Изобретение относится к биотрансформации соединений колхициноидов в соответствующие 3-O-гликозилпроизводные, осуществляемой посредством выбранных микробных штаммов. Способ данного изобретения обеспечивает получение соединений колхициноидов, гликозидированных исключительно по С-3 положению ароматического цикла А, исходя из колхицина, тиоколхицина или их производных, с высокими выходами и чистотой.
Соединения колхициноидов, гликозидированные исключительно по С-3 положению бензольного кольца имеют важное фармакологическое значение по причине их высокой эффективности или возможности получения новых лекарственных препаратов.
В частности, тиоколхикозид (3-О-глюкозилтиоколхицин) является в высшей степени важным активным ингредиентом, применяемым в области фармации, особенно в терапии скелетно-мышечной системы и в качестве исходного материала для получения новых противоопухолевых, иммунодепрессивных, противопсориазных и противовоспалительных лекарственных средств. Ранее был предпринят ряд попыток получения соединений 3-гликозилколхициноидов либо посредством химических реакций, либо посредством биотрансформации.
Химический путь состоит в последовательном ряде комплексных, неспецифических реакций, приводящих к смеси гликозидированных производных, некоторые из которых неактивны. По этой причине конверсионные выходы эффективного продукта, особенно гликозидированного по С-3 положению ароматического кольца, очень низкие.
Биологический подход по существу связан с биотрансформацией тиоколхицина с помощью культуры Centella Asiatica в моногликозидированные производные по С-2 и С-3 ароматического кольца; такие трансформации не являются высоко селективными и сопровождаются невысокими выходами и низкой продуктивностью (Solet J.M., et al., Phytochemistry 33, 4, 817-820, 1993).
Другие попытки биотрансформировать соединения колхициноидов приводят просто к деметилированию метоксигрупп, связанных с ароматическим кольцом (по С-2 и С-3), и в любом случае характеризуются ограниченными выходами и низкой региоселективностью.
Так, Hufford C.D. et al. [J.Pharm. Sc., 68, 10, 1239-1242, 1979), используя Streptomycts griseus и/или Streptomyces spectabilis, и Bettet P. et al. (GB- 923421, 1959), используя различные штаммы Streptomyces и других видов бактерий и грибков, пытались превратить колхицин и его производные в соответствующие 3-деметилированные производные. Результаты этих известных способов подтверждают сказанное выше в отношении неселективности вовлеченных в процесс микробных ферментов, к примеру, по С-2, С-3 или С-10 алкалоидной молекулы. Кроме того, уровни продуктивности указанных каталитических систем достаточно низкие вследствие низких конверсионных выходов, пониженных концентраций субстрата, которые можно использовать, и часто происходящего разрушения кольца трополона.
Недавно, Poulev et al. (J.Ferment. Bioeng, 79, 1, 33-38, 1995) добились специфической биотрансформации бактериальных микроорганизмов, но все еще с довольно низкими выходами и продуктивностью.
Активный комплекс ферментов из микроорганизмов, подобных вышеуказанным (Streptomyces, Bacillus, и т.д.), был использован для биотрансформации других соединений, таких как (maytansinoids) мэйтанзиноиды (патент США, 4 361 650: Izawa, М., et al., J.Antibiotics, 34, 12, 1587-1590, 1981). В этом случае также катализируемая реакция состоит исключительно в деметилировании, характеризуемом низкими конверсионными выходами и низкой продуктивностью.
Гликозилтрансферазная активность α-амилазы из Bacillus megaterium описана (Brum P.J., et al., Starch, 43, 8, 319-323, 1991); причем акцепторная специфичность реакции трансферазы (исключительно глюкоза или глюкозиды) особенно высока. Циклодекстрин-глюкозилтрансферазы, продуцируемые тем же самым микробным источником, катализируют α-1,4-трансглюкозилирование (rubusoside) рубузозида (β-D-глюкозиловый эфир 13-O-β-D-глюкозил-стевиола), исходя из крахмала. Кроме того, в этих биопревращениях акцептором реакции трансферазы является фракция глюцидного субстрата (Darise, М., et al., Agric. Bioel. Chem. , 48, 10, 2483-2488, 1984). Циклодекстрин-глико-зилтрансферазы ранее использовались для получения циклодекстринов G6, G7 и G8 из крахмала (Kitahata, S., Okada, S., Agric. Biol. (Chem., 38, 12, 2413-2417, 1974).
