Перекрестная ссылка на связанные заявки
Настоящая заявка заявляет приоритет по патентным заявкам США с серийными номерами 62/346717 (подана 7 июня 2016 года; рассматривается) и 62/432328 (подана 9 декабря 2016 года; рассматривается), каждая из которых включена в настоящий документ ссылкой полностью.
Ссылка на перечень последовательностей:
Эта заявка включает один или несколько перечней последовательностей в соответствии с 37 CFR 1.821 et seq., которые раскрыты на машиночитаемом носителе (название файла: 1301_0145PCT_ST25.txt, создан 19 мая 2017 года и имеет размер 59 748 байт), файл включен в настоящее описание ссылкой во всей своей полноте.
Предшествующий уровень изобретения
Область техники, к которой относится изобретение
Настоящее изобретение направлено на способы применения биспецифических связывающих молекул, которые обладают сайтом связывания, специфичным для эпитопа CD32B, и сайтом связывания, специфичным для эпитопа CD79B, и, таким образом, которые способны одновременно связываться с CD32B и CD79B. Изобретение, в частности, относится к таким молекулам, которые являются биспецифичными антителами или биспецифичными диателами (и особенно такими диателами, которые дополнительно включают домен Fc). Изобретение направлено на применение таких молекул и на применение фармацевтических композиций, которые содержат такие молекулы, при лечении воспалительных заболеваний или состояний.
Описание предшествующего уровня техники
I. Рецепторы Fcγ и CD32B
Взаимодействие комплексов антитело-антиген с клетками иммунной системы приводит к широкому спектру ответов, начиная от эффекторных функций, таких как антителозависимая цитотоксичность, дегрануляция тучных клеток и фагоцитоз, до иммуномодулирующих сигналов, таких как регулирование пролиферации лимфоцитов и секреция антител. Все эти взаимодействия инициируются посредством связывания домена Fc антител или иммунных комплексов со специализированными клеточными рецепторами клеток на гематопоэтических клетках. Разнообразие клеточных ответов, вызванных антителами и иммунными комплексами, обусловлено структурной гетерогенностью рецепторов Fc. Рецепторы Fc имеют общие структурно родственные лиганд-связывающие домены, которые предположительно опосредуют внутриклеточную передачу сигналов.
Рецепторы Fc являются представителями генного суперсемейства иммуноглобулиновых белков. Это поверхностные гликопротеины, которые могут связывать Fc-часть иммуноглобулиновых молекул. Каждый представитель семейства распознает иммуноглобулины одного или нескольких изотипов при помощи домена узнавания на α-цепи рецептора Fc.
Рецепторы Fc определяются их специфичностью к подтипам иммуноглобулинов (см. Ravetch JV et al. (1991) “Fc Receptors”, Annu. Rev. Immunol. 9:457-92; Gerber J.S. et al. (2001) “Stimulatory And Inhibitory Signals Originating From The Macrophage Fcγ Receptors”, Microbes and Infection, 3:131-139; Billadeau D.D. et al. (2002) “ITAMs Versus ITIMs: Striking A Balance During Cell Regulation”, J. Clin. Invest. 2(109):161-168l; Ravetch J.V. et al. (2000) “Immune Inhibitory Receptors”, Science 290:84-89; Ravetch J.V. et al. (2001) “IgG Fc Receptors”, Annu. Rev. Immunol. 19:275-90; Ravetch J.V. (1994) “Fc Receptors: Rubor Redux”, Cell, 78(4): 553-60).
Рецепторы Fc, которые способны связываться с антителами IgG, называются «FcγR». Каждый представитель этого семейства представляет собой интегральный мембранный гликопротеин, обладающий внеклеточными доменами, родственными C2-наборам иммуноглобулин-родственных доменов, одним трансмембранным доменом и внутрицитоплазматическим доменом переменной длины. Существует три известных FcγR, обозначенные как FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16). Три рецептора кодируются разными генами; однако обширные гомологии между тремя представителями семейства позволяют предположить, что они возникли из общего прародителя, возможно, путем дупликации генов.
Белки FcγRII (CD32) представляют собой интегральные мембранные гликопротеины с молекулярной массой 40 кДа, которые связывают только комплексированный IgG из-за низкой аффинности к мономерному Ig (106 М-1). Этот рецептор является наиболее широко экспрессируемым FcγR, присутствующим на всех кроветворных клетках, включая моноциты, макрофаги, B-клетки, NK-клетки, нейтрофилы, тучные клетки и тромбоциты. FcγRII имеет только два иммуноглобулиноподобных участка в своей иммуноглобулин-связывающей цепи и, следовательно, гораздо более низкую аффинность к IgG, чем FcγRI. Существует три гена человеческого FcγRII (FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIC (CD32C)), каждый из которых связывает IgG в агрегатах или иммунных комплексах.
Явные различия в цитоплазматических доменах FcγRIIA и FcγRIIB создают два функционально гетерогенных ответа на рецепторное лигирование. Принципиальное отличие заключается в том, что при связывании с участком Fc из IgG изоформа FcγRIIA инициирует внутриклеточную передачу сигналов, приводящую к активации иммунной системы (например, фагоцитоза, респираторного взрыва и т.д.), тогда как при связывании с участком Fc из IgG изоформа FcγRIIB инициирует сигналы, которые приводят к ослаблению или ингибированию иммунной системы (например, ингибированию активации B-клеток и т.д.).
Такие активирующие и ингибирующие сигналы передаются через FcγR после лигирования с участком Fc IgG. Эти диаметрально противоположные функции являются результатом структурных различий между различными изоформами рецептора. Два разных домена в цитоплазматических сигнальных доменах рецептора, которые называются иммунорецепторными тирозиновыми активирующими мотивами (ITAM) или иммунорецепторными тирозиновыми ингибирующими мотивами (ITIM), объясняют различные ответы. Рекрутирование различных цитоплазматических ферментов в эти структуры диктует результат опосредованных FcγR клеточных ответов. ITAM-содержащие комплексы FcγR включают FcγRI, FcγRIIA, FcγRIIIA, тогда как ITIM-содержащие комплексы включают только FcγRIIB.
Человеческие нейтрофилы экспрессируют ген FcγRIIA. Кластеризация FcγRIIA через иммунные комплексы или специфическое перекрестное связывание антителами служит для агрегации ITAM с рецептор-ассоциированными киназами, которые способствуют фосфорилированию ITAM. Фосфорилирование ITAM служит местом стыковки Syk-киназы, активация которой приводит к активации нижележащих субстратов (например, PI3K). Клеточная активация приводит к высвобождению провоспалительных медиаторов.
Ген FcγRIIB экспрессируется на В-лимфоцитах; его внеклеточный домен на 96% идентичен FcγRIIA и связывает комплексы IgG неразличимым образом. Присутствие ITIM в цитоплазматическом домене FcγRIIB определяет этот ингибирующий подкласс FcγR. Недавно была установлена молекулярная основа этого ингибирования. При совместном лигировании с активирующим FcγR ITIM в FcγRIIB становится фосфорилированным и притягивает домен SH2 инозитполифосфат-5'-фосфатазы (SHIP), который гидролизует мессенджеры-фосфоинозитолы, высвобождаемые в результате активации тирозинкиназы, опосредованной ITAM-содержащим FcγR, следовательно, предотвращая приток внутриклеточного Ca++. Таким образом, такое перекрестное связывание FcγRIIB и активирующего рецептора ослабляет активность активирующего рецептора и, таким образом, ингибирует клеточную респонсивность. Таким образом, в случае В-клеток активация В-клеток, пролиферация В-клеток и секреция антител ослабляются или прерываются. Таким образом, в начале обнаружения антигена происходит связывание мономерного антигена IgG, и области Fc связанных антител связываются с ITAM активирующих FcγR, для обеспечения активации иммунной системы. По мере прогрессирования реакции хозяина образуются мультимерные иммунные комплексы IgG-антиген, которые способны связываться с FcγRIIB (таким образом, связывая такие комплексы с активирующим рецептором), что приводит к ослаблению и окончательному прекращению иммунного ответа (см., например, пат. Соединенных Штатов № 8 445 645; 8 217 147; 8 216 579; 8 216 574; 8 193 318; 192 737; 8 187 593; 8 133 982; 8 044 180; 8 003 774; 7 960 512; 7 786 270; 7 632 497; 7 521 542; 7 425 619; 7 355 008; и публикации пат. Соединенных Штатов № 2012/0276094; 2012/0269811; 2012/0263711; 2012/0219551; 2012/0213781; 2012/0141476; 2011/0305714; 2011/0243941; 2010/0322924; 2010/0254985; 2010/0196362; 2010/0174053; 2009/0202537; 2009/0191195; 2009/0092610; 2009/0076251; 2009/0074771; 2009/0060910; 2009/0053218; 2009/0017027; 2009/0017026; 2009/0017023; 2008/0138349; 2008/0138344; 2008/0131435; 2008/0112961; 2008/0044429; 2008/0044417; 2007/0077246; 2007/0036799; 2007/0014795; 2007/0004909; 2005/0260213; 2005/0215767; 2005/0064514; 2005/0037000; 2004/0185045).
II. В-клеточный рецептор и CD79B
В-клетки - это клетки иммунной системы, которые отвечают за выработку антител. Кроме того, В-клетки презентируют антигены и секретируют цитокины. В-клеточный ответ на антиген является важным компонентом нормальной иммунной системы. В-клетки обладают специализированными рецепторами клеточной поверхности (В-клеточные рецепторы; «BCR»). Если В-клетка встречается с антигеном, способным связываться с BCR этой клетки, В-клетка будет стимулирована для пролиферации и продуцирования антител, специфичных к связанному антигену. Для получения эффективного ответа на антигены также необходимы BCR-ассоциированные белки и помощь T-клеток. Комплекс антиген/BCR интернализуется, и антиген подвергается протеолитической обработке. Небольшая часть антигена остается в комплексе с основными молекулами комплекса гистосовместимости-II («МНС-II») на поверхности В-клеток, где комплекс может распознаваться Т-клетками. Т-клетки, активированные такой презентацией антигена, секретируют CD40L и различные лимфокины, которые вызывают созревание B-клеток.
Передача сигналов через BCR играет важную роль в генерации антител, аутоиммунности и в установлении иммунологической толерантности (Gauld, SB et al. (2002) “B Cell Antigen Receptor Signaling: Roles In Cell Development And Disease”, Science 296(5573):1641-1642). Незрелые В-клетки, которые связывают аутоантигены еще в костном мозге, удаляются путем апоптоза. Напротив, связывание антигена со зрелыми В-клетками приводит к активации, пролиферации, анергии и апоптозу. Конкретный наблюдаемый функциональный ответ зависит от того, получает ли B-клетка костимулирующие сигналы через другие активируемые поверхностные рецепторы и специфические пути передачи сигнала.
BCR состоит из мембранного иммуноглобулина, который вместе с нековалентно связанными α и β субъединицами CD79 («CD79a» и «CD79B» соответственно) образует комплекс BCR. CD79a и CD79B являются субъединицами, передающими сигналы, которые содержат консервативный иммунорецепторный мотив активации на основе тирозина («ITAM»), необходимый для передачи сигнала (Dylke, J. et al. (2007) “Role of the extracellular and transmembrane domain of Ig-alpha/beta in assembly of the B cell antigen receptor (BCR)”, Immunol. Lett. 112(1):47-57; Cambier, J.C. (1995) “New Nomenclature For The Reth Motif (or ARH1/TAM/ARAM/YXXL)”, Immunol. Today 16:110). Агрегация комплекса BCR мультивалентным антигеном инициирует трансфосфорилирование ITAM CD79a и CD79B и активацию рецептор-ассоциированных киназ (DeFranco, A.L. (1997) “The Complexity Of Signaling Pathways Activated By The BCR”, Curr. Opin. Immunol. 9:296-308; Kurosaki, T. (1997) “Molecular Mechanisms In B-Cell Antigen Receptor Signaling”, Curr. Opin. Immunol. 9:309-318; Kim, K.M. et al. (1993) “Signalling Function Of The B-Cell Antigen Receptors”, Immun. Rev. 132:125-146). Фосфорилированные ITAM привлекают дополнительные эффекторы, такие как PI3K, PLC-γ и участники пути Ras/MAPK. Эти сигнальные события ответственны как за пролиферацию B-клеток, так и за повышенную экспрессию маркеров активации (таких как MHC-II и CD86), которые необходимы для инициации B-клеток для их последующих взаимодействий с T-хелперными («Th») клетками.
III. Воспалительные заболевания или состояния
Воспаление - это процесс, посредством которого лейкоциты и химические вещества организма защищают наш организм от заражения инородными веществами, такими как бактерии и вирусы. Обычно этот процесс характеризуется болью, припухлостью, горячностью и покраснением пораженного участка. Химические вещества, известные как цитокины и простагландины, контролируют этот процесс и высвобождаются в упорядоченном и самоограничивающемся каскаде в кровь или пораженные ткани. Этот выброс химических веществ увеличивает приток крови к области травмы или инфекции и может привести к покраснению и горячности. Некоторые из химических веществ вызывают утечку жидкости в ткани, что приводит к припухлости. Этот защитный процесс может стимулировать нервы и вызывать боль. Эти изменения, происходящие в течение ограниченного периода времени в соответствующей области, приносят пользу организму.
Воспалительные заболевания или состояния отражают атаку иммунной системы на собственные клетки и ткани организма (т.е. «аутоиммунный» ответ). Есть много различных аутоиммунных расстройств, которые по-разному влияют на организм. Например, мозг поражен у людей с рассеянным склерозом, кишечник поражен у людей с болезнью Крона, а синовия, кость и хрящ различных суставов - у людей с ревматоидным артритом. По мере того как аутоиммунные нарушения приводят к разрушению одного или нескольких типов тканей организма, это может привести к аномальному росту органа или изменениям в функции органа. Аутоиммунное расстройство может затрагивать только один орган или тип ткани или может затрагивать несколько органов и тканей. Органы и ткани, которые обычно подвержены аутоиммунным нарушениям, включают эритроциты, кровеносные сосуды, соединительные ткани, эндокринные железы (например, щитовидную железу или поджелудочную железу), мышцы, суставы и кожу. Примеры аутоиммунных расстройств включают, без ограничения указанным, болезнь Аддисона, аутоиммунный гепатит, аутоиммунное заболевание внутреннего уха, миастению, болезнь Крона, дерматомиозит, семейный аденоматозный полипоз, болезнь трансплантат против хозяина (GvHD), болезнь Грейвса, тиреоидит Хашимото, красную волчанку, рассеянный склероз (MS); пернициозную анемию, синдром Рейтера, ревматоидный артрит (RA), синдром Шегрена, системную красную волчанку (SLE), диабет 1 типа, первичный васкулит (например, ревматическую полимиалгию, гигантоклеточный артериит, болезнь Бехчета), пемфигус, оптикомиелит, анти-NMDA-рецепторный энцефалит, синдром Гийена-Барре, хроническую воспалительную демиелинизирующую полиневропатию (CIDP), офтальмопатию Грейвса, связанные с IgG4 заболевания, идиопатическую тромбоцитопеническую пурпуру (ITP) и язвенный колит.
Воспалительные заболевания или состояния могут также возникать, когда обычно защитная иммунная система организма наносит ущерб путем нападения на инородные клетки или ткани, присутствие которых полезно для организма (например, отторжение трансплантатов (болезнь хозяин против хозяина)) или из-за отторжения клеток иммуносупрессированного хозяина иммунокомпетентными клетками введенного трансплантата (болезнь трансплантат против хозяина) (DePaoli, AM et al. (1992) “Graft-Versus-Host Disease And Liver Transplantation,” Ann. Intern. Med. 117:170-171; Sudhindran, S. et al. (2003) “Treatment Of Graft-Versus-Host Disease After Liver Transplantation With Basiliximab Followed By Bowel Resection,” Am J Transplant. 3:1024-1029; Pollack, M.S. et al. (2005) “Severe, Late-Onset Graft-Versus-Host Disease In A Liver Transplant Recipient Documented By Chimerism Analysis,” Hum. Immunol. 66:28-31; Perri, R. et al. (2007) “Graft Vs. Host Disease After Liver Transplantation: A New Approach Is Needed,” Liver Transpl. 13:1092-1099; Mawad, R. et al. (2009) “Graft-Versus-Host Disease Presenting With Pancytopenia After En Bloc Multiorgan Transplantation: Case Report And Literature Review,” Transplant Proc. 41:4431-4433; Akbulut, S. et al. (2012) “Graft-Versus-Host Disease After Liver Transplantation: A Comprehensive Literature Review,” World J. Gastroenterol. 18(37): 5240-5248).
Несмотря на последние достижения в лечении таких заболеваний или состояний, по-прежнему существует потребность в композициях, способных лечить или предотвращать воспалительные заболевания или состояния.
IV. Биспецифичные связывающие молекулы
A. Биспецифические антитела
Способность немодифицированного природного антитела (например, IgG) связывать эпитоп антигена зависит от присутствия и взаимодействия вариабельных доменов на легкой и тяжелой цепях иммуноглобулина (т.е. его вариабельного домена легкой цепи (домен VL) и его вариабельного домена тяжелой цепи (домен VH) для формирования эпитопсвязывающих сайтов антитела. Вследствие присутствия только одного вида легкой цепи и одного вида тяжелой цепи природные антитела способны связываться только с одним видом эпитопа (т.е. они являются моноспецифичными), хотя они могут связывать несколько копий этого вида (т. е. демонстрировать двухвалентность или мультивалентность).
Однако, в данной области техники удалось получить биспецифичные антитела, например, путем совместной экспрессии двух пар тяжелой цепи и легкой цепи иммуноглобулина, имеющих различную эпитопную специфичность, с последующей очисткой искомой молекулы с помощью аффинной хроматографии, как описано Milstein et al., (1983) “Hybrid Hybridomas And Their Use In Immunohistochemistry,” Nature 305:537-39, WO 93/08829. В другом подходе вариабельные домены антител с искомой специфичностью связывания (сайты объединения антитело-антиген) были слиты с последовательностями константного домена иммуноглобулина, например, константного домена тяжелой цепи, включающего, по меньшей мере, часть шарнирных областей, областей CH2 и CH3. Нуклеиновые кислоты, кодирующие эти слияния, могут быть вставлены в один и тот же или разные экспрессирующие векторы и экспрессироваться в подходящем организме-хозяине. Биспецифичные антитела рассматриваются, например, в Traunecker et al. (1991) “Bispecific Single Chain Molecules (Janusins) Target Cytotoxic Lymphocytes On HIV Infected Cells,” EMBO J. 10:3655-3659; Zhukovsky, E.A. et al. (2016) “Bispecific Antibodies And Cars: Generalized Immunotherapeutics Harnessing T Cell Redirection,” Curr. Opin. Immunol. 40:24-35; Kiefer, J.D. et al. (2016) “Immunocytokines And Bispecific Antibodies: Two Complementary Strategies For The Selective Activation Of Immune Cells At The Tumor Site,” Immunol. Rev. 270(1):178-192; Solimando, A.G. et al. (2016) “Targeting B-Cell Non-Hodgkin Lymphoma: New And Old Tricks,” Leuk. Res. 42:93-104; Fan, G. et al. (2015) “Bispecific Antibodies And Their Applications,” J. Hematol. Oncol. 8:130; Grandjenette, C. et al. (2015) “Bispecific Antibodies: An Innovative Arsenal To Hunt, Grab And Destroy Cancer Cells,” Curr. Pharm. Biotechnol. 16(8):670-683; Nuñez-Prado, N. et al. (2015) “The Coming Of Age Of Engineered Multivalent Antibodies,” Drug Discov. Today 20(5):588-594; and Kontermann, R.E. et al. (2015) “Bispecific Antibodies,” Drug. Discov. Today 20(7):838-847.
В дополнение к интактным биспецифическим антителам в данной области разработаны производные биспецифических одноцепочечных антител (например, привлекающие T-клетки биспецифические активаторы (BiTE)), которые состоят из одной полипептидной цепи, имеющей домен VL и VH для первой связывающей молекулы и домен VL и VH для второй связывающей молекулы (например, патенты США № 7 112 324 7 235 641 7 575 923 7 919 089; Wu, J. et al. (2015) “Blinatumomab: A Bispecific T Cell Engager (BiTe) Antibody Against CD19/CD3 For Refractory Acute Lymphoid Leukemia,” J. Hematol. Oncol. 8:104; Lutterbuese, R. et al. (2008) “Conversion Of Cetuximab, Panitumumab, Trastuzumab And Omalizumab Into T-Cell-Engaging BiTE Antibodies Creates Novel Drug Candidates Of High Potency,” Proc. Am. Assoc. Cancer Res 99:Abs 2402; Baeuerle, P.A. et al. (2009) “Bispecific T-Cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944.
B. Биспецифичные диатела
В данной области анонсирована возможность продуцирования диател, которые отличаются от природных антител, способных связывать два или более различных видов эпитопов (т.е. проявлять биспецифичность или мультиспецифичность в дополнение к двухвалентности или мультивалентности). Конструкция диатела основана на одноцепочечной конструкции Fv (scFv), которая имеет домен VL и соответствующий домен VH, разделенные промежуточным линкером, который позволяет таким доменам взаимодействовать друг с другом. Когда такое взаимодействие доменов VL и VH становится невозможным из-за применения линкера недостаточной длины (менее чем около 12 аминокислотных остатков), две такие конструкции scFv могут взаимодействовать друг с другом с образованием молекулы двухвалентного диатела, в которой домен VL одной цепи ассоциируется с доменом VH другой (рассматривается в Marvin et al. (2005) “Recombinant Approaches To IgG-Like Bispecific Antibodies,” Acta Pharmacol. Sin. 26:649-658, Holliger et al. (1993) “’Diabodies’: Small Bivalent And Bispecific Antibody Fragments,” Proc. Natl. Acad. Sci. (U.S.A.) 90:6444-6448; US 2004/0058400 (Holliger et al.); US 2004/0220388 (Mertens et al.) ; Mertens, N. et al., “New Recombinant Bi- and Trispecific Antibody Derivatives,” In: Novel Frontiers In The Production Of Compounds For Biomedical Use, A. VanBroekhoven et al. (Eds.), Kluwer Academic Publishers, Dordrecht, The Netherlands (2001), pages 195-208; Alt et al. (1999) FEBS Lett. 454(1-2):90-94; Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672; WO 02/02781 (Mertens et al.); Olafsen, T. et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Protein Eng. Des. Sel. 17(1):21-27; Wu, A. et al. (2001) “Multimerization Of A Chimeric Anti-CD20 Single Chain Fv-Fc Fusion Protein Is Mediated Through Variable Domain Exchange,” Protein Engineering 14(2):1025-1033; Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Region,” Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588; Baeuerle, P.A. et al. (2009) “Bispecific T-Cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944).
Получение немоноспецифических диател обеспечивает существенное преимущество: способность колигировать и колокализовать клетки, которые экспрессируют различные эпитопы. Таким образом, двухвалентные диатела имеют широкое применение, включая терапию и иммунодиагностику. Бивалентность обеспечивает большую гибкость в дизайне и разработке диатела в различных применениях, обеспечивая повышенную авидность к мультимерным антигенам, сшивание различных антигенов и направленное нацеливание на конкретные типы клеток, основанное на присутствии обоих целевых антигенов. Из-за их повышенной валентности, низких скоростей диссоциации и быстрого клиренса из кровотока (для небольших размеров, около или ниже ~50 кДа), молекулы диател, известные в данной области, также показали особое применение в области формирования опухоли (Fitzgerald et al. (1997) “Improved Tumour Targeting By Disulphide Stabilized Diabodies Expressed In Pichia pastoris,” Protein Eng. 10:1221-1225). Особое значение имеет колигирование различных клеток, например, сшивание цитотоксических Т-клеток с опухолевыми клетками (Staerz et al. (1985) “Hybrid Antibodies Can Target Sites For Attack By T Cells,” Nature 314:628-631, and Holliger et al. (1996) “Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody,” Protein Eng. 9:299-305).
Эпитопсвязывающие домены диатела также могут быть направлены на поверхностную детерминанту любой иммунной эффекторной клетки, такой как CD3, CD16, CD32 или CD64, которые экспрессируются на Т-лимфоцитах, клетках естественных киллерах (NK) или других мононуклеарных клетках. Во многих исследованиях было также обнаружено, что связывание диатела с детерминантами эффекторных клеток, например, рецепторами Fcγ (FcγR), активирует эффекторные клетки (Holliger et al. (1996) “Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody,” Protein Eng. 9:299-305; Holliger et al. (1999) “Carcinoembryonic Antigen (CEA)-Specific T-cell Activation In Colon Carcinoma Induced By Anti-CD3 × Anti-CEA Bispecific Diabodies And B7 × Anti-CEA Bispecific Fusion Proteins,” Cancer Res. 59:2909-2916; WO 2006/113665; WO 2008/157379; WO 2010/080538; WO 2012/018687; WO 2012/162068). Обычно активация эффекторных клеток запускается связыванием антигенсвязанного антитела с эффекторной клеткой посредством взаимодействия Fc-FcγR; таким образом, в этом отношении молекулы диатела по изобретению могут проявлять Ig-подобную функциональность независимо от того, содержат ли они домен Fc (например, выявляемый в любом анализе эффекторной функции, известном в данной области, или представленном в качестве примера в настоящем документе (например, в анализе ADCC)). Путем сшивки опухолевых и эффекторных клеток диатело не только помещает эффекторные клетки в непосредственную близость к опухолевым клеткам, но и приводит к эффективному уничтожению опухоли (см., например, Cao et al. (2003) “Bispecific Antibody Conjugates In Therapeutics,” Adv. Drug. Deliv. Rev. 55:171-197).
Однако вышеупомянутые преимущества имеют значительную цену. Формирование таких немоноспецифических диател требует успешной сборки двух или более различных и разных полипептидов (т.е. такое формирование требует, чтобы диатела образовывались путем гетеродимеризации различных видов полипептидной цепи). Этот факт контрастирует с моноспецифическими диателами, которые образуются путем гомодимеризации идентичных полипептидных цепей. Поскольку, по меньшей мере, два разнородных полипептида (т.е. два вида полипептидов) должны быть предоставлены для образования немоноспецифического диатела, а также потому, что гомодимеризация таких полипептидов приводит к образованию неактивных молекул (Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588), получение таких полипептидов должно осуществляться таким образом, чтобы предотвратить ковалентную связь между полипептидами одного и того же вида (Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588). Таким образом, в данной области сообщается о нековалентной ассоциации таких полипептидов (см., например, Olafsen et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Prot. Engr. Des. Sel. 17:21-27; Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain,” Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588; Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672).
Однако, в данной области признается, что биспецифические диатела, состоящие из нековалентно связанных полипептидов, являются неустойчивыми и легко диссоциируют на нефункциональные мономеры (см., например, Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672).
Ввиду этой проблемы в данной области удалось разработать стабильные, ковалентно связанные гетеродимерные немоноспецифические диатела (см., например, WO 2006/113665; WO 2008/157379; WO 2010/080538; WO 2012/018687; WO 2012/162068; Johnson, S. et al. (2010) “Effector Cell Recruitment With Novel Fv-Based Dual-Affinity Re-Targeting Protein Leads To Potent Tumor Cytolysis And In Vivo B-Cell Depletion,” J. Molec. Biol. 399(3):436-449; Veri, M.C. et al. (2010) “Therapeutic Control Of B Cell Activation Via Recruitment Of Fcgamma Receptor IIb (CD32B) Inhibitory Function With A Novel Bispecific Antibody Scaffold,” Arthritis Rheum. 62(7):1933-1943; Moore, P.A. et al. (2011) “Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T-Cell Killing Of B-Cell Lymphoma,” Blood 117(17):4542-4551; Chen, X. et al. (2016) “Mechanistic Projection Of First In Human Dose For Bispecific Immuno-Modulatory P-Cadherin LP-DART - An Integrated PK/PD Modeling Approach,” Clin. Pharmacol. Ther. doi: 10.1002/cpt.393; Tsai, P. et al. (2016) “CD19xCD3 DART Protein Mediates Human B-Cell Depletion In Vivo In Humanized BLT Mice,” Mol. Ther. Oncolytics. 3:15024; Root et al. (2016) “Development of PF-06671008, a Highly Potent Anti-P-cadherin/Anti-CD3 Bispecific DART Molecule with Extended Half-Life for the Treatment of Cancer,” Antibodies 5:6; Sloan, D.D. et al. (2015) “Targeting HIV Reservoir in Infected CD4 T Cells by Dual-Affinity Re-targeting Molecules (DARTs) that Bind HIV Envelope and Recruit Cytotoxic T Cells,” PLoS Pathog. 11(11):e1005233; Al-Hussaini, M. et al. (2016) “Targeting CD123 In Acute Myeloid Leukemia Using A T-Cell-Directed Dual-Affinity Retargeting Platform,” Blood 127(1):122-131); Chichili, G.R. et al. (2015) “A CD3xCD123 Bispecific DART For Redirecting Host T Cells To Myelogenous Leukemia: Preclinical Activity And Safety In Nonhuman Primates,” Sci. Transl. Med. 7(289):289ra82; Zanin, M. et al. (2015) “An Anti-H5N1 Influenza Virus FcDART Antibody Is A Highly Efficacious Therapeutic Agent And Prophylactic Against H5N1 Influenza Virus Infection,” J. Virol. 89(8):4549-4561). Такие подходы включают создание одного или нескольких остатков цистеина в каждой из используемых полипептидных соединений. Например, было показано, что добавление остатка цистеина к С-концу таких конструкций позволяет образовать дисульфидную связь между полипептидными цепями, стабилизируя полученный гетеродимер, при этом не влияя на характеристики связывания двухвалентной молекулы.
Опираясь на такой успех, в данной области было создано MGD010, биспецифичное, двухвалентное диатело DART®, которое колигирует ингибирующий Fcγ-рецептор IIb (CD32B) и компонент B-клеточного рецептора (BCR), CD79B, на B-клетках, тем самым одновременно связывая CD32B и CD79B (Chen, W. (2014) “Development Of Human B-Lymphocyte Targeted Bi-Specific DART® Molecules For The Treatment Of Autoimmune Disorders,” J. Immunol. 192(1 Supp.):200.9) (фигура 1A). Настоящее изобретение относится к улучшенным способам применения и введения MGD010 и других биспецифичных молекул CD32B × CD79B, в частности, таких биспецифичных молекул, которые включают домен Fc.
Сущность изобретения:
Настоящее изобретение направлено на способы применения биспецифических связывающих молекул, которые обладают сайтом связывания, специфичным для эпитопа CD32B, и сайтом связывания, специфичным для эпитопа CD79B, и, таким образом, способны одновременно связываться с CD32B и CD79B. Изобретение, в частности, относится к таким молекулам, которые представляют собой биспецифичные антитела (т.е. «антитела CD32B × CD79B») или биспецифичные диатела (т.е. «диатела CD32B × CD79B»), и особенно такие диатела, которые дополнительно включают домен Fc (т.е. «Fc-диатела CD32B × CD79B»). Изобретение направлено на применение таких молекул и на применение фармацевтических композиций, которые включают такие молекулы, при лечении воспалительных заболеваний или состояний.
В развёрнутой форме изобретение относится к способу лечения воспалительного заболевания или состояния, который включает введение терапевтически эффективного количества связывающей молекулы CD32B × CD79B объекту, нуждающемуся в этом, где связывающая молекула CD32B × CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B, и где связывающую молекулу CD32B × CD79B вводят в дозе от около 3 до около 30 мг/кг и в режиме дозирования от одной дозы в неделю до одной дозы в 8 недель.
Изобретение, кроме того, относится к способу снижения или ингибирования иммунного ответа, который включает введение терапевтически эффективного количества связывающей молекулы CD32B × CD79B объекту, нуждающемуся в этом, где связывающая молекула CD32B × CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B, и в котором связывающая молекула CD32B × CD79B вводится в дозе от около 3 мг/кг до около 30 мг/кг и в режиме дозирования от одной дозы в неделю до одной дозы в 8 недель.
Изобретение также относится к воплощениям таких способов, в которых связывающая молекула CD32B × CD79B вводится в дозе около 3 мг/кг, где связывающая молекула CD32B × CD79B вводится в дозе около 10 мг/кг или где связывающая молекула CD32B × CD79B вводится в дозе около 30 мг/кг.
Изобретение также относится к воплощениям таких способов, в которых режим дозирования составляет одну дозу за 2 недели (Q2W), где режим дозирования составляет одну дозу за 3 недели (Q3W) или где режим дозирования составляет одну дозу за 4 недели (Q4W)).
Изобретение также относится к воплощениям таких способов, в которых связывающая молекула CD32B × CD79B представляет собой биспецифичное антитело, которое связывает эпитоп CD32B и эпитоп CD79B, или молекулу, которая включает CD32B- и CD79B-связывающие домены биспецифического антитела.
Изобретение также относится к воплощениям таких способов, в которых связывающая молекула CD32B × CD79B представляет собой биспецифичное диатело CD32B × CD79B, которое связывает эпитоп CD32B и эпитоп CD79B, и, в частности, где биспецифическое диатело CD32B x CD79B представляет собой биспецифичное Fc-диатело CD32B × CD79B.
Изобретение также относится к воплощениям таких способов, в которых воспалительное заболевание или состояние представляет собой аутоиммунное заболевание и, в частности, где аутоиммунное заболевание выбрано из группы, которая включает: болезнь Аддисона, аутоиммунный гепатит, аутоиммунное заболевание внутреннего уха, миастению, болезнь Крона, дерматомиозит, семейный аденоматозный полипоз, болезнь трансплантат против хозяина (GvHD), болезнь Грейвса, тиреоидит Хашимото, красную волчанку, рассеянный склероз (MS); пернициозную анемию, синдром Рейтера, ревматоидный артрит (RA), синдром Шегрена, системную красную волчанку (SLE), диабет 1 типа, первичный васкулит (например, ревматическую полимиалгию, гигантоклеточный артериит, болезнь Бехчета), пемфигус, оптикомиелит, анти-NMDA-рецепторный энцефалит, синдром Гийена-Барре, хроническую воспалительную демиелинизирующую полиневропатию (CIDP), офтальмопатию Грейвса, связанные с IgG4 заболевания, идиопатическую тромбоцитопеническую пурпуру (ITP) и язвенный колит. Изобретение особенно касается воплощений таких способов, в которых воспалительное заболевание или состояние представляет собой GvHD, MS, RA или SLE.
Изобретение также относится к воплощениям таких способов, в которых уровень иммуноглобулина в сыворотке снижается на 36 день после введения первой дозы связывающей молекулы CD32B × CD79B. Изобретение особенно касается воплощений таких способов, где иммуноглобулин представляет собой IgM, IgA или IgG.
Изобретение также касается воплощений таких способов, в которых BCR-опосредованная активация периферических B-клеток ингибируется через 24 часа после введения однократной дозы связывающей молекулы CD32B × CD79B, где активация B-клеток определяется ex vivo анализом мобилизации кальция. Изобретение, в частности, касается воплощений таких способов, в которых BCR-опосредованная активация B-клеток ингибируется, по меньшей мере, на 50%, и где ингибирование поддерживается в течение, по меньшей мере, 6 дней.
Изобретение также относится к воплощениям таких способов, в которых, по меньшей мере, 20% сайтов связывания CD32B × CD79B на периферической B-клетке заняты через 6 часов после введения первой дозы связывающей молекулы CD32B × CD79B.
Изобретение также относится к воплощениям таких способов, в которых объектом является человек.
Изобретение особенно касается воплощений всех таких способов, в которых связывающая молекула CD32B × CD79B содержит:
(A) домен VLCD32B, который включает аминокислотную последовательность SEQ ID NO: 30;
(B) домен VHCD32B, который включает аминокислотную последовательность SEQ ID NO: 31;
(C) домен VLCD79B, который включает аминокислотную последовательность SEQ ID NO: 32;
(D) домен VHCD79B, который включает аминокислотную последовательность SEQ ID NO: 33.
Изобретение также относится к воплощениям таких способов, в которых Fc-диатело CD32B × CD79B включает:
(А) первую полипептидную цепь, которая включает аминокислотную последовательность SEQ ID NO: 39;
(B) вторую полипептиднцю цепь, которая включает аминокислотную последовательность SEQ ID NO: 41;
(С) третью полипептидную цепь, которая включает аминокислотную последовательность SEQ ID NO: 44.
Краткое описание чертежей
На фигуре 1 представлена схема типичного ковалентно связанного диатела, имеющего два эпитопсвязывающих сайта, состоящих из двух полипептидных цепей, каждая из которых имеет E-спираль или K-спираль, способствующие гетеродимеризации (ниже приведены альтернативные домены, способствующие гетеродимеризации). Остаток цистеина может присутствовать в линкере и/или в домене, способствующем образованию гетеродимера, как показано на фигурах 3A/3B. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с применением единой схемы штриховки или заливки.
