ОБЛАСТЬ ТЕХНИКИ
[0001] Настоящее изобретение относится к области иммунотерапии аутоиммунных заболеваний, и относится к лекарственному средству на основе гуманизированного моноклонального антитела против IL17A и его применению.
УРОВЕНЬ ТЕХНИКИ
[0002] IL17A, также обычно называемый IL-17, представляет собой воспалительный цитокин, состоящий из 155 аминокислот с молекулярным весом 35 кДа. Он в основном секретируется Т-хелперами 17, в дополнение к Т-клеткам 17, клетки CD4+, CD8+ Т и γδ-Т также могут секретировать IL-17. Семейство IL-17 включает шесть членов, IL17A, IL-17B, IL-17C, IL-17D (IL-27), IL-17E (IL-25) и IL-17F (Gu, Wu и др. 2013), среди которых IL17A и IL-17F являются наиболее важными членами. Имея 55% гомологию по аминокислотам, IL17A и IL-17F могут образовывать гомодимеры или гетеродимеры, связанные дисульфидными связями (Dubin и Kolls 2009). При связывании с различными экспрессирующими IL-17RA клетками семейства рецепторов IL-17, такими как: макрофаги, дендритные клетки, гемопоэтические клетки, остеобласты, фибробласты и т.д., димер IL17A/A или IL-17A/F может активировать NFκB, C/EBPs, MAPK и другие сигнальные пути в рецепторных клетках, побуждающие эти клетки секретировать факторы воспаления и хемокины (IL-6, IL-8, CXCL1 и т.д.), рекрутируют нейтрофилы и опосредуют развитие воспалительных реакций (Mitra, Raychaudhuri и др. 2014). Возникновение и развитие многих связанных с воспалением аутоиммунных заболеваний, таких как псориаз, псориатический артрит, ревматоидный артрит и анкилозирующий спондилоартрит, тесно связаны с путем IL-17 (Wang, Suzuki и др. 2017), со значительным повышением уровня экспрессии IL-17 в сыворотке пациентов (Marinoni, Ceribelli и др. 2014), что приводит к стойкой воспалительной реакции. Дополнительно IL-17 также может непосредственно воздействовать на эндотелиальные клетки, эпителиальные клетки, фибробласты и кератиноциты кожи, усиливая высвобождение множественных воспалительных факторов и вызывая патологические изменения кожи (Mitra, Raychaudhuri и др. 2014, Brembilla, Senra и др. 2018). Таким образом, блокирование путей ИЛ-17 обеспечивает возможность ингибирования процесса аутоиммунных заболеваний.
[0003] В настоящее время продаваемыми препаратами на основе антител, нацеленных на IL-17, являются COSENTYX®/Секукинумаб (US 7807155 B2) от Novartis и Taltz®/Иксекизумаб (US 7838638 B2/CN 101326195 B) от Eli Lilly, основной механизм их действия следующий: посредством их связывания с IL17A и IL17A/F ингибируется связывание IL17A с рецептором (IL-17RA/C), последовательно блокируется высвобождение воспалительных факторов и хемокинов, тем самым эффективно облегчаются аутоиммунные заболевания (Fala 2016, Liu, Lu и др. 2016. Секукинумаб одобрен для лечения бляшечного псориаза, псориатического артрита и анкилозирующего спондилита. Иксекизумаб одобрен для лечения бляшечного псориаза и псориатического артрита. Гуманизированное моноклональное антитело против IL17A согласно настоящему изобретению способно специфически связывать белок IL17A с высокой аффинностью, обладает сильной способностью блокировать связывание IL17A, IL17A/F с рецептором IL17RA и обладает превосходной или эквивалентной способностью блокировать EL17A, IL17A/F, предотвращая индукцию секреции воспалительных цитокинов эпидермальных фибробластов человека HFF, по сравнению с сопоставимыми препаратами; в модели псориаза на мышах указанное антитело также демонстрирует значительно лучшую фармакодинамическую активность in vivo, чем сопоставимые препараты, со значительно более низкими показателями PASI после введения дозы; указанное гуманизированное антитело демонстрирует благоприятные фармакокинетические профили у яванских макаков, включая быстрое всасывание после подкожной инъекции, длительный период полувыведения и лучшую экспозицию лекарственного средства, закладывая основу для более длительных циклов дозирования. Антитело согласно настоящему изобретению планируется использовать для, в том числе, лечения псориаза, но не ограничивается им.
Краткое описание
[0004] Настоящее изобретение удовлетворяет вышеуказанную потребность путем разработки связывающего IL17A антитела с новой структурой. Настоящее изобретение раскрывает гуманизированное моноклональное антитело, которое специфически связывает IL17A с высокой аффинностью и обладает сильной способностью блокировать связывание IL17A, IL17A/F с рецептором IL17RA. По сравнению с другими сопоставимыми лекарственными средствами антитело согласно настоящему изобретению обладает превосходной или эквивалентной способностью блокировать IL17A и IL17A/F, предотвращая индукцию секреции воспалительных цитокинов эпидермальными фибробластами HFF человека; в модели псориаза на мышах указанное антитело также продемонстрировало существеннное преимущество фармакологической активности in vivo по сравнению с сопоставимыми препаратами и значительно снизило показатели PASI после введения дозы; в фармакокинетическом исследовании на яванских макаках, указанное гуманизированное антитело продемонстрировало превосходные фармакокинетические характеристики, в том числе быстрое всасывание после подкожной инъекции, длительный период полувыведения, лучшую экспозицию лекарственного средства и т.д., заложив основу для более длительных циклов дозирования. Гуманизированное моноклональное антитело согласно настоящему изобретению может применяться для лечения псориаза.
[0005] В одном аспекте настоящее изобретение обеспечивает изолированное антитело против IL17A или его антиген-связывающий фрагмент, содержащий вариабельный участок тяжелой цепи, включающий участок CDR1 тяжелой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 13, участок CDR2 тяжелой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 14, и участок CDR3 тяжелой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 15; и вариабельный участок легкой цепи, включающий участок CDR1 легкой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 10, участок CDR2 легкой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 11, и участок CDR3 легкой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 12. (6 CDR, общие для антитела М069 мыши и гуманизированного антитела Н069, идентичны у мыши и человека.)
[0006] В одном варианте реализации указанное антитело против IL17A или его антиген-связывающий фрагмент содержат вариабельный участок тяжелой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 22, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности SEQ ID NO: 22; и вариабельный участок легкой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 23, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности SEQ ID NO: 23. (Последовательность аминокислот вариабельных участков тяжелой цепи и легкой цепи гуманизированного антитела Н069)
[0007] В одном варианте реализации указанное антитело против IL17A или его антиген-связывающий фрагмент представляет собой гуманизированное антитело или химерное антитело.
[0008] В одном варианте реализации указанное антитело против IL17A дополнительно включает константный участок тяжелой цепи и константный участок легкой цепи, предпочтительно указанный константный участок тяжелой цепи представляет собой константный участок тяжелой цепи IgG1, имеющий последовательность аминокислот, указанную в SEQ ID NO: 24, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности SEQ ID NO: 24; и/или указанный константный участок легкой цепи представляет собой константный участок легкой цепи каппа человека, имеющий последовательность аминокислот, указанную в SEQ ID NO: 25, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности SEQ ID NO: 25. (Последовательность аминокислот тяжелой цепи и константный участок легкой цепи гуманизированного антитела Н069).
[0009] В одном варианте реализации указанное антитело против IL17A дополнительно включает сигнальный пептид, присоединенный к вариабельному участку тяжелой цепи и/или сигнальный пептид, присоединенный к вариабельному участку легкой цепи, предпочтительно указанный сигнальный пептид, присоединенный к вариабельному участку тяжелой цепи, представляет собой последовательность аминокислот, указанную в SEQ ID NO: 20 или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности SEQ ID NO: 20; и/или указанный сигнальный пептид, присоединенный к вариабельному участку легкой цепи, представляет собой последовательность аминокислот, указанную в SEQ ID NO: 21 или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности SEQ ID NO: 21. (Последовательности аминокислот сигнальных пептидов тяжелой цепи и легкой цепи гуманизированного антитела Н069).
[0010] В одном варианте реализации указанное антитело против IL17A или его антиген-связывающий фрагмент представляет собой антитело IgG, предпочтительно антитело IgG1.
[0011] В одном варианте реализации указанное антитело против IL17A или его антиген-связывающий фрагмент представляет собой моноклональное антитело.
[0012] В одном варианте реализации аффинность связывания KD указанного антитела против IL17A или его антиген-связывающего фрагмента с рекомбинантным белком IL17A человека составляет 0,1-10E-11 М, предпочтительно 0,5-5Е-11 М, и более предпочтительно 2,88Е-11 М.
[0013] В одном варианте реализации аффинность связывания KD указанного антитела против IL17A или его антиген-связывающего фрагмента с рекомбинантным белком IL17A/F человека составляет 0,1-10Е-10 М, предпочтительно 0,5-5Е-10 М, и более предпочтительно 5,37Е-10 М.
[0014] В одном варианте реализации указанный антиген-связывающий фрагмент представляет собой Fv, Fab, Fab', Fab'-SH, F(ab')2, фрагмент Fd, фрагмент Fd', молекулу одноцепочечного антитела или однодоменное антитело; при этом указанной молекулой одноцепочечного антитела предпочтительно является scFv, ди-scFv, три-scFv, диатело или scFab.
[0015] В другом аспекте настоящее изобретение обеспечивает конъюгат антитела и лекарственного средства, содержащий антитело против IL17A или его антиген-связывающий фрагмент, описанный в настоящем документе и дополнительный терапевтический агент, при этом предпочтительно указанное антитело против IL17A или его антиген-связывающий фрагмент связаны с дополнительным терапевтическим агентом посредством линкера.
[0016] В еще одном аспекте, настоящее изобретение обеспечивает нуклеиновую кислоту, колирующую антитело против IL17A или его антиген-связывающий фрагмент, описанный в настоящем документе.
[0017] В одном варианте реализации указанная нуклеиновая кислота включает последовательность нуклеотидов, указанную в SEQ ID NO: 30, кодирующую вариабельный участок тяжелой цепи и/или последовательность нуклеотидов, указанную в SEQ ID NO: 31, кодирующую вариабельный участок легкой цепи.
[0018] В еще одном аспекте, настоящее изобретение обеспечивает вектор экспрессии, содержащий указанную нуклеиновую кислоту, описанную в настоящем документе.
[0019] В еще одном аспекте, настоящее изобретение обеспечивает клетку-хозяина, содержащую указанную нуклеиновую кислоту, описанную в настоящем документе или указанный вектор экспрессии, описанный в настоящем документе.
[0020] В еще одном аспекте, настоящее изобретение обеспечивает способ получения антитела против IL17A или его антиген-связывающего фрагмента, описанных в настоящем документе, включающий выращивание указанной клетки-хозяина, описанной в настоящем документе, в условиях, подходящих для экспрессии антитела, и выделение экспрессированного антитела из культуральной жидкости.
[0021] В еще одном аспекте, настоящее изобретение обеспечивает фармацевтическую композицию, содержащую антитело против IL17A или его антиген-связывающий фрагмент, описанный в настоящем документе, или конъюгат антитела и лекарственного средства, описанный в настоящем документе, или указанную нуклеиновую кислоту, описанную в настоящем документе, или указанный вектор экспрессии, описанный в настоящем документе, и фармацевтически приемлемый носитель.
[0022] В еще одном аспекте, настоящее изобретение обеспечивает антитело против IL17A или его антиген-связывающий фрагмент, описанные в настоящем документе, или конъюгат антитела и лекарственного средства, описанный в настоящем документе, или фармацевтическую композицию, описанную в настоящем документе, для лечения псориаза.
[0023] В еще одном аспекте, настоящее изобретение обеспечивает способ для лечения псориаза, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества антитела против IL17A или его антиген-связывающего фрагмента, описанных в настоящем документе, или конъюгата антитела и лекарственного средства, описанного в настоящем документе, или фармацевтической композиции, описанной в настоящем документе, излечивая таким образом псориаз.
[0024] В еще одном аспекте, настоящее изобретение обеспечивает применение антитела против IL17A или его антиген-связывающего фрагмента, описанных в настоящем документе, или конъюгата антитела и лекарственного средства, описанного в настоящем документе или фармацевтической композиции, описанной в настоящем документе, для получения лекарственного средства для лечения псориаза.
[0025] В еще одном аспекте, настоящее изобретение обеспечивает фармацевтическую композицию, содержащую антитело против IL17A или его антиген-связывающий фрагмент, описанный в настоящем документе, или конъюгат антитела и лекарственного средства, описанный в настоящем документе, или фармацевтическую композицию, описанную в настоящем документе, и один или более дополнительный терапевтический агент.
[0026] В еще одном аспекте, настоящее изобретение обеспечивает набор, содержащий антитело против IL17A или его антиген-связывающий фрагмент, описанный в настоящем документе, или конъюгат антитела и лекарственного средства, описанный в настоящем документе, или фармацевтическую композицию, описанную в настоящем документе, и предпочтительно дополнительно содержащую устройство для введения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0027] На Фиг. 1 показано, что IL17A-M069 блокирует индуцированную IL17A секрецию IL-6 клетками HFF.
[0028] На Фиг. 2 показано связывание IL17A-Н069 с рекомбинантным белком IL17A человека.
[0029] На Фиг. 3 показано связывание IL17A-H069 с рекомбинантным белком IL17A/F человека.
[0030] На Фиг. 4 показано межвидовое перекрестное связывание между IL17A-H069 и белком IL17A мыши.
[0031] На Фиг. 5 показано, что IL17A-H069 блокирует связывание IL17RA с белком IL17A.
[0032] На Фиг. 6 показано, что IL17A-H069 блокирует связывание IL17RA с белком IL17A/F.
[0033] На Фиг. 7 показано, что IL17A-H069 блокирует вызванную IL17A секрецию IL-6 клетками HFF.
[0034] На Фиг. 8 показано влияние IL17A-H069 на область псориаза и Индекс выраженности заболевания PASI в модели псориаза на мышах.
[0035] На Фиг. 9 показана кривая зависимости концентрацию лекарственного средства от времени после однократного подкожного введения IL17A-H069 у яванских макаков.
ПОДРОБНОЕ ОПИСАНИЕ
[0036] Различные аспекты настоящего изобретения относятся к изолированному антителу против IL17A или его антиген-связывающему фрагменту, конъюгату антитела и лекарственного средства, содержащему указанное антитело или его антиген-связывающий фрагмент, нуклеиновой кислоте и вектору экспрессии, кодирующим указанное антитело или его антиген-связывающий фрагмент, и клетке-хозяину, содержащей указанную нуклеиновую кислоту или вектор экспрессии, способу получения указанного антитела против IL17A или его антиген-связывающего фрагмента, фармацевтической композиции, содержащей указанное антитело против IL17A или его антиген-связывающий фрагмент, и способу применения указанного антитела против IL17A или его антиген-связывающего фрагмента для лечения псориаза.
[0037] Определения
[0038] Если не указано иное, все используемые здесь технические и научные термины имеют значения, обычно понятные специалистам в данной области техники, к которой относится настоящее изобретение. Для целей настоящего изобретения следующие термины определяются в соответствии со значениями, обычно понимаемыми в данной области техники.
[0039] При использовании в настоящем описании и в прилагаемой формуле изобретения формы единственного числа «один», «другой» и «указанный» включают обозначение объекта во множественном числе, если контекст явно не указывает на иное.
[0040] Термин «антитело» относится к молекуле иммуноглобулина и относится к любой форме антитела, которая проявляет желаемую биологическую активность. К ним относятся, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела и мультиспецифические антитела (например, биспецифические антитела), и даже фрагменты антитела, но не ограничиваются ими. Обычно, структуры полноразмерных антител предпочтительно включают четыре цепи полипептидов, две тяжелых (Н) цепи и две легких (L) цепи, обычно взаимосвязанных дисульфидными связями. Каждая тяжелая цепь содержит вариабельный участок тяжелой цепи и константный участок тяжелой цепи. Каждая легкая цепь содержит вариабельный участок легкой цепи и константный участок легкой цепи. В дополнение к этой обычной структуре полноразмерного антитела, указанная структура также включает другие производные формы.
[0041] Указанные вариабельный участок тяжелой цепи и вариабельный участок легкой цепи могут быть дополнительно подразделяться на более консервативные участки (называемые каркасными участками (FR)) и гипервариабельные участки (называемые участками, определяющими комплементарность (CDR)), разделенные промежутками.
[0042] Термин «участок, определяющий комплементарность» (CDR, например, CDR1, CDR2 и CDR3) относится к таким остаткам аминокислот в указанном вариабельном участке антитела, чье присутствие необходимо для связывания антигена. Каждый вариабельный участок обычно содержит три участка CDR, обозначенные как CDR1, CDR2 и CDR3. Каждый участок, определяющий комплементарность, может содержать остатки аминокислот из «участка, определяющего комплементарность» в соответствии с нумерацией Kabat (Kabat и др., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Бетесда, Мэриленд, 1991) и/или остатки из "гипервариабельной петли" (Chothia и Lesk; J MolBiol 196: 901-917(1987)).
[0043] Термин остатки «каркаса» или «FR» означает остатки в указанном вариабельном участке, отличающиеся от остатков CDR, как определено в настоящем документе.
[0044] Каждый вариабельный участок тяжелой цепи и вариабельный участок легкой цепи обычно содержит 3 CDR и до 4 FR, при этом указанные CDR и FR расположены от аминоконца к карбоксильному концу в следующем порядке, например: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
[0045] Участок, определяющий комплементарность, (CDR) и каркасный участок (FR) указанного антитела могут быть определены с использованием системы Kabat (Kabat и др.: Sequences of Proteins of Immunological Interest, 5th edition, US Department of Health and Human Services, PHS, NIH, NIH Publication No. 91- 3242, 1991).
[0046] Термин «константный участок» относится к такой последовательности аминокислот в легкой и тяжелой цепях антитела, которая впрямую не вовлечена в связывание указанного антитела с антигеном, но влияет на различные эффекторные функции, такие как вызываемая антителом цитотоксичность.
[0047] В соответствии с антигенными различиями последовательности аминокислот константного участка, указанная тяжелая цепь антитела может быть отнесена к одному из пяти классов: α, β, ε, γ, и μ. Когда она образует полноразмерное антитело с легкой цепью, оно может быть отнесено к одному из пяти классов: IgA, IgD, IgE, IgG и IgM, которые могут быть дополнительно отнесены к пяти подклассам (изотипам), таким как IgG1, IgG2, IgG3, IgG4, IgAn IgA2. На основании последовательности аминокислот его константного участка, легкая цепь антитела может быть классифицирована как κ и λ.
[0048] «Антиген-связывающий фрагмент антитела» означает часть интактной молекулы антитела, у которой остается по меньшей мере некоторая специфичность связывания исходного антитела и обычно включает по меньшей мере часть антиген-связывающего участка или вариабельный участок (например, один или более CDR) исходного антитело. Примеры антиген-связывающих фрагментов включают фрагмент Fv, Fab, Fab', Fab'-SH, F(ab')2, Fd, Fd', молекулы одноцепочечных антител (например, scFv, ди-scFv или три-scFv, диагело или scFab), однодоменные антитела, но не ограничиваются ими.
[0049] Термин «фрагмент антитела» относится к молекуле неполноразмерного антитела, у которой остаются по меньшей мере некоторые биологические свойства исходного антитела, включая фрагмент Fc, в дополнение к фрагментам, описанным выше как «антиген-связывающие фрагменты», но не ограничиваются ими.
[0050] Термин «конъюгат антитела и лекарственного средства» или «ADC» относится к связывающему белку, такому как антитело или его антиген-связывающий фрагмент, который химически связан с одним или более химическим средством (также обозначенном в настоящем документе как «агент»), которое необязательно может быть терапевтическим агентом или цитотоксическим агентом. В одном из предпочтительных вариантов реализации ADC включает антитело, цитотоксическое или терапевтическое лекарственное средство, и линкер, с помощью которого указанное лекарственное средство может быть связано указанным антителом или может образовывать с ним конъюгат. ADC обычно могут содержать от 1 до 8 лекарственных средств, образующих конъюгат с указанным антителом, включая 2, 4, 6 или 8 лекарственных средств. Неограничивающими примерами лекарственных средств, которые могут быть включены в ADC, являются митотические ингибиторы, противоопухолевые антибиотики, иммуномодуляторы, векторы для генной терапии, алкилирующие агенты, агенты против IL17A, антиметаболиты, борсодержащие агенты, химиотерапевтические защитные агенты, гормоны, антигормональные агенты, кортикостероиды, фотоактивные терапевтические агенты, олигонуклеотиды, радионуклидные агенты, ингибиторы топоизомеразы, ингибиторы тирозинкиназы и радиосенсибилизаторы.
[0051] Термин «химерные антитело» относится к антителу, в котором часть указанной тяжелой цепи и/или легкой цепи получена из одного конкретного источника или вида, а оставшаяся часть получена из другого источника или вида. Указанное «химерное антитело» также может быть функциональным фрагментом, описанным выше. «Гуманизированное антитела» являются подмножеством «химерных антител».
[0052] Термин «гуманизированное антитело» или «гуманизированный антиген-связывающий фрагмент» в настоящем документе означает антитело или фрагмент антитела, который: (i) получен из источника, не являющегося человеком, (например, трансгенной мыши, имеющей гетерологичную иммунную систему) и основан на последовательности зародышевой линии человека; или (ii) является химерным антителом, в котором вариабельный участок имеет происхождение не из человека, а константный участок происходит из человека; или (iii) трансплантат CDR, в котором указанный CDR вариабельного участка имеет происхождение не из человека, один или более каркасных участков вариабельного участка происходят из человека и константный участок, если есть, происходит из человека. Целью «гуманизации» является устранение иммуногенности антител нечеловеческого происхождения в организме человека при сохранении максимально возможной аффинности. В качестве шаблона для гуманизации целесообразно выбрать последовательность каркаса человека, которая наиболее похожа на последовательность каркаса антитела, происходящего их источника, не являющегося человеком. В некоторых случаях может оказаться необходимым заменить одну или несколько аминокислот в каркасной последовательности человека соответствующими остатками в конструкции не из человека, чтобы избежать потери аффинности.
[0053] Термин «моноклональное антитело» относится к антителу, полученному из по существу гомогенной популяции антител, т.е. каждое отдельное антитело, входящее в указанную популяцию, является идентичным, за исключением возможных мутаций (например, природных мутаций), которые могут присутствовать в очень маленьких количествах. Таким образом, термин «моноклональное» указывает на природу рассматриваемого указанного антитела, т.е., а не на смесь неродственных антител. В отличие от препаратов поликлональных антител, которые обычно включают разные антитела против разных эпитопов, все моноклональные антитела в препаратах моноклональных антител направлены против одного эпитопа антигена. В дополнение к его специфичности, препараты моноклональных антител имеют то преимущество, что они обычно не загрязнены другими антителами. Термин «моноклональные» не следует понимать как требующее производства указанного антитела каким-либо конкретным способом.
[0054] Указанное антитело «специфически связывается» с целевым антигеном, таким как ассоциированный с опухолью пептидный антиген-мишень (в данном случае, IL17A), т.е. связывает указанный антиген с достаточной аффинностью, чтобы сделать возможным использование указанного антитело в качестве терапевтического агента, нацеливаясь на клетку или ткань, экспрессирующую указанный антиген, и не дает значительной перекрестной реакции с другими белками или не дает значительной перекрестной реакции с белками, отличными от гомологов и вариантов белков-мишеней, упомянутых выше (например, мутантные формы, варианты сплайсинга, или укороченные формы белка, полученные в результате гидролиза).
[0055] Термин «аффинность связывания» относится к силе суммы нековалентных взаимодействий между отдельными сайтами связывания молекулы и ее партнерами по связыванию. Если не указано иное, «аффинность связывания» при использовании в настоящем документе относится к внутренней аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающей пары (например, антитело и антиген). В настоящем тексте термин «KD» относится к равновесной константе диссоциации взаимодействия антитело-антиген. В настоящем тексте термин «kon» относится к константе скорости, при которой антитело связывается с антигеном. В настоящем тексте термин «koff» относится к константе скорости, при которой антитело диссоциирует из комплекса антитело/антиген. «KD», «константа скорости связывания kon» и «константа скорости диссоциации koff» обычно используются для описания сродства между молекулой (например, антителом) и ее партнером по связыванию (например, антигеном). Аффинность, т.е., степень силы, при которой лиганд связывается с определенным белком. На аффинность связывания влияют нековалентные межмолекулярные взаимодействия, такие как водородные связи, электростатические взаимодействия, гидрофобные и ван-дер-ваальсовы силы между двумя молекулами. Кроме того, на аффинность связывания между лигандом и его целевой молекулой может влиять присутствие других молекул. Аффинность может быть проанализирована традиционными методами, известными в данной области техники, включая ИФА, описанный в настоящем документе.
[0056] Термин «эпитоп» включает любой кластер белковой детерминанты, который специфически связывается с антителом или Т-клеточным рецептором. Кластеры эпитопной детерминанты обычно состоят из химически активных поверхностных групп молекулы (например, боковых цепей аминокислот или сахаров или их комбинации) и часто имеют специфические трехмерные структурные признаки, а также специфические характеристики заряда.
[0057] Термин «изолированное» антитело представляет собой антитело, которое было идентифицировано и выделено из компонентов клетки, где экспрессируется указанное антитело. Изолированные антитела включают антитела in situ внутри рекомбинантных клеток, где меньше по мере один компонент в естественной среде указанного антитела отсутствует. Однако, как правило, изолированное антитело получают посредством по меньшей мере одной стадии очистки.
[0058] «Идентичность последовательностей» между двумя последовательностями полипептидов или нуклеиновых кислот указывает количество остатков, которые идентичны между указанными последовательностями в процентах от общего числа остатков, и рассчитывается на основе размера меньшей из сравниваемых молекул. При вычислении идентичности в процентах сравниваемые последовательности сопоставляются таким образом, чтобы получить максимальное совпадение между указанными последовательностями, при этом пробелы при сравнении (если они есть) разрешаются с помощью определенного алгоритма. Предпочтительные компьютерные программные методы для определения идентичности между двумя последовательностями включают наборы программ GCG, включая GAP, BLASTP, BLASTN и FASTA (Altschul и др., 1990, J. Mol. Biol. 215: 403-410), но не ограничиваются ими. Вышеупомянутые процедуры общедоступны в Международном центре биотехнологической информации (NCBI) и других источниках. Известный алгоритм Смита-Уотермана также может быть использован для определения идентичности.
[0059] Термин «Fc рецептор» или «FcR» относится к рецептору, который связывается с участком Fc антитела. Предпочтительны природные последовательности FcR человека и предпочтительно рецепторы, которые связываются с IgG антителами (гамма-рецепторы), которые включают FcyRI, FcyRII и изоформы FcyRIII, а также варианты из этих рецепторов. Все остальные FcR включены в термин «FcR». Термин также включает неонатальный рецептор (FcRn), который отвечает за транспорт материнского IgG к плоду (Guyer и др., Journal of Immunology 117: 587 (1976) и Kim и др., Journal of Immunology 24: 249 (1994)).
[0060] Термин «неонатальный Fc-рецептор», сокращенно «FcRn», связывается с участком Fc IgG-антитела. Неонатальный Fc-рецептор (FcRn) играет важную роль в метаболической судьбе IgG-подобных антител in vivo. FcRn функционирует для спасения IgG от пути лизосомной деградации, тем самым снижая его клиренс в сыворотке и удлиняя период его полувыведения. Таким образом, свойства связывания FcRn /характеристики IgG in vitro указывают на его фармакокинетические свойства в кровотоке in vivo.
[0061] Термин «эффекторная функция» относится к той биологической активности, которая приписывается участку Fc антитела, которая варьирует от изотипа к изотипу. Примеры эффекторной функции антитела включают связывание C1q и комплемент-зависимую цитотоксичность (CDC), связывание рецептора Fc, антитело-зависимую клеточно-опосредованная цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз (ADCP), секреция цитокинов, опосредованное иммунными комплексами поглощение антигена антигенпрезентирующими клетками, подавление рецепторов клеточной поверхности (например, рецепторов В-клеток) и активация В-клеток.
[0062] Термин «эффекторная клетка» относится к клетке, которая экспрессирует один или несколько FcR и выполняет эффекторные функции. В одном аспекте указанные эффекторные клетки экспрессируют в меньшей степени FcγRIII и выполняют эффекторные функции ADCC. Примеры клеток человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови, (РВМС), натуральные киллеры (NK), моноциты, цитотоксические Т-клетки и нейтрофилы. Эффекторные клетки могут быть выделены из природных источников, например, крови. Эффекторные клетки обычно представляют собой лимфоциты, ассоциированные с эффекторной фазой и функционирующие для продукции цитокинов (хелперные Т-клетки), уничтожения клеток, инфицированных патогенами (цитотоксические Т-клетки) или секреции антитела (дифференцированные В-клетки).
[0063] «Иммунные клетки» включают клетки, которые имеют гемопоэтическое происхождение и играют роль в иммунном ответе. Иммунные клетки включают: лимфоциты, такие как В-клетки и Т-клетки; естественные клетки-киллеры; и миелоидные клетки, такие как моноциты, макрофаги, эозинофилы, тучные клетки, базофилы и гранулоциты.
[0064] «Антитело-зависимая клеточно-опосредованная цитотоксичность» или «ADCC» относится к форме цитотоксичности, при которой секретируемый Ig связывается с рецепторами Fcγ, представленными на определенных цитотоксических клетках (например, NK-клетках, нейтрофилах и макрофагах), что позволяет этим цитотоксическим эффекторным клеткам специфически связываться с клетками-мишенями, несущими антигены, и последующего уничтожения указанных клеток-мишеней с использованием, например, цитотоксина. Для оценки ADCC-активности целевого антитела могут быть выполнены анализы ADCC in vitro, такие как анализы ADCC in vitro, задокументированные в патенте США №5,500,362 или 5,821,337 или патенте США №6,737,056 (Presta). Полезные эффекторные клетки для применения в таких случаях включают РВМС и NK-клетки.
[0065] «Комплемент-зависимая цитотоксичность» или «CDC» относится к лизису клеток-мишеней в присутствии комплемента. Классический путь активации комплемента инициируется связыванием первого компонента системы комплемента (C1q) с антителом (соответствующего подкласса), которое связывается с соответствующим антигеном. Для оценки активации комплемента проводят анализ CDC, такой как анализ CDC, описанный в Gazzano-Santoro и др., J. Immunol Methods 202: 163 (1996). Например, в патенте США №6,194,551 В1 и WO 1999/51642, описаны варианты полипептидов, содержащие измененную последовательность аминокислот участка Fc (полипептиды, содержащие вариант участка Fc) и варианты полипептидов, имеющие повышенное или пониженное связывание C1q.
[0066] Термины «биоэквивалент COSENTYX» и «биоэквивалент Taltz» относятся к антителам, полученным в соответствии со структурами COSENTYX и Taltz, соответственно.
[0067] Последовательность аминокислот указанного антитела согласно настоящему изобретению
[0068] В настоящем изобретении использовали рекомбинантный белок IL17A человека для иммунизации мышей, а затем получали клон антитела IL17A-M069, которое специфически связывается с рекомбинантным белком IL17A человека посредством скрининга библиотеки фагового дисплея. Указанную последовательность нуклеотидов, кодирующую вариабельные участки тяжелой и легкой цепи антитела IL17A-M069 scFv затем вводили посредством ПЦР в векторы pSTEP2, содержащие последовательность нуклеотидов, кодирующую константный участок IgG1 мыши или константный участок легкой цепи каппа мыши, соответственно, и выращивали для экспрессии. Антитела высокой степени очистки получали посредством очистки на колонке с белком А. ИФА показал, что указанное антитело мыши было способно блокировать индуцируемую IL17A секрецию IL-6 клетками HFF.
[0069] Затем, используя классический метод гуманизированной трансплантации CDR, в качестве матрицы были выбраны вариабельные участки легкой цепи или тяжелой цепей антитела человека, последовательность которых ближе к указанной последовательности вариабельных участков легкой или тяжелой цепей мыши, гуманизированный вариабельный участок легкой цепи (VL) и вариабельный участок тяжелой цепи (VH) по очереди путем вставки каждого из трех CDR (Таблица 1) легкой цепи или тяжелой цепи антитела кролика в вариабельные участки указанного антитела человека. Поскольку ключевые сайты каркасного участка антитела мыши необходимы для поддержания стабильности активности CDR, ключевые сайты были мутированы обратно в соответствующую последовательность антитела мыши. Векторы экспрессии легкой цепи/тяжелой цепи IL17A-H069 получали путем полного синтеза гена, трансфицировали его в дефицитные клетки CHO-K1-GS и культивировали для экспрессии, клоны, имеющие наиболее высокую степень экспрессии антитела отбирали для дальнейшего выращивания, в результате чего получали антитело IL17A-Н069 высокой степени очистки и высокого качества.
[0070] Нуклеиновые кислоты согласно настоящему изобретению
[0071] Настоящее изобретение также относится к молекулам нуклеиновой кислоты, кодирующей антитела или их фрагменты согласно настоящему изобретению. Последовательности этих молекул нуклеиновый кислоты включают SEQ ID NO: 2-7, 26-33, 36-37, 40-41 и 43, но не ограничиваются ими.
[0072] Указанные молекулы нуклеиновой кислоты согласно настоящему изобретению не ограничены указанными последовательностями, описанными в настоящем документе, но также включают их варианты. Варианты в настоящем изобретении могут быть описаны со ссылкой на их физические свойства при гибридизации. Специалисту в данной области техники понятно, что с использованием методов гибридизации нуклеиновых кислот, нуклеиновые кислоты может применяться для идентификации их комплементов, а также их эквивалентов и гомологов. Также понятно, что гибридизация может происходить при комплементарности, меньшей 100%. Однако при соответствующем выборе условий методы гибридизации могут применяться для различения указанных последовательностей ДНК на основе структурной релевантности к последовательности ДНК конкретного зонда. Руководство по таким условиям смотри Sambrook и др., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, Cold Spring Harbor, N.Y., 1989 и Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Sedman, J.G., Smith, J. А. и Struhl, K. eds. (1995). Current Protocols in Molecular Biology. New York: John Wiley и Sons.
[0073] Рекомбинантные векторы и экспрессия
[0074] Согласно настоящему изобретению также предложены рекомбинантные конструкции, содержащие одну или более последовательностей нуклеотидов согласно настоящему изобретению. Указанную рекомбинантную конструкцию согласно настоящему изобретению получают посредством вставки указанной молекулы нуклеиновой кислоты, кодирующей указанное антитело согласно настоящему изобретению, в вектор, такой как плазмида, фагмида, фаг или вирусный вектор.
[0075] Антитела, предлагаемые в настоящем документе, могут быть получены посредством рекомбинантной экспрессии последовательности нуклеотидов, кодирующей легкую и тяжелую цепи или их части в клетке-хозяине. Для рекомбинантной экспрессии указанного антитела, указанная клетка-хозяин может быть трансфицирована одним или более рекомбинантными векторами экспрессии, несущими последовательность нуклеотидов, кодирующую легкую и/или тяжелую цепи или их части, так, что указанная легкая и тяжелая цепи экспрессируются в указанной клетке-хозяине. Стандартную методологию рекомбинантных ДНК используют для получения нуклеиновых кислот, кодирующих тяжелую и легкую цепи, для включения указанных нуклеиновых кислот в рекомбинантные векторы экспрессии и для введения указанных векторов в клетки-хозяева, например, Sambrook, Fritsch и Maniatis (eds.), Molecular Cloning; A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y., (1989), Ausubel, F.M. и др. (eds.) Current Protocols in Molecular Biology, Greene Publishing Associates, (1989) и описанную в патенте США №4,816,397 Boss и др.
[0076] Подходящими клетками-хозяевами являются прокариотические и эукариотические клетки. Примерами прокариотических клеток-хозяев являются бактерии, а примерами эукариотических клеток-хозяев являются клетки дрожжей, насекомых или млекопитающего. Следует понимать, что дизайн векторной экспрессии, включая выбор регуляторной последовательности, определяется рядом факторов, таких как выбор клетки-хозяина, уровень экспрессии желаемого белка, и является ли экспрессия конститутивной или индуцируемой.
[0077] Бактериальная экспрессия
[0078] Вектор экспрессии для применения в бактерии конструируют путем вставки структурной последовательности ДНК, кодирующей желаемое антитело вместе с соответствующими сигналами инициации и терминации трансляции и функциональными промоторами в действующую рамку считывания. Вектор содержит один или несколько маркеров фенотипической селекции и точку начала репликации, чтобы обеспечить сохранение вектора и обеспечить амплификацию в хозяине по мере необходимости. Подходящие прокариотические хозяева для трансформации включают несколько видов Е. coli, Bacillus subtilis, Salmonella typhimurium, а также Pseudomonas, Streptomyces и Staphylococcus.
[0079] Указанный бактериальный вектор может быть на основе, например, фага, плазмиды или фагмиды. Эти векторы могут содержать маркеры селекции и бактериальные точки начала репликации, которые получены из коммерчески доступных плазмид, которые обычно содержат элементы хорошо известного вектора клонирования pBR322 (АТСС 37017). после трансформации соответствующего штамма-хозяина и выращивания штамма-хозяина до соответствующей плотности клеток выбранный промотор дерепрессируют/индуцируют соответствующим методом (например, изменением температуры или химической индукцией), и клетки культивируют в течение еще некоторого времени. Клетки обычно собирают центрифугированием, разрушают физическими или химическими методами, а полученный неочищенный экстракт сохраняют для дополнительной очистки.
[0080] В бактериальной системе множество векторов экспрессии могут быть выгодно подобранными в соответствии с предполагаемым использованием экспрессированного белка. Например, когда необходимо произвести большое количество таких белков для производства антител или для скрининга библиотеки пептидов, например, может потребоваться вектор, который позволяет осуществить высокоуровневую экспрессию слитого белка продукта для легкой очистки.
[0081] Экспрессия в клетках млекопитающих и очистка
[0082] Предпочтительные регуляторные последовательности для экспрессии в клетках-хозяевах млекопитающих включают вирусные элементы, которые позволяют осуществить высокоуровневую экспрессию белка в клетках млекопитающего, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV) (например, промотор/энхансер CMV), промоторы и/или энхансеры вируса обезьян 40 (SV40) (например, промотор/энхансер SV40), промоторы и/или энхансеры аденовируса (например, главный поздний промотор аденовируса (AdMLP)) и промоторы и/или энхансеры вируса полиомы. Для дополнительного описания вирусных регуляторных элементов и их последовательностей, см., например, патент США №5,168,062, Stinski, патент США №4,510,245, Bell и др., и патента США №4,968,615, Schaffner и др. Указанные рекомбинантные векторы экспрессии также могут включать точку начала репликации и маркер селекции (см., например, патенты США №4,399,216, №4,634,665 и №5,179,017, Axel и др.), подходящие маркеры селекции включают гены, которые обеспечивают устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клеткам-хозяевам, в которые введен указанный вектор. Например, ген дигидрофолатредуктазы (DHFR) обеспечивает устойчивость к метотрексату, в то время как ген neo обеспечивает устойчивость к G418.
[0083] Трансфекцию указанного вектора экспрессии в клетки-хозяева можно проводить с использованием стандартных методов, таких как электропорация, осаждение с фосфатом кальция и трансфекция DEAE-декстраном.
[0084] Подходящие клетки-хозяева млекопитающих для экспрессии представленного в настоящем документе антитела включают клетки яичника китайского хомячка (клетки СНО) [включая клетки dhfr-CHO, описанные Urlaub и Chasin, (1980) Proc. Natl. Acad. Sci. USA 77:4216-4220, с применением маркера селекции DHFR, как описано, например, в R.J. Kaufman и Р.А. Sharp (1982) Mol. Biol. 159:601-621], клетки миеломы NSO, клетки COS и клетки SP2.
[0085] Антитела согласно настоящему изобретению могут быть выделены и очищены от культуры рекомбинантных клеток известными способами, включая осаждение сульфатом аммония или этанолом, кислотную экстракцию, аффинную хроматографию на белке А, аффинную хроматографию на белке G, анионо- или катионообменную хроматография, хроматографию на фосфоцеллюлозе, хроматографию гидрофобного взаимодействия, аффинную хроматографию, хроматографию на гидроксиапатите, хроматографию на лектинах, но не ограничиваются ими. Высокоэффективная жидкостная хроматография (ВЭЖХ) также может применяться для очистки. См., например, Colligan, Current Protocols in Immunology, или Current Protocols in Protein Science, John Wiley & Sons, NY, N.Y., (1997-2001), например, главы 1, 4, 6, 8, 9, и 10, каждая из которых полностью включена в настоящий документ посредством ссылки.
[0086] Характеристики и функции антитела согласно настоящему изобретению
[0087] Осуществляли анализ характеристик и функциональный анализ гуманизированного антитела IL17A-Н069 согласно настоящему изобретению. Анализ показал, что указанное антитело согласно настоящему изобретению обладает следующими преимуществами:
[0088] (1) Лучшее специфическое связывание с рекомбинантным белком IL17A человека, чем биоаналог COSENTYX (EC50 гуманизированного антитела IL17A-H069 составляет 46 нг/мл, при этом ЕС50 биоаналога COSENTYX составляет 74,8 нг/мл) (Пример 4.1.1).
[0089] (2) Лучшее специфическое связывание с рекомбинантным димерным белком IL17A/F человека, чем у биоаналога COSENTYX (ЕС50 гуманизированного антитело IL17A-H069 составляет 36,3 нг/мл; в то время как ЕС50 биоаналога COSENTYX составляет 63,9 нг/мл) (Пример 4.1.2).
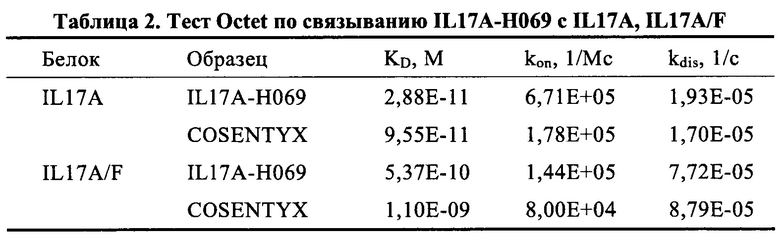
[0090] (3) Хорошая аффинность связывания (намного лучшая, чем у биоаналога COSENTYX; 2,88Е-11 М против 9,55-11 М) и благоприятный коэффициент ассоциации (намного быстрее, чем у биоаналога COSENTYX; 6,71Е+05 M-1с-1 против 1,78Е+05 М-1с-1) с рекомбинантным белком IL17A человека, хорошая аффинность связывания (выше, чем у биоаналога COSENTYX; 5,37Е-10 М против 1,10-09 М) и благоприятный коэффициент ассоциации (быстрее, чем у биоаналога COSENTYX; 1,44Е+05 M-1с-1 против 8,00Е+04 М-1с-1) (Пример 4.1.3).
[0091] (4) Отсутствие активности по перекрестному связыванию с белком mIL17A мыши (Пример 4.1.4).
[0092] (5) Эффективно связывает рекомбинантный белок IL17A человека и эффективно ингибирует связывание белка IL17A с рецептором IL17RA, намного лучше, чем биоаналог COSENTYX, но сравнимо с биоаналогом Taltz (IC50: 0,50 мкг/мл против 2,99 мкг/мл против 0,50 мкг/мл; максимальная степень ингибирования: 85,4% против 73,5% против 89,5% (Пример 4.2.1).
[0093] (6) Эффективно связывает рекомбинантный димерный белок IL17A/F человека и эффективно ингибирует связывание белка IL17A с рецептором IL17RA, лучше, чем биоаналог COSENTYX и биоаналог Taltz (IC50: 1,02 мкг/мл против 1,2 мкг/мл против 1,35 мкг/мл; максимальная степень ингибирования: 92,3%, 87,9% и 75%) (Пример 4.2.2).
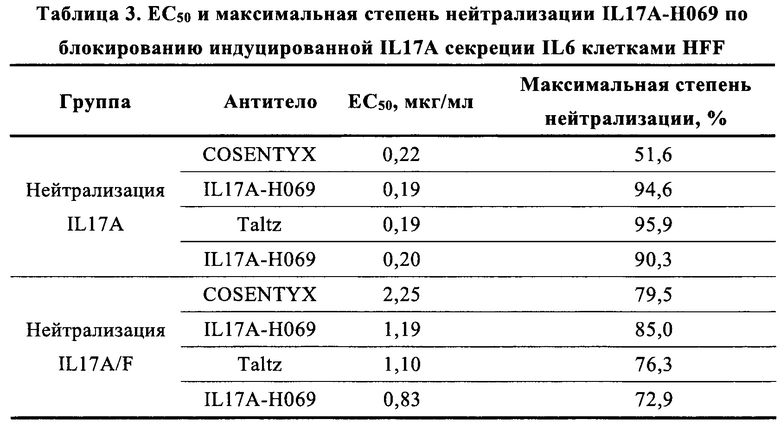
[0094] (7) Блокирует индуцируемую IL17A, IL17A/F секрецию IL-6 клетками HFF; активность по нейтрализации IL17A намного выше, чем у биоаналога COSENTYX (ЕС50 0,19 мкг/мл против 0,22 мкг/мл, максимальная степень нейтрализации 94,6% портив 51,6%) и активность по нейтрализации IL17A близка к биоаналогу Taltz (ЕС50 0,20 мкг/мл против 0,19 мкг/мл, максимальная степень нейтрализации 90,3% против 95,9%); активность по нейтрализации IL17A/F также выше, чем у биоаналога COSENTYX (ЕС50 1,19 мкг/мл против 2,25 мкг/мл, максимальная степень нейтрализации 85,0% против 79,5%); и активность по нейтрализации IL17A/F лучше, чем у биоаналога Taltz при низких концентрациях, но близка к биоаналогу Taltz при высоких концентрациях (ЕС50: 0,83 мкг/мл против 1,10 мкг/мл, максимальная степень нейтрализации: 72,90% против 76,3%) (Против 5.1).
[0095] (8) Будучи проверенным на модели псориаза у мышей, антитело согласно настоящему изобретению эффективно облегчает прогрессирование псориаза и уменьшает симптомы со значительно большей эффективностью, чем биоаналог Taltz. (Пример 6).
[0096] (9) Фармакокинетически антитело согласно настоящему изобретению имеет более быструю абсорбцию после подкожной инъекции и более длительный период полувыведения (Пример 7).
[0097] Применения
[0098] Антитело согласно настоящему изобретению может применяться для лечения рака толстой и прямой кишки. Антитело настоящего изобретения также может быть использовано для получения лекарственного средства для лечения указанных заболеваний.
[0099] Фармацевтические композиции
[00100] Антитела согласно настоящему изобретению могут быть получены с по меньшей мере одним другим агентом (например, стабильным соединением) с образованием фармацевтической композиции, содержащий антитело согласно настоящему изобретению и один или более фармацевтически приемлемые носители, разбавители или вспомогательные вещества. Указанная фармацевтическая композиция необязательно может содержать дополнительные терапевтические агенты.
[00101] Наборы
[00102] Настоящее изобретение также относится к фармацевтическому набору и набору, содержащему один или более контейнеров, где указанные контейнеры содержат указанные выше фармацевтические композиции согласно настоящему изобретению. К таким контейнерам могут прилагаться спецификации по форме, установленной государственным органом, регулирующим производство, использование или распространение лекарственного средства или биологического продукта, которые отражают одобрение его введения человеку указанным органом страны, в которой указанный продукт производится, используется или распространяется.
[00103] Получение и хранение
[00104] Указанная фармацевтическая композиция согласно настоящему изобретению может быть получена способом, известным в данной области техники, например, традиционными методами смешивания, растворения, грануляции, приготовления пастилок, измельчения, эмульгирования, инкапсуляции, заливки или лиофилизации.
[00105] Будучи приготовленными, фармацевтические композиции, содержащие соединения согласно настоящему изобретению, изготовленные в виде лекарственной формы в приемлемом носителе, они могут быть помещены в соответствующие контейнеры и помечены для лечения указанного состояния. Такая маркировка должна включать количество, частоту и способы введения препарата.
[00106] Комбинации
[00107] Указанная фармацевтическая композиция, содержащая антитела согласно настоящему изобретению, описанные выше, также может быть скомбинирована с одним или более другими терапевтическими агентами, такими как противоопухолевые агенты, если при этом полученная комбинация не вызывает неприемлемых побочных эффектов.
ПРИМЕРЫ
[00108] Настоящее изобретение будет лучше понято со ссылкой на следующие неограничивающие экспериментальные примеры. Все экспериментальные методы в последующих примерах, если не указано иное, являются обычными методами. Экспериментальные материалы, использованные в последующих примерах, если не указано иное, были приобретены у обычных дистрибьюторов биохимических реагентов.
[00109] Пример 1: Скрининг антител против IL17A
[00110] 1.1 Иммунизация мышей
[00111] Мышей иммунизировали белком IL17A в соответствии с методом, описанным StGroth и др. (de StGroth и Scheidegger 1980) с подходящими модификациями. Рекомбинантный белок IL17A человека (от SinoBiological, Inc, кат. №10247-Н07В) использовали для иммунизации мышей. Последовательность аминокислот белка ПЛ7А (UniprotKB Q16552) представляет собой последовательность Met1-Ala155 (SEQ ID NO: 1). Рекомбинантный белок IL17A человека смешивали с адъювантомна основе фосфата алюминия (при этом первую и четвертую иммунизации осуществляли с дополнительным полным адъювантом Фрейнда CFA в виде эмульсии в фосфатно-солевом буфере, и мышей иммунизировали подкожно в нескольких местах в дозировке 50 мкг на дозу указанной смеси в течение 4 раз с интервалами 2 недели, 3 недели, и 3 недели, соответственно. Начиная с третьей иммунизации, кровь собирали через семь дней после каждой иммунизации через медиальные кантальные сплетения глаз. Титр антитела против IL17A мыши в сыворотке измеряли с помощью ИФА с использованием покрытия рекомбинантным белком IL17A человека. Титре в сыворотке после четвертой иммунизации достиг желаемого уровня (ИФА, OD>1,0) после разбавления 1:8000, и этим мышах вводили бустерную дозу внутривенно в количестве 25 мкг рекомбинантного белка IL17A человека через 75 суток после четвертой иммунизации. Через 4 суток мышей умерщвляли, ткань селезенки удаляли и замораживали в жидком азоте.
[00112] 1.2 Конструирование и скрининг библиотеки фагового дисплея антител
[00113] РНК экстрагировали из ткани селезенки мыши с использованием TriPure Isolation Reagent (от Roche, кат. №11 667 165 001), и получали кДНК обратной транскрипцией РНК с использованием набора для обратной транскрипции (от Invitrogen Кат. №18080-051). Конструировали 2 пар праймеров для амплификации последовательности вариабельного участка легкой цепи антитела мыши и 1 пару праймеров для амплификации последовательности вариабельного участка тяжелой цепи в соответствии с методом, описанным в Jones и Bendig 1991. Указанные последовательности, кодирующие вариабельные участки легкой и тяжелой цепи антитела мыши объединяли с последовательностью нуклеотидов, кодирующей scFv, с помощью ПЦР с удлинением перекрывающихся участков, затем эти две последовательности нуклеотидов соединяли с помощью линкера (SEQ ID NO: 2) для объединения с последовательностью нуклеотидов, кодирующей scFv; затем ферментативно лигировали в фаговый вектор pComb3x (Sino Biological, Inc.) с помощью эндонуклеазы рестрикции SƒiI (Fermentas), и электротрансформировали в компетентные клетки Х-Blue для получения библиотеки фагового дисплея антител scFv мыши; размер библиотеки представляет собой (sic). Фаговую библиотеку, обогащенную положительными антителами против IL17A, получали скринингом с помощью метода ИФА способом фаговой сортировки антител (Aitken 2002). Антитела scFv, которые специфически связывались с рекомбинантным белком IL17A человека, экспрессировали отдельными колониями фагов из обогащенной библиотеки и тестировали на их связывание с рекомбинантным белком IL17А человека с помощью ИФА. Последовательность нуклеотидов антитела scFv одной колонии отсеквенировали как SEQ ID NO: 3, и антитело, названное IL17A-М069, получали из этой колонии после стадий в Примере 1.3.
[00114] 1.3 Получение моноклонального антитела мыши против IL17A
[00115] Последовательность нуклеотидов, кодирующую вариабельный участок тяжелой цепи (SEQ ID NO: 4) с последовательностью сигнального пептида тяжелой цепь (SEQ ID NO: 43) и последовательность константного участка тяжелой цепи IgG1 мыши (SEQ ID NO: 6) указанного антитела scFv, амплифицировали и собирали с помощью ПЦР с удлинением перекрывающихся участков, вводили в вектор pSTEP2, расщепленный HindIII+XbaI (Fermentas), получая таким образом вектор экспрессии полной тяжелой цепи (SEQ ID NO: 36); аналогично, последовательность нуклеотидов, кодирующую вариабельный участок легкой цепи (SEQ ID N0: 5) с последовательностью сигнального пептида легкой цепи (SEQ ID NO: 29) и последовательностью константного участка легкой цепи каппа мыши (SEQ ID NO: 7) указанного антитела scFv, амплифицировали и собирали с помощью ПЦР с удлинением перекрывающихся участков, и вводили в вектор pSTEP2, расщепленный ферментами рестрикции HindIII+XbaI (Fermentas), получая таким образом вектор экспрессии полной легкой цепи (SEQ ID NO: 37). Праймеры для сборки сигнального пептида тяжелой цепи, вариабельного участка тяжелой цепи и константного участка тяжелой цепи IgG1 следующие:

[00116] Праймеры для сборки сигнального пептида легкой цепи, вариабельного участка легкой цепи и константного участка легкой цепи каппа мыши следующие:

[00117] Клетки 293Е (АТСС) пересевали в среде SCD4-4-TC2 (SinoBiological, Inc.) в колбе объемом 200 мл на колбу с начальной плотностью засева 0,3~0,4*106 клеток/мл, и колбу культивировали в качалке с СО2 со скоростью вращения 175 об/мин при 37°С, до достижения плотности клеток 1,5~3*106 клеток/мл. Затем, плазмиды, кодирующие легкую цепь и тяжелую цепь смешивали в соотношении 1:1, и 100 мкг смешанной ДНК плазмид и 800 мкл реагента для трансфекции TF2 добавляли в колбу для культивирования, которую затем культивировали в качалке со скоростью вращения 175 об/мин при 37°С в течение 7 суток для последующего сбора. Культуральную жидкость центрифугировали при 4000 об/мин в течение 25 мин, супернатант собирали и добавляли буфер для хранения. После уравновешивания колонки для хроматографии с белком А объемом 1/5 объема супернатанта с помощью фосфатно-солевого буфера в количестве 5-10 объемов колонки, отфильтрованный супернатант наносили на указанную хроматографическую колонку и снова уравновешивали с помощью 5-10 объемов колонки, затем содержимое колонки элюировали с помощью буфера ацетата натрия для сбора образца. Образец нейтрализовали с помощью Трис, в результате чего получали моноклональное антитело высокой степени очистки в нейтральном растворе.
[00118] Пример 2: Функциональный анализ моноклонального антитела мыши против IL17A
[00119] 2.1 Антитело IL17A-M069 мыши блокирует индуцированную IL17A секрецию IL-6 клетками HFF
[00120] Как описано у Beerli, Bauer и др. 2014, IL17A стимулирует секрецию цитокинов IL-6 фибробластами крайней плоти человека HFF в условиях in vitro. Антитело против IL17A добавляли в эту систему для оценки нейтрализующего эффекта антитело против IL17A в отношении IL17A путем определения секреции IL-6 клетками HFF. Клетки HFF (АТСС, SCRC-1041) инокулировали в 96-луночный планшет при плотности клеток 1×104 на лунку и выращивали в течение ночи в среде ДМЭМ, содержащей 15% ФБС. Различные концентрации антитела IL17A-M069 и положительного контроля биоаналога Taltz добавляли соответственно в количестве 10 мкл на лунку на следующий день, затем белок IL17A в конечной концентрации 50 нг/мл добавляли в количестве 10 мкл на лунку. Указанный 96-луночный планшет инкубировали при 37°С, 5% СО2 в инкубаторе клеток в течение 48 ч, и использовали пустую лунку В (без клеток), отрицательный контроль М' (клетки инокулировали, без образца антитела, добавлен IL17A) и М (клетки инокулировали, без образца антитела и без IL17A). После инкубации, супернатант собирали и секрецию IL-6 измеряли с помощью ИФА. Из уровня секреции DL-6 в лунках с образцом и в группе лунок М' вычитали уровень секреции IL-6 в группе лунок М, соответственно, для расчета степени ингибирования. Степень ингибирования % = (1 - секреция IL-6 в лунках с образцом) / секреция IL-6 в группе лунок М' × 100%. Стандартную кривую рассчитывали с использованием статистического программного обеспечения, принимая значение концентрации образца антитело в качестве горизонтальной координаты и секрецию IL-6 в качестве вертикальной координаты. Результаты показаны на Фиг. 1, антитело IL17A-М069 мыши может блокировать индуцированную IL17A секрецию IL-6 клетками HFF, и максимальная степень ингибирования и средняя концентрация ингибирования IL17A-М069 белка IL17A были схожи со значениями биоаналога Taltz, использованного в качестве положительного контроль, максимальная степень ингибирования антитела IL17A-M069 мыши и биоаналога Taltz составляла 96,33% и 97,35% соответственно, а значение EC50 было 243,3 нг/мл и 246,6 нг/мл, соответственно. Следовательно, IL17A-М069 представляет собой подходящее антитело с хорошей активностью in vitro, последующую модификацию гуманизации и функциональный анализ осуществляли именно с IL17A-M069.
[00121] Пример 3: Гуманизация и получение антитела IL17A-M069 против IL17A
[00122] На основании результатов функционального анализа антитела IL17A-M069 мыши в Примере 2, гуманизацию и получение осуществляли соответствующим образом.
[00123] 3.1 Определение последовательностей CDR легкой и тяжелой цепей антитела IL17A-M069 против IL17A
[00124] Последовательность аминокислот вариабельных участков тяжелой цепи и легкой цепи антитела IL17A-M069-scFv получали из последовательности нуклеотидов антитела IL17A-M069-scFv, определенной в Примере 1.3, смотри SEQ ID NO: 8/9.
[00125] Последовательность аминокислот каждой из трех CDR легкой и тяжелой цепей антитела IL17A-M069-scFv мыши определяли со ссылкой на индекс Кабата (Abhinandan и Martin 2008, Dondelinger, Filee и др. 2018) и нумерацию системы IMGT (Lefranc 2014), смотри Таблицу 1 и SEQ ID NO: 10-15. Указанные выше соответствующие три CDR легкой цепи и тяжелой цепи трансплантировали на последующих стадиях и сохраняли в конечном полученном гуманизированном антителе IL17A-H069, смотри Примеры 3.2 и 3.3.

[00126] 3.2 Трансплантация CDR антитела мыши
[00127] Гуманизацию антитела мыши осуществляли с использованием классического метода гуманизации трансплантацией CDR (Kettleborough, Saldanha и др. 1991). Вариабельные участки легкой или тяжелой цепи антитела, человека, чьи последовательности являются наиболее близкими указанным последовательностям вариабельных участков легкой или тяжелой цепи мыши (сходство > 50%), выбирали в качестве матрицы, и последовательности каждого из трех CDR (SEQ ID NO: 10-15) из легкой или тяжелой цепи мыши вводили в вариабельный участок антитело человека, в результате чего получали гуманизированные последовательности аминокислот вариабельного участка легкой цепи (VL) или вариабельного участка тяжелой цепи (VH), соответственно. Матрицей антитела человека для вариабельного участка легкой цепи IL17A-M069 было антитело IGKV4-1*01, имеющее гомологию 75,2% с легкой цепью IL17A-M069, а матрицей антитела человека для вариабельного участка было антитело IGHV1-69-2*01, имеющее гомологию 65,3% с тяжелой цепью IL17A-M069.
[00128] 3.3 Обратные мутации в каркасном участке гуманизированного вариабельного участка
[00129] Поскольку некоторые ключевые аминокислоты в каркасном участке мыши необходимы для поддержания активности CDR, ключевые аминокислоты были обратно мутированы в соответствующие последовательности аминокислот антитела мыши, следующие сайты были обратно мутированы: в соответствии с системой индексов Кабата в легкой цепи позиция 48 была обратно мутирована в V, позиция 49 была обратно мутирована в D, и позиция 87 была обратно мутирована в F; при этом, в тяжелой цепи позиция 24 была обратно мутирована в А, и позиция 43 была обратно мутирована в Н. Гуманизированное антитело IL17A-H069 получали путем гуманизированной трансплантации CDR и обратных мутаций в каркасном участке, и последовательности аминокислот его тяжелой и легкой цепей показаны в SEQ ID NO: 16/17, соответственно; последовательности аминокислот его тяжелой и легкой цепей в форме, содержащей сигнальные пептиды, показаны соответственно в SEQ ID NO: 18/19, где последовательности сигнальных пептидов, соединенных с тяжелой/легкой цепями приведены в SEQ ID NO: 20/21; последовательности вариабельных участков тяжелой цепи / легкой цепи гуманизированного антитела показаны в SEQ ID NO: 22/23; последовательности константного участка тяжелой цепи IgG1 / константного участка легкой цепи каппа человека гуманизированного антитела приведены в SEQ ID NO: 24/25), соответственно.
[00130] 3.4 Получение гуманизированного моноклонального антитела IL17A-Н069
[00131] Последовательность нуклеотидов (SEQ ID NO: 27), кодирующую легкую цепь и сигнальный пептид антитела IL17A-H069, которая содержит следующую последовательность нуклеотидов, кодирующую сигнальный пептид легкой цепи (SEQ ID NO: 29), вариабельный участок легкой цепи (SEQ ID NO: 31) гуманизированного антитела и константный участок легкой цепи каппа антитела человека (SEQ ID NO: 33), соединенные в этом порядке, амплифицировали с помощью ПЦР и вводили в самостоятельно разработанный вектор pGS (KpnI+XbaI) методом инфузии, и правильность последовательности плазмиды проверяли секвенированием. Аналогично, последовательность нуклеотидов (SEQ ID NO: 26), кодирующую тяжелую цепь антитела IL17A-H069, содержащую сигнальный пептид, которая содержит следующую последовательность нуклеотидов, кодирующую сигнальный пептид тяжелой цепи (SEQ ID NO: 28), вариабельный участок тяжелой цепи гуманизированного антитела (SEQ ID NO: 30) и константный участок тяжелой цепи IgG1 антитела человека (SEQ ID NO: 32), соединенные в этом порядке, амплифицировали с помощью ПЦР и вводили в вектор GS (NheI+NotI), который затем проверяли на содержание правильного гена легкой цепи методом инфузии, и правильность сборки векторов, экспрессирующих как легкую, так и тяжелую цепи IL17A-Н069, проверяли секвенированием. Эти векторы экспрессии являются эукариотическими векторами экспрессии, содержащими ген GS в качестве маркера селекции, а также легкую и тяжелую цепи указанного антитела в качестве элементов экспрессии. Эти векторы экспрессии трансфицировали в дефицитные клетки CHO-K1-GS и линии клеток с высокой экспрессией IL17A-H069 получали скринигом MSX. Клоны с высокой экспрессией антитела отбирали с помощью ИФА, и эти линии клеток с высокой экспрессией выбирали, принимая во внимание как статус роста клеток, так и ключевые характеристики качества антитела в качестве медикамента. Бессывороточные суспензионные культуры использовали для выращивания линии клеток СНО, продуцирующей IL17A-H069, в результате чего получали антитела IL17A-H069 высокой чистоты и качества.
[00132] Пример 4: Анализ антиген-связывающей аффинности гуманизированного антитела IL17A-H069
[00133] 4.1 Анализ аффинности связывания гуманизированного антитела с белком IL17A
[00134] 4.1.1 Связывание IL17A-H069 с рекомбинантным белком IL17A человека
[00135] Рекомбинантный белок IL17A человека (SinoBiological, Inc.) в различных концентрациях наносили в качестве покрытия на 96-луночный планшет в течение ночи при 4°С в количестве 100 мкл на лунку. Планшет промывали на следующий день и блокировали при комнатной температуре в течение 1 ч. После инкубации с 100 мкл 2 мкг/мл биоаналога COSENTYX (SinoCelltech Co., Ltd.) и IL17A-H069 (SinoCelltech Co., Ltd.) соответственно, планшет промывали для удаления несвязавшегося антитела, затем инкубировали с Fc/HRP козы против IgG человека и промывали несколько раз, а затем добавляли раствор хромогенного субстрата до появления окраски. OD450 измеряли после завершения образования окраски. Используя концентрацию рекомбинантного белка IL17A человека в качестве горизонтальной координаты и значение OD450 в качестве вертикальной координаты, использовали программное обеспечение graphPad Prism 6.0 для анализа данных и строили кривую доза-эффективность, из которой рассчитывали значение средней эффективной концентрации ЕС50.
[00136] Результаты, показанные на Фиг. 2, демонстрируют, что значение ЕС50 связывания биоаналога COSENTYX с рекомбинантным белком IL17A человека составляет 74,8 нг/мл, R2=0,9993; значение ЕС50 связывания IL17A-H069 с рекомбинантным белком IL17A человека составляет 46 нг/мл, R2=0,9958. Это указывает на то, способность IL17A-H069 связываться с рекомбинантным белком IL17A человека немного лучше, чем у биоаналога COSENTYX.
[00137] 4.1.2 Связывание IL17A-H069 с рекомбинантным белком IL17A/IL17F человека
[00138] Рекомбинантный димерный белок IL17A/F человека (SinoBiological, Inc., CT047-HNAE) при различных концентрациях наносили в качестве покрытия на 96-луночный планшет в течение ночи при 4°С в количестве 100 мкл на лунку. Планшет промывали на следующий день и блокировали при комнатной температуре в течение 1 ч. После инкубации с 100 мкл 2 мкг/мл биоаналога COSENTYX (SinoCelltech Co., Ltd.) и IL17A-H069 (SinoCelltech Co., Ltd.) соответственно, планшет промывали для удаления несвязавшегося антитела, затем инкубировали с Fc/HRP козы против IgG человека и промывали несколько раз, а затем добавляли раствор хромогенного субстрата до появления окраски. OD450 измеряли после завершения образования окраски. Используя концентрацию рекомбинантного белка IL17A/F человека в качестве горизонтальной координаты и значение OD450 в качестве вертикальной координаты, использовали программное обеспечение graphPad Prism 6.0 для анализа данных и строили кривую доза-эффективность, из которой рассчитывали значение средней эффективной концентрации ЕС50.
[00139] Результаты, показанные на Фиг. 3, демонстрируют, что значение ЕС50 связывания биоаналога COSENTYX с рекомбинантным белком IL17A/F человека составляет 63,9 нг/мл, R2=0,9999; значение ЕС50 связывания IL17A-H069 с рекомбинантным белком IL17A/F человека составляет 36,3 нг/мл, R2=1,0. Это указывает на то, что способность IL17A-H069 связываться с рекомбинантным димерным белком IL17A/F человека немного лучше, чем у биоаналога COSENTYX.
[00140] 4.1.3 Тест на аффинность связывания IL17A-H069 с рекомбинантным белком IL17A человека и рекомбинантным белком IL17A/TL17F человека
[00141] Аффинности IL17A-H069 при различных концентрациях (0,42 нМ, 0,90 нМ, 1,74 нМ и 3,47 нМ) и положительного контроля COSENTYX (Norvatis, SHM12) при различных концентрациях (0,90 нМ, 1,74 нМ, 3,47 нМ, 6,94 нМ и 13,9 нМ) с биотинилированными белками IL7A или IL17A/F определяли соответственно с использованием системы Octet Biomolecular Interaction Assay. Результаты в Таблице 2 показывают, что значение аффинности связывания KD IL17A-Н069 с рекомбинантным белком IL17A человека составляло 2,88Е-11 М, константа скорости ассоциации коп составляла 6,71Е+05 M-1с-1 и константа скорости диссоциации koff составляла 1,93Е-05 м-1; в то время как значение аффинности связывания KD COSENTYX с белком IL17A составляло 9,55Е-11 М, константа скорости ассоциации коп составляла 1,78Е+05 М-1с-1 и константа скорости диссоциации koff составляла 1,70Е-05 с-1. Значение аффинности связывания KD IL17A-H069 с рекомбинантным белком IL17A/F человека составляло 5,37Е-10 М, константа скорости ассоциации kon составляла 1,44E+0,5M-1с-1 иконстанта скорости диссоциации koff составляла 7,72Е-05 с-1; в то время как значение аффинности связывания KD COSENTYX с белком IL17A/F составляло 1,10Е-09 М, константа скорости ассоциации коп составляла 8,00Е+04 M-1с-1 и константа скорости диссоциации koff составляла 8,79Е-05 с-1. Результаты демонстрируют, что IL17A-H069 связывается с белком IL17A с большей аффинностью, чем COSENTYX, аффинность IL17A-H069 примерно в 3,32 раза выше, чем у COSENTYX, и IL17A-H069 обладает более высокой скоростью ассоциации, так что IL17A-H069 имеет более сильную связывающую активность с белком IL17A, чем COSENTYX; IL17A-H069 связывается с белком IL17A/F с более сильной аффинностью, чем COSENTYX, аффинность IL17A-H069 примерно 2,05 раза выше аффинности COSENTYX, и IL17A-H069 обладает более высокой скоростью ассоциации, так что IL17A-H069 имеет более сильную связывающую активность с белком IL17A/F, чем COSENTYX.
[00142] 4.1.4 Определение видоспецифичной кросс-реактивности IL17A-H069 с белком IL17A мыши
[00143] Рекомбинантный белок EL17A человека (Sino Biological, Inc.) и белок mIL17A мыши (Sino Biological, Inc.) при различных концентрациях соответственно наносили на 96-луночный планшет в количестве 100 мкл на лунку в течение ночи при 4°С. Планшет промывали на следующий день, блокировали при комнатной температуре в течение 1 ч. Добавляли 100 мкл 2 мкг/мл IL17A-H069 (Sino Biological, Inc.), COSENTYX (Norvatis, SUM 12) в качестве положительного контроля и антитело H7N9-R1 (SinoCelltech Co., Ltd.) в качестве отрицательного контроля соответственно и инкубировали. Планшет промывали для удаления несвязавшегося антитела. Планшет инкубировали с Fc/HRP козы против IgG человека (Sino Biological, Inc.) и затем несколько раз промывали, и добавляли раствор хромогенного субстрата до появления окраски. После завершения образования окраски измеряли OD450. Результаты, показанные на Фиг. 4 демонстрируют, что IL17A-H069 не имело перекрестного связывания с белком mIL17A мыши.
[00144] 4.2 IL17A-H069 блокирует связывание белка IL17A и белка IL17A/F с рецептором IL17RA
[00145] 4.2.1 IL17A-H069 блокирует связывание белка IL17A с рецептором IL17RA
[00146] Белок IL17A при концентрации 0,4 мкг/мл наносили на 96-луночный планшет в количестве 100 мкл на лунку в течение ночи при 4°С. Планшет промывали на следующий день, блокировали при комнатной температуре в течение 1 ч. Добавляли 100 мкл 2 мкг/мл биотинилированного белка IL17RA-His-биотин (Sino Biological, Inc.) в каждую лунку, затем добавляли различные концентрации IL17A-H069 (SinoCelltech Co., Ltd.), COSENTYX (Norvatis, SHM12) в качестве положительного контроля, Taltz (Eli Lilly) в качестве положительного контроля и антитело H7N9-R1 (SinoCelltech Co., Ltd.) в качестве отрицательного контроля, соответственно, и инкубировали. Планшет промывали для удаления несвязавшегося антитела. Планшет инкубировали со стрептавидин/HRP (Beijing ZSGB-Bio Co., Ltd., SA-5004), затем несколько раз промывали, и добавляли раствор хромогенного субстрата до появления окраски. После завершения образования окраски измеряли OD450 в каждой исследованной группе в двух повторах.
[00147] Принимали концентрацию антитела в качестве горизонтальной координаты и степень ингибирования % в качестве вертикальной координаты, программное обеспечение graphPad Prism 6.0 использовали для анализа данных и построения графика, рассчитывали значения IC50. Степень ингибирования % = (OD контроль - OD образец) / OD контроль × 100%, где OD контроль обозначает значение OD лунки только с добавленным белком IL17RA-His-биотин, но без добавки антитела, OD образец обозначает значение OD лунки как с добавленным белком IL17RA-His-биотин, так и с добавленным антителом.
[00148] Результаты, показанные на Фиг. 5 демонстрируют, что биотинилированный белок IL17RA может эффективно связываться с нанесенным на планшет рекомбинантный белком IL17A человека, и антитело IL17A-H069 может ингибироватъ связывание белка IL17А с рецептором IL17RA со значительно более лучшим профилем, представленным его кривой ингибирования, чем COSENTYX, использованного в качестве положительного контроля, но при этом профиль IL17A-H069 по ингибированию связывания белка IL17A с рецептором IL17RA был близок к профилю Taltz, использованного в качестве положительного контроля. Значения IC50 IL17A-H069, COSENTYX и Taltz были 0,50 мкг/мл, 2,99 мкг/мл, и 0,50 мкг/мл соответственно, а максимальная степень ингибирования составляла 85,4%, 73,5% и 89,5%, соответственно.
[00149] 4.2.2 IL17A-H069 блокирует связывание белка IL17A/F с рецептором IL17RA
[00150] Белок IL17RA-Fc (Sino Biological, Inc.) при концентрации 5 мкг/мл наносили на 96-луночный планшет в количестве 100 мкл на лунку в течение ночи при 4°С. Планшет промывали на следующий день, блокировали при комнатной температуре в течение 1 ч. Добавляли 100 мкл 0,8 мкг/мл белка IL17A/F-биотин (Sino Biological, Inc.) в каждую лунку, затем добавляли различные концентрации IL17A-Н069 (SinoCelltech Co., Ltd.), биоаналог COSENTYX (SinoCelltech Co., Ltd.) в качестве положительного контроля, биоаналог Taltz (SinoCelltech Co., Ltd.) в качестве положительного контроля и антитело H7N9-R1 (SinoCelltech Co., Ltd.) в качестве отрицательного контроля, соответственно, и инкубировали. Планшет промывали для удаления несвязавшегося антитела. Планшет инкубировали со стрептавидин/HRP (Beijing ZSGB-Bio Co., Ltd., SA-5004) и затем несколько раз промывали, и добавляли раствор хромогенного субстрата до появления окраски. OD450 измеряли после стабилизации появления окраски.
[00151] Принимали концентрацию антитела в качестве горизонтальной координаты и степень ингибирования % в качестве вертикальной координаты, программное обеспечение graphPad Prism 6.0 использовали для анализа данных и построения кривой, рассчитывали значения IC50. Степень ингибирования % = (OD контроль - OD образец) / OD контроль × 100%, где OD контроль обозначает значение OD лунки только с добавленным белком IL17A/F-биотин без добавления антитела, OD образец обозначает значение OD лунки как с добавленным белком IL17A/F-биотин, так и добавленным антителом.
[00152] Результаты, показанные на Фиг. 6 демонстрируют, что белок IL17A/F-биотин может эффективно связываться с покрытием рекомбинантного белка IL17RA-Fc человека, а добавление антитела IL17A-H069 может эффективно ингибировать связывание белка IL17A/F с рецептором IL17RA-Fc. IL17A-H069 обладает более сильным эффектом ингибирования связывания белка IL17A/F с рецептором IL17RA-Fc, чем биоаналог COSENTYX, использованный в качестве положительного контроля, и биоаналог Taltz, использованный в качестве положительного контроля. Значения IC50 IL17A-H069, биоаналога COSENTYX и биоаналога Taltz были 1,02 мкг/мл, 1,20 мкг/мл и 1,35 мкг/мл, соответственно, а максимальные степени ингибирования были 92,3%, 87,9% и 75%, соответственно.
[00153] Пример 5: Функциональный анализ гуманизированного антитела IL17A-Н069
[00154] 5.1 IL17A-H069 блокирует индуцированную IL17A или IL17A/F секрецию IL-6 клетками HFF
[00155] Клетки HFF инокулировали в 96-луночный планшет при плотности клеток 1×104 на лунку и выращивали в течение ночи в среде ДМЭМ, содержащей 15% ФБС.Различные концентрации IL17A-H069 (SinoCelltech Co., Ltd.) и COSENTYX (Norvatis) в качестве положительного контроля или Taltz (Eli Lilly) в качестве положительного контроля добавляли соответственно в количестве 10 мкл на лунку на следующий день. Затем в каждую лунку добавляли 10 мкл белка IL17A (Sino Biological, Inc. 12047-HNAS) в конечной концентрации 50 нг/мл или белка IL17A/F (Sino Biological, Inc. CT047-HNAE) в конечной концентрации 1 мкг/мл, соответственно. 96-луночный планшет инкубировали при 37°С, 5% СО2 в инкубаторе клеток в течение 48 ч, и использовали контрольную лунку В (без клеток), отрицательный контроль М (с добавлением клеток, без добавления образца антитела, с добавлением IL17A или IL17A/F) и М' (с добавлением клеток, без добавления образца антитела и без добавления EL17A или IL17A/F). После инкубации, супернатант собирали и измеряли секрецию IL-6 с помощью ИФА. Из значения секреции IL-6 в лунках с образцом и лунках группы М вычитали значение секрецию IL-6 в лунках группы М' соответственно для расчета степени ингибирования. Степень ингибирования % = (1 - секреция IL-6 в лунках с образцом) / секреции IL-6 в лунках группы М × 100%. Стандартную кривую рассчитывали с использованием статистического программного обеспечения, принимая концентрацию образца антитела в качестве горизонтальной координаты и секрецию IL-6 в качестве вертикальной координаты, и использовали уравнение логистической регрессии с 4 параметрами для построения стандартной кривой "S" для расчета половины максимальной эффективной концентрации (ЕС50) образца указанного антитела.
[00156] В описанных выше измерениях, как показано на Фиг. 7 и в Таблица 3, активность IL17A-Н069 в нейтрализации IL17A (EC50: 0,19 мкг/мл, максимальная степень нейтрализации: 94,6%) была намного выше, чем у COSENTYX, использованного в качестве положительного контроля (ЕС50: 0,22 мкг/мл, максимальная степень нейтрализации: 51,6%) (Фиг. 7А); в сравнении с Taltz, использованным в качестве положительного контроля, активность IL17A-H069 по нейтрализации IL17A (ЕС50: 0,20 мкг/мл, максимальная степень нейтрализации: 90,3%) была близкой к нейтрализующей активности Taltz (ЕС50: 0,19 мкг/мл, максимальная степень нейтрализации: 95,9%) (Фиг. 7С); активность IL17A-H069 в нейтрализации IL17A/F (ЕС50: 1,19 мкг/мл, максимальная степень нейтрализации: 85,0%) также была слегка выше, чем у COSENTYX (ЕС50: 2,25 мкг/мл, максимальная степень нейтрализации: 79,5%) (Фиг. 7В). В сравнении Taltz, использованным в качестве положительного контроля, активность IL17A-H069 по нейтрализации IL17A/F при высокой концентрации (ЕС50: 0,83 мкг/мл, максимальная степень нейтрализации: 72,90%) была близка к Taltz (ЕС50: 1,10 мкг/мл, максимальная степень нейтрализации: 76,3%), и превосходной по отношению к Taltz, использованному в качестве положительного контроля, при низких концентрациях (Фиг. 7D). В итоге, IL17A-H069 имеет лучшую биологическую активность по нейтрализации IL17A и IL17A/F.
[00157] Пример 6: Эффективность гуманизированного антитела in vivo в мышах
[00158] 6.1 Эффективность IL17A-H069 in vivo в модели псориаза (PsO) с иммунно восстановленными hPBMC мышей
[00159] Используя hPBMC из 3 доноров, получали в общей сложности 60 B-NDG мышей (Biocytogen Pharmaceuticals (Beijing) Co., Ltd.) с гуманизированной иммунной системой (20 мышей на одного донора hPBMC). Периферическую кровь собирали через одну неделю и измеряли процент клеток, полученных от человека, с помощью клеточной цитометрии. 20 мышей имели процент клеток, полученных от человека, в диапазоне 0,04-1,5%, 31 мышь имела процент в диапазоне 1,5-7%, и 8 мышей имели процент >7%. Мышей с процентом 1,5-7% выбирали для создания IMQ (имихимод)-индуцированной модели псориаза на мышах, на основании которой оценивали эффективность IL17A-H069.
[00160] Выбранные мыши были сгруппированы в соответствии с данной стратегией: 5 немоделированных мышей были сгруппированы в нормальную контрольную группу G1; 5 мышей с моделированным псориазом, которым не вводили препарат, были объединены в группу G2, т.е., модельную группу псориаза; 7 мышей с моделированным псориазом, которым вводили IL17A-H069, были объединены в группу G3; и 7 мышей с моделированным псориазом, которым вводили положительный контроль Taltz, были объединены в группу G4. Спины всех мышей выбривали, чтобы образовалась открытая область размером примерно 2 см × 3 см, и на спину каждой мыши в группах G2, G3 и G4 наносили 100 мг крема IMQ, а на уши наносили 10 мг крема IMQ каждой мыши в группах G2, G3 и G4 ежедневно в течение 10 дней. Ежедневно регистрировали показатели PASI мышей. В соответствии с критерием оценки PASI, как показано в Таблице 4, мышам присваивали 0-4 балла за степень эритемы, шелушение и утолщение кожи на спине на задней стороне очага поражения, соответственно, и суммировали три балла, в результате чего оценивали общее количество баллов. Для групп G3 и G4 введение антитела начинали в 1-й день применения крема IMQ в дозе 10 мкг/кг еженедельно.
[00161] Результаты показаны на Фиг. 8. По сравнению с мышами в нормальной контрольной группе показатель PASI был значительно выше у мышей в группе с моделью псориаза, отражая, что эта модель мыши может в некоторой степени характеризовать псориаз; с 5-го дня показатели PASI были значительно ниже в группе, получавшей IL17A-H069, по сравнению с группой модели псориаза; а по сравнению с контрольной группой Taltz, IL17A-H069 продемонстрировал более высокую эффективность in vivo в снижении показателей псориаза у мышей, таким образом, IL17A-H069 эффективен в облегчении дебюта псориаза (PsO) и уменьшении симптомов псориаза у мышей с моделью псориаза.
[00162] Пример 7: Фармакокинетика гуманизированного антитела in vivo
[00163] 7.1 Фармакокинетика однократного подкожного введения IL17A-H069 у яванских макаков
[00164] В этом примере однократные подкожные инъекции антитела IL17A-H069 вводили яванским макакам в дозировке 1 мг/кг. Сыворотку собирали до введения и через 1 ч, 2 ч, 4 ч, 6 ч, 8 ч, 24 ч, 48 ч, 3 дня, 4 дня, 7 дней, 10 дней, 14 дней, 17 дней, 21 день, 24 дня, 28 дней, 31 день и 35 дней после введения, соответственно. Разработанный способ ИФА использовали для измерения концентрации лекарственного средства IL17A-H069 в сыворотке мышей, фармакокинетические параметры рассчитывали с использованием программного обеспечения некомпартментального анализа (NCA)Phoenix-WinNonlin (Pharsight) 6.4. Исследовали динамическую картину изменения профиля препарата in vivo после однократного подкожного введения IL17A-H069 для изучения динамических изменений препарата в организме после однократного подкожного введения IL17A-Н069.
[00165] Изменения концентрации лекарственного средства IL17A-H069 с течением времени показана в Таблице 6 и на Фиг. 9. Не было существенной тендерной разницы в Cmax и AUClast между самками и самцами мышей (данные не показаны), а время полувыведения t1/2 IL17A-H069 было 353,66 ч, значение Tmax составляло 34 ч. В терминах экспозиции in vivo, AUClast IL17A-H069 составляла 3846,86 ч*мкг/мл.
[00166] При дозировке 1 мг/кг, IL17A-H069 более короткое Tmax и более длинное t1/2, так что IL17A-H06 демонстрирует превосходную фармакокинетику, включая быструю абсорбцию после подкожной инъекции, большое время полувыведения и лучшую экспозицию лекарственного средства и т.д., тем самым закладывая основу для более длительного цикла дозирования.

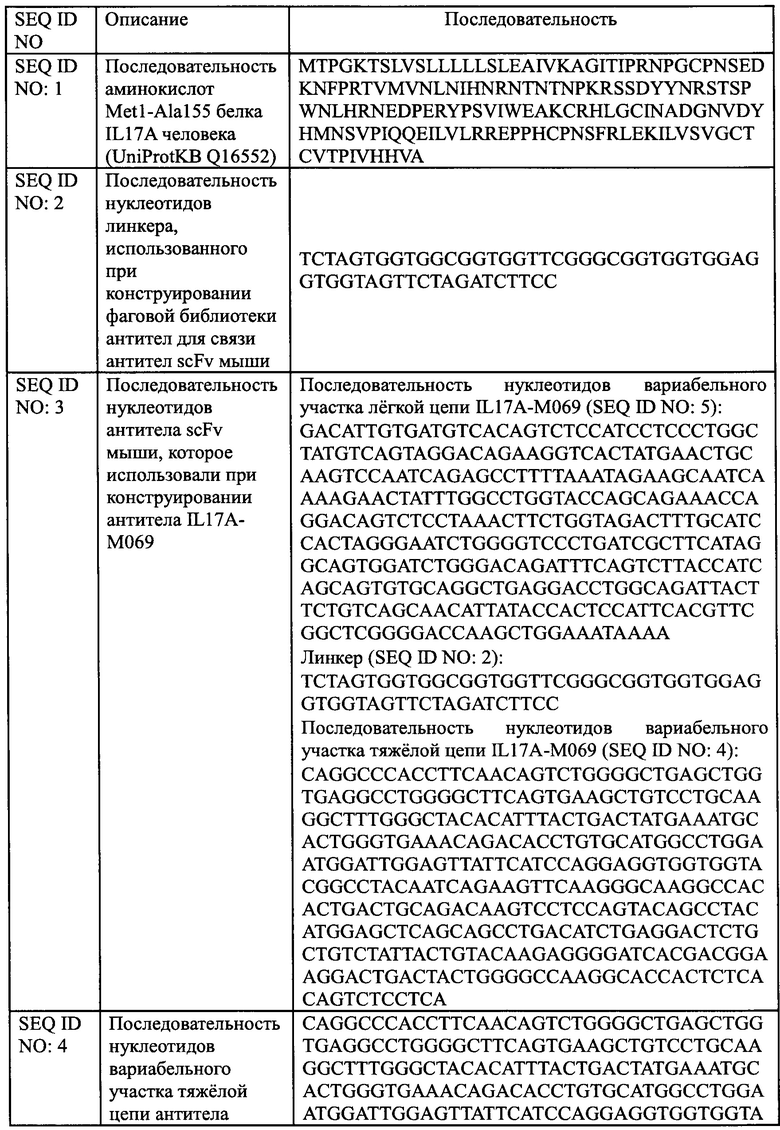

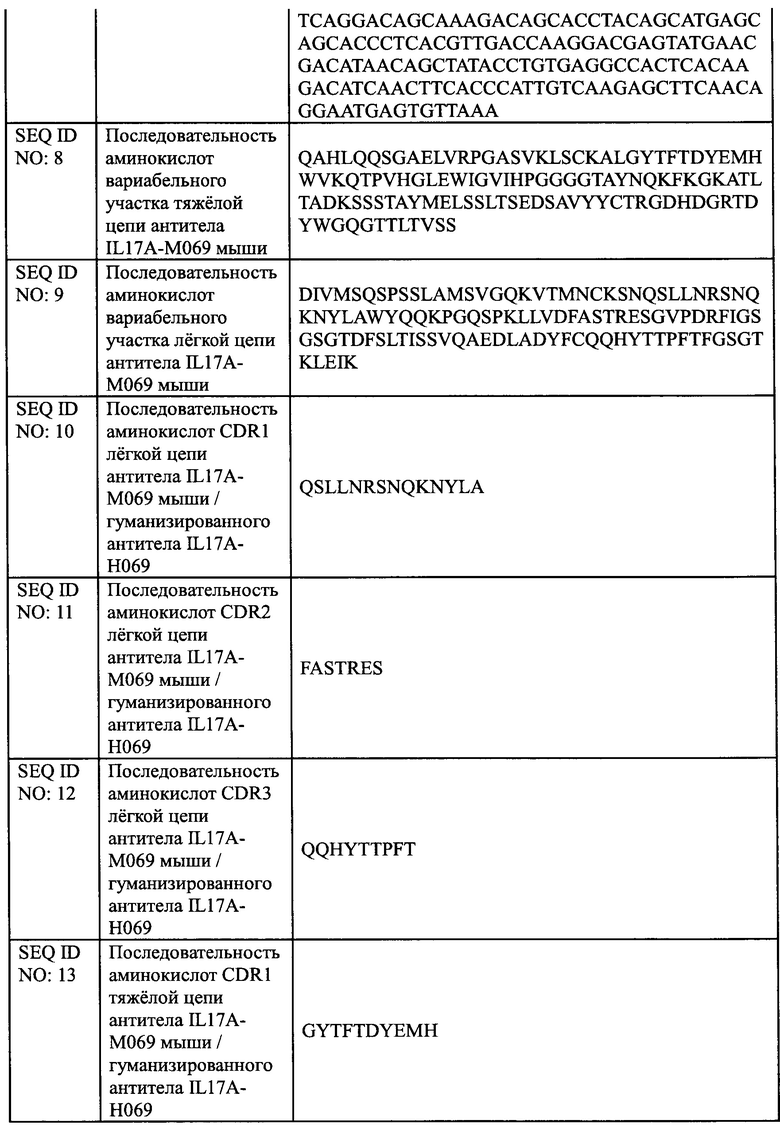
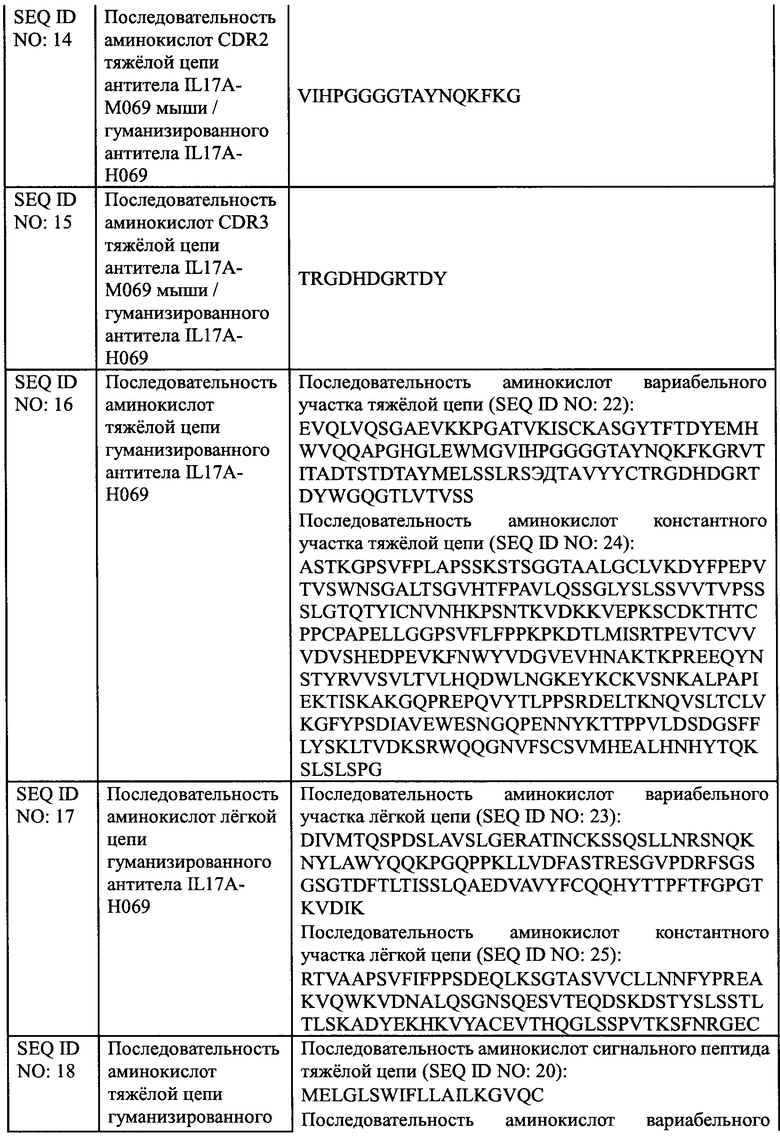
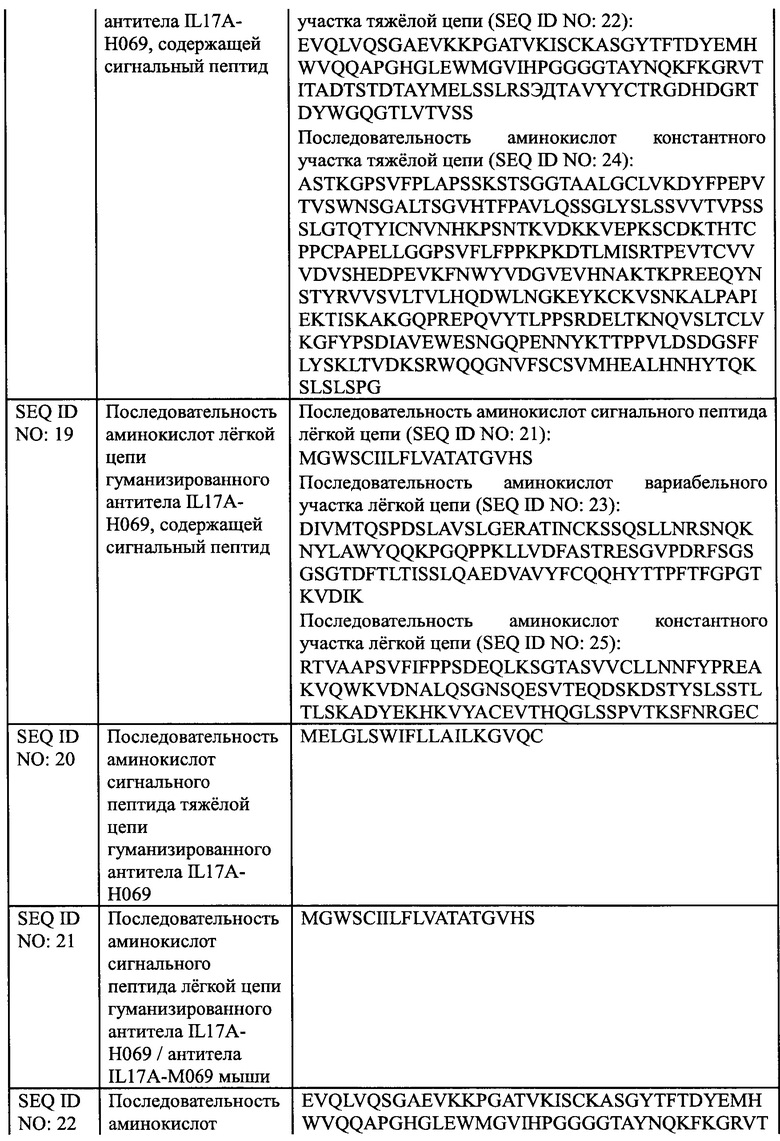
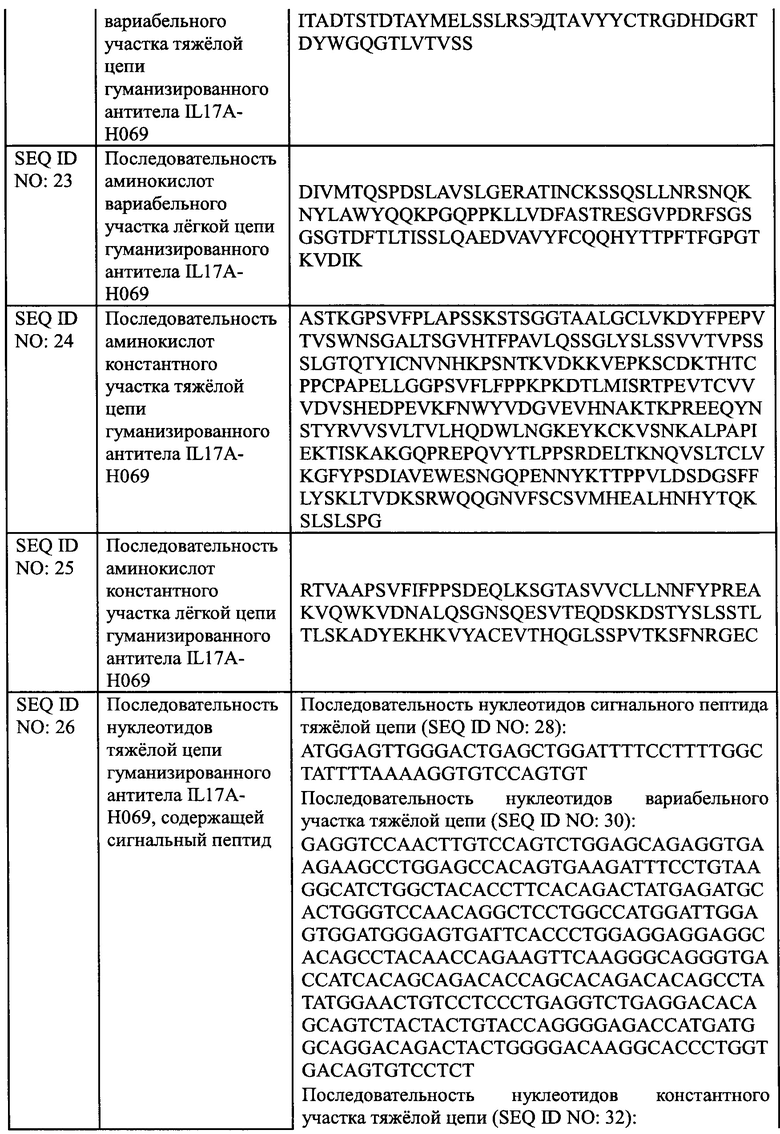
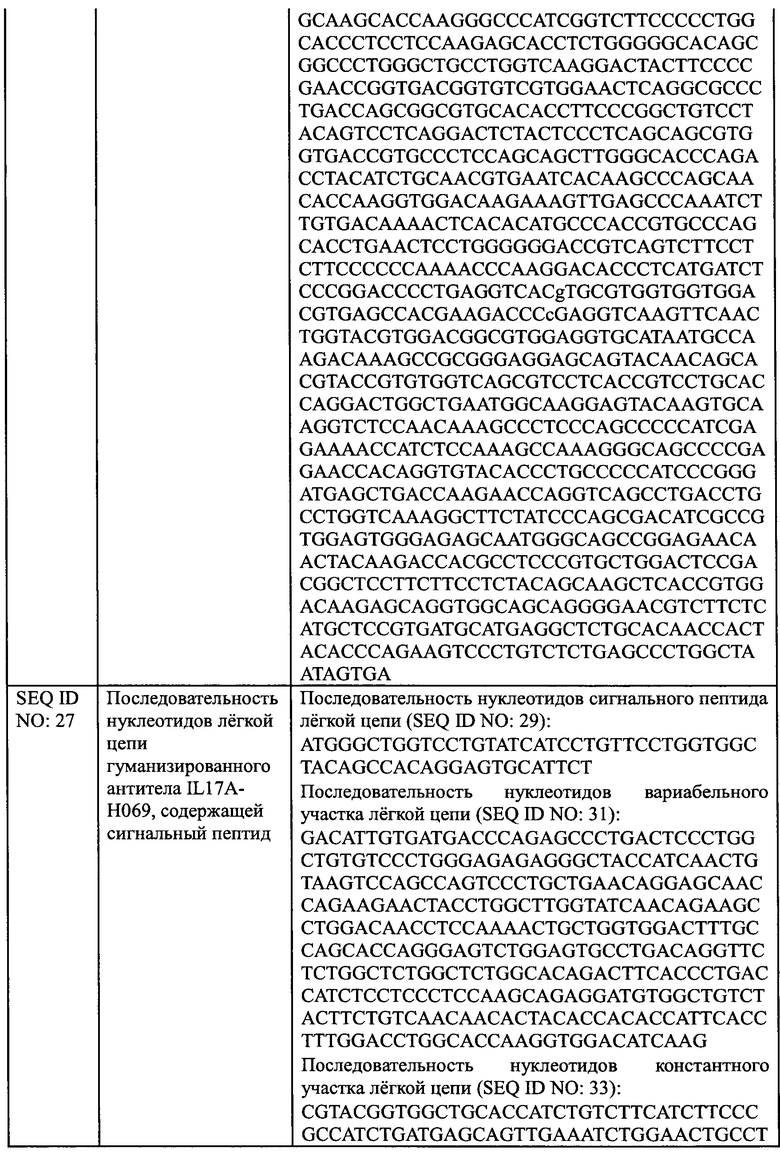
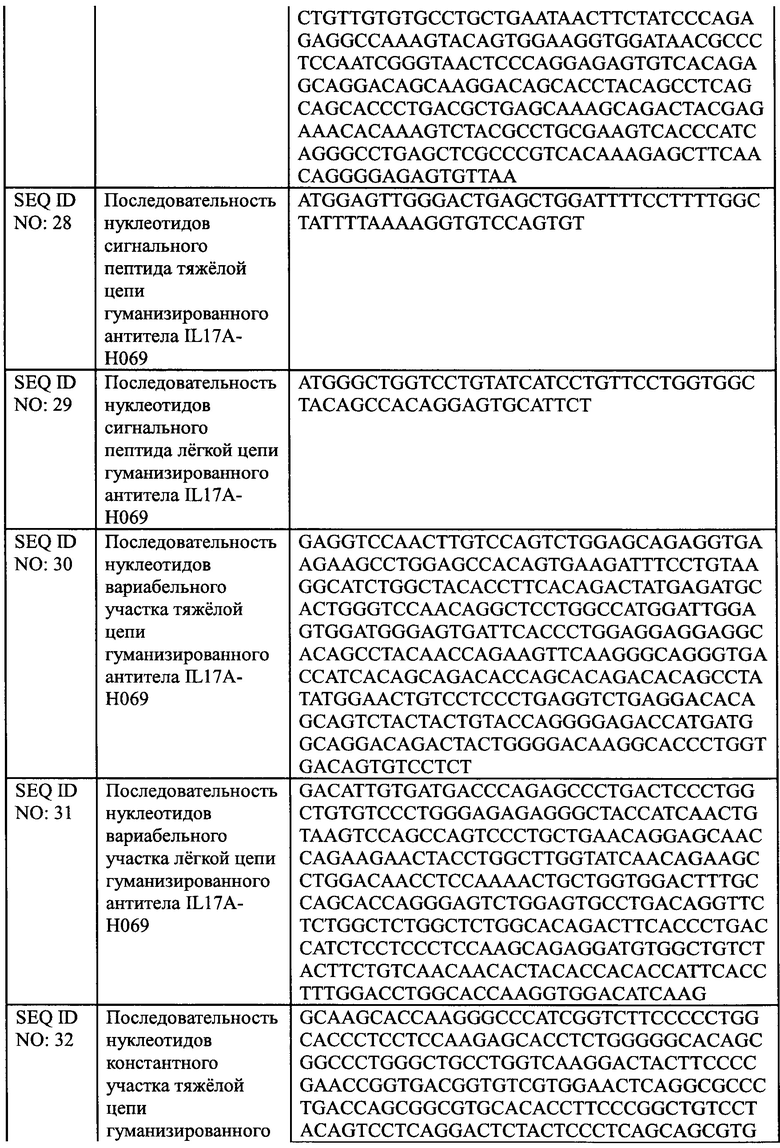
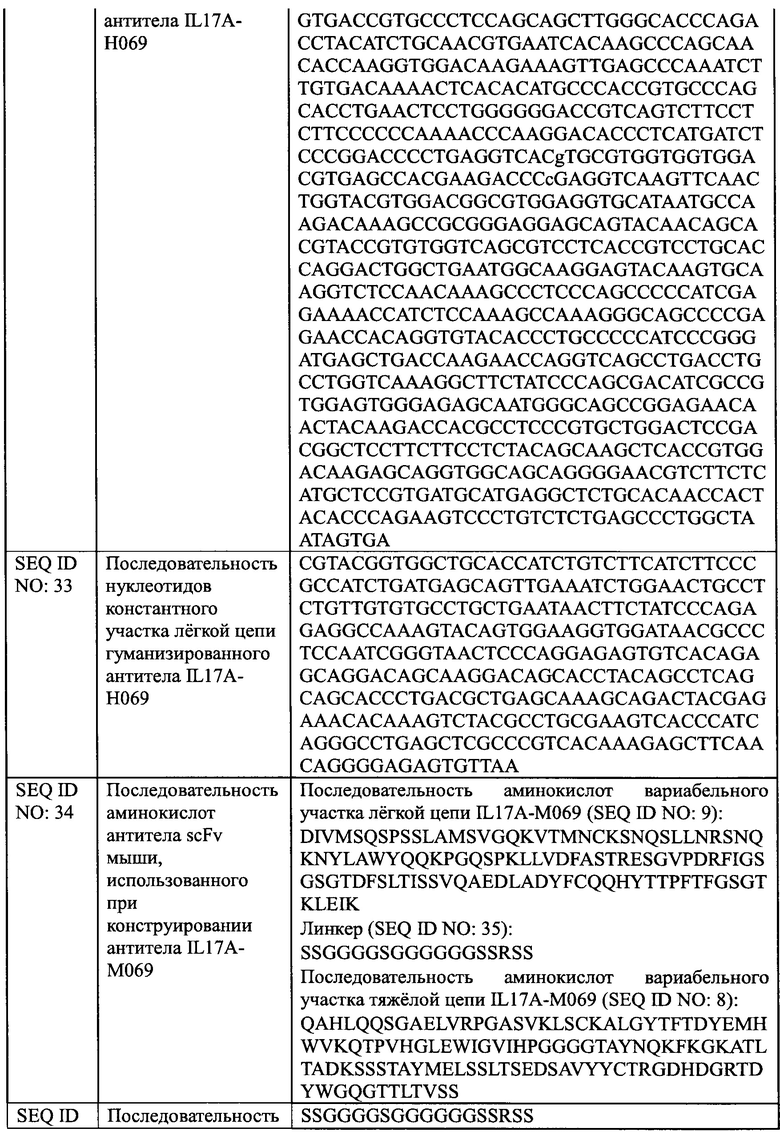
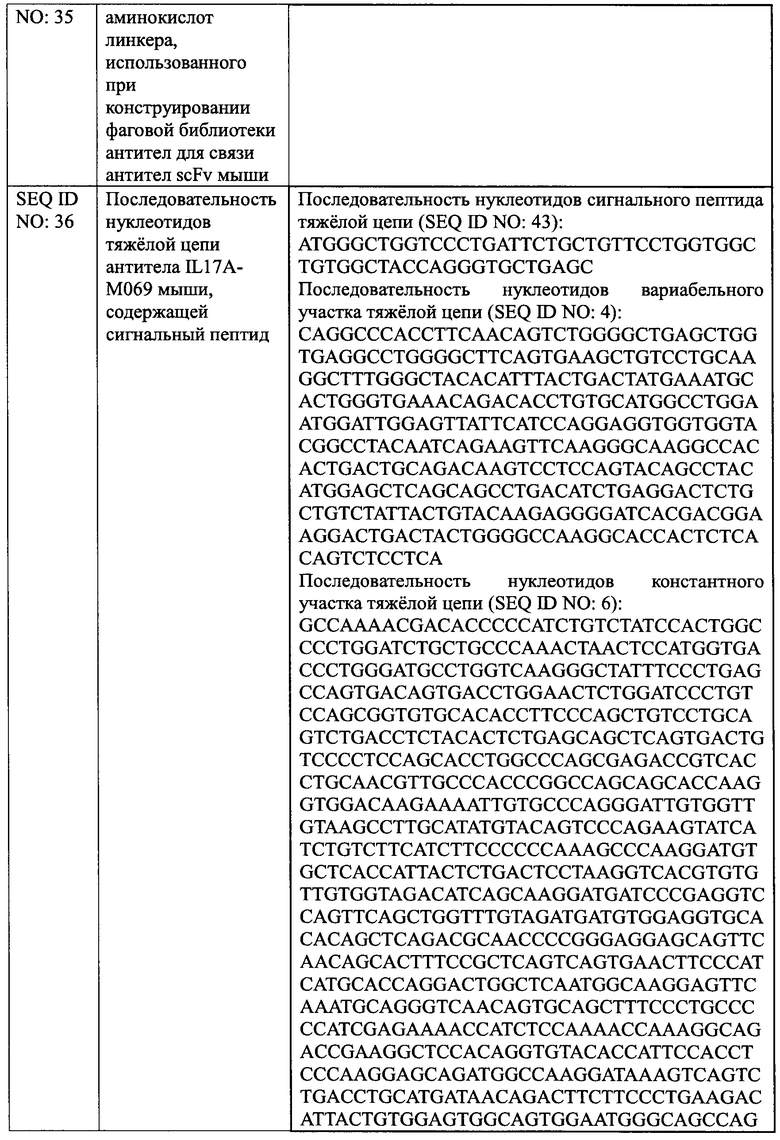
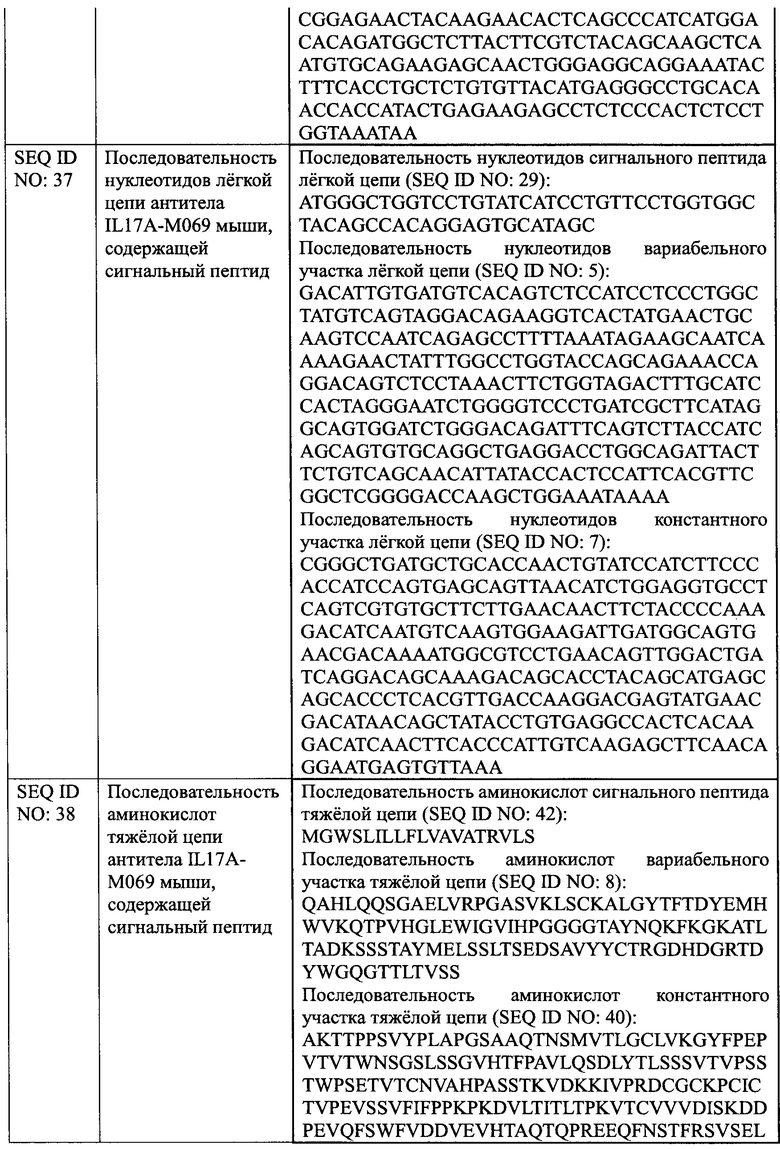
Цитированная литература
Abhinandan, K.R. и А.С. Martin (2008). "Analysis and improvements to Kabat and structurally correct numbering of antibody variable domains." Mol Immunol 45(14): 3832-3839.
Aitken, P. M. О. B. (2002). Antibody Phage Display Methods and Protocols, Humana Press.
Beerli, R.R., M. Bauer, A. Fritzer, L.B. Rosen, R.B. Buser, M. Hanner, M. Maudrich, M. Nebenfuehr, J. A. S. Toepfer, S. Mangold, A. Bauer, S.M. Holland, S.K. Browne и A. Meinke (2014). "Mining the human autoantibody repertoire: isolation of potent IL17A-neutralizing monoclonal antibody from a patient with thymoma." mAbs 6(6): 1608-1620.
Brembilla, N.C., L. Senra и W.-H. Boehncke (2018). "The IL-17 family of cytokines in psoriasis: IL-17A and beyond." Frontiers in immunology 9.
Bryksin, A.V. и I. Matsumura (2010). "Overlap extension PCR cloning: a simple and reliable way to create recombinant plasmids." BioTechniques 48(6): 463-465.
de StGroth, S.F. и D. Scheidegger (1980). "Production of monoclonal antibody: strategy and tactics." J Immunol Methods 35(1-2): 1-21.
Dondelinger, M., P. Filee, E. Sauvage, B. Quinting, S. Muyldermans, M. Galleni и M.S. Vandevenne (2018). "Understanding the Significance and Implications of Antibody Numbering and Antigen-Binding Surface/Residue Definitions." Frontiers in immunology 9: 2278-2278.
Dubin, P.J. и J.K. Kolls (2009). "Interleukin-17A and Interleukin-17F: A Tale of Two Cytokines." Immunity 30(1): 9-11.
Fala, L. (2016). "Cosentyx (Secukinumab): First IL-17A Antagonist Receives FDA Approval for Moderate-to-Severe Plaque psoriasis." American health & drug benefits 9(Spec Feature): 60-63.
Gu, C., L. Wu и X. Li (2013). "IL-17 family: cytokines, receptors and signaling." Cytokine 64(2): 477-485.
Jones, S.Т. и M.M. Bendig (1991). "Rapid PCR-cloning of full-length mouse immunoglobulin variable regions." Biotechnology (N Y) 9(6): 579.
Kettleborough, C.A., J. Saldanha, V.J. Heath, C.J. Morrison и M.M. Bendig (1991). "Humanization of a mouse monoclonal antibody by CDR-grafting: the importance of framework residues on loop conformation." Protein Eng 4(7): 773-783.
Lefranc, M.-P. (2014). "Immunoglobulin and T Cell Receptor Genes: IMGT(®) and the Birth and Rise of Immunoinformatics." Frontiers in immunology 5: 22-22.
Liu, L., J. Lu, B.W. Allan, Y. Tang, J. Tetreault, C.-K. Chow, B. Barmettler, J. Nelson, H. Bina, L. Huang, V.J. Wroblewski и K. Kikly (2016). "Generation and characterization of Ixekizumab, a humanized monoclonal antibody that neutralizes interleukin-17A." Journal of inflammation research 9: 39-50.
Marinoni, В., A. Ceribelli, M.S. Massarotti и С.Selmi (2014). "The Тh17 axis in psoriatic disease: pathogenetic and therapeutic implications." Auto-immunity highlights 5(1): 9-19.
Mitra, A., S. Raychaudhuri и S.P. Raychaudhuri (2014). "IL-17 and IL-17R: an auspicious therapeutic target for psoriatic disease." Actas dermo-sifiliograficas 105: 21-33.
Saldanha, J.W., A.C. Martin и О.J. Leger (1999). "A single back mutation in the human kIV framework of a previously unsuccessfully humanized antibody restores binding activity and increases the secretion in cos cells." Mol Immunol 36(11-12): 709-719.
Wang, E.A., E. Suzuki, E. Maverakis и I.E. Adamopoulos (2017). "Targeting IL-17 in psoriatic arthritis." European journal of rheumatology 4(4): 272-277.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SinoCellTech Ltd.
<120> ГУМАНИЗИРОВАННОЕ АНТИТЕЛО ПРОТИВ IL17A И ЕГО ПРИМЕНЕНИЕ
<130> CIE200074PCT
<160> 43
<170> BiSSAP 1.3.6
<210> 1
<211> 155
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот Met1-Ala155 белка IL17A человека
(UniProtKB Q16552)
<400> 1
Met Thr Pro Gly Lys Thr Ser Leu Val Ser Leu Leu Leu Leu Leu Ser
1 5 10 15
Leu Glu Ala Ile Val Lys Ala Gly Ile Thr Ile Pro Arg Asn Pro Gly
20 25 30
Cys Pro Asn Ser Glu Asp Lys Asn Phe Pro Arg Thr Val Met Val Asn
35 40 45
Leu Asn Ile His Asn Arg Asn Thr Asn Thr Asn Pro Lys Arg Ser Ser
50 55 60
Asp Tyr Tyr Asn Arg Ser Thr Ser Pro Trp Asn Leu His Arg Asn Glu
65 70 75 80
Asp Pro Glu Arg Tyr Pro Ser Val Ile Trp Glu Ala Lys Cys Arg His
85 90 95
Leu Gly Cys Ile Asn Ala Asp Gly Asn Val Asp Tyr His Met Asn Ser
100 105 110
Val Pro Ile Gln Gln Glu Ile Leu Val Leu Arg Arg Glu Pro Pro His
115 120 125
Cys Pro Asn Ser Phe Arg Leu Glu Lys Ile Leu Val Ser Val Gly Cys
130 135 140
Thr Cys Val Thr Pro Ile Val His His Val Ala
145 150 155
<210> 2
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов линкера, использованного при
конструировании фаговой библиотеки антител для связи с антителом scFv мыши
<400> 2
tctagtggtg gcggtggttc gggcggtggt ggaggtggta gttctagatc ttcc 54
<210> 3
<211> 747
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов антитела scFv мыши, использованного при
конструировании антитела IL17A-M069
<400> 3
gacattgtga tgtcacagtc tccatcctcc ctggctatgt cagtaggaca gaaggtcact 60
atgaactgca agtccaatca gagcctttta aatagaagca atcaaaagaa ctatttggcc 120
tggtaccagc agaaaccagg acagtctcct aaacttctgg tagactttgc atccactagg 180
gaatctgggg tccctgatcg cttcataggc agtggatctg ggacagattt cagtcttacc 240
atcagcagtg tgcaggctga ggacctggca gattacttct gtcagcaaca ttataccact 300
ccattcacgt tcggctcggg gaccaagctg gaaataaaat ctagtggtgg cggtggttcg 360
ggcggtggtg gaggtggtag ttctagatct tcccaggccc accttcaaca gtctggggct 420
gagctggtga ggcctggggc ttcagtgaag ctgtcctgca aggctttggg ctacacattt 480
actgactatg aaatgcactg ggtgaaacag acacctgtgc atggcctgga atggattgga 540
gttattcatc caggaggtgg tggtacggcc tacaatcaga agttcaaggg caaggccaca 600
ctgactgcag acaagtcctc cagtacagcc tacatggagc tcagcagcct gacatctgag 660
gactctgctg tctattactg tacaagaggg gatcacgacg gaaggactga ctactggggc 720
caaggcacca ctctcacagt ctcctca 747
<210> 4
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка тяжёлой цепи
антитела IL17A-M069 мыши
<400> 4
caggcccacc ttcaacagtc tggggctgag ctggtgaggc ctggggcttc agtgaagctg 60
tcctgcaagg ctttgggcta cacatttact gactatgaaa tgcactgggt gaaacagaca 120
cctgtgcatg gcctggaatg gattggagtt attcatccag gaggtggtgg tacggcctac 180
aatcagaagt tcaagggcaa ggccacactg actgcagaca agtcctccag tacagcctac 240
atggagctca gcagcctgac atctgaggac tctgctgtct attactgtac aagaggggat 300
cacgacggaa ggactgacta ctggggccaa ggcaccactc tcacagtctc ctca 354
<210> 5
<211> 339
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка лёгкой цепи
антитела IL17A-M069 мыши
<400> 5
gacattgtga tgtcacagtc tccatcctcc ctggctatgt cagtaggaca gaaggtcact 60
atgaactgca agtccaatca gagcctttta aatagaagca atcaaaagaa ctatttggcc 120
tggtaccagc agaaaccagg acagtctcct aaacttctgg tagactttgc atccactagg 180
gaatctgggg tccctgatcg cttcataggc agtggatctg ggacagattt cagtcttacc 240
atcagcagtg tgcaggctga ggacctggca gattacttct gtcagcaaca ttataccact 300
ccattcacgt tcggctcggg gaccaagctg gaaataaaa 339
<210> 6
<211> 975
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка тяжёлой цепи IgG1
мыши
<400> 6
gccaaaacga cacccccatc tgtctatcca ctggcccctg gatctgctgc ccaaactaac 60
tccatggtga ccctgggatg cctggtcaag ggctatttcc ctgagccagt gacagtgacc 120
tggaactctg gatccctgtc cagcggtgtg cacaccttcc cagctgtcct gcagtctgac 180
ctctacactc tgagcagctc agtgactgtc ccctccagca cctggcccag cgagaccgtc 240
acctgcaacg ttgcccaccc ggccagcagc accaaggtgg acaagaaaat tgtgcccagg 300
gattgtggtt gtaagccttg catatgtaca gtcccagaag tatcatctgt cttcatcttc 360
cccccaaagc ccaaggatgt gctcaccatt actctgactc ctaaggtcac gtgtgttgtg 420
gtagacatca gcaaggatga tcccgaggtc cagttcagct ggtttgtaga tgatgtggag 480
gtgcacacag ctcagacgca accccgggag gagcagttca acagcacttt ccgctcagtc 540
agtgaacttc ccatcatgca ccaggactgg ctcaatggca aggagttcaa atgcagggtc 600
aacagtgcag ctttccctgc ccccatcgag aaaaccatct ccaaaaccaa aggcagaccg 660
aaggctccac aggtgtacac cattccacct cccaaggagc agatggccaa ggataaagtc 720
agtctgacct gcatgataac agacttcttc cctgaagaca ttactgtgga gtggcagtgg 780
aatgggcagc cagcggagaa ctacaagaac actcagccca tcatggacac agatggctct 840
tacttcgtct acagcaagct caatgtgcag aagagcaact gggaggcagg aaatactttc 900
acctgctctg tgttacatga gggcctgcac aaccaccata ctgagaagag cctctcccac 960
tctcctggta aataa 975
<210> 7
<211> 325
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка лёгкой цепи каппа
мыши
<400> 7
cgggctgatg ctgcaccaac tgtatccatc ttcccaccat ccagtgagca gttaacatct 60
ggaggtgcct cagtcgtgtg cttcttgaac aacttctacc ccaaagacat caatgtcaag 120
tggaagattg atggcagtga acgacaaaat ggcgtcctga acagttggac tgatcaggac 180
agcaaagaca gcacctacag catgagcagc accctcacgt tgaccaagga cgagtatgaa 240
cgacataaca gctatacctg tgaggccact cacaagacat caacttcacc cattgtcaag 300
agcttcaaca ggaatgagtg ttaaa 325
<210> 8
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка тяжёлой цепи
антитела IL17A-M069 мыши
<400> 8
Gln Ala His Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Leu Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile His Pro Gly Gly Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Gly Asp His Asp Gly Arg Thr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 9
<211> 113
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка лёгкой цепи
антитела IL17A-M069 мыши
<400> 9
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Met Ser Val Gly
1 5 10 15
Gln Lys Val Thr Met Asn Cys Lys Ser Asn Gln Ser Leu Leu Asn Arg
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Val Asp Phe Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln
85 90 95
His Tyr Thr Thr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 10
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR1 лёгкой цепи антитела IL17A-M069
мыши / гуманизированного антитела IL17A-H069
<400> 10
Gln Ser Leu Leu Asn Arg Ser Asn Gln Lys Asn Tyr Leu Ala
1 5 10
<210> 11
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR2 лёгкой цепи антитела IL17A-M069
мыши / гуманизированного антитела IL17A-H069
<400> 11
Phe Ala Ser Thr Arg Glu Ser
1 5
<210> 12
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR3 лёгкой цепи антитела IL17A-M069
мыши / гуманизированного антитела IL17A-H069
<400> 12
Gln Gln His Tyr Thr Thr Pro Phe Thr
1 5
<210> 13
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR1 тяжёлой цепи антитела IL17A-M069
мыши / гуманизированного антитела IL17A-H069
<400> 13
Gly Tyr Thr Phe Thr Asp Tyr Glu Met His
1 5 10
<210> 14
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR2 тяжёлой цепи антитела IL17A-M069
мыши / гуманизированного антитела IL17A-H069
<400> 14
Val Ile His Pro Gly Gly Gly Gly Thr Ala Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 15
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR3 тяжёлой цепи антитела IL17A-M069
мыши / гуманизированного антитела IL17A-H069
<400> 15
Thr Arg Gly Asp His Asp Gly Arg Thr Asp Tyr
1 5 10
<210> 16
<211> 447
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот тяжёлой цепи гуманизированного антитела
IL17A-H069
<400> 16
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Gln Gln Ala Pro Gly His Gly Leu Glu Trp Met
35 40 45
Gly Val Ile His Pro Gly Gly Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Gly Asp His Asp Gly Arg Thr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 17
<211> 220
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот лёгкой цепи гуманизированного антитела
IL17A-H069
<400> 17
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Arg
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Val Asp Phe Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
His Tyr Thr Thr Pro Phe Thr Phe Gly Pro Gly Thr Lys Val Asp Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 18
<211> 466
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот тяжёлой цепи гуманизированного антитела
IL17A-H069, содержащей сигнальный пептид
<400> 18
Met Glu Leu Gly Leu Ser Trp Ile Phe Leu Leu Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asp Tyr Glu Met His Trp Val Gln Gln Ala Pro Gly His Gly Leu
50 55 60
Glu Trp Met Gly Val Ile His Pro Gly Gly Gly Gly Thr Ala Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Gly Asp His Asp Gly Arg Thr Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
225 230 235 240
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
275 280 285
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Pro Gly
465
<210> 19
<211> 239
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот лёгкой цепи гуманизированного антитела
IL17A-H069, содержащей сигнальный пептид
<400> 19
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val
20 25 30
Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Arg Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Val Asp Phe Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe
100 105 110
Cys Gln Gln His Tyr Thr Thr Pro Phe Thr Phe Gly Pro Gly Thr Lys
115 120 125
Val Asp Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro
130 135 140
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
145 150 155 160
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
165 170 175
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
180 185 190
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
195 200 205
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
210 215 220
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 20
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот сигнального пептида тяжёлой цепи
гуманизированного антитела IL17A-H069
<400> 20
Met Glu Leu Gly Leu Ser Trp Ile Phe Leu Leu Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys
<210> 21
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот сигнального пептида лёгкой цепи
гуманизированного антитела IL17A-H069 / антитела мыши IL17A-M069
<400> 21
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser
<210> 22
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка тяжёлой цепи
гуманизированного антитела IL17A-H069
<400> 22
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Gln Gln Ala Pro Gly His Gly Leu Glu Trp Met
35 40 45
Gly Val Ile His Pro Gly Gly Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Gly Asp His Asp Gly Arg Thr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 23
<211> 113
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка лёгкой цепи
гуманизированного антитела IL17A-H069
<400> 23
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Arg
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Val Asp Phe Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
His Tyr Thr Thr Pro Phe Thr Phe Gly Pro Gly Thr Lys Val Asp Ile
100 105 110
Lys
<210> 24
<211> 329
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот константного участка тяжёлой цепи
гуманизированного антитела IL17A-H069
<400> 24
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 25
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот константного участка лёгкой цепи
гуманизированного антитела IL17A-H069
<400> 25
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 26
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов тяжёлой цепи гуманизированного антитела
IL17A-H069, содержащей сигнальный пептид
<400> 26
atggagttgg gactgagctg gattttcctt ttggctattt taaaaggtgt ccagtgtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggcat ctggctacac cttcacagac tatgagatgc actgggtcca acaggctcct 180
ggccatggat tggagtggat gggagtgatt caccctggag gaggaggcac agcctacaac 240
cagaagttca agggcagggt gaccatcaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtaccag gggagaccat 360
gatggcagga cagactactg gggacaaggc accctggtga cagtgtcctc tgcaagcacc 420
aagggcccat cggtcttccc cctggcaccc tcctccaaga gcacctctgg gggcacagcg 480
gccctgggct gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca 540
ggcgccctga ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac 600
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc 660
aacgtgaatc acaagcccag caacaccaag gtggacaaga aagttgagcc caaatcttgt 720
gacaaaactc acacatgccc accgtgccca gcacctgaac tcctgggggg accgtcagtc 780
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcacg 840
tgcgtggtgg tggacgtgag ccacgaagac cccgaggtca agttcaactg gtacgtggac 900
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac 960
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag 1020
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa 1080
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaccaag 1140
aaccaggtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 1200
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 1260
gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggg 1320
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac ccagaagtcc 1380
ctgtctctga gccctggcta atagtga 1407
<210> 27
<211> 720
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов лёгкой цепи гуманизированного антитела
IL17A-H069, содержащей сигнальный пептид
<400> 27
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcattctgac 60
attgtgatga cccagagccc tgactccctg gctgtgtccc tgggagagag ggctaccatc 120
aactgtaagt ccagccagtc cctgctgaac aggagcaacc agaagaacta cctggcttgg 180
tatcaacaga agcctggaca acctccaaaa ctgctggtgg actttgccag caccagggag 240
tctggagtgc ctgacaggtt ctctggctct ggctctggca cagacttcac cctgaccatc 300
tcctccctcc aagcagagga tgtggctgtc tacttctgtc aacaacacta caccacacca 360
ttcacctttg gacctggcac caaggtggac atcaagcgta cggtggctgc accatctgtc 420
ttcatcttcc cgccatctga tgagcagttg aaatctggaa ctgcctctgt tgtgtgcctg 480
ctgaataact tctatcccag agaggccaaa gtacagtgga aggtggataa cgccctccaa 540
tcgggtaact cccaggagag tgtcacagag caggacagca aggacagcac ctacagcctc 600
agcagcaccc tgacgctgag caaagcagac tacgagaaac acaaagtcta cgcctgcgaa 660
gtcacccatc agggcctgag ctcgcccgtc acaaagagct tcaacagggg agagtgttaa 720
<210> 28
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов сигнального пептида тяжёлой цепи
гуманизированного антитела IL17A-H069
<400> 28
atggagttgg gactgagctg gattttcctt ttggctattt taaaaggtgt ccagtgt 57
<210> 29
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов сигнального пептида лёгкой цепи
гуманизированного антитела IL17A-H069
<400> 29
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcattct 57
<210> 30
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка тяжёлой цепи
гуманизированного антитела IL17A-H069
<400> 30
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg catctggcta caccttcaca gactatgaga tgcactgggt ccaacaggct 120
cctggccatg gattggagtg gatgggagtg attcaccctg gaggaggagg cacagcctac 180
aaccagaagt tcaagggcag ggtgaccatc acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtac caggggagac 300
catgatggca ggacagacta ctggggacaa ggcaccctgg tgacagtgtc ctct 354
<210> 31
<211> 339
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка лёгкой цепи
гуманизированного антитела IL17A-H069
<400> 31
gacattgtga tgacccagag ccctgactcc ctggctgtgt ccctgggaga gagggctacc 60
atcaactgta agtccagcca gtccctgctg aacaggagca accagaagaa ctacctggct 120
tggtatcaac agaagcctgg acaacctcca aaactgctgg tggactttgc cagcaccagg 180
gagtctggag tgcctgacag gttctctggc tctggctctg gcacagactt caccctgacc 240
atctcctccc tccaagcaga ggatgtggct gtctacttct gtcaacaaca ctacaccaca 300
ccattcacct ttggacctgg caccaaggtg gacatcaag 339
<210> 32
<211> 996
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка тяжёлой цепи
гуманизированного антитела IL17A-H069
<400> 32
gcaagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacgt gcgtggtggt ggacgtgagc cacgaagacc ccgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacc 960
cagaagtccc tgtctctgag ccctggctaa tagtga 996
<210> 33
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка лёгкой цепи
гуманизированного антитела IL17A-H069
<400> 33
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttaa 324
<210> 34
<211> 249
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот антитела scFv мыши, использованного при
конструировании антитела IL17A-M069
<400> 34
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Met Ser Val Gly
1 5 10 15
Gln Lys Val Thr Met Asn Cys Lys Ser Asn Gln Ser Leu Leu Asn Arg
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Val Asp Phe Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln
85 90 95
His Tyr Thr Thr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser
115 120 125
Arg Ser Ser Gln Ala His Leu Gln Gln Ser Gly Ala Glu Leu Val Arg
130 135 140
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Leu Gly Tyr Thr Phe
145 150 155 160
Thr Asp Tyr Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu
165 170 175
Glu Trp Ile Gly Val Ile His Pro Gly Gly Gly Gly Thr Ala Tyr Asn
180 185 190
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
195 200 205
Thr Ala Tyr Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
210 215 220
Tyr Tyr Cys Thr Arg Gly Asp His Asp Gly Arg Thr Asp Tyr Trp Gly
225 230 235 240
Gln Gly Thr Thr Leu Thr Val Ser Ser
245
<210> 35
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот линкера, использованного при
конструировании фаговой библиотеки антител для связи с антителом scFv мыши
<400> 35
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser Arg
1 5 10 15
Ser Ser
<210> 36
<211> 1386
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов тяжёлой цепи антитела IL17A-M069 мыши,
содержащей сигнальный пептид
<400> 36
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagccag 60
gcccaccttc aacagtctgg ggctgagctg gtgaggcctg gggcttcagt gaagctgtcc 120
tgcaaggctt tgggctacac atttactgac tatgaaatgc actgggtgaa acagacacct 180
gtgcatggcc tggaatggat tggagttatt catccaggag gtggtggtac ggcctacaat 240
cagaagttca agggcaaggc cacactgact gcagacaagt cctccagtac agcctacatg 300
gagctcagca gcctgacatc tgaggactct gctgtctatt actgtacaag aggggatcac 360
gacggaagga ctgactactg gggccaaggc accactctca cagtctcctc agccaaaacg 420
acacccccat ctgtctatcc actggcccct ggatctgctg cccaaactaa ctccatggtg 480
accctgggat gcctggtcaa gggctatttc cctgagccag tgacagtgac ctggaactct 540
ggatccctgt ccagcggtgt gcacaccttc ccagctgtcc tgcagtctga cctctacact 600
ctgagcagct cagtgactgt cccctccagc acctggccca gcgagaccgt cacctgcaac 660
gttgcccacc cggccagcag caccaaggtg gacaagaaaa ttgtgcccag ggattgtggt 720
tgtaagcctt gcatatgtac agtcccagaa gtatcatctg tcttcatctt ccccccaaag 780
cccaaggatg tgctcaccat tactctgact cctaaggtca cgtgtgttgt ggtagacatc 840
agcaaggatg atcccgaggt ccagttcagc tggtttgtag atgatgtgga ggtgcacaca 900
gctcagacgc aaccccggga ggagcagttc aacagcactt tccgctcagt cagtgaactt 960
cccatcatgc accaggactg gctcaatggc aaggagttca aatgcagggt caacagtgca 1020
gctttccctg cccccatcga gaaaaccatc tccaaaacca aaggcagacc gaaggctcca 1080
caggtgtaca ccattccacc tcccaaggag cagatggcca aggataaagt cagtctgacc 1140
tgcatgataa cagacttctt ccctgaagac attactgtgg agtggcagtg gaatgggcag 1200
ccagcggaga actacaagaa cactcagccc atcatggaca cagatggctc ttacttcgtc 1260
tacagcaagc tcaatgtgca gaagagcaac tgggaggcag gaaatacttt cacctgctct 1320
gtgttacatg agggcctgca caaccaccat actgagaaga gcctctccca ctctcctggt 1380
aaataa 1386
<210> 37
<211> 721
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов лёгкой цепи антитела IL17A-M069 мыши,
содержащей сигнальный пептид
<400> 37
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcatagcgac 60
attgtgatgt cacagtctcc atcctccctg gctatgtcag taggacagaa ggtcactatg 120
aactgcaagt ccaatcagag ccttttaaat agaagcaatc aaaagaacta tttggcctgg 180
taccagcaga aaccaggaca gtctcctaaa cttctggtag actttgcatc cactagggaa 240
tctggggtcc ctgatcgctt cataggcagt ggatctggga cagatttcag tcttaccatc 300
agcagtgtgc aggctgagga cctggcagat tacttctgtc agcaacatta taccactcca 360
ttcacgttcg gctcggggac caagctggaa ataaaacggg ctgatgctgc accaactgta 420
tccatcttcc caccatccag tgagcagtta acatctggag gtgcctcagt cgtgtgcttc 480
ttgaacaact tctaccccaa agacatcaat gtcaagtgga agattgatgg cagtgaacga 540
caaaatggcg tcctgaacag ttggactgat caggacagca aagacagcac ctacagcatg 600
agcagcaccc tcacgttgac caaggacgag tatgaacgac ataacagcta tacctgtgag 660
gccactcaca agacatcaac ttcacccatt gtcaagagct tcaacaggaa tgagtgttaa 720
a 721
<210> 38
<211> 461
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот тяжёлой цепи антитела IL17A-M069 мыши,
содержащей сигнальный пептид
<400> 38
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Ala His Leu Gln Gln Ser Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Leu Gly Tyr Thr Phe
35 40 45
Thr Asp Tyr Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu
50 55 60
Glu Trp Ile Gly Val Ile His Pro Gly Gly Gly Gly Thr Ala Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Gly Asp His Asp Gly Arg Thr Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser
130 135 140
Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val
145 150 155 160
Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro
195 200 205
Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro
210 215 220
Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly
225 230 235 240
Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile
245 250 255
Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys
260 265 270
Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln
275 280 285
Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln
290 295 300
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu
305 310 315 320
Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg
325 330 335
Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
340 345 350
Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro
355 360 365
Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr
370 375 380
Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln
385 390 395 400
Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly
405 410 415
Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu
420 425 430
Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn
435 440 445
His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
450 455 460
<210> 39
<211> 239
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот лёгкой цепи антитела IL17A-M069 мыши,
содержащей сигнальный пептид
<400> 39
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Met
20 25 30
Ser Val Gly Gln Lys Val Thr Met Asn Cys Lys Ser Asn Gln Ser Leu
35 40 45
Leu Asn Arg Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Ser Pro Lys Leu Leu Val Asp Phe Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Ser Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Asp Tyr Phe
100 105 110
Cys Gln Gln His Tyr Thr Thr Pro Phe Thr Phe Gly Ser Gly Thr Lys
115 120 125
Leu Glu Ile Lys Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro
130 135 140
Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe
145 150 155 160
Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp
165 170 175
Gly Ser Glu Arg Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp
180 185 190
Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys
195 200 205
Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys
210 215 220
Thr Ser Thr Ser Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
225 230 235
<210> 40
<211> 324
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот константного участка тяжёлой цепи
антитела IL17A-M069 мыши
<400> 40
Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala
1 5 10 15
Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro
100 105 110
Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu
115 120 125
Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser
130 135 140
Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln
210 215 220
Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val
225 230 235 240
Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val
245 250 255
Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln
260 265 270
Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn
275 280 285
Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val
290 295 300
Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His
305 310 315 320
Ser Pro Gly Lys
<210> 41
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот константного участка лёгкой цепи
антитела IL17A-M069 мыши
<400> 41
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 42
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот сигнального пептида тяжёлой цепи
антитела IL17A-M069 мыши
<400> 42
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser
<210> 43
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов сигнального пептида тяжёлой цепи
антитела IL17A-M069 мыши
<400> 43
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagc 57
22
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ IL-17A И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2816204C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ PD-L1 и LAG-3 | 2017 |
|
RU2784388C2 |
| АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2739208C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| СВЯЗЫВАЮЩИЙ RGMa БЕЛОК И ЕГО ИСПОЛЬЗОВАНИЕ | 2016 |
|
RU2809500C2 |
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| БИСПЕЦИФИЧЕСКИЙ БЕЛОК | 2019 |
|
RU2784486C1 |
| АНТИТЕЛО К B7-H4, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2792748C2 |
| АНТИТЕЛО К EphA4 | 2020 |
|
RU2816371C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ИНДУЦИРУЮЩАЯ ИММУННЫЙ ОТВЕТ НА АНТИГЕН-МИШЕНЬ | 2012 |
|
RU2722829C2 |
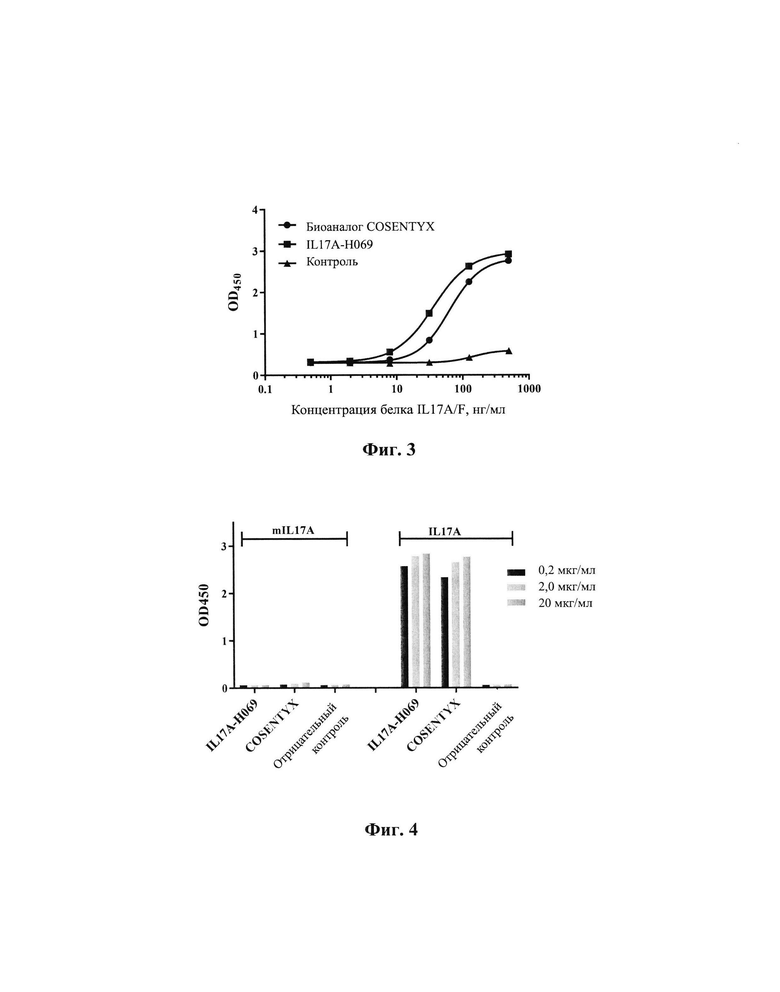
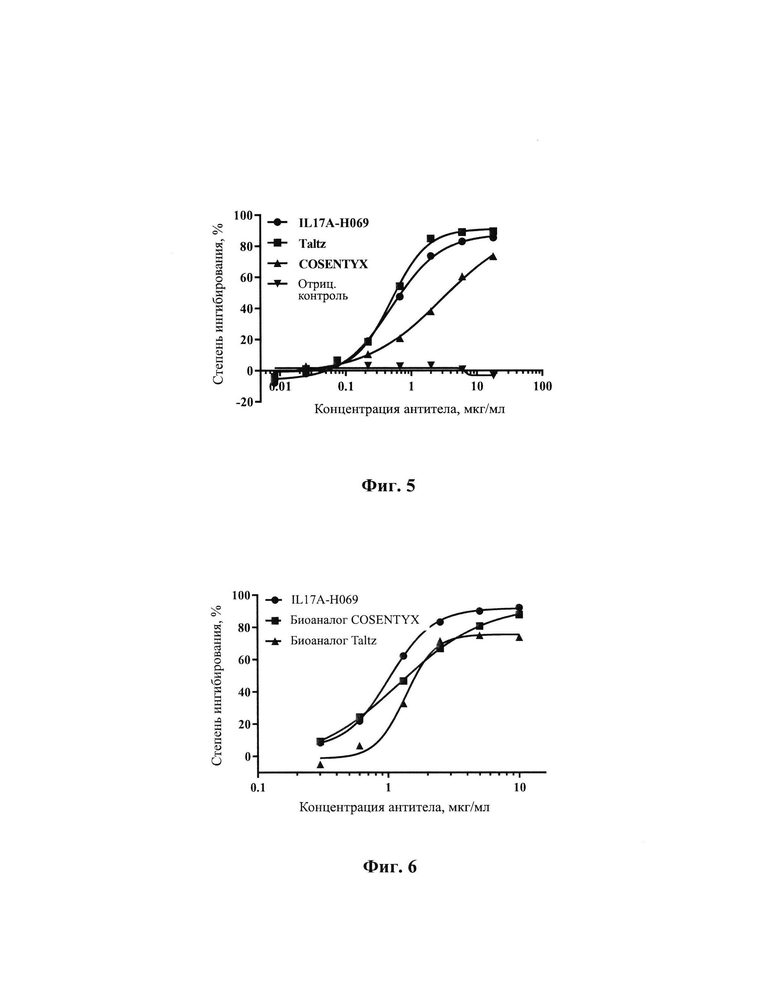
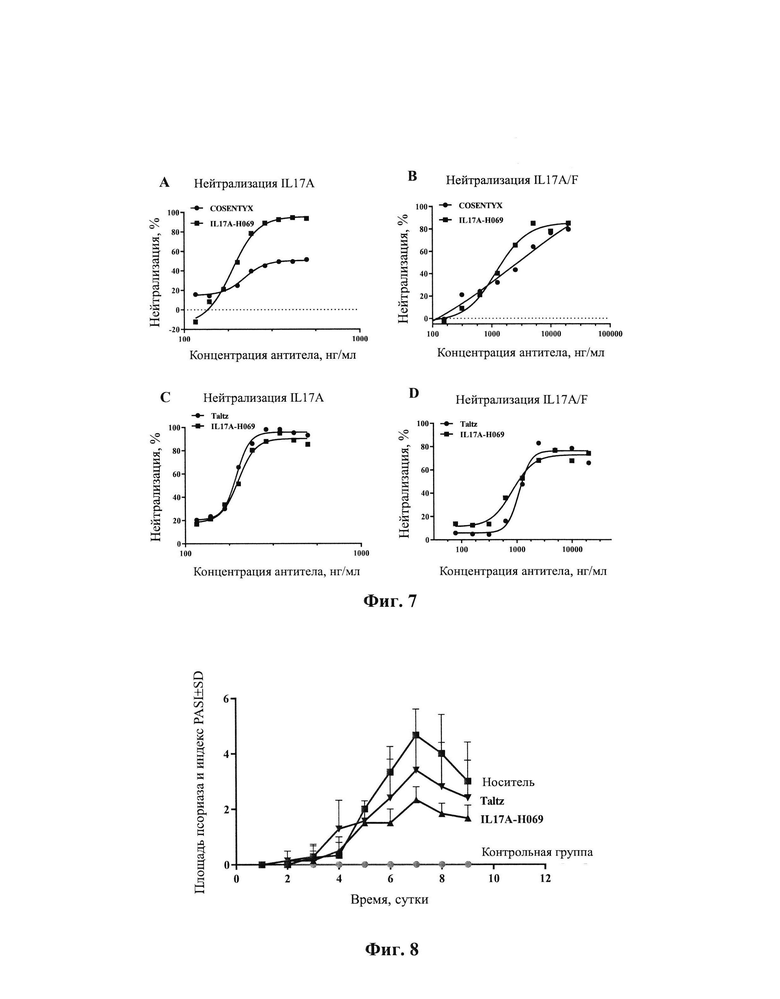
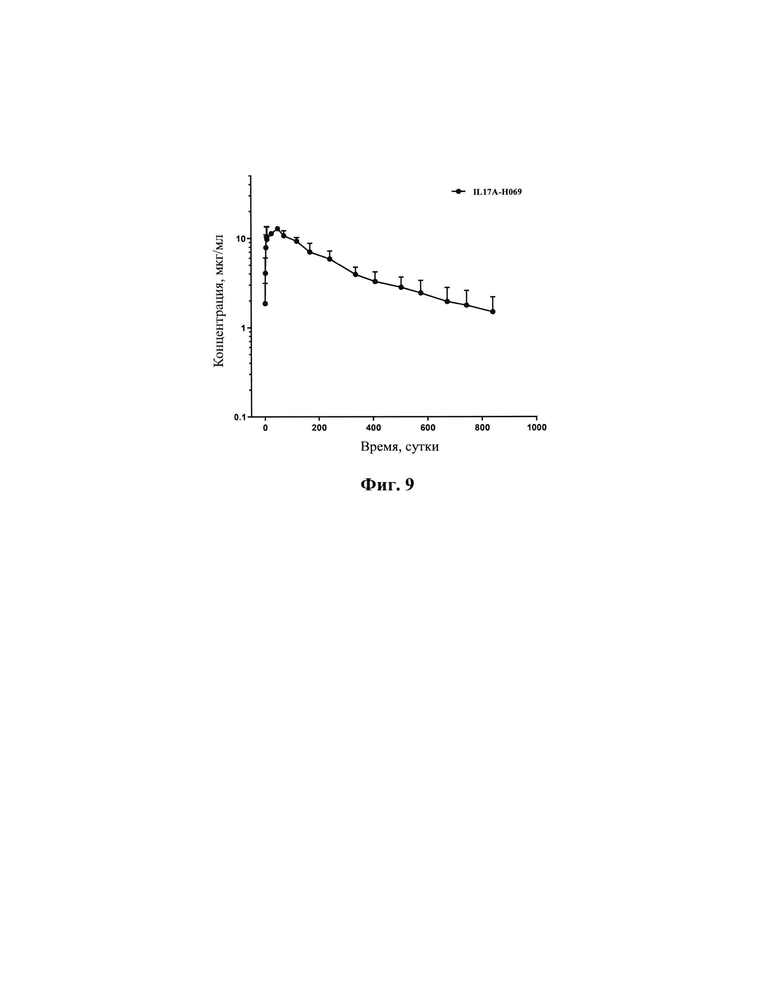
Изобретение относится к области биотехнологии. Предложено моноклональное антитело, которое связывается с IL17A. Настоящее изобретение также раскрывает нуклеиновую кислоту, кодирующую указанное антитело, вектор экспрессии и клетку-хозяина, содержащие последовательность указанной нуклеиновой кислоты, способ получения антитела. Кроме того, изобретение относится к конъюгату антитела и лекарственного средства, фармацевтической композиции и набору, которые применяются для лечения псориаза. Указанное моноклональное антитело специфически связывает белок IL17A с высокой аффинностью, обладает превосходной или эквивалентной способностью блокировать индуцированную IL17A и IL17A/F секрецию воспалительных цитокинов эпидермальными фибробластами HFF человека по сравнению с сопоставимыми препаратами, а также улучшенной эффективностью in vivo. 12 н. и 10 з.п. ф-лы, 9 ил., 5 табл., 7 пр.
1. Изолированное антитело против IL17A или его антиген-связывающий фрагмент, содержащий
вариабельный участок тяжелой цепи, содержащий участок CDR1 тяжелой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 13, участок CDR2 тяжелой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 14, и участок CDR3 тяжелой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 15; и
вариабельный участок легкой цепи, содержащий участок CDR1 легкой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 10, участок CDR2 легкой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 11, и участок CDR3 легкой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 12.
2. Антитело против IL17A или его антиген-связывающий фрагмент по п. 1, содержащий
вариабельный участок тяжелой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 22, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности SEQ ID NO: 22; и
вариабельный участок легкой цепи, имеющий последовательность аминокислот, указанную в SEQ ID NO: 23, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности SEQ ID NO: 23.
3. Антитело против IL17A или его антиген-связывающий фрагмент по п. 1, отличающееся тем, что указанное антитело против IL17A или его антиген-связывающий фрагмент представляет собой гуманизированное антитело или химерное антитело.
4. Антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-3, дополнительно содержащее
константный участок тяжелой цепи и константный участок легкой цепи, где, предпочтительно, указанный константный участок тяжелой цепи представляет собой константный участок тяжелой цепи IgG1, имеющий последовательность аминокислот, указанную в SEQ ID NO: 24, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности SEQ ID NO: 24; и/или
константный участок легкой цепи представляет собой константный участок легкой цепи каппа человека, имеющий последовательность аминокислот, указанную в SEQ ID NO: 25, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности SEQ ID NO: 25.
5. Антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-4, дополнительно содержащее
сигнальный пептид, присоединенный к вариабельному участку тяжелой цепи, и/или сигнальный пептид, присоединенный к вариабельному участку легкой цепи,
где, предпочтительно, указанный сигнальный пептид, присоединенный к вариабельному участку тяжелой цепи представляет собой последовательность аминокислот, указанную в SEQ ID NO: 20, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности SEQ ID NO: 20; и/или
указанный сигнальный пептид, присоединенный к вариабельному участку легкой цепи, представляет собой последовательность аминокислот, указанную в SEQ ID NO: 21, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности SEQ ID NO: 21.
6. Антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-5, отличающееся тем, что указанное антитело против IL17A или его антиген-связывающий фрагмент представляет собой антитело IgG, предпочтительно антитело IgG1.
7. Антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-6, отличающееся тем, что указанное антитело против IL17A или его антиген-связывающий фрагмент представляет собой моноклональное антитело.
8. Антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-7, отличающееся тем, что аффинность связывания KD указанного антитела против IL17A или его антиген-связывающего фрагмента с рекомбинантным белком IL17A человека составляет 0,1-10Е-11 М, предпочтительно 0,5-5Е-11 М и более предпочтительно 2,88Е-11 М.
9. Антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-7, отличающееся тем, что аффинность связывания KD указанного антитела против IL17A или его антиген-связывающего фрагмента с рекомбинантным белком IL17A/F человека составляет 0,1-10Е-10 М, предпочтительно 0,5-5Е-10 М и более предпочтительно 5,37Е-10 М.
10. Антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-9, отличающееся тем, что указанный антиген-связывающий фрагмент представляет собой Fv, Fab, Fab’, Fab’-SH, F(ab’)2, фрагмент Fd, фрагмент Fd’, молекулу одноцепочечного антитела или однодоменное антитело; при этом молекула одноцепочечного антитела предпочтительно представляет собой scFv, ди-scFv, три-scFv, диатело или scFab.
11. Конъюгат антитела и лекарственного средства для применения в лечении псориаза, содержащий антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-10 и дополнительный терапевтический агент, где предпочтительно указанное антитело против IL17A или его антиген-связывающий фрагмент связан с указанным дополнительным терапевтическим агентом посредством линкера.
12. Нуклеиновая кислота, кодирующая антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-10.
13. Нуклеиновая кислота по п. 12, содержащая последовательность нуклеотидов, указанную в SEQ ID NO: 30, кодирующую вариабельный участок тяжелой цепи, и/или последовательность нуклеотидов, указанную в SEQ ID NO: 31, кодирующую вариабельный участок легкой цепи.
14. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 12 или 13.
15. Клетка-хозяин для получения антитела против IL17A или его антиген-связывающего фрагмента по любому из пп. 1-10, содержащая нуклеиновую кислоту по п. 12 или 13 или вектор экспрессии по п. 14.
16. Способ получения антитела против IL17A или его антиген-связывающего фрагмента по любому из пп. 1-10, включающий выращивание клетки-хозяина по п. 15 в условиях, подходящих для экспрессии антитела, и выделения экспрессированного антитела из культуральной жидкости.
17. Фармацевтическая композиция для применения в лечении псориаза, содержащая антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-10, или конъюгат антитела и лекарственного средства по п. 11, и фармацевтически приемлемый носитель.
18. Применение антитела против IL17A или его антиген-связывающего фрагмента по любому из пп. 1-10, или конъюгата антитела и лекарственного средства по п. 11, или фармацевтической композиции по п. 17 в лечении псориаза.
19. Способ лечения псориаза, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества антитела против IL17A или его антиген-связывающего фрагмента по любому из пп. 1-10, или конъюгата антитела и лекарственного средства по п. 11, или фармацевтической композиции по п. 17 для лечения псориаза.
20. Применение антитела против IL17A или его антиген-связывающего фрагмента по любому из пп. 1-10, или конъюгата антитела и лекарственного средства по п. 11, или фармацевтической композиции по п. 17 для получения лекарственного средства для лечения псориаза.
21. Фармацевтическая комбинация для применения в лечении псориаза, содержащая антитело против IL17A или его антиген-связывающий фрагмент по любому из пп. 1-10, или конъюгат антитела и лекарственного средства по п. 11, или фармацевтическую композицию по п. 17, и один или более дополнительный терапевтический агент.
22. Набор для применения в лечении псориаза, содержащий антитело против IL17A или антиген-связывающий фрагмент по любому из пп. 1-10, или конъюгат антитела и лекарственного средства по п. 11, или фармацевтическую композицию по п. 17, дополнительно содержащий устройство для введения.
| WO 2018236728 A1, 27.12.2018 | |||
| WO 2014161570 A1, 09.10.2014 | |||
| BAUER E | |||
| et al., Brodalumab-an IL-17RA monoclonal antibody for psoriasis and psoriatic arthritis, Expert Opinion on Biological Therapy, 2015, vol.15(6), pp.883-893 | |||
| БАКУЛЕВ А.Л., Стратегия "лечение до достижения цели" при псориазе | |||
| Актуальные вопросы устойчивости к биологической терапии, |