Эти примеры подтверждают высокую субстратную специфичность активности гликозилтрансферазы, экспрессированной Bacillus magaterium, которая затрагивает только акцепторы глюцида, не вовлекая таким образом ни в какие реакции вторичные метаболиты, имеющие другую, сложную молекулярную структуру (такие как колхициноиды). Действительно, не известно примеров использования указанных микроорганизмов для ферментативной конверсии колхициноидов до 3-гликозилпроизводных.
В настоящее время найдено, что штаммы Bacillus megaterium, способные к росту в присутствии высоких концентраций колхицина и тиоколхицина, обладают крайне высокой, очень специфической активностью биотрансформации колхициноидных субстратов в производные, гликозидированные исключительно по С-3 положению ароматического кольца. Такие превращения происходят за очень короткое время и характеризуются необычайно высокими выходами.
Таким образом, изобретение относится к способу получения соединений 3-О-гликозилколхициноидов формулы (I)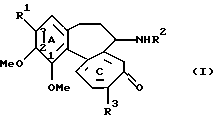
где R1 - остаток О-гликозида; R2 - водород или С1-С7 ацил; R3 - С1-С6 алкокси или C1-C6 тиоалкил, включающий биотрансформацию соединений, в которых R1 - ОН или метокси, посредством Bacillus megaterium.
Bacillus megaterium представляет собой грамположительную споргенерирующую бактерию с диаметром клеток более 1 мкм; растущую аэробно на ряде культурных сред; катализа-позитивную; гидролизующую желатин.
Как подтверждается исследованиями роста и микроскопическим анализом, штаммы Bacillus megaterium, которые могут быть использованы согласно изобретению, обладают удовлетворительной способностью к росту и сохранению жизнеспособности также при высоких концентрациях колхицина и/или тиоколхицина (свыше 3 г/л).
У родственных видов, таких как Bacillus cereus, уже при концентрациях субстрата 1,5 г/л проявляются трудности в росте (абсорбционная способность 15-20% от контроля), которые становятся еще более явными при концентрациях 3 г/л, когда наблюдается очень сильный автолиз. Выбранные культуры Bacillus megaterium, напротив, при вышеуказанных концентрациях могут достигать значительно больших уровней роста (удваивается или утраивается) по сравнению с Bacillus cereus.
Высокая селективность и эффективность биотрансформации поразительна и необычна, поскольку уровни выходов находятся в пределах от 80 до 100%, а обычно около 90-95%.
Кроме того, используемые в биоконверсии микроорганизмы способны постоянно поддерживать каталитическую активность, даже в повторных стадиях ферментации, обеспечивая вследствие этого специфическую биоконверсию периодическим и непрерывным способами. Кроме того, этот способ обеспечивает высокие уровни продуктивности и воспроизводимости.
Указанная реакция региоселективности обеспечивает, вдобавок к значительным выходам продукта, высокое качество и чистоту образующегося продукта, обеспечивая таким образом его 100% чистоту с помощью простой обработки в нисходящем потоке.
Кроме того, важными преимуществами являются пониженная опасность на стадии очистки и выделения продукта, экономичность способа, адекватность (воспроизводимость) и безопасность применения.
Последовательность операций по селекции бактериальных штаммов, применяемых в способе по изобретению, включает:
A) Селекцию культур Bacillus megaterium, способных к росту в присутствии высоких концентраций колхицинового субстрата, из природных источников или из коллекции штаммов.
B) Отбор изолята из А) для подтверждения активности по трансформации колхицинов в соответствующие 3-О-гликозилпроизводные путем биоконверсионных испытаний на специфических субстратах, вводимых в последовательно возрастающих концентрациях.
C) Микробиологическую идентификацию штаммов, отобранных в В).
D) Последовательное увеличение выхода биотрансформации путем мишень-специфической селекции бактериальной популяции из В).
Е) Изучение и оптимизацию критических параметров ферментации для оптимизации биотрансформации.
F) Изучение и оптимизацию способов конверсии высокопродуктивных культур для гарантии стабильной, гомогенной инокуляции с целью продуктивного применения в промышленном масштабе.
G) Увеличение масштабов реакции в ферментере, при единовременной загрузке, при периодическом и непрерывном способах.
Н) Отработку и оптимизацию способов обработки в нагнетательном потоке и способов выделения продукта.
В частности, пригодные для использования по данному изобретению микроорганизмы могут быть выбраны из коллекции культур, полученной из центров-депозиториев штаммов, или из почвенных образцов разнообразного происхождения путем селективного выделения на различных агаровых средах, содержащих органический источник азота (пептоны, дрожжевые экстраты, мясные экстракты, аспарагин и т.д.), углеродный источник (глицерин, крахмал, мальтозу, глюкозу и т. д. ), с рН 5-8, предпочтительно 6-7. Температура инкубации изменяется в пределах от 20 до 45oС, предпочтительно 28-40oС.
Способность культуры к росту в присутствии токсических концентраций трансформируемого колхицинового субстрата оценивают по методикам последовательного разведения и параллельного посева на различных содержащих агар субстратах, часть из которых содержит предварительно добавленный колхицин или тиоколхицин, при концентрациях 0,1-3,0 г/л (такой, чтобы подавить рост основной части микроорганизмов).
Колонии, способные к росту в описанных условиях, извлекают, соблюдая стерильность, и помещают на различные содержащие агар среды для подтверждения их чистоты и гомогенности роста.
Культуральные среды, используемые для сохранения культур, являются обычными микробиологическими субстратами, содержащими органические источники азота (пептоны, дрожжевые экстракты, триптон, мясные экстракты и т.д.), источник углерода (глюкоза, мальтоза, глицерин и т.д.), при рН 5-8, предпочтительно 6-7. Интервалы температуры инкубации составляют от 20 до 45oС, предпочтительно 28-40oС.
Выбранные микроорганизмы испытывают затем на способность к росту в условиях погружения в культуру, в присутствии соединений колхициноидов, и способность трансформации последних в соответствующие 3-гликозилпроизводные.
Указанные испытания проводят в 100 мл колбах, содержащих 20 мл жидкой среды (с различными составами сред), включающей органические источники азота (один или более) (дрожжевые экстракты, пептоны, триптон, казеиновые гидролизаты, мясной экстракт, гидролизат из початков кукурузы и т.д.), источники углерода (один или более) (глюкоза, глицерин, крахмал, сахароза и т.д.), неорганические источники фосфора и азота и неорганические соли различных ионов (К+, Na+, Мg++, Са++, Fe++, Mn++ и т.д.).
Образцы культур могут необязательно быть подвергнуты мутагенным обработкам с применением общепринятых мутагенных методик (облучение УФ-лучами и т.д.) для индуцирования мутантов, обладающих специфической активностью биоконверсии, которая может быть оценена теми же способами, что указаны выше.
Образцы культур из каждого источника на биоконверсию анализируют для оценки продуцирования 3-гликозилпроизводных с помощью ТСХ- и ЖХВР-хроматографий.
Способность отобранных микроорганизмов к трансформации колхициноидных субстратов в соответствующие 3-гликозилпроизводные подтверждается путем биоконверсионных испытаний в колбах объемом 300 мл в тех же культуральных бульонах, что используются на стадии отбора.
Дающие положительные реакции микроорганизмы используют в испытаниях по оптимизации биоконверсии, в различных культуральных бульонах, в 300-мл объемах. Основными изучаемыми культуральными и ферментативными параметрами являются следующие: органические источники азота, источники углерода, неорганические соли, температура, перемешивание-аэрация, рН, время инкубации, коэффициент инокулята, стадии субкультуры, время введения трансформируемого субстрата.
Отобранные бактериальные микроорганизмы, способные ускорять биотрансформацию по данному изобретению, могут расти как на твердых, так и на жидких культуральных субстратах, содержащих один или более органических источников азота, предпочтительно дрожжевой экстракт, мясной экстракт, пептон, триптон, казеиновые гидролизаты, гидролизат из початков кукурузы и т.д. Источниками углерода, используемыми для роста и биотрансформации, являются глюкоза, фруктоза, сахароза, глицерин, солодовый экстракт и т.д., предпочтительно глюкоза, фруктоза и глицерин. Культуральная среда содержит, кроме того, неорганические источники фосфора и соли К+, Na+, Мg++, NH4 + и т.д.
Отобранные микроорганизмы могут расти при 20-45oС, предпочтительно 28-40oС, при рН между 5 и 8, предпочтительно 6-7. В тех же условиях рассматриваемые микроорганизмы способны к трансформации соединений колхициноидов в соответствующие 3-гликозилпроизводные. Указанная трансформация происходит в условиях погружения в культуру, в колбах, инкубируемых на ротационном вибраторе, со скоростью перемешивания 150-250 об/мин.
Благодаря связанным с микробным ростом особенностям кинетики рассматриваемой биотрансформации оптимальными условиями для биотрансформации являются те же условия, которые оптимальны для роста. Поэтому культуральные среды, полезные для стимулирования быстрого микробного роста, такие как перечисленные выше среды, основанные на органических и неорганических компонентах, также полезны для высокой активации биотрансформации на описанном субстрате. Последний добавляют к культуре на начальной стадии ферментации.
Биотрансформация по изобретению основана на ферментативной конверсии, которая начинается во время фазы экспоненциального роста и продолжается с параллельной прогрессией этой стадии роста; максимальные уровни конверсии до 3-гликозилпроизводного (очень высокая: достигает до 95-100%) достигаются за первые 24-30 ч. Региоселективность биотрансформации является абсолютной: доказательством служит то, что 2-гликозилпроизводные никогда не присутствуют. Образующиеся продукты являются исключительно внеклеточными.
Биотрансформация по изобретению может быть проведена в увеличенных до размеров ферментера масштабах, особенно при поддержании неизменных культуральных условий, что касается культуральной среды, температуры и когда речь идет о временах процессов. Для достижения быстрого роста важны адекватные уровни перемешивания-аэрации, в частности требуются уровни аэрации 1-2 л воздуха на 1 л культуры в минуту (об/об.мин) предпочтительно 1,5-2 (об/об. мин).
Полученные в ходе биоконверсии продукты экстрагируют из культуральных бульонов после отделения биомасс от жидкой фракции путем центрифугирования и выделения супернатанта или микрофильтрацией и выделением пермеата. Культура может быть обработана спиртами с целью оптимального выделения продукта.
Очистка и выделение продуктов биотрансформации может быть проведена с применением хроматографических способов разделения на адсорбционных смолах и элюирования спиртами, преимущественно метанолом. Воднометанольные растворы, содержащие продукт, могут быть дополнительно очищены экстракцией метилен-хлоридом. После дополнительных обработок смесями спиртов и органических растворителей продукт может быть выделен в чистом виде из полученных спиртовых растворов путем кристаллизации.
Способ биотрансформации является специфическим для субстратов, содержащих группу трополона, и может быть применен к ряду соединений колхициноидов, таких как колхицин, тиоколхицин, 3-деметилколхицин, 3-деметилтиоколхицин, N-дезацетил-тиоколхицин и другие разнообразно замещенные колхицины.
Другие природные соединения, не имеющие трополона, не гликозидируются с помощью Bacillus megaterium.
Глюкоза может быть заменена другими сахарами, такими как фруктоза или галактоза, что не приводит к потере активности гликозилтрансферазы.
Последующие примеры описывают изобретение более подробно.
Пример 1. Аликвоты культур Bacillus meqaterium, выделенных из сельскохозяйственной почвы, ресуспендируют в 20 мл стерильного раствора соли и последовательно разбавляют до коэффициента разведения 1:10.000.000. Суспензии при различных разведениях высевают на чашки с LB-агар-культуральной средой и на LB-агар, дополненный соответственно колхицином или тиоколхицином, до конечной концентрации 2 г/л. Культуры инкубируют при 28oС в течение 3 дней в темноте. Растущие на выбранной среде колонии дополняют колхициноидом, выделяют и очищают путем высеивания на неселективную среду; указанные образцы инкубируют, как указано выше, но в течение более короткого периода времени (24 ч). В дальнейшем, культуры переносят в ту же агаровую среду, в пробирку, и инкубируют, как указано выше, в течение 24 ч.
Аликвоты культур, отобранные как описано, используют для инокуляции 100 мл колб Эрленмейера, содержащих 20 мл культуральной среды ST, дополненной колхицином или тиоколхицином, до конечной концентрации 0,4 мг/мл. Указанные культуры инкубируют в течение ночи при 28oС на роторном вибраторе при 200 об/мин.
Трансформацию колхицинового субстрата контролируют анализами аликвот культуральных бульонов, отбираемыми каждые 3-4 часа, путем ТСХ на силикагеле с применением ацетон:этилацетат:вода 5:4:1 элюентной системы.
После 4 дней инкубации аликвоты культур, демонстрирующих очевидную каталитическую активность в отношении 3-гликозилпроизводного, извлекают для высеивания на чашки посредством описанного выше последовательного разведения для получения нового инокулята в пробирке. Испытание на биотрансформацию в колбе повторяют в тех же условиях, что описаны выше, но используя значительно большие конечные концентрации колхицина и тиоколхицина (1 мг/мл). Отдельные наиболее активные культуры (конверсия субстрата равна или выше 80%) используют для получения инокулята в замороженных криоскопических пробирках, как описано в примере 3.
Состав культуральной среды, г/л:
1) LB-агар (Стерилизация: 121oС • 20') - рН 7
Триптон - 10
Дрожжевой экстракт - 5
NaCl - 10
Агар-Агар - 15
2) ST -бульон (Стерилизация: 121oС • 20') - рН 7
Глюкоза - 20
Глицерин - 10
Пептон - 15
Дрожжевой экстракт - 5
NaCl - 3
NH4Cl - 3
К2НРO4 - 8
КН2РО4 - 3
МgSO4•7Н20 - 0,5
Пример 2. Повторяют способ, описанный в примере 1, исходя из культур Bacillus megaterium, получаемых из следующей коллекции штаммов (Deutsch Sammlung von Mikroorganismen, Branscherig, Germany): DSM 90, 509, 322, 333, 1667, 1670, 1671.
Культуру, отобранную, как в примере 1, и дополненную тиоколхицином (1 мг/мл), инкубируют в течение 4 дней в жидкой культуре: ТСХ - анализ свидетельствует о протекании трансформации субстрата в тиоколхикозид с конверсионными выходами, изменяющимися от 50% (штамм DSM1671) до 70% (штамм DSM 90), до 80% и выше (штаммы DSM 333, DSM 509, DSM 1667, DSM 1670).
Пример 3. Аликвоты образцов культур в пробирке, отобранные, как описано в приведенном выше примере, используют для инокуляции 100 мл колб Эрленмейера, содержащих 20 мл бульона ST.
Обе культуры инкубируют при 30oС на роторном вибраторе при 200 об/мин в течение ночи. После инкубации культуры дополняют стерильным раствором глицерина до конечной концентрации 20%. Затем культуры распределяют по 2 мл криоскопическим пробиркам и сразу же погружают в жидкий азот.
Через несколько дней 10% культур быстро размораживают при 37oС. Аликвоты каждой криоскопической пробирки используют для инокуляции 100 мл колб Эрленмейера, содержащих 20 мл ST среды, которые впоследствии инкубируют при 28oС в течение ночи (прекультура) при 200 об/мин. После инкубации по 2 мл каждой прекультуры переносят в стерильных условиях в 20 мл свежей ST среды, дополненной к этому времени колхицином или тиоколхицином, до конечной концентрации 1 г/л. Биотрансформацию проводят и контролируют в описанных в примере 1 условиях. Анализ подтверждает, что трансформация субстрата в 3-гликозилпроизводное осуществляется с указанными выше количественными выходами (80% и выше), доказывая таким образом каталитическую стабильность замороженных культур.
Параллельные контрольные опыты бульонных культур, высеянных на LB-Агар сразу же после размораживания, подтверждают жизнеспособность, гомогенность и чистоту замороженных культур.
Пример 4. Аликвоты культур в криоскопической пробирке после размораживания используют для инокуляции 300 мл колб Эрленмейера, содержащих 50 мл ST среды (прекультура). После инкубации в течение ночи при 30oС, 250 об/мин, 5 мл прекультуры переносят в 50 мл той же среды, дополненной колхицином до конечной концентрации 1 г/л. Культуры инкубируют в течение 2 дней в тех же условиях, что описаны выше. Каждые 4 ч отбирают образцы для оценки уровня роста (измерение абсорбции при 600 нм), образования колхикозида (ТСХ и ЖХВР), стерильности (на LB-Агаре) и для микроскопического морфологического исследования.
Для ЖХВР анализа к 1 мл-фракциям культуральных бульонов добавляют 9 мл метанола и центрифугируют при 13.000 об/мин в течение 2 мин. Содержание в колкикозиде супернатанта анализируют обращенно-фазовой ЖХВР с изократным элюированием с помощью элюентной системы вода:ацетонитрил 80:20.
ЖХВР анализ подтверждает, что конверсия колхицина в колхикозид следует параллельной прогрессии роста. После порядка 26 ч инкубации биоконверсия завершается.
Конечный выход колхикозида находится в пределах от 80% до 85%.
Пример 5. Повторяют методику, описанную в примере 4, добавляя к культурам вместо колхицина тиоколхицин, при той же самой конечной концентрации (1 г/л).
Рост и реакции продуцирования культур такие же, как описаны для колхицина, с выходами тиоколхикозида порядка 90%.
Пример 6. Повторяют методику, описанную в примере 4, добавляя к культуре вместо колхицина 3-деметилтиоколхицин, при той же самой конечной концентрации (1 г/л). Рост и реакции продуцирования культур такие же, как описаны выше, с выходами тиоколхикозида порядка 90%.
Пример 7. Повторяют методику, описанную в примере 4, добавляя к культуре вместо колхицина N-формилтиоколхицин, при той же самой конечной концентрации (1 г/л). Рост и реакции продуцирования культур такие же, как описаны выше, с выходами N-деацетил-N-формилтиоколхикозида порядка 90%.
Пример 8. Один литр ST-бульона в колбе Эрленмейера инокулиругот культурой штамма DSM 1670 из криоскопической пробирки. Колбы инкубируют в течение ночи при 30oС, 250 об/мин. Инокулят переносят в стерильных условиях в 14 л-ферментер, содержащий 9 л стерильного ST бульона, дополненного тиоколхицином до конечной концентрации 1 г/л. Ферментацию проводят, поддерживая подходящие уровни перемешивания-аэрации (перемешивание до 900 об/мин; перемешивание от 1 до 1,5 (об/мин) в зависимости от культурального роста). Каждые 2 ч отбирают образцы из культуральных бульонов и приводят следующие анализы:
- Оптическая плотность (OD) при 600 нм,
- Анализы на стерильность и чистоту штамма на LB агаре;
- Микроскопическая морфология (Gram-окрашенные);
- Анализ на содержание тиоколхикозида, ТСХ и ЖХВР.
После 28 часов ферментации трансформация в тиоколхикозид почти завершается. Конечный выход порядка 90%.
Пример 9. Повторяют способ, описанный в примере 8, но после 28 ч ферментации, но только 90% культуральных бульонов выделяют для экстракции продукта (фракция 1). Оставшиеся 10% стерильно вносят в ферментер с 9 л свежей стерильной ST среды, содержащей 10 г тиоколхицина. Ферментацию проводят, как описано в примере 8. Спустя 26 ч отбирают 9 л культуральных бульонов и экстрагируют (фракция 2). Оставшийся объем культуральных бульонов стерильно дополняют еще 9 л свежей стерильной ST среды, содержащей свежий тиоколхицин (10 г). Ферментацию проводят, как указано выше. Спустя 26 ч культуральный бульон полностью отбирают и экстрагируют (фракция 3). Биотрансформационная активность штамма остается стабильной во всех трех партиях с конверсионными выходами порядка 90% и по существу с трехкратным общим выходом тиоколхикозида по сравнению с выходом, получаемым в отдельном периодическом способе.
Пример 10. Конечный культуральный бульон из ферментации (общий объем: порядка 27 л) подвергают микрофильтрации в поперечном потоке, используя набор 0,22 мкм керамических картриджей (пластин) для фильтрования, с целью выделения клеток из бульона. Проникающие продукты (пермеаты) абсорбируются на колонке, заполненной HP 21, Mitsubishi абсорбционной смолой. После промывания водой продукт элюируют метанолом. Метанольный элюат концентрируют досуха в вакууме, затем повторно растворяют в метаноле. После повторных экстракций метиленхлоридом спиртовую фракцию концентрируют досуха и вновь растворяют в этанол-метиленхлориде, 1: 1 смеси. После очистки силикагелем раствор концентрируют в вакууме, затем метиленхлорид заменяют этанолом. Образовавшуюся суспензию концентрируют и оставляют кристаллизоваться. Повторную кристаллизацию с этанолом проводят после дополнительных стадий повторного растворения твердого продукта в смесях этанол-хлороформ и очистки на силикагеле.
Полученный продукт, проанализированный путем ЖХВР, С-ЯМР, Н-ЯМР и масс-спектроскопии оказался таким же, как стандартный тиоколхикозид.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИОТРАНСФОРМАЦИЯ КОЛХИЦИНОИДНЫХ СОЕДИНЕНИЙ | 2004 |
|
RU2346050C2 |
| СПОСОБ БИОТРАНСФОРМАЦИИ КОЛХИКОНОВОГО СОЕДИНЕНИЯ В СООТВЕТСТВУЮЩЕЕ 3-0-ГЛИКОЗИЛЬНОЕ ПРОИЗВОДНОЕ | 1998 |
|
RU2218409C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПРОИЗВОДНЫХ СТЕРОИДНЫХ ГЛИКОЗИДОВ RUSCUS ACULEATUS | 2000 |
|
RU2249043C2 |
| ПРОИЗВОДНЫЕ КОЛХИЦИНА И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ИХ ОСНОВЕ | 1996 |
|
RU2163241C2 |
| Штамм Bacillus megaterium 2-06-TS1 - продуцент кортизола | 2023 |
|
RU2817849C1 |
| ПРОИЗВОДНЫЕ ТИОКОЛХИЦИНА, СПОСОБЫ ИХ ПОЛУЧЕНИЯ, СОЕДИНЕНИЕ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1998 |
|
RU2190598C2 |
| Способ получения комплексного микробиологического препарата санитарно-гигиенического назначения | 2017 |
|
RU2654604C1 |
| СПОСОБ ГЛИКОЗИДИРОВАНИЯ КОЛХИЦИНА И ТИОКОЛХИЦИНА | 2009 |
|
RU2488589C2 |
| Штамм бактерий Bacillus megaterium, обладающий способностью продуцировать пробиотические и антимикробные вещества класса органических кислот | 2021 |
|
RU2757086C1 |
| ПРОИЗВОДНЫЕ КОЛХИЦИНА, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ НА ИХ ОСНОВЕ | 1997 |
|
RU2181354C2 |
Соединения колхициноидов трансформируют в соответствующие 3-О-гликозилпроизводные посредством штаммов Bacillus megaterium. Биотрансформацию проводят в ферментерах при аэрации с использованием глюкозы, фруктозы или глицерина в качестве источников углерода при 28-40oС и рН 6-7. Способ биотрансформации является специфическим и позволяет получать разнообразно замещенные колхицины с высокой степенью конверсии (до 95-100%). 4 з.п. ф-лы.
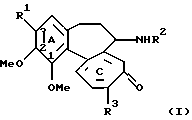
где R1 - остаток О-гликозида;
R2 - водород или С1-С7-ацил;
R3 - С1-С6-алкокси или С1-С6-тиоалкил,
включающий биотрансформацию производных колхицина формулы (I), в которой R1 - ОН или метокси, а R2 и R3 определены выше, посредством Bacillus megaterium.
| SOLET J.M | |||
| et al | |||
| Glucosylation of thiocolchicine by a cell suspension culture of Centella asiatica | |||
| Phytochemistry | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| FR 1344157 А, 21.10.1963 | |||
| POULEV et al | |||
| J | |||
| Ferment | |||
| Bioeng | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