На фигуре 2 представлена схема типичной ковалентно связанной молекулы диатела, имеющей два эпитопсвязывающих сайта, состоящих из двух полипептидных цепей, каждый из которых имеет домен CH2 и CH3, так что связанные цепи образуют весь или часть домена Fc. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с применением единой схемы штриховки или заливки.
На фигурах 3А-3Е представлены схемы типичной молекулы ковалентно связанного диатела, включающего два эпитопсвязывающих участка, состоящих из трех полипептидных цепей. Показаны две ориентации доменов CH2-CH3 (фигуры 3A/3B и фигуры 3C/3D). Две полипептидные цепи обладают доменами CH2 и CH3, так что связанные цепи образуют весь или часть домена Fc. Полипептидные цепи, содержащие домены VL и VH, дополнительно включают домен, способствующий гетеродимеризации, и ковалентно связаны друг с другом через дисульфидную связь, образованную между остатками цистеина, присутствующими в линкере (фигуры 3А и 3С) или в домене, способствующем гетеродимеризации (фигуры 3D и 3B). Фигура 3E иллюстрирует структуру и функцию примерного Fc-диатела CD32B x CD79B, имеющего ориентацию доменов, показанных на фигуре 3A. Диатело, показанное на фигуре 3E, представляет собой ковалентно связанный комплекс, который включает три полипептидные цепи диатела, представленного на фигуре 3A, но без необязательно присутствующего домена, способствующего гетеродимеризации. Комплекс содержит домен Fc, который включает домен тяжелой цепи CH2 и CH3 из IgG, и домены связывания, специфичные для CD32B и для CD79B. Домены VL и VH, которые распознают один и тот же эпитоп, показаны с применением единой схемы штриховки или заливки.
Фигура 4 иллюстрирует примерный механизм, посредством которого диатела по настоящему изобретению могут опосредовать ингибирование иммунной системы. Как показано на фигуре, диатело по изобретению способно одновременно связываться с молекулой CD79B из BCR и с молекулой CD32B из B-клетки, таким образом, связывая такие молекулы друг с другом. Такое колигирование служит для фософрилирования ITIM молекулы CD32B и привлечения домена SH2 инозит-полифосфат-5'-фосфатазы (SHIP), которая гидролизует мессенджеры-фосфоинозитолы, высвобождаемые в результате ITAM-опосредованной активации тирозинкиназы. Такой гидролиз ингибирует сигнал активации ITAM и, тем самым служит, для ослабления активации B-клеток.
На фигуре 5 показана способность предпочтительного диатела CD32B × CD79B Fc снижать ксеногенный GvHD in vivo на мышиной модели.
На фигуре 6 показана фармакокинетика in vivo типичной связывающей молекулы CD32B × CD79B при введении людям.
Фигура 7 суммирует ex vivo анализ проточной цитометрией связывания in vivo с периферическими В-клетками типичной связывающей молекулы CD32B × CD79B при введении объектам-людям в течение курса исследования.
На фигурах 8A-8D показаны периферические популяции В- и Т-клеток после введения примерной связывающей молекулы CD32B × CD79B людям в течение курса исследования, что определено анализом проточной цитометрии ex vivo.
На фигурах 9A-9D показаны способы и результаты исследований функции B-клеток. Фигуры 9А-9В иллюстрируют экспериментальную процедуру ex vivo и способы анализа данных теста мобилизации кальция ex vivo, который был использован для оценки функции В-клеток у реципиентов примерной связывающей молекулы CD32B × CD79B. Фигуры 9C-9D показывают уменьшение пикового ответа (фигура 9C) и устойчивое снижение общего отклика, измеренного по площади под кривой (AUC; фигура 9D), после введения примерной связывающей молекулы CD32B × CD79B людям в ходе исследования.
На фигурах 10A-10C показано, что введение связывающих молекул CD32B × CD79B подавляет экспрессию BCR на B-клетках памяти CD27+ людей в ходе исследования. Фигура 10А: мембраносвязанный IgG (mIgG); фигура 10В: мембраносвязанный IgM; фигура 10С: мембраносвязанный IgD (mIgD). Данные представлены как Среднее ± SEM.
На фигурах 11A-11C показано, что введение связывающих молекул CD32B × CD79B подавляет экспрессию BCR на CD27- наивных B-клетках людей в ходе исследования. Фигура 11А: мембраносвязанный IgD (mIgD); фигура 11В: мембраносвязанный IgM; фигура 11С: процентное изменение мембраносвязанного IgM (mIgM). Уровни мембраносвязанного иммуноглобулина определяли проточной цитометрией. Данные представлены как Среднее ± SEM.
На фигурах 12A-12C показано, что введение связывающих молекул CD32B × CD79B модулирует уровни сывороточного Ig у людей в течение курса исследования. Фигура 12А: сывороточный IgM; фигура 12В: сывороточный IgA; фигура 12С: сывороточный IgG. Уровни иммуноглобулина IgA, IgG и IgM в сыворотке определяли методом тИФА. Данные представлены как Среднее ± SEM.
На фигуре 13 показано, что введение связывающих молекул CD32B × CD79B снижает уровень молекулы костимуляции CD40, что определяется с помощью анализа проточной цитометрии ex vivo поверхностных костимулирующих молекул периферических В-клеток. Данные представлены как Среднее ± SEM.
На фигурах 14A-14B показаны данные (в двух разных диапазонах концентраций) для исследования ex vivo Emax PK/PD связывания B-клеток примерного Fc-диатела CD32B × CD79B из Примера 1. Данные были графически оценены на линейно-линейной и логарифмической линейной шкале.
На фигурах 15A-15F показаны доклинические целевые концентрации с наложенными фармакокинетическими профилями связывающей молекулы CD32B × CD79B у людей для определения доз, которые достигли бы целевых концентраций. На фигурах 15A-15F оси Y обозначают концентрацию связывающей молекулы CD32B × CD79B [нг/мл], а оси X обозначают время в часах, верхняя, средняя и нижняя горизонтальные линии представляют собой, соответственно, значения концентрации связывающей молекулы для исследования связывания/ингибирования В-клеток in vitro, значения концентрации связывающей молекулы для ЕС50 связывания В-клеток в текущем исследовании, и значения концентрации связывающей молекулы для in vivo ингибирования IgG и IgM на гуманизированной мышиной модели.
Фигуры 16A-16D показывают моделирование средней концентрации связывающей молекулы CD32B × CD79B в дозах 0,3 мг/кг (фигура 16A), 1 мг/кг (фигура 16B), 3 мг/кг (фигура 16C) и 10 мг/кг (фигура 16D) массы тела объекта для режимов дозирования один раз в 2 недели (Q2W), один раз в 3 недели (Q3W) и один раз в 4 недели (Q4W). Были использованы фактические моменты времени и концентрации и номинальные дозы. На фигурах 16A-16D оси Y обозначают концентрацию связывающей молекулы CD32B × CD79B [нг/мл], а оси X - время в часах.
Фигуры 17A-17D показывают спрогнозированную изменчивость (с SD) в смоделированных профилях фигур 16A-16D.
На фигуре 18 показана концентрация HAV-специфического IgG, присутствующего в сыворотке здоровых людей на 57-й день после вакцинации HAV. Эти данные показывают, что введение связывающих молекул CD32B × CD79B снижает HAV-специфические уровни IgG у людей, вакцинированных HAV.
На фигуре 19 показано, что связывающие молекулы CD32B × CD79B блокируют CD40-зависимые B-клеточные ответы, что определяется путем детекции in vitro CD40-зависимой секреции IgG B-клетками. Человеческие В-клетки культивировали со стимуляторами или без них (CD40-лиганд (500 нг/мл), IL-4 (100 нг/мл) и IL-21 (20 нг/мл)), применяя в неразбавленном виде или в серии из 3 раз разведений (1, 1/3, 1/9 и 1/27) в присутствии или в отсутствие примерной связывающей молекулы CD32B × CD79B (20 мкг/мл) в течение 5 дней и определяли секретируемый IgG анализом тИФА.
Подробное описание изобретения
Настоящее изобретение направлено на способы применения биспецифических связывающих молекул, которые обладают сайтом связывания, специфичным для эпитопа CD32B, и сайтом связывания, специфичным для эпитопа CD79B, и, таким образом, способны одновременно связываться с CD32B и CD79B. Изобретение, в частности, относится к таким молекулам, которые представляют собой биспецифичные антитела (т.е. «антитела CD32B × CD79B») или биспецифичные диатела (т.е. «диатела CD32B × CD79B»), и особенно такие диатела, которые дополнительно включают домен Fc (т.е. «Fc-диатела CD32B × CD79B»). Изобретение направлено на применение таких молекул и на применение фармацевтических композиций, которые содержат такие молекулы.
Как обсуждалось выше, CD79B и CD32B (FcγRIIB) оба экспрессируются B-клетками, которые пролиферируют в ответ на распознавание антигена. Биспецифичные связывающие молекулы по изобретению способны иммуноспецифически связываться с обеими молекулами и, следовательно, способны колигировать молекулы. Такое колигирование (см., например, фигуру 4) позволяет ITIM молекуле CD32B фосфорилироваться и привлекать домен SH2 инозит-полифосфат-5'-фосфатазы (SHIP), которая гидролизует мессенджеры-фосфоинозитолы, которые высвобождаются вследствие опосредуемой тирозинкиназой активации CD79B ITAM. Такой гидролиз ингибирует ITAM-активирующий сигнал в CD79B и, таким образом, служит для ослабления активации B-клеток. Таким образом, биспецифичные связывающие молекулы по изобретению обладают способностью ингибировать или ослаблять иммунную систему хозяина в ответ на нежелательную активацию B-клеток, пролиферацию B-клеток и секрецию антител и могут применяться для лечения воспалительных заболеваний и расстройств и, в частности, системной красной волчанки (SLE), рассеянного склероза (MS) и болезни трансплантат против хозяина (GvHD).
I. Признаки и структура антител
Используемый в данном документе термин «антитело» относится к молекуле иммуноглобулина, способной к иммуноспецифическому связыванию с полипептидом или белком или небелковой молекуле благодаря присутствию на такой молекуле определенного домена или фрагмента или конформации («эпитоп»). Эпитопсодержащая молекула может иметь иммуногенную активность, так что она вызывает реакцию продуцирования антитела у животного; такие молекулы называются «антигенами». Эпитопсодержащие молекулы не обязательно должны быть иммуногенными.
Природные антитела (такие как антитела IgG) состоят из двух легких цепей в комплексе с двумя тяжелыми цепями. Каждая легкая цепь естественного антитела (такого как антитело IgG) содержит вариабельный домен (домен VL) и константный домен (домен CL). Каждая тяжелая цепь природного антитела содержит вариабельный домен тяжелой цепи (домен VH), три константных домена (домены CH1, CH2 и CH3), и «шарнир» (Домен («H»), расположенный между доменами CH1 и CH2. Таким образом, основной структурной единицей естественных иммуноглобулинов (например, IgG) является тетрамер, имеющий две легкие цепи и две тяжелые цепи, обычно экспрессируемые в виде гликопротеина около 150 000 Да. Аминоконцевая («N-концевая») часть каждой цепи включает вариабельный домен, содержащий от 100 до 110 или более аминокислот, в основном ответственных за распознавание антигена. Карбоксиконцевая («С-концевая») часть каждой цепи определяет константный участок, причем легкие цепи имеют один константный домен, а тяжелые цепи, обычно имеют три константных домена и шарнирный домен. Таким образом, структура легких цепей молекулы IgG представляет собой n-VL-CL-c, а структура тяжелых цепей IgG представляет собой n-VH-CH1-H-CH2-CH3-c (где n и c представляют, соответственно, N-конец и С-конец полипептида). Способность антитела связывать эпитоп антигена зависит от наличия и аминокислотной последовательности доменов VL и VH антитела. Взаимодействие легкой цепи антитела и тяжелой цепи антитела и, в частности, взаимодействие его доменов VL и VH образует один из двух эпитопсвязывающих сайтов природного антитела. Природные антитела способны связываться только с одним видом эпитопа (т.е. они являются моноспецифичными), хотя они могут связывать несколько копий этого вида (т.е. проявлять двухвалентность или мультивалентность). Вариабельные домены молекулы IgG состоят из участков, определяющих комплементарность («CDR»), которые включают остатки в контакте с эпитопом и сегменты без CDR, называемые каркасными фрагментами («FR»), которые в целом поддерживают структуры и определяют расположение петель CDR, чтобы обеспечить такое контакт (хотя некоторые каркасные остатки также могут контактировать с антигеном). Таким образом, VL- и VH-домены имеют структуру n-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4-c. Полипептиды, которые являются (или могут служить в качестве) первого, второго и третьего CDR из легкой цепи антитела, в данном документе соответственно обозначены как домен CDRL1, домен CDRL2 и домен CDRL3. Аналогичным образом, полипептиды, которые являются (или могут служить в качестве) первого, второго и третьего CDR из тяжелой цепи антитела, соответственно, обозначены в данном документе как домен CDRH1, домен CDRH2, и домен CDRH3. Таким образом, термины Домен CDRL1, Домен CDRL2, Домен CDRL3, Домен CDRH1, домен CDRH2 и домен CDRH3 направлены на полипептиды, которые при включении в белок приводят к тому, что этот белок может связываться с определенным эпитопом независимо от того, является ли такой белок антителом, имеющим легкие и тяжелые цепи, или является диателом или одноцепочечной связывающей молекулой (например, scFv, BiTe и т.д.) или является другим типом белка. Соответственно, используемый в данном документе термин «эпитопсвязывающий домен», относится к той части молекулы, связывающей эпитоп, которая отвечает за способность такой молекулы иммуноспецифически связывать эпитоп. Эпитопсвязывающий фрагмент может содержать 1, 2, 3, 4, 5 или все 6 доменов CDR такого антитела и, хотя и способен иммуноспецифически связываться с таким эпитопом, может проявлять иммуноспецифичность, аффинность или селективность по отношению к такому эпитопу, который отличается от указанного такого антитела. Однако предпочтительно, чтобы эпитопсвязывающий фрагмент содержал все 6 доменов CDR такого антитела. Эпитопсвязывающий фрагмент антитела может быть одной полипептидной цепью (например, scFv) или может содержать две или более полипептидных цепей, каждая из которых имеет аминоконец и карбоксильный конец (например, диатело, фрагмент Fab, фрагмент Fab 2 и т.д.).
Используемый в данном документе термин «антитело» охватывает моноклональные антитела, полиспецифические антитела, антитела человека, гуманизированные антитела, синтетические антитела, химерные антитела, поликлональные антитела, верблюжьи антитела, одноцепочечные Fv (scFv), одноцепочечные антитела, иммунологически активные фрагменты антител (например, фрагменты антител, способные связываться с эпитопом, например, фрагменты Fab, фрагменты Fab', фрагменты F(ab')2, фрагменты Fv, фрагменты, содержащие домены VL и/или VH, или содержащие 1, 2 или 3 участков, определяющих комплементарность (CDR) такого домена VL (т.е. CDRL1, CDRL2 и/или CDRL3) или домена VH (т.е. CDRH1, CDRH2 и/или CDRH3)), которые специфически связывают антиген и т.д., бифункциональные или многофункциональные антитела, дисульфид-связанные биспецифичные Fv (sdFv), интратела и диатела и эпитопсвязывающие фрагменты любого из вышеуказанного. В частности, термин «антитело» предназначен для охвата молекул иммуноглобулина и иммунологически активных фрагментов молекул иммуноглобулина, то есть молекул, которые включают эпитопсвязывающий сайт. Молекулы иммуноглобулина могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса (см. например, патентные публикации Соединенных Штатов №: 20040185045; 20050037000; 20050064514; 20050215767; 20070004909; 20070036799; 20070077246; и 20070244303). В последние несколько десятилетий наблюдается возрождение интереса к терапевтическому потенциалу антител, и антитела стали одним из ведущих классов лекарственных средств, полученных биотехнологически (Chan, CE et al. (2009) “The Use Of Antibodies In The Treatment Of Infectious Diseases,” Singapore Med. J. 50(7):663-666). Более 200 лекарственных средств на основе антител были одобрены для применения или находятся в стадии разработки.
Термин «химерное антитело» относится к антителу, где часть тяжелой и/или легкой цепи идентична или гомологична антителу одного вида (например, мыши) или классу или подклассу антитела, в то время как оставшаяся часть идентична антителу другого вида (например, человека) или классу или подклассу антитела, при условии, что они проявляют искомую биологическую активность. Представляющие интерес химерные антитела в данном документе включают «приматизированные» антитела, содержащие антигенсвязывающие последовательности вариабельного домена, полученные из приматов, не являющихся человеком (например, обезьян Старого света, человекообразных обезьян и др.), и последовательности константных областей человека.
Используемый в данном документе термин «моноклональное антитело» относится к антителу популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных антител, обладающих природными мутациями, которые могут присутствовать в незначительных количествах, а термин «поликлональное антитело» в контексте настоящего описания относится к антителу, полученному из популяции гетерогенных антител. Термин «моноклональное» указывает на характер антитела как, по существу, на гомогенную популяцию антител, и его не следует истолковывать как требующий продуцирования антитела каким-либо конкретным способом (например, гибридомой, отбором фагов, рекомбинантной экспрессией, трансгенными животными и т.д.). Термин включает целые иммуноглобулины, а также фрагменты и т.д., описанные выше под определением «антитело». Способы получения моноклональных антител известны в данной области. Одним из способов, который может быть использован, является способ Kohler, G. et al. (1975) “Continuous Cultures Of Fused Cells Secreting Antibody Of Predefined Specificity,” Nature 256:495-497 или его модификации. Обычно моноклональные антитела вырабатывают в мышах, крысах или кроликах. Антитела получают путем иммунизации животного иммуногенным количеством клеток, клеточных экстрактов или белковых препаратов, которые включают искомый эпитоп. Иммуноген может представлять собой, без ограничения указанным, первичные клетки, культивируемые клеточные линии, злокачественные опухолевые клетки, белки, пептиды, нуклеиновые кислоты или ткань. Клетки, используемые для иммунизации, могут культивироваться в течение периода времени (например, по меньшей мере, 24 часа) до их применения в качестве иммуногена. Клетки могут быть использованы в качестве иммуногенов сами по себе или в сочетании с неденатурирующим адъювантом, таким как Ribi (см., например, Jennings, V.M. (1995) “Review of Selected Adjuvants Used in Antibody Production,” ILAR J. 37(3):119-125). Как правило, клетки должны быть сохранены интактными и предпочтительно должны быть жизнеспособными при использовании в качестве иммуногенов. Интактные клетки могут позволить антигенам лучше детектироваться иммунизированными животными, чем в случае антигенов поврежденных клеток. Применение денатурации или жестких адъювантов, например, адъюванта Фрейда, может привести к разрыву клеток и, следовательно, не рекомендуется. Иммуноген можно вводить несколько раз с периодическими интервалами, такими как, дважды в неделю или еженедельно, или может вводиться таким образом, чтобы поддерживать жизнеспособность животного (например, в тканевом рекомбинанте). В ином случае, существующие моноклональные антитела и любые другие эквивалентные антитела, которые являются иммуноспецифическими для искомого патогенного эпитопа, могут быть секвенированы и получены рекомбинантно любыми способами, известными в данной области. В одном воплощении такое антитело секвенировано и полинуклеотидная последовательность затем клонируется в вектор для экспрессии или размножения. Последовательность, кодирующая антитело, может поддерживаться в векторе в клетке-хозяине, и затем клетка-хозяин может быть размножена и заморожена для будущего применения. Полинуклеотидную последовательность таких антител можно использовать для генетической манипуляции для получения моноспецифических или мультиспецифических (например, биспецифических, триспецифических и тетраспецифических) молекул по изобретению, а также оптимизированного по аффинности, химерного антитела, гуманизированного антитела и/или канинизированного антитела, для улучшения аффинности или других характеристик антитела.
Термин «scFv» относится к одноцепочечным фрагментам вариабельных доменов. Молекулы scFv получают путем связывания вариабельного домена легкой и/или тяжелой цепи с использованием коротко связывающего пептида. Bird et al. (1988) (“Single-Chain Antigen-Binding Proteins,” Science 242:423-426) описывает пример линкерных пептидов, которые формируют мостик длиной около 3,5 нм между карбоксильным концом одного вариабельного домена и аминоконцом другого вариабельного домена. Были разработаны и использованы линкеры с другими последовательностями (Bird et al. (1988) “Single-Chain Antigen-Binding Proteins,” Science 242:423-426). В свою очередь, линкеры могут быть модифицированы для дополнительных функций, таких как прикрепление лекарственных средств или прикрепление к твердым подложкам. Одноцепочечные варианты могут быть получены либо рекомбинантно, либо синтетически. Для синтетического продуцирования scFv может использоваться автоматизированный синтезатор. Для рекомбинантного продуцирования scFv подходящая плазмида, содержащая полинуклеотид, который кодирует scFv, может быть введена в подходящую клетку-хозяина, либо эукариотическую, такую как клетки дрожжей, растений, насекомых или млекопитающих, или прокариотическую, такуб как E. coli. Полинуклеотиды, кодирующие представляющие интерес scFv, могут быть получены с помощью обычных манипуляций, таких как лигирование полинуклеотидов. Полученный scFv может быть выделен с применением стандартных методов очистки белка, известных в данной области.
Термин «гуманизированное» антитело относится к гибридной молекуле, обычно полученной с применением рекомбинантных методик, имеющей эпитопсвязывающий сайт иммуноглобулина из видов, не являющихся человеком, и остальную структуру иммуноглобулиновой молекулы, которая основана на структуре и/или последовательности иммуноглобулина человека. Эпитопсвязывающий сайт, может включать либо полные вариабельные домены, слитые с константными доменами, либо только CDR, привитые на соответствующие каркасные области в вариабельных доменах. Домены, связывающие эпитоп, могут быть дикого типа или модифицироваться одной или несколькими аминокислотными заменами. Это исключает константный участок в качестве иммуногена у людей, но оставляет возможность иммунного ответа на чуждый вариабельный домен (LoBuglio, AF et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224). Другой подход фокусируется не только на предоставлении константных участков человеческого происхождения, но и на изменении вариабельных доменов для того, чтобы как можно более точно изменить их на человеческую форму. Известно, что вариабельные домены как тяжелых, так и легких цепей содержат три участка, определяющих комплементарность (CDR), которые изменяются в зависимости от рассматриваемых антигенов и определяют способность связывания, фланкированные четырьмя каркасными областями (FR), которые относительно консервативные у данных видов и которые предположительно обеспечивают поддерживающий каркас для CDR. Когда антитела нечеловеческого происхождения получают к определенному антигену, вариабельные домены могут быть «изменены» или «гуманизированы» путем прививки CDR, полученных из антител нечеловеческого происхождения, на FR, присутствующие в человеческом антителе, подлежащем модификации. Применение этого подхода к различным антителам сообщалось Sato, K. et al. (1993) “Reshaping A Human Antibody To Inhibit The Interleukin 6-Dependent Tumor Cell Growth,” Cancer Res 53:851-856. Riechmann, L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; Kettleborough, C. A. et al. (1991) “Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformation,” Protein Engineering 4:773-3783; Maeda, H. et al. (1991) “Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity,” Human Antibodies Hybridoma 2:124-134; Gorman, S. D. et al. (1991) “Reshaping A Therapeutic CD4 Antibody,” Proc. Natl. Acad. Sci. (U.S.A.) 88:4181-4185; Tempest, P.R. et al. (1991) “Reshaping A Human Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo,” Bio/Technology 9:266-271; Co, M. S. et al. (1991) “Humanized Antibodies For Antiviral Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 88:2869-2873; Carter, P. et al. (1992) “Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 89:4285-4289; и Co, M.S. et al. (1992) “Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen,” J. Immunol. 148:1149-1154. В некоторых воплощениях гуманизированные антитела сохраняют все последовательности CDR (например, гуманизированное мышиное антитело, которое содержит все шесть CDR из мышиных антител). В других воплощениях гуманизированные антитела имеют один или несколько CDR (один, два, три, четыре, пять или шесть), которые изменены в их аминокислотной последовательности (последовательностях) относительно исходного антитела, которые также называют одним или несколькими CDR, полученными из одного или нескольких CDR исходного антитела (т.е. полученные из таких CDR, полученные на основании информации об аминокислотных последовательностях таких CDR и т.д.). Полинуклеотидная последовательность, которая кодирует вариабельный домен антитела, может быть использована для получения таких производных и для улучшения аффинности или других характеристик таких антител. Общий принцип гуманизации антитела включает сохранение основной последовательности эпитопсвязывающей части антитела, при замене нечеловеческого остатка антитела на последовательности антитела человека. Существует четыре общих стадии гуманизации моноклонального антитела. Это: (1) определение нуклеотидной и спрогнозированной аминокислотной последовательности исходных вариабельных доменов легких и тяжелых цепей антитела (2), конструирование гуманизированного антитела или канинизированного антитела, т.е. решение о том, какой каркасный участок антитела будет использоваться процесса гуманизации или канинизации, (3) фактические методологии/методы гуманизации или канинизации и (4) трансфекция и экспрессия гуманизированного антитела. См., например, пат. США № 4816567; 5807715; 5866692; и 6331415.
Как указано выше, биспецифические связывающие молекулы по настоящему изобретению обладают, по меньшей мере, двумя эпитопсвязывающими доменами. Каждый из таких эпитопсвязывающих доменов способен связываться с эпитопами «иммуноспецифичным» образом. В настоящем описании антитело, диатело или другая биспецифическая связывающая молекула по настоящему изобретению, согласно сообщениям, «иммуноспецифически» связывает (или проявляет «специфическое» связывание с) областью другой молекулы (т.е. эпитопа), если оно реагирует или ассоциируется чаще, быстрее, с большей продолжительностью и/или с большей аффинностью с этим эпитопом по сравнению с альтернативными эпитопами. Например, антитело, которое специфически связывается с эпитопом CD32B (или эпитопом CD79B), представляет собой антитело, которое связывает такой эпитоп с большей аффинностью, авидностью, более легко и/или с большей продолжительностью, чем оно связывается с другими эпитопами CD32B (или с другими эпитопами CD79B) или с эпитопом молекулы, отличной от CD32B (или CD79B). Также понятно, прочитав это определение, что, например, антитело (или фрагмент или эпитоп), которое иммуноспецифически связывается с первой мишенью, может или не может специфически или предпочтительно связываться со второй мишенью. Таким образом, «иммуноспецифическое связывание» необязательно требует (хотя и может включать) исключительного связывания. Как правило, но необязательно, ссылка на связывание означает «специфическое» связывание. Способность антитела иммуноспецифически связываться с эпитопом может быть определена, например, с помощью иммуноанализа.
Эпитопсвязывающий домен гуманизированных молекул по настоящему изобретению может содержать полный вариабельный домен, слитый с константным доменом, или только участки, определяющие комплементарность (CDR), такого вариабельного домена, привитые в соответствующие каркасные области. Эпитопсвязывающий домен может быть дикого типа или может быть модифицирован одной или несколькими аминокислотными заменами, например, для уменьшения способности любого константного домена молекулы служить иммуногеном у людей. Хотя это может устранить константную область в качестве иммуногена у людей, возможность иммунного ответа на чужеродный вариабельный домен остается (LoBuglio, AF et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224). Другой подход фокусируется не только на предоставлении константных участков человеческого происхождения, но и на изменении вариабельных доменов для того, чтобы как можно более точно изменить их на человеческую форму. Известно, что вариабельные домены как тяжелых, так и легких цепей содержат три участка, определяющих комплементарность (CDR), которые изменяются в зависимости от рассматриваемых антигенов и определяют способность связывания, фланкированные четырьмя каркасными областями (FR), которые относительно консервативные у данных видов и которые предположительно обеспечивают поддерживающий каркас для CDR. Когда антитела нечеловеческого происхождения получают к определенному антигену, вариабельные домены могут быть «изменены» или «гуманизированы» путем прививки CDR, полученных из антител нечеловеческого происхождения, на FR, присутствующие в человеческом антителе, подлежащем модификации. Применение этого подхода к различным антителам сообщалось Sato, K. et al. (1993) Cancer Res 53:851-856. Riechmann, L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; Kettleborough, C. A. et al. (1991) “Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformation,” Protein Engineering 4:773-3783; Maeda, H. et al. (1991) “Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity,” Human Antibodies Hybridoma 2:124-134; Gorman, S. D. et al. (1991) “Reshaping A Therapeutic CD4 Antibody,” Proc. Natl. Acad. Sci. (U.S.A.) 88:4181-4185; Tempest, P.R. et al. (1991) “Reshaping A Human Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo,” Bio/Technology 9:266-271; Co, M. S. et al. (1991) “Humanized Antibodies For Antiviral Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 88:2869-2873; Carter, P. et al. (1992) “Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 89:4285-4289; и Co, M.S. et al. (1992) “Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen,” J. Immunol. 148:1149-1154. В некоторых воплощениях гуманизированные антитела сохраняют все последовательности CDR (например, гуманизированное мышиное антитело, которое содержит все шесть CDR из мышиных антител). В других воплощениях гуманизированные антитела имеют один или несколько CDR (один, два, три, четыре, пять или шесть), которые отличаются по последовательности от исходного антитела.
Было описано несколько молекул гуманизированного антитела, содержащих эпитопсвязывающий сайт, полученный из иммуноглобулина, отличного от человека, включая гибридные антитела, содержащие вариабельный домен из грызунов или модифицированный вариабельный домен из грызунов, и связанные с ними области, определяющие комплементарность (CDR), слитые с константными доменами человека (см., например, Winter et al. (1991) “Man-made Antibodies,” Nature 349:293-299; Lobuglio et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224 (1989), Shaw et al. (1987) “Characterization Of A Mouse/Human Chimeric Monoclonal Antibody (17-1A) To A Colon Cancer Tumor-Associated Antigen,” J. Immunol. 138:4534-4538, and Brown et al. (1987) “Tumor-Specific Genetically Engineered Murine/Human Chimeric Monoclonal Antibody,” Cancer Res. 47:3577-3583). Другие источники описывают CDR грызунов, привитые в поддерживающую каркасную область (FR) человека перед слиянием с соответствующим константным доменом антитела человека (см., например, Riechmann L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; and Jones et al. (1986) “Replacing The Complementarity-Determining Regions In A Human Antibody With Those From A Mouse,” Nature 321:522-525). В другом источнике описываются CDR грызунов, поддерживаемые рекомбинантно венированными каркасными областями грызунов. См., например, Европейскую патентную публикацию №519596. Эти «гуманизированные» молекулы предназначены для минимизации нежелательной иммунологической реакции на молекулы грызунов против антител человека, которые ограничивают продолжительность и эффективность терапевтических применений этих фрагментов у реципиентов-людей. Другие способы гуманизации антител, которые также могут быть использованы, раскрыты Daugherty et al. (1991) “Polymerase Chain Reaction Facilitates The Cloning, CDR-Grafting, And Rapid Expression Of A Murine Monoclonal Antibody Directed Against The CD18 Component Of Leukocyte Integrins,” Nucl. Acids Res. 19: 2471-2476 и в пат. США № 6180377; 6054297; 5997867; и 5866692.
II. Константные участки антитела
Предпочтительным доменом CL является домен CL каппа из IgG человека. Аминокислотной последовательностью примерного домена CL-каппа человека является (SEQ ID NO: 1):
RTVAAPSVFI FPPSDEQLKS GTASVVCLLN NFYPREAKVQ WKVDNALQSG NSQESVTEQD SKDSTYSLSS TLTLSKADYE KHKVYACEVT HQGLSSPVTK SFNRGEC
В ином случае примерный домен CL является доменом CL лямбда из IgG человека. Аминокислотная последовательность примерного домена CL Лямбда человека представляет собой (SEQ ID NO: 2):
QPKAAPSVTL FPPSSEELQA NKATLVCLIS DFYPGAVTVA WKADSSPVKA GVETTPSKQS NNKYAASSYL SLTPEQWKSH RSYSCQVTHE GSTVEKTVAP TECS
Примерный домен CH1 является доменом CH1 из IgG1 человека. Аминокислотная последовательность примерного домена CH1 из IgG1 человека представляет собой (SEQ ID NO: 3):
ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YICNVNHKPS NTKVDKRV
Примерный домен CH1 является доменом CH1 из IgG2 человека. Аминокислотная последовательность примерного домена CH1 из IgG2 человека представляет собой (SEQ ID NO: 4):
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSNFGTQT YTCNVDHKPS NTKVDKTV
Примерный домен CH1 является доменом CH1 из IgG4 человека. Аминокислотная последовательность примерного домена CH1 из IgG4 человека представляет собой (SEQ ID NO: 5):
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTKT YTCNVDHKPS NTKVDKRV
Один примерный шарнирный участок представляет собой шарнирный участок из IgG1 человека. Аминокислотная последовательность примерного шарнирного участка из IgG1 человека представляет собой (SEQ ID NO: 6): EPKSCDKTHTCPPCP.
Другой пример шарнирного участка представляет собой шарнирный участок из IgG2 человека. Аминокислотная последовательность примерного шарнирного участка из IgG2 человека представляет собой (SEQ ID NO: 7): ERKCCVECPPCP.
Другой пример шарнирного участка представляет собой шарнирный участок из IgG4 человека. Аминокислотная последовательность примерного шарнирного участка из IgG4 человека представляет собой (SEQ ID NO: 8): ESKYGPPCPSCP. Как описано в данном документе, шарнирный участок из IgG4 может включать стабилизирующую мутацию, такую как замена S228P. Аминокислотная последовательность типичного стабилизированного шарнирного участка IgG4 представляет собой (SEQ ID NO: 9): ESKYGPPCPPCP.
Домены СН2 и СН3 тяжелых цепей антител взаимодействуют с образованием области Fc, которая включает домен Fc, распознаваемый клеточными рецепторами Fc, включая, без ограничения указанным, гамма-рецепторы Fc (FcγR), такие как CD32B. Аминокислотная последовательность домена CH2-CH3 типичного IgG1 человека представляет собой (SEQ ID NO: 10):

согласно нумерации по индексу EU, представленном у Kabat, где X является лизином (K) или отсутствует.
Аминокислотная последовательность домена CH2-CH3 примерного IgG2 человека представляет собой (SEQ ID NO: 14):

согласно нумерации по индексу EU, представленном у Kabat, где X является лизином (K) или отсутствует.
Аминокислотная последовательность домена CH2-CH3 примерного IgG3 человека представляет собой (SEQ ID NO: 12):


согласно нумерации по индексу EU, представленном у Kabat, где X является лизином (K) или отсутствует.
Аминокислотная последовательность домена CH2-CH3 примерного IgG4 человека представляет собой (SEQ ID NO: 13):

согласно нумерации по индексу EU, представленном у Kabat, где X является лизином (K) или отсутствует.
В настоящем описании нумерация остатков в константной области тяжелой цепи IgG соответствует нумерации индекса EU, как в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, NH1, MD (1991) (“Kabat”), явно включенного в данный документ ссылкой. Термин «индекс EU по Kabat» относится к нумерации человеческого антитела IgG1 EU. Аминокислоты из вариабельных доменов зрелых тяжелых и легких цепей иммуноглобулинов обозначаются положением аминокислоты в цепи. Kabat описал многочисленные аминокислотные последовательности для антител, идентифицировал аминокислотную консенсусную последовательность для каждой подгруппы и присвоил номер остатка каждой аминокислоте, и CDR идентифицируется, как определено Kabat (понятно, что CDRH1, определенный в Chothia, C. & Lesk, AM ((1987) “Canonical structures for the hypervariable regions of immunoglobulins,”. J. Mol. Biol. 196: 901-917) начинается на пять остатков ранее). Схема нумерации Kabat расширяется до антител, не включенных в его сборник, путем выравнивания рассматриваемого антитела с одной из консенсусных последовательностей в Kabat по отношению к консервативным аминокислотам. Этот способ присвоения номеров остатков стал стандартным в данной области и легко идентифицирует аминокислоты в эквивалентных положениях в разных антителах, включая гибридные или гуманизированные варианты. Например, аминокислота в положении 50 легкой цепи человеческого антитела занимает эквивалентное положение аминокислоты в положении 50 легкой цепи мышиного антитела.
Полиморфизмы наблюдались в нескольких разных положениях в константных участках антитела (например, в положениях в Fc, включая, без ограничения указанным, положения 270, 272, 312, 315, 356 и 358, согласно нумерации по индексу EU, указанному в Kabat), и, таким образом, могут существовать небольшие различия между представленной последовательностью и последовательностями в известном уровне техники. Хорошо известны полиморфные формы иммуноглобулинов человека. В настоящее время известны 18 аллотипов Gm: G1m (1, 2, 3, 17) или G1m (a, x, f, z), G2m (23) или G2m (n), G3m (5, 6, 10, 11, 13, 14, 15, 16, 21, 24, 26, 27, 28) или G3m (b1, c3, b3, b0, b3, b4, s, t, g1, c5, u, v, g5) (Lefranc, et al., «The Human IgG Subclasses: Molecular Analysis Of Structure, Function And Regulation.». Pergamon, Oxford, pp. 43-78 (1990); Lefranc, G. et al., 1979, Hum. Genet.: 50, 199-211). В частности, предполагается, что антитела по настоящему изобретению могут включать любой аллотип, изоалотип или гаплотип любого гена иммуноглобулина и не ограничены аллотипом, изоаллотипом или гаплотипом последовательностей, представленных в данном документе. Кроме того, в некоторых системах экспрессии C-концевой аминокислотный остаток (выделенный жирным шрифтом) домена CH3 может быть посттрансляционно удален. Соответственно, С-концевой остаток домена СН3 является необязательным аминокислотным остатком в связывающих молекулах по изобретению, содержащих домен Fc. Настоящим изобретением специально охватывается связывающие молекулы по настоящему изобретению, в которых отсутствует С-концевой остаток домена СН3. Также, в частности, в настоящее изобретение включены такие конструкты, которые содержат С-концевой остаток лизина в домене СН3.
III. Предпочтительные связывающей молекулы CD32B x CD79B по настоящему изобретению
Настоящее изобретение относится к биспецифическим связывающим молекулам, которые способны связываться с эпитопом CD32B и эпитопом Cd79b, способны одновременно связываться с такими молекулами, которые нативно расположены (т.е. без рекомбинантно индуцированной сверхэкспрессии) на поверхности B-клетки. Такие специфически связывающиеся молекулы могут состоять из одной полипептидной цепи (например, BiTe) или могут состоять из двух, трех, четырех, пяти или более полипептидных цепей, которые вместе образуют ковалентно связанный комплекс, предпочтительно посредством присутствия множества дисульфидных связей между отдельными полипептидными цепями связывающей молекулы CD32B × CD79B. Предпочтительно, такие молекулы будут способны иммуноспецифически связываться с CD32B, по существу, не влияя или не препятствуя способности молекулы CD32B связываться с Fc-доменом антитела или диатела, содержащего Fc-домен.
A. Биспецифичные ScFv и антитела
В первом предпочтительном воплощении связывающие молекулы CD32B × CD79B по настоящему изобретению представляют собой одноцепочечные молекулы, такие как BiTe, которые обладают доменом VLCD32B, доменом VLCD79B, доменом VHCD32B и доменом VLCD79B и в которых домены разделены молекулами пептидного линкера, которые позволяют домену VLCD32B взаимодействовать с доменом VHCD32B, чтобы сформировать домен, связывающий эпитоп CD32B, и которые позволяют домену VLCD79B взаимодействовать с доменом VHCD79B, чтобы сформировать домен, связывающий эпитоп CD79B.
Во втором предпочтительном воплощении связывающие молекулы CD32B × CD79B по настоящему изобретению представляют собой биспецифичные антитела или их эпитопсвязывающие фрагменты, которые обладают доменом VLCD32B, доменом VLCD79B, доменом VHCD32B и доменом VLCD79B, тем самым образуя CD32B-эпитопсвязывающий домен и CD79B-эпитопсвязывающий домен. Такие антитела могут содержать домен Fc.
B. Биспецифичные диатела
1. Биспецифичные диатела, не содержащие домен Fc
В следующем предпочтительном воплощении связывающие молекулы CD32B × CD79B по настоящему изобретению представляют собой биспецифичные моновалентные диатела, которые состоят из двух, трех, четырех, пяти или более полипептидных цепей.
Например, на фигуре 1 показано биспецифическое моновалентное диатело CD32B × CD79B, состоящее из двух полипептидных цепей, которые ковалентно связаны друг с другом через дисульфидную связь. Домен VL первой полипептидной цепи взаимодействует с доменом VH второй полипептидной цепи, формируя первый функциональный сайт связывания антигена, который специфичен для первого антигена (т.е. либо CD32B, либо CD79B). Аналогично, домен VL второй полипептидной цепи взаимодействует с доменом VH первой полипептидной цепи, формируя второй функциональный сайт связывания антигена, который специфичен для второго антигена (т.е. либо CD79B, либо CD32B, в зависимости от идентичности первого антигена). Таким образом, отбор доменов VL и VH первой и второй полипептидных цепей скоординирован таким образом, что две полипептидные цепи совместно включали домены VL и VH, способные связываться с CD32B и CD79B (т.е. они содержат VLCD32B/VHCD32B и VLCD79B/VHCD79B) (фигура 1). В совокупности каждый такой домен VL и VH и промежуточный линкер, который их разделяет, называются антигенсвязывающим доменом молекулы.
Первая полипептидная цепь предпочтительного биспецифического моновалентного диатела CD32B × CD79B содержит (в направлении от N-конца к C-концу): амино-конец, домен VL моноклонального антитела, способного связываться либо с CD32B, либо с CD79B (т.е. либо VLCD32B либо VLCD79B), промежуточный спейсерный пептид (линкер 1), домен VH моноклонального антитела, способного связываться либо с CD79B (если такая первая полипептидная цепь содержит VLCD32B), либо с CD32B (если такая первая полипептидная цепь содержит VLCD79B), промежуточный спейсерный пептид (линкер 2), домен, способствующий гетеродимеризации, необязательный дополнительный домен для обеспечения улучшенной стабилизации домена, способствующего гетеродимеризации, и С-конец (фигура 1).
Вторая полипептидная цепь такого предпочтительного CD32B × CD79B биспецифического моновалентного Fc-диатела включает (в направлении от N-конца к C-концу): аминоконец, домен VL моноклонального антитела, способного связываться с любым CD79B или CD32B (т.е. либо VLCD79B, либо VLCD32B, в зависимости от домена VL, выбранного для первой полипептидной цепи диатела), промежуточный линкерный пептид (линкер 1), домен VH моноклонального антитела, способного связываться с любым из CD32B (если такой второй полипептид цепь содержит VLCD79B) или CD32B (если такая вторая полипептидная цепь содержит VLCD32B), промежуточный спейсерный пептид (линкер 2), домен, способствующий гетеродимеризации, и С-конец (фигура 1).
Наиболее предпочтительно, если длина линкера 1, который разделяет такие домены VL и VH, выбирается так, чтобы по существу или полностью предотвращать связывание таких доменов VL и VH друг с другом (например, состоит из 1, 2, 3, 4, 5, 6 7, 8 или 9 аминокислотных остатков). Таким образом, домены VL и VH первой полипептидной цепи по существу или полностью неспособны связываться друг с другом. Аналогично, Домены VL и VH второй полипептидной цепи по существу или полностью неспособны связываться друг с другом. Предпочтительный промежуточный спейсерный пептид (линкер 1) имеет последовательность (SEQ ID NO: 14): GGGSGGGG.
Целью линкера 2 является отделение домена VH полипептидной цепи от необязательно присутствующего домена, способствующего гетеродимеризации, этой же полипептидной цепи. Любой из множества линкеров может использоваться в качестве линкера 2. Предпочтительная последовательность для такого линкера 2 имеет аминокислотную последовательность: ASTKG (SEQ ID NO: 15), которая получена из домена CH1 из IgG, или GGCGGG (SEQ ID NO: 16), который включает остаток цистеина, который может быть использован для ковалентного связывания первой и второй полипептидных цепей друг с другом через дисульфидную связь. Поскольку линкер 2, ASTKG (SEQ ID NO: 15) не обладает таким цистеином, применение такого линкера 2 предпочтительно связано с использованием цистеинсодержащего домена, способствующего гетеродимеризации, такого как E-спираль SEQ. ID NO: 23 или K-спираль SEQ ID NO: 24 (см. ниже). Таким образом, в одном воплощении линкер 2 полипептидных цепей содержит остаток цистеина (чтобы ковалентно связывать первую и вторую полипептидные цепи друг с другом). В другом воплощении линкер 2 полипептидных цепей не обладает цистеином, и домены, способствующие гетеродимеризации, таких полипептидных цепей содержат такой остаток цистеина, чтобы, тем самым, ковалентно связывать первую и вторую полипептидные цепи друг с другом.
Образование гетеродимеров из первой и второй полипептидных цепей может быть обусловлено включением доменов, способствующих гетеродимеризации. Такие домены включают GVEPKSC (SEQ ID NO: 17) или VEPKSC (SEQ ID NO: 18) на одной полипептидной цепи и GFNRGEC (SEQ ID NO: 19) или FNRGEC (SEQ ID NO: 20) на другой полипептидной цепи (US2007/0004909).
Более предпочтительно, однако, если домены, способствующие гетеродимеризации, по настоящему изобретению образованы из одного, двух, трех или четырех тандемно повторяющихся спиральных доменов с противоположным зарядом, которые содержат последовательность, по меньшей мере, из шести, по меньшей мере, семи или, по меньшей мере, восьми заряженных аминокислотных остатков (Apostolovic, B. et al. (2008) “pH-Sensitivity of the E3/K3 Heterodimeric Coiled Coil,” Biomacromolecules 9:3173-3180; Arndt, K.M. et al. (2001) “Helix-stabilized Fv (hsFv) Antibody Fragments: Substituting the Constant Domains of a Fab Fragment for a Heterodimeric Coiled-coil Domain,” J. Molec. Biol. 312:221-228; Arndt, K.M. et al. (2002) “Comparison of In Vivo Selection and Rational Design of Heterodimeric Coiled Coils,” Structure 10:1235-1248; Boucher, C. et al. (2010) “Protein Detection By Western Blot Via Coiled-Coil Interactions,” Analytical Biochemistry 399:138-140; Cachia, P.J. et al. (2004) “Synthetic Peptide Vaccine Development: Measurement Of Polyclonal Antibody Affinity And Cross-Reactivity Using A New Peptide Capture And Release System For Surface Plasmon Resonance Spectroscopy,” J. Mol. Recognit. 17:540-557; De Crescenzo, G.D. et al. (2003) “Real-Time Monitoring of the Interactions of Two-Stranded de novo Designed Coiled-Coils: Effect of Chain Length on the Kinetic and Thermodynamic Constants of Binding,” Biochemistry 42:1754-1763; Fernandez-Rodriquez, J. et al. (2012) “Induced Heterodimerization And Purification Of Two Target Proteins By A Synthetic Coiled-Coil Tag,” Protein Science 21:511-519; Ghosh, T.S. et al. (2009) “End-To-End And End-To-Middle Interhelical Interactions: New Classes Of Interacting Helix Pairs In Protein Structures,” Acta Crystallographica D65:1032-1041; Grigoryan, G. et al. (2008) “Structural Specificity In Coiled-Coil Interactions,” Curr. Opin. Struc. Biol. 18:477-483; Litowski, J.R. et al. (2002) “Designing Heterodimeric Two-Stranded α-Helical Coiled-Coils: The Effects Of Hydrophobicity And α-Helical Propensity On Protein Folding, Stability, And Specificity,” J. Biol. Chem. 277:37272-37279; Steinkruger, J.D. et al. (2012) “The d'--d--d' Vertical Triad is Less Discriminating Than the a'--a--a' Vertical Triad in the Antiparallel Coiled-coil Dimer Motif,” J. Amer. Chem. Soc. 134(5):2626-2633; Straussman, R. et al. (2007) “Kinking the Coiled Coil - Negatively Charged Residues at the Coiled-coil Interface,” J. Molec. Biol. 366:1232-1242; Tripet, B. et al. (2002) “Kinetic Analysis of the Interactions between Troponin C and the C-terminal Troponin I Regulatory Region and Validation of a New Peptide Delivery/Capture System used for Surface Plasmon Resonance,” J. Molec. Biol. 323:345-362; Woolfson, D.N. (2005) “The Design Of Coiled-Coil Structures And Assemblies,” Adv. Prot. Chem. 70:79-112; Zeng, Y. et al. (2008) “A Ligand-Pseudoreceptor System Based On de novo Designed Peptides For The Generation Of Adenoviral Vectors With Altered Tropism,” J. Gene Med. 10:355-367).
Такие повторяющиеся спиральные домены могут быть точными повторами или могут иметь замены. Например, домен, способствующий гетеродимеризации, первой полипептидной цепи может содержать последовательность из восьми отрицательно заряженных аминокислотных остатков, а домен, способствующий гетеродимеризации, второй полипептидной цепи может содержать последовательность из восьми отрицательно заряженных аминокислотных остатков. Неважно, какая спираль предлагается для первой или второй полипептидным цепям, при условии, что спираль с противоположным зарядом используется для другой полипептидной цепи. Однако предпочтительное биспецифическое моновалентное диатело CD32B × CD79B по настоящему изобретению имеет первую полипептидную цепь с отрицательно заряженной спиралью. Положительно заряженная аминокислота может представлять собой лизин, аргинин, гистидин и т.д. и/или отрицательно заряженная аминокислота может представлять собой глутаминовую кислоту, аспарагиновую кислоту и др. Положительно заряженная аминокислота предпочтительно представляет собой лизин, и/или отрицательно заряженная аминокислота предпочтительно представляет собой глутаминовую кислоту. Возможно применение только одного домена, способствующего гетеродимеризации (поскольку такой домен будет ингибировать гомодимеризацию и, следовательно, способствовать гетеродимеризации), однако предпочтительно, чтобы как первая, так и вторая полипептидные цепи диател по настоящему изобретению содержали домены, способствующие гетеродимеризации.
В предпочтительном воплощении один из доменов, способствующих гетеродимеризации, будет содержать четыре тандемных «E-спиральных» геликоидальных домена (SEQ ID NO:21: 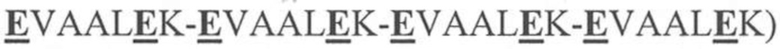 ? ? ? чьи глутаматные остатки будут образовывать отрицательный заряд при рН 7, в то время как другой из доменов, способствующих гетеродимеризации, будет содержать четыре тандемных «K-спиральных» домена (SEQ ID NO: 22:
? ? ? чьи глутаматные остатки будут образовывать отрицательный заряд при рН 7, в то время как другой из доменов, способствующих гетеродимеризации, будет содержать четыре тандемных «K-спиральных» домена (SEQ ID NO: 22: 
остатки лизина которого будут образовывать положительный заряд при pH 7. Наличие таких заряженных доменов способствует ассоциации между первым и вторым полипептидами и, таким образом, способствует гетеродимеризации. Особенно предпочтительным является домен, способствующий гетеродимеризации, где один из четырех тандемных «E-спиральных» геликоидальных доменов SEQ ID NO: 21 был модифицирован путем включения остатка цистеина: 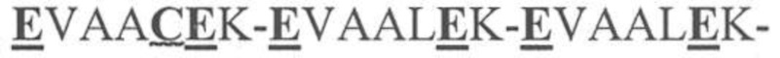
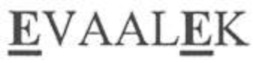 (SEQ ID NO:23). Аналогичным образом, особенно предпочтительным является домен, способствующий гетеродимеризации, в котором один из четырех «K-спиральных» геликоидальных доменов (SEQ ID NO: 22) был модифицирован путем включения остатка цистеина:
(SEQ ID NO:23). Аналогичным образом, особенно предпочтительным является домен, способствующий гетеродимеризации, в котором один из четырех «K-спиральных» геликоидальных доменов (SEQ ID NO: 22) был модифицирован путем включения остатка цистеина: 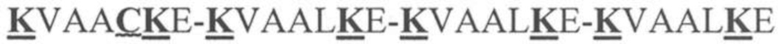 (SEQ ID NO: 24).
(SEQ ID NO: 24).
2. Биспецифичные диатела, содержащие домен Fc
В следующем предпочтительном воплощении изобретения диатела CD32B × CD79B по настоящему изобретению дополнительно включают область Fc. Область Fc таких Fc-содержащих диател по настоящему изобретению может представлять собой либо полный участок Fc (например, полный участок Fc из IgG), либо только фрагмент полного участка Fc. Хотя домен Fc биспецифических моновалентных Fc-диател по настоящему изобретению может обладать способностью связываться с одним или несколькими Fc-рецепторами (например, FcγR), более предпочтительно, чтобы такой домен Fc обладал существенным снижением или отсутствием способности связываться с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (относительно связывания, проявляемого участком Fc дикого типа). Таким образом, домен Fc молекул, содержащих домен Fc, по настоящему изобретению, может включать часть или весь домен CH2 и/или часть или весь домен CH3 полной области Fc или может включать последовательность вариантного CH2 и/или вариантного CH3 (который может включать, например, одну или несколько вставок и/или одну или несколько делеций по отношению к доменам CH2 или CH3 полной области Fc). Домен Fc биспецифических моновалентных Fc-диател по настоящему изобретению может содержать участки, не принадлежащие полипептиду Fc или может содержать участки не полностью естественных участков Fc или может содержать не встречающиеся в природе ориентации доменов CH2 и/или CH3 (такие как, например, два домена CH2 или два домена CH3, или в направлении от N-конца к C-концу, домен CH3, связанный с доменом CH2 и т.д.).
Такие антитела, содержащие Fc-область, по настоящему изобретению могут содержать две полипептидные цепи (например, фигура 2) или могут содержать три (например, фигуры 3А-3Е) или более полипептидных цепей. На фигуре 2 показано диатело, имеющее структуру, аналогичную описанной выше, за исключением того, что соответствующие домены, способствующие гетеродимеризации, заменены доменами CH2-CH3. Предпочтительно, чтобы область Fc, образованная такими полипептидными цепями, имела существенно сниженную способность или вообще не связывалась с активирующим FcγR, таким как FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (относительно связывания, демонстрируемого участком Fc дикого типа).
На фигуре 3А-3Е показаны альтернативные Fc-диатела CD32B × CD79B, состоящие из трех полипептидных цепей, среди которых первая и вторая полипептидные цепи ковалентно связаны друг с другом, и первая и третья полипептидные цепи ковалентно связаны друг с другом. Как и в описанных выше диателах, домен VL первой полипептидной цепи взаимодействует с доменом VH второй полипептидной цепи для образования первого функционального сайта связывания антигена, который специфичен для первого антигена (т.е. либо CD32B, либо CD79B). Аналогично, домен VL второй полипептидной цепи взаимодействует с доменом VH первой полипептидной цепи, чтобы сформировать второй функциональный сайт связывания антигена, который специфичен для второго антигена (т.е. либо CD79B, либо CD32B, в зависимости от идентичности первого антигена). Таким образом, выбор из VL и VH доменов первой и второй полипептидных цепей осуществляется таким образом, что две полипептидные цепи в совокупности включают домены VL и VH, способные связываться с CD32B и CD79B (т.е., они содержат VLCD32B/VH CD32B и VLCD79B/VH CD79B). В совокупности каждый такой домен VL и VH и промежуточный линкер, который их разделяет, называют антигенсвязывающим доменом молекулы.
В воплощении биспецифического Fc-диатела CD32B × CD79B, показанного на фигуре 3А и фигуре 3В, первая полипептидная цепь содержит (в направлении от N-конца к С-концу): аминоконец, цистеинсодержащий пептид (пептид 1), домен Fc из IgG, состоящий из всех или части доменов CH2 и CH3 участка Fc антитела, промежуточный пептидный линкер (линкер 3), домен VL моноклонального антитела, способного связываться либо с CD32B, либо с CD79B (т.е. с VLCD32B или VLCD79B), промежуточный пептид (линкер 1), домен VH моноклонального антитела, способного связываться либо с CD79B (если такая первая полипептидная цепь содержит VLCD32B), либо с CD32B (если такая первая полипептидная цепь содержит VLCD79B), промежуточный спейсерный пептид (линкер 2), домен, способствующий гетеродимеризации, необязательный четвертый спейсерный пептид (линкер 4) для обеспечения улучшенной стабилизации домена, способствующего гетеродимеризации, и С-конец.
Вторая полипептидная цепь таких воплощений биспецифического Fc-диатела CD32B × CD79B содержит (в направлении от N-конца к C-концу): амино-конец, домен VL моноклонального антитела, способного связываться либо с CD79B, либо с CD32B (т.е. либо VLCD79B либо VLCD32B, в зависимости от домена VL, выбранного для первой полипептидной цепи диатела), промежуточный линкерный пептид (линкер 1), домен VH моноклонального антитела, способный связываться с любым CD32B (если такая вторая полипептидная цепь содержит VLCD79B) или CD32B (если такая вторая полипептидная цепь содержит VLCD32B), промежуточный спейсерный пептид (линкер 2), домен, способствующий гетеродимеризации, и С-конец.
Третья полипептидная цепь такого предпочтительного биспецифического Fc-диатела CD32B × CD79B включает (в направлении от N-конца к C-концу): амино-конец, цистеинсодержащий пептид (пептид 1), домен Fc из IgG (предпочтительно домены CH2 и СН3 участка Fc антитела), имеющие тот же изотип, что и у домена Fc первой полипептидной цепи и С-конец.
Воплощение биспецифического Fc-диатела CD32B × CD79B, показанного на фигуре 3А, отличается от показанного на фигуре 3В тем, что промежуточный спейсерный пептид (линкер 2) первой и второй полипептидных цепей не содержит остаток цистеина, такой остаток цистеина теперь является частью доменов, способствующих гетеродимеризации, из этих полипептидных цепей.
В воплощении биспецифического Fc-диатела CD32B × CD79B, показанного на фигуре 3C, первая полипептидная цепь содержит (в направлении от N-конца к C-концу): аминоконец, домен VL моноклонального антитела, способного связываться с CD32B или CD79B (т.е. либо VLCD32B, либо VLCD79B), промежуточный пептид (линкер 1), домен VH моноклонального антитела, способного связываться либо с CD79B (если такая первая полипептидная цепь содержит VLCD32B), либо с CD32B (если такой первый полипептид цепь содержит VLCD79B), цистеинсодержащий промежуточный спейсерный пептид (линкер 2), домен, способствующий гетеродимеризации, цистеинсодержащий пептид (пептид 1), домен Fc из IgG, состоящий из всех или части доменов CH2 и CH3 участка Fc антитела, и С-конец.
Вторая полипептидная цепь таких воплощений биспецифического Fc-диатела CD32B × CD79B содержит (в направлении от N-конца к C-концу): амино-конец, домен VL моноклонального антитела, способного связываться либо с CD79B, либо с CD32B (т.е. либо VLCD79B или VLCD32B, в зависимости от домена VL, выбранного для первой полипептидной цепи диатела), промежуточный линкерный пептид (линкер 1), домен VH моноклонального антитела, способного связываться с любым CD32B (если такая вторая полипептидная цепь содержит VLCD79B) или CD32B (если такая вторая полипептидная цепь содержит VLCD32B), цистеинсодержащий промежуточный спейсерный пептид (линкер 2), домен, способствующий гетеродимеризации, и С-конец.
Третья полипептидная цепь биспецифического Fc-диатела CD32B × CD79B на фигуре 3C и фигуре 3D содержит (в направлении от N-конца к C-концу): аминоконец, цистеинсодержащий пептид (пептид 1), домен Fc из IgG (предпочтительно домены СН2 и СН3 участка Fc антитела), имеющие тот же изотип, что и у домена Fc первой полипептидной цепи, и С-конец.
Воплощение биспецифического Fc-диатела CD32B × CD79B, показанного на фигуре 3D, отличается от показанного на фигуре 3C тем, что промежуточный спейсерный пептид (линкер 2) первой и второй полипептидных цепей не содержит остатка цистеина, такой остаток цистеина теперь является частью доменов, способствующих гетеродимеризации, этих полипептидных цепей.
Цистеинсодержащий пептид (пептид 1) первой и третьей полипептидных цепей может состоять из одной и той же аминокислотной последовательности или из разных аминокислотных последовательностей и содержать 1, 2, 3 или более остатков цистеина. Особенно предпочтительный пептид 1 имеет аминокислотную последовательность (SEQ ID NO: 25): DKTHTCPPCP или (SEQ ID NO: 26) GGGDKTHTCPPCP. Предпочтительный промежуточный линкерный пептид (линкер 3) содержит аминокислотную последовательность (SEQ ID NO: 27): APSSS и более предпочтительно имеет аминокислотную последовательность (SEQ ID NO: 28): APSSSPME. Предпочтительный пептид четвертого спейсера (линкер 4) имеет последовательность GGG или представляет собой SEQ ID NO: 29: GGGNS.
Предпочтительно, чтобы Fc-домен, образованный первой и третьей полипептидными цепями Fc-содержащих диател по изобретению, существенно слабее связывался или не имел способности связываться с активирующим FcγR, таким как FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (относительно связывания, проявляемого участком Fc дикого типа). Домены Fc, имеющие мутации, которые уменьшают или устраняют связывание с такими рецепторами, хорошо известны в данной области и включают аминокислотные замены в положениях 234 и 235, замену в положении 265 или замену в положении 297 (см., например, патент США №.5624821, включенный в настоящее описание ссылкой). В предпочтительном воплощении домен СН2 и СН3 включает замену в положении 234 на аланин и 235 на аланин.
Домены СН2 и/или СН3 первой и третьей полипептидных цепей Fc-содержащих диател по изобретению не обязательно должны быть идентичными и преимущественно модифицированы, чтобы способствовать комплексообразованию между двумя полипептидами. Например, замещение аминокислоты (предпочтительно замещение аминокислотой, содержащей громоздкую боковую группу, образующую «выступ», например триптофан), может быть введена в домен CH2 или CH3, так что стерическая интерференция предотвратит взаимодействие с аналогично мутированным доменом и обязывает мутированный домен спариваться с доменом, где была спроектирована комплементарная или вмещающая мутация, т.е. «углубление» (например, замена на глицин). Такие наборы мутаций могут быть сконструированы в любой паре полипептидов, содержащих молекулу Fc-диатела, и, кроме того, встроены в любую часть цепей полипептидов указанной пары. Способы белковой инженерии, способствующие гетеродимеризации, а не гомодимеризации, хорошо известны в данной области, в частности, в отношении инженерии иммуноглобулиноподобных молекул и охватываются данным документом (см., например, Ridgway et al. (1996) “‘Knobs-Into-Holes’ Engineering Of Antibody CH3 Domains For Heavy Chain Heterodimerization,” Protein Engr. 9:617-621, Atwell et al. (1997) “Stable Heterodimers From Remodeling The Domain Interface Of A Homodimer Using A Phage Display Library,” J. Mol. Biol. 270: 26-35, and Xie et al. (2005) “A New Format Of Bispecific Antibody: Highly Efficient Heterodimerization, Expression And Tumor Cell Lysis,” J. Immunol. Methods 296:95-101; каждый из которых полностью включен в настоящее описание ссылкой). Предпочтительно «выступ» встроен в домены CH2-CH3 первой полипептидной цепи, а «углубление» встроено в домены CH2-CH3 третьей полипептидной цепи. Таким образом, «выступ» поможет предотвратить гомодимеризацию первой полипептидной цепи через ее домены CH2 и/или CH3. Поскольку третья полипептидная цепь предпочтительно содержит замену «углубление», она будет гетеродимеризоваться с первой полипептидной цепью, а также гомодимеризоваться сама с собой. Предпочтительный выступ создается путем изменения участка Fc из нативного IgG так, чтобы он содержал модификацию T366W. Предпочтительное углубление создается путем модификации участка Fc из нативного IgG так, чтобы он содержал модификации T366S, L368A и Y407V. Чтобы облегчить очистку гомодимера третьей полипептидной цепи, несущей отверстие, от конечного биспецифического моновалентного Fc-диатела, сайт связывания протеина А в доменах СН2 и СН3, несущих углубление, третьей полипептидной цепи предпочтительно мутирован путем аминокислотной замены в положении 435 (H435R). Чтобы помочь очистить гомодимер третьей полипептидной цепи от конечного биспецифического моновалентного Fc-диатела, содержащего первую, вторую и третью полипептидные цепи, сайт связывания протеина А доменов СН2 и СН3 третьей полипептидной цепи предпочтительно мутируют путем замены аминокислот. Таким образом, третий гомодимер полипептидной цепи не будет связываться с протеином А, тогда как биспецифическое моновалентное диатело Fc сохранит свою способность связывать протеин А через сайт связывания протеина А на первой полипептидной цепи.
3. Типичные биспецифичные диатела CD32B × CD79B
Типичное биспецифичное диатело CD32B × CD79B по изобретению будет содержать две или более полипептидных цепей и будет включать:
(1) домен VL антитела, связывающего CD32B (VLCD32B), такой как домен VLCD32B, имеющий последовательность (SEQ ID NO: 30):
DIQMTQSPSS LSASVGDRVT ITCRASQEIS GYLSWLQQKP GKAPRRLIYA ASTLDSGVPS RFSGSESGTE FTLTISSLQP EDFATYYCLQ YFSYPLTFGG GTKVEIK
(2) домен VH антитела, которое связывает CD32B (VHCD32B), такого как домен VHCD32B, имеющий последовательность (SEQ ID NO: 31):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS DAWMDWVRQA PGKGLEWVAE IRNKAKNHAT YYAESVIGRF TISRDDAKNS LYLQMSSRA EDTAVYYCGA LGLDYWGQGT LVTVSS
(3) домен VL антитела, связывающего CD79B (VLCD79B), такой как домен VLCD79B, имеющий последовательность (SEQ ID NO: 32):
DVVMTQSPLS LPVTLGQPAS ISCKSSQSLL DSDGKTYLNW FQQRPGQSPN RLIYLVSKLD SGVPDRFSGS GSGTDFTLKI SRVEAEDVGV YYCWQGTHFP LTFGGGTKLE IK
(4) домен VH антитела, связывающего CD79B (VHCD79B), такой как домен VHCD79B, имеющий последовательность (SEQ ID NO: 33):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT SYWMNWVRQA PGQGLEWIGM IDPSDSETHY NQKFKDRVTM TTDTSTSTAY MELRSLRSDD TAVYYCARAM GYWGQGTTVT VSS
Первое примерное биспецифичное диатело CD32B x CD79B по изобретению имеет две полипептидные цепи. Первая полипептидная цепь такого примерного диатела имеет структуру, в направлении от N-конца к C-концу: N-конец, вышеуказанный домен VLCD32B, Линкер 1, вышеуказанный домен VHCD79B, цистеинсодержащий линкер 2, E-спиральный домен и C-конец. Аминокислотная последовательность такого предпочтительного полипептида представляет собой (SEQ ID NO: 34):
DIQMTQSPSS LSASVGDRVT ITCRASQEIS GYLSWLQQKP GKAPRRLIYA ASTLDSGVPS RFSGSESGTE FTLTISSLQP EDFATYYCLQ YFSYPLTFGG GTKVEIKGGG SGGGGQVQLV QSGAEVKKPG ASVKVSCKAS GYTFTSYWMN WVRQAPGQGL EWIGMIDPSD SETHYNQKFK DRVTMTTDTS TSTAYMELRS LRSDDTAVYY CARAMGYWGQ GTTVTVSSGG CGGGEVAALE KEVAALEKEV AALEKEVAAL EK
В SEQ ID NO: 34 аминокислотные остатки 1-107 представляют собой домен VL антитела, которое связывает CD32B (VLCD32B) (SEQ ID NO: 30), аминокислотные остатки 108-115 представляют собой линкер 1 (SEQ ID NO: 14), аминокислотные остатки 116-228 являются доменом VH антитела, которое связывает CD79B (VH CD79B) (SEQ ID NO: 33), аминокислотные остатки 229-234 представляют собой цистеинсодержащий линкер 2 (SEQ ID NO: 16) аминокислотные остатки 235-262 являются E-спиральным доменом, способствующим гетеродимеризации (SEQ ID NO: 21).
Вторая полипептидная цепь такого примерного диатела имеет аминокислотную последовательность в направлении от N-конца к C-концу (SEQ ID NO: 35):
DVVMTQSPLS LPVTLGQPAS ISCKSSQSLL DSDGKTYLNW FQQRPGQSPN RLIYLVSKLD SGVPDRFSGS GSGTDFTLKI SRVEAEDVGV YYCWQGTHFP LTFGGGTKLE IKGGGSGGGG EVQLVESGGG LVQPGGSLRL SCAASGFTFS DAWMDWVRQA PGKGLEWVAE IRNKAKNHAT YYAESVIGRF TISRDDAKNS LYLQMNSLRA EDTAVYYCGA LGLDYWGQGT LVTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 35 аминокислотные остатки 1-112 представляют собой домен VL антитела, которое связывает CD79B (VLCD79B) (SEQ ID NO: 32), аминокислотные остатки 113-120 представляют собой линкер 1 (SEQ ID NO: 14), аминокислотные остатки 121-236 представляют собой домен VH антитела, которое связывает CD32B (VHCD32B) (SEQ ID NO: 31), аминокислотные остатки 237-242 представляют собой цистеинсодержащий линкер 2 (SEQ ID NO: 16) и аминокислотные остатки 243-270 являются K-спиральным доменом, способствующим гетеродимеризации (SEQ ID NO: 22).
Второе иллюстративное биспецифическое диатело CD32B x CD79B по изобретению имеет две полипептидные цепи, в которых первая полипептидная цепь имеет структуру в направлении от N-конца к C-концу: N-конец, указанный выше домен VLCD32B, Линкер 1, вышеуказанный домен VHCD79B, линкер 2, цистеинсодержащий E-спиральный домен и C-конец. Аминокислотная последовательность такого предпочтительного полипептида представляет собой (SEQ ID NO: 36):
DIQMTQSPSS LSASVGDRVT ITCRASQEIS GYLSWLQQKP GKAPRRLIYA ASTLDSGVPS RFSGSESGTE FTLTISSLQP EDFATYYCLQ YFSYPLTFGG GTKVEIKGGG SGGGGQVQLV QSGAEVKKPG ASVKVSCKAS GYTFTSYWMN WVRQAPGQGL EWIGMIDPSD SETHYNQKFK DRVTMTTDTS TSTAYMELRS LRSDDTAVYY CARAMGYWGQ GTTVTVSSAS TKGEVAACEK EVAALEKEVA ALEKEVAALE K
В SEQ ID NO: 36 аминокислотные остатки 1-107 представляют собой домен VL антитела, которое связывает CD32B (VLCD32B) (SEQ ID NO: 30), аминокислотные остатки 108-115 представляют собой линкер 1 (SEQ ID NO: 14), аминокислотные остатки 116-228 представляют собой домен VH антитела, которое связывает CD79B (VHCD79B) (SEQ ID NO: 33), аминокислотные остатки 229-233 представляют собой линкер 2 (SEQ ID NO: 15), аминокислотные остатки 234-261 представляют собой цистеинсодержащий Е-спиральный домен, способствующий гетеродимеризации (SEQ ID NO: 23).
Вторая полипептидная цепь такого второго примерного диатела имеет аминокислотную последовательность в направлении от N-конца к C-концу (SEQ ID NO: 37):
DVVMTQSPLS LPVTLGQPAS ISCKSSQSLL DSDGKTYLNW FQQRPGQSPN RLIYLVSKLD SGVPDRFSGS GSGTDFTLKI SRVEAEDVGV YYCWQGTHFP LTFGGGTKLE IKGGGSGGGG EVQLVESGGG LVQPGGSLRL SCAASGFTFS DAWMDWVRQA PGKGLEWVAE IRNKAKNHAT YYAESVIGRF TISRDDAKNS LYLQMNSLRA EDTAVYYCGA LGLDYWGQGT LVTVSSASTK GKVAACKEKV AALKEKVAAL KEKVAALKE
В SEQ ID NO: 37 аминокислотные остатки 1-112 представляют собой домен VL антитела, которое связывает CD79B (VLCD79B) (SEQ ID NO: 32), аминокислотные остатки 113-120 представляют собой Линкер 1 (SEQ ID NO: 14), аминокислотные остатки 121-236 представляют собой домен VH антитела, которое связывает CD32B (VHCD32B) (SEQ ID NO: 31), аминокислотные остатки 237-241 представляют собой линкер 2 (SEQ ID NO: 15) и аминокислотные остатки 242-269 представляют собой цистеинсодержащий К-спиральный домен, способствующий гетеродимеризации (SEQ ID NO: 24).
4. Типичные биспецифичные Fc-диатела CD32B × CD79B
Первое примерное биспецифическое Fc-диатело CD32B × CD79B по изобретению имеет три полипептидные цепи (фигура 3А). Первая полипептидная цепь будет содержать домены CH2 и CH3 выступ-содержащего участка Fc из IgG, имеющего последовательность (SEQ ID NO: 38):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK
Таким образом, первая полипептидная цепь такого примерного Fc-диатела имеет структуру в направлении от N-конца к C-концу: пептид 1, домен CH2-CH3 участка Fc из IgG, линкер 1, домен VL антитела, которое связывает CD32B (VLCD32B), цистеинсодержащий линкер 2, домен VH антитела, которое связывает CD79B (VHCD79B), линкер 3, E-спиральный домен, линкер 4 и C-конец. Аминокислотная последовательность такого предпочтительного полипептида представляет собой (SEQ ID NO: 39):
DKTHTCPPCP APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGKAPS SSPMEDIQMT QSPSSLSASV GDRVTITCRA SQEISGYLSW LQQKPGKAPR RLIYAASTLD SGVPSRFSGS ESGTEFTLTI SSLQPEDFAT YYCLQYFSYP LTFGGGTKVE IKGGGSGGGG QVQLVQSGAE VKKPGASVKV SCKASGYTFT SYWMNWVRQA PGQGLEWIGM IDPSDSETHY NQKFKDRVTM TTDTSTSTAY MELRSLRSDD TAVYYCARAM GYWGQGTTVT VSSGGCGGGE VAALEKEVAA LEKEVAALEK EVAALEKGGG NS
В SEQ ID NO: 39 аминокислотные остатки 1-10 представляют собой пептид 1 (SEQ ID NO: 25), аминокислотные остатки 11-227 представляют собой домены CH2 и CH3 выступ-содержащего участка Fc из IgG (SEQ ID NO: 38), аминокислотные остатки 228-235 представляют собой линкер 3 (SEQ ID NO: 28), аминокислотные остатки 236-342 представляют собой домен VL антитела, которое связывает CD32B (VLCD32B) (SEQ ID NO: 30), аминокислотные остатки 343-350 представляют собой линкер 1 (SEQ ID NO: 14), аминокислотные остатки 351-463 представляют собой домен VH антитела, которое связывает CD79B (VHCD79B) (SEQ ID NO: 33), аминокислотные остатки 464-469 представляют собой цистеинсодержащий линкер 2 (SEQ ID NO: 16), аминокислотные остатки 470-497 представляют собой E-спиральный домен, способствующий гетеродимеризации (SEQ ID NO: 21), а аминокислотные остатки 498-502 представляют собой линкер 4 (SEQ ID NO: 29)
Предпочтительный полинуклеотид, который кодирует первую полипептидную цепь, имеет последовательность (SEQ ID NO: 40):
gacaaaactc acacatgccc accgtgccca gcacctgaag ccgcgggggg accgtcagtc ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggagga gatgaccaag aaccaggtca gcctgtggtg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggg aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc ctctccctgt ctccgggtaa agccccttcc agctccccta tggaagacat ccagatgacc cagtctccat cctccttatc tgcctctgtg ggagatagag tcaccatcac ttgtcgggca agtcaggaaa ttagtggtta cttaagctgg ctgcagcaga aaccaggcaa ggcccctaga cgcctgatct acgccgcatc cactttagat tctggtgtcc catccaggtt cagtggcagt gagtctggga ccgagttcac cctcaccatc agcagccttc agcctgaaga ttttgcaacc tattactgtc tacaatattt tagttatccg ctcacgttcg gaggggggac caaggtggaa ataaaaggag gcggatccgg cggcggaggc caggttcagc tggtgcagtc tggagctgag gtgaagaagc ctggcgcctc agtgaaggtc tcctgcaagg cttctggtta cacctttacc agctactgga tgaactgggt gcgacaggcc cctggacaag ggcttgagtg gatcggaatg attgatcctt cagacagtga aactcactac aatcaaaagt tcaaggacag agtcaccatg accacagaca catccacgag cacagcctac atggagctga ggagcctgag atctgacgac acggccgtgt attactgtgc gagagctatg ggctactggg ggcaagggac cacggtcacc gtctcctccg gaggatgtgg cggtggagaa gtggccgcac tggagaaaga ggttgctgct ttggagaagg aggtcgctgc acttgaaaag gaggtcgcag ccctggagaa aggcggcggg aactct
Вторая полипептидная цепь такого примерного Fc-диатела имеет структуру в направлении от N-конца к C-концу: домен VL антитела, который связывает CD79B (VLCD79B), Линкер 1, домен VH антитела, который связывает CD32B (VH CD32B), цистеинсодержащий линкер 2, К-спиральный домен, способствующий гетеродимеризации, и C-конец.
Предпочтительной последовательностью для второй полипептидной цепи является (SEQ ID NO: 41):
DVVMTQSPLS LPVTLGQPAS ISCKSSQSLL DSDGKTYLNW FQQRPGQSPN RLIYLVSKLD SGVPDRFSGS GSGTDFTLKI SRVEAEDVGV YYCWQGTHFP LTFGGGTKLE IKGGGSGGGG EVQLVESGGG LVQPGGSLRL SCAASGFTFS DAWMDWVRQA PGKGLEWVAE IRNKAKNHAT YYAESVIGRF TISRDDAKNS LYLQMNSLRA EDTAVYYCGA LGLDYWGQGT LVTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 41 аминокислотные остатки 1-112 представляют собой домен VL антитела, которое связывает CD79B (VLCD79B) (SEQ ID NO: 32), аминокислотные остатки 113-120 представляют собой линкер 1 (SEQ ID NO: 14), аминокислотные остатки 121-236 представляют собой домен VH антитела, которое связывает CD32B (VH CD32B) (SEQ ID NO: 31), аминокислотные остатки 237-242 представляют собой цистеинсодержащий линкер 2 (SEQ ID NO: 16) и аминокислотные остатки 243-270 являются K-спиральным доменом, способствующим гетеродимеризации (SEQ ID NO: 22).
Предпочтительный полинуклеотид, который кодирует вторую полипептидную цепь, имеет последовательность (SEQ ID NO: 42):
gatgttgtga tgactcagtc tccactctcc ctgcccgtca cccttggaca gccggcctcc atctcctgca agtcaagtca gagcctctta gatagtgatg gaaagacata tttgaattgg tttcagcaga ggccaggcca atctccaaac cgcctaattt atctggtgtc taaactggac tctggggtcc cagacagatt cagcggcagt gggtcaggca ctgatttcac actgaaaatc agcagggtgg aggctgagga tgttggggtt tattactgct ggcaaggtac acattttccg ctcacgttcg gcggagggac caagcttgag atcaaaggag gcggatccgg cggcggaggc gaagtgcagc ttgtggagtc tggaggaggc ttggtgcaac ctggaggatc cctgagactc tcttgtgccg cctctggatt cacttttagt gacgcctgga tggactgggt ccgtcaggcc ccaggcaagg ggcttgagtg ggttgctgaa attagaaaca aagctaaaaa tcatgcaaca tactatgctg agtctgtgat agggaggttc accatctcaa gagatgacgc caaaaacagt ctgtacctgc aaatgaacag cttaagagct gaagacactg ccgtgtatta ctgtggggct ctgggccttg actactgggg ccaaggcacc ctggtgaccg tctcctccgg aggatgtggc ggtggaaaag tggccgcact gaaggagaaa gttgctgctt tgaaagagaa ggtcgccgca cttaaggaaa aggtcgcagc cctgaaagag
Такое примерное биспецифическое Fc-диатело CD32B × CD79B будет иметь третью полипептидную цепь, которая будет содержать домены CH2 и CH3 содержащего отверстие участка Fc из IgG, имеющей аминокислотную последовательность (SEQ ID NO: 43)
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE ALHNRYTQKS LSLSPGK
Таким образом, аминокислотная последовательность третьей полипептидной цепи такого примерного биспецифического Fc-диатела CD32B × CD79B представляет собой SEQ ID NO: 44:
DKTHTCPPCP APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE ALHNRYTQKS LSLSPGK
В SEQ ID NO: 44 аминокислотные остатки 1-10 представляют собой пептид 1 (SEQ ID NO: 25), а аминокислотные остатки 11-227 представляют собой домены CH2 и CH3 содержащего отверстие участка Fc из IgG (SEQ ID NO: 43).
Предпочтительный полинуклеотид, который кодирует третью полипептидную цепь, имеет последовательность (SEQ ID NO: 45):
gacaaaactc acacatgccc accgtgccca gcacctgaag ccgcgggggg accgtcagtc ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggagga gatgaccaag aaccaggtca gcctgagttg cgcagtcaaa ggcttctatc ccagcgacat cgccgtggag tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc gacggctcct tcttcctcgt cagcaagctc accgtggaca agagcaggtg gcagcagggg aacgtcttct catgctccgt gatgcatgag gctctgcaca accgctacac gcagaagagc ctctccctgt ctccgggtaa a
Второе примерное биспецифичное Fc-диатело CD32B × CD79B по изобретению также имеет три полипептидные цепи (фигура 3B). Первая полипептидная цепь содержит СН2 и СН3 домены выступ-содержащего участка Fc из IgG, и имеет аминокислотную последовательность SEQ ID NO: 38.
Таким образом, первая полипептидная цепь такого второго примерного Fc-диатела имеет структуру в направлении от N-конца к C-концу: пептид 1, домен CH2-CH3 выступ-содержащего участка Fc из IgG, линкер 1, домен VL антитела, которое связывает CD32B (VLCD32B), линкер 2, домен VH антитела, связывающего CD79B (VH CD79B), линкер 3, цистеинсодержащий E-спиральный домен, линкер 4 и C-конец. Аминокислотная последовательность такого предпочтительного полипептида представляет собой (SEQ ID NO: 46):
DKTHTCPPCP APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGKAPS SSPMEDIQMT QSPSSLSASV GDRVTITCRA SQEISGYLSW LQQKPGKAPR RLIYAASTLD SGVPSRFSGS ESGTEFTLTI SSLQPEDFAT YYCLQYFSYP LTFGGGTKVE IKGGGSGGGG QVQLVQSGAE VKKPGASVKV SCKASGYTFT SYWMNWVRQA PGQGLEWIGM IDPSDSETHY NQKFKDRVTM TTDTSTSTAY MELRSLRSDD TAVYYCARAM GYWGQGTTVT VSSASTKGEV AACEKEVAAL EKEVAALEKE VAALEKGGGN S
В SEQ ID NO: 46 аминокислотные остатки 1-10 представляют собой пептид 1 (SEQ ID NO: 25), аминокислотные остатки 11-227 представляют собой домены CH2 и CH3 выступ-содержащего участка Fc из IgG (SEQ ID NO: 38).), аминокислотные остатки 228-235 представляют собой линкер 3 (SEQ ID NO: 28), аминокислотные остатки 236-342 представляют собой домен VL антитела, которое связывает CD32B (VLCD32B) (SEQ ID NO: 30), аминокислотные остатки 343-350 представляют собой линкер 1 (SEQ ID NO: 14), аминокислотные остатки 351-463 представляют собой домен VH антитела, которое связывает CD79B (VHCD79B) (SEQ ID NO: 33), аминокислотные остатки 464-468 представляют собой линкер 2 (SEQ ID NO: 15), аминокислотные остатки 469-496 представляют собой цистеинсодержащий E-спиральный домен, способствующий гетеродимеризации (SEQ ID NO: 23), а аминокислотные остатки 497-501 представляют собой Линкер 4 (SEQ ID NO: 29).
Вторая полипептидная цепь такого второго примерного Fc-диатела имеет структуру в направлении от N-конца к C-концу: домен VL антитела, которое связывает CD79B (VLCD79B), линкер 1, домен VH антитела, которое связывает CD32B (VHCD32B), линкер 2, цистеинсодержащий К-спиральный домен, способствующий гетеродимеризации, и C-конец.
Предпочтительной последовательностью для второй полипептидной цепи является (SEQ ID NO: 47):
DVVMTQSPLS LPVTLGQPAS ISCKSSQSLL DSDGKTYLNW FQQRPGQSPN RLIYLVSKLD SGVPDRFSGS GSGTDFTLKI SRVEAEDVGV YYCWQGTHFP LTFGGGTKLE IKGGGSGGGG EVQLVESGGG LVQPGGSLRL SCAASGFTFS DAWMDWVRQA PGKGLEWVAE IRNKAKNHAT YYAESVIGRF TISRDDAKNS LYLQMNSLRA EDTAVYYCGA LGLDYWGQGT LVTVSSASTK GKVAACKEKV AALKEKVAAL KEKVAALKE
В SEQ ID NO: 47 аминокислотные остатки 1-112 представляют собой домен VL антитела, которое связывает CD79B (VLCD79B) (SEQ ID NO: 32), аминокислотные остатки 113-120 представляют собой линкер 1 (SEQ ID NO: 14), аминокислотные остатки 121-236 представляют собой домен VH антитела, которое связывает CD32B (VHCD32B) (SEQ ID NO: 31), аминокислотные остатки 237-241 представляют собой линкер 2 (SEQ ID NO: 15) и аминокислотные остатки 242-269 представляют собой цистеинсодержащий К-спиральный домен, способствующий гетеродимеризации (SEQ ID NO: 24).
Третья полипептидная цепь такого второго примерного Fc-диатела будет содержать домены CH2 и CH3 участка IgG, содержащего углубление (SEQ ID NO: 43).
Таким образом, аминокислотная последовательность третьей полипептидной цепи такого примерного биспецифического Fc-диатела CD32B × CD79B представляет собой SEQ ID NO: 48:
DKTHTCPPCP APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE ALHNRYTQKS LSLSPGK
В SEQ ID NO: 48 аминокислотные остатки 1-10 представляют собой пептид 1 (SEQ ID NO: 25), а аминокислотные остатки 11-227 представляют собой домены CH2 и CH3 содержащего углубление участка Fc из IgG (SEQ ID NO: 43).
Альтернативная молекула биспецифического моновалентного Fc-диатела CD32B x CD79B по настоящему изобретению схематически показана на фигуре 3C. Такие альтернативные молекулы Fc-диатела CD32B × CD79B содержат три полипептидные цепи, из которых первая и вторая полипептидные цепи ковалентно связаны друг с другом, а первая и третья полипептидные цепи связаны друг с другом. Альтернативные молекулы биспецифического моновалентного Fc-диатела CD32B × CD79B отличаются по порядку своих доменов относительно порядка, присутствующего в предпочтительных молекулах биспецифического моновалентного Fc-диатела CD32B × CD79B. Однако, как и в случае предпочтительного Fc-диатела CD32B × CD79B, домен VL первой полипептидной цепи альтернативного биспецифического моновалентного Fc-диатела CD32B × CD79B взаимодействует с доменом VH второй полипептидной цепи альтернативного биспецифичного моновалентного Fc-диатела CD32B × CD79B для образования первого функционального сайта связывания антигена, который специфичен для первого антигена (т.е. либо CD32B, либо CD79B). Аналогично, домен VL второй полипептидной цепи альтернативного биспецифического моновалентного Fc-диатела CD32B × CD79B взаимодействует с доменом VH первой полипептидной цепи альтернативного биспецифического моновалентного Fc-диатела CD32B × CD79B с образованием второго функционального антигенсвязывающего сайта, который специфичен ко второму антигену (т.е. CD79B или CD32B, в зависимости от идентичности первого антигена). Таким образом, отбор доменов VL и VH первой и второй полипептидных цепей скоординирован таким образом, что две полипептидные цепи совместно включают домены VL и VH, способные связываться с CD32B и CD79B (т.е. они содержат VLCD32B/VHCD32B и VLCD79B/VHCD79B) (фигура 3C). В совокупности каждый такой домен VL и VH и промежуточный линкер, который их разделяет, называются антигенсвязывающим доменом молекулы.
Первая полипептидная цепь такого альтернативного Fc-диатела CD32B × CD79B содержит в направлении от N-конца к C-концу: аминоконец, домен VL моноклонального антитела, способного связываться либо с CD32B, либо с CD79B (т.е. с VLCD32B, или VLCD79B), промежуточный спейсерный пептид (линкер 1), домен VH моноклонального антитела, способного связываться либо с CD79B (если такая первая полипептидная цепь содержит VLCD32B), либо с CD32B (если такая первая полипептидная цепь содержит VLCD79B), цистеинсодержащий третий промежуточный спейсерный пептид (линкер 2), домен, способствующий гетеродимеризации, необязательный четвертый спейсерный пептид (линкер 4) для обеспечения улучшенной стабилизации домена, способствующего гетеродимеризации (предпочтительно E-спиральный домен), цистеин-содержащий пептид (пептид 1), домен Fc из IgG (предпочтительно, домены СН2 и СН3 выступ-содержащего участка Fc из IgG) и С-конец. Предпочтительно, чтобы домен Fc первой полипептидной цепи имел существенно сниженную способность или вообще не связывался с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (относительно связывания, демонстрируемого участком Fc дикого типа) (фигура 3C).
Вторая полипептидная цепь такого альтернативного Fc-диатела CD32B × CD79B содержит, в направлении от N-конца к C-концу, амино-конец, домен VL моноклонального антитела, способного связываться либо с CD79B, либо с CD32B (т.е. с VLCD79B или VLCD32B, в зависимости от домена VL, выбранного для первой полипептидной цепи диатела), промежуточный пептидный линкер (Линкер 1), домен VH моноклонального антитела, способного связываться с любым CD32B (если такая вторая полипептидная цепь содержит VLCD79B) или CD32B (если такая вторая полипептидная цепь содержит VLCD32B), цистеинсодержащий спейсерный пептид (Линкер 2), домен, способствующий гетеродимеризации (предпочтительно К-спиральный домен) и C-конец (фигура 3C).
Третья полипептидная цепь предпочтительного Fc-диатела CD32B × CD79B содержит, в направлении от N-конца к C-концу, амино-конец, цистеинсодержащий пептид (пептид 1), домен Fc из IgG (предпочтительно, домены CH2 и CH3, содержащего углубление участка Fc из IgG), имеющие тот же изотип, что и у домена Fc первой полипептидной цепи, и С-конец. Предпочтительно, чтобы домен Fc третьей полипептидной цепи имел существенно пониженную способность или вообще не связывался с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (относительно связывания, демонстрируемого участком Fc дикого типа) (фигура 3C).
На фигуре 3D показан вариант такого Fc-диатела, где цистеинсодержащий линкер 2 (например, GGCGGG (SEQ ID NO: 16)) был заменен не цистеинсодержащим линкером (например, ASTKG (SEQ ID NO: 15)) и в котором соответствующие домены,
способствующие гетеродимеризации, содержат остатки цистеина (например, 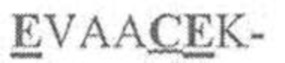
 (SEQ ID NO: 23) и
(SEQ ID NO: 23) и
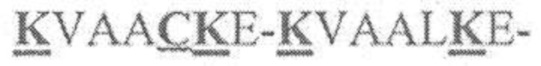
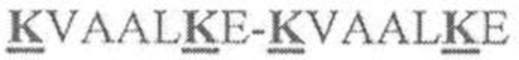 (SEQ ID NO: 24).
(SEQ ID NO: 24).
Фармацевтические композиции
Композиции по изобретению включают нерасфасованные лекарственные композиции, пригодные для изготовления фармацевтических композиций (например, неочищенных или нестерильных композиций) и фармацевтических композиций (т.е. композиций, которые подходят для введения объекту или пациенту), которые могут быть использованы в подготовке стандартных лекарственных форм. Такие композиции содержат профилактически или терапевтически эффективное количество связывающих молекул CD32B × CD79B по настоящему изобретению, и, в частности, любое из диател CD32B × CD79B или Fc-диател по изобретению или комбинацию таких агентов и фармацевтически приемлемый носитель. Предпочтительно композиции по изобретению содержат профилактически или терапевтически эффективное количество одной или нескольких молекул по изобретению и фармацевтически приемлемый носитель.
Изобретение также охватывает фармацевтические композиции, содержащие такие связывающие молекулы CD32B × CD79B и, в частности, любое из диател CD32B × CD79B или Fc-диател по изобретению и второе терапевтическое антитело (например, антигенспецифическое моноклональное антитело против аутоиммунного заболевания или воспалительного заболевания), которое является специфическим для конкретного антигена аутоиммунного или воспалительного заболевания, и фармацевтически приемлемый носитель.
В конкретном воплощении термин «фармацевтически приемлемый» означает одобренный регулирующим органом федерального правительства или правительства штата или перечисленный в Фармакопее США или другой общепризнанной фармакопее для применения на животных и, более конкретно, на людях. Термин «носитель» относится к разбавителю, адъюванту (например, адъюванту Фрейнда (полному и неполному), эксципиенту или носителю, с которым вводится терапевтическое средство. Такими фармацевтическими носителями могут быть стерильные жидкости, такие как вода и масла, включая масла нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное. Вода является предпочтительным носителем, когда фармацевтическую композицию вводят внутривенно. Солевые растворы и водные растворы декстрозы и глицерина также могут быть использованы в качестве жидких носителей, особенно для инъекционных растворов. Подходящие фармацевтические эксципиенты включают, например, крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропилен, гликоль, вода, этанол и тому подобное. Композиция, при необходимости, может также содержать незначительные количества смачивающих или эмульгирующих агентов или рН-буферных агентов. Эти композиции могут принимать форму растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков, составов с замедленным высвобождением и тому подобного.
Как правило, ингредиенты композиций по изобретению поставляются либо отдельно, либо смешиваются вместе в стандартной дозированной форме, например, в виде сухого лиофилизированного порошка или концентрата без воды в герметично закрытом контейнере, таком как ампула или саше, на которых указано количество активного агента. В случае, когда композиция должна вводиться путем инфузии, ее можно распределять из инфузионной емкости, содержащей стерильную воду или физиологический раствор фармацевтической категории. Когда композицию вводят путем инъекции, может быть предусмотрена ампула стерильной воды для инъекций или физиологического раствора, так чтобы ингредиенты могли быть смешаны до введения.
Композиции по изобретению могут быть составлены в виде нейтральных или солевых форм. Фармацевтически приемлемые соли включают, без ограничения указанным, соли, образованные с помощью анионов, таких как соли, полученные из соляной, фосфорной, уксусной, щавелевой, винной кислот и т.д., и соли, образованные с катионами, такие как соли, полученные из натрия, калия, аммония, кальция, гидроксида железа, изопропиламина, триэтиламина, 2-этиламиноэтанола, гистидина, прокаина и др.
Изобретение также относится к фармацевтической упаковке или набору, содержащему один или несколько контейнеров, заполненных связывающими молекулами CD32B × CD79B, и, в частности, любым из диател CD32B × CD79B или Fc-диател по изобретению отдельно или с таким фармацевтически приемлемым носителем. Кроме того, один или несколько других профилактических или терапевтических агентов, полезных для лечения заболевания, также могут быть включены в фармацевтический пакет или набор. Изобретение также относится к фармацевтической упаковке или набору, содержащему один или несколько контейнеров, заполненных одним или несколькими ингредиентами фармацевтических композиций по изобретению. Необязательно связанным с таким контейнером(ами) может быть уведомление в форме, предписанной правительственным учреждением, регулирующим производство, применение или продажу фармацевтических препаратов или биологических продуктов, которое отражает одобрение агентства по производству, использованию или продаже для введения людям.
Настоящее изобретение предлагает наборы, которые могут использоваться в вышеуказанных способах. В одном воплощении набор содержит одну или несколько молекул по изобретению. В другом воплощении набор дополнительно содержит один или несколько других профилактических или терапевтических агентов, полезных для лечения аутоиммунного или воспалительного заболевания, в одном или нескольких контейнерах. В другом воплощении набор дополнительно включает одно или несколько антител, которые связывают один или несколько антигенов аутоиммунного или воспалительного заболевания, связанных с аутоиммунным или воспалительным заболеванием. В определенных воплощениях другое профилактическое или терапевтическое средство представляет собой химиотерапевтическое средство. В других воплощениях профилактическое или терапевтическое средство представляет собой биологическое или гормональное терапевтическое средство.
Применения композиций по изобретению
Связывающие молекулы CD32B × CD79B и, в частности, любые из диател CD32B × CD79B или Fc-диател по изобретению обладают способностью лечить любое заболевание или состояние, связанное или характеризуемое экспрессией CD79B или имеющее В-клеточный компонент в болезни. Таким образом, без ограничения, фармацевтические композиции, содержащие такие молекулы, могут использоваться для диагностики или лечения аутоиммунных или воспалительных заболеваний или состояний. Таким образом, изобретение может быть использовано для лечения, предотвращения, замедления прогрессирования и/или ослабления симптома опосредованных B-клетками заболеваний или нарушений, включая отторжение трансплантата, болезнь «трансплантат против хозяина» (GvHD), ревматоидный артрит (RA).), рассеянный склероз (MS) и системная красная волчанка (SLE). На фигуре 5 показана способность предпочтительного Fc-диатела CD32B × CD79B снижать ксеногенный GvHD у мыши (см. WO 2015/021089, включенную в настоящий документ ссылкой). Подобным образом, связывающие молекулы CD32B × CD79B по изобретению могут быть использованы для снижения или ингибирования опосредованных B-клетками иммунных ответов (например, ответа на антигены, включая аутоантигены), для ослабления активации B-клеток и/или для уменьшения или ингибирования пролиферации B-клеток.
Способы введения
Композиции по настоящему изобретению могут быть предоставлены для лечения, профилактики и ослабления одного или нескольких симптомов, связанных с заболеванием, расстройством или инфекцией, путем введения объекту эффективного количества фармацевтической композиции по изобретению. В предпочтительном аспекте такие композиции по существу очищаются (т.е. практически не содержат веществ, которые ограничивают их действие или вызывают нежелательные побочные эффекты). В конкретном воплощении объектом является животное, предпочтительно млекопитающее, такое как не примат (например, бык, лошадь, кошка, собачий, грызун и т.д.) или примат (например, обезьяна, такая как яванский макак, человек и т.д.). В предпочтительном воплощении объект является человеком.
Доза введения и «режим дозирования» (частота введения) связывающих молекул CD32B × CD79B по изобретению могут быть уменьшены или изменены путем усиления поглощения и проникновения в ткани таких связывающих молекул посредством таких модификаций, как, например, липидизация. В одном воплощении один уровень дозировки (см. ниже) будет вводиться один или несколько раз в течение курса терапии. Во втором воплощении дозировка, обеспечиваемая в течение курса лечения, будет варьировать, например, в режиме с возрастающей дозировкой или в режиме с уменьшающейся дозировкой. Вводимая доза может быть дополнительно откорректирована, чтобы отразить терпимость объекта к терапии и степень терапевтического успеха, связанного с терапией.
Доза связывающих молекул CD32B × CD79B по изобретению, которая будет эффективной при лечении, профилактике или ослаблении одного или нескольких симптомов, связанных с расстройством, может быть определена стандартными клиническими методами. Точная доза, которая будет использоваться в составе, будет также зависеть от способа введения и от тяжести состояния и должна определяться в соответствии с суждением практикующего врача и обстоятельствами каждого пациента. Эффективные дозы могут быть экстраполированы из кривых доза-ответ, полученных из in vitro тест-систем или тест-систем на животных. Однако связывающие молекулы CD32B × CD79B по изобретению предпочтительно вводят в дозировке, которая обычно составляет, по меньшей мере, около 0,1 мг/кг, по меньшей мере, около 0,2 мг/кг, по меньшей мере, около 0,3 мг/кг, по меньшей мере, около 0,5 мг./кг, по меньшей мере, около 1,0 мг/кг, по меньшей мере, около 3,0 мг/кг, по меньшей мере, около 5,0 мг/кг, по меньшей мере, около 7,5 мг/кг, по меньшей мере, около 10,0 мг/кг, по меньшей мере, около 15 мг/кг, по меньшей мере, около 20 мг/кг или более от массы тела объекта. В частности, связывающей молекулы CD32B × CD79B по изобретению вводят в дозировке, которая составляет около 1,0 мг/кг, около 3,0 мг/кг, около 10,0 мг/кг, около 20,0 мг/кг или около 30,0 мг/кг. Связывающие молекулы CD32B × CD79B и, в частности, любые из диател CD32B × CD79B или Fc-диател по изобретению предпочтительно упаковывают в герметически закрытый контейнер, такой как ампула или саше с указанием количества таких молекул. При использовании в данном документе дозировка указывается как «около» относительно указанной дозировки, если описана в значимых цифрах, используемых для описания указанной дозировки (например, дозировка составляет около 0,1 мг/кг, если она составляет ± 0,05 мг/кг такой дозы, и доза составляет около 15 мг/кг, если она составляет ± 0,5 мг/кг такой дозы).
Частота введения связывающих молекул CD32B × CD79B по изобретению может существенно варьироваться, в зависимости, например, от реакции пациента или способа введения. Таким образом, композиции по изобретению могут вводиться один раз в день, два раза в день или три раза в день, один раз в неделю, два раза в неделю, один раз каждые две недели, один раз каждые три недели, один раз каждые четыре недели, один раз в месяц раз в шесть недель, раз в двенадцать недель, раз в два месяца, раз в три месяца, два раза в год, раз в год и т.д. Также следует понимать, что эффективная дозировка молекул, используемых для лечения, может увеличиваться или уменьшаться в течение определенного лечения.
Однако предпочтительно вводить связывающую молекулу CD32B × CD79B по изобретению в ходе курса лечения 1 недели, 2 недели, 3 недели, 4 недели, 5 недель, 6 недель, 7 недель, 8 недель, 9 недель, 10 недели, 11 недель, 12 недель, 13 недель, 14 недель, 15 недель или более 15 недель, и такой однократный курс лечения может повторяться 1, 2, 3, 4, 5 или более раз. В предпочтительном воплощении объекта лечат курсом терапии молекулами по изобретению один раз в 2 недели (Q2W), один раз в 3 недели (Q3W), один раз в 4 недели (Q4W), один раз в 5 недели (Q5W), один раз в 6 недель (Q6W), один раз в 7 недель (Q7W), один раз в 8 недель (Q8W), один раз в 9 недель (Q9W), один раз в 10 недель (Q10W), один раз в 11 недель (Q11W) или один раз в 12 недель (Q2W). Особенно предпочтительно вводить связывающей молекулы CD32B × CD79B по изобретению в ходе курса терапии один раз в 2 недели (Q2W), один раз в 3 недели (Q3W) или один раз в 4 недели (Q4W). По завершении любого такого единственного курса терапии терапия может быть возобновлена при той же схеме дозирования или при другой схеме дозировки и может включать ту же дозу или другую дозу введенной связывающей молекулы CD32B x CD79B. Таким образом, лечение объекта терапевтически или профилактически эффективным количеством связывающей молекулы CD32B × CD79B по изобретению может включать один курс лечения или может включать несколько курсов лечения, которые могут быть такими же или отличаться от любого предшествующего курса лечения.
В предпочтительных воплощениях связывающую молекулу CD32B × CD79B, связывающие молекулы CD32B × CD79B и, в частности, любое из диател CD32B × CD79B или Fc-диател по изобретению вводят в дозе около 3,0 мг/кг, около 10,0 мг/кг, около 20,0 мг/кг или около 30,0 мг/кг на курс терапии Q2W, Q3W или Q4W.
В одном воплощении Fc-диатела CD32B × CD79B по изобретению поставляются в виде сухого стерилизованного лиофилизированного порошка или безводного концентрата в герметически закрытом контейнере и могут быть восстановлены, например, водой или солевым раствором до подходящей концентрации для введения объекту. Предпочтительно, связывающие молекулы CD32B × CD79B и, в частности, любые из диател CD32B × CD79B или Fc-диател по изобретению поставляются в виде сухого стерильного лиофилизированного порошка в герметически закрытом контейнере в стандартной дозировке, по меньшей мере, 1 мг, более предпочтительно, по меньшей мере, 2 мг, по меньшей мере, 3 мг, по меньшей мере, 5 мг, по меньшей мере, 10 мг, по меньшей мере, 20 мг, по меньшей мере, 30 мг, по меньшей мере, 50 мг, по меньшей мере, 100 мг, по меньшей мере, 200 мг, по меньшей мере, 300 мг, по меньшей мере, 500 мг или, по меньшей мере, 1000 мг, так что, например, при добавлении подходящего объема носителя может быть подготовлена вводимая дозировка 0,1 мг/кг, 0,3 мг/кг, 1 мг/кг или 10 мг/кг.
Лиофилизированные связывающие молекулы CD32B × CD79B и, в частности, любые из диател CD32B × CD79B или Fc-диател по изобретению следует хранить при температуре от 2°C до 8°C в их исходном контейнере, и молекулы следует вводить в течение 12 часов, предпочтительно в течение 6 часов, в течение 5 часов, в течение 3 часов или в течение 1 часа после восстановления. В альтернативном воплощении такие молекулы поставляются в жидкой форме в герметично закрытом контейнере, указывающем количество и концентрацию молекулы, гибридного белка или конъюгированной молекулы. Предпочтительно жидкая форма связывающих молекул CD32B × CD79B по изобретению поставляется в герметически закрытом контейнере, где молекулы присутствуют в концентрации, по меньшей мере, 1 мкг/мл, более предпочтительно, по меньшей мере, 2,5 мг/мл, по меньшей мере, 5 мг/мл, по меньшей мере, 10 мг/мл, по меньшей мере, 50 мг/мл, по меньшей мере, 100 мг/мл или, по меньшей мере, 200 мг/мл.
В одном воплощении дозировку связывающих молекул CD32B × CD79B по изобретению, вводимых пациенту, можно рассчитать для применения в виде терапии одним агентом. В другом воплощении связывающие молекулы по изобретению используются в комбинации с другими терапевтическими композициями, и дозировка, вводимая пациенту, ниже, чем, когда такие связывающие молекулы используются в виде терапии с одним агентом.
Предпочтительные способы введения связывающих молекул CD32B × CD79B и, в частности, любого из диател CD32B × CD79B или Fc-диател по изобретению, включают, без ограничения указанным, парентеральное введение (например, внутрикожное, внутримышечное, внутрибрюшинное, внутривенное и подкожное), эпидуральное и через слизистую (например, интраназальным и пероральным путями). В конкретном воплощении изобретения молекулы по изобретению вводят внутримышечно, внутривенно или подкожно. Композиции можно вводить любым удобным способом, например, путем инфузии или болюсной инъекцией, путем абсорбции через эпителиальные или слизистые выстилки (например, слизистую оболочку полости рта, прямой кишки и слизистую оболочку кишечника и т.д.) и могут быть введены вместе с другими биологически активными агента. Введение может быть системным или местным.
В конкретном воплощении может быть желательным вводить фармацевтические композиции по изобретению локально в область, нуждающуюся в лечении; это может быть достигнуто, например, а не путем ограничения, локальной инфузией, инъекцией или с помощью имплантата, причем указанный имплантат выполняется из пористого, непористого или желатинового материала, включая мембраны, такие как мембраны sialastic или волокна. Предпочтительно, при введении молекулы по изобретению следует проявлять осторожность при использовании материалов, которые могут абсорбировать молекулу.
Воплощения изобретения
Ниже приведены неограничивающие примеры некоторых воплощений изобретения.
Воплощение 1. Способ лечения воспалительного заболевания или состояния, который включает введение терапевтически эффективного количества связывающей молекулы CD32B × CD79B объекту, нуждающемуся в этом, где указанная связывающая молекула CD32B × CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B, и где указанную связывающую молекулу CD32B × CD79B вводят в дозе от около 1 мг/кг до около 30 мг/кг и в режиме дозирования от одной дозы в неделю до одной дозы в 8 недель.
Воплощение 2. Способ снижения или ингибирования В-клеточных иммунных ответов, который включает введение терапевтически эффективного количества связывающей молекулы CD32B × CD79B объекту, нуждающемуся в этом, где указанная связывающая молекула CD32B × CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B и где указанную связывающую молекулу CD32B × CD79B вводят в дозе от около 1 мг/кг до около 30 мг/кг и в режиме дозирования от одной дозы в неделю до одной дозы в 8 недель.
Воплощение 3. Способ ослабления активации B-клеток, который включает введение терапевтически эффективного количества связывающей молекулы CD32B × CD79B объекту, нуждающемуся в этом, где указанная связывающая молекула CD32B × CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B и где указанную связывающую молекулу CD32B × CD79B вводят в дозе от около 1 мг/кг до около 30 мг/кг и в режиме дозирования от одной дозы в неделю до одной дозы в 8 недель.
Воплощение 4. Способ уменьшения или ингибирования пролиферации B-клеток, который включает введение терапевтически эффективного количества связывающей молекулы CD32B × CD79B объекту, нуждающемуся в этом, где указанная связывающая молекула CD32B × CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B, и где указанную связывающую молекулу CD32B × CD79B вводят в дозе от около 1 мг/кг до около 30 мг/кг и в режиме дозирования от одной дозы в неделю до одной дозы в 8 недель.
Воплощение 5. Способ по любому из воплощений 1-4, где указанную связывающую молекулу CD32B × CD79B вводят в дозе около 1 мг/кг.
Воплощение 6. Способ по любому из воплощений 1-4, где указанную связывающую молекулу CD32B × CD79B вводят в дозе около 3 мг/кг.
Воплощение 7. Способ по любому из воплощений 1 или 4, где указанную связывающую молекулу CD32B × CD79B вводят в дозе около 10 мг/кг.
Воплощение 8. Способ по любому из воплощений 1-7, где указанный режим дозирования составляет одну дозу за 2 недели (Q2W).
Воплощение 9. Способ по любому из воплощений 1-7, где указанный режим дозирования составляет одну дозу за 3 недели (Q3W).
Воплощение 10. Способ по любому из воплощений 1-7, где указанный режим дозирования составляет одну дозу в 4 недели (Q4W).
Воплощение 11. Способ по любому из воплощений 1-10, где указанная связывающая молекула CD32B × CD79B представляет собой биспецифическое антитело, которое связывает эпитоп CD32B и эпитоп CD79B, или молекулу, которая включает CD32B- и CD79B-связывающие домены указанного антитела.
Воплощение 12. Способ по любому из воплощений 1-11, где указанная связывающая молекула CD32B × CD79B представляет собой биспецифичное диатело CD32B × CD79B, которое связывает эпитоп CD32B и эпитоп CD79B.
Воплощение 13. Способ по воплощению 12, где указанное биспецифичное диатело CD32B × CD79B представляет собой Fc-диатело CD32B × CD79B.
Воплощение 14. Способ по любому из воплощений 1 или 5-13, где указанное воспалительное заболевание или состояние представляет собой аутоиммунное заболевание.
Воплощение 15. Способ по воплощению 14, где указанное аутоиммунное заболевание выбрано из группы, которая включает: болезнь Аддисона, аутоиммунный гепатит, аутоиммунное заболевание внутреннего уха, миастению, болезнь Крона, дерматомиозит, семейный аденоматозный полипоз, болезнь трансплантат против хозяина (GvHD), болезнь Грейвса, тиреоидит Хашимото, красную волчанку, рассеянный склероз (MS); пернициозную анемию, синдром Рейтера, ревматоидный артрит (RA), синдром Шегрена, системную красную волчанку (SLE), диабет 1 типа, первичный васкулит (например , ревматическую полимиалгию, гигантоклеточный артериит, болезнь Бехчета), пемфигус, оптикомиелит, анти-NMDA-рецепторный энцефалит, синдром Гийена-Барре, хроническую воспалительную демиелинизирующую полиневропатию (CIDP), офтальмопатию Грейвса, связанные с IgG4 заболевания, идиопатическую тромбоцитопеническую пурпуру (ITP) и язвенный колит.
Воплощение 16. Способ по воплощению 15, где указанное воспалительное заболевание или состояние представляет собой GvHD, RA, MS или SLE.
Воплощение 17. Способ по любому из воплощений 1-16, где уровень иммуноглобулина в сыворотке снижается к 36 дню после введения первой дозы указанной связывающей молекулы CD32B × CD79B.
Воплощение 18. Способ по воплощению 17, где указанным иммуноглобулином является IgM, IgA или IgG.
Воплощение 19. Способ по воплощению 18, где указанным иммуноглобулином является IgM.
Воплощение 20. Способ по любому из воплощений 1-19, где BCR-опосредуемая активация периферических B-клеток ингибируется через 24 часа после введения первой дозы указанной связывающей молекулы CD32B × CD79B, где указанная активация B-клеток определяется ex vivo анализом мобилизации кальция.
Воплощение 21. Способ по воплощению 20, где указанная BCR-опосредованная активация B-клеток ингибируется, по меньшей мере, на 50%, и где указанное ингибирование сохраняется в течение, по меньшей мере, 6 дней.
Воплощение 22. Способ по любому из воплощений 1-21, где, по меньшей мере, 20% сайтов связывания CD32B × CD79B на периферической B-клетке заняты через 6 часов после введения первой дозы указанной связывающей молекулы CD32B × CD79B.
Воплощение 23. Способ по любому из воплощений 1-22, где:
(А) экспрессия CD40 на В-клетках подавляется; и/или
(B) CD40-опосредованная секреция IgG ингибируется.
Воплощение 24. Способ по любому из воплощений 1-23, где указанный объект представляет собой человека.
Воплощение 25. Способ по любому из воплощений 1-24, где указанная связывающая молекула CD32B × CD79B содержит:
(A) домен VLCD32B, который включает аминокислотную последовательность SEQ ID NO: 30; и
(B) домен VHCD32B, который включает аминокислотную последовательность SEQ ID NO: 31.
Воплощение 26. Способ по любому из воплощений 1-24, где указанная связывающая молекула CD32B × CD79B содержит:
(A) домен VLCD79B, который включает аминокислотную последовательность SEQ ID NO: 32; и
(B) домен VHCD79B, который включает аминокислотную последовательность SEQ ID NO: 33.
Воплощение 27. Способ по любому из воплощений 1-24, где указанная связывающая молекула CD32B × CD79B содержит:
(A) домен VLCD32B, который включает аминокислотную последовательность SEQ ID NO: 30;
(B) домен VHCD32B, который включает аминокислотную последовательность SEQ ID NO: 31;
(C) домен VLCD79B, который включает аминокислотную последовательность SEQ ID NO: 32; и
(D) домен VHCD79B, который включает аминокислотную последовательность SEQ ID NO: 33.
Воплощение 28. Способ по любому из воплощений 25-27, где указанная связывающая молекула CD32B × CD79B представляет собой биспецифическое антитело или его биспецифический антигенсвязывающий фрагмент.
Воплощение 29. Способ по любому из вариантов 25-27, где указанная связывающая молекула CD32B × CD79B представляет собой биспецифичное диатело CD32B × CD79B.
Воплощение 30. Способ по воплощению 29, где указанное биспецифичное диатело CD32B × CD79B представляет собой Fc-диатело CD32B × CD79B.
Воплощение 31. Способ по воплощению 30, где указанное Fc-диатело CD32B × CD79B включает:
(А) первую полипептидную цепь, которая включает аминокислотную последовательность SEQ ID NO: 39;
(B) вторую полипептидную цепь, которая включает аминокислотную последовательность SEQ ID NO: 41; и
(С) третью полипептидную цепь, которая включает аминокислотную последовательность SEQ ID NO: 44.
Теперь, имея в целом описанное изобретение, то же самое будет легче понять со ссылкой на представленные далее примеры, которые предлагаются в качестве иллюстрации и не предназначены для ограничения настоящего изобретения, если не указано иное.
Примеры
Пример 1
Конструирование биспецифических моновалентных Fc-диател CD32B × CD79B и контрольных диател
Fc-диатело CD32B × CD79B было приготовлено и использовано в качестве примера связывающей молекулы CD32B × CD79B по изобретению. Fc-диатело CD32B × CD79B содержало три полипептидные цепи, имеющие аминокислотные последовательности, показанные в таблице 1.
Таблица 1
(SEQ ID NO: 39)
SEQ ID NO: 38
SEQ ID NO: 28
SEQ ID NO: 30
SEQ ID NO: 14
SEQ ID NO: 33
SEQ ID NO: 16
SEQ ID NO: 21
SEQ ID NO: 29
(SEQ ID NO: 41)
SEQ ID NO: 14
SEQ ID NO: 31
SEQ ID NO: 16
SEQ ID NO: 22
(SEQ ID NO: 44)
SEQ ID NO: 43
Было обнаружено, что описанное выше Fc-диатело CD32B × CD79B способно одновременно связываться с CD32B и с CD79B. Способы получения биспецифических моновалентных диател представлены в WO 2006/113665, WO 2008/157379, WO 2010/080538, WO 2012/018687, WO 2012/162068 и WO 2012/162067.
Пример 2
Оценка in vivo введения биспецифичных Fc-диател CD32B × CD79B
Для оценки безопасности и переносимости связывающих молекул CD32B × CD79B по настоящему изобретению Fc-диатело CD32B × CD79B из Примера 1 вводили здоровым людям в возрасте 18-50 лет с BMI 18-30 кг/м2. Объекты не включали беременных женщин или женщин с детородным потенциалом. Кроме того, объекты не включали индивидуумов, имеющих серьезные острые или хронические заболевания, индивидуумов, которые принимали какие-либо рецептурные лекарства в течение 4 недель после введения дозы или которые принимали лекарства, отпускаемые без рецепта, в течение 1 недели после введения дозы, индивидуумов, которые курят более 10 сигарет в день, индивидуумов, страдающих туберкулезом, инфекциями гепатита В, гепатита С или ВИЧ-инфекциями, индивидуумов, имеющих известную историю аутоиммунных или сосудистых заболеваний, или индивидуумов, имеющих положительный результат теста на наркотики. Объекты дополнительно не включали индивидуумов с корректированным QT (QTc) более 450 мсек, частотой сердечных сокращений менее 45 ударов в минуту или более 120 ударов в минуту, систолическим артериальным давлением (САД) более 140 мм рт.ст. или диастолическим артериальным давлением (ДАД) более 90 мм рт. Базовая демография объектов, вовлеченных в исследование, представлена в таблице 2.
Таблица 2
Оценка включала использование 6 дозовых когорт (каждая из которых состояла из 8 объектов, из которых объектам 1-6 вводили Fc-диатело CD32B × CD79B, а объектам 7-8 назначали плацебо). Внутри каждой когорты введение Fc-диатела CD32B × CD79B было осуществлено в шахматном порядке, так что объект 2 получал лечение через 24 часа после объекта 1, объекты 3-5 получали лечение через 24 часа после объекта 2, а объекты 7-8 получали лечение через 24 часа после объектов 3-5. Дозовые когорты описаны в таблице 3.
Таблица 3
Объекты после введения подвергались мониторингу на наличие побочных эффектов, связанных с Fc-диателом CD32B × CD79B: сердечная блокада второй или третьей степени, желудочковая аритмия (включая трепетание-мерцание) или ≥ побочных эффектов 3 степени согласно Руководству по шкале оценки токсичности для здоровых взрослых и подростков-добровольцев, отобранных в клиническое исследование профилактической вакцины. Для неохваченных проблем (например, реакция, связанная с инфузией) был использован NCI CTCAE v4.03. Было отмечено пятнадцать побочных эффектов, из которых 4 считались связанными с введением Fc-диатела CD32B × CD79B. Таблица 4 суммирует наблюдаемые побочные эффекты.
Таблица 4
(Предпочтительный термин MedRa)
А. Фармакодинамические эффекты Fc-диатела на гуморальные иммунные реакции
На фигуре 6 показана фармакокинетика типичной связывающей молекулы CD32B × CD79B при введении людям. Концентрации связывающей молекулы измеряли с помощью валидированного анализа ELISA, а параметры PK рассчитывали из не-компартментных анализов. Как показано на фигуре, объекты получали Fc-диатело CD32B × CD79B в дозировках 0,01 мг/кг, 0,1 мг/кг, 0,3 мг/кг, 1 мг/кг, 3 мг/кг или 10 мг/кг массы тела, и концентрация диатела в сыворотке крови измеряли в течение максимум 57 дней после введения. Обнаружено, что концентрация сыворотки диатела, максимальная концентрация сыворотки (Cmax) и AUC (площадь под кривой) увеличиваются с увеличением концентрации дозы. Период полувыведения диатела составлял от 4 до 8 дней. Быстрое распределение диатела наблюдалось при самой низкой дозе (0,01 мг/кг). Среднее время очищения (CL) варьировалось от 1,426 мл/ч/кг (доза 0,01 мг/кг) до 0,350 мл/ч/кг (доза 10 мг/кг), и соотношение между CL и дозой было нелинейным, с CL, снижающимся с увеличением дозы. Установлено, что объем распределения в стационарном состоянии (Vss), являющийся показателем распределения диатела в объеме крови, не зависит от дозы. Период полувыведения из сыворотки (T1/2) варьировал от 92 часов (~4 дня) для диатела, вводимого в дозе 0,03 мг/кг, до 191 часа (~8 дней) для диатела, вводимого в дозе 10 мг/кг, и увеличивается с увеличением дозы, что согласуется с дозозависимым снижением CL. Были отмечены нарушения в профилях PK, что предполагает наличие антител против лекарственного средства (ADA). Фармакокинетические данные приведены в таблице 5.
Таблица 5
(мг/кг)
(мкг/мл)
(ч*мкг/мл)
(мл/ч/кг)
(мл/кг)
(ч)
В. Способность связываться с CD32B и CD79B на периферических B-клетках
Было обнаружено, что введенная связывающая молекула CD32B × CD79B способна связываться с периферическими B-клетками, демонстрируя максимальную загрузку ≥1 мг/кг массы тела, при устойчивой загрузке 50% B-клеток при более высоких уровнях дозы. Устойчивая 20% загрузка B-клеток наблюдалась при ≥0,3 мг/кг. Фигура 7 суммирует анализ проточной цитометрией ex vivo на связывание с периферическими В-клетками в течение курса исследования.
С. Оценка статуса активации периферических B-клеток и подмножеств B-клеток
Проточный цитометрический анализ был проведен для того, чтобы оценить, было ли введение диатела связано с устойчивыми изменениями в количестве B-клеток (фигура 8A), количестве T-клеток (фигура 8B), соотношении B-клеток и T-клеток (фигура 8C) и отношение CD4 + T-клеток к CD8 + T-клеток (фигура 8D). Как показано на фигурах 8А-8В, устойчивых изменений в популяциях периферических Т-клеток не наблюдалось, временное уменьшение популяций В-клеток наблюдалось при более высоких дозах в этом исследовании. Как показано на фигурах 8C-8D, устойчивых изменений соотношений таких периферических B- и T-клеточных популяций не наблюдалось.
D. Оценка ответа периферических В-клеток на стимуляцию BCR ex vivo
Ex vivo анализ мобилизации кальция был проведен с целью оценки функции В-клеток реципиентов в ответ на примерную связывающую молекулу CD32B × CD79B. Вкратце, РВМС были свежевыделены из образцов крови, собранных в различные моменты времени до и после введения дозы, и поток Ca++ индуцировали лигированием BCR. Иономицин был введен для того, чтобы вызвать максимальный поток Са++. Значения AUC или пика были нормализованы с помощью значений, полученных с помощью иономицина, так что отношение (AUC) = AUC (IgM)/AUC (иономицин). Фигуры 9А-9В иллюстрируют экспериментальную процедуру и способы анализа данных, используемые в этом исследовании.
Обнаружено, что обработка примерной связывающей молекулы CD32B × CD79B снижает поток кальция в ответ на лигирование BCR анти-IgM антителом, демонстрируя, таким образом, ингибирующую активность связывающих молекул CD32B × CD79B по настоящему изобретению на периферических B-клетках (фигура 9C -9D).
E. Связывающие молекулы CD32B × CD79B подавляют экспрессию BCR на CD27+ B-клетках памяти
Чтобы дополнительно оценить эффект от введения примерной связывающей молекулы CD32B × CD79B, проточную цитометрию использовали для определения уровней мембраносвязанного иммуноглобулина. На фигурах 10A-10C показано, что введение связывающих молекул CD32B × CD79B подавляет экспрессию BCR на CD27+ B-клетках памяти, что определяется по экспрессии мембраносвязанного IgG (mIgG) (фигура 10A), мембраносвязанного IgM (mIgM) (фигура 10В) и мембраносвязанного IgD (mIgD) (фигура 10С).
Аналогичные исследования показали, что введение связывающих молекул CD32B × CD79B подавляет экспрессию BCR на наивных CD27-В-клетках памяти (фигура 11A: мембраносвязанный IgD (mIgD); фигура 11B: мембраносвязанный IgM; фигура 11C: процентное изменение мембраносвязанного IgM (mIgM)).
F. Связывающие молекулы CD32B × CD79B модулируют уровни сывороточных Ig
Оценивали влияние введения связывающей молекулы CD32B × CD79B на сывороточные уровни иммуноглобулинов IgM, IgA и IgG. Было обнаружено, что связывающая молекула модулирует сывороточные уровни этих иммуноглобулинов, при этом уровни IgM, демонстрируют наибольшее снижение, а уровень IgG в основном сохраняется (фигуры 12A-12C, соответственно). Сберегание IgG желательно. Снижение уровня сывороточного IgM предполагает воздействие на плазмобласты. Эти результаты согласуются с экспрессией CD79B, который не экспрессируется на плазматических клетках.
G. Связывающие молекулы CD32B × CD79B снижают уровень костимулирующей молекулы, CD40
Было обнаружено, что введение связывающей молекулы CD32B × CD79B снижает поверхностные уровни экспрессии CD40 на В-клетках (фигура 13А), что определяется с помощью проточного цитометрического анализа поверхностных костимулирующих молекул периферических В-клеток.
Однократная внутривенная доза типичной связывающей молекулы (Fc-диатело CD32B × CD79B из Примера 1) существенно не уменьшала количество периферических B-клеток, что указывает на то, что связывающие молекулы по настоящему изобретению не истощают B-клетки. Проточный цитологический анализ продемонстрировал, что связывающей молекулы по настоящему изобретению способны связываться с периферическими В-клетками. Полное насыщение таких сайтов связывания на периферических В-клетках наблюдалось при уровнях ≥1 мг/кг. Было установлено, что продолжительность такого связывания связана с используемым уровнем дозы.
Связывающие молекулы по настоящему изобретению опосредуют множество фармакодинамических дозозависимых эффектов, включая:
1. снижение активности in vivo BCR-индуцированной мобилизации Ca++;
2. отрицательную регуляцию поверхностной экспрессии IgG-BCR на подмножестве CD27+ В-клеток памяти;
3. уменьшение популяции IgM+ наивных В-клеток;
4. отрицательную регуляцию поверхностной экспрессии IgD-BCR на наивных В-клетках; и
5. отрицательную регуляцию экспрессии CD40 на В-клетках.
Связывающие молекулы по настоящему изобретению опосредуют множество благоприятных фармакодинамических характеристик, подходящих для клинического дозирования. Периферические В-клетки полностью насыщаются примерным Fc-диателом CD32B × CD79B из Примера 1 при уровнях дозы ≥1 мг/кг, а продолжительность связывания коррелирует с повышением уровня дозировки при устойчивой 20% загрузке, наблюдаемой при уровнях дозы ≥0,3 мг/кг. Связывающие молекулы по настоящему изобретению не истощают периферические B-клетки, и было обнаружено, что они отрицательно модулируют активность B-клеток дозозависимым образом:
1. снижение BCR-опосредованного притока Ca2+;
2. отрицательная регуляция поверхностной экспрессии иммуноглобулина;
3. снижение уровня сывороточного IgM; и
4. отрицательная регуляция экспрессии CD40.
Данные подтверждают полезность настоящего изобретения для лечения воспалительных заболеваний, состояний и расстройств и, в частности, для лечения аутоиммунных заболеваний.
Пример 3
Исследование фармакокинетических и фармакодинамических свойств связывающих молекул CD32B × CD79B
Модель Emax (максимальный эффект) PK/PD была использована для более полного исследования фарамкинетических (PK) и фармакодинамических (PD) свойств связывающих молекул CD32B × CD79B по изобретению. Вкратце, В-клетки инкубировали в присутствии различных концентраций связывающей молекулы, и определяли концентрации связывающей молекулы, приводящие к 50%, 60%, 80% и 90% ингибированию Emax.
Пациенты-реципиенты показали исходные значения, которые варьировались от 6,3% до 15,6% (среднее значение = 9,4%). Концентрация, дающая 50% максимального ответа (ЕС50), составила 1677 нг/мл. Когда концентрация введенной связывающей молекулы увеличилась, процент связывания B-клеток достиг плато, демонстрируя максимальный ответ (Emax) для % связывания B-клеток при 73,1%. Данные приведены в таблице 6. Исходя из in vivo модели гуманизированных мышей, прогнозируемая целевая концентрация, демонстрирующая полное ингибирование IgG и IgM, составляет 1500 нг/мл. Отношение PK/PD в исследовании предполагает, что концентрация ≥EC50 может быть потенциальной целевой концентрацией для терапевтического применения.
Таблица 6
На фигурах 14A-14B показаны данные (в двух разных диапазонах концентраций). На фигурах 15А-15F показаны доклинические целевые концентрации с наложенными фармакокинетическими профилями связывающей молекулы CD32B × CD79B у людей, для определения доз, которые достигли бы целевых концентраций (достижение целевой концентрации при дозировке 0,01 мг/кг (фигура 15А); при дозировке 0,1 мг/кг (фигура 15B); в дозировке 0,3 мг/кг (фигура 15C); в дозировке 1 мг/кг (фигура 15D); в дозировке 3 мг/кг (фигура 15E); в дозировке 10 мг/кг (фигура 15F)).
Были смоделированы фармакокинетические данные индивидуального объекта, и наилучшие оценки параметров модели были использованы для прогнозирования профилей для многократного введения при введении один раз в 2 недели (Q2W), один раз в 3 недели (Q3W) и один раз в 4 недели (Q4W). Исследованные дозы составляли 1, 3 и 10 мг/кг. Было проведено сравнение параметров воздействия (Cmax, Cmin и AUC) и исследованы в одно- и двухкамерной модели инфузии. Результаты такого моделирования показаны для доз 0,3 мг/кг массы тела объекта (фигура 16А), 1 мг/кг массы тела объекта (фигура 16В), массы тела объекта 3 мг/кг (фигура 16С) и 10 мг/кг. Масса тела объекта (фигура 16D). Масса тела объекта при приеме одной дозы в 2 недели (Q2W), одной дозы в 3 недели (Q3W) и одной дозы в 4 недели (Q4W). Предсказанная изменчивость в смоделированных выше профилях представлена (с SD) на фигурах 17A-17D.
Сводка статистики для параметров экспозиции после первой дозы (CMax1, Cmin1 и AUC1) и в стационарном состоянии (Сmaxss, Сminss и AUCss) приведены в таблицах 7, 8 и 9 для 1, 3 и 10 мг/кг дозы соответственно. Все 3 дозы вводились по схеме Q2W, Q3W и Q4W. В таблицах 7, 8, 9 и 10 AUC определяли в интервалах дозирования 14, 21 и 28 дней для режимов Q2W, Q3W и Q4W, соответственно.
Таблица 7. Моделируемые параметры экспонирования для дозы 0,3 мг/кг
(нг/мл)
(нг/мл)
(ч*нг/мл)
(нг/мл)
(нг/мл)
Таблица 8. Моделируемые параметры экспонирования для дозы 1 мг/кг
(нг/мл)
(нг/мл)
(ч*нг/мл)
(нг/мл)
(нг/мл)
(ч*нг/мл)
Таблица 9. Моделируемые параметры экспонирования для дозы 3 мг/кг
(нг/мл)
(нг/мл)
(ч*нг/мл)
(нг/мл)
(нг/мл)
(ч*нг/мл)
Таблица 10. Моделируемые параметры экспонирования для дозы 10 мг/кг
(нг/мл)
(нг/мл)
(ч*нг/мл)
(нг/мл)
(нг/мл)
(ч*нг/мл)
Как указано в таблицах 7, 8, 9 и 10 в отношении данных о первой дозе, значения среднего Cmax1 были одинаковыми для режимов Q2W, Q3W и Q4W (т.е. для режима дозирования 1 мг/кг - около 22 мкг/мл; для режима дозирования 3 мг/кг - около 54 мкг/мл и для режима дозирования 10 мг/кг - около 187 мкг/мл). Незначительные различия наблюдались в средних значениях AUC1 между 3 режимами. Было отмечено, что среднее Cmin1 (минимальная концентрация) была выше для режима Q2W (т.е. для режима дозирования 1 мг/кг, около 1,1 мкг/мл; для режима дозирования 3 мг/кг, около 5 мкг/мл и для режима дозирования 10 мг/кг, около 24 мкг/мл), который имел более короткий интервал дозирования (14 дней) по сравнению с режимом Q4W (т.е. для режима дозирования 1 мг/кг, около 0,14 мкг/мл; для режима дозирования 3 мг/кг - около 0,95 мкг/мл и для режима дозирования 10 мг/кг - около 5 мкг/мл), который имел более длительный интервал дозирования 28 дней. Было отмечено, что среднее значение Cmin1 для режима Q3W является промежуточным по сравнению с режимами Q2W и Q4W (т.е. для режима дозирования 1 мг/кг около 0,38 мкг/мл; для режима дозирования 3 мг/кг около 2,2 мкг/мл и для режима дозирования 10 мг/кг около 5 мкг/мл).
Как указано в таблицах 7, 8, 9 и 10 в отношении стационарного состояния, наблюдалось, что среднее значение Cmaxss, Cminss и AUCs численно выше по сравнению со значениями первой дозы и в соответствии с данными первой дозы средняя Cmaxss схем Q2W, Q3W и Q4W была одинаковой (разница <13%). Кроме того, в соответствии с данными первой дозы наблюдались небольшие различия в средних значениях AUCss между 3 режимами. Однако интервал дозирования для режима Q4W составляет 28 дней, и за тот же интервал количество доз, вводимых для режима Q2W, в 2 раза выше, чем в режиме Q4W. Таким образом, в течение 28-дневного интервала дозирования, AUCss для режима Q2W как ожидается, будет около в 2 раза выше, чем у режима Q4W. Независимо от режима введения параметры воздействия в стационарном состоянии предполагают минимальное накопление связывающей молекулы CD32B × CD79B по сравнению с данными первой дозы.
В целом, связывающие молекулы по настоящему изобретению вводили людям в дозах до 10 мг/кг и они продемонстрировали диапазон клинических доз от 0,3 мг/кг до 10 мг/кг. Однако более высокие дозы могут быть еще более эффективными. Для схем Q2W дозы ≥1 мг/кг должны достигать целевой концентрации. Для схем Q3W дозы ≥3 мг/кг должны достигать целевой концентрации. Однако более низкие дозы (например, 0,3 мг/кг) могут быть эффективными для достижения искомой биологической активности. Было обнаружено, что введение является безопасным для здоровых людей и демонстрирует иммуномодулирующую активность. Введение связывающих молекул по настоящему изобретению не сопровождалось или не связывалось с нежелательным высвобождением цитокинов. Независимо от введенной дозы, фармакокинетический/фармакодинамический анализ (PK/PD) и анализ моделирования и симуляции (M & S) показывают, что значения первой дозы или Cmax в стационарном состоянии аналогичны для режимов Q2W, Q3W и Q4W. В стационарном состоянии AUCss в течение 28-дневного интервала (интервал дозирования для режима Q2W), как ожидается, будет примерно в 2 раза выше для режима Q2W по сравнению с режимом Q4W. Значения Cmin для первой дозы или стационарного состояния для режимов Q2W и Q3W с более короткими интервалами дозирования (14 и 21 день соответственно) выше по сравнению с режимом Q4W, который имеет более длительный интервал дозирования 28 дней (более длительный период вымывания).
Хотя время полужизни в сыворотке (T1/2) связывающих молекул CD32B × CD79B по изобретению варьировалось от около 4-8 дней, даже после того, как закончился первый период полувыведения, было обнаружено, что значительная доля введенных связывающих молекул CD32B × CD79B остается связанной с периферическими В-клетками. Например, что касается Fc-диатела CD32B × CD79B из Примера 1, то загрузка более 20% наблюдалась в течение, по меньшей мере, 8 дней в режиме дозирования 0,3 мг/кг и в течение от 30 до почти 50 дней в режиме дозирования 3 мг/кг и 10 мг/кг.
Ингибирование активации периферических В-клеток (особенно отраженное в измерениях AUC) приводит к возвращению к исходному уровню примерно к 30 дню. Связывающие молекулы CD32B × CD79B также снижают активность мишени BCR на В-клетках памяти (фигуры 10А-10С) и на наивных В-клетках (фигуры 11А-11С), и эта отрицательная регуляция также сохраняется в течение примерно 27 дней или более. Аналогично, модуляция сывороточных уровней Ig (фигуры 12A-12C) сохраняется в течение, по меньшей мере, 57 дней даже при самой низкой дозе. Эти результаты получены от введения однократной дозы. Соответственно, предпочтительная схема дозировки может основываться на периоде полувыведения, биологической активности пролонгированного действия связывающих молекул CD32B × CD79B или на периоде полувыведения и такой биологической активности длительного действия.
Пример 4
Оценка in vivo введения биспецифичных Fc-диател CD32B × CD79B на гуморальные иммунные ответы
Как описано в данном документе, связывающие молекулы CD32B × CD79B, в частности биспецифичные Fc-диатела по настоящему изобретению предназначены для ингибирования активированных B-клеток путем запуска физиологической отрицательной петли обратной связи, которая основана на сопряжении активации-ингибирования. Может быть исследовано ингибирующее действие связывающих молекул CD32B × CD79B на гуморальные иммунные ответы в ответ на вакцинацию иммуногеном, таким как гемоцианин лимфы улитки (KLH). В частности, на основе экстраполяции данных PK от приматов, не являющихся человеком, при введении в дозе 0,3 мг/кг циркулирующего Fc-диатела CD32B × CD79B будет сохраняться уровень выше, для поддержания 20% загрузки сайтов связывания F32 CD32B × CD79B на В-клетках периферической крови в течение 6 дней. Основываясь на исследованиях in vitro, можно предположить, что такие уровни загрузки поддерживают как минимум 50% ингибирование BCR-опосредованной активации B-клеток в течение не менее 6 дней. Принимая во внимание, что иммунные ответы на вакцинацию KLH у людей занимают от около 4 до 6 дней, ожидается, что осуществление вакцинации KLH у подвергнутых введению связывающих молекул CD32B × CD79B в дозах ≥,3 мг/мл приведет к обнаруживаемой фармакодинамической активности, о чем свидетельствует ингибирование В-клеточных ответов на вакцинацию антигеном KLH. Соответственно, KLH можно вводить в подкожной (SC) дозе 1,0 мг в день 2 для всех объектов, получивших дозу связывающих молекул CD32B × CD79B 0,3 мг/кг или выше. Кроме того, вакцинация KLH может проводиться не менее чем через 24 часа после инфузии связывающей молекулы CD32B × CD79B, чтобы установить безопасность и переносимость такой связывающей молекулы CD32B × CD79B для каждого объекта до вакцинации.
Титры анти-KLH IgG и IgM и их процентное изменение ингибирования по сравнению с исходным уровнем, а также доля объектов, у которых были установлены количественные ответы против KLH, IgG и IgM после иммунизации, будут сведены в таблицу и суммированы в зависимости от дозы и времени. Различия в иммунных ответах между обработкой связывающей молекулой CD32B × CD79B и плацебо будут оцениваться с использованием описательной статистики. Дополнительные анализы (например, анализ воздействие-ответ) могут быть проведены, если это будет сочтено целесообразным.
Пример 5
Оценка воздействия in vivo введения биспецифичных Fc-диател CD32B × CD79B на гуморальные иммунные ответы
Как описано выше, могут быть оценены ингибирующие эффекты связывающих молекул CD32B × CD79B на гуморальные иммунные ответы в ответ на вакцинацию. Инактивированная вакцина против гепатита А (HAV) является признанным нео-антигеном, используемым для оценки реакции иммунизации при нарушениях иммунитета (Valdez H, et al. (2000) “Response To Immunization With Recall And Neoantigens After Prolonged Administration Of An Hiv-1 Protease Inhibitor-Containing Regimen,” ACTG 375 team. AIDS clinical trials group. AIDS 14: 11-21) и среди пациентов, которые используют терапию с истощением B-клеток (Van Der Kolk LE, et al.(2002) “Rituximab Treatment Results In Impaired Secondary Humoral Immune Responsiveness,” Blood 100:2257-9. В этом исследовании ингибирующие эффекты примерной связывающей молекулы CD32B × CD79B, описанной выше, оценивали в ответ на введение HAV нормальным здоровым добровольцам, имеющим отрицательный серологический титр гепатита А. Вкратце, однократную дозу связывающей молекулы CD32B × CD79B или плацебо вводили в дозе 3 мг/кг или 10 мг/кг посредством в.в. инфузии, объекты также получали однократную дозу HAV (VAQTA® (вакцина против гепатита A, инактивированная, Merck, 50 Ед/мл) внутримышечно в день 2. Влияние введения связывающей молекулы CD32B × CD79B на иммунные ответы на однократную дозу HAV оценивали путем мониторинга появления сывороточного IgG-специфического анти-HAV-специфического антитела у вакцинированных объектов. Для выявления наличия HAV-специфического IgG был использован анализ ARCHITECT HAVAb-IgG (Abbott Laboratories). Анализ ARCHITECT HAVAb-IgG представляет собой хемилюминесцентный иммуноанализ микрочастиц (CMIA) для качественного определения антител IgG к HAV в сыворотке или плазме человека. Также были использованы количественные анализы с модифицированной системой Эбботта.
Таблица 11 суммирует начальные результаты для 10 объектов, получавших плацебо или примерную связывающую молекулу CD32B × CD79B в дозе 3 мг/кг или 10 мг/кг. Таблица 12 суммирует результаты на 56 день после вакцинации от всех объектов исследования. Средняя концентрация HAV-специфического IgG (анти-HAV IgG), присутствующего в сыворотке каждой вакцинированной группы, суммирована в таблице 13. Концентрация анти-HAV IgG, присутствующего в сыворотке каждого из вакцинированных объектов, показана на фигуре 18. Результаты этого исследования показывают, что введение связывающих молекул CD32B × CD79B ингибирует активированные В-клетки и гуморальные иммунные ответы в ответ на вакцинацию.
Таблица 11
Таблица 12
Таблица 13
Пример 6
Связывающие молекулы CD32B × CD79B блокируют CD40-зависимые B-клеточные ответы
Как описано выше, введение связывающих молекул CD32B × CD79B снижает уровни костимулирующей молекулы CD40. Активность типичной связывающей молекулы CD32B × CD79B в отношении CD40-зависимых ответов оценивали in vitro. В этих исследованиях используется система анализа, которая имитирует процесс дифференцировки В-клеток в клетки, секретирующие антитела (например, IgG) в присутствии сигналов стимуляции, предоставляемых фолликулярными CD4-хелперами, что происходит в герминативном центре вторичных или территориальных лимфоидных органов. Вкратце, В-клетки человека очищали из цельной периферической крови здоровых доноров с использованием набора для отрицательного отбора. Очищенные В-клетки человека культивировали в полной среде RPMI1640 со стимуляторами или без них (CD40-лиганд (500 нг/мл), IL-4 (100 нг/мл) и IL-21 (20 нг/мл)), использованных в неразбавленном виде, или в виде 3-кратного серийного разведения (1, 1/3, 1/9 и 1/27) в присутствии или в отсутствие примерной связывающей молекулы CD32B × CD79B, описанной выше (20 мкг/мл) в 5% СО2 37°С в инкубаторе в течение 5 дней. Культуральные надосадочные жидоксти собирали и секретируемый человеческий IgG определяли с помощью ELISA. Результаты этого исследования представлены на фигуре 19.
Как показано на фигуре 19, связывающие молекулы CD32B × CD79B могут снижать секрецию IgG, указывая на то, что связывающей молекулы CD32B × CD79B могут ингибировать опосредованный CD40 путь, связанный с выработкой IgG.
Все публикации и патенты, упомянутые в этом описании, включены в данный документ ссылкой в той же степени, как если бы каждая отдельная публикация или патентная заявка были конкретно и индивидуально указаны для включения в качестве ссылки в полном объеме. Хотя изобретение было описано в связи с его конкретными воплощениями, следует понимать, что имеется возможность дальнейших модификаций, и эта заявка предназначена для охвата любых изменений, применений или адаптаций по изобретению, следуя, в общем, принципам изобретения и включая такие отклонения от настоящего раскрытия, которые входят в известную или обычную практику в области, к которой относится изобретение, и которые могут быть применены к основным признакам, изложенным выше.
--->
Список последовательностей
<110> MacroGenics, Inc.
Chen, Wei
Moore, Paul A.
Pandya, Namish Bharat
Bonvini, Ezio
<120> Methods for the Use of CD32B x CD79B-Binding Molecules in the
Treatment of Inflammatory Diseases and Disorders
<130> 1301.0145PCT
<150> US 62/432,328
<151> 2016-12-09
<150> US 62/346,717
<151> 2016-06-07
<160> 48
<170> PatentIn version 3.5
<210> 1
<211> 107
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(107)
<223> Human IgG CL Kappa Domain
<400> 1
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 2
<211> 104
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(104)
<223> Human IgG CL Lambda Domain
<400> 2
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
50 55 60
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
65 70 75 80
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
85 90 95
Thr Val Ala Pro Thr Glu Cys Ser
100
<210> 3
<211> 98
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(98)
<223> Human IgG1 CH1 Domain
<400> 3
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 4
<211> 98
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(98)
<223> Human IgG2 CH1 Domain
<400> 4
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val
<210> 5
<211> 98
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(98)
<223> Human IgG4 CH1 Domain
<400> 5
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 6
<211> 15
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(15)
<223> Human IgG1 Hinge Region
<400> 6
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 7
<211> 12
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(12)
<223> Human IgG2 Hinge Region
<400> 7
Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro
1 5 10
<210> 8
<211> 12
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(12)
<223> Human IgG4 Hinge Region
<400> 8
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
1 5 10
<210> 9
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Human IgG4 Hinge Region with S228P Substitution
<400> 9
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 10
<211> 217
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(217)
<223> CH2-CH3 Domain of Human IgG1
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> XAA is Lysine (K) or is absent
<400> 10
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 11
<211> 216
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(216)
<223> CH2-CH3 Domain of Human IgG2
<220>
<221> MISC_FEATURE
<222> (216)..(216)
<223> XAA is Lysine (K) or is absent
<400> 11
Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
1 5 10 15
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
20 25 30
Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
35 40 45
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
50 55 60
Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln
65 70 75 80
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
85 90 95
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro
100 105 110
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
115 120 125
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
130 135 140
Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
145 150 155 160
Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
165 170 175
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
180 185 190
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
195 200 205
Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 12
<211> 217
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(217)
<223> CH2-CH3 Domain of Human IgG3
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> XAA is Lysine (K) or is absent
<400> 12
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Xaa
210 215
<210> 13
<211> 217
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(217)
<223> CH2-CH3 Domain of Human IgG4
<220>
<221> MISC_FEATURE
<222> (217)..(217)
<223> XAA is Lysine (K) or is absent
<400> 13
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Xaa
210 215
<210> 14
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Preferred Intervening Spacer Peptide (Linker 1)
<400> 14
Gly Gly Gly Ser Gly Gly Gly Gly
1 5
<210> 15
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker 2
<400> 15
Ala Ser Thr Lys Gly
1 5
<210> 16
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker 2
<400> 16
Gly Gly Cys Gly Gly Gly
1 5
<210> 17
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Heterodimer-Promoting Domain
<400> 17
Gly Val Glu Pro Lys Ser Cys
1 5
<210> 18
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Heterodimer-Promoting Domain
<400> 18
Val Glu Pro Lys Ser Cys
1 5
<210> 19
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Heterodimer-Promoting Domain
<400> 19
Gly Phe Asn Arg Gly Glu Cys
1 5
<210> 20
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Heterodimer-Promoting Domain
<400> 20
Phe Asn Arg Gly Glu Cys
1 5
<210> 21
<211> 28
<212> PRT
<213> Artificial Sequence
<220>
<223> "E-coil" Heterodimer-Promoting Domain
<400> 21
Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
1 5 10 15
Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys
20 25
<210> 22
<211> 28
<212> PRT
<213> Artificial Sequence
<220>
<223> "K-coil" Heterodimer-Promoting Domain
<400> 22
Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val
1 5 10 15
Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
20 25
<210> 23
<211> 28
<212> PRT
<213> Artificial Sequence
<220>
<223> Cysteine-Containing "E-Coil" Heterodimer-Promoting Domain
<400> 23
Glu Val Ala Ala Cys Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
1 5 10 15
Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys
20 25
<210> 24
<211> 28
<212> PRT
<213> Artificial Sequence
<220>
<223> Cysteine-Containing "K-Coil" Heterodimer-Promoting Domain
<400> 24
Lys Val Ala Ala Cys Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val
1 5 10 15
Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
20 25
<210> 25
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Cysteine-Containing Peptide (Peptide 1)
<400> 25
Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10
<210> 26
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Cysteine-Containing Peptide (Peptide 1)
<400> 26
Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10
<210> 27
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Intervening Linker Peptide (Linker 3)
<400> 27
Ala Pro Ser Ser Ser
1 5
<210> 28
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Intervening Linker Peptide (Linker 3)
<400> 28
Ala Pro Ser Ser Ser Pro Met Glu
1 5
<210> 29
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Spacer Peptide (Linker 4)
<400> 29
Gly Gly Gly Asn Ser
1 5
<210> 30
<211> 107
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(107)
<223> VL Domain of Anti-Human CD32B Antibody
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 31
<211> 116
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(116)
<223> VH Domain of Anti-Human CD32B Antibody
<400> 31
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ala
20 25 30
Trp Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Glu Ile Arg Asn Lys Ala Lys Asn His Ala Thr Tyr Tyr Ala Glu
50 55 60
Ser Val Ile Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Gly Ala Leu Gly Leu Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 32
<211> 112
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(112)
<223> VL Domain of Anti-Human CD79B Antibody
<400> 32
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 33
<211> 113
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(113)
<223> VH Domain of Anti-Human CD79B Antibody
<400> 33
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
100 105 110
Ser
<210> 34
<211> 262
<212> PRT
<213> Artificial Sequence
<220>
<223> First Polypeptide Chain of First Exemplary CD32B x CD79B
Bispecific Diabody
<400> 34
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Lys Phe Lys Asp Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val
210 215 220
Thr Val Ser Ser Gly Gly Cys Gly Gly Gly Glu Val Ala Ala Leu Glu
225 230 235 240
Lys Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu
245 250 255
Val Ala Ala Leu Glu Lys
260
<210> 35
<211> 270
<212> PRT
<213> Artificial Sequence
<220>
<223> Second Polypeptide Chain of First Exemplary CD32B x CD79B
Bispecific Diabody
<400> 35
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Cys Gly
225 230 235 240
Gly Gly Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
245 250 255
Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
260 265 270
<210> 36
<211> 261
<212> PRT
<213> Artificial Sequence
<220>
<223> First Polypeptide Chain of Second Exemplary CD32B x CD79B
Bispecific Diabody
<400> 36
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Glu Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Trp Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Ile Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn
165 170 175
Gln Lys Phe Lys Asp Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser
180 185 190
Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val
210 215 220
Thr Val Ser Ser Ala Ser Thr Lys Gly Glu Val Ala Ala Cys Glu Lys
225 230 235 240
Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
245 250 255
Ala Ala Leu Glu Lys
260
<210> 37
<211> 269
<212> PRT
<213> Artificial Sequence
<220>
<223> Second Polypeptide Chain of Second Exemplary CD32B x CD79B
Bispecific Diabody
<400> 37
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
225 230 235 240
Gly Lys Val Ala Ala Cys Lys Glu Lys Val Ala Ala Leu Lys Glu Lys
245 250 255
Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
260 265
<210> 38
<211> 217
<212> PRT
<213> Artificial Sequence
<220>
<223> CH2-CH3 Domain of Knob-Bearing IgG Fc Region
<400> 38
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 39
<211> 502
<212> PRT
<213> Artificial Sequence
<220>
<223> First Polypeptide Chain of First Exemplary CD32B x CD79B
Bispecific Fc Diabody
<400> 39
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys Ala Pro Ser Ser Ser Pro Met Glu Asp Ile Gln Met Thr
225 230 235 240
Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile
245 250 255
Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr Leu Ser Trp Leu Gln
260 265 270
Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile Tyr Ala Ala Ser Thr
275 280 285
Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Glu Ser Gly Thr
290 295 300
Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr
305 310 315 320
Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu Thr Phe Gly Gly Gly
325 330 335
Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly Gly Gly Gly Gln Val
340 345 350
Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala Ser Val
355 360 365
Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Met
370 375 380
Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly Met
385 390 395 400
Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn Gln Lys Phe Lys Asp
405 410 415
Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr Met Glu
420 425 430
Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys Ala Arg
435 440 445
Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
450 455 460
Gly Cys Gly Gly Gly Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala
465 470 475 480
Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu
485 490 495
Lys Gly Gly Gly Asn Ser
500
<210> 40
<211> 1506
<212> DNA
<213> Artificial Sequence
<220>
<223> Polynucleotide Encoding First Polypeptide Chain of First
Exemplary CD32B x CD79B Bispecific Fc Diabody
<400> 40
gacaaaactc acacatgccc accgtgccca gcacctgaag ccgcgggggg accgtcagtc 60
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca 120
tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac 180
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac 240
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag 300
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa 360
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggagga gatgaccaag 420
aaccaggtca gcctgtggtg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 480
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 540
gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggg 600
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc 660
ctctccctgt ctccgggtaa agccccttcc agctccccta tggaagacat ccagatgacc 720
cagtctccat cctccttatc tgcctctgtg ggagatagag tcaccatcac ttgtcgggca 780
agtcaggaaa ttagtggtta cttaagctgg ctgcagcaga aaccaggcaa ggcccctaga 840
cgcctgatct acgccgcatc cactttagat tctggtgtcc catccaggtt cagtggcagt 900
gagtctggga ccgagttcac cctcaccatc agcagccttc agcctgaaga ttttgcaacc 960
tattactgtc tacaatattt tagttatccg ctcacgttcg gaggggggac caaggtggaa 1020
ataaaaggag gcggatccgg cggcggaggc caggttcagc tggtgcagtc tggagctgag 1080
gtgaagaagc ctggcgcctc agtgaaggtc tcctgcaagg cttctggtta cacctttacc 1140
agctactgga tgaactgggt gcgacaggcc cctggacaag ggcttgagtg gatcggaatg 1200
attgatcctt cagacagtga aactcactac aatcaaaagt tcaaggacag agtcaccatg 1260
accacagaca catccacgag cacagcctac atggagctga ggagcctgag atctgacgac 1320
acggccgtgt attactgtgc gagagctatg ggctactggg ggcaagggac cacggtcacc 1380
gtctcctccg gaggatgtgg cggtggagaa gtggccgcac tggagaaaga ggttgctgct 1440
ttggagaagg aggtcgctgc acttgaaaag gaggtcgcag ccctggagaa aggcggcggg 1500
aactct 1506
<210> 41
<211> 270
<212> PRT
<213> Artificial Sequence
<220>
<223> Second Polypeptide Chain of First Exemplary CD32B x CD79B
Bispecific Fc Diabody
<400> 41
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Cys Gly
225 230 235 240
Gly Gly Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
245 250 255
Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
260 265 270
<210> 42
<211> 810
<212> DNA
<213> Artificial Sequence
<220>
<223> Polynucleotide Encoding Second Polypeptide Chain of First
Exemplary CD32B x CD79B Bispecific Fc Diabody
<400> 42
gatgttgtga tgactcagtc tccactctcc ctgcccgtca cccttggaca gccggcctcc 60
atctcctgca agtcaagtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttcagcaga ggccaggcca atctccaaac cgcctaattt atctggtgtc taaactggac 180
tctggggtcc cagacagatt cagcggcagt gggtcaggca ctgatttcac actgaaaatc 240
agcagggtgg aggctgagga tgttggggtt tattactgct ggcaaggtac acattttccg 300
ctcacgttcg gcggagggac caagcttgag atcaaaggag gcggatccgg cggcggaggc 360
gaagtgcagc ttgtggagtc tggaggaggc ttggtgcaac ctggaggatc cctgagactc 420
tcttgtgccg cctctggatt cacttttagt gacgcctgga tggactgggt ccgtcaggcc 480
ccaggcaagg ggcttgagtg ggttgctgaa attagaaaca aagctaaaaa tcatgcaaca 540
tactatgctg agtctgtgat agggaggttc accatctcaa gagatgacgc caaaaacagt 600
ctgtacctgc aaatgaacag cttaagagct gaagacactg ccgtgtatta ctgtggggct 660
ctgggccttg actactgggg ccaaggcacc ctggtgaccg tctcctccgg aggatgtggc 720
ggtggaaaag tggccgcact gaaggagaaa gttgctgctt tgaaagagaa ggtcgccgca 780
cttaaggaaa aggtcgcagc cctgaaagag 810
<210> 43
<211> 217
<212> PRT
<213> Artificial Sequence
<220>
<223> CH2-CH3 Domain of Hole-Containing IgG Fc Region
<400> 43
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 44
<211> 227
<212> PRT
<213> Artificial Sequence
<220>
<223> Third Polypeptide Chain of First Exemplary CD32B x CD79B
Bispecific Fc Diabody
<400> 44
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 45
<211> 681
<212> DNA
<213> Artificial Sequence
<220>
<223> Polynucleotide Encoding Third Polypeptide Chain of First
Exemplary CD32B x CD79B Bispecific Fc Diabody
<400> 45
gacaaaactc acacatgccc accgtgccca gcacctgaag ccgcgggggg accgtcagtc 60
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca 120
tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac 180
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac 240
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag 300
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa 360
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggagga gatgaccaag 420
aaccaggtca gcctgagttg cgcagtcaaa ggcttctatc ccagcgacat cgccgtggag 480
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 540
gacggctcct tcttcctcgt cagcaagctc accgtggaca agagcaggtg gcagcagggg 600
aacgtcttct catgctccgt gatgcatgag gctctgcaca accgctacac gcagaagagc 660
ctctccctgt ctccgggtaa a 681
<210> 46
<211> 501
<212> PRT
<213> Artificial Sequence
<220>
<223> First Polypeptide Chain of Second Exemplary CD32B x CD79B
Bispecific Fc Diabody
<400> 46
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys Ala Pro Ser Ser Ser Pro Met Glu Asp Ile Gln Met Thr
225 230 235 240
Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile
245 250 255
Thr Cys Arg Ala Ser Gln Glu Ile Ser Gly Tyr Leu Ser Trp Leu Gln
260 265 270
Gln Lys Pro Gly Lys Ala Pro Arg Arg Leu Ile Tyr Ala Ala Ser Thr
275 280 285
Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Glu Ser Gly Thr
290 295 300
Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr
305 310 315 320
Tyr Tyr Cys Leu Gln Tyr Phe Ser Tyr Pro Leu Thr Phe Gly Gly Gly
325 330 335
Thr Lys Val Glu Ile Lys Gly Gly Gly Ser Gly Gly Gly Gly Gln Val
340 345 350
Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala Ser Val
355 360 365
Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Met
370 375 380
Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly Met
385 390 395 400
Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn Gln Lys Phe Lys Asp
405 410 415
Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr Met Glu
420 425 430
Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys Ala Arg
435 440 445
Ala Met Gly Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala
450 455 460
Ser Thr Lys Gly Glu Val Ala Ala Cys Glu Lys Glu Val Ala Ala Leu
465 470 475 480
Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys
485 490 495
Gly Gly Gly Asn Ser
500
<210> 47
<211> 269
<212> PRT
<213> Artificial Sequence
<220>
<223> Second Polypeptide Chain of Second Exemplary CD32B x CD79B
Bispecific Fc Diabody
<400> 47
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Asn Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly
115 120 125
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
130 135 140
Ser Gly Phe Thr Phe Ser Asp Ala Trp Met Asp Trp Val Arg Gln Ala
145 150 155 160
Pro Gly Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Asn Lys Ala Lys
165 170 175
Asn His Ala Thr Tyr Tyr Ala Glu Ser Val Ile Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Gly Ala Leu Gly Leu Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
225 230 235 240
Gly Lys Val Ala Ala Cys Lys Glu Lys Val Ala Ala Leu Lys Glu Lys
245 250 255
Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
260 265
<210> 48
<211> 227
<212> PRT
<213> Artificial Sequence
<220>
<223> Third Polypeptide Chain of Second Exemplary CD32B x CD79B
Bispecific Fc Diabody
<400> 48
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ СНИЖЕНИЯ ИММУНОГЕННОСТИ С ПОМОЩЬЮ НЕ ПРИВОДЯЩИХ К ДЕПЛЕЦИИ В-КЛЕТОЧНЫХ ИНГИБИТОРОВ | 2020 |
|
RU2821892C2 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ ADAM9, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783619C2 |
| МОЛЕКУЛЫ СО СПЕЦИФИЧНОСТЬЮ К CD45 И CD79 | 2015 |
|
RU2747980C2 |
| КОВАЛЕНТНЫЕ ДИАНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2744176C2 |
| МОЛЕКУЛЫ СО СПЕЦИФИЧНОСТЬЮ В ОТНОШЕНИИ CD79 И CD22 | 2015 |
|
RU2743590C2 |
| БИСПЕЦИФИЧНЫЕ МОЛЕКУЛЫ, ИММУНОРЕАКТИВНЫЕ С ИММУННЫМИ ЭФФЕКТОРНЫМИ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ АКТИВИРУЮЩИЙ РЕЦЕПТОР | 2014 |
|
RU2721707C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2733315C2 |
| БИСПЕЦИФИЧЕСКИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ БЕЛОК EGFR/CD16 | 2019 |
|
RU2792240C2 |
| АНТИТЕЛА ПРОТИВ CD79b, КОНЪЮГАТЫ С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2805251C2 |
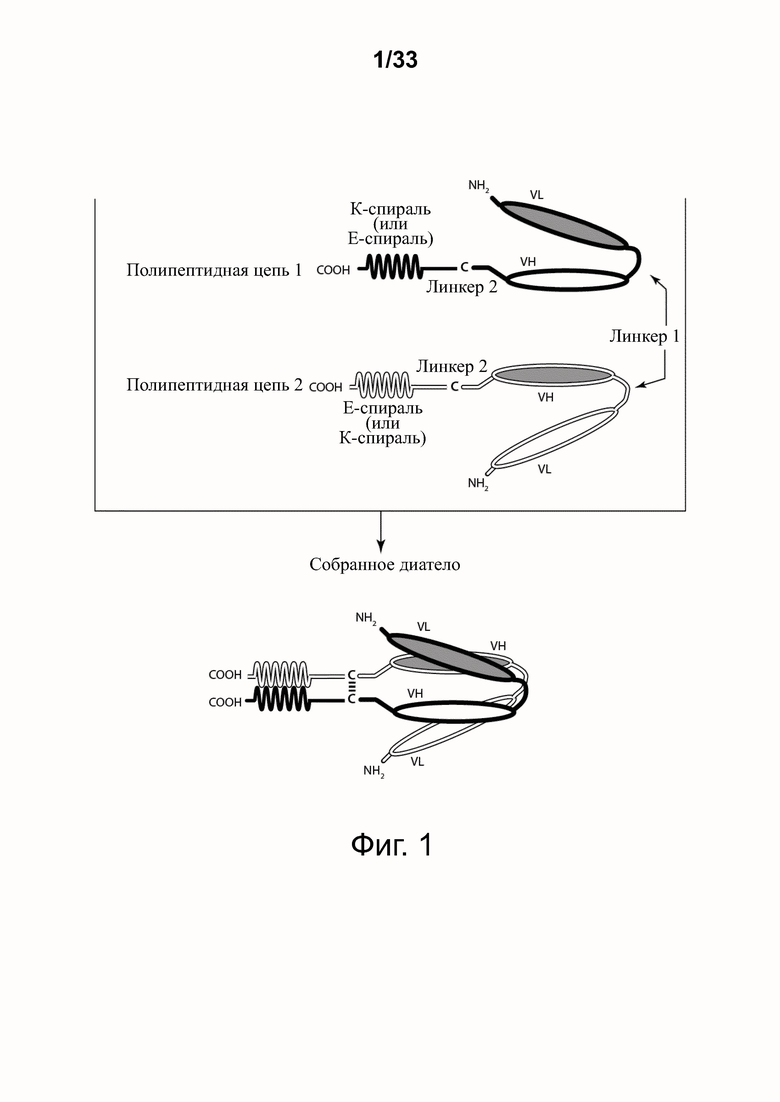
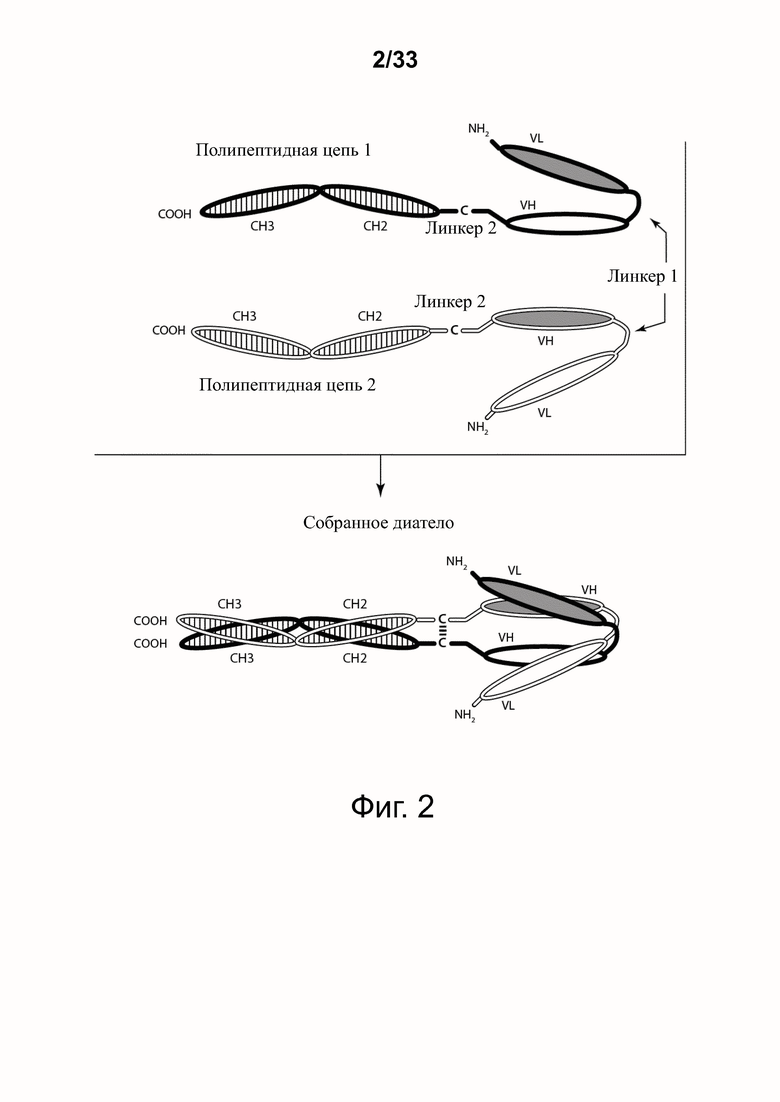
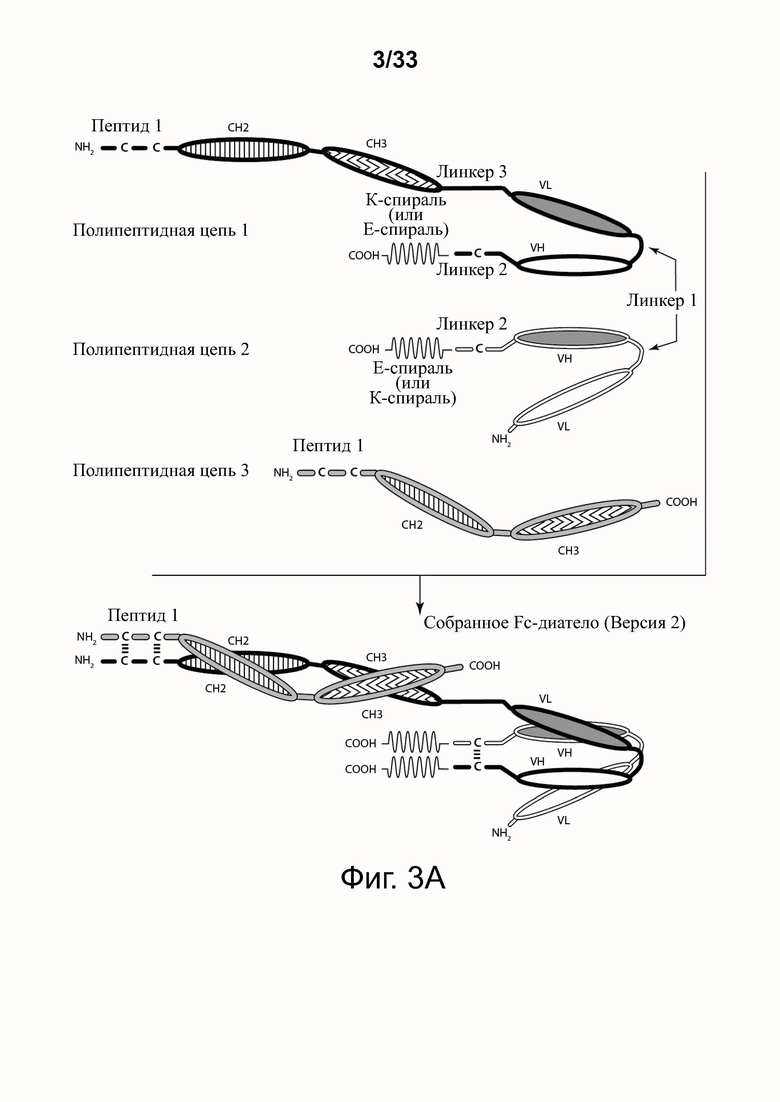
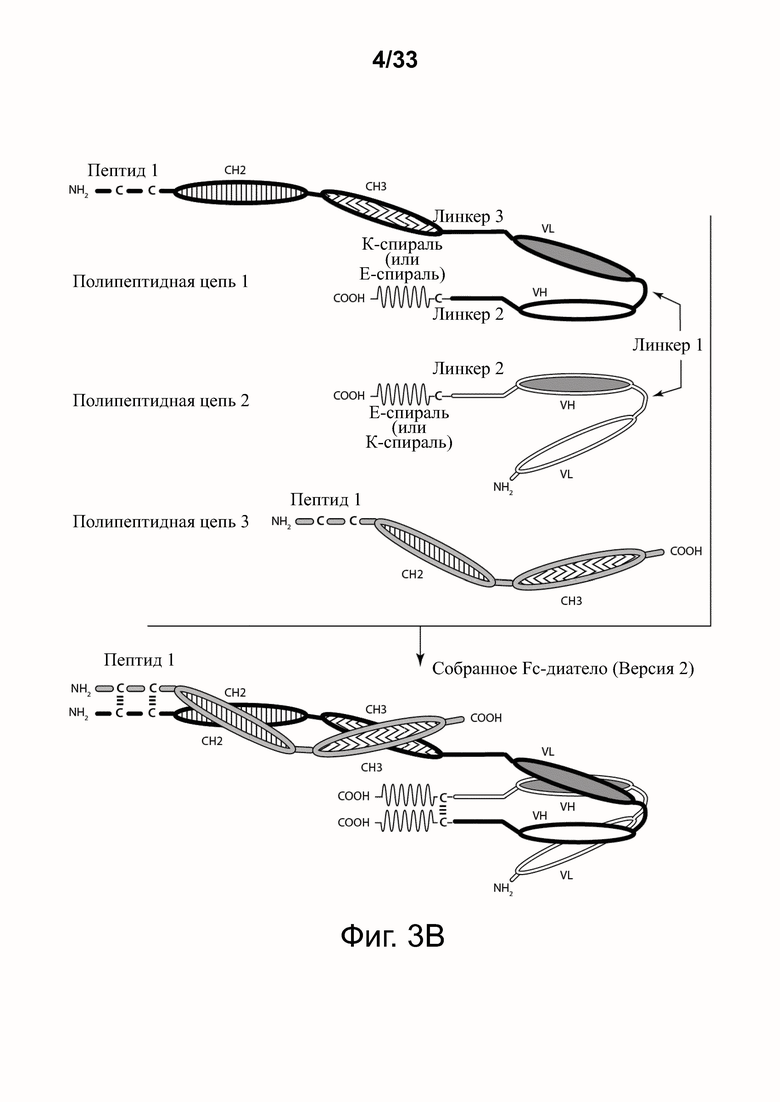

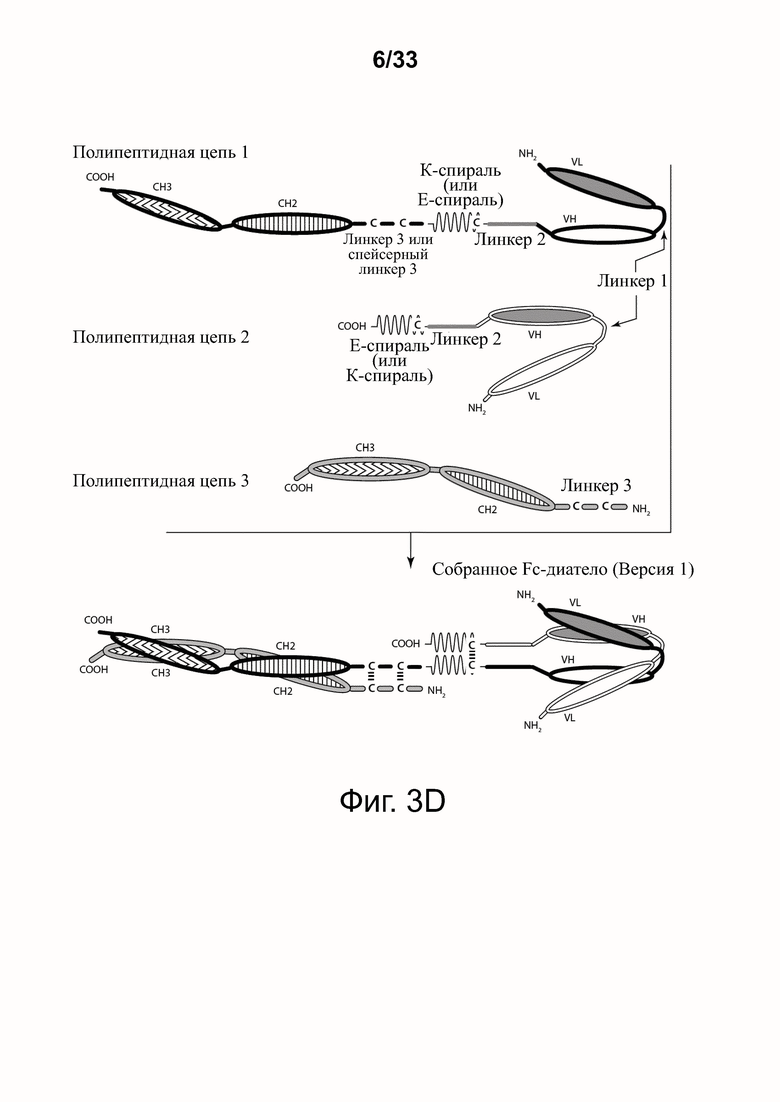
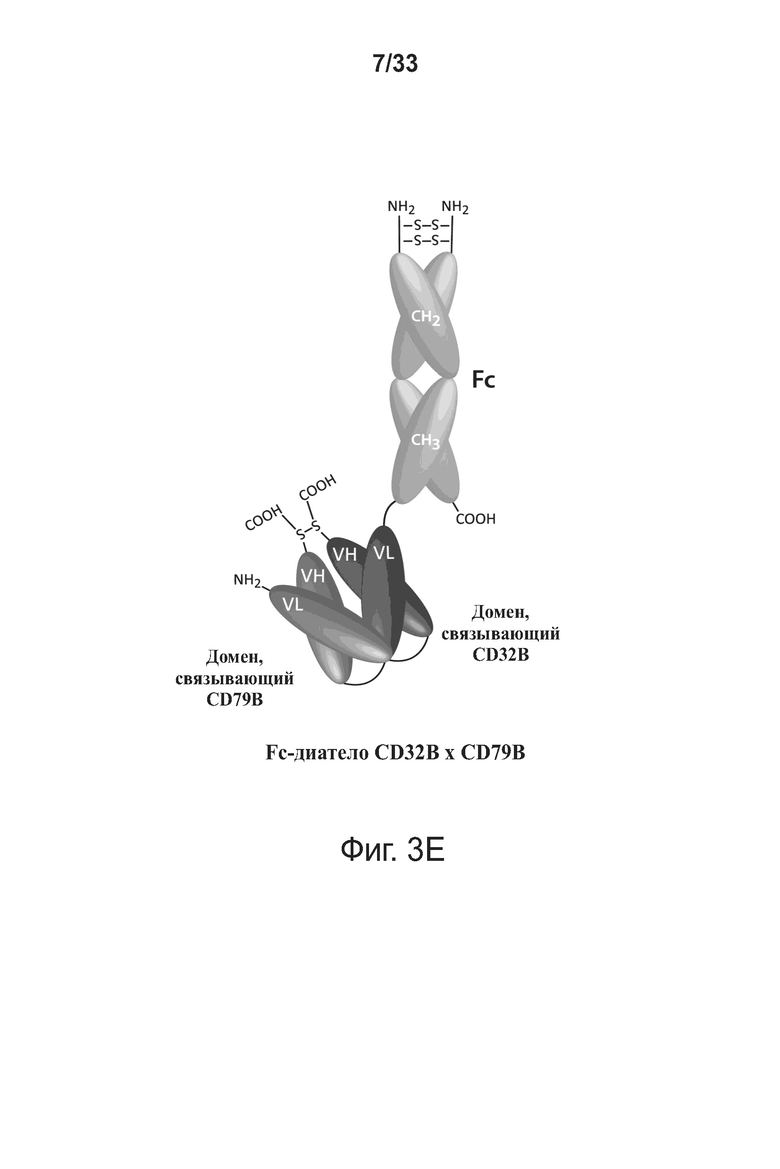
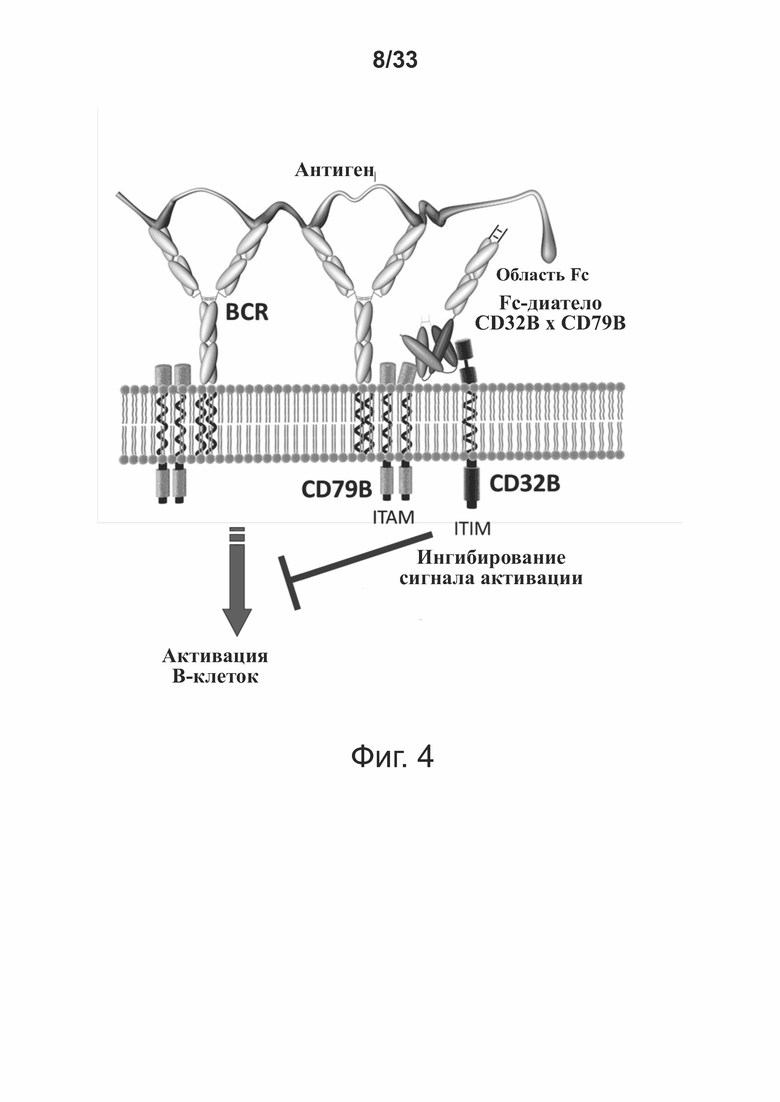
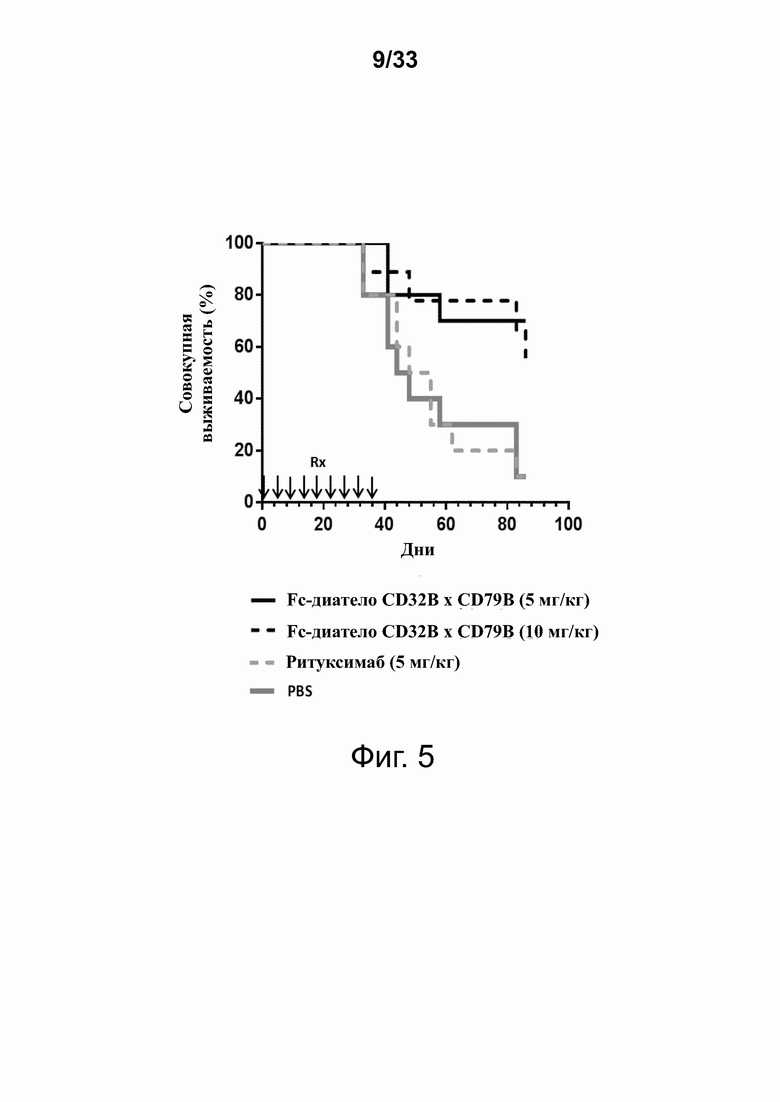
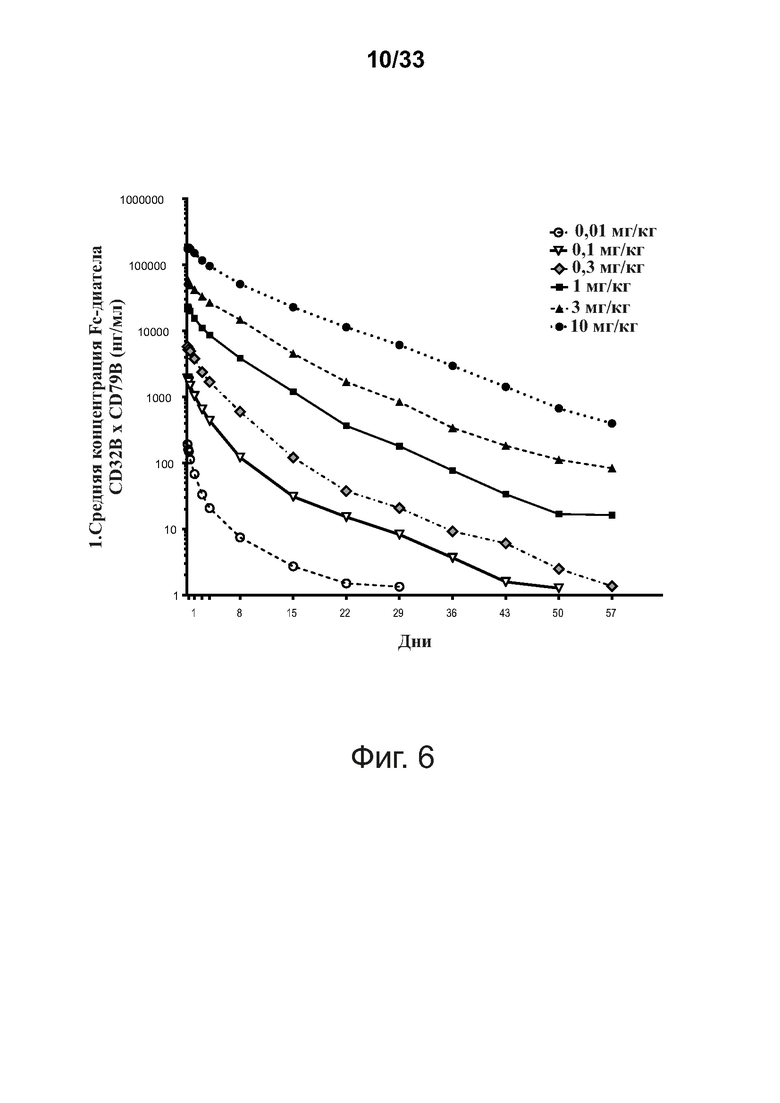
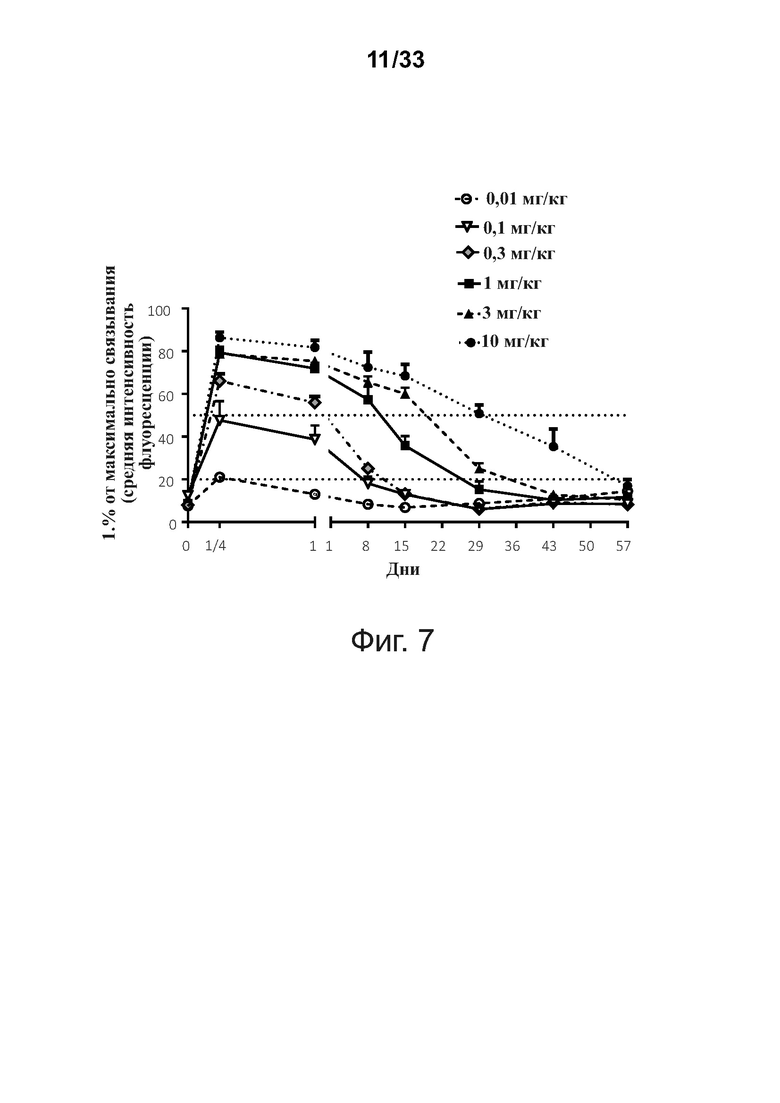
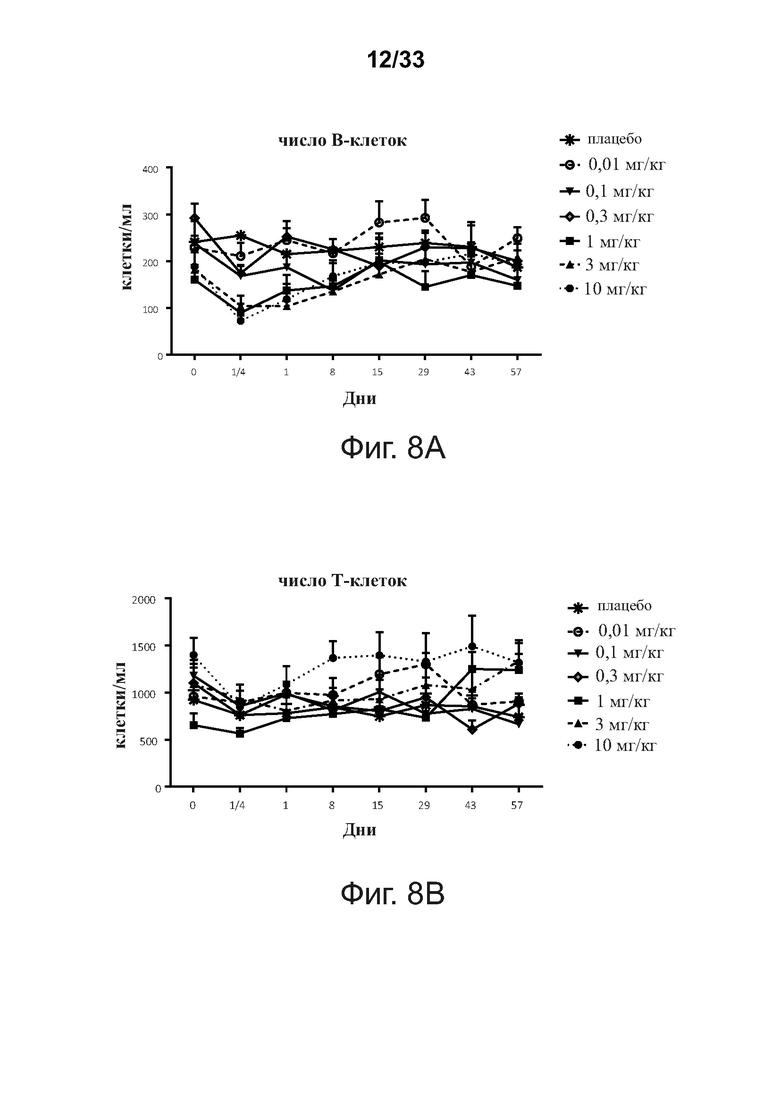
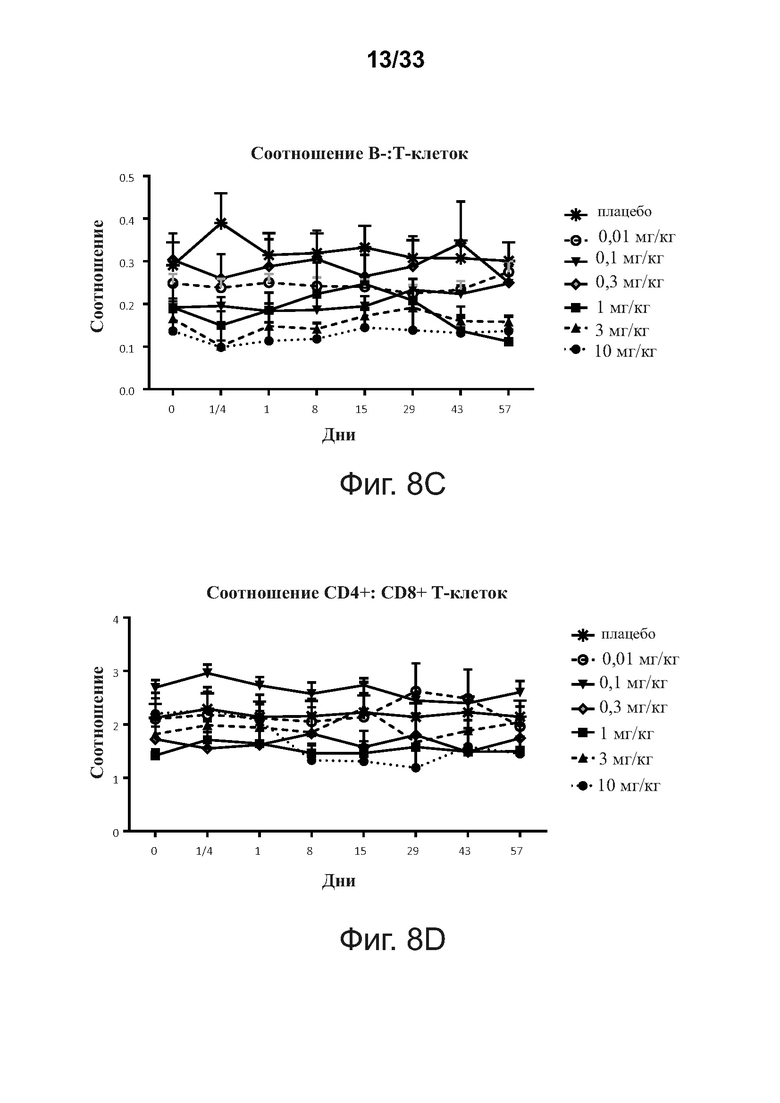
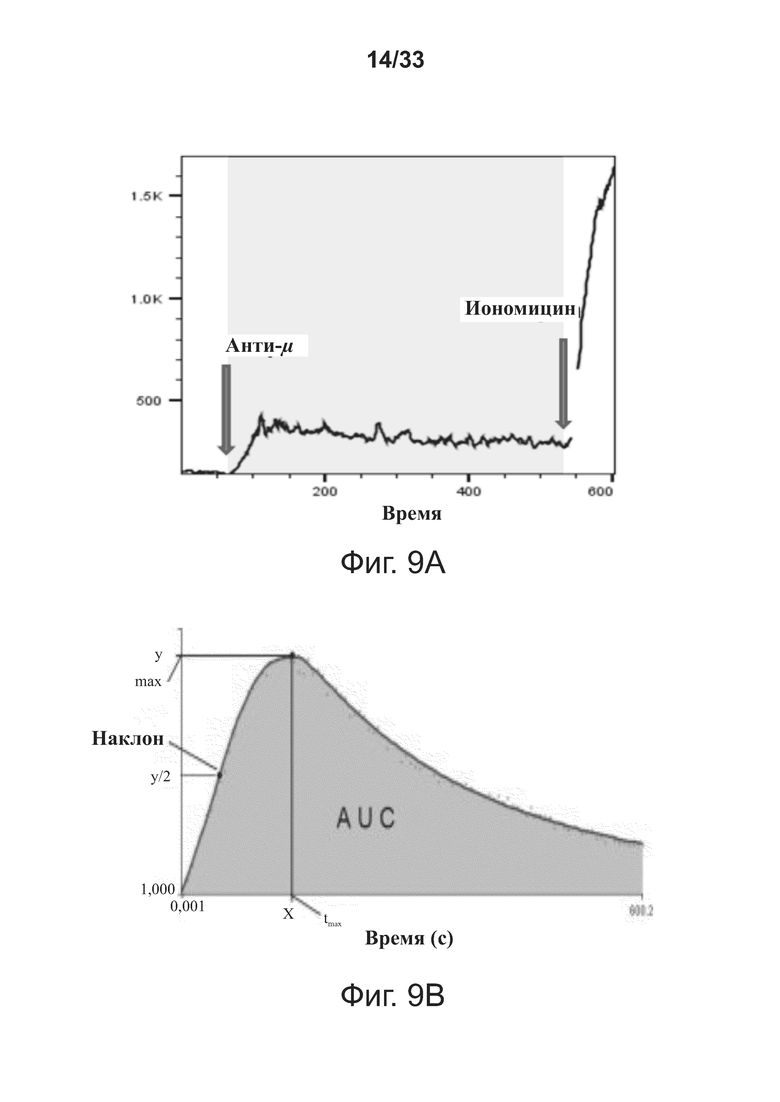
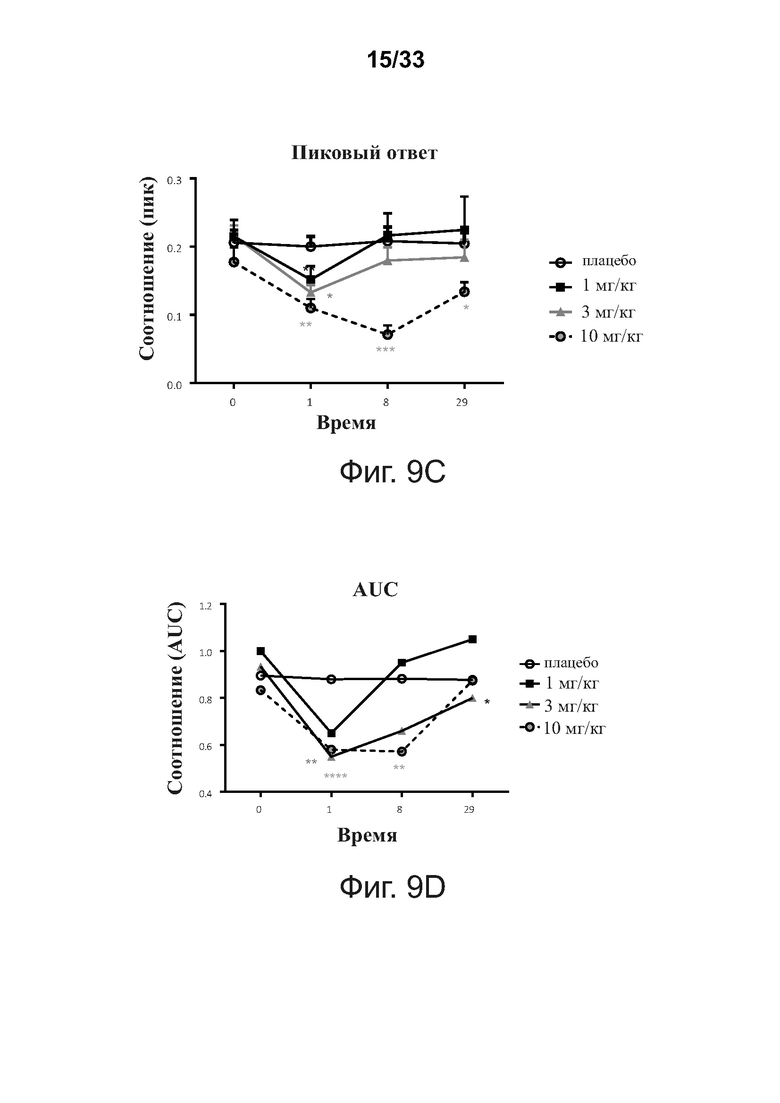
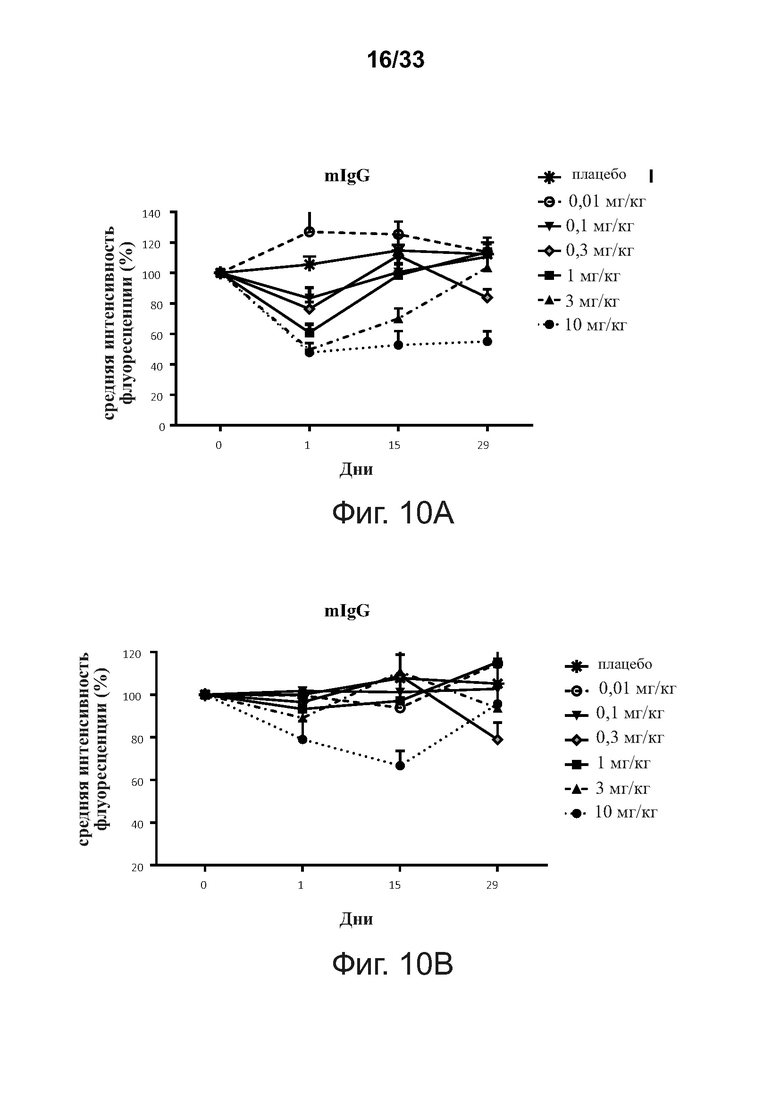
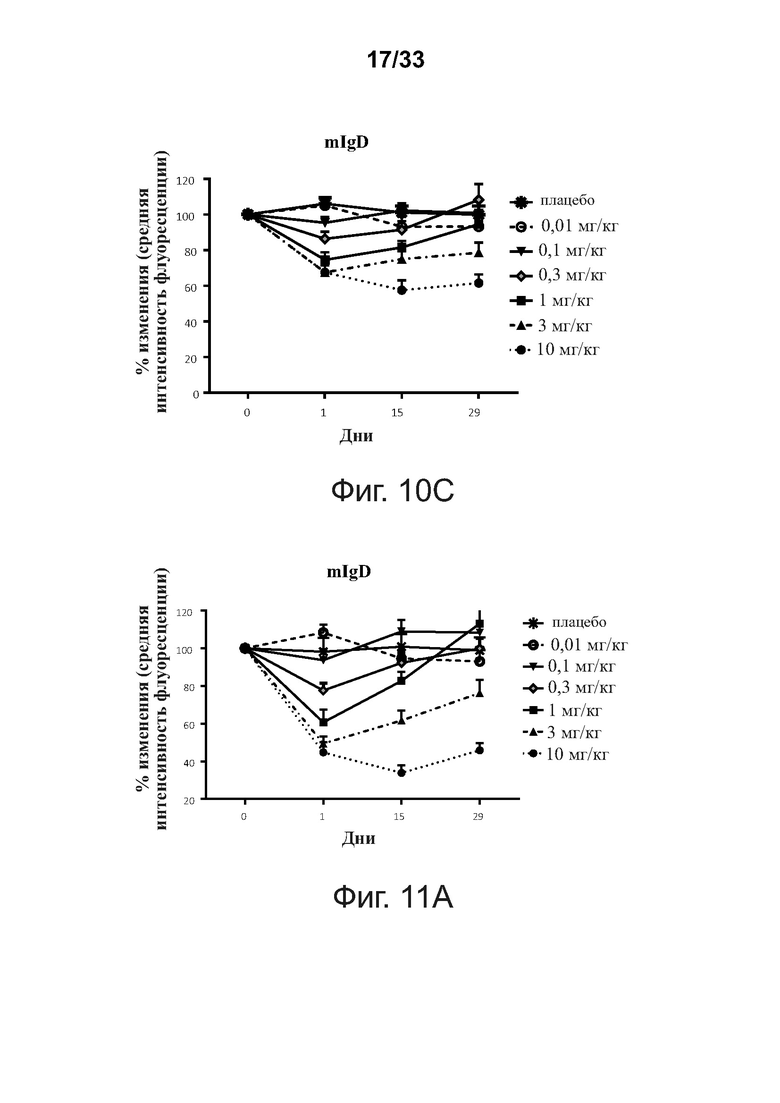

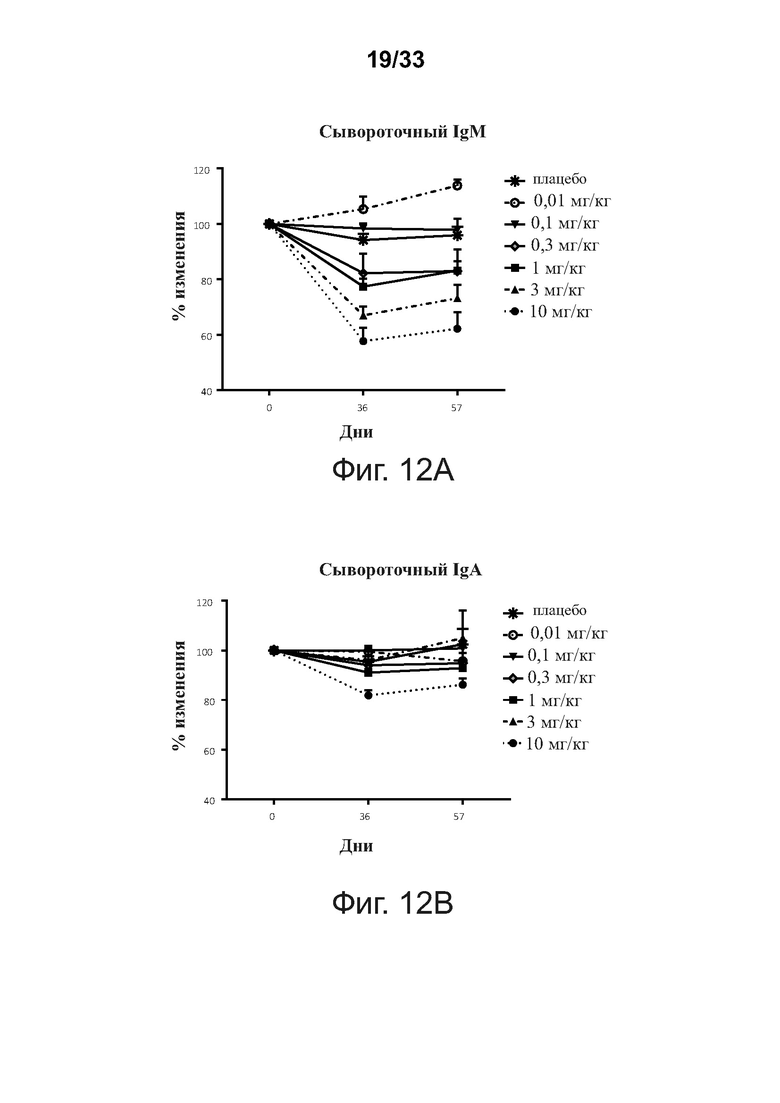
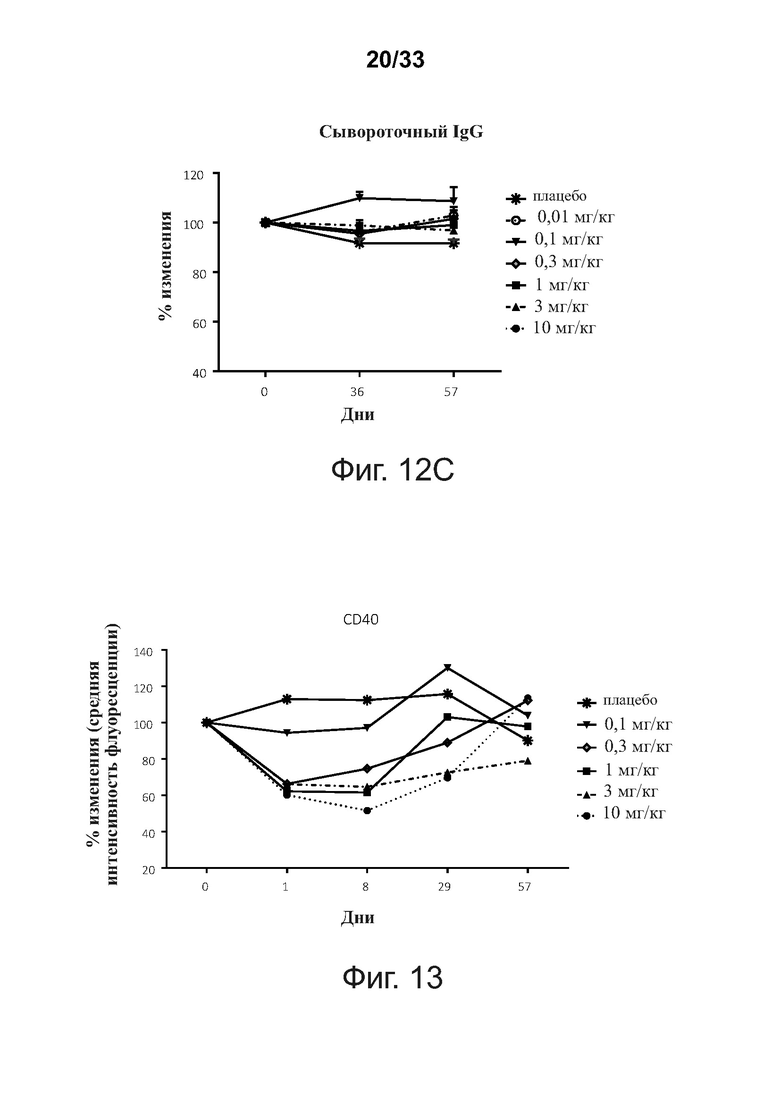
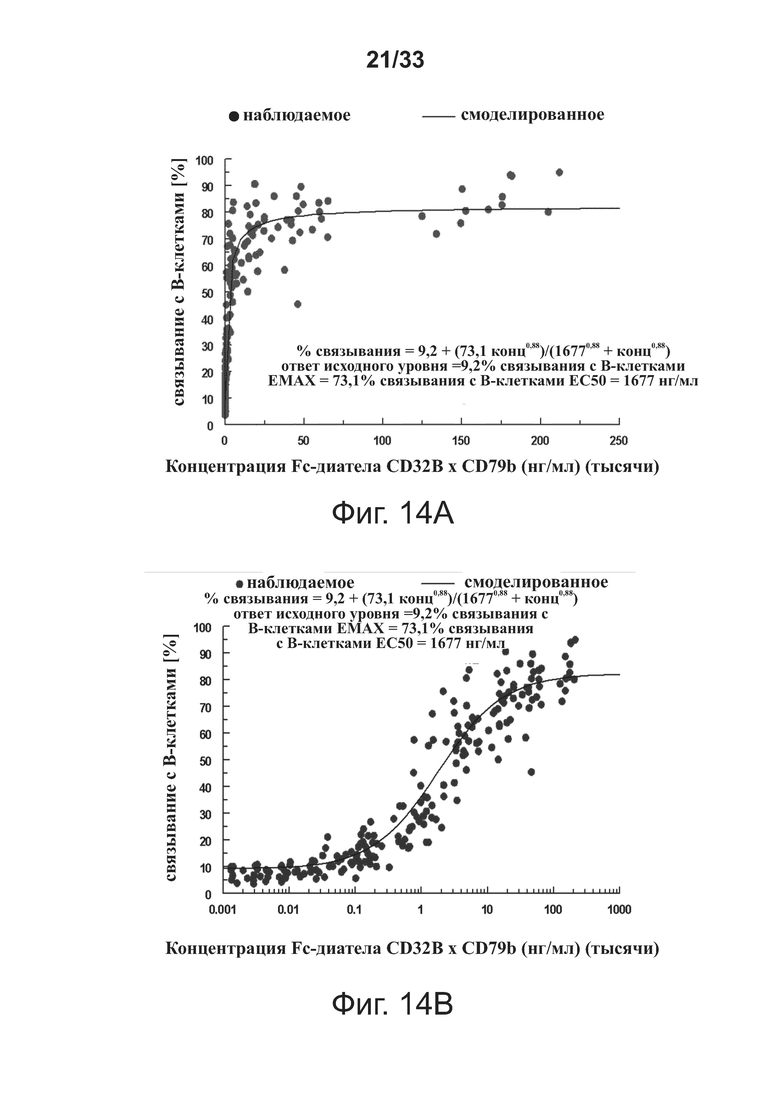
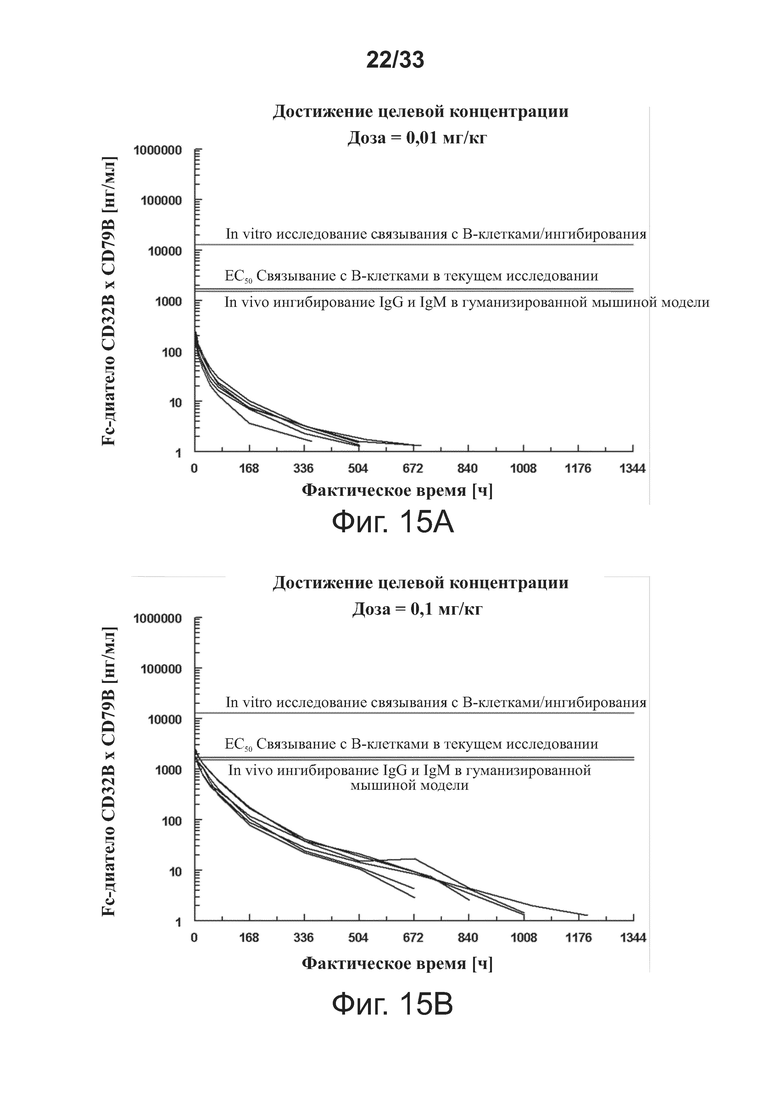
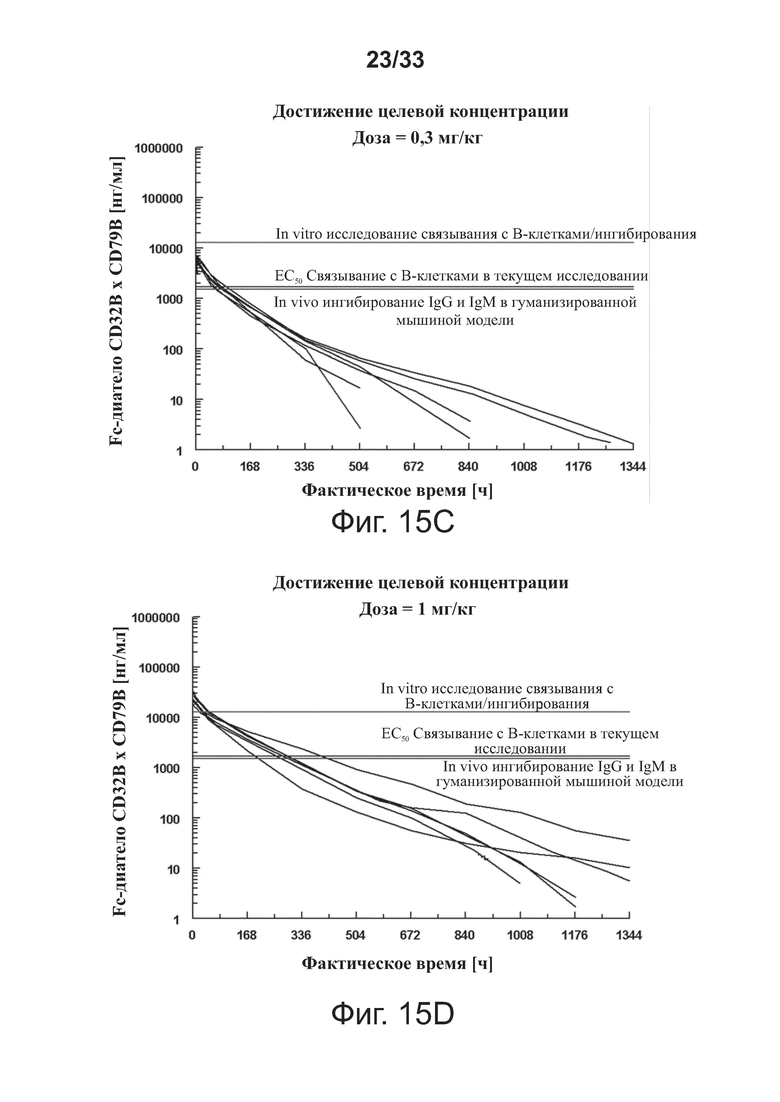
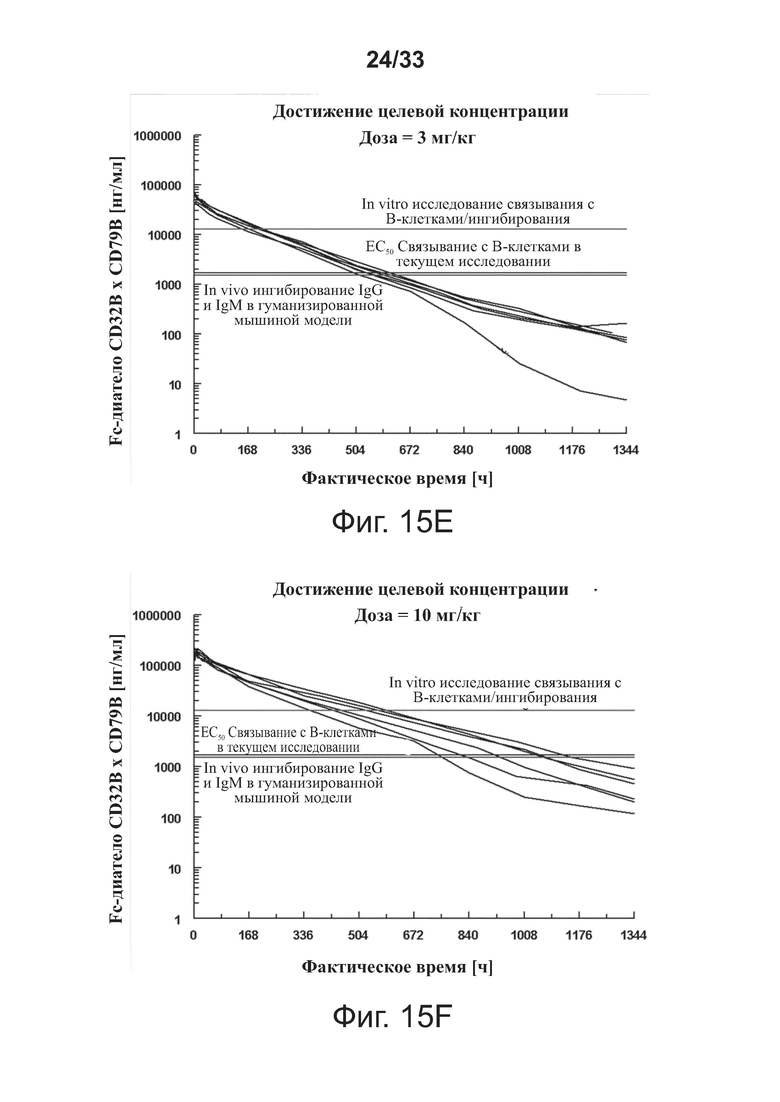
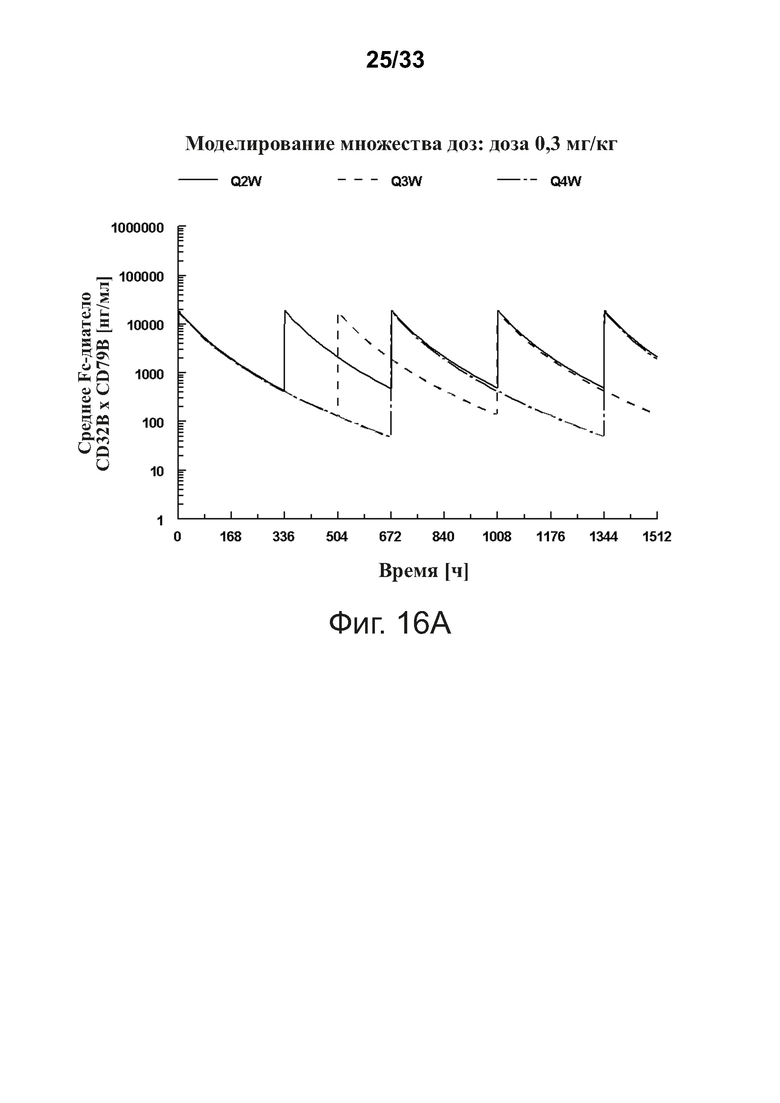
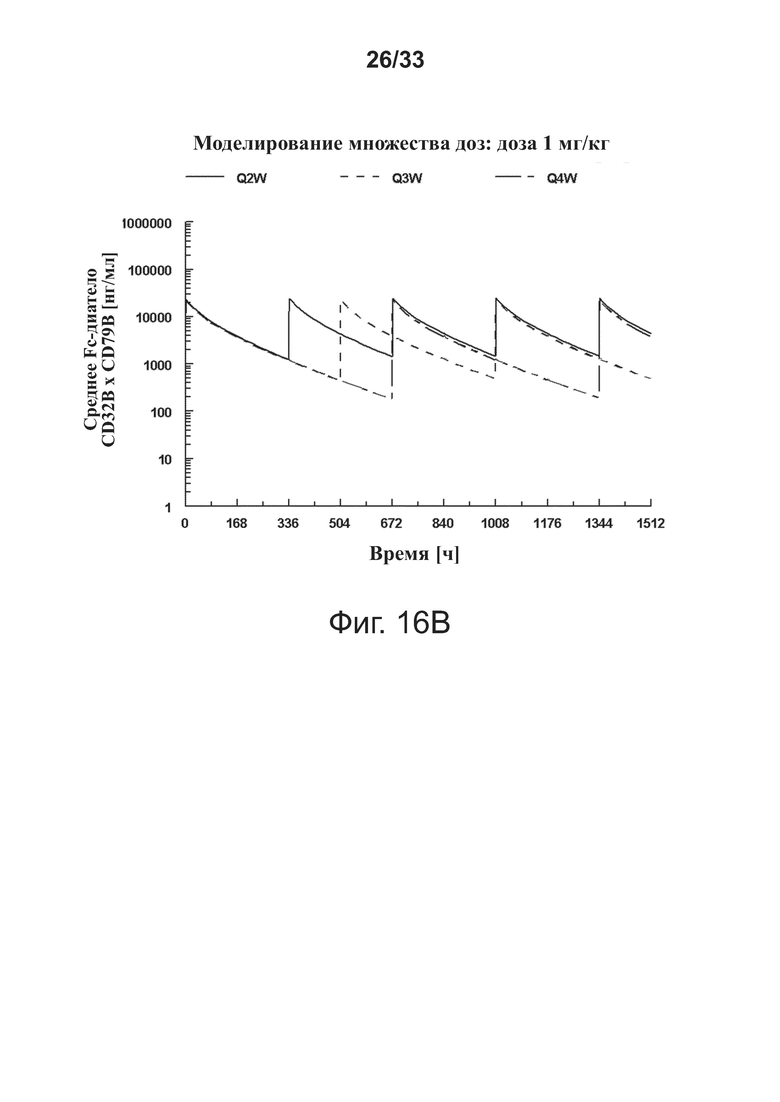
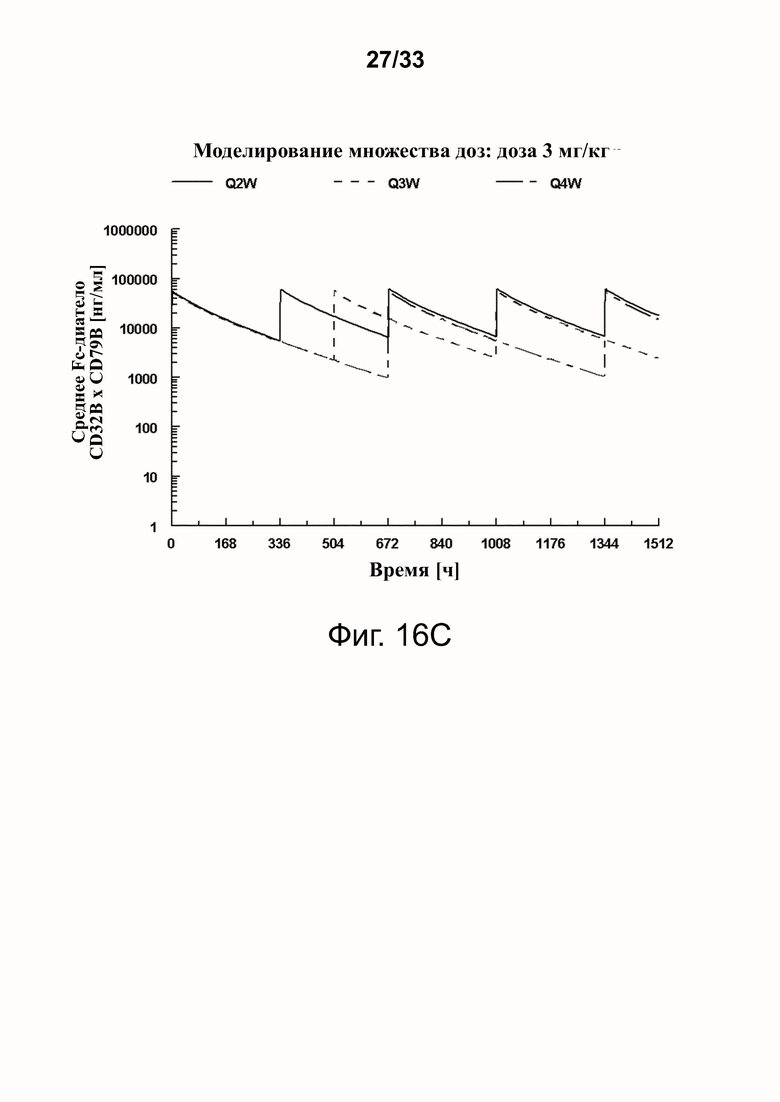
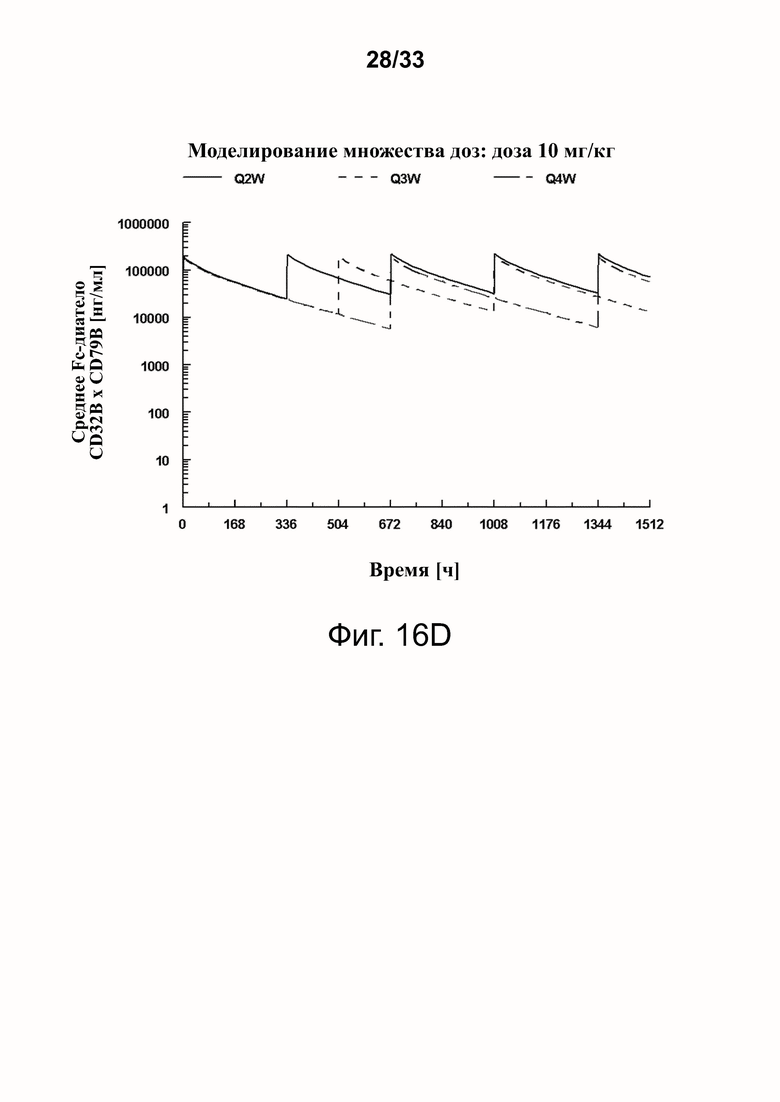
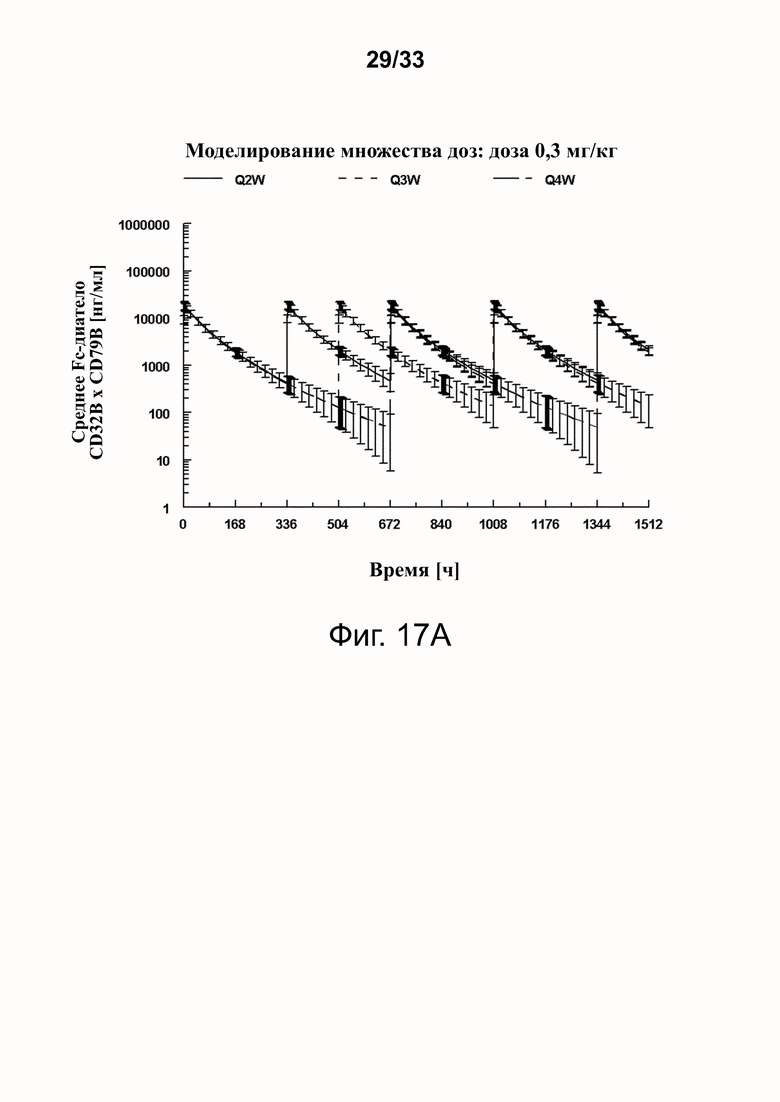
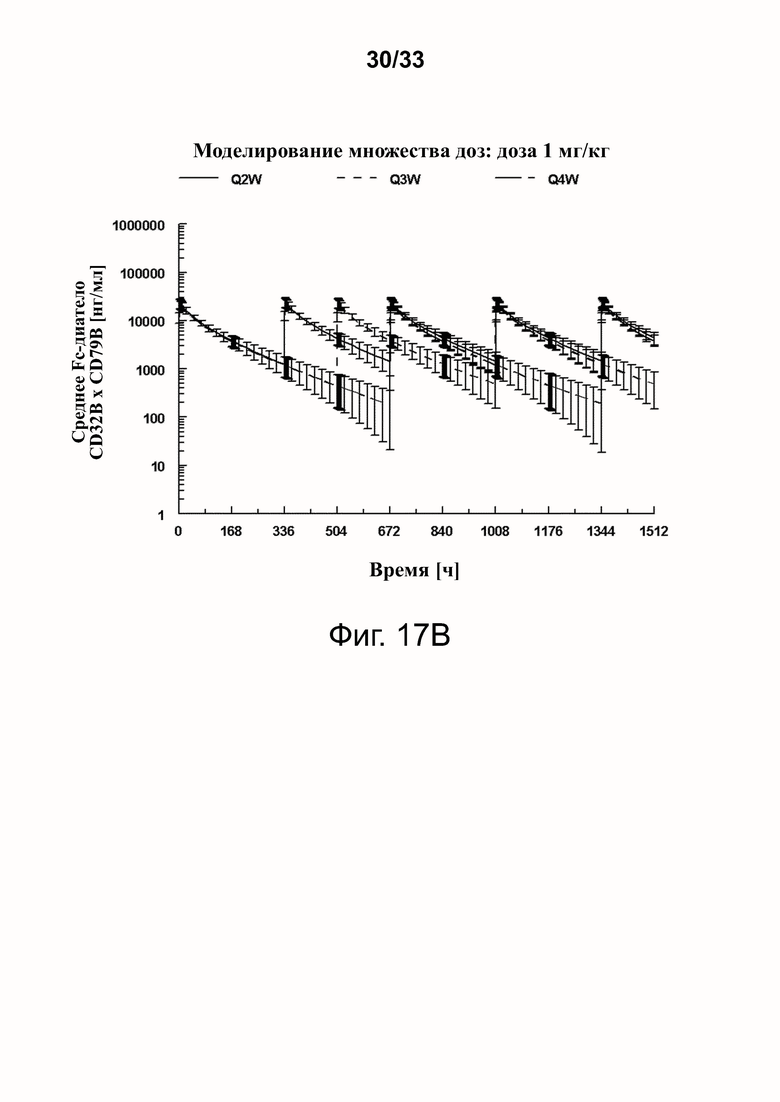
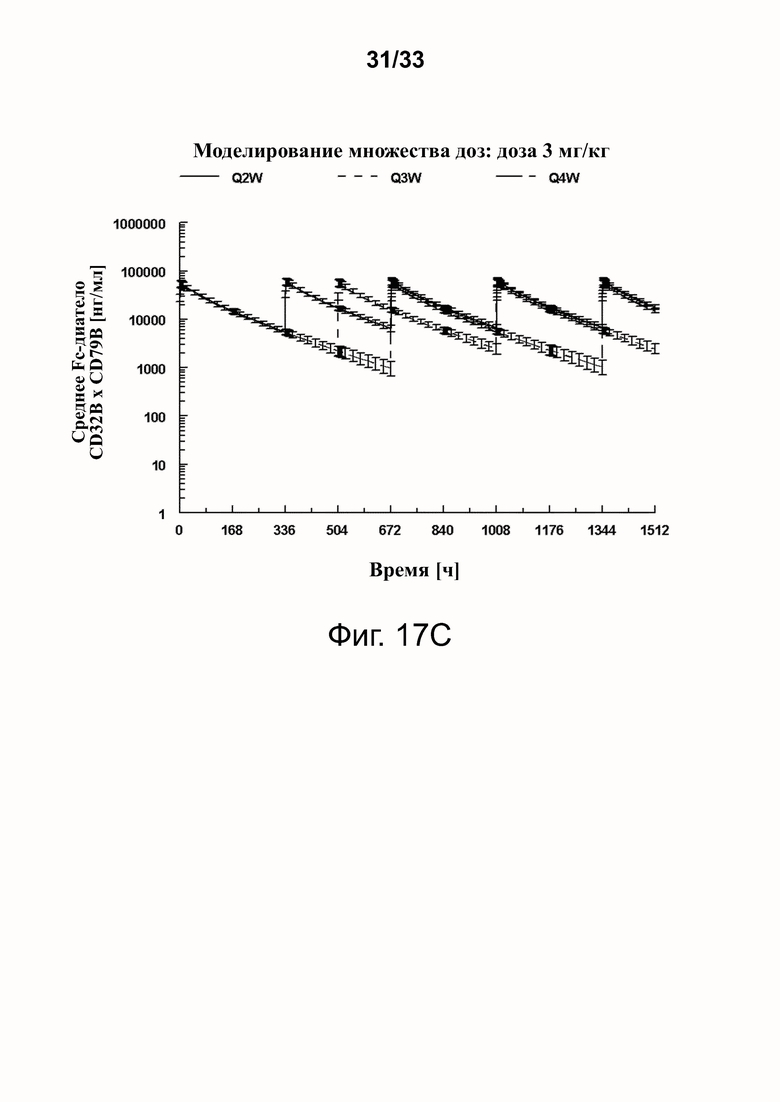
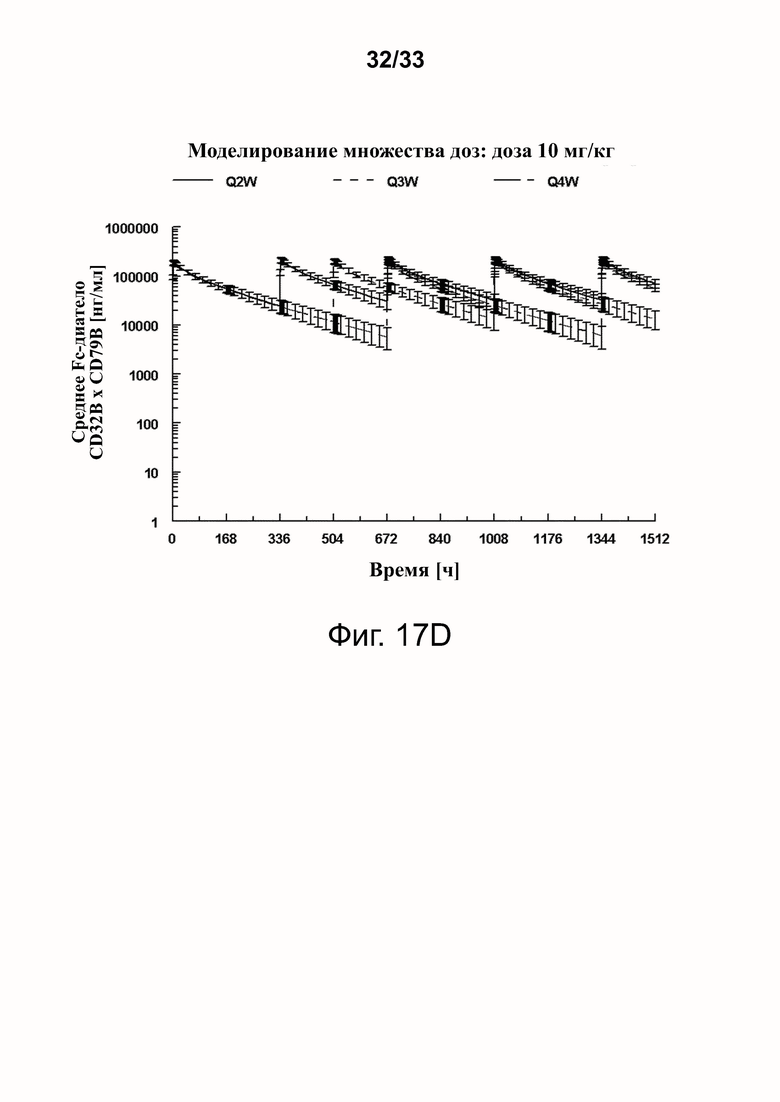
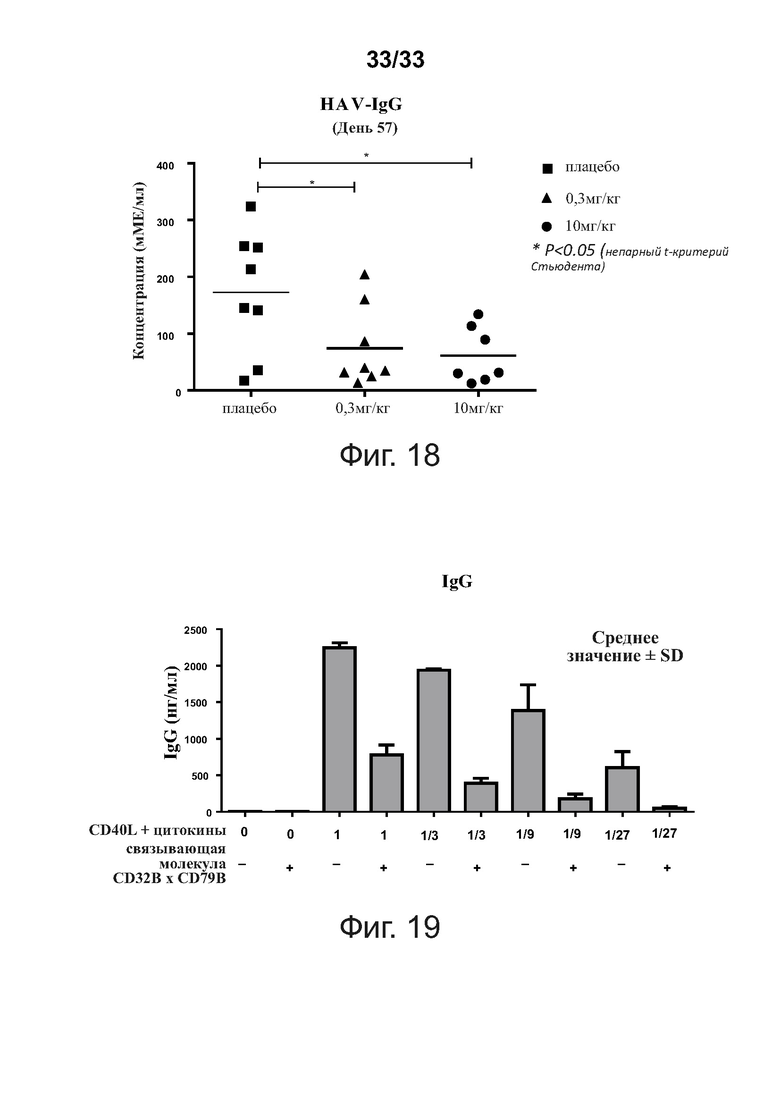
Группа изобретений относится к области медицины и фармацевтики, а именно к способам лечения воспалительного заболевания или состояния и уменьшения или ингибирования иммунного ответа, которые включают введение терапевтически эффективного количества связывающей молекулы CD32B х CD79B, которая способна иммуноспецифично связывать эпитоп CD32B и эпитоп CD79B и включает: (A) домен VLCD32B, который содержит аминокислотную последовательность SEQ ID NO: 30; (B) домен VHCD32B, который содержит аминокислотную последовательность SEQ ID NO: 31; (C) домен VLCD79B, который содержит аминокислотную последовательность SEQ ID NO: 32; (D) домен VHCD79B, который содержит аминокислотную последовательность SEQ ID NO: 33; и указанную связывающую молекулу CD32B x CD79B вводят в дозе от около 3 до около 10 мг/кг и в режиме дозирования одна доза каждые 3 недели (Q3W). Также группа изобретений относится к применению указанной связывающей молекулы CD32B х CD79B для лечения воспалительного заболевания или состояния и для снижения или ингибирования иммунного ответа. Группа изобретений обеспечивает линейную фармакокинетику средства, которое демонстрируют более 50%-ное связывание В-клеток в течение длительного времени. 4 н. и 10 з.п. ф-лы, 47 ил., 13 табл., 6 пр.
1. Способ лечения воспалительного заболевания или состояния, который включает введение терапевтически эффективного количества связывающей молекулы CD32B х CD79B субъекту, нуждающемуся в этом, где указанная связывающая молекула CD32B х CD79B способна иммуноспецифично связывать эпитоп CD32B и эпитоп CD79B, и где:
I. указанная связывающая молекула CD32B x CD79B включает:
(A) домен VLCD32B, который содержит аминокислотную последовательность SEQ ID NO: 30;
(B) домен VHCD32B, который содержит аминокислотную последовательность SEQ ID NO: 31;
(C) домен VLCD79B, который содержит аминокислотную последовательность SEQ ID NO: 32; а также
(D) домен VHCD79B, который содержит аминокислотную последовательность SEQ ID NO: 33; и
II. указанную связывающую молекулу CD32B x CD79B вводят в дозе от около 3 до около 10 мг/кг и в режиме дозирования одна доза каждые 3 недели (Q3W).
2. Способ уменьшения или ингибирования иммунного ответа, который включает введение терапевтически эффективного количества связывающей молекулы CD32B x CD79B субъекту, нуждающемуся в этом, где указанная связывающая молекула CD32B x CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B, и где:
I. указанная связывающая молекула CD32B x CD79B включает:
(A) домен VLCD32B, который содержит аминокислотную последовательность SEQ ID NO: 30;
(B) домен VHCD32B, который содержит аминокислотную последовательность SEQ ID NO: 31;
(C) домен VLCD79B, который содержит аминокислотную последовательность SEQ ID NO: 32; а также
(D) домен VHCD79B, который содержит аминокислотную последовательность SEQ ID NO: 33; и
II. указанную связывающую молекулу CD32B x CD79B вводят в дозе от около 3 до около 10 мг/кг и в режиме дозирования одна доза каждые 3 недели (Q3W).
3. Применение связывающей молекулы CD32B x CD79B для лечения воспалительного заболевания или состояния у субъекта, нуждающегося в этом, где указанная связывающая молекула CD32B x CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B, и при этом:
I. указанная связывающая молекула CD32B x CD79B включает:
(A) домен VLCD32B, который содержит аминокислотную последовательность SEQ ID NO: 30;
(B) домен VHCD32B, который содержит аминокислотную последовательность SEQ ID NO: 31;
(C) домен VLCD79B, который содержит аминокислотную последовательность SEQ ID NO: 32; а также
(D) домен VHCD79B, который содержит аминокислотную последовательность SEQ ID NO: 33; и
II. указанная связывающая молекула CD32B x CD79B вводится в дозе от примерно 3 мг/кг до примерно 10 мг/кг и в режиме дозирования одна доза каждые 3 недели (Q3W).
4. Применение связывающей молекулы CD32B x CD79B для снижения или ингибирования иммунного ответа у субъекта, нуждающегося в этом, при этом указанная связывающая молекула CD32B x CD79B способна иммуноспецифически связывать эпитоп CD32B и эпитоп CD79B, и где:
I. указанная связывающая молекула CD32B x CD79B включает:
(A) домен VLCD32B, который содержит аминокислотную последовательность SEQ ID NO: 30;
(B) домен VHCD32B, который содержит аминокислотную последовательность SEQ ID NO: 31;
(C) домен VLCD79B, который содержит аминокислотную последовательность SEQ ID NO: 32; а также
(D) домен VHCD79B, который содержит аминокислотную последовательность SEQ ID NO: 33; и
II. указанная связывающая молекула CD32B x CD79B вводится в дозе от примерно 3 мг/кг до примерно 10 мг/кг, и в режиме дозирования одна доза каждые 3 недели (Q3W).
5. Способ по любому из пп. 1 или 2 или применение по любому из пп. 3 и 4, где указанную связывающую молекулу CD32B х CD79B вводят в дозе около 3 мг/кг.
6. Способ по любому из пп. 1 или 2 или применение по любому из пп. 3 и 4, где указанную связывающую молекулу CD32B х CD79B вводят в дозе около 10 мг/кг.
7. Способ по любому из пп. 1, 2 и 5-6 или применение по любому из пп. 3-6, где указанная связывающая молекула CD32B x CD79B представляет собой биспецифическое антитело, которое связывает эпитоп CD32B и эпитоп CD79B, или молекулу, которая включает CD32B- и CD79B-связывающие домены указанного биспецифического антитела.
8. Способ по любому из пп. 1, 2 и 5-7 или применение по любому из пп. 3-7, где указанная связывающая молекула CD32B x CD79B представляет собой биспецифическое диатело CD32B x CD79B.
9. Способ по п. 8 или применение по п. 8, где указанное биспецифическое диатело CD32B x CD79B представляет собой Fc-диатело CD32B x CD79B.
10. Способ по любому из пп. 1, 2 или 5-9 или применение по любому из пп. 3-9, где указанное воспалительное заболевание или состояние представляет собой аутоиммунное заболевание.
11. Способ по п. 10 или применение по п. 10, где указанное аутоиммунное заболевание выбрано из группы, которая состоит из болезни Аддисона, аутоиммунного гепатита, аутоиммунного заболевания внутреннего уха, миастении, болезни Крона, дерматомиозита, семейного аденоматозного полипоза, болезни трансплантата против хозяина (GvHD), болезни Грейвса, тиреоидита Хашимото, красной волчанки, рассеянного склероза (MS); пернициозной анемии, синдрома Рейтера, ревматоидного артрита (RA), синдрома Шегрена, системной красной волчанки (SLE), диабета 1 типа, первичного васкулита (например, ревматической полимиалгии, гигантоклеточного артериита, болезнь Бехчета), пемфигуса, оптикомиелита, анти-NMDA-рецепторного энцефалита, синдрома Гийена-Барре, хронической воспалительной демиелинизирующей полиневропатии (CIDP), офтальмопатии Грейвса, связанных с IgG4 заболеваний, идиопатической тромбоцитопенической пурпуры (ITP) и язвенного колита.
12. Способ по п. 11 или применение по п. 11, где указанное аутоиммунное заболевание представляет собой GvHD, RA, MS или SLE.
13. Способ по любому из пп. 1, 2 или 5-12 или применение по любому из пп. 3-12, где указанный субъект представляет собой человека.
14. Способ по любому из пп.1, 2 и 5-13 или применение по любому из пп. 3-13, где указанная связывающая молекула CD32B x CD79B представляет собой Fc-диатело, включающее:
(А) первую полипептидную цепь, которая включает аминокислотную последовательность SEQ ID NO: 39;
(B) вторую полипептидную цепь, которая включает аминокислотную последовательность SEQ ID NO: 41; и
(С) третью полипептидную цепь, которая включает аминокислотную последовательность SEQ ID NO: 44.
| WO2015021089A1, 12.02.2015 | |||
| ЯКУБКЕ Х.-Д | |||
| и др | |||
| Аминокислоты, пептиды, белки: Пер | |||
| с нем | |||
| - М: Мир, 1985 | |||
| Подвижной рельс для пересечений железнодорожных путей | 1922 |
|
SU456A1 |
| LLOYD C | |||
| et al | |||
| УСТРОЙСТВО ДЛЯ ИСПОЛЬЗОВАНИЯ ЭНЕРГИИ МОРСКИХ ВОЛН | 1923 |
|
SU1011A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Vol | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |

