 нг, 0,1 нг, 0,01 нг/мл) в течение 24 часов. После инкубирования в течение 24 часов добавляли еще IT9302, а через 1 час в клеточные культуры добавляли r-hIL-lβ (1 нг/мл). Супернатанты собирали после всего 48-часового периода инкубирования и концентрацию секретированного IL-8 измеряли с помощью EUSA-анализа IL-8 с использованием набора для IL-8-ELISA (Dainippon Pharmaceutical Co., Ltd., Osaka, Japan). Вкратце, стандарты и клеточные супернатанты инкубировали в течение одного часа в дубликатах при 20oС на микропланшет-шейкере. После промывки добавляли вторичное антитело в течение одного часа, а затем инкубировали в течение одного часа меченным пероксидазой козьим антителом против кроличьих IgG. После промывки, реакционную смесь проявляли O-фенилендиамином. Через 30 минут реакцию прекращали путем добавления 1,6 н. серной кислоты. Оптическую плотность (ОП) измеряли на ELISA-ридере при 490 нм.
Концентрацию IL-8 вычисляли с помощью калибровочной кривой зависимости оптической плотности от концентраций IL-8-стандарта.
Определение концентрации IRAP
РВМС очищали, как описано выше. РВМС культивировали в среде RPMI 1640 с 10% стерильной инактивированной нагреванием фетальной сывороткой теленка (включая пенициллин, 10000 МЕ/мл, стрептомицин, 10 мг/мл, гентамицин, 2,5 мг/мл) так, чтобы концентрация клеток составляла 5•106 клеток/мл. Затем моноциты очищали стандартным способом путем адгезии к пластику. Эти моноциты культивировали в RPMI 1640 с 2% FCS (2,5•106 клеток/мл) и с различными разведениями rhIL-10 или IT9302. Клетки стимулировали в течение 24 часов и супернатанты собирали для определения IRAP. ELISA-анализ IRAP проводили с использованием набора для иммуноанализа Human IL-Ira Quantikin Immune-assay Kit от фирмы R&D Systems Europe Ltd. (кат. DRA 00, Abingdon, Oxon, UK).
Определение пролиферации Т-клеток
Анализ на пролиферацию
РВМС (2•105) культивировали в 200 мкл среды RPMI, содержащей 10% FCS, в течение 72 часов вместе с РНА (0,5 мкл/мл) и rhIL-10 (1, 10, 100 нг/мл) или IT9302 (0,1, 1, 10 нг/мл) в трех дубликатах. В последние 18 часов добавляли 3H-тимидин при 0,5 мкКи/лунку (Amersham, Denmark). Клетки собирали на фильтре (Glass Microfibre Filters, Whatman, кат. 1822 849) и добавляли сцинтилляционную жидкость (Ultima, Gold, MV, Packard). Подсчет импульсов проводили на модели Tri-Carb 1600 TR, Packard.
Определение экспрессии антигена МНС класса II на моноцитах
Экспрессия HLA-DR на моноцитах человека.
Моноциты выделяли из свежей гепаринизированной крови путем адгезии к пластику при 37oС в RPMI 1640, содержащей 10% FCS. После инкубирования супернатант удаляли, добавляли раствор Хэнкса (4oС с 1% FCS) и клетки отделяли путем охлаждения при -20oС в течение 15 минут, и легкого постукивания об стол. Моноциты стимулировали (2•106 клеток/мл) в RPMI 1640, содержащей 2% FCS, путем добавления IFNγ (10 нг/мл) либо добавления rhIL-10 (100, 10, 1 нг/мл), а затем IFNγ, либо добавления IT9302 (10, 1, 0,1 нг/мл), а затем IFNγ в течение 40 часов. Клетки отделяли от лунок путем охлаждения, как указано выше. Свежие нефиксированные клетки были использованы для типирования поверхности антителом против HLA-DR человека. Клетки суспендировали в растворе Хэнкса с 1% FCS при концентрации 1•106 клеток/мл и добавляли в течение 45 минут FITC-конъюгированные мышиные антитела против HLA-DR, DP, DK человека (F 0817, DAKO, Denmark). Клетки три раза промывали в растворе Хэнкса и FACS-анализ проводили на проточном цитометре Coulter-Epics XL-MCL при длине волны 488 нм. Неспецифическое связывание определяли с помощью нерелевантного FITC-конъюгированного антитела (мышиное антикозье антитело DAKO F 479).
Альтернативно, клетки фиксировали в 10% ДМСО, 40% RPMI 1640 и 50% стерильной FCS, хранили при -80oС и использовали для типирования ДНК.
Определение апоптоза в моноцитах
Типирование ДНК стимулированных моноцитов.
Фиксированные клетки инкубировали в 70% этанола в течение 60 минут и дважды промывали в растворе Хэнкса. 1•106 Клеток ресуспендировали в 250 мкл РНКазы, 1 мкг/мл в 1,12% цитрата натрия, рН 8,4 (Рибонуклеаза A, Phannacia 17-0442-01) и инкубировали в течение 3 минут при 37oС. После этого добавляли 250 мкл иодида пропидия (50 мкг/мл в растворе Хэнкса) и клетки инкубировали в темноте в течение 30 минут. Затем клетки дважды промывали в растворе Хэнкса (Jensen et al., [110]). Содержание ДНК измеряли с помощью проточной цитометрии на Coulter-Epics XL-MCL при длине волны 550 нм.
Результаты
Регуляция пролиферации Т-клеток под действием IL-10 и IT9302
РВМС стимулировали с помощью РНА, как описано выше, а затем в течение 72 часов добавляли rhIL-10 и IT9302 и включение 3H-тимидина измеряли в течение последних 18 часов. rhIL-10 и IT9302 негативно регулировали пролиферацию клеток при оптимальной концентрации 100 нг/мл rhIL-10 и 10 нг/мл IT9302, как показано на Фиг.11.
Экспрессия HLA-DR на моноцитах человека
Моноциты очищали, как описано выше, и стимулировали в течение 40 часов, добавляя IFNγ (10 нг/мл), и/или rhIL-10 и/или IT9302 за 30 минут до добавления IFNγ. Экспрессию антигена МНС II исследовали путем инкубирования с ПТС-конъюгированным мышиным антителом против HLA-DR, DP, DK человека. IFNγ позитивно регулировал экспрессию антигена МНС II, как показано на фиг.12, а 10 нг/мл rhIL-10 и 1 нг/мл IT9302 негативно регулировали экспрессию антигена МНС II аналогичным способом.
Типирование ДНК стимулированных моноцитов
Моноциты, стимулированные IFNγ, как описано выше, инкубировали с иодидом пропидия и содержание ДНК измеряли с помощью проточной цитометрии. Измеряли фракцию клеток, экспрессирующую ДНК в фазе G1 или G2 пролиферации клеток, а также измеряли фракцию, экспрессирующую апоптоз. Как видно на фиг.12, не стимулированные моноциты экспрессируют 6,6%-ный апоптоз, а IFNγ-стимуляция подавляет апоптоз до 4,1%, в то время как IL-10-, IFNγ- и IT9302 + IFNγ-стимуляция клеток индуцирует фракцию апоптоза до 10,3% и 9,3% соответственно.
Определение продуцирования IL-4 CD4+-T-лимфоцитами
Клеточные культуры
CD4+-T-лимфoциты выделяли путем очистки из гепаринизированной крови здорового человека. После градиентного центрифугирования с использованием LymphoprepTM (Nycomed Pharma, Oslo, Norway), мононуклеарные клетки сортировали при 4oС с использованием гранул Dynabeads (Dynal AS, Norway), покрытых моноклональными антителами против CD4. Гранулы отделяли путем добавления поликлонального антимышиного антитела (Dynal, AS, Norway). Чистота позитивно отобранных клеток составляла выше 99%, на что указывал FACS-анализ. При оценивании продуцирования de novo IL-4 IL-8-стимулированными Т-клетками, Т-клетки культивировали при 5•106 клеток/мл в среде RPMI 1640 (Gibco кат. 61870-010), не содержащей LPS и включающей 1%-ную стерильно отфильтрованную инактивированную нагреванием фетальную сыворотку теленка (FCS), пенициллин (10000 МЕ/мл), стрептомицин (10 мг/мл) и гентамицин (2,5 мг/мл).
Т-клетки стимулировали в течение 3 дней с использованием rIL-8 (100 нг/мл), rIL-10 (100 нг/мл), IT9302 (10 нг/мл) и IFNγ (10 нг/мл). Рекомбинантный IL-8 человека (rhIL-8) был любезно предоставлен фирмой Dainippon Pharmaceutical Co. Ltd. , Osaka, Japan, a IFNγ был закуплен у фирмы Boehringer Ingelheim Am Rhein, Germany. Для получения специфического ингибирования стимуляции IL-8 было использовано нейтрализующее моноклональное антитело против IL-8 (WS.4) (любезно предоставленное фирмой Dr.K.Matsushima, Japan). Рекомбинантный IL-10 поставлялся фирмой Pepro Tech. Inc. (London, England).
Получение клеточного материала и супернатанта культуры для гель-электрофореза
Культивированные Т-клетки и культуральные среды отделяли путем центрифугирования при 2000 об/мин в течение 5 минут. Супернатанты сушили вымораживанием, а затем растворяли в 100 мкл буфера для лизиса. Клетки ресуспендировали непосредственно в 100 мкл гелевого буфера для лизиса [9]. Полученный материал выдерживали при -80oС до тех пор, пока не была проведена дополнительная оценка.
ЭХЛ-Вестерн-блот-анализ белков, происходящих от CD4+-Т-клеток
Клетки или супернатанты лиофилизованной клеточной культуры использовали для определения содержания белка IL-4. Белки из одномерных 15% ДСН-ПААГ-гелей переносили посредством блоттинга на нитроцеллюлозные мембраны Hybond-ЭХЛ (Amersham RPN 2020D, UK) и блокировали 5% альбумином бычьей сыворотки (Sigma) в забуференном Трисом физиологическом растворе с рН 7,8, содержащем 0,1% Твин-20. Затем блоты инкубировали с поликлональным козьим антителом против человеческого IL-4 (R & D Systems, UK), после чего инкубировали со вторичным антителом, меченным пероксидазой хрена (кат. RPN 2106 ЭХЛ, Amersham, UK), и иммунное окрашивание детектировали путем экспонирования пленки (Kodak X-OMAT-S, USA) в течение 90 секунд.
Пример 1
Ингибирование спонтанного продуцирования IL-8 на моноцитах человека
Тест осуществляли, как описано в разделе "Продуцирование IL-8 нормальными мононуклеарными клетками периферической крови человека (РВМС)". Моноциты очищали методом адгезии к пластику и 3,0•106 клеток/мл стимулировали в течение 40 часов. Как показано на фиг.1, IТ9302 ингибировал продуцирование IL-8 моноцитами, а при 0,1 нг/мл IT9302, продуцирование IL-8 ингибировалось до 35% от спонтанного продуцирования in vitro. Жизнеспособность клеток всегда превышала 80% после 1-дневного культивирования, а добавление IT9302 ни в этом, ни в следующих примерах не влияло на их жизнеспособность при любой концентрации IT9302 от 0,1 до 1000 нг/мл (мол. масса IT9302: 1127 Дальтон, предсказанная мол. масса rhIL-10: 18400 Дальтон).
Пример 2
Ингибирование IL-1β-индуцированного продуцирования IL-8 мононуклеарными клетками периферической крови человека (РВМС)
Тест проводили, как описано в разделе "Продуцирование IL-8 нормальными мононуклеарными клетками периферической крови человека (РВМС)". Как показано на фиг. 2, IT9302 дозозависимым способом ингибировал IН-1β-индуцированное продуцирование IL-8 мононуклеарными клетками периферической крови человека in vitro. Супрессия продуцирования IL-8 обнаруживала "плато" при концентрациях IT9302 между 0,01 и 100 нг/мл.
Пример 3
Продуцирование белка-антагониста рецептора интерлейкина-1 (IRAP) на моноцитах человека
Тест проводили, как описано в разделе "Определение концентрации IRAP". Как показано на фиг.3, IT9302 дозозависимым способом индуцировал продуцирование IRAP на моноцитах человека. Продуцирование резко увеличивается при использовании концентраций IT9302, составляющих выше 10 нг/мл. На фиг.4 показано индуцирование IRAP рекомбинатным hIL-10, поскольку hIL-10 приблизительно в 20 раз больше, чем IT9302, то 5 нг/мл IT9302 равно 100 нг/мл IL-10 в молярном отношении. Следовательно, эффективности IT9302 и rhIL-10 отношении индуцирования IRAP являются сравнимыми и при низких концентрациях приблизительно равны. При концентрациях IT9302, превышающих 10 нг/мл, индуцирование IRAP увеличивается и достигает максимального уровня 60 нг/мл. Кроме того, была протестирована специфичность этого индуцирования антителом, которое специфически направлено против IT9302. В отдельном эксперименте, где моноциты спонтанно продуцировали 3,5 нг/мл IRAP и они были стимулированы 1-10 нг/мл IT9302 до макисмального продуцирования IRAP 10,6+0,6 нг/мл, это продуцирование должно быть блокировано поликлональным антителом для IT9302, добавленным в количестве 2 мкг/мл за 30 минут до добавления 10 нг/мг IT9302 так, чтобы уровень IRAP снижался до 2,9+0,3 нг/мл. Эти результаты противоположны результатам, полученным с антителом, направленным против IL-10, 19F1, которое добавляли аналогичным способом (2 мкг/мл IT9302 за 30 минут до добавления 10 нг/мл IT9302) и которое не блокировало продуцирование IRAP, а, напротив, стимулировало это продуцирование до 22 нг/мл. Этот результат можно объяснить тем, что это антитело способно нейтрализовать эндогенный IL-10, но не IT9302. Спонтанно продуцированный IL-10 в клеточной культуре оказывает негативное саморегулирующее действие на 1Т9302-продуцирование IRAP.
Пример 4
Хемотаксическое действие на CD8+-Т-лимфоциты человека
Эксперимент проводили, как описано в разделе "Хемотаксический анализ лейкоцитов". Как показано на фиг.5, IT9302 индуцировал хемотаксическую миграцию СD8+-Т-лимфоцитов человека in vitro, но не оказывал влияния на СD4+-Т-клетки (данные не показаны). И снова, эффективность IT9302, показанная в этом эксперименте, является сравнимой с эффективностью rhIL-10, показанной ранее [14].
Пример 5
Десенсибилизация CD8+-Т-клеток человека, приводящая к невосприимчивости к rhIL-10
Эксперимент проводили, как описано в разделе "Хемотаксический анализ лейкоцитов". К суспензии СD8+-Т-клеток в течение 30 минут добавляли IT9302, а затем эти клетки тестировали на их хемотаксический ответ к rhIL-10. Как показано на фиг.6, предварительное инкубирование клеток с IT9302 приводило к супрессии восприимчивости СD8+-Т-клеток к rhIL-10. Это указывает на то, что IT9302 может влиять на связывание rhIL-10 с рецептором IL-10.
Пример 6
Супрессия хемотаксического ответа CD4+-Т-лимфоцитов по отношению к IL-8
Эксперимент проводили, как описано в разделе "Хемотаксический анализ лейкоцитов". Как показано на фиг.1, IT9302, добавленный к суспензии CD4+-T-лимфoцитoв человека, дозозависимым образом ингибирует ответ CD4+-T-клeтoк на IL-8.
Пример 7
Супрессия хемотаксического ответа моноцитов человека на MCAF/MCP-1
Эксперимент проводили, как описано в разделе "Хемотаксис моноцитов". Как показано на фиг.8, IT9302, добавленный к суспензии моноцитов человека, дозозависимым образом ингибирует хемотаксический ответ моноцитов на MCAF/MCP-1.
Пример 8
Ингибирование экспрессии молекулы МНС класса И на моноцитах человека при IFNγ-стимуляции
В новой экспериментальной модели (проточной цитометрии) было показано, что IFNγ-стимулированные моноциты стимулируют их экспрессию антигена МНС II, и это индуцирование может быть блокировано IL-10 и IT9302 аналогичным способом. В то же время оба пептида осуществляют позитивную регуляцию числа клеток, обнаруживающих апоптоз в моноцитах (см. фиг.12). Эти эксперименты указывают на то, что IL-10 и IT9302 могут ингибировать моноцит/макрофагзависимую пролиферацию Т-клеток путем уменьшения антиген-представляющей способности моноцитов посредством негативной регуляции экспрессии МНС класса II (см. фиг.12).
РНА-стимулированные РВМС инициируют пролиферацию, как показано путем включения 3H-тимидина, и могут также супрессироваться интерлейкином IL-10 и пептидом IT9302. РНА стимулирует пролиферацию Т-клеток посредством активации Са2+-зависимых каналов. Потоки Са2+ в Т-клеточных клонах человека уменьшались, если клетки были предварительно инкубированы с IL-10 (Таgа, К. et al., 1992). Настоящий эксперимент показал, что пролиферация Т-клеток негативно регулируется IT9302, а также регулируется под контролем Са2+-зависимых каналов.
Обсуждение экспериментов
Полученные данные демонстрируют дозозависимое ингибирующее действие синтетического нонапептида, IT9302 на процессы, которые отражают провоспалительную активность, включая продуцирование IL-8 и миграцию моноцитов и/или Т-клеток. Таким образом, IT9302 обладает способностью ингибировать спонтанное продуцирование IL-8 моноцитами человека, культивированными в течение ночи. Это может быть объяснено непосредственным ингибирующим действием на продуцирование мРНК IL-8 и/или последующее продуцирование и/или высвобождение белка. Другой механизм может быть объяснен тем фактом, что моноциты, культивированные in vitro, будут экспрессировать и продуцировать IL-1, который, в свою очередь, будет затем индуцировать продуцирование IL-8. Это подтверждается тем фактом, что, как было продемонстрировано, IT9302 в значительной степени индуцирует продуцирование IRAP моноцитами. Следовательно, IT9302 может также ингибировать спонтанное продуцирование IL-8 путем блокирования активности IL-1. Очевидно, наблюдаемое IТ9302-индуцирование IRAP стимулирует биологически активный IRAP, поскольку IT9302, добавленный к культурам, препятствует IL-1-индуцированному продуцированию IL-8, но только в том случае, если добавление проводят, по крайней мере, за 16 часов до добавления IL-1 к культурам. Это может означать, что IT9302 ингибирует IL-1-индуцированное продуцирование IL-8 путем стимуляции продуцирования IRAP, который затем, в свою очередь, блокирует активность IL-1 посредством его рецептора. Если IT9302 непосредственно ингибирует продуцирование IL-8, то следует ожидать, что добавление IT9302 к культурам за один час до добавления IL-1 должно ингибировать продуцирование IL-8, что не имело места (данные не приводятся). Поэтому наблюдаемое ингибирование IT9302 продуцирования IL-8, очевидно, обусловлено индуцированием продуцирования IRAP, а не прямым ингибированием продуцирования IL-8. IT9302 специфически блокирует рецептор IL-1 путем индуцирования IRAP и тем самым индуцирования других цитокинов, которые стимулируются интерлейкином IL-1, TNFα, IL-8 и, вероятно, многих других цитокинов и факторов. Специфичность индуцирования подтверждается тем, что наше антитело против IT9302 может блокировать индуцирование IRAP. Другое антитело против IL-10, 19F1, не блокирует IТ9302-индуцированное продуцирование IRAP.
IT9302 также имитирует активность IL-10 путем супрессии способности CD4+-T-клeтoк мигрировать в ответ на IL-8. Поскольку IL-8 ассоциируется со многими различными воспалительными состояниями и поскольку СD4+-Т-клетки появляются довольно рано в инфильтрате Т-клеточно-опосредованного иммунного воспаления, такого как аллергия кожи, то можно утверждать, что эта особенность имеет существенное терапевтическое значение для регуляции иммунного воспаления, опосредованного Т-клетками.
Продемонстрированная хемотаксическая активность IT9302 по отношению к СD8+-Т-клеткам является также аналогичной активности IL-10 и, кроме того, IT9302 может также активировать популяции Т-клеток посредством супрессорной активности, способствующей прекращению Т-клеточно-опосредованного иммунного воспаления. Поэтому в соответствии с примерами, продемонстрированными выше, IT9302 обладает терапевтическим действием при заболеваниях, в которых IL-10 и/или IRAP могут также оказывать терапевтическое действие. Кроме того, IT9302 может также обладать терапевтической активностью при заболеваниях, при которых IL-8 и/или MCAF и/или IL-1 играют, очевидно, патогенную роль. В настоящем изобретении описаны аналоги IT9302, то есть вещества или пептиды, имеющие, по крайней мере, некоторые из вышеописанных свойств.
Пример 9
Индуцирование продуцирования IL-4 в CD4+-Т-лимфоцитах
Предварительное общее описание
Подобно IL-10 IL-4 представляет собой продукт CD4+-T-клеток типа Тн2. По наблюдениям было установлено, что рекомбинантный IL-10 человека индуцирует продуцирование IL-4 культивированными CD4+-T-клeткaми человека. Это означает, что IL-10 помимо своих собственных иммуносупрессорных функций также индуцирует продуцирование другого иммуносупрессорного цитокина IL-4. Поэтому были также проведены испытания для того, чтобы убедиться в том, что IT9302 также индуцирует продуцирование IL-4 CD4+-T-клeткaми.
Для этого СD4+-Т-клетки, очищенные, как описано в главе "Методы для хемотаксиса Т-клеток", и культивированные, как описано в разделе "Определение продуцирования IL-4 CD4+-T-лимфоцитами", были в течение 3 дней подвергнуты стимуляции пептидом IT9302 (10 нг/мл) или интерлейкином IL-10 (100 нг/мл). Цитозольные фракции собирали и анализировали на содержание в них IL-4 методом Вестерн-блот-анализа (фиг. 9) с использованием козьего поликлонального антитела против IL-4 человека.
Как продемонстрировано на фиг.9, наблюдения показали, что IL-10, а также IT9302 индуцируют продуцирование IL-4 культивированными нормальными CD4+-T-клeткaми человека.
Пример 10
Инаибирование продуцирования TNF-α в процессе реакции смешанной культуры лейкоцитов (MLR)
Было продемонстрировано, что реакция смешанной культуры лейкоцитов частично зависит от продуцирования TNFα во время реакции. Было показано, что IT9302 не вызывает значительного снижения MLR, однако было обнаружено, что при этом имеет место значительное снижение продуцирования TNF-α в процессе MLR.
Таким образом, MLR осуществляли путем очистки лейкоцитов человека с последующим культивированием в течение 4 дней 1 миллиона клеток/мл, взятых от аллогенных доноров. Перед получением культуры одну группу клеток в течение 2 минут облучали бета-излучением. Цитозольную фракцию белков очищали, как описано в разделе "Определение продуцирования IL-4 CD4+-Т-лимфоцитами", а Вестерн-блот-анализ осуществляли с использованием кроличьего антитела против TNF-α человека.
Как продемонстрировано на фиг.10, значительное снижение продуцирования TNFα наблюдалось в процессе реакции смешанной культуры лимфоцитов.
Пример 11
Модуляция LPS-индуцированного шока и лейкопении у свиней
Поскольку было обнаружено, что IT9302 модулирует продуцирование цитокинов, включая TNFα и IL-8, что было подтверждено опубликованной последовательностью IL-10 свиньи [88], то был проведен тест на способность IT9302 модулировать развитие LPS-индуцированной лейкопении у свиней (фиг.13).
В предварительном эксперименте был проведен тест для того, чтобы определить, как внутривенная инъекция 0,1 мг/кг IT9302 модулирует действие внутривенной инъекции 2 мкг/кг LPS у свиней (N=3). IT9302 инъецировали за 30 минут до инъекции LPS, после чего брали пробы крови, как описано на фиг.13. Затем определяли полное число лейкоцитов, а также число дифференциальных клеток и исходя из этих результатов подсчитывали полное число нейтрофильных лейкоцитов.
Как показали наблюдения, инъекция LPS вызывала кратковременную лейкопению. Однако инъекция IT9302 способствовала предупреждению лейкопении, как показано на фиг.13.
Пример 12
Влияние мышиного IT9302, гомологичного человеческому IT9302, на высвобождение TNFα в плазме после введения LPS мышам
Было продемонстрировано на животных, что рекомбинантный мышиный IL-10 может защищать мышей от летальной эндотоксемии [89] и, кроме того, было продемонстрировано, что IL-10, введенный человеку, обладает ингибирующим действием на Т-клетки и подавляет продуцирование провоспалительных цитокинов [90] . Целью этого исследования было определить, может ли IT9302 подавлять продуцирование TNFα у мышей, подвергнутых эндотоксин-индуцированному шоку после введения LPS (фиг.14).
Для этого эксперимента было использовано 8-недельные мыши BALB/C (полученные из Centre for Research Animals, Bomhoitgaard," Bomholtvej 10, DK-8680 Ry, Denmark). LPS от e.coli Были получены от Sigma (партия 3444114) а мышиный IT9302 был получен от Schafer-N, Lers⊘  42, DK-2100 Copenhagen 0, Denmark.
42, DK-2100 Copenhagen 0, Denmark.
Мышам внутрибрюшинно инъецировали 100 мкг LPS в 1 мл 0,9 М NaCl. За 60 минут до инъекции LPS мышам вводили IT9302, эквивалетный 0,1 мкг, 1 мкг и 10 мкг человеческого IL-10. Группы из 5 животных анестезировали (Imobilon, Pherrovet,  Sweden) через 1, 2, 3 и 4 часа соответственно, после чего посредством пункции в область сердца брали максимум цельной крови. Эту кровь центрифугировали в условиях холода, сыворотку собирали и хранили при -70oС для последующего анализа на содержание TNFα с помощью ELISA (TNFα Mouse Immuno Assay Diagnostic kits, Genzyme, Cambridge, MA, USA).
Sweden) через 1, 2, 3 и 4 часа соответственно, после чего посредством пункции в область сердца брали максимум цельной крови. Эту кровь центрифугировали в условиях холода, сыворотку собирали и хранили при -70oС для последующего анализа на содержание TNFα с помощью ELISA (TNFα Mouse Immuno Assay Diagnostic kits, Genzyme, Cambridge, MA, USA).
Как показали наблюдения, 10 мкг мышиного IT9302 явно ингибировали TNFα в мышиной сыворотке по сравнению с контрольной группой, которой не вводили mIТ9302 (см. табл. 3).
Пример 13
Модуляция острого панкреатита, индуцированного желчной кислотой, с использованием мышиного IT9302
В качестве in vivo-моделей часто используются грызуны, мыши и кролики, а недавно был разработан метод Genzyme Diagnostic с использованием набора для оценки перекрестной реактивности цитокиновых продуктов (Cytokine Research Products). В частности, с использованием набора для иммуноанализа, проводимого на мышах, можно измерить кроличьи TNFα и IFNγ. Поскольку при изучении патофизиологической роли IL-8 в экспериментальном остром панкреатите в качестве рабочей модели мы использовали кроличью модель, то нами было проведено исследование действия мышиного IT9302 на индуцированную лейкопению (фиг. 14). Наша гипотеза состояла в том, что даже IL-10 могут быть идентичными у мышей и кроликов и способны влиять на IL-8-индуцированную лейкоцитарную инвазию в поджелудочной железе во время острого панкреатита.
Экспериментальная модель
Белые новозеландские кролики (Oryctofagus cuniculus) весом 1,7-4,0 кг содержались на голодной диете в течение 18-24 часов. Рабочие процедуры проводили в соответствии с описанием Banerjee et al., 1994 (91) и Hong et al., 1962 (92), а острый панкреатит у кроликов индуцировали 5% желчной кислотой, введенной в проток поджелудочной железы. Мышиный IT9302 (100 мкг/кг) вводили через центральную вену за 30 минут до введения желчной кислоты и сразу после введения этой кислоты подкожно в дозе 100 мкг на кг веса кролика. Образцы плазмы собирали как показано на фиг.14.
Как показали наблюдения, инъекция IT9302 кроликам до индуцирования панкреатита способствовала предупреждению лейкопении, как продемонстрировано на фиг. 14.
Пример 14
Модуляция острого панкреатита у кроликов, индуцированного желчной кислотой, с использованием IT9302 человека
Была использована такая же экспериментальная модель, что и в примере 13, за исключением того, что мышиный IT9302 был заменен на человеческий IT9302. Острый панкреатит индуцировали путем внутрипротоковой инъекции 2,0 мл 5% хемодезоксихолина (желчной кислоты) (10 животным). Другую группу животных обрабатывали человеческим IT9302 (8 животных), при этом первая доза (100 мкг/кг) была введена подкожно, а вторая доза (100 мкг/мл) внутривенно за полчаса до индуцирования острого панкреатита (см табл. 4, 5). Для измерения уровня цитокинов собирали образцы сыворотки в интервалы времени через 0, 1, 3, 6 и 12 часов после индуцирования острого панкреатита.
Методы измерения уровня TNFα и IL-8 в сыворотке кроликов
В модифицированной версии метода измерения уровней TNFα в сыворотке кроликов был использован набор для ELISA мышиного TNFα (Genzyme, code 80-3807-00). Этот набор содержал иммобилизованное моноклональное антитело хомячка против мышиного TNFα, которое захватывает присутствующий в сыворотке TNFα; конъюгированное с пероксидазой хрена козье антитело против мышиного TNFα; субстрат и хромаген ТМВ. Время инкубирования для сыворотки и TNFα-стандарта составляло 4 часа, а время проявления при инкубировании субстрата составляло 30 минут.
Измерение уровня кроличьих IL-8 в сыворотке кроликов проводили с использованием специального набора для IL-8-ELISA, любезно предоставленного профессором Kouji Matsushima, Tokyo, Japan. Для этого в качестве иммобилизованного (для захвата) антитела, вторичного антитела и идентифицирующего антитела использовали моноклональное антитело против IL-8, антитело морской свинки против кроличьих IL-8, и конъюгировнные со щелочной фосфатазой кроличьи антитела против иммуноглобулинов G морской свинки соответственно. Методы см., Ikeda et al. [106].
Результат
Краткий обзор
Максимальное индуцирование TNFα достигалось за 3 часа, а максимальные уровни IL-8 наблюдались через 6 ч. Уровни TNFα и IL-8 в кровотоке заметно снижались в течение 0-12 часов. Были также измерены уровни панкреатических ферментов в крови (амилазы, липазы и триптазы), и все они обнаруживали пик через 3-6 часов после индуцирования острого панкреатита, но ни один из этих ферментов не обнаруживал воздействия IТ9302-обработки. Гистологически, аналогичные панкреатические изменения наблюдались в двух группах, но при этом имело место значительное уменьшение интерстициальной нейтрофильной инфильтрации в легких.
Заключение
IT9302 ингибирует продуцирование TNFα и IL-8 в модели острого панкреатита, индуцированного желчной кислотой, и блокирует нейтрофильную инфильтрацию в легких обработанных кроликов, что способствует предупреждению развития РДСВ (респираторного дистресс-синдрома взрослых) - подобного синдрома у этих животных, и через 12 часов приводит к снижению смертности на 60-0%. Интерлейкин-lβ считается важным индуктором острого панкреатита [107], и это подтверждает нашу гипотезу, что IT9302 может блокировать все индуцибельные эффекты IL-1. Соображения, по которым человеческий IT9302 может быть использован в кроличьей модели, могут быть объяснены наблюдением, сделанным Dan Gaur et al. и опубликованным в этом году в Nature [108], где он показал, что филогенетически белки кроликов (Lagomorpha) являются более близкими к белкам приматов (человека), чем к белкам грызунов (мышей, крыс,...).
Пример 15
Использование IТ9302 для лечения рака и предупреждения метастазов
Недавно было показано [111], что системное введение клеточного IL-10 индуцирует эффективный специфический и продолжительный иммунный ответ in vivo на развившиеся опухоли у мышей. Было предположено, что этот эффект может быть частично объяснен известным действием IL-10 на клетки различных типов, включая ко-стимуляцию пролиферации Т-клеток, хемоатракцию СD8+-Т-клеток, и стимуляцию активности активированных лимфокинами клеток-киллеров. Наблюдения показали также, что человеческий IL-10 может способствовать реверсии локального иммуносупрессорного действия вирусного IL-10. Другая группа исследователей подтвердила потенциальную терапевтическую роль введения IL-10 при раке [112], поскольку они наблюдали, что IL-10 ингибирует метастазы посредством природного опосредованного клетками-киллерами механизма в in vivo-эксперименте на мышах, имеющих различные опухоли, включая метастазирующую злокачественную меланому. Kundi et al. (1996) [113] сообщали, что IL-10 обнаруживает антиметастатическое и противоопухолевое действие в мышиной модели рака молочной железы человека.
Эти наблюдения свидетельствуют о возможной роли введения человеческого IL-10 и IT9302 в биологической терапии рака.
Пример 16
IT9302 в качестве иммунного адъюванта при лечении вирусных инфекций
Известно, что IL-10 обладает определенным антивирусным действием. Так, например, Kollmann et al. (1996) [114] обнаружили, что IL-10 ингибирует in vivo острую ВИЧ-инфекцию у SCID-мышей (с тяжелым комбинированным иммунодефицитом) с человеческим фетальным тимусом и печенью. Мы наблюдали, что двое из заявителей/исследователей с использованием IT9302-содержащего препарата в виде крема (полная доза 400-500 мкг) при окклюзии с помощью пластичного бинта на коже спины в течение 24 часов, добивались у себя увеличения числа СD8+-T-клеток (60% и 90% соответственно) и увеличение концентрации IRAP в сыворотке от 1 нг/мл до 2 нг/мл. У семи необработанных волонтеров концентрация IRAP никогда не превышала 1,1 нг/мл. Случайным и неожиданным был тот факт, что оба исследователя наблюдали на коже одинаковые бугорки (вследствие кожной HPV-инфекции), явно свидетельствующие о воспалении, причем краснота и зуд вокруг этих бугорков появлялась через 3-4 дня после применения IТ9302-содержащего крема. Одновременно воспалительная реакция наблюдалась на нескольких пальцах одного из десяти индивидуумов, а у другого индивидуума наблюдался лишь один бугорок на пальце. В обоих случаях размер этих бугорков быстро и постепенно снижался в последующие 4-7 дней так, что через 10-11 дней не оставалось никаких клинических признаков от этой инфекции в виде бугорков. При последующем контроле, проводимом 2 месяца спустя, не было обнаружено рецидива инфекции.
Поэтому нами было установлено, что IT9302, возможно, посредством системной активации клеток-киллеров и/или активности цитотоксических СD8+-Т-клеток обладает способностью индуцировать латентную иммунную реакцию в ответ на кожную HPV-инфекцию, что в конечном приводит к клинической ремиссии вирусной инфекции. Таким образом, IT9302 является, возможно, альтернативным терапевтическим агентом, который может быть использован для лечения вирусных инфекций, таких как ВИЧ- и HPV-инфекции.
Пример 17
IT9302 в качестве иммунного адъюванта при лечении воспалительных болезней суставов (артритов)
Очевидно, что анти-IL-1-терапия, а также анти-TNFα-терапия имеет большое клиническое значение для лечения артритов (Maini, 1996) [115]. Как описано в настоящем документе, нами было обнаружено, что IT9302 является ингибитором продуцирования TNFα, а также стимулятором IRAP (белок-антагонист рецептора IL-1) мононуклеарных клеток человека. Так, например, IT9302 является потенциальным агентом, оказывающим терапевтическое воздействие при артритах. Это было подтверждено наблюдением одного из заявителей настоящего изобретения, который после нанесения IТ9302-содержащего крема (приблизительно 500 мкг) на кожу в течение 24 часов наблюдал значительное ослабление хронической боли в суставе, вызываемой артритом. Эти наблюдения проводили три раза, и в каждом случае после прекращения нанесения IТ9302 симптомы периодически снова возникали в течение последующей недели. Таким образом, нами было найдено in vivo - подтверждение аргументов в пользу использования IT9302 для лечения острых или хронических воспалительных реакций, таких как артриты или другие аутоиммунные болезни.
REFERENCES
1. Bendtzen К. Lymphokines in inflammation. Inflammation Basic Mechanisms Tissue Injuring Principles and Clinical Models (P Venge & A Lindbom eds) 1985; Almqvist & Wiksell International. Stockholm: 187-217.
2. Bendtzen К. Interleukin-1, Interleukin-6, and tumor necrosis factor in infection, inflammation and immunity. Immunol Lett 1988; 19:183-192.
3. Larsen C.G. Leukocyte activating and chemotactic cytokines in cell-mediated immune reactions of the human skin. Acta Dermatovenerol. 1991; Suppl. 160:1-48.
4. Fiorentino D.F., M.W.Bond, and T.R.Mosmann. 1989. Two types of mouse helper T cell. IV. Th2 clones secrete a factor that inhibits cytokine production by Thl clones. J. Exp. Med., 170:2081.
5. Viera P., R. de Wall-Malefyt. M.-N.Dang, K.E.Johnson, R.Kastelein, D. F. Fiorentiono, J. E. de Vries, M.-G.Roncarolo, T.R.Mosmann, and K.W.Moore. 1991. Isolation and expression of human cytokine synthesis inhibitory factor (CSIF/IL-10) cDNA clones: homology to Epstein-Barr virus open reading frame BCRFI. Proc. Natl. Acad. Sci. (USA), 88:1172.
6. Moore, K. W. , O'Garra A., de Waal Malefyt R., Vieira, Mosmann T.R. 1993. Incerleukin-10. Annu Rev. Immunol, 11:165-90.
7. Kim, J. M., Brannan, C.I. Copeland N.G., Jenkins, N.A., Khan, T.A., Moore, K.W. 1992. Structure of the mouse interleukin-10 gene and chromosomal localization of the mouse and human genes. J. Immunol 148:3618-23.
8. Carter, D.B., Deibel, M.R-Jr, Dunn, C.J. et al. 1990. Purification, cloning, expression and biological characterization of an interleukin-1 receptor antagonist protein. NATURE 344:633-638.
9. Hannum, C. H.. Wilcox. C.J., Arend, W.P. et al. 1990. Interleukin-1 receptor antagonist activity of a human interleukin-1 inhibitor. Nature 343: 336-40.
10. Firescein, G.S., Boyle, D.L., Yu, C., et al. 1994. Synovial interleukin-1 receptor antagonist and interleukin-1 balance in rheumatoid arthritis. Arthritis Rheum 37:644-652.
11. Fisher, C. J.-Jr., Slotman, G.J., Opal, S.M., Pribble, J.P. et al. 1994. Initial evaluation of recombinant interleukin-1 receptor antagonist in the treatment of sepsis syndrome: a randomized, open-label, placebo-controlled multicenter trial. The IL-1RA Sepsis Syndrome Study Group. Crit-Care-Med. 22:12-21.
12. de Waal-Malefyt, R., Haanen J., Spits, H., et al. 1991. IL-10 and viral IL-10 strongly reduce antigen-specific human T cell proliferation by diminishing the antigen-presenting capacity of monocytes via down-regulation of class II MHC expression. J. Exp. Med. 174:915-24.
13. Gazzinelli, R.T., Oswald, I.P., James, S.L., Sher, A., 1992. IL-10 inhibits parasite killing and nitric oxide production by IFN-/-activated macrophages. J. Immunol. 148:1792-96.
14. Jinquan, Т., Larsen, C.G., Gesser, В., Matsushima, K., Thestrup-Pedersen, K. 1993. Human IL-10 is a chemoattractant for CD8+ Т lymphocytes and an inhibitor of IL-8- induced CD4+ Т lymphocyte Migration. Journal of Immunology, 151:4545-4551.
15. Rousset F., E.Garcia, T.Defrance, C.Peronne. D.-H.Hsu, R.Kastelein, K. W.Moore, and J.Banchereau. 1992. IL-10 is a potent growth and differentiation factor for activated human B lymphocytes. Proc. Natl. Acad. Sci. USA, 175:671.
16. Howard, M., O'Garra, A., Ishida, H., de Waal Malefyt. R., de Vries, J. 1992. Biological properties of Interleukin-10. J. Clin. Immunol 12:239-47.
17. Kuhn, R. , Lohler, J., Rennick, D., Rajewsky, K., Muller, W. 1993. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 75: 263-74.
18. Sher, A. , Fiorencino, D.F., Caspar, P., Pearce. E., Mosmann, T. 1991. Production of IL-10 by CD4+ lymphocytes correlates with down-regulation of Th1 cytokine synthesis in helminth infection. J. Immunol. 147:2713-16.
19. Clerici, M., Shearer, G.M. 1993 Immunology Today. 14:107-111.
20. Bry, К., Lappalainen, U. 1994. Interleukin-4 and transforming growth factor-beta 1 modulate the production of incerleukin-1 receptor antagonist and prostaglandin E2 by decidual cells. Am-J-obstet-Gynecol 170 (4):1194-1198.
21. Firestein, G., S., Boyle, D.L., Yu. C., Paine, M.M., Whisenand, T.D. , Zvaifler, N.J., Arend, W.P. 1994. Synovial interleukin-1 receptor antagonist and interleukin-1 balance in rheumatoid arthritis. Arthritis-Rheum, 37/5: 644-652.
22. Roberge, С.J., De-Medicis, R., Dayer, J.M., Rola-Pleszcyczynski, M., Naccahe, P. H., Poubelle, P.E. 1994. Crystal-induced neutrophil activation: V. Differential production of biologically active IL-1 receptor antagonist. J. Immunol 152/11: 5485-5494.
23. McCall, R.D., Haskill, S., Zimmermann, E.M., Lund, P.K., Thompson, R. C. , Sartor, R.B. 1994. Tissue interleukin 1 and interkeukin-1 receptor antagonist expression in entercolitis in resistant and susceptible rats. Gastroenterology (4):960-72.
25. Kimble, R.В., Vannice, J.L. Bloedow, D.C., Thompson, R.C., Hopfer. W. , Kung. V.Т., Brownfield, C., Pacifici, R. 1994. Interleukin-1 receptor antagonise decreases bone loss and bone resorption in ovariectomized rats. J. Clin Invest. 93/5:1959-1967.
26. Kline, J.N., Geist, L.J., Monick, M.M., Stinski, M.F., Hunninghake, G.W., 1994. J. Immunol. 152 (5): 2351-7.
27. Tompkins, R.G. 1994. Human recombinant interleukin-1 receptor antagonist in the treatment of sepsis syndrome (editorial; comment). Crit-Care-Med. 22 (l): 3, 22 (1):12-21.
28. Everaerdt, В. , Brouckaert, P., Fiers, W. 1994. Recombination IL-1 receptor antagonist protects against TNF-induced lethality in mice. J. Immunol. 152/10: 5041-5049.
29. Fischer, С.J.Jr., Slotman, G.J., Opal. S.M., Pribble, J.P., Bone, R. C. , Emmanuel, G., Ng, D., Bloedow, D.C., Cacalano, M.A. 1994. Initial evaluation of human recombination interleukin-l receptor antagonist in the treatment of sepsis syndrome: a randomized, open-label, placebo-controlled multicencer trial. The IL-1RA Sepsis Syndrome Study Group (see comments). Crit-Care-Med. 22(1): 12-21, 22(1): 3.
30. Gomez-Reino-Carnoco, J.J. 1994. New terapies in rheumatoid arthritis. Med-Clin 543-545.
32. Nishihara, Т. , Ohsaki, Y., Ueda. N., Saito, N., Mundy, G.R. 1994. Mouse interleukin-l receptor antagonist induced by actinobacillus actinomycetemcomitans lipo-polysaccharide blocks the effects of interleukin-l om bone resorption and osteoclast-like cell formation. Infect-Immun. 62(2): 390-7.
33. Simon, C., Frances, A., Piquette, G.N., el-Danasouri, I., Zurawski, G., Dang, W., Polan, M.L. 1994. Embryonic implantation in mice is blocked by interleukin-l receptor antagonist (see comments). Endocrinology. 134(2): 521-8, 134(2): 519-20.
34. Baergen, R., Benirschke, К., Ulich, Т.R., 1994. Cytokine expression in the placenta. The role of interleukin 1 and interleukin 1 receptor antagonise expression in chorioamnionitis and parturition. Arch-Pathol-Lab-Med. 118(1): 52-5.
35. Tang. W. W., Feng, L., Vannice. J.L., Wilson, С.В. 1994. Interleukin-1 receptor antagonist ameliorates experimental antiglomerular basement membrane antibody-associated flomeulonephritis. J. Clin-Invest. 93 (l): 279-9.
36. Cassatella, M.A., Meda, L., Gasperini, S., Calzetti, F., Bonara. S. 1994.
37. Inierleukin 10 (IL-10) upregulates IL-1 receptor antagonist production from lipopolysaccharide-stimulated human polymorphonuclear leukocytes by delaying mRNA degradation. J. Exp-Med. 179/5: 1695-1699.
38. Mancini, R. , Bendetti, A., Jezequel, A.M. 1994. An interleukin-1 receptor antagonist decreases fibrosis induced by dimethylnitrorsamine in rat liver. Virchows-Arch. 424/1: 25-31.
39. Lukacs, N.W., Kunkel, S.L., Burdick, M.D., Lincoln, P.M., Stricter, R.M. 1993.
40. Interleukin-l receptor antagonist blocks chemokine production in the mixed lymphocyte reaction. Blood. 82(12): 3668-74.
41. Bandara, G. , Mueller, G.M., Galea-Lauri, J., Tindal, M.H., Georgescu, H. I. , Suchanek. M. K., Hung, G.L., Gloriso, J.C., Robbins, P.D., Evans., C.H. 1993.
42. Incraarticular expression of biologically active interleukin l-receptor-antagonist protein by ex vivo transfer. Proc-Natl-Acad-Sci-U-S-A. 90(22): 10764-8.
43. Dinarello, С. А. 1994. Anci-incerleukin-l strategies in the treatment of the septic shock syndrome. Can-J-infect-Dis. 5 (suppl. A): 9A-16A.
44. Oelmann, E. , Topp, M.S., Reufi, В., Papadimitriiou, C., Koeningsmann, М. , Oberberg, D. , Thiel, E., Berdel, W.E. 1994. Int-J-Oncol. 4/3: 555-558.
45. Estrov, z. 1993. Interruption of autocrine and paracri-ne growth-stimolatory mechaisms: a new therapeutic stracefy for chronic myelogenous leukemia. Semin-Hema-tol. 30(3 suppi 3): 35-6.
46. Wooley, P.H., Whalen, J.D., Chapman, D.L., Berger, A.E., Richard, K. A. , Aspar, D.G., Staice. N.D. 1993. The effect of an interleukin-1 receptor antagonist protein on type II collagen-induced arthritis and antigen-induced arthritis in mice. Arthritis Rheum. 36 (9): 1305-1314.
47. Peterson, C. M., Hales, H.A., Hatasaka, H.H., Mitchell, M.D., Rittenhouse. L. , Jones. K.P. 1993. Interleukin-1 beta (IL-l beta) modulates prostaglandin production and the natural IL-l receptor antagonist inhibits ovulation in the optimally stimulated rat ovarian perfusion model. Endocrinology 133 (5): 2301-2306.
48. Estrov, Z., Kurzrock, R., Talpaz, M. 1993. Role of interleukin-1 inhibitory molecules in therapy of acute and chronic myelogenous leukemia. Leuk. Lymphoma 10 (6): 407-418.
49. Chensue, S.W., Bienkowski, M., Eessalu, Т.Е., Warmington, K.S., Hershey, S.D., Lukacs, N.W., Kunkel, S.L. 1993. nous IL-l receptor antagonist protein (IRAP) regulates schistosome egg granuloma formation and the regional lymphoid response. J. Immunol. 151 (7): 3654-3662.
50. Bowyer, J.F., Davies, D.L., Schmued, L., Broening, H.W., Newport, G. D. , Slikker. W. Jr. , Holson, R.R. 1994. Further studies of the role of hyperthermia in methamhetamine neurotoxicity. J. Pharmacol. Exp. Ther. 268/3: 1571-1580.
51. Cole, O.F., Sullivan, M.H.F., Elder, M.G. 1993. The 'interleukin-1 receptor antagonist' is a partial agonise of prostaglandin synthesis by human decidual cells. Prostaglandins 46/6: 493-498.
52. Jenkins, J.K., Arend. W.P. 1993. Interleukin l receptor antagonist production in human monocyces is induced by IL-lalpha. IL-3, and IL-4 and GM-CSF. Cytokine 5/5: 407-415.
53. Coceani, F., Lees, J., Redford, J., Bishai, I. 1992. Incerleukin-l receptor antagonist: effectiveness against incerleukin-1 fever. Can. J. Phannacol. 70 (12): 1590-1596.
54. Schiro, R., Longoni. D., Rossi, V., Maglia, O., Doni, A., Arsura, M. , Carrara. G., Masera, G., Vannier, E., Dinarello, C.A., Rambaldi, A., Biondi, A. 1994. Suppression of juvenile chronic myelogenous leukemia colony growth by interleukin-1 receptor antagonist. Blood 83/2: 460-465.
55. Watson, M.L., Smith, D., Bourne, A.D., Thompson, R.C., Westwick, J. 1993. Cytokines contribute to airway dysfunction hyperreactivity, pulmonary eosinophil accumulation and tumor necrosis factor generation by pretreatment with an interleukin-1 receptor antagonist. Am. J. Respir. Cell Mol. Biol. 8 (4): 365-369.
56. Abhyankar. S., Gilliland, D.G., Ferrara, J.L.M. 1993. Interleukin-1 is a critical effector molecule during cytokine dysregulation in graft-versus-host disease to minor histocompatibility antigens. Transplantation 56/6: 1518-1523.
57. Lan, H. Y., Nikolic Paterson, D.J., Zarama, M., Vannice, J.L., Atkins, R.C. 1993. Suppression of experimental crescentic glomerulonephitis by the interleukin-1 receptor antagonist. Kidney Inc. 43 (2): 479-485.
58. Herve, P. 1993. Prevention and treatment of acute GvHD New modalities. Nouv. Rev. Fr. Hematol. 35/3: 295-297.
59. Conti, P., Panara, M.R., Barbacane. R.C., Placido, F.C., Bongrazio, M., Reale, M., Dempsey, R.A., Fiore. S. 1992. Blocking the incerleukin-1 receptor inhibits leukotriene B4 and prostaglandin E2 generation in human monocyte cultures. Cell Immunol. 145 (I): 199-209.
60. Kristensen, M., Deleuran. В., Eedy, D.J., Feldmann. M., Breathnach, S. M., Brennan, F.M. 1992. Distribution of interleukin-1 receptor antagonist protein (IRAP), interleukin-1 receptor, and incerleukin-1 alpha in normal and psoriatic skin. Decreased expression of IRAP in psoriatic lesional epidermis. Br. J. Dermatol. 127 (4): 305-311.
61. Romero, R. , Sepuiveda, W., Mazor, M., Brandt, F., Cotton, D.B., Dinarello, C. A: . Mitchell, M.D. 1992. The natural interleukin-l receptor antagonist in term and pre-term parturition. Am. J. Obstet. Gynecol. 167 (4 Pt 1): 863-872.
62. Dinarello, C.A. 1992. Reduction of inflammation by decreasing production of interleukin-l or by specific receptor antagonism. Int. J. Tissue. React. 14 (2): 65-75.
63. Conti, P., Panara, M.R., Barbacane, R.C., Bongrazio, M:, Dempsey, R. A., Reale, M. 1993. Human recombinant IL.l receptor antagonist (IL-IRa) inhibits leukotriene B4 generation from human monocyte suspensions stimulaced by lipopolysaccharide (LPS). Clin. Exp. Immunol. 91/3: 526-531.
64. DeForge, L.E., Tracey, D.E., Kenney, J.S., Remick, D.G. 1992. Interleukin-1 receptor antagonist protein inhibits incerleukin-8 expression in lipopolysaccharide-stirouled human whole blood. Am. J. Pathol. 140 (5): 1045-1054.
65. Porat, R. , Poutsiaka, D.D., Miller, L.C., Granowicz, E.V., Dinarello, C. A. 1992. Interleukin-l (IL-1) receptor blockade reduces endotoxin and Borrealia burgdorferi-stimulated IL-8 synthesis in human monoclear cells. Faseb. J. 6 (7): 2482-2486.
66. Boermeester, M.A., van Leeuwen, P.A.M., Schneider, A.J., Houdijk, A. P. J. , Ferwerda, C.C., Wesdorp, R.I.C. 1993. Interleukin-l receptor antagonist: A new therapeutic agent in the treatment of septic syndrome. Ned. Tijdschr. Geneesks. 137/7: 337-342.
67. Smith, R.J., Chin, J.E., Sam, L.M., Justen, .J.M. 1991. Biologic effects of an interleukin-l receptor antagonist protein on interleukin-l-stimulated cartilage erosion and chondrocyte responsiveness. Archsicis Rheum. 34 (1): 78-83.
68. Conti, P. Barbacane, R.C., Panara, M.R., Reale, M., Placido, F.C., Fridas, S. , Bongrazio, M., Dempsey, R.A. 1992. Human recombinant incerleukin-1 receptor antagonist (hrIL-lra) enhances the stimulatory effect of interleukin-2 on natural killer cell activity against MOLT-4 target cells. Int. J. Iinmunopharm. 14/6: 987-993.
69. Selig, W., Tocker, J. 1992. Effect of interleukin-1 receptor antagonist on antigen-induced pulmonary responses in guinea pigs. Eur. J. Pharmacol. 213/3: 331-336.
70. McCarthy, P.L. Jr., Abhyankar, S., Neben, S., Newman, G., Sieff, C., Thompson, R. C., Burakoff, S.J., Ferrara. J.L.M. 1991. Inhibition of interleukin-l by an interleukin-1 receptor antagonist prevents graft-versus-host diseases. Blood 78/8: 1915-1918.
71. Estrov, Z, Kurzrock, R., Wetzler, M., Kantarjian, H., Blake, M., Harris. D., Gutterman, J.U., Talpaz, M. 1991. Suppression of chronic myelongenous leukemia colony growth by interleukin-l (IL-1) receptor antagonist and soluble IL-1 receptors: A novel application for inhibitors of IL-1 activity. Blood 78/6: 1476-1484.
72. Thomas, Т.К., Will, Р.С., Srivastava, A., Wilson, C.L., Harbison. M. , Little, J., Chesonis, R.S., Pignatello. M., Schmolze, D., Symington. J., Kilin, P. L. , Thompson, R.C. 1991. Evaluation of an interleukin-l receptor antagonist in the rat acetic acid-induced colitis model. Agents Actions 34/1-2: 187-190.
73. Carter, D.B., Deibel, M.R. Jr., Dunn, C.J., Tomich, C.S.C., Laborde, A. L., Slightom, J.L., Berger, A.E., Bienkowski, M.J., Sun, F.F., McEwan, R. N. , Harris, P.K.W., Yem, A.W., Waszak. G.A., Chosay, J.G., Sieu. L.C., Hardee. M.M., Zurcher Neely, H.A., Reardon. I.M., Heinrikson, R.L. et al. 1990. Purification, cloning expression and biological characterization of an interleukin-l recepcor antagonist protein. Nature 344/6267: 633-638.
74. Larsen C.G, Anderson A.O., Apella E., Oppenheim J.J., Matsushima K., 1989. Science 243:1464;
75. Larsen C. G., Jinquan Т., Deleurant В., Thestrup-Pedersen K. 1993, IL-10 is a potent regulator of the chemotactic response of mononuclear cells, but not of granulocytes. J. Invest. Dennatol. Vol 100, No 6.
76. Sankoff and Kruskal in chapter l of "Time Warps, String Edits, and Macromolecules: The Theory and Practice of Sequence Comparison" (Addison-Wesley, Reading, Mass. 1983).
77. Berzofsky, Science 229, (1985) 932-940.
78. Bowie еt а1., Science 247, (1990) 1306-1310.
79. Wassennan et al., J. Immunol. 87. 1961. 290-295.
80. Levine et al., Methods in Enzymolocy 11, 1967, 928-936.
81. Lewis et al., Biochemistry 22, 1983, 948-954.
82. Rene de Waal Maletyt, John Abrahams, Bruce Bennet, Carl G. Figdor and Jan E. de Vries (1991), Interleukin 10 (IL-10) Inhibits Cytokine Synthesis by Human Monocytes: An Autoregulatory Role of IL-10 Produced by Monocytes. J. Exp. Med. 174, 1209-1220.
83. Szoka ее al., Ann. Rev. Biophys. Bioeng. 9, 1980, 467.
84. US Patent No. 4235871.
85. US 4501728.
86. US 4837028.
87. Walter H. Gotlieb. John S. Abrams, Joanna M. Watson, Thierry J. Velu, Jonathan S. Berek, Oconiel Martinez-Meza (1992), Presence of incerleukin 10 (IL-10) in the ascites of patients with ovarian and other intra-abdominal cancers. Cytokine 4, No. 5, 385-390.
88. Blanche G., P. Gianello, Sh. Germana, M. Baetscher, D.H. Sachs and Chr. LeGuern. (1995), Molecular identification of porcine incerleukin 10: Regulation of expression in kidney allograf model. Proc. Natl. Acad. Sci. USA 92, 2800-2804.
89. Howard, M., Т. Muchamuel, S. Andrade, S. Menon (1993), Interleukin 10 protects mice from lethal endotoxemia. J. Exp. Med. 177, 1205-1208.
90. Chernoff, A. E., E.V. Granowitz, L. Shapiro. E. Vannier, G. Lonnemann. J. B. Angel, J. S. Kennedy, A.R. Rabson, S. Wolff, C.A. Dinarello (1995), A randomized, controlled trial of IL-10 in humans. Inhibition of inflammatory cytokine production and immune responses. J. Immunol. 154. 5492-5499.
91. Banerjee, А. К., S.W. Galloway and A.N. Kingsnorth (1994), Experimental models of acute pancreatitis. Br.J. Surg. 81, 1096-1103.
92. Hong, S.S., D.S. Chin, T.S. Cho, S.E. Kirn (1962), Experimental pancreatitis induced by alcohol and bile in rabbits. Annals of Surgery 156(6), 929-939.
93. Bodansky, M. (1984), Principles or Peptide Synthesis. Springer-Verlag, Berlin.
94. Pelton, J.T., et al., Proc. Natl. Acad. Sci. USA 82, 233-239.
95. Dyson, H., et al. (1988). Annual Review of Biophysics and Biophysical Chemistry 17, 305-324).
96. Nakanishi, H., et al. (1993), Peptidomimetics of the immunoglobulin supergene family a review. Gene 137, 51-56.
97. US 5446128.
98. Walter et al. (1995), Biochemistry 34, 12118-25.
99. Marshall. G.R. (1993), Tetrahedron 49, 3547-3558.
100. Merrifield, R.B. (1963), J. Amer. Chem. Soc. 85, 2149-2154).
101. Kent, S.B.H. (1988), Annu. Rev. Biochem. 57. 957-989.
102. Carpino, L.A. and Han, G.Y. (1972), J. Org. Chem. 37. 3404-3409.
103. Stewart, J.M. and Young, J.D. (1983), "Solid Phase Peptide Synthesis", Pierce Chemical Company, Rockford. Illinois.
104. Atherton, E. and Sheppard, R. C. (1989), "Solid Phase Pepcide Synthesis". IRL Press at Oxford University Press.
105. Pennington, M.W. and Dunn, B.M. (eds.) (1994), "Peptide Synthesis Protocols", Humana Press, Totowa, New Jersey.
106. Ikeda, N. et al. (1995), Infection and. Immunity. 4812-4817.
107. Fink, G.W. and Norman, J.G. (1996), "Intrapancreatic Incerleukin-1β Gene Expression by Specific Leukocyte Populations during Acute Pancreacitis", J. Surgical Research 63. 369-373.
108. Gaur. D. et al. (1996), "Phylogenetic position of the order Lagomorpha (rabbits, hares and allies)". Nature, 379, 333-335.
109. Poli, G. ее а1. (1994), Proc. Natl. Acad. Sci. USA 91, 108-112.
110. Jensen, I.M. et al. (1993), Analyt. Cell. Pathol. 5, 213-223.
111. Berman, R.M., Suzuki, T. et al. (1996), "Systemic administration of cellular IL-10 induces an effective, specific, longlived immune response against established tumors in mice", J. Immunol. 157, 231-238.
112. Zheng, L.M., Ojcius. D.M. et al. (1996), "IL-10 inhibits tumor metastasis through an NK cell-dependent mechanism", J. Exp. Med. 184, 579-584.
113. Kundu, N., Beaty, T.L. et al. (1996), "Antimetastatic and anti-tumor activities of IL-10 in a murine model of breast cancer", J. Natl. Cancer Inst. 88, 479-480.
114. Kollmann, T.R., Pettoello-Mantovani. M. et al. (1996), "Inhibition of acute in vivo HIV infection by human IL-10 treatment of SCID mice implanted with human fetal thymus and liver", Proc. Natl. Acad. Sci. USA 93, 3126-3131.
115. Maini, R.N. (1996), "A perspective on anti-cytokine and anti T cell directed therapies in rheumatoid arthritis", Clin. Exp. Rheumatol. 13, suppl. 12. S35-40.
Синтетические аналоги
Указанные ниже синтетические пептидные аналоги IT9302 были синтезированы профессором Arne Holm в Королевском Университете Ветеринарии и Сельского Хозяйства, Копенгаген, и испытаны авторами письма на их способность индуцировать IRAP в культурах клеток, что было измерено твердофазным иммуноферментным анализом ELISA (Quantikine Immunoassay, Human IL-Ira DROO, R&D Systems, UK).
Условия эксперимента
Пептиды были восстановлены в стерильном отфильтрованном растворяющем буфере PBS с рН 7,4 и/или PBS с рН 7,4, содержащем 4% альбумина бычьей сыворотки (Sigma, A-9647). После этого 1•106-2•106 очищенных моноцитов были стимулированы 0, 1, 10 и 100 нг/мл пептидов, разведенных до эквимолярной концентрации по отношению к rhIL-10, в RPMI, содержащем 2% фетальной телячьей сыворотки, в течение 24 часов (FCS, North American, Life Technology, cat. Nr. 16000044).
Перечень синтетических пептидов, которые были испытаны, приведен в табл. 6.
А= L-аланин, Y= L-тирозин, М=L-метионин, Т=l-треонин, K-L-лизин, I-L-изолейцин, R=L-аргинин, N=L-аспарагин, m=D-метионин, а=D-аланин, у=D-тирозин, t= D-треонин, k=D-лизин, i=D-изолейцин, r=D-аргинин, n=D-аспарагин, МЕА=метоксиэтиламин (пептоид), IВUА=1-бутиламин (пептоид), βА=бета-аланин, Сhа=циклогексилаланин, Руа=пиридилаланин, Met(O)=метионин-S-оксид, Nle=нopлeйцин, Nуа=норвалин, Огn=орнитин, Dab=2,4-диаминомасляная кислота.
Димерный пептид синтезировали, как указано в статье: Ligand Presenting Assembly. A Method for C-and N-terminal antigen presentation, A.Holm, R.M. Jorgensen, S.Ostergaard, and M.Theisen. J.Peptide Res. (2000) 56, 105-113.
Метод LPA дает возможность связывания свободных α-аминокислотных групп на аминоконцевой части двух пептидов IT9302, тогда как две пептидных цепи с полной защитой боковой цепи с дикарбоновой кислотой остаются связанными с синтетической смолой.
Испытание на растворимость
Отвешивали части пептидов примерно 1 мг и растворяли в 1 мг/мл забуференного фосфатом физиологического раствора (PBS) с рН 7,4 и хранили в течение ночи при температуре -80oС. После этого отбирали образец 100 мкл и анализировали на содержание аланина (или другой аминокислоты) для определения растворимости пептидов. После первого испытания тест на концентрацию показал, что несколько из синтетических пептидов являются труднорастворимыми, поэтому было решено перед добавлением PBS с рН 7,4 добавить к каждому пептиду 10 мкл ДМСО (Диметилсульфоксид, Merck 1.02931) для растворения агрегатов. Затем профессор Arne Holm снова провел испытание на концентрацию (см. табл. 7).
Дополнительные измерения
Продуцирование IRAP нг/мл - Исп.4
Нестимулированные клетки - 15,8±0,9
Клетки, стимулированные 1 нг/мл rIL-10 - 22,3±0,6
10 нг/мл - 44,0±0,3
100 нг/мл - 25,7±0,4
Испытание 4 осуществляли с добавлением 10 мкл ДМСО, но без испытания на концентрацию.
Аналоги с эквимолярной концентрацией по отношению к rIL-10 приведены в табл. 8.
Выводы:
На основании первых трех испытаний с индуцированном IRAP было предложено 6 аналогов, обладающих, по меньшей мере, такой же активностью, как и исходный пептид (см. табл. 9).
Профессор A. Holm сделал следующее заключение.
Met в положении x4 может быть замещен норвалином Nva как редкой аминокислотой. Туr (Y) в положении x2 также демонстрирует такую же возможность. Такие замещения могут придать стабильность против протеазной активности. В N-концевой части имеются специальные возможности для замещения. А1а (А) может быть заменен на IBua, который является N-бутилглицином или глутаминовой кислотой IBua (E-1Bua). Такие замещения могут придать стабильность против пептидазной активности. Модификация с 1Bua также предполагает, что могут быть предпочтительными аналоги, являющиеся более липофильными. Липофильный аналог может проявить более длительную устойчивость на участке применения и тем самым продлить время активации. Вопрос о С-концевой стабилизации еще не решен.
В конце был также синтезирован IT9302 димер для целей стабилизации. Димерный пептид показал такой же минимальный уровень активности, что и 6 выбранных аналогов.
IL-10 и участки связывания рецептора IL-10
Кристаллическая структура IL-10 и его растворимый рецептор IL-l0Rα показали димер IL-10, связывающий два растворимые рецептора, A.Zdanov et al. (1996) Protein Science 5: 1955-1962.
Позднее на втором рецепторе IL-10 был обнаружен IL-l0Rβ, являющийся важной субъединицей рецептора IL-10, S.D.Spencer et al. (1998) J.Exp.Med. vol. 187. No. 4571-578.
Картирование сайтов IL-10/рецептора IL-10 показало, что СООН концевая часть связывается с субъединицей IL-10 Rα. U. Reineke et а1. (1998) Protein Science, 7: 951-960. Был сконструирован мономер IL-10 человека, и в отличие от IL-10 дикого типа он показал 1:1 комплексы с растворимым IL-10R (Rα). Связывания IL-10 мономера с IL-10 Rα было достаточно для рекрутинга цепи рецептора сигнальной трансдукции (IL-l0Rβ) в сигнальный комплекс и выявления IL-10 клеточных ответов. K. Josephson et а1. (2000) Vol. 275, No. 18, 13552-13557.
Были получены и проанализированы пептиды, перечисленные ниже, и их идентичность была подтверждена методом масс-спектрометрии по времени пролета с лазерной десорбцией/ионизацией на матрице (MALDI-TOF-MS) с использованием оборудования для время-пролетной линейно-лазерной десорбционной спектрометрии Fisons Tofspec E MALDI. Все измерения соответствовали расчетным массам в пределах рабочих характеристик оборудования (таблица 10).
Список последовательностей приведен после формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИД С IL-10 ПОДОБНЫМИ СВОЙСТВАМИ (ВАРИАНТЫ), КОНЪЮГАТ ПОЛИПЕПТИДА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБ МОДУЛЯЦИИ ИММУННОГО ОТВЕТА У МЛЕКОПИТАЮЩЕГО | 1995 |
|
RU2218348C2 |
| АНТИТЕЛА И ПОЛИПЕПТИДЫ, НАПРАВЛЕННЫЕ ПРОТИВ CD127 | 2017 |
|
RU2769352C2 |
| РЕКОМБИНАНТНЫЙ ХИМЕРНЫЙ БЕЛОК ANTH1, КОДИРУЮЩАЯ ЕГО НУКЛЕИНОВАЯ КИСЛОТА И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2322455C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ НА CD127 | 2015 |
|
RU2734076C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО ПРОТИВ IL17A И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2804963C2 |
| СПОСОБЫ ПРИМЕНЕНИЯ CD32B X CD79B-СВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРИ ЛЕЧЕНИИ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ И РАССТРОЙСТВ | 2017 |
|
RU2772434C2 |
| АНТИТЕЛО ПРОТИВ ИНТЕРФЕРОН-АЛЬФА/БЕТА-СВЯЗЫВАЮЩЕГО БЕЛКА II (IFNAB-BPII) | 1995 |
|
RU2362781C2 |
| АНТИТЕЛА ПРОТИВ VSIG4 ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2776638C1 |
| МОДИФИЦИРОВАННЫЕ ГЕМОПОЭТИЧЕСКИЕ СТВОЛОВЫЕ КЛЕТКИ/КЛЕТКИ-ПРЕДШЕСТВЕННИКИ И Не-Т ЭФФЕКТОРНЫЕ КЛЕТКИ, И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2733652C2 |
| АНТИТЕЛО ПРОТИВ ИНТЕРФЕРОН-α/β-СВЯЗЫВАЮЩЕГО БЕЛКА I (IFNAB-BPI) | 1995 |
|
RU2363705C2 |
















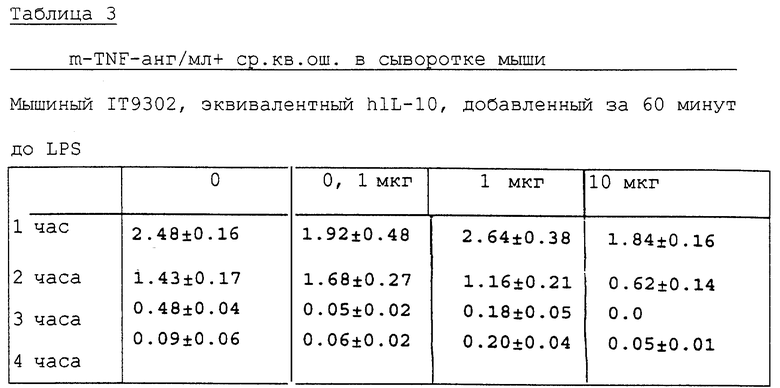


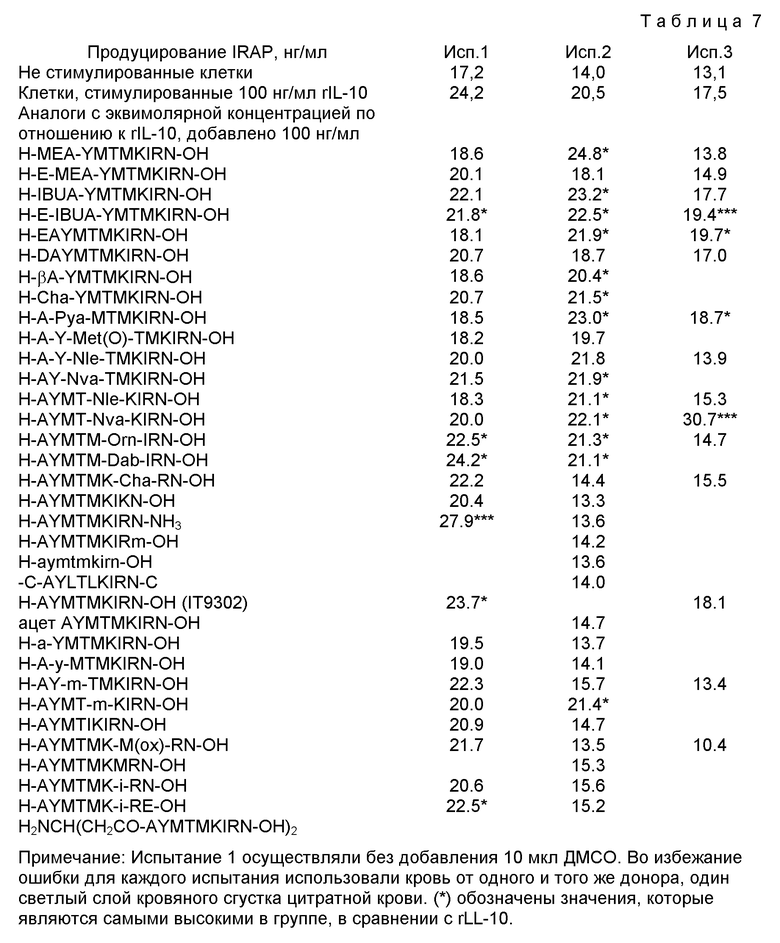



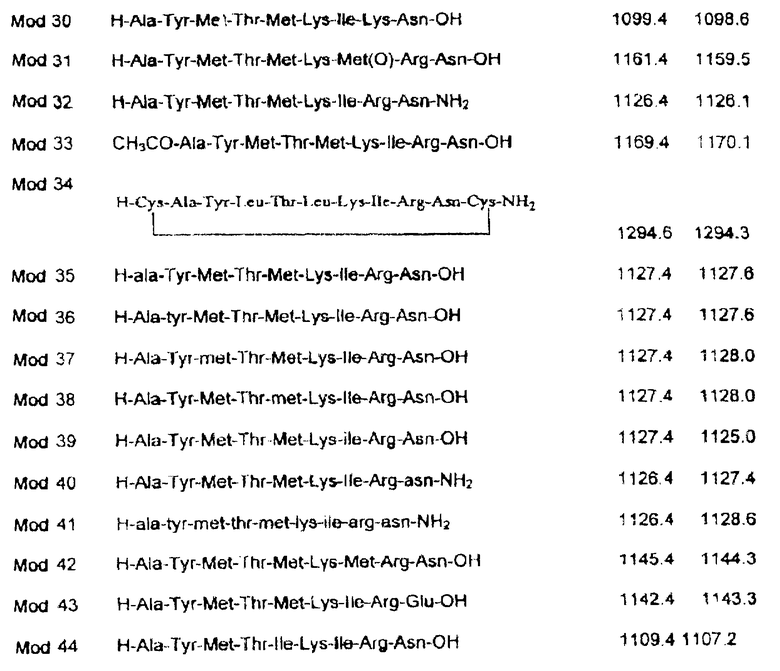

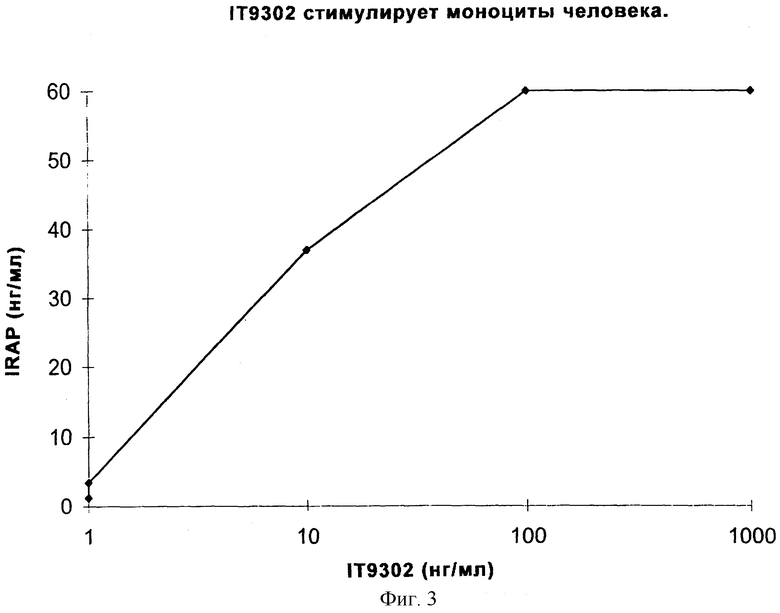
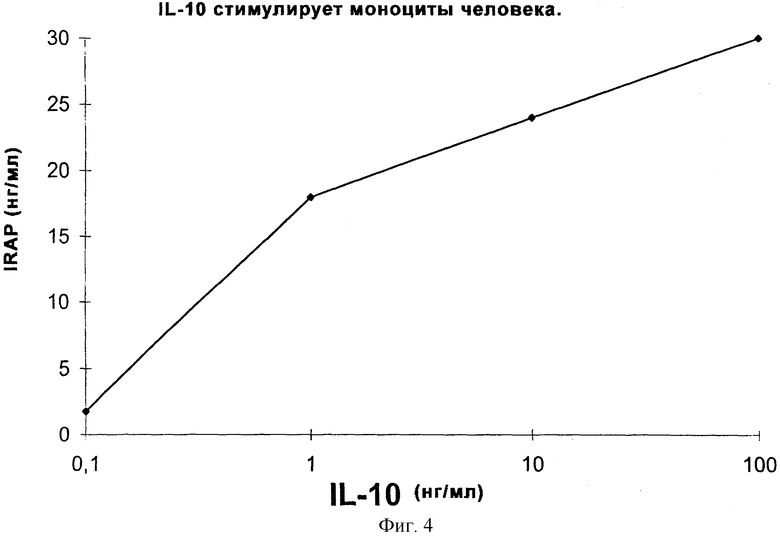

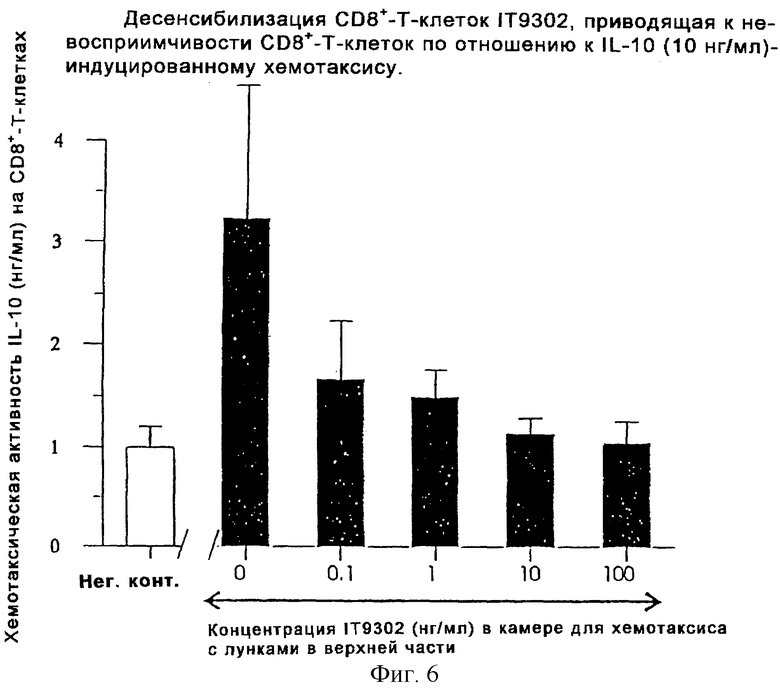
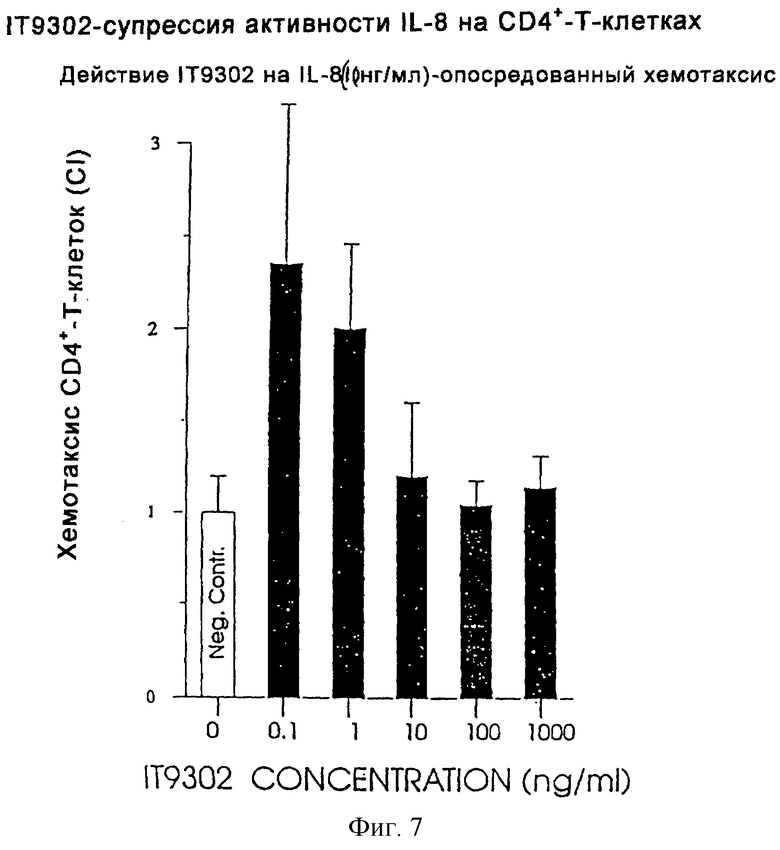
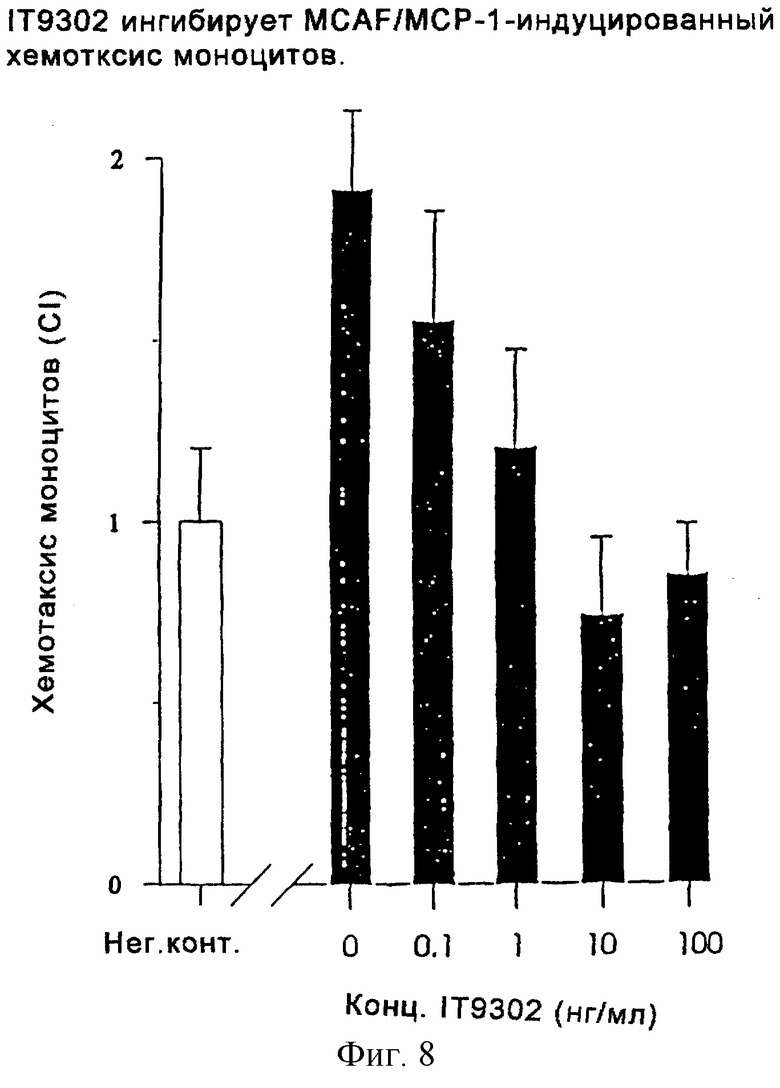
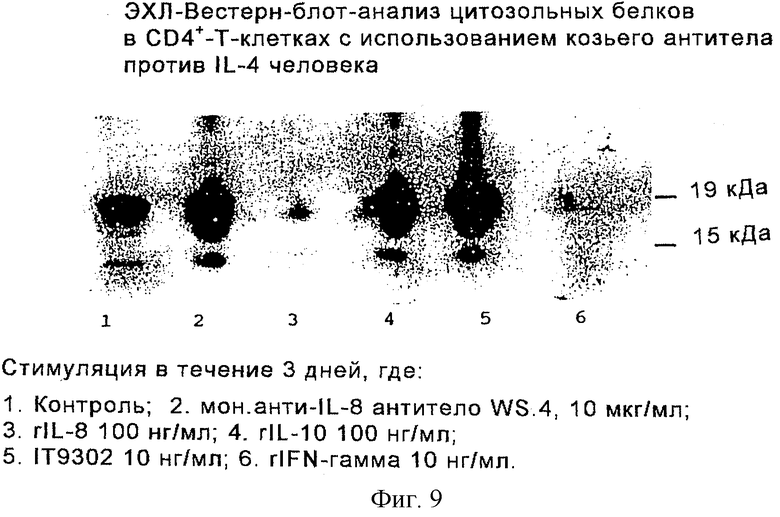
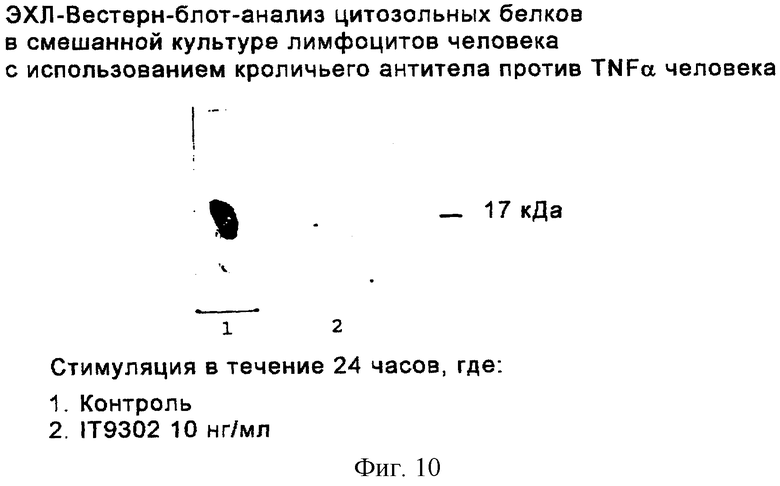

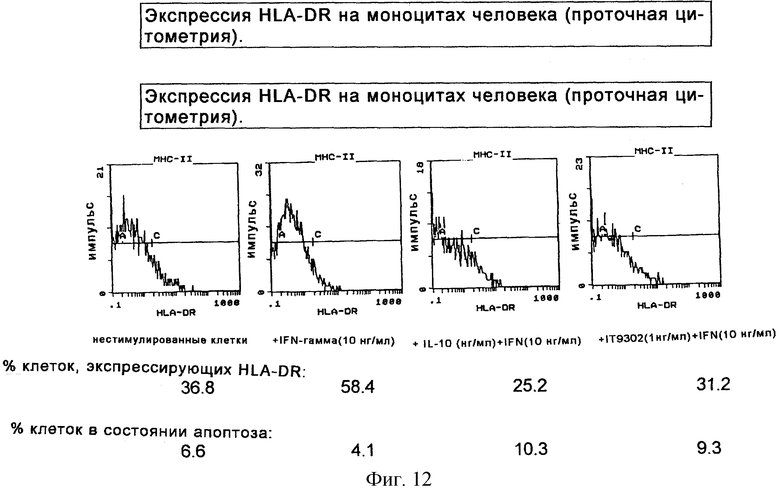
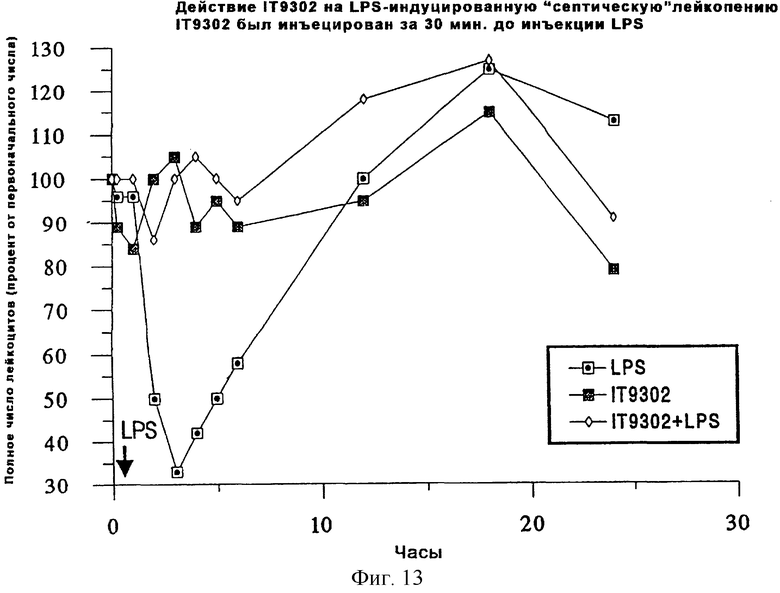
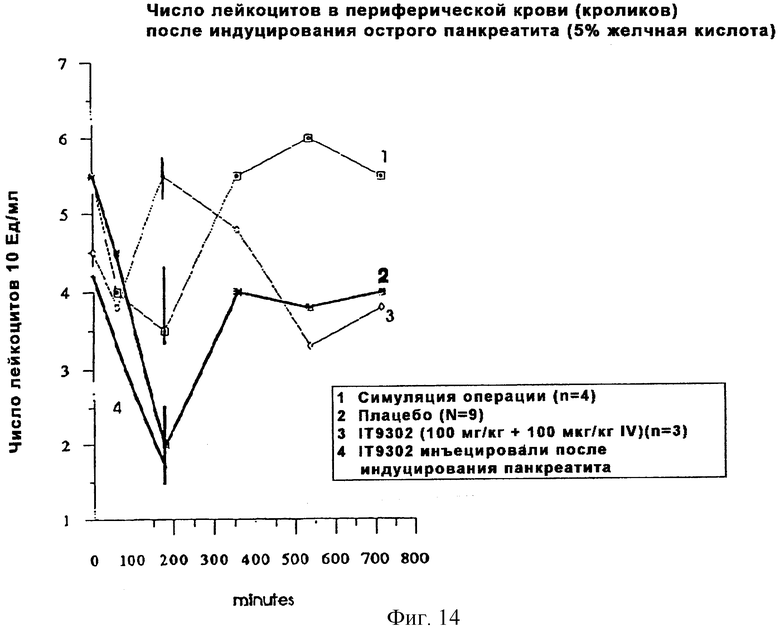
Изобретение относится к полипептидам, включающим следующую последовательность: Thr-X4-Lys-X5-Arg-X6, где X4 и Х5 - независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu. Соединения по изобретению обладают IL-10 - подобной активностью и могут использоваться в качестве агонистов или антагонистов IL-10. Описаны также фармацевтическая композиция, содержащая указанный полипептид, а также способ его получения. 3 с. и 20 з.п.ф-лы, 14 ил., 10 табл.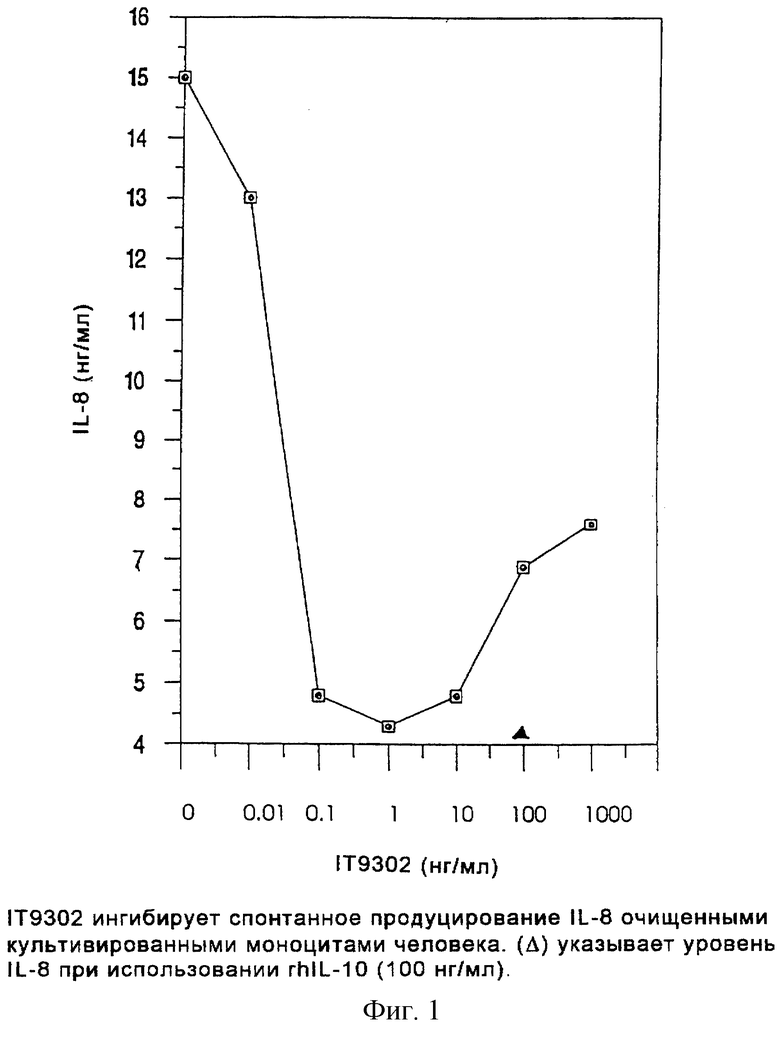
Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 19),
где Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu, где по крайней мере один из Х4, Х5, Х6, Thr, Lys и Arg I) независимо замещен не встречающимися в природе или редкими аминокислотами; и/или II) полипептид является циклизованным; и/или III) полипептид является стабилизированным, и/или IV) амино-концевой аминокислотный остаток является ацилированным, и/или V) карбокси-концевой аминокислотный остаток является амидированным; причем указанный полипептид обладает по крайней мере одним из следующих свойств:
а) индуцирует ингибирование спонтанного продуцирования IL-8 моноцитами человека;
b) индуцирует ингибирование IL-1β-индуцированное продуцирование IL-8 мононуклеарными клетками периферической крови человека (РВМС);
с) индуцирует продуцирование белка, являющегося антагонистом рецептора интерлейкина-1 (IRAP), моноцитами человека;
d) индуцирует хемотаксическую миграцию CD8+-Т-лимфоцитов человека in vitro,
е) десенсибилизирует CD8+-Т-клетки человека, что приводит к их невосприимчивости по отношению к rhIL-10;
f) супрессирует хемотаксический ответ CD4+-Т-лимфоцитов человека по отношению к IL-8;
g) супрессирует хемотаксический ответ моноцитов человека по отношению к MCAF/МCP-1;
h) ингибирует экспрессию молекулы МНС (главного комплекса гистосовместимости) класса II на моноцитах человека, стимулированных IFN-γ;
i) индуцирует продуцирование IL-4 культивированными нормальными CD4+-Т-клетками;
j) снижает продуцирование TNFα в реакции смешанной культуры лейкоцитов человека;
k) осуществляет негативную регуляцию продуцирования IL-8 и TNFα в кроличьей модели острого панкреатита, индуцированного желчной кислотой, а также способствует снижению уровня нейтрофильной инфильтрации в легких обработанных кроликов.
Х3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 20),
где Х3, Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu, где I) по крайней мере один из Х3, Х4, Х5, Х6, Thr, Lys и Arg независимо замещен не встречающимися в природе или редкими аминокислотами; и/или II) полипептид является циклизованным; и/или III) полипептид является стабилизированным, и/или IV) амино-концевой аминокислотный остаток является ацилированным, и/или V) карбокси-концевой аминокислотный остаток является амидированным.
Х2-Х3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 21),
где Х2 представляет Tyr или Phe,
Х3, Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu, где I) по крайней мере один из Х2, Х3, Х4, Х5, Thr, Lys и Arg независимо замещен не встречающимися в природе или редкими аминокислотами; и/или II) полипептид является циклизованным; и/или III) полипептид является стабилизированным, и/или IV) амино-концевой аминокислотный остаток является ацилированным, и/или V) карбокси-концевой аминокислотный остаток является амидированным.
Х1-Х2-Х3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 22),
где Х1 представляет Ala или Gly,
X2 представляет Tyr или Phe,
Х3, Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu, где I) по крайней мере один из Х1, Х2, Х3, Х4, Х5, Х6, Thr, Lys и Arg независимо замещен не встречающимися в природе или редкими аминокислотами; и/или II) полипептид является циклизованным; и/или III) полипептид является стабилизированным, и/или IV) амино-концевой аминокислотный остаток является ацилированным, и/или V) карбокси-концевой аминокислотный остаток является амидированным.
Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 19),
где Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu, где по крайней мере один из Тhr, Lys и Arg независимо замещен не встречающейся в природе или редкой аминокислотой.
Х1-Х2-Х3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 22),
где Х1 представляет Ala или Gly,
Х2 представляет Tyr или Phe,
Х3, Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu, где по крайней мере один из Х1, Х2, Х3, Х4, Х5, Х6, Thr, Lys и Arg независимо замещен не встречающейся в природе или редкой аминокислотой.
Х2-Х3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 21),
где Х2 представляет Tyr или Phe,
Х3, Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, включающей Asn, Asp, Gln и Glu, где по крайней мере один из Х2, Х3, Х4, Х5, Х6, Thr, Lys и Arg независимо замещен не встречающейся в природе или редкой аминокислотой.
Х3-Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 20),
где Х3, Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu, где по крайней мере один из Х3, Х4, Х5, Х6, Thr, Lys и Arg независимо замещен не встречающейся в природе или редкой аминокислотой.
Thr-X4-Lys-X5-Arg-X6(SEQ ID NO: 19),
где Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, состоящей из Asn, Asp, Gln и Glu, где по крайней мере один из Х4, Х5, Х6, Thr, Lys и Arg независимо замещен не встречающейся в природе или редкой аминокислотой.
Ala-Tyr-Met-Thr-Met-Lys-Ile-Arg-Asn (SEQ ID NO: 1),
где 1) по крайней мере одна из аминокислот независимо замещена не встречающейся в природе или редкой аминокислотой; и/или II) полипептид является циклизованным; и/или III) полипептид является стабилизированным; и/или IV) аминоконцевой аминокислотный остаток является ацилированным, и/или V) карбокси-аминокислотный остаток является амидированным.
Thr-X4-Lys-X5-Arg-X6 (SEQ ID NO: 19),
где Х4 и Х5 независимо выбраны из группы, состоящей из Met, Ile, Leu и Val, и
Х6 выбран из группы, включающей из Asn, Asp, Gln и Glu, где сродство связывания полипептида с I1-10 рецептором увеличивается или уменьшается в сравнении с полипептидом, имеющим аминокислотную последовательность
Ala-Tyr-Met-Thr-Met-Lys-Ile-Arg-Asn (SEQ ID NO: 1)
22. Фармацевтическая композиция, содержащая полипептид, по любому из пп. 1-21, в качестве агонистов или антагонистов интерлейкина-10.
а) ковалентное связывание С-концевой аминокислоты, взятой в форме N-альфа-защищенного с необязательно защищенными боковыми цепями реакционноспособного производного, либо непосредственно, либо посредством подходящей линкерной группы с твердым носителем;
b) удаление N-альфа-защитной группы;
с) постадийное присоединение следующих защищенных аминокислот в соответствии с нужной последовательностью;
d) удаление защитных групп, если они имеются, с боковых цепей;
е) после синтеза полной пептидной цепи отщепление этого пептида от смолы; и необязательно
f) циклизация и/или стабилизация пептида, и/или ацилирование амино-концевого аминокислотного остатка, и/или амидирование карбокси-концевого аминокислотного остатка.
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| RU 2051922 С1, 10.01.1996 | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| US 5328989 А, 12.07.1994 | |||
| Benjamin D., Cancer Treat | |||
| Res | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |

