Настоящая заявка является частичным продолжением заявки на патент США рег. 08/656984, поданной 6 июня 1996 и находящейся в настоящее время на рассмотрении, и частичным продолжением заявки на патент США рег. 08/481130, поданной 7 июня 1995 и находящейся в настоящее время на рассмотрении, которая является частичным продолжением заявки на патент США рег. 08/245295, поданной 18 мая 1994 и находящейся в настоящее время на рассмотрении, которая в свою очередь является частичным продолжением заявки на патент США рег. 08/102852, поданной 5 августа 1993 и в настоящее время отозванной, которая является частичным продолжением заявки на патент США рег. 08/009266, поданной 22 января 1993 и в настоящее время отозванной, которая является частичным продолжением заявки на патент США рег. 07/894061, поданной 5 июня 1992 и в настоящее время отозванной, которая является частичным продолжением заявки на патент США рег. 07/889724, поданной 26 мая 1992 и в настоящее время отозванной, которая является частичным продолжением одновременно рассматриваемой заявки на патент США рег. 07/827689, поданной 27 января 1992 и в настоящее время отозванной.
Область, к которой относится изобретение
В общих чертах настоящее изобретение относится к молекулам клеточной адгезии, а более конкретно к клонированию и экпрессии ДНК, кодирующей не известный до настоящего времени полипептид, обозначенный "ICAM-4", который обладает структурным сходством с молекулами межклеточной адгезии ICAM-1, ICAM-2 и ICAM-R.
Предпосылки создания изобретения
Исследования, проводимые в последнее десятилетие, пролили свет на молекулярные события, сопутствующие межклеточным взаимодействиям в организме, а особенно на те события, которые вызывают миграцию и активацию клеток иммунной системы, при этом совсем недавно было выявлено, что они обеспечивают развитие и нормальную физиологическую функцию клеток нервной системы. См, в основном, работу Springer, Nature, 346: 425-434 (1990), относящуюся к клеткам иммунной системы, и работы Yoshihara et al., Neurosci. Res. 10: 83-105 (1991) и Sonderegger & Rathjien, J.Cell.Biol., 119: 1387-1394 (1992), относящиеся к клеткам нервной системы. Белки клеточной поверхности, а особенно так называемые молекулы клеточной адгезии ("CAMs") являются, соответственно, объектами фармацевтических исследований и разработок, целью которых является проникновение в процессы экстравазации лейкоцитов в места воспаления и миграции лейкоцитов в различные ткани-мишени, а также дифференцировку нейронов и образование сложной нервной сети. Выделение и характеризация молекул клеточной адгезии, клонирование и экпрессия ДНК-последовательностей, кодирующих эти молекулы, и получение терапевтических и диагностических агентов, влияющих на воспалительные процессы и развитие функции нервной системы, были также объектом многочисленных заявок США и иностранных патентных заявок. См. Edwards, Current Opinion in Therapeutic Patents, 1(11): 1617-1630 (1991), и в частности, опубликованные "патентные документы", цитируемые в настоящей заявке.
Из работ, предшествующих настоящей заявке, особый интерес представляет предварительная идентификация и характеризация некоторых медиаторов событий клеточной адгезии, "лейкоинтегринов", LFA-1, МАС-1, и gp 150,95 (обозначенных в номенклатуре WHO CD18/CD11a, CD18/CD11b, и CD18/CD11c, соответственно), которые образуют подсемейство гетеродимерных белков клеточной поверхности, "интегринов", присутствующих на В-лимфоцитах, Т-лимфоцитах, моноцитах и гранулоцитах. См. например, Таблицу 1 Springer, см. выше, на стр. 429. Представляют также интерес и другие одноцепочечные адгезивные молекулы (CAMs), участвующие в активации, адгезии, подвижности лейкоцитов и т.п., которые являются событиями, сопровождающими воспалительный процесс. Например, в настоящее время предполагается, что перед экстравазацией лейкоцитов, которая характеризует воспалительные процессы, происходит активация интегринов, конститутивно экспрессированных на лецкоцитах, с последующим прочным взаимодействием лиганд/рецептор между интегринами (например, LFA-1), и одной или обеих из двух различных молекул клеточной адгезии (ICAMs), обозначенных IСАМ-1 и ICAM-2, которые экспрессируются на поверхности эндотелиальных клеток кровеносных сосудов и на других лейкоцитах.
Подобно другим CAMs, охарактеризованным до настоящего времени [например, молекуле васкулярной адгезии (VCAM-1), описанным в РСТ WO 90/13300, опубликованной 15 ноября 1990; и молекуле клеточной адгезии эндотелиальных тромбоцитов (РЕСАМ-1), описанной в работе Newman et al., Science, 247: 1219-1222 (1991) и в РСТ WO91/10683, опубликованной 25 июля, 1991], IСАМ-1 и ICAM-2 являются структурно гомологичными другим членам суперсемейства генов иммуноглобулинов в той внеклеточной части, каждая из которых состоит из серии доменов, имеющих аналогичный карбоксиконцевой мотив. "Типичный" иммуноглобулинподобный домен содержит петлевую структуру, обычно "заякоренную" дисульфидной связью между двумя цистеинами на концах каждой петли. IСАМ-1 включает пять иммуноглобулинподобных доменов; ICAM-2, который отличается от IСАМ-1 по своему распределению в клетке, включает два таких домена; РЕСАМ-1 включает шесть доменов; a VCAM включает шесть или семь доменов в зависимости от вариантов сплайсинга, и т.п. Более того, обычно CAMs включают гидрофобную "трансмембранную" область, очевидно участвующую в ориентации молекулы на клеточной поверхности, и карбоксиконцевую "цитоплазматическую" область. Графические модели правильного расположения CAMs обычно показывают, что эта молекула заякорена в клеточной мембране в трансмембранной области цитоплазматическим "хвостом", простирающимся в цитоплазму клетки, и одним или несколькими иммуноглобулинподобными петлями, простирающимися кнаружи от клеточной поверхности.
Ряд нервных клеток экпрессирует на своей поверхности рецепторы с внеклеточными lg-подобными доменами, структурно схожими с ICAMs. Например, Yoshihara et al., см. выше. Помимо lg-подобных доменов, многие адгезивные молекулы нервной системы также содержат тандемно повторяющиеся фибронектинподобные последовательности во внеклеточном домене.
Был предложен ряд терапевтических применений молекул межклеточной адгезии, включая использование, основанное на предполагаемой способности IСАМ-1 связываться с риновирусом человека. Европейская патентная заявка 468257 А, опубликованная 29 января 1992, направлена, например, на разработку мультимерных конфигураций и форм IСАМ-1 (включая полноразмерные и усеченные молекулярные формы), которые, предположительно, обладают повышенной лиганд/рецепторсвязывающей активностью, особенно при связывании с вирусами, лимфоцитассоциированными антигенами и патогенами, такими как Plasmodium falciparum.
Аналогичным образом был предложен ряд применений для белков, иммунологически связывающихся с молекулами межклеточной адгезии. Заявка WO91/16928, опубликованная 14 ноября 1991, например, направлена на "очеловеченные" химерные антитела против IСАМ-1 и на их использование для лечения специфических и неспецифических воспалений, вирусных инфекций и астмы. В WO92/04034, опубликованной 19 марта 1992, описаны антитела против IСАМ-1 и их фрагменты, используемые для лечения эндотоксического шока. В заявке W092/06119, опубликованной 16 апреля 1992, описано ингибирование IСАМ-1-зависимого воспалительного ответа с использованием антиидиотипических антител против IСАМ-1.
Несмотря на фундаментальное представление о явлениях клеточной адгезии, которое было сформировано благодаря идентификации и характеризации белков внеклеточной алгезии, таких как IСАМ-1, и лимфоцитарные интерактивные интегрины, такие как LFA-1, картина этого явления далека от завершения. В основном, очевидно, что в воспалительных процессах и направленной миграции лимфоцитов по всему организму принимает участие множество других белков. Так, например, в заявке на патенты США рег. 07/827689, 07/889724, 07/894061 и 08/009266 и в соответствующей опубликованной заявке РСТ WO 93/14776 (опубликованной 5 августа 1993) описано клонирование и экспрессия белка, родственного ICAM, ICAM-R. Описание этих заявок вводится в настоящее описание посредством ссылки, а ДНК-последовательности и аминокислотные последовательности ICAM-R представлены в настоящем описании в SEQ ID No:4. Было обнаружено, что этот новый лиганд экспрессируется на лимфоцитах, моноцитах и гранулоцитах человека.
Особый интерес для настоящей заявки представляет тот факт, что была идентифицирована другая ICAM-подобная поверхностная молекула, которая, в отличие от любых известных молекул ICAM, имеет тканеспецифическую экспрессию. В работе Моri et al. , [Рrос. Natl. Acad. Sci. (USA) 84: 3921-3925 (1987)] сообщалось об идентификации специфичного для конечного мозга антигена головного мозга кролика, обладающего специфической иммунореактивностью с моноклональным антителом 271А6. Этот поверхностный антиген назван теленцефалином. В работе Imamura et al., [Neurosci. Letts. 119:118-121 (1990)] с использованием поликлонального антитела для оценки локализованной экспрессии, было высказано мнение, что экспрессия теленцефалина, при визуальном осмотре коры головного мозга кошек, обнаруживает изменения в слоях ткани, а также, что экспрессия рассматриваемого теленцефалина является функцией стадии развития. Затем, Oka et al., [Neuroscience 35:93-103 (1990)] сообщили о выделении теленцефалина с использованием моноклонального антитела 271А6. В этой публикации сообщается, что молекулярная масса этой поверхностной молекулы составляет около 500 кДа и что эта молекула состоит из четырех субъединиц, каждая из которых имеет нативную молекулярную массу 130 кДа и приблизительно 100 кДа после обработки N-гликаназой. Yoshihiro et al., [Neurosclence, Reserch Supplement 18, p.S83 (1994)] на 17-м Ежегодном заседании Японского общества по нейронаукам, проведенном 7-9 декабря 1993 в Нагое (Nagoya, Япония) и на 23-м Ежегодном заседании Общества по нейронаукам, проведенном 9 ноября 1993 в Вашингтоне D.C. [Society for Neuroscience Abstracts 19 (1-3) р.646 (1993)] сообщили кДНК-последовательность и аминокислотную последовательность для кроличьего теленцефалина. Сообщенная выведенная аминокислотная последовательность дает основание предположить, что 130 кДа-теленцефалин представляет собой интегральный мембранный белок с девятью внеклеточными иммуноглобулин(lg)-подобными доменами. Из этих доменов восемь дистальных доменов обнаружили гомологию с другими lg-подобными доменами ICAM. Те же самые сведения были сообщены Yoshihara et al., в Neuron 12: 543-553 (1994).
Таким образом очевидно, что специалистам необходимо продолжать поиск других белков, участвующих в межклеточных взаимодействиях в организме человека, а особенно необходимо получить информацию, которая может быть использована для специфической идентификации и характеризации таких белков с точки зрения их аминокислотной последовательности. Более того, такие молекулы, до определенной степени, могут служить основой для получения терапевтических и диагностических агентов, а главное, для выявления ДНК, кодирующих эти молекулы. Такая основополагающая информация должна обеспечить, inter alia, крупномасшабное продуцирование белков; идентификацию клеток, продуцирующих эти белки в природных условиях; и получение антител или новых специфически реагирующих с ними связывающих белков; и/или ингибирование реакций связывания лиганд/рецептор, в которых они участвуют.
Сущность изобретения
В одном из своих аспектов настоящее изобретение относится к очищенным и выделенным полинуклеотидам (например, к ДНК-последовательностям, РНК-транскриптам и их антисмысловым олигонуклеотидам), кодирующим новый полипептид "IСАМ-4", а также их новые полипептидные варианты (включая их фрагменты и аналоги с делециями, заменами и добавлениями), обладающие одной или более биологическими активностями связывания лиганд/рецептор, и/или иммунологические свойства, специфичные для IСАМ-4. Биологические ICAM-4-специфические активности связывания лиганд/рецептор охватывают взаимодействия как внеклеточного, так и цитоплазматического IСАМ-4 с другими молекулами (например, в процессах межклеточной адгезии и/или передаче сигнала). Предпочтительные ДНК-последовательности настоящего изобретения включают геномные и кДНК-последовательности, а также полностью или частично химически синтезированные ДНК-последовательности. Рассматриваемый в настоящей заявке предпочтительный полинуклеотид представлен в SEQ ID No:1 и кодирует крысиный вид ICAM-4. Рассматриваются биологические реплики выделенных ДНК-последовательностей настоящего изобретения (то есть копии ДНК-последовательностей, полученные in vitro или in vivo). Также получены автономно реплицирующиеся рекомбнантные конструкции, такие как плазмидные и вирусные ДНК-векторы, включающие последовательности ICAM-4, а особенно векторы, где ДНК, кодирующая IСАМ-4 или вариант IСАМ-4, правильно соединена с эндогенной или экзогенной ДНК-последовательностью, регулирующей экспрессию.
В соответствии с другим аспектом настоящего изобретения клетки-хозяева, особенно клетки одноклеточных хозяев, такие как прокариотические и эукариотические клетки, стабильно трансформируют ДНК-последовательности настоящего изобретения способом, обеспечивающим экспрессию в этих клетках нужных полипептидов.
Клетки-хозяева, экспрессирующие продукты IСАМ-4 и вариантов IСАМ-4, могут служить для различных целей. Учитывая то, что экспрессированные продукты "обнаруживаются" на поверхности клеток-хозяев, можно сказать, что эти клетки могут представлять собой ценный иммуноген для разработки антител, вступающих в специфические иммунные реакции с IСАМ-4 и вариантами IСАМ-4. Очевидно, что клетки-хозяева настоящего изобретения могут быть использованы в способах крупномасштабного продуцирования IСАМ-4 и вариантов IСАМ-4, где эти клетки выращивают в подходящей культуральной среде, и нужные полипептидные продукты выделяют из клеток и из среды, в которой эти клетки были культивированы.
Новые IСАМ-4 настоящего изобретения могут быть получены в виде изолятов из природных клеточных источников, но, наряду с продуктами ICAM-4-варианта, их, предпочтительно, продуцируют с помощью рекомбинантной техники, предусматривающей использование клеток настоящего изобретения. Рассматриваемая в настоящей заявке аминокислотная последовательность для полипептида IСАМ-4 представлена в SEQ ID No:2. Эти продукты могут быть получены полностью или частично в гликозилированной, частично или полностью в дегликозилированной или в негликозилированной форме в зависимости от клетки-хозяина, выбранной для рекомбинантного продуцирования, и/или от обработки после выделения. Варианты IСАМ-4 настоящего изобретения могут содержать водорастворимые или не растворимые в воде мономерные, мультимерные или циклические фрагменты IСАМ-4, которые включают все или часть одного или нескольких участков доменов, определенных выше и обладающих биологическими или иммунологическими свойствами IСАМ-4, включая, например, способность связываться с партнером связывания IСАМ-4 и/или ингибировать связывание IСАМ-4 с его природным партнером по связыванию. Варианты IСАМ-4 настоящего изобретения могут также содержать полипептидные аналоги, где одна или несколько специфических аминокислот делетированы или заменены: (1) без потери, а предпочтительно с усилением одной или нескольких биологических активностей или иммунологических свойств, специфичных для ICAM-4; или (2) без специфического повреждения конкретной функции связывания лиганд/рецептор. Рассматриваются также полипептиды-аналоги, включающие дополнительные аминокислотные (например, лизин или цистеин) остатки, которые способствуют образованию мультимера.
Настоящее изобретение также относится к антителам (например, моноклональным и поликлональным антителам, фрагментам антител, одноцепочечным антителам, химерным антителам, CDR-привитым антителам и т.п.) и к другим связывающим белкам (например, полипептидам и пептидам), которые являются специфичными для ICAM-4, или вариантов ICAM-4 (то есть не способными вступать в реакцию с молекулами межклеточной адгезии ICAM-1, ICAM-2 и ICAM-R, с которыми ICAM-4 имеет структурное сходство). Настоящее изобретение также относится к гибридомной клеточной линии, которая специфически секретирует моноклональные антитела настоящего изобретения. Предпочтительными гибридомами настоящего изобретения являются гибридомы, обозначенные 127А, 127Н, 173Е, 1791 и 179Н. Молекулы антител могут быть получены с использованием природных или рекомбинантных ICAM-4, или вариантов ICAM-4, или клеток, экспрессирующих такие продукты на своей поверхности. Связывающие белки настоящего изобретения, кроме того, могут быть использованы для характеризации структуры сайта связывания (например, эпитопов и/или восприимчивости связывающих свойств к модификациям в аминокислотной последовательности ICAM-4).
Связывающие белки могут быть использованы в свою очередь для иммунизации, а также для очистки полипептидов настоящего изобретения и идентификации клеток, имеющих эти полипептиды на своей поверхности. Очевидно также, что они могут быть использованы в модуляции (то есть блокировании, ингибировании или стимуляции) биологической активности связывания лиганд/рецептор, включая ICAM-4, а особенно тех эффекторных функций ICAM-4, которые участвуют в специфических и неспецифических иммунных системных ответах. Рассматриваются также антиидиотипические антитела, специфичные к молекулам антител против ICAM-4, и использование этих антиидиотипических антител для модуляции иммунного ответа. Кроме того, настоящее изобретение относится к способам выявления нейропатологии у индивидуумов, включающих стадии: а) получения жидкой пробы от индивидуума; b) контактирования этой пробы с антителом, являющимся специфически иммунореактивным для ICAM-4; с) количественной оценки уровня связывания ICAM-4/антитела в пробе; и d) сравнения уровня связывания ICAM-4/антитела в пробе с уровнем связывания ICAM-4/антитела у индивидуумов (контроли), о которых известно, что они не имеют нейропатологии. Анализы на детекцию и количественную оценку IСАМ-4 на клеточных поверхностях и в физиологической жидкости организма, таких как сыворотка или цереброспинальная жидкость, могут включать, например, одно антитело или множество антител в анализе формата типа "сэндвича". При детекции IСАМ-4 в физиологической жидкости организма антитела настоящего изобретения также могут быть использованы для оценки наличия нейропатологии, которая может коррелировать с повышенными уровнями IСАМ-4 в кровотоке. Такими нейропатологиями являются, но не ограничиваются ими, церебральная ишемия (т.е. шок), вызванная различными заболеваниями, включая, например, тромбоз, эмболию, церебральную геморрагию, вызванную аневризмой сосуда, вазоспазм и т.п. Количественная оценка циркулирующего IСАМ-4 позволяет также дифференцировать различные формы эпилепсии и, кроме того, позволяет определить стадии прогрессирования СПИД'а. Другими нейродегенеративными расстройствами, для диагностики которых могут быть использованы измерения циркулирующих IСАМ-4, являются различные формы болезни Альцгеймера и другие корковые деменции (такие как болезнь Пика, болезнь диффузных корковых телец Леви и дегенерация лобной доли) субкорковая деменция (включая болезнь Паркинсона, болезнь Гентингтона и прогрессирующее поражение надъядерной зоны), ряд первичных психических расстройств (таких как депрессия, шизофрения и психоз), а также приобретенная деменция, вызванная, например, инфекциями, васкулитами и алиментарными расстройствами (например, недостаточной функцией щитовидной железы, дефицитом витамина В12), сосудистыми расстройствами (обширным инфарктом, лакунарным состоянием, болезнью Бинсвангера), токсическим энцефалитом (например, воздействием моноокиси углерода, тяжелых металлов или другими промышленными загрязнениями) и опухолями.
Научное значение информации, получаемой благодаря определению ДНК и аминокислотных последовательностей, является очевидным. В качестве одного ряда примеров можно указать тот факт, что знание последовательности кДНК для ICAM-4 дает возможность путем ДНК/ДНК-гибридизации выделить геномные последовательности ДНК, кодирующие ICAM-4, и конкретно определить регуляторные последовательности, контролирующие экспрессию ICAM-4, такие как промоторы, операторы и т.п. Предполагается, что процессы ДНК/ДНК-гибридизации, осуществляемые с использованием ДНК-последовательностей настоящего изобретения в строгих условиях, являются теми процессами, которые позволяют выделить ДНК, кодирующие аллельные варианты ICAM-4, другие структурно родственные белки, имеющие одно или несколько общих биологических и/или иммунологических свойств, характерных для ICAM-4, и белки, гомологичные ICAM-4 от других видов. ДНК настоящего изобретения могут быть использованы в анализе методом ДНК/ДНК-гибридизации для выявления способности клеток синтезировать ICAM-4. Настоящее изобретение также относится к антисмысловым полинуклеотидам, подходящим для регуляции экспрессии ICAM-4 в тех клетках, которые обычно экспрессируют IСАМ-4. В качестве второго ряда примеров можно отметить, что знание ДНК и аминокислотных последовательностей IСАМ-4 дает возможность генерировать рекомбинантными методами варианты IСАМ-4, такие как гибридные слитые белки (называемые иногда "иммуноадгезионными"), характеризующиеся присутствием последовательностей белка IСАМ-4 и константных участков тяжелой цепи иммуноглобулина и/или шарнирных областей. См., Capon et al., Nature, 337: 525-531 (1989); Ashkenazi et al, , P.N.A.S (USA), 88: 10535-10539 (1991); и РСТ WO 89/02922, опубликованной 6 апреля 1989. Слитые белки вариантов ICAM-4 могут также включать, например, выбранные внеклеточные домены ICAM-4 и части других молекул клеточной адгезии.
ДНК настоящего изобретения также позволяет идентифицировать нетранслируемые ДНК-последовательности, которые специфически стимулируют экспрессию полинуклеотидов, правильно соединенных с промоторными областями. Идентификация и использование таких промоторных последовательностей являются особенно предпочтительными в случаях, например, переноса генов, когда может потребоваться специфическая экспрессия гетерологичных генов в ограниченных условиях среды окружающей нервной ткани. В настоящем изобретении также рассматриваются векторы, содержащие промоторы настоящего изобретения, а также химерные генные конструкции, которые включают промотор настоящего изобретения, правильно соединеннный с гетерологичной полинуклеотидной последовательностью и с сигналом терминации транскрипции.
Информация о ДНК-последовательности и об аминокислотных последовательностях, полученная с помощью настоящего изобретения, также дает возможность проводить систематические анализы структуры и функции IСАМ-4 и выявить те молекулы, с которыми этот белок взаимодействует на внеклеточном и внутриклеточном уровнях. Идиотипы моноклональных антител против IСАМ-4 настоящего изобретения являются репрезентативными для таких молекул и могут имитировать природные связывающие белки (пептиды и полипептиды), посредством которых модулируется внутриклеточная и внеклеточная активность IСАМ-4 или посредством которых IСАМ-4 модулирует внутриклеточные и внеклеточные события. Альтернативно они могут представлять новый класс модуляторов активности IСАМ-4. Антиидиотипические антитела в свою очередь могут представлять новый класс биологически активных эквивалентов ICAM-4. In vitro-анализ для идентификации антител и других соединений, которые модулируют активность IСАМ-4, может предусматривать, например, иммобилизацию ICAM-4 или природного лиганда, с которым связывается ICAM-4, детектируемое мечение неиммобилизованного партнера связывания, инкубирование партнеров связывания вместе и определение влияния тестируемого соединения на количество связанной метки, где уменьшение количества связанной метки в присутствии тестируемого соединения по сравнению с количеством связанной метки в отсутствие тестируемого соединения указывает на то, что тестируемый агент является ингибитором связывания ICAM-4.
Информация о ДНК-последовательности, представленная в настоящем изобретении, дает возможность разработать стратегии гомологичной рекомбинации или стратегии "нокаута" [см., например, Kapecchi, Science, 244: 1288-1292 (1989)] для грызунов, которые не экспрессируют функциональный белок ICAM-4 или которые экспрессируют вариантный белок ICAM-4. Такие грызуны могут быть использованы в качестве моделей для исследования активности ICAM-4 или модуляторов ICAM-4 in vivo.
Подробное описание изобретения
Описание родственной заявки на патент США рег. 08/102852, поданной 5 августа 1993, вводится в настоящее описание посредством ссылки. Примеры этой заявки направлены, inter alia, на получение и конструирование олигонуклеотидных зондов для PCR-амплификации ICAM- ДНК; использование этих зондов для амплификации человеческого геномного фрагмента, который гомологичен ДНК, кодирующим IСАМ-1 и ICAM-2, но отличается от этих ДНК; скрининг кДНК-библиотеки с использованием этого геномного фрагмента в целях выделения дополнительных ICAM-R-кодирующих последовательностей; скрининг кДНК-библиотек для выделения полноразмерной кДНК-последовательности, кодирующей ICAM-R; характеризацию данных о ДНК и аминокислотных последовательностях для ICAM-R, особенно последовательностей, родственных IСАМ-1 и ICAM-2; продуцирование клеток-хозяев млекопитающих, экспрессирующих ICAM-R; оценку критериев участия ICAM-R в событиях клеточной адгезии, включая СD18-зависимый и СD18-независимый пути метаболизма; ингибирование клеточной адгезии с ICAM-R пептидами, происходящими от ICAM-R; экспрессию вариантов ICAM-R; получение и характеризацию антител против ICAM-R и его фрагментов; картирование эпитопов ICAM-R, распознаваемых моноклональными антителами против ICAM-R; анализ распределения и биохимическую характеризацию ICAM-R и РНК, кодирующей ICAM-R; оценку ICAM-R на гомотипическую межклеточную адгезию и иммунную активацию/пролиферацию клеток; характеризацию моноклональных антител против ICAM-R; оценку дифференциального фосфорилирования и цитоскелетных ассоциаций цитоплазматического домена ICAM-R. Описана также идентификация ДНК, кодирующей IСАМ грызунов, которая, как считалось, является крысиным гомологом человеческого ICAM-R, и использование этой ДНК для конструирования и экспрессии ДНК, кодирующей слитые белки глутатион-S-трансферазы. Более подробное описание идентификации этой ДНК грызунов можно найти в родственной заявке (США рег. 08/102852) в Примере 6, и это описание изложено в настоящей заявке в Примере 1. По мере идентификации все больших частей ICAM-кодирующей последовательности грызунов стало очевидным, что ДНК ICAM грызунов не кодирует крысиный гомолог человеческого ICAM-R, а фактически кодирует новый полипептид ICAM, который в настоящем описании назван ICAM-4. Для выявления событий, которые привели к идентификации ICAM-4, ниже подробно описана хронология этих событий.
Сначала была идентифицирована геномная последовательность ICAM-4 грызунов, которая кодирует область, гомологичную домену 2 (в настоящем описании SEQ ID No:3, а в заявке США рег. 08/102852 SEQ ID No:23) человеческого ICAM-R (в настоящем описании SEQ ID No:4). Затем была также идентифицирована перекрывающаяся геномная ДНК (в настоящем описании SEQ ID No:5, а в заявке США рег. 08/102852, SEQ ID No:26), кодирующая область домена 2 SEQ ID No:3, и последовательность для IСАМ-1. С использованием SEQ ID No:3 в качестве зонда была идентифицирована кДНК селезенки грызунов (в настоящем описании SEQ ID No:6, а в заявке США рег. 08/102852, SEQ ID No:25), кодирующая домены 2-5, а также пятый домен, который ранее не наблюдался как домен ICAM. В это же самое время было установлено, что только что идентифицированные ДНКs грызунов кодируют гомолог грызунов для человеческого ICAM-R, однако сравнение первичных последовательностей 3'областей этих ДНКs с ДНК других ICAM оказалось достаточно трудным.
Последующее выделение 1 т.п.о. - кДНК-клона из библиотеки крысиной селезенки и амплификация RT-PCR-фрагмента, показала, что часть кДНК и геномного клонов не была секвенирована. Другой продукт RT-PCR-амплификации (SEQ ID No: 7) подтвердил это упущение. Было установлено, что фрагмент 177 п.о. был вырезан из геномного и кДНК-клонов путем EcoRI-гидролиза клонов и выделения этих последовательностей из фага λ для исследования ДНК путем секвенирования. Повторный анализ SEQ ID No:5 и 6 с рассмотрением этих других последовательностей позволил идентифицировать более точные и полные последовательности для первоначально выявленных геномных и кДНК-клонов, которые в их исправленной форме представлены в данном описании как SEQ ID No:8 и 9.
Для идентификации полной кодирующей последовательности для ICAM-4 была выделена кДНК головного мозга крысы (SEQ ID No:10) и была определена 5'-концевая последовательность путем быстрой 5'-амплификации концов кДНК (5'-RACE); продукт амплификации представлен в SEQ ID No:11. Суммарная информация, полученная для RT-PCR-клона (SEQ ID No:7), кДНК головного мозга (SEQ ID No: 10) и продукта RACE-амплификации (SEQ ID No:11), позволила идентифицировать полную кодирующую последовательность для ICAM-4(SEQ ID No:1).
Настоящее изобретение проиллюстрировано нижеследующими примерами. Более конкретно, в Примере 1 описано клонирование неполной ДНК ICAM-4. В Примере 2 описан Нозерн-блот-анализ транскрипции ICAM-4 грызуна. В Примере 3 описано выделение полноразмерной кДНК ICAM-4 грызуна. Пример 4 относится к in situ-гибридизации ICAM-4 в ткани головного мозга. В Примере 5 описано продуцирование слитых белков ICAM-4 в прокариотах. В примере 6 описано продуцирование моноклональных антител, специфичных для гибридных белков "крысиный ICAM-4/GST". В Примере 7 описана экспрессия растворимых белков крысиных ICAM-4 в бакуловирусной экспрессирующей системе. В Примере 8 описано продуцирование моноклональных антител, специфичных для крысиного ICAM-4, экспрессированного в бакуловирусной системе. В Примере 9 описан иммуноцитохимический анализ экспрессии крысиного ICAM-4. В Примере 10 описано клонирование геномной ДНК, кодирующей ICAM-4 человека. В Примере 11 описано клонирование кДНК, кодирующей ICAM-4 человека. В Примере 12 описан Нозерн-блот-анализ экспрессии ICAM-4 человека. В примере 13 описано продуцирование гибридных белков "ICAM-4 человека/GST". В Примере 14 описано продуцирование моноклональных антител, иммуноспецифичных для ICAM-4 человека. В Примере 15 описана разработка анализа методом "захвата" для определения концентрации растворимого ICAM-4 в индивидуальной жидкости. В примере 16 описан метод анализа методом захвата, используемый для оценки концентрации ICAM-4 в сыворотве пациентов с инсультом. Пример 17 относится к оценке транскрипции ICAM-4 в крысиной модели эпилепсии. В Примере-18 описано измерение концентрации циркулирующего ICAM-4 для оценки различных нейродегенеративных расстройств. В Примере 19 описано клонирование промоторной области для ICAM-4 человека.
ПРИМЕР 1
Клонирование ДНК для IСАМ крысы
А. Выделение геномной ДНК для домена 2 IСАМ крысы
Крысиную геномную библиотеку, сконструированную в λ EMBL3, скринировали [32Р]-меченным зондом, генерированным с помощью PCR из ДНК, кодирующей домен 2 IСАМ-3 человека. Последовательность этого зонда представлена в SEQ ID No: 12. Бляшки библиотеки переносили на найлоновые мембраны Hybond N+ (Amersham, Arlington Heights, IL). Скринирование всех ДНК и геномных библиотек осуществляли в соответствии со стандартными протоколами. Предварительную гибридизацию и гибридизацию проводили в растворе 40-50% формамида, 5х Денхардта, 5Х SSPE и 1,0% ДСН при 42oС. Зонды ([32P]-меченные) добавляли в раствор для гибридизации в концентрации 105-106 об/мин/мл. После проведения гибридизации в течение 16-18 часов найлоновые мембраны интенсивно промывали при комнатной температуре в 2Х SSPE с 0,1% ДСН, а затем экспонировали с рентгеновской пленкой в течение ночи при -80oС. Для получения клонального фага позитивные бляшки подвергали одному или нескольким циклам гибридизации. ДНК, полученную из лизата позитивных клонов, субклонировали в pBS+ и секвенировали.
Первый геномный клон, кодирующий домен 2 крысиного IСАМ, идентифицировали на установление его гомологичности по отношению к участкам домена 2 в других членах семейства IСАМ (см., например, Таблицу 1 заявки на патент США рег. 08/102852), который все же отличается от ранее установленных нуклеотидных последовательностей для крысиного ICAM-1 [Kita, et al., Biochem. Biophys. Acta 1131: 108-110 (1992)] или мышиного ICAM-2 [Xu et al., J. Immunol. 149; 2560-2565 (1992)]. Последовательности нуклеиновой кислоты и выведенные аминокислотные последовательности для этого клона описаны в одновременно рассматриваемых заявках, родственных настоящей заявке, как соответствующие вариантные формы крысиного ICAM-R, и были представлены как SEQ ID No:23 и 24, соответственно, в заявке США 08/102/852. В данной заявке эти же самые последовательности представлены в SEQ ID No:3 и 13, соответственно.
Второй перекрывающийся клон был также идентифицирован с использованием тех же зондов и оценен на присутствие в нем последовательности домена 2 ICAM SEQ ID No:3 и 5'-ДНК, кодирующей, по крайней мере, часть крысиного ICAM-1. Нуклеиновокислотная последовательность для этого клона представлена в одновременно рассматриваемой родственной заявке как SEQ ID No:26, а в настоящей заявке она представлена как SEQ ID No:5. Этот второй клон указывает на то, что фрагмент гена ICAM первого клона и ген, кодирующий крысиный ICAM-1, расположены на той же самой крысиной хромосоме на расстоянии друг от друга в 5 т.п.о.
В. Выделение кДНК для ICAM крысы
Для идентификации более полной белок-кодирующей последовательности для полипептида IСАМ [32Р]-меченную ДНК, кодирующую последовательность домена 2 из крысиного геномного клона, идентифицированного в разделе A (SEQ ID No:3, см. выше), использовали для скрининга ряда библиотек кДНК от различных типов клеток крысы и мыши, включая макрофаг (Clontech, Palo Alto, CA), лимфоцит периферической крови (PBL)(Clontech), Т-клетки (сконструированные на месте) и селезенку (Clontech) крысы, и PBL (Clontech), Т-клетки (сконструированные на месте) и В клетки (сконструированные на месте) мыши.
Единичный клон был идентифицирован в библиотеке кДНК селезенки крысы (Clontech), которая содержала пять lg-подобных доменов, четыре из которых были гомологичны доменам 2-5 как в IСАМ-1, так и в ICAM-R. Более того, этот клон включал 3'ДНК, кодирующую очевидный пятый lg-подобный домен, который до этого не был идентифицирован в каком-либо другом полипептиде ICAM. Кроме того, этот клон содержал необычную 3'последовательность, которая позднее была определена как неполный интрон (обсуждаемый ниже), локализованный между доменами 4 и 5, что дает основание предположить, что этот клон является продуктом незрелого или неправильно сплайсированного транскрипта. Присутствие этого уникального домена и определение того, что 3' область не имеет точного сходства с другими известными ICAM, наводит на мысль о том, что эта относящаяся к ICAM ДНК возможно кодирует новый крысиный полипептид ICAM. Нуклеиновокислотная последовательность для этого клона представлена в заявке, родственной настоящей заявке, как SEQ ID No:25; а в настоящей заявке нуклеиновокислотная последовательность для этого клона кДНК селезенки представлена в SEQ ID No:6.
С. поворотный анализ кДНК и геномной кДНК крысы
Согласно заявке на патент США рег. 08/102852, поданной 5 августа 1993 г. , было определено, что в неполном клоне кДНК селезенки крысы (SEQ ID No:25 в родственной заявке и SEQ ID No:6 в настоящей заявке) и в геномном клоне печени крысы (SEQ ID No:26 в родственной заявке, и SEQ ID No:5 в настоящей заявке) отсутствует внутренний EcoRI-фрагмент в 177 п.о, который являлся частью каждого из этих клонов, но был утерян при осуществлении стадии субклонирования, когда вставки библиотеки были удалены из вектора λ путем EcoRI-гидролиза и лигированы в секвенирующий вектор. Тот факт, что кДНК и геномные клоны могут терять кодирующий фрагмент, стал очевидным после сравнения первичной геномной последовательности и кДНК-последовательности крысы с различными продуктами RT-PCR-амплификаци, включая SEQ ID No:7, которая обнаруживает брешь в крысиной последовательности.
Последующее выделение и сравнение первичных последовательностей кДНК из библиотеки селезенки с использованием кДНК-клона селезенки (SEQ ID No:6) в качестве зонда было первым свидетельством того, что часть кДНК селезенки и геномных клонов не была секвенирована. Дальнейшее подтверждение этого вывода появилось после амплификации RT-PCR-фрагмента, охватывающего домены 3-5, с использованием 5'-праймера (RRD3 5' Xho, содержащий рестрикционный 5'-Хлоl-сайт для облегчения клонирования), представленного в SEQ ID No:14, и 3'-праймера (RRD5 3' HindIII, содержащий рестрикционный НindIII-сайт для облегчения клонирования), представленного в SEQ ID No:15.
GAACTCGAGGCCATGCCTCCACTTTCC (SEQ ID No:14) \
CCATAAGCTTTATTCCACCGTGACAGCCAC (SEQ ID No:15)
Сравнение первичных последовательностей этих двух ДНК ясно показало, что в кДНК- и геномном клонах был потерян фрагмент перед секвенированием; этот факт был еще раз подтвержден после секвенирования ДНК с помощью RT-PCR, как обсуждается ниже. Был сделан вывод, что рестрикционный гидролиз ферментом EcoRI для удаления кДНК и геномных фрагментов перед секвенированием приводит к вырезанию 177 п.о.-фрагмента, который не был обнаружен визуально при разделении в агарозном геле клонов от последовательностей фага λ. Последующий анализ последовательности подтвердил локализацию двух EcoRI-сайтов, фланкирующих 117 п.о.-фрагмент в обоих исходных клонах.
EcoRI-177 п.о.-фрагмент расположен между нуклеотидами 719 и 896 в неполном крысином кДНК-клоне, показанном в SEQ ID No:9, и между нуклеотидами 2812 и 2989 в неполном крысином кДНК-клоне, показанном в SEQ ID No:8.
D. ДНК, выделенная путем RT-PCR-клонирования
Для получения более полной информации для гена, относящегося к ICAM крысы, использовали RT-PCR. Информацию о последовательности геномного клона (SEQ ID No: 3) использовали для конструирования смысловых праймеров, комплементарных 5' участку белок-кодирующей области, как было определено из кДНК-клона, и антисмысловых праймеров, комплементарных кодирующим последовательностям и 3'-участкам до кодирующей последовательности в кДНК-клоне (SEQ ID No:6).
кДНК-матрицу для PCR-реакций получали следующим образом. Приблизительно 2 мкг poly A+PHK, выделенной из клеток крысиной селезенки, денатурировали путем нагревания при 65oС в объеме 10 мкл. После денатурации добавляли 0,1 мкл РНКазина (Invitrogen, San Diego, CA), 5 мкл 5Х буфера RT (BRL, Bethesda, MD), 2 мкл рандомизированного гексамера (pd(N)6 при 100 мкг/мл) (Pharmacia, Piscataway, NJ), 6 мкл dNTP (2 мМ каждого) и 2 мкл обратной транскриптазы AMV (BRL) и реакцию инкубировали в течение 60-90 минут при 42oС. Реакционные смеси хранили при -20oС, до тех пор, пока они не потребуются.
Предварительные серии экспериментов проводили в целях идентификации пар олигонуклеотидных праймеров, продуцирующих продукт амплификации в PCR-реакциях с использованием кДНК селезенки крысы в качестве матрицы. Различные 5' смысловые праймеры в PCR были использованы в паре с 3' праймером, который был сконструирован так, чтобы он был комплементарен внутренней кодирующей последовательности; 3' праймер был обозначен RRD2 3-1 и представлен в SEQ ID No:16.
AACGTGCGGAGCTGTCTG (SEQ ID No: 16) (В конечном выделенном RT-PCR-продукте, SEQ ID No:7, см. ниже, праймер RRD2 3-1 соответствовал нуклеотидам 719-736). Аналогичным образом различные 3' антисмысловые праймеры были использованы в паре с 5' праймером, который был сконструирован так, чтобы он был комплементарен другой внутренней кодирующей последовательности, где 5' праймер в этих реакциях был обозначен RGen3900S и представлен в SEQ ID No: 17.
ACGGAATTCGAAGCCATCAACGCCAGG (SEQ ID No:17)
(В SEQ ID No:7, см. ниже, праймер RGen3900S соответствовал нуклеотидам 1719-1736). Исходя из размеров продуктов амплификации и способности этих продуктов к гибридизации с неполным кДНК-клоном одна пара праймеров была определена как наиболее эффективная и была использована в последующих PCR-амплификациях. 5'-праймер был обозначен RGen780S (SEQ ID No:18), а 3'-праймер был обозначен RGen4550S (SEQ ID No:19):
CATGAATTCCGAATCTTGAGTGGGATG (SEQ ID No:18)
ATAGAATTCCTCGGGACACCTGTAGCC (SEQ ID No:19)
(в SEQ ID No:7, см. ниже, праймер RGen780S соответствовал нуклеотидам 1-18, а праймер RGen4550AS соответствовал нуклеотидам 2197-2214).
Эта пара праймеров была использована в PCR в ряде условий для оптимизации амплификации. Все из 15 различных PCR-буферов, которые варьировались по рН и концентрации Мg++, были использованы при двух различных температурах отжига, а образец продукта от каждой реакции разделяли в 1% агарозном геле. Поскольку продукт амплификации не мог быть обнаружен путем визуального наблюдения окрашенного этидийбромидом геля исходя из каких-либо условий реакции, то для обнаружения PCR-продуктов была применена более чувствительная Саузернгибридизация.
Аликвоты амплифицированной ДНК были получены путем электрофореза, перенесены на найлоновую мембрану Hybond N+ капиллярным методом стандартного Саузернблоттинга, и гибридизованы с полной кДНК крысы, которая была помечена [32P]. Условия гибридизации были, в основном, аналогичными условиям, описанным для процедуры скрининга библиотеки в Разделе А, см. выше. Авторадиография показала, что в двух из этих реакций было генерировано небольшое количество ДНК приблизительно в 2,2 т.п.о., а остальная часть продукта амплификации, полученная в двух реакциях, была выделена на агарозном геле. 2,2 т.п. о. -область была элюирована из геля, даже несмотря на то, что после визуального осмотра не было обнаружено какой-либо полосы, и использована в качестве матрицы в другой PCR-реакции с использованием тех же самых праймеров (SEQ ID No: 18 и 19), буфера Трис-HCl, рН 8,0, содержащего 1 мМ Мg++, и температуры отжига 55oС. Продукт амплификации от вторичной PCR был визуально обнаружен в геле и был элюирован и клонирован в плазмиду pBS+ (Stratagene, La Jolla, CA) для анализа последовательности.
Полученный RT-PCR-клон оценивали на содержание 2214 п.о., представленных в SEQ ID No:7. Этот клон кодировал домены 2-6, обнаруженные в кДНК-клоне селезенки крысы, дополнительный аминоконцевой домен 1, дополнительный карбоксиконцевой домен 7, и 164 п.о., которыми очевидно является другой карбоксиконцевой домен 8. Непосредственно 5'-последовательностью до домена 1 была дополнительная 144 п. о.-последовательность, предположительно происходящая от интрона, находящегося между лидерной последовательностью и первым доменом. Этот клон не содержал 5'-лидерную последовательность или 3'трансмембранные и цитоплазматические области. Помимо предварительно идентифицированного домена 6 в клоне кДНК селезенки крысы, присутствие 7-ого и 8-ого доменов в RT-PCR-клоне подтвердило гипотезу о том, что этот клон представляет собой новый ICAM грызунов.
ПРИМЕР 2
Нозерн-блот-анализ
Для последующего изучения вероятности того, что ICAM-клоны, идентифицированные в Примере 1, кодируют новый полипептид ICAM, как было предположено благодаря идентификации уникальных lg-подобных доменов, была оценена тканеспецифическая экспрессия с помощью Нозерн-блот-анализа для сравнения с ранее описанными характерами экспрессии человеческих ICAM [ICAM-1, Dustin, et al. , J. Immunol, 137: 245-254 (1986); ICAM-2; Staunton, et al. Nature 339: 61-64 (1989); ICAM-R, de Fourgerolles and Springer, J.Exp.Med. 175: 185-190 (1992)].
Полную клеточную РНК, происходящую от легких, головного мозга, позвоночника, печени, пищеварительного тракта, тимуса, лимфатических узлов и селезенки крысы, получали с использованием реагента для выделения РНК STAT60 (Tel-тест "В", Inc, Friendswood, Texas) в соответствии с инструкциями производителя. poly(A)+PHK очищали из полной РНК на колонках с олиго-dТ-целлюлозой. Приблизительно 5 мкг РНК, происходящей от каждой ткани, выделяли на формальдегидсодержащем 1% агарозном геле и переносили на нитроцеллюлозные мембраны Hybond-C (Amersham).
Фрагмент кДНК селезенки крысы, полученный как описано в Примере 1, соответствующий доменам 2-4 (нуклеотиды 1-724 в SEQ ID No:6), субклонировали в pBluescript SK+ (Stratagene), и антисмысловой рибозонд генерировали путем in vitro-транскрипции с использованием 32Р-меченного UTP и приблизительно 500 нг линеаризованной матрицы в соответствии с инструкцией, предлагаемой производителями (Boehringer Mannhiem, Indianapolis, IN). Мембрано-ассоциированную РНК предварительно гибридизовали в растворе, содержащем 50% формамид, 5X SSC, 1Х РЕ (50 мМ Трис-HCl, рH 7,5, 0,1% пирофосфат натрия, 0,2% поливинилпирролидон, 0,2% фиколл, 5 мМ EDTA, 1% ДСН), и 150 мкг/мл денатурированной ДНК спермы лосося. Радиоактивный зонд денатурировали путем кипячения и добавления в раствор для предварительной гибридизации до конечной концентрации 1•106 имп./мл. Гибридизацию проводили в течение 16-18 часов при 65oС. Затем мембраны промывали при 65oС в 2X SSC, содержащем 0,1% ДСН, а затем экспонировали с рентгеновской пленкой в течение 3-16 часов.
Нозерн-блот-анализ показал, что ICAM-кДНК, идентифицированная в Примере 1, экспрессировалась только в головном мозге крысы, причем о тканеспецифичности каких-либо других полипептидов ICAM ранее не сообщалось. Эта картина экспрессии, в комбинации с уникальными lg-подобными доменами, ранее не наблюдалась в полипептидах ICAM, что свидетельствует о том, что данный ICAM-клон является новым членом семейства белков ICAM, и этот новый клон был обозначен ICAM-4.
Тот факт, что первоначально идентифицированные кДНК-клоны были детектированы в библиотеке селезенки крысы, дает основание предположить, что субпопуляция клеток в селезенке может экспрессировать ICAM-4 на низких уровнях. Однако правильно сплайсированный клон не мог быть детектирован в многочисленных гемопоэтических кДНК-библиотеках, что внушало сомнение относительно того, действительно ли белок ICAM-4 экспрессируется в других тканях, кроме головного мозга. Одним из объяснений детекции кДНК ICAM-4 в селезенке может служить то, что чувствительность PCR позволяет амплифицировать следовые количества транскрипта, даже если эти ткани не экспрессируют кодированный белок.
ПРИМЕР 3
Выделение полноразмерной кДНК IСАМ-4 крысы
А. Идентификация кДНК-клона головного мозга крысы
Исходя из тканеспецифической экспрессии IСАМ-4, в целях выделения полноразмерной кДНК, кодирующей IСАМ-4, была использована мРНК ткани головного мозга. Два зонда, один из которых комплементарен доменам 1-2, а второй комплементарен доменам 3-5 клона кДНК селезенки, идентифицированным в Примере 1 (SEQ ID No:7), были радиоактивно помечены и использованы для скрининга библиотеки кДНК головного мозга крысы в λgt10, которая была предварительно сконструирована на месте. Условия гибридизации были такими, как они описаны в Примере 1, а позитивные бляшки подвергали одному или нескольким циклам скрининга с получением клонального фага.
Было идентифицировано девять позитивных клонов, два из которых гибридизовались с обоими зондами. Самый длинный из этих двух клонов, обозначенный клоном 7, содержал 2550 п.о., кодирующие четыре из пяти lg-подобных доменов, обнаруженных в кДНК зонда. Помимо этого, клон 7 кодировал четыре других lg-подобных домена, которые не были обнаружены в этом зонде. Были идентифицированы предполагаемые трансмембранные и цитоплазматические домены, за которыми следовали стопкодон, сигнал полиаденилирования и poly-A-хвост. В клоне 7 отсутствовал, по крайней мере, один 5' lg-домен, как было определено путем сравнения с RT-PCR-клоном (SEQ ID No:7), а также отсутствовала лидерная последовательность; при этом повторное секвенирование
библиотеки не давало каких-либо более длинных клонов, которые содержали бы эти последовательности. Нуклеотидтная последовательность для клона 7 представлена в SEQ ID No: 10.
В. Определение 5'-конца
Для выделения домена 1 и других 5' последовательностей была применена PCR-техника, названная быстрой амплификацией 5'-концов кДНК (RACE= Rapid Amplification of cDNA Ends) [PCR Protocols: A Guide to Methods and Application, Innis, et al., (eds) Academic Press:
New York (1990) pp:28-38], с использованием набора 5'-RACE (Clontech). Эта техника позволяет использовать внутренний праймер, спаренный со вторым праймером, комплементарным последовательности адаптора, лигированного с 5'-концом молекул библиотеки кДНК. Следовательно, PCR с использованием этой пары праймеров будет амплифицировать и облегчать идентификацию интронов. Затем данные о перекрывающейся последовательности могут быть использованы для генерирования полной последовательности гена.
Уже готовую кДНК, полученную из головного мозга крысы с помощью RACE (поставляемую вместе с набором), использовали в PCR с олигонуклеотидом, имеющимся в наборе, и антисмысловым праймером, полученным на основе внутренней последовательности ICAM-4. 3'-антисмысловой праймер, обозначенный Spot714AS, был сконструирован в соответствии с последовательностью домена 4 ICAM-4 и представлен в SEQ ID No:20.
CARGGTGACAAGGGCTCG (SEQ ID No:20)
Продукт амплификации, полученный из этой пары праймеров, затем подвергали вторичной PCR с использованием того же самого 5'-праймера из набора, спаренного с 3'-праймером, комплементарным области в домене 1 ICAM-4. Второй 3'-праймер был обозначен RRACE2 и представлен в SEQ ID No:21.
TATGAATTCAGTTGAGCCACAGCGAGC (SEQ ID No:21)
Для облегчения клонирования полученного продукта амплификации в pBS+(Stratagene) каждый праймер, используемый во вторичной PCR, содержал ЕсоRI-сайт. Полученную плазмидную ДНК, которая содержала 5'-конец гена, идентифицировали путем гибридизации с зондом доменов 1 и 2 ICAM-4 крысы, соответствующим нуклеотидам 1-736 в SEQ ID No:7. Данные о неполной последовательности для домена 1 и гиброфобной лидерной последовательности определяли исходя из полученного продукта амплификации.
Продукт, полученный методом 5' RACE, представлял собой ДНК-фрагмент длиной в 222 п.о., содержащий выше (в направлении 3'-->5') 60 п.о. инициирующего метионинового остатка, 82 п.о. лидерной последовательности и 80 п.о. последовательности домена 1. Продукт амплификации представлен в SEQ ID No: 11.
С. Полноразмерная последовательность ICAM-4 крысы
Составной клон полноразмерного ICAM-4 был сконструирован исходя из данных о последовательности, полученной методом 5'-RACE (SEQ ID No:11), RT-PCR-клона (SEQ ID No:7) и клона 7 кДНК головного мозга (SEQ ID No:10). Было определено, что полноразмерный ген для ICAM-4 крысы содержит 2985 п.о. с одной открытой рамкой считывания, кодирующей выведенный белок из 917 аминокислот. Предполагаемая последовательность Kozak расположена выше метионинового остатка в лидерной последовательности. За гидрофобной лидерной последовательностью из 27 аминокислот следуют девять lg-подобных доменов, трансмембранная область и цитоплазматический хвост из 58 аминокислот. Составная кДНК IСАМ-4 представлена в SEQ ID No:1, а выведенная аминокислотная последовательность представлена в SEQ ID No:2.
Подобно другим полипептидам ICAM, ICAM-4 содержит внеклеточный, трансмембранный и цитоплазматический домены. Во внеклеточном домене аминоконец ICAM-4 представляет собой лидерную последовательность, содержащую аминокислоты 1-27, за которой следуют девять иммуноглобулин(lg)-подобных доменов, что является характерным уникальным признаком для ICAM-4, если учесть, что ICAM-1, ICAM-2 и ICAM-R содержат пять, два и пять внеклеточных lg-подобных доменов соответственно. В ICAM-4, домен 1 содержит аминокислоты 28-118; домен 2 содержит аминокислоты 119-224; домен 3 содержит аминокислоты 225-321; домен 4 содержит аминокислоты 322-405; домен 5 содержит аминокислоты 406-488; домен 6 содержит аминокислоты 489-569; домен 7 содержит аминокислоты 570-662; домен 8 содержит аминокислоты 663-742; а домен 9 содержит аминокислоты 743-830. Внутри каждого домена характерная структура "петли" образована дисульфидной связью между цистеиновыми остатками, расположенными, в основном, на противоположных концах аминокислотной последовательности домена. Другими структурными особенностями ICAM-4 является трансмембранная область, содержащая аминокислоты 831-859, и цитоплазматическая область, содержащая аминокислоты 860-917.
Сравнение гомологии аминокислотной последовательности каждого домена ICAM-4 крысы с другими членами семейства ICAM было ограничено соответствующими последовательностями ICAM-1, IСАМ-2 и ICAM-R человека, поскольку данные о последовательности для всех трех гомологов грызунов ранее не сообщались. В первом домене IСАМ-4 грызунов обнаруживал 21%-, 30%-, и 28%-ную идентичность с ICAM-1, ICAM-2 и ICAM-R человека соответственно. Второй домен был более консервативным, причем, процент аминокислотной идентичности составлял 60, 42 и 62 по отношению к ICAM-1, -2, и -3 соответственно. Домены 3-5 показали 48%, 49%, и 40%-ную идентичность с ICAM-1 и 60, 59, и 29 для ICAM-R соответственно. Интересно отметить, что домены 6-8 крысиного ICAM-4 являются наиболее гомологичными домену 5 (идентичность в пределах 29-42%), что вероятно вызвано дупликацией генного сегмента. Девятый и последний внеклеточный домен имеет плохое сходство с другими доменами ICAM, но имеет 22%-ную идентичность с 3-м и 6-м доменами VCAM-1 человека, другим членом lg-семейства белка, который участвует в клеточной адгезии. Цитоплазматический хвост имеет длину в 58 аминокислот. Он является более длинным, чем другие члены семейства ICAM, где человеческие ICAM-1, -2, и -3 содержат 28, 26 и 37 аминокислот соответственно. Как и в случае девятого домена, цитоплазматический хвост крысиного ICAM-4 является наиболее гомологичным цитоплазматическому хвосту VCAM-1 человека и содержит только 19 аминокислот. Мембранные проксимальные 19 аминокислот крысиного ICAM-4 имеют общие 7 аминокислотных остатков с VCAM-1 (37%).
И наконец, было картировано функциональное связывание с LFA-1 (CD11a/CD18) для первого домена в ICAM. Vonderheide et al., [J.Cell. Вiоl., 125: 21 5-222 (1994)] идентифицировали мотив последовательности, играющий соответствующую роль в связывании с интегрином. Несмотря на то, что в домене 1, крысиный ICAM-4 и другие ICAM имеют относительно низкую гомологию, этот мотив последовательности связывания является консервативным, что дает основание предположить, что крысиный ICAM-4 может служить лигандом для LFA-1 и, вероятно, других интегринов.
ПРИМЕР 4
In situ-гибридизация в ткани головного мозга
Для локализации участка специфической ткани головного мозга, который экспрессирует ICAM-4, была проведена in situ-гибридизация с антисмысловыми рибозондами для домена 1 ICAM-4 и для доменов 3-4 ICAM-4. Эти зонды были помечены путем in vitro-транскрипции с использованием 35S-меченного UTP.
Замороженные срезы ткани нормального головного мозга крысы фиксировали в 4% параформальдегиде в течение 20 минут, промывали и дегидратировали, и фиксированную РНК денатурировали в течение 2 минут в 2•SSC, 70% формамиде при 70oС перед началом гибридизации. Срезы ткани гибридизовали в течение ночи при 50oС в растворе, содержащем 50% формамид, 0,3 М NaCl, 20 мМ Трис-HCl, рН 7,4, 5 мМ EDTA, 10% сульфат декстрана, 1 • раствор Денхгардта, 0,5 мг/мл дрожжевой РНК, 100 мМ DTT и зонд в концентрации 50000 имп./мкл. Предметные стекла один раз промывали в 4Х SSC, 10 мМ DTT при комнатной температуре в течение 60 минут, один раз в 50% формамиде, 2Х SSC, 10 мМ DTT при 60oС в течение 40 минут и один раз в каждом из 2Х SSC и 1Х SSC в течение 30 минут при комнатной температуре. Специфичность гибридизации определяли в параллельных экспериментах, проводимых в соответствии с тем же самым протоколом, но с использованием также более жесткой промывки в 50% формамиде, 1•SSC, 10 мМ DTT при 60oС в течение 40 минут. После промывки предметные стекла опускали в эмульсию NTB2 (Kodak, Rochester, NY) и экспонировали в течение периода времени от 2 до 21 дня, после чего проявляли и подвергали контрастному окрашиванию. Помимо рибозонда вируса иммунодефицита человека (ВИЧ-1) в качестве негативных контролей использовали смысловые зонды, генерированные из домена 1 ICAM-4, и смысловые рибозонды, генерированные из доменов 3-4 ICAM-4.
Сигнал, детектированный в ткани головного мозга, находился, главным образом, в сером веществе, причем наиболее сильный сигнал детектировался в коре головного мозга и гиппокампе. Профиль гибридизации соответствовал экспрессии ICAM-4, главным образом, в нейронах головного мозга.
ПРИМЕР 5
Генерирование гибридных белков ICAM-4
Для продуцирования моноклональных антител против специфических фрагментов полипептида ICAM-4 гибридные белки "ICAM-4 крысы/глутатион-S-трансфераза" (GST) были генерированы с использованием экспрессирующего вектора прокариотов pGEX (Pharmacia, Alameda, CA).
Для амплификации ДНК-фрагментов, кодирующих отдельные домены, были использованы PCR-праймеры, соответствующие 5'- и 3'-концам домена 1 и 5'- и 3' -концам домена 2. Полученные фрагменты отдельно клонировали в EcoRI-сайт pGEX-2T; анализ ДНК-последовательности подтверждал наличие правильной ориентации и рамки считывания. Затем трансформанты были скринированы на их способность продуцировать гибридный белок с соответствующей молекулярной массой.
После того как бактерии были лизированы путем обработки ультразвуком в PBS, содержащем 1% ДСН, оба гибридных белка "домен 1 ICAM-4/GST" и "домен 2 ICAM-4/GST" оставались в нерастворимой фракции. Нерастворимую белковую фракцию из 100 мл культуры кипятили в загрузочном красителе с ДСН и разделяли в 10% препаративном полиакриламидном геле с ДСН. Этот гель окрашивали в охлажденном льдом 0,4 М KCl и полосы гибридного белка вырезали. Гибридные белки подвергали электроэлюции из гелевых срезов в трубках для диализа, в буфере, содержащем 25 мМ Трис-НСl и 192 мМ глицин. Приблизительную концентрацию белка определяли по ОП280, а чистоту препарата определяли в ПААГ-ДСН, окрашенном кумасси синим.
ПРИМЕР 6
Продуцирование моноклональных антител против гибридных белков "IСАМ-4 крысы/GST"
Мышей Balb/c иммунизировали путем подкожной инъекции 40-50 мкг гибридного белка "домен 2 ICAM-4/GST" (описанного в Примере 5), эмульгированного в полном адъюванте Фрейнда (ПАФ). Через две недели мышей снова иммунизировали путем подкожной инъекции того же самого белка, но эмульгированного в неполном адъюванте Фрейнда. Две конечные внутрибрюшинные иммунизации, проведенные через две недели после второй иммунизации, осуществляли с использованием растворимого антигена без адъюванта с двухнедельными интервалами. Сыворотку, полученную от каждой иммунизованной мыши, анализировали с помощью ELISA на способность к специфической реакции с крысиным IСАМ-4, продуцированным бакуловирусной экспрессирующей системой, описанной выше.
У мыши #1654 удаляли селезенку в стерильных условиях и помещали в 10 мл бессывороточную среду RPMI-1640. Моноклеточную суспензию приготавливали путем измельчения ткани селезенки между замороженными концами двух предметных стекол микроскопа, погруженных в бессывороточную среду RPMI-1640 (Gibco, Burlingen, Ottawa, Canada), в которую были добавлены 2 мМ L-глутамина, 1 мМ пирувата натрия, 100 ед. /мл пенициллина, и 100 мкг/мл стрептомицина. Эту клеточную суспензию фильтровали через стерильное клеточное сито Nitex в 70 меш (Becton Dickinson, Parsippany, NJ), и два раза промывали RPMI с последующим центрифурированием при 200хg в течение 5 минут. Осадок, полученный после конечной промывки, ресуспендировали в 20 мл бессывороточной среды RPMI. Тимоциты, взятые от трех неиммунизованных мышей BALB/c, получали аналогичным образом.
Перед слиянием миеломные клетки NS-1 поддерживали в логарифмической фазе роста в среде RPMI, содержащей 11% сыворотку Fetaclone (FBS) (Hyclone Laboratories, Logan, Utab) в течение трех дней. После сбора клетки центрифугировали при 200xg в течение 5 минут, и осадок два раза промывали, как описано в предыдущем параграфе. После промывки клеточную суспензию доводили до конечного объема 10 мл в бессывороточной среде RPMI. Затем брали 20 мкл аликвоты, разводили 1:50 бессывороточной средой RPMI, из этого разведения брали 20 мкл аликвоты, смешивали ее с 20 мкл 0,4% трипанового синего в 85% физиологический раствор (Gibco), загружали в гемацитометр (Baxter Helthcare, Deerfield, IL), и клетки подсчитывали. Приблизительно 2,425•108 клеток селезенки объединяли с 4,85•107 клетками NS-1, смесь центрифугировали, и супернатант удаляли. Полученный осадок удаляли путем промывания пробирки водопроводной водой, и в течение 1 минуты, размешивая, добавляли 2 мл 50% ПЭГ 1500 в 75 мМ Hepes, pH 8,0 (Boehringer Mannhiem, Indianapolis, IN). Затем в течение более 7 минут добавляли еще 14 мл бессывороточной RPMI. Клеточную суспензию центрифугировали при 200xg в течение 10 минут, и супернатант отбрасывали. Осадок ресуспендировали в 200 мл RPMI, содержащей 15% FBS, 100 мкМ натрийсодержащего гипоксантина, 0,4 мкМ аминоптерина, 16 мкМ тимидина (HAT) (Gibco), 25 ед. /мл IL-6 (Boehringer Mannhiem) и 1,5•106 тимоцитов/мл. Сначала суспензию промещали в 225 см2-колбу (Corning, Essex, United Kingdom) при 37oС в течение четырех часов, а затем распределяли по десяти 96-луночным плоскодонным планшетам для культивирования тканей (Corning) в концентрации 200 мкл/лунку. Клетки в планшетах подпитывали на дни 3, 4, 5 и 6 после слияния путем отсасывания иглой калибра 20 G (Becton Dickinson) из каждой лунки приблизительно 100 мкл среды и добавления 100 мкл на лунку среды для культивирования, описанной выше, за исключением того, что эта среда содержала 10 ед./мл IL-6 и не содержала тимоцитов.
Планшеты для слияния сначала скринировали с помощью ELISA методом захвата антигена следующим образом. 4 планшета Immulon (Dynatech, Cambridge, МА) сенсибилизировали в течение ночи при 4oС 100 нг/лунку либо гибридного белка "домен 1-GST", либо гибридного белка "домен 2 - GST" в 50 мМ карбонатном буфере. Эти планшеты блокировали 100 мкл/лунку 0,5% желатина рыбьей чешуи (Sigma, St. Louis, МО) в PBS в течение 30 минут при 37oС. После блокирования планшеты промывали 3 раза PBS, содержащим 0,05% Твина 20 (PBST), и добавляли 50 мкл/лунку супернатанта гибридомы от каждого слияния. После инкубирования в течение 30 минут при 37oС планшеты промывали, как описано выше, и добавляли 50 мкл разведения 1:3500 конъюгированных с пероксидазой хрена козьих антимышиных IgG (Fc) (Jackson Immuno-Research, West Grove, Pensylvania). Затем планшеты снова инкубировали в течение 30 минут и промывали 4 раза PBST. После этого добавляли 100 мкл/лунку субстрата, содержащего 1 мг/мл о-фенилендиамина (Sigma) и 0,1 мкл/мл 30% H2O2 в 100 мМ цитрате, рН 4,5. Цветная реакция проходила в течение 10 минут, после чего ее гасили добавлением 50 мкл/лунку 15% H2SO4. Затем определяли оптическую плотность при 490 нм на автоматическом ридере (Dynatech).
Лунки, которые были позитивными для белка "домен 2-GST", но не для белка "домен 1-GST", скринировали с помощью ELISA против супернатанта бакуловируса (как описано выше). ELISA осуществляли как описано выше, за исключением того, что 4 планшета Immulon были сначала в течение ночи сенсибилизированы супернатантом бакуловируса, разведенного 1:4 в 50 мМ карбонатном буфере. Три лунки (103А, 103В и 103F) клонировали от двух до трех раз последовательно путем дублирования разведения в RPMI, 15% FBS, 100 мкМ гипоксантина натрия, 16 мкМ тимидина, и 10 ед./мл IL-6. Лунки из клонированных планшетов оценивали визуально через 4 дня, и число колоний в лунках с наименьшей плотностью регистрировали. Лунки, отобранные от каждого клонирования, снова оценивали с помощью ELISA через 7-10 дней либо на белок "домен 1 - GST" и белок "домен 2-GST", либо на супернатант бакуловируса.
Моноклональные антитела, продуцированные гибридомами, были изотипированы с помощью ELISA. 4 планшета Immulon (Dynatech) сенсибилизировали при 4oC 50 мкл/лун. козьими антителами против мышиных IgA, IgG или IgM (Organon Teknika, Durham, NC), разведенными при 1:5000 в 50 мМ карбонатном буфере, рН 9,6. Лунки блокировали в течение 30 минут при 37oС 1% BSA в PBS, а затем трижды промывали PBST. В каждый планшет добавляли 1:10 разведение супернатанта гибридомной культуры (50 мкл), инкубировали и промывали, как описано выше. После удаления последней промывки добавляли 50 мкл конъюгированных с пероксидазой хрена кроличьих антител против мышиных IgG1, G2a. G2b или G3 (Zymed, San Francisco, СА)(разведенных 1:1000 в PBST с 1% нормальной козьей сывороткой). Планшеты инкубировали, как описано выше, промывали 4 раза PBST и добавляли 100 мкл субстрата. Цветную реакцию гасили через 5 минут путем добавления 50 мкл 15% H2SO4, и оптическую плотность определяли при 490 нм на планшет-ридере (Dynatech).
Результаты показали, что все антитела 103А, 103В, и 103F представляли собой изотип IgG1. Затем эти антитела были использованы в иммуноцитохимических анализах в Вестернблоттинге и для очистки белка, экспрессированного в бакуловирусе.
ПРИМЕР 7
Бакуловирусная экспрессия IСАМ-4 крысы
Для получения растворимого белка, соответствующего доменам 1-6 ICAM-4, была использована бакуловирусная экспрессирующая система (Invitrogen). Поскольку лидерная последовательность для IСАМ-4 в настоящее время не известна, была создана экспрессионная конструкция, содержащая кодирующую последовательность для IСАМ-4, слитую у 3' с лидерной последовательностью IСАМ-1 с сохранением рамки считывания. Конкретные подробные данные, относящиеся к плазмиде, экспрессирующей конструкцию IСАМ-1/IСАМ-4, приведены ниже.
IСАМ-ДНК крысы, кодирующую пять lg-подобных доменов, амплифицировали посредством PCR с использованием праймеров, которые включали некоторые отличительные особенности для облегчения конструирования гибридной плазмиды. Помимо консенсусной последовательности Kozak, расположенной выше первого метионина в лидерной последовательности, 5'-олигонуклеотидный праймер включал HindIII и Bg/II-сайты. 3'-Олигонуклеотидный праймер включал кодирующую последовательность для шести гистидинов, за которой следовали стопкодон и HindIII-сайт клонирования. Продукт PCR-амплификации клонировали в HindIII-гидролизованный вектор pBS+, и анализ последовательности подтверждал соответствующую конструкцию. Был осуществлен гидролиз во внутреннем Smal-caйтe в лидерной последовательности IСАМ-1 и в другом Smal-сайте в области множественного клонирования вектора (3' до lg-подобного домена 5 IСАМ-1), что приводило к удалению большей части последовательности, кодирующей IСАМ-1. После проведения этих манипуляций линеаризованный затупленный по концам вектор содержал часть расположенной выше области множественного клонирования (рестрикционные сайты у 5' относительно первоначального Hindlll-сайта в области множественного клонирования), последовательность Kozak и наибольшую часть лидерной последовательности IСАМ-1.
Кодирующую последовательность для доменов 1-6 крысиного ICAM-4 амплифицировали с помощью PCR с использованием праймеров, сконструированных в целях клонирования этой последовательности в линеаризованный вектор, описанный выше. 5'-олигонуклеотидный праймер включал EcoRV-сайт и кодоны, необходимые для завершения лидерной последовательности IСАМ-1. 3'-олигонуклеотидный праймер включал кодоны для шести гистидиновых остатков, стопкодон и рестрикционные HindIII- и EcoRV-сайты. Продукт PCR-амплификации гидролизовали EcoRV с получением затупленной по концам последовательности, которую затем лигировали в затупленный по концам Smal-гидролизованный линеаризованный вектор pBS+. Полную последовательность, содержащую лидерную последовательность IСАМ-1 от 5'-конца до доменов 1-6 ICAM-4, удаляли из конструкции путем гидролиза ферментами BgIII и HindIII, и очищенную гибридную последовательность IСАМ-1/IСАМ-4 клонировали непосредственно в BgIII/HindIII-гидролизованный вектор pBluesac III (Invitrogen).
Продуцирование белка с помощью рекомбинантного вируса анализировали посредством ELISA, сначала с использованием иммунных сывороток от мышей, иммунизированных гибридным белком "домен 2 крысиного ICAM-4/GST", как описано в Примере 5. В более поздней работе моноклональные антитела, полученные от этих мышей, были использованы для очистки белка ICAM-4, продуцированного рекомбинантным бакуловирусом в клетках SF9.
ПРИМЕР 8
Продуцирование моноклональных антител против экспрессированного бакуловирусом крысиного IСАМ-4
Домены 1-6 крысиного ICAM-4 были экспрессированы в бакуловирусной экспрессирующей системе, как описано в Примере 7. Рекомбинантный белок очищали с использованием моноклонального антитела 103А (как описано в Примере 6).
Вкратце, 30 мг очищенного моноклонального антитела 103А (в 100 мМ бората натрия, 500 мМ хлорида натрия) связывали с тремя граммами активированной бромцианом Сефарозы 4В (Pharmacia, Piscataway, NJ). Бакуловирусный супернатант, содержащий рекомбинантный крысиный ICAM-4 (домены 1-6), загружали на ночь на колонку с Сефарозой при 4oС. Колонку промывали забуференным фосфатом физиологическим раствором, не содержащим кальция-магния (CMF-PBS), и связанный материал элюировали 50 мМ лимонной кислотой, 500 мМ NaCl, pH 4,0. Образец нейтрализовали 1/10 объемами Триса, pH 10 и хранили при -20oС. Очищенный белок, разделенный путем электрофореза в ПААГ-ДСН, имел чистоту более чем 90% и мигрировал приблизительно при 80 кДа.
Мышей иммунизировали очищенным крысиным рекомбинантным белком ICAM-4 с 1-6 доменами способом, аналогичным описанному в Примере 6. Для получения гибрида #127 использовали селезенку от мыши #1945. Протокол слияния описан в Примере 6. Лунки для слияния скринировали с помощью ELISA на рекомбинантный белок ICAM-4. Вторичный скрининг проводили иммуноцитохимическим методом на срезах головного мозга крысы (как описано ниже в Примере 9). Исходя из этого слияния были клонированы четыре дополнительных антитела, специфичных для ICAM-4 крысы: 127А, 127Е, 127F и 127Н. Картина иммуноцитохимического окрашивания каждым антителом на срезах головного мозга крысы была аналогична картине, наблюдаемой с использованием моноклонального антитела 103А (см. Пример 9). Моноклональные антитела были тестированы на их способность связываться со гибридными белками D1/GST и D2/GST (описанными в Примере 5). Моноклональное антитело 127А распознавало гибридный белок D1/GST, а моноклональное антитело 127H распознавало гибридный белок D2/GST. Эти две различные специфичности связывания моноклональных антител, наряду с другими антителами, которые не связываются ни с одним белком GST, дают основание предположить, что, по крайней мере, 3 различных эпитопа распознаются панелью антител. Гибридомы 127А и 127Н были депонированы 31 мая 1995 г. и 1 июня 1995 г. соответственно Американской коллекцией типовых культур, 12301 Parklawn Drive, Rockville, Maryland 20852, и им были присвоены номера доступа НВ11905 и НВ11911 соответственно.
ПРИМЕР 9
Иммуноцитохимия экспрессии IСAM-4 крысы
Иммуноцитохимию с использованием моноклонального антитела 103А осуществляли для локализации продуцирования белка в головном мозге крысы.
Головной мозг брали от нормальной взрослой самки крысы Lewis, сагиттально рассекали и промывали 1 раз PBS, не содержащим РНКазы, на льду в течение 30 минут. Затем срезы головного мозга помещали на криоформы Tissue Tek II (Miles Laboratories, Inc., Naperville, IL) с небольшими количеством соединения О. С. Т. (Miles, Inc., Elkhart, IN). Головной мозг центрировали на криоформах, эти криоформы заполняли соединением ОСТ, а затем помещали в контейнер с 2-метилбутаном (Aldrich Chemical Company, Inc., Milwaukee, Wl), и контейнер помещали в жидкий азот. После того как ткань и ОСТ в криоформах были заморожены, блоки хранили при -80oС до тех пор, пока их не брали для приготовления срезов.
Ткань иссекали с получением срезов толщиной 6 мкм, фиксировали на предметных стеклах, покрытых Vectabond (Vector Laboratories, Inc, Burlingame, CA), и оставляли на ночь для осушки на воздухе при комнатной температуре вплоть до их использования. Эти срезы фиксировали в этиловом эфире (Malinckrodt, Paris, KY) в течение 5 минут при комнатной температуре. После удаления предметных стекол из эфира реагент оставляли для испарения. Каждый срез ткани блокировали 150 мкл 50% нормальной крысиной сыворотки (Sigma) и 2% альбумина бычьей сыворотки (BSA) (Sigma) в 1Х PBS (приготовленного только с использованием фосфата натрия) при комнатной температуре в течение 30 минут. После блокирования раствор мягко блотировали со срезов и антитело 103А, очищенное из супернатанта (1,65 мг/мл), разводили 1:10 в блокирующем растворе и 150 мкл этого раствора наносили на каждый срез ткани. Предметные стекла помещали в увлажняющую камеру и инкубировали в течение ночи при 4oС.
На следующий день раствор антитела мягко блотировали со среза, и предметные стекла три раза промывали в 1Х PBS в течение четырех минут в каждой промывке. Избыток PBS отсасывали с предметного стекла и на ткани наносили 100 мкл второго крысиного антимышиного конъюгированного с биотином антитела (Jackson Immuno-Research Laboratories), разведенного 1:100 в растворе 10% сыворотки нормальной крысы и 2% BSA в 1Х PBS. Инкубирование осуществляли в течение одного часа при комнатной температуре. Срезы промывали два раза в 1Х PBS в течение четырех минут в каждой промывке, а затем на каждый срез наносили 100 мкл реагента АВС из набора, содержащего IgG крысы и вектазин (Elite Rat IgG Vectastain ABC kit) (Vector Laboratories, Inc., Burlingame, CA), полученного в соответствии со вставкой продукта. Инкубирование проводили в течение 30 минут при комнатной температуре. После инкубирования предметные стекла дважды промывали в 1Х PBS (каждая промывка по четыре минуты) и на каждый срез приблизительно в течение десяти минут наносили 150 мкл раствора субстрата пероксидазы и вектора VIP (Vector Laboratories, Inc., Burlingame, CA). После проявления окраски срезы промывали под струей водопроводной воды в течение пяти минут, подвергали контрастному окрашиванию гематоксилином Mayer (Sigma) в течение 20 секунд и снова промывали под медленной струей водопроводной воды в течение пяти минут. Предметные стекла дегидратировали с использованием этанола высшего сорта, пропускали через ксилол и помещали на устройстве Accumount 60 (Stephens Scientific, Riverdale, NJ).
Иммуногистохимический анализ срезов головного мозга крысы, окрашенных моноклональным антителом (mAb) 103A, показал, что крысиный ICAM-4 экспрессировался в нервных клетках гиппокампа. Характер окрашивания дает основание предположить, что данный белок может быть ограничен отростками нейронов (дендритов). Срезы головного мозга, окрашенные аналогичным образом, только нерелевантным антителом или реагентом второй стадии не давали четкой картины экспрессии, наблюдаемой в случае Маb 103А.
ПРИМЕР 10
Клонирование геномной ДНК IСАМ-4 человека
В процессе клонирования крысиного IСАМ-4 из геномной ДНК было выявлено, что IСАМ-4 и IСАМ-1 расположены на расстоянии 5 т.п.о. друг от друга, и эта информация была использована при попытке клонировать человеческий гомолог ICAM-4.
С помощью геномных систем (Genome Systems Inc.) (St. Louis, МО) были амплифицированы фрагменты в библиотеке Р1 человека посредством PCR с использованием праймеров для домена 3 IСАМ-1 человека; сконструированного смыслового праймера, комплементарного домену 3 IСАМ-1 человека (H-1/D3S); и сконструированного антисмыслового праймера, комплементарного домену 3 IСАМ-1 человека (H-3/D3 AS). Эти праймеры представлены в SEQ ID No:22 и 23, соответственно.
CCGGGTCCTAGAGGTGGACACGCA (SEQ ID No:22)
TGCAGTGTCTCCTGGCTCTGGTTC (SEQ ID No:23)
Два клона, обозначенные 1566 и 1567, были идентифицированы и использованы для дальнейшего анализа. Оба клона Р1 содержали геномные ДНК-вставки размером приблизительно 75-95 т. п. о. Эти клоны гидролизовали ферментом SamH1, разделяли путем электрофореза в агарозном геле и блотировали на найлоновые мембраны. Саузерн-блот-гибридизацию осуществляли либо в условиях низкой жесткости (30% формамид), либо в условиях высокой жесткости (60% формамид) при 42oC с использованием радиоактивно меченных зондов для IСАМ-1 человека, ICAM-3 человека или ICAM-4 крысы; другие составляющие раствора для гибридизации были такими, как они описаны в Примере 1. Партию для гибридизации при низкой жесткости промывали при комнатной температуре в 2•SSPE, содержащем 0,1% ДСН. Партию для гибридизации с высокой жесткостью промывали при 65oС в 0,2•SSPE, содержащем 0,1% ДСН. Промытые мембраны экспонировали с рентгеновской пленкой в течение 3,5 часа.
Дифференциальная гибридизация показала, что человеческий IСАМ-1 находился на 5,5 т.п.о.-SamH1-фрагменте, в то время как человеческий IСАМ-3 находился на 4,0 т.п.о. - и 1,5 т.п.о.-BamHI-фрагментах. Фрагменты человеческих IСАМ-1 и ICAM-R субклонировали в pBS+ и их идентичность была подтверждена анализом лимитированных последовательностей.
7,0 т. п. о.-BamH1-фрагмент, который гибридизовался с крысиным ICAM-4 в условиях высокой жесткости, субклонировали, а затем фрагментировали путем гидролиза рестриктазой Rsal. Были идентифицированы три Rsal-фрагмента, которые гибридизировались с крысиным ICAM-4, и их последовательности были определены. Исходя из гомологии с крысиным ICAM-4 очевидно, что эти фрагменты содержат домены 2, 3, 4, 5 и часть домена 6.
ПРИМЕР 11
Клонирование кДНК ICAM-4 человека
Фрагменты геномной ДНК, соответствующей доменам 2-5 человеческого ICAM-4 (описанных в Примере 10), были использованы в качестве зондов для скрининга кДНК-библиотеки гиппокампа человека λgt10 (Clonetech, Palo Alto, CA). Протокол скрининга библиотеки был, в основном, описан в Примере 1.
Наиболее длинный клон IСАМ-4 человека (#18), который был обнаружен в этой библиотетке, составлял всего 992 п.о. (SEQ ID No:24) и соответствовал, по грубой оценке, середине предсказанного 3 т.п.о. -гена. Эту 992 п.о. - ДНК-вставку из клона 18 (SEQ ID No:24) использовали в качестве зонда для скрининга кДНК-библиотеки гиппокампа человека в λZAPII (Stratagene, La Jolla, CA). Эта библиотека дала ряд позитивных клонов. Наиболее длинный клон, # 34, составлял 2775 п.о. (SEQ ID No:25). Исходя из сравнения его первичной последовательности с полноразмерной последовательностью крысиного ICAM-4, было предположено, что в этом клоне отсутствует лидерная последовтельность и приблизительно 30 п. о. в 5'-конце домена 1. Poly(A)+PHK-xвocт на 3'-конце отсутствовал, но присуствовал кодон терминации трансляции.
Фрагмент ДНК, соответствующий первым 3 доменам (нуклеотидам 1-840 в клоне # 34) использовали в качестве зонда для скрининга кДНК-библиотеки λgt10, полученной из коры головного мозга человека (Clonetech, Palo Alto, CA). Один клон, 16-1 (SEQ ID No:26) был идентифицирован как клон, имеющий 1557 п.о. и включающий 39 п. о. 5'-нетранслируемой ДНК, лидерную последовательность и данные о последовательности пятого домена. Перекрывающиеся клоны #34 (SEQ ID No: 25) и 16-1 (SEQ ID No:26) были использованы для генерирования составных частей полноразмерной последовательности ICAM-4 человека (SEQ ID No:27).
Полноразмерный ген имеет длину 2927 п.о. и кодирует белок в 924 аминокислоты. Нуклеотидная последовательность ICAM-4 представлена в SEQ ID No:27, а аминокислотная последовательность представлена в SEQ ID No:28. Сравнение первичной последовательности с полноразмерным геном крысиного ICAM-4 (SEQ ID No: 11) выявило, что полная ДНК-последовательность имеет 82%-идентичность, а аминокислотная последовательность имеет 85%-идентичность. Видимая структура 9 lg-подобного внеклеточного домена белка крысы и человека является консервативной. Лидерная последовательность простирается от аминокислоты 1 до аминокислоты 28; домен 1 простирается от аминоксилоты 29 до аминокислоты 117; домен 2 простирается от аминокислоты 118 до аминокислоты 224; домен 3 простирается от аминокислоты 225 до аминокислоты 320; домен 4 простирается от аминокислоты 321 до аминокислоты 405; домен 5 простирается от аминокислоты 406 до аминокислоты 488; домен 6 простирается от аминокислоты 489 до аминокислоты 570; домен 7 простирается от аминокислоты 571 до аминокислоты 663; домен 8 простирается от аминокислоты 664 до аминокислоты 743; домен 9 простирается от аминокислоты 744 до аминокислоты 837; трансмембранная область простирается от аминокислоты 838 до аминокислоты 857; цитоплазматический хвост простирается от аминокислоты 858 до 924.
ICAM-4 человека (HulCAM-4), помимо своего генетического сходства с IСАМ-1 и ICAM-R, также обнаруживает общие структурные особенности с белками, которые позволяют классифицировать их как семейство молекул. Сравнение первичных последовательностей доменов HulCAM-4 с другими членами семейства ICAM-4 обнаружило различную степень гомологии. Аминокислотная последовательность домена 1 HulCAM-4 имеет 21, 30 и 26% идентичности с доменом 1 IСАМ-1, 2 и 3 соответственно. Домен 2 HulCAM-4 имеет 61, 39 и 62% идентичности с ICAM 1, 2 и 3 соответственно. Домен 3 HulCAM-4 имеет 50 и 65% идентичности с IСАМ 1 и 3 соответственно. Домен 4 HulCAM-4 имеет 54 и 64% идентичности с ICAM 1 и 3 соответственно. Домены 5-8 HulCAM-4 имеют наибольшую гомологию с пятыми доменами ICAM 1 и 3, при этом процент их идентичности составляет в пределах 33-47% для домена 5 IСАМ-1 и 21-31% для домена 5 ICAM-R. Последовательность девятого домена HulCAM-4 мало соответствует другим членам семейства ICAM-4, но обнаруживает гомологию с доменами 3 (24% идентичности) и 6 (23% идентичности) HulCAM-1.
ПРИМЕР 12
Нозерн-блот-анализ экспрессии ICAM-4 человека
Два Нозерн-блота множества тканей человека (MTN) было получено от Clonetech (Palo Alto, CA). Они содержали, по крайней мере, 2 мкг poly(A)+PHK от 16 различных тканей человека (как показано в Таблице 1), которые были подвергнуты электрофорезу в денатурирующем формальдегидсодержащем 1,2% агарозном геле и перенесены на найлоновую мембрану. Эти блоты подвергали предварительной гибридизации в течение трех часов при 42oС в 10 мл раствора, содержащего 5Х SSPE, 10 • раствора Денхардта, 50% формамида, 2% ДСН и 100 мкг/мл денатурированной ДНК спермы лосося. Эти блоты гибридизовали в вышеуказанном растворе с радиоактивно меченным зондом ICAM-4 человека (клон #18, SEQ ID No: 24) в течение 16 часов при 42oС. На следующий день эти блоты промывали в растворе 0,1Х SSC/0,1% ДСН при комнатной температуре, а затем промывали при 50oС. Блоты экспонировали с рентгеновской пленкой в течение 24 часов при -80oС. Результаты анализа представлены в Таблице 1.
Только дорожка, содержащая РНК от головного мозга, гибридизованную с ICAM-4-зондом, давала одну полосу приблизительно 3 т.п.о. Более продолжительное экспонирование (пять дней) подтвердило, что только головной мозг обнаруживает детектируемый уровень транскрипта. Для того чтобы определить, содержат ли все дорожки сравнимые количества РНК при сравнимом качестве, тот же самый блот был гибридизован с контрольным β-=актиновым зондом. Блоты были отделены от ICAM-4-зонда путем 15-минутной обработки кипящим раствором 0,1% ДСН, а затем зондированы аналогичным образом β-актиновым зондом в соответствии с указаниями производителя. За исключением лишь небольших вариаций в количествах все дорожки обнаруживали РНК хорошего качества.
От Clonetech были получены два дополнительных Нозерн-блота, которые содержали poly A+ РНК от 16 различных субобластей головного мозга человека (как показано в Таблице 2). Эти блоты зондировали способом, аналогичным способу, использованному для анализа тканей, и результаты представлены в Таблице 2. Количество и качество загруженной РНК проверяли путем зондирования блотов β-актиновым зондом.
Все области, которые обнаруживали экспрессию ICAM-4, являлись частью конечного мозга, за исключением таламуса, который считается частью промежуточного мозга. Гиппокамп и кора головного мозга обнаруживали самый высокий уровень экспрессии. Размер транскрипта во всех случаях был таким же и составлял 3 т.п.о. Явно выраженное распределение экспрессии ICAM-4 в тканях дает основание предположить, что промоторная область может содержать элементы, которые обеспечивают стадие- и пространственно-специфическую экспрессию генного продукта. Использование этой информации может дать общее представление о регуляции экспрессии генов нервных тканей.
ПРИМЕР 13
Генерирование гибридных белков IСДМ-4 человека/lgG
Для использования в анализе адгезии в целях идентификации потенциальных лигандов IСАМ-4 были сконструированы плазмиды, экспрессирующие гибридные белки IСАМ-4 человека/lgG. Были получены две конструкции: первая конструкция включала ДНК, кодирующую домены 1-3 HulCAM-4, а вторая конструкция включала домены 4-8. Обе эти конструкции были соединены с Fc-областью человеческого lgG1 в векторе pDCS1, который содержал промотор цитомегаловируса (CMV) для регулирования экспрессии и сигнальную последовательность от lgG4 для стимуляции экспрессии молекул.
Для генерирования нужных ДНК-фрагментов для субклонирования были сконструированы PCR-праймеры (показанные ниже в SEQ ID No:29-32). Для достраивания 30 п.о. домена 1, отсутствующих в клоне #34, был сконструирован "смысловой" праймер для 5'-конца домена 1 (H14-D1, SEQ ID No:29). Праймеры H14-D1(S)(SEQ ID No: 29) и H14-D3(AS)(SEQ ID No:30) были использованы для генерирования ДНК-фрагмента, кодирующего домены 1-3 человеческого ICAM-4, соответствующего области в SEQ ID No:1 от нуклеотида 130 до нуклеотида 996. Праймеры Н14-D3(S)(SEQ ID No:31) и H14-D8(AS) (SEQ ID No:32) были использованы для генерирования ДНК-фрагмента, кодирующего домены 4-8 человеческого ICAM-4 и соответствующего области в SEQ ID No:30 от нуклеотида 997 до нуклеотида 2268. Для облегчения субклонирования у 5' относительно гена lgG1 каждый 5'-праймер кодировал рестрикционный SamH1-сайт (GGATCC, показанный жирным шрифтом в Таблице А), а каждый 3' (антисмысловой) праймер содержал Xhol-сайт (CTCGAG, показанный жирным шрифтом в Таблице А). Для осуществления рестрикционного гидролиза, все олигонуклеотиды содержали спейсерные нуклеотиды (подчеркнутые, см. в Таблице А) у 5'-конца.
PCR-реакции осуществляли в объеме 50 мкл с использованием буферов, поставляемых Perkin Elmer, с ферментом AmpliTaq. Праймеры добавляли с конечной концентрацией 10 мкг/мл, а все четыре dNTP имели концентрацию 2 мМ. Реакции продолжали в течение 30 циклов: денатурации (94oС, четыре минуты), отжига (50oС, две минуты), и удлинения (72oС, одна минута). PCR-продукты визуализировали на агарозных гелях и аликвоты каждой реакции использовали для субклонирования PCR-продуктов в вектор pCRII (Invitrogen, San Diego, CA). Для обнаружения возможных ошибок, возникших в результате процедуры амплификации, и для подтверждения правильной ориентации был проведен анализ последовательности. Соответствующие клоны гидролизовали ферментами ВаmH1 и Xhol, и фрагменты разделяли путем электрофореза в агарозном геле. Очищенные фрагменты лигировали в вектор pDCS1, предварительно гидролизованный ферментами BamH1 и Хhol, и полученные плазмиды секвенировали для подтверждения правильной ориентации и рамки считывания.
Гибридные белки "домены 1-3 и 4-8 ICAM-4 человека/lgG1" были получены вследствие кратковременной трансфекции экспрессирующих плазмид в клетки COS 7 и выделения секретированного белка из культуральных сред. Трансфекцию осуществляли следующим образом. Адгезивные клетки COS 7 приблизительно при 50-60% конфлюэнтности промывали CMF-PBS, а затем подвергали контакту с 10-15 мкг плазмидной ДНК в 7,5 мл бессывороточной среды DMEM (Gibco, Gaithers-burg, MD), содержащей 6 мкл 0,25 М хлорохина (Sigma, St. Louis, МО). Затем добавляли еще 7,5 мл бессывороточной среды, содержащей 150 мкл декстрана-DEAE (50 мг/мл) (Sigma, St. Louis, МО), и эти планшеты инкубировали 2-3 часа, а затем среды удаляли и меняли на 10% ДМСО (Mallinckrodt, McGaw Park, Illinois) в PBS. После инкубирования в течение одной минуты раствор ДМСО удаляли и заменяли свежей средой, содержащей 5% FBS. Каждую трансфекцию проводили во множестве планшетов, и среды от клеток, экспрессирующих тот же самый белок, объединяли для выделения этого белка.
Среды собирали каждые три дня, получая порядка 3-4 урожаев. Белки очищали на 0,4-0,8 мл-колонке Procep A (Bioprocessing Ltd., England), предварительно уравновешенной 35 мМ трисом, 150 мМ NaCl, pH 7,5. Культуральные среды наносили на колонку дважды при скорости потока менее чем 60 колоночных объемов в 1 час. Колонку один раз промывали (каждым) 20 колоночными объемами буфера трис/NaCl, 20 колоночными объемами 0,55 М диэтаноламина, pH 8,5, и 20 колоночными объемами 50 мМ лимонной кислоты, pH 5,0. Гибридные белки элюировали в 1 мл-фракции с использованием 50 мМ лимонной кислоты, pH 3,0, и каждую фракцию нейтрализовали 1/10 объемами 1М триса, pH 9,5. Концентрацию белка определяли по ОП280, а чистоту определяли с помощью электрофореза в ПААГ-ДСН.
Были отмечены значительные примеси бычьего IgG (присутствующего в FBS). Хотя, как было предсказано, этот гибридный белок с доменами 1-3 имеет меньший размер, чем гибридный белок с доменами 4-8, однако оба этих белка мигрировали приблизительно при 90 кДа. Одним из возможных объяснений этого факта является то, что меньший гибридный белок с доменами 1-3 может более интенсивно гликозилироваться, чем более крупные гибридные белки с доменами 4-8.
Помимо использования очищенных белков для продуцирования моноклонального антитела, описанного ниже, эти белки могут быть также использованы в анализах на адгезию для идентификации лигандов IСАМ-4.
ПРИМЕР 14
Продуцирование моноклональных антител
Очищенный белок, описанный в Примере 13, использовали для генерирования моноклональных антител в соответствии с протоколом иммунизации, описанным в Примере 6.
Селезенку от мыши # 2250 (иммунизированной HulCAM-4D1-3/-lgG1) использовали для слияния 172, а селезенку мыши #2272 (иммунизированной HulCAM-4 D4-8/lgG1) использовали для слияния 173. Протокол слияния использовали, как описано в Примере 6. Планшеты для слияния скринировали с помощью ELISA (в основном, как описано в Примере 6) с использованием каждого из гибридных белков HulCAM-4/lgG1. Для клонирования рассматривали только супернатанты лунок для слияния, которые распознавали иммуногенный белок, и никакие другие. В качестве вторичного скрининга использовали иммуноцитохимический анализ на срезах гиппокампа человека.
Один первичный клон от каждого гибрида был положительным, на что указывал иммуноцитохимический анализ, и был клонирован. Поскольку роста одного из двух клонов не наблюдалось после клонирования, то оставался только один кандидат для исследования, а именно, клон 173Е, который происходил от мыши, иммунизованной HulCAM D4-8/lgG1. Гибридома 173Е была депонирована 1 июня 1995 Американской коллекцией типовых культур, 12301 Parklawn Drive, Rockville, Maryland 20852 и зарегистрирована под номером допуска НВ11912.
Было обнаружено, что из других гибридов, происходящих от мыши, иммунизованной фрагментом растворимого ICAM-4, соответствующим доменам 1-3, шесть клонов (179А, 179В, 179D, 179Н, 1791, и 179К) являются специфичными для доменов 1-3 (D1-3) HulCAM4. Все шесть антител в серии 179 связываются с дендритными отростками в зубчатой извилине, а также слоях полиморфных и пирамидных клеток. Помимо дендритных отростков, моноклональное антитело 179А окрашивало тела нервных клеток из этих участков. Гибридомные клеточные линии, продуцирующие антитела 1791 и 179Н, были депонированы 10 июня 1996 г. в Американской коллекции типовых культур, 12301 Parklawn Drive, Rockville Maryland, 20852 и зарегистрированы под номерами допуска НВ 12123 и НВ 12124 соответственно.
Аналогичным образом были проведены другие слияния в целях генерирования антител, являющихся иммунореактивными по отношению к конкретным участкам ICAM-4.
ПРИМЕР 15
Разработка анализа, проводимого методом захвата
Шесть моноклональных антител от гибрида 179 тестировали в различных комбинациях на их способность захватывать и обнаруживать растворимый ICAM-4 в растворе. Этот анализ, описанный ниже, был разработан для оценки уровней растворимого ICAM-4 в жидкостях организма здорового человека и человека, страдающего заболеванием.
96-луночные планшеты Immulon 4 (Dynatech) сенсибилизировали
антителом 1791 в концентрации 3 мкг/мл, 125 мкл/лунку в течение двух часов при 37oС. Раствор антитела удаляли путем отсасывания, и лунки блокировали в течение 30 минут при комнатной температуре 300 микролитрами блокирующего раствора, содержащего 5% желатин Teleostean в PBS, не содержащего кальция и магния (CMF-PBS). Блокирующий раствор удаляли путем отсасывания, после чего в каждую лунку добавляли 100 мкл жидкой пробы, разбавленной в разбавителе Omni (CMF-PBS, 1% желатин и 0,05% Твин-20), и полученную смесь инкубировали в течение 30 минут при 37oС. Планшеты три раза промывали PBST (CMF-PBS, 0,05% Твин-20). Антитело 179Н биотинилировали при 1,5 мг/мл с использованием NHS-LC-биотина (Pierce) в соответствии с протоколом, предлагаемым производителями, разводили 1:2000 и добавляли в лунки (100 мкл/лунку). Полученную смесь инкубировали в течение 30 минут при 37oС, и планшеты три раза промывали PBST. В каждую лунку добавляли стрептавидин-пероксидазу хрена (HRP) (Pierce) (100 мкл, 0,25 мкг/мл) и эту смесь инкубировали в течение 30 минут при 37oС. Планшеты четыре раза промывали PBST, а затем добавляли 100 мкл тетраметилбензидина (Sigma)(10 мг/мл исходного продукта в ДМСО), разведенного 1: 100 в забуференном субстрате (13,6 г/л тригидрата ацетата натрия, рН доводили до 5,5 путем добавления 1М лимонной кислоты, 150 мкл/л 30% перекиси водорода непосредственно перед проявлением). Реакцию проявляли в темноте в течение 30 минут при комнатной температуре, после чего, реакцию прекращали путем добавления 50 мкл/лунку 15% H2SO4. Оптическую плотность считывали при 450 нм.
Результаты показали, что этот анализ позволяет детектировать растворимый рекомбинантный белок HulCAM-4 D1-3 при концентрации вплоть до 5-10 нг/мл, причем линейный участок кривой составляет в пределах 10-100 нг/мл. В том случае, когда эту область белка тестировали при 1 и 10 мкг/мг, какой-либо перекрестной реактивности с HulCAM-4 D4-8 не наблюдалось.
ПРИМЕР 16
Оценка растворимого IСАМ-4 в сыворотке пациента с шоком
Для оценки роли IСАМ-4 в нервных болезнях и состояниях сыворотку от двадцати восьми пациентов, страдающих острым инсультом, и от двадцати молодых здоровых волонтеров (возраст не имеет значения) исследовали, как описано выше, на различия в концентрации сыворотки растворимого IСАМ-4.
Результаты показали, что сыворотка от здоровых волонтеров не обнаруживала детектируемого уровня IСАМ-4. Однако двадцать из двадцати восьми пациентов с острым инсультом обнаруживали детектируемые уровни растворимого IСАМ-4. Сигнал от позитивных пациентов с инсультом составлял в пределах от 5 до 38 нг/мг стандарта (растворимый рекомбинантный белок IСАМ-4 D1-3).
ПРИМЕР 17
Уровни мРНК IСАМ-4в гиппокапме в крысиной модели эпилепсии
Уровни экспрессированной крысиной мРНК IСАМ-4 оценивали в гиппокампе крыс, обработанных так, чтобы получить in vivo-модель со стимуляцией эпилептогенеза [Lothman, et al., Brain Res. 360: 83-91 (1095)]. В этой модели гиппокамп крысы стимулировали серией субконвульсивных электрошоков через электрод, имплантированный в области мозга, который постепенно вызывают тяжелые приступы поведенческих расстройств. Этот способ возбуждения предусматривает двенадцать стимуляций в день, проводимых через день в течение восьми дней. После всей процедуры возбуждения один раздражитель может вызвать поведенческие приступы и гистологические изменения, которые обычно характерны для эпилепсии человека. Для полного возбуждения крысы получали две стимуляции в день в течение двух недель, а затем, через 24 часа после последней стимуляции, животные были умерщвлены. Гипокамп удаляли и иссекали для получения РНК-препарата.
От каждого образца получали полную РНК путем экстракции с использованием гуанидиния/фенола/хлороформа [Chomezynski & Sacchi, Anal.Biochem. 162:156-159 (1987)] . РНК выделяли на денатурирующем формальдегид-содержащем агарозном геле, переносили на найлоновые мембраны и гибридизовали с радиоктивно меченными ДНК-зондами, специфичными для крысиного ICAM-4 и глицеральдегид-3-фосфат-дегидрогеназы (GAPDH). GAPDH, в основном, экспрессируется геном, который обычно используется в качестве контроля для детекции изменения количества РНК, загруженного на гель для разных дорожек. Флуктуации в отношении ICAM-4/GAPDH интерпретируются как изменения в уровнях экспрессии ICAM-4. Полосы гибридизации для ICAM-4 и GAPDH были количественно оценены с помощью фосфорного визуализатора изображений, и было определено отношение ICAM-4/GAPDH.
Отношение ICAM-4/GAPDH для контрольных животных, которые не были стимулированы (n=5), было значительно выше по сравнению со стимулированной тестированной группой (n= 5), что дает основание предположить, что ICAM-4 ингибируется в результате процесса возбуждения. Однако при этом следует отметить, что контрольная группа не была подвергнута какой-либо ложной обработке такой, которая могла бы приводить к модулированию уровней мРНК IСАМ-4 в ответ на хирургическую обработку, ассоциированную с возбуждением.
ПРИМЕР 18
Концентрация IСДМ-4 в сыворотке как показатель нейродегенеративных расстройств
Циркулирующие концентрации IСАМ-4 в сыворотке оценивали как возможный показатель различных нейродегенеративных расстройств. Пробы сыворотки и/или плазмы, взятые от анонимных доноров, оценивали, как описано в выше в Примере 16, и сравнивали с пробами, взятыми от контрольных доноров, у которых ранее не было диагностировано каких-либо неврологических расстройств.
Контрольные доноры
Для установления средних фоновых значений уровней циркулирующих ICAM-4 у нормальных здоровых индивидуумов были исследованы пробы сыворотки от 100 доноров. Полученные результаты показали, что у двенадцати индивидуумов (12%) уровни циркулирующего ICAM-4 превышали 10 нг/мл. Из этих двенадцати проб в пяти пробах концентрация ICAM-4 составляла в среднем 20-100 нг/мл, три пробы имели конценцентрации ICAM-4 в среднем 20-100 нг/мл; две пробы обнаруживали уровни ICAM-4 в пределах 100-500 нг/мл и две пробы содержали ICAM-4 в концентрации, превышающей 500 нг/мл.
Для оценки стабильности наблюдаемых уровней со временем у обоих доноров с очень высокими уровнями пробы брали одновременно в различные промежутки времени в течение восьми месяцев. Наблюдения показали, что в течение этих месяцев зарегистрированные уровни давали флуктуации. Получение каких-либо медицинских данных, касающихся этих доноров, которые позволили бы провести корреляцию с уровнями IСАМ-4 и физическим состоянием этих доноров, не представлялось возможным. Как в случае проб сыворотки, так и в случае проб плазмы, полученных от одного и того же индивидуума, какой-либо разницы в уровнях присутствующего IСАМ-4 не наблюдалось.
Эти наблюдения свидетельствуют о том, что анализ на растворимый IСАМ-4 должен иметь широкое применение как в случае сыворотки, так и в случае плазмы. Кроме того, результаты показали, что IСАМ-4 является очень стабильным в крови, что дает основание предположить, что повышенный уровень IСАМ-4, который является результатом некоторых патологических состояний, по всей вероятности не может быть кратковременным. И наконец, вследствие видимой стабильности IСАМ-4 в системе кровообращения в анализе на растворимый IСАМ-4 могут быть использованы образцы банка крови, что тем самым снижает потребность в свежей крови для каждого анализа. Для того чтобы определить, влияют ли методы сбора и/или хранения на наблюдения, оценивали стабильность ICAM-4 в сыворотке путем обработки проб, взятых от одного индивидуума с самым высоким уровнем циркулирующего ICAM-4, различными способами с последующим измерением уровней ICAM-4. Ни инкубирование в течение 24 часов при 37oС, ни проведение от одного до трех циклов замораживания/оттаивания не изменили уровень детектируемого ICAM-4 в сыворотке.
Доноры с эпилепсией
Концентрацию IСАМ-4 в пробах сыворотки, взятых от двадцати пациентов с височной эпилепсией (ВЭ), измеряли и сравнивали с пробами сыворотки, взятыми от контрольной группы пациентов, которые участвовали в экспериментах "Большие эпилептические припадки" (38 различных пациентов) и "Синкопе" (8 пациентов), или от нормальных здоровых доноров (20 индивидуумов). В этом анализе снова использовали метод, описанный в Примере 15, и результаты выражали как нг/мл по отношению к стандарту, используемому в данном анализе, растворимому рекомбинантному белку HulCAM-4 D1-3 (описанному в Примере 13).
Сыворотка от всех 20 пациентов с ВЭ содержала детектируемые уровни ICAM-4, которые в среднем составляли 140 нг/мл. В пробах сыворотки от всех 3 контрольных групп, включая группы "Большого эпилептического припадка", средняя концентрация ICAM-4 была ниже 10 нг/мл. Эти наблюдения дают основание предположить, что уровень ICAM-4 в сыворотке для отдельного индивидуума может служить биохимическим показателем, способным дифференцировать фокальные эпилептические припадки, подобные тем, которые наблюдались в экспериментах с ВЭ, и более генерализованные большие эпилептические припадки.
Доноры со СПИД'ом
Была также исследована концентрация ICAM-4 в сыворотках, взятых от ограниченного числа пациентов со СПИД'ом. Эти пациенты были разделены на группы в соответствии с оценкой CD4 и в соответствии с присутствием любых признаков деменции. Первая группа содержала шестнадцать пациентов на ранней стадии, у которых отсутствовали симптомы заболевания и у которых уровень CD4 составлял более 500. Ко второй группе принадлежали семь пациентов на более поздней стадии заболевания, у которых уровень CD4 составлял менее 300; причем признаки деменции у пациентов этой группы обнаружены не были. И к последней группе принадлежали девять пациентов на поздней стадии СПИД'а, у каждого из которых наблюдались признаки деменции.
Эти результаты показали, что пробы сыворотки от четырех из шестнадцати (25%) пациентов на бессимптомной ранней стадии обнаруживали детектируемые уровни растворимого IСАМ-4; три из четырех проб имели концентрацию IСАМ-4, превышающую 500 нг/мл. Четыре из семи (57%) проб сыворотки, взятой у пациентов на более поздней стадии, также были положительными в отношении IСАМ-4, причем две из этих четырех проб имели концентрации IСАМ-4, превышающие 500 нг/мл. Пробы, взятые от пациентов на поздней стадии, обнаруживающих признаки деменции, не содержали детектируемых уровней IСАМ-4. Результаты этого предварительного исследования дают основание предположить, что IСАМ-4 может служить ранним показателем нейродегенеративных расстройств, ассоциированных с деменцией, вызываемой СПИД'ом.
Доноры с другими нейродегенеративными заболеваниями
Результаты исследований сыворотки, полученной от доноров с эпилепсией и СПИД'ом, дают основание предположить, что уровни IСАМ-4 в крови могут служить показателем разрушения нейронов, которые их экспессируют в нормальных условиях. Существует ряд других нервных болезней, при которых также обнаруживается, как часть их этиологии, разрушение ICAM-4-экспрессирующих нейронов, что может приводить к изменениям сывороточной концентрации IСАМ-4 на периферии.
Так, например, болезнь Альцгеймера ассоциируется с обширным разрушением нейронов в областях конечного мозга, в которых экспрессируется IСАМ-4. Оценка уровней IСАМ-4 в сыворотке пациентов с врожденными формами этого заболевания с ранним проявлением, а также пациентов со спорадической формой заболевания, может служить показателем различных стадий заболевания, и тем самым позволяет проводить возможное терапевтическое лечение.
В качестве другого примера можно указать тот факт, что другие корковые деменции, такие как болезнь Пика, болезнь диффузных телец Леви и дегенерация лобной доли, могут быть иногда ошибочно приняты за болезнь Альцгеймера, но эти заболевания могут быть отдифференцированы друг от друга и от болезни Альцгеймера с помощью анализа сыворотки на IСАМ-4. В качестве другого примера можно указать на тот факт, что концентрация IСАМ-4 в сыворотке пациентов, страдающих от подкорковой деменцией, включая болезнь Паркинсона, болезнь Гентингтона и прогрессирующее поражение надъядерной зоны, может быть увеличена в результате общих патологических показаний у этого класса расстройств.
Другим примером может служить тот факт, что ряд первичных психических расстройств, таких как депрессия, шизофрения и психозы, характеризуются отчасти степенью нейродегенерации, которая может быть ассоциирована с детектируемыми уровнями IСАМ-4 в крови.
Другим примером может служить тот факт, что повышенные уровни IСАМ-4 могут ассоциироваться с рядом приобретенных деменций, развивающихся вследствии инфекций, васкулитов, метаболических и алиментарных расстройств (например, недостаточностью поджелудочной железы, недостатком витамина В12), сосудистых заболеваний (обширного инфаркта, лакунарного состояния, болезни Бинсвангера), токсического энцефалита (вызываемого, например, воздействием моноокиси углерода, тяжелых металлов, или другими промышленными загрязнениями окружающей среды) и опухолей.
ПРИМЕР 19
Клонирование и анализ регуляторной ДНК, расположенной выше IСДМ-4 человека
Экспрессия гена IСАМ-4 является пространственно и стадиеспецифически регулируемой, причем эта экспрессия ограничена, главным образом, передней или вентральной областью головного мозга, конечного мозга. В попытке идентифицировать генные последовательности, ответственные за ограниченную регуляцию транскрипции IСАМ-4, была исследована нуклеотидная область от 5'-конца до кодирующих последовательностей IСАМ-4 человека.
Секвенировали BamHI/Pstl-фрагмент в 2607 п.о., полученный из геномного 7,0 T.n.o.-BamHI-фрагмента (описанного в Примере 10), и было обнаружено, что он содержит 1684 нуклеотидов, расположенных выше старткодона ATG. Полная последовательность для этой расположенной выше области представлена в SEQ ID No:33. Что касается положения ICAM-4-кодирующей последовательности, то "А" в старткодоне ATG (пронумерованный в SEQ ID No:33 как нуклеотиды 1685-1687) обозначали нуклеотидом +1, а нуклеотид, расположенный непосредственно за 5' и до нуклеотида А+1, обозначали -1. Таким образом, целая последовательность простирается от нуклеотида -1684 до нуклеотида +3, что соответствует нумерации Списка последовательностей от нукдеотида 1 до нуклеотида 1687.
Исходя из геномной последовательности HulCAM-4 были синтезированы олигонуклеотиды и использованы в PCR для генерирования ДНК-молекул различной длины в вышерасположенной регуляторной области. Каждый олигонуклеотид, представленный в Таблице 3, содержит спейсерную область (показана курсивом) приблизительно в 6-10 п.о. для ферментативного гидролиза PCR-продукта, рестрикционный Nhel- или НindIII-сайт (показаны в рамке), и праймерную последовательность для специфической гибридизации (подчеркнута). Названия этих олигонуклеотидов содержат числа, которые обозначают их расположение внутри расположенной выше регуляторной области. При PCR-амплификации, олигонуклеотиды спариваются, как показано в Таблице 4, с образованием ДНК-фрагментов, содержащих специфические области расположенной выше регуляторной области.
Рестрикционные сайты и спейсерная область, генерированные внутри каждого олигонуклеотида, обеспечивают ферментативный гидролиз и последующее направленное клонирование отдельных PCR-продуктов в основной вектор pGL3 (Promega, Madison, WI), который содержит генрепортер люциферазы, расположенный непосредственно ниже сайта множественного клонирования (MCS). Активность промотора, клонированного в MCS-область этого вектора, регулирует экспрессию гена-репортера люциферазы в трансфецированных клеточных линиях, и излучение света от экспрессированной люциферазы может быть измерено как показатель промоторной активности. Основной вектор pGL3 не имеет промотора, а поэтому он служит в качестве негативного контроля, тогда как вектор pGL3, содержащий промотор SV40, служит в качестве позитивного контроля. Последовательность каждой экспрессирующей конструкции проверяли с помощью рестрикционного анализа и секвенирования ДНК.
Плазмиды, содержащие амплифицированные последовательности, описанные в Таблице 4, переносили в клетки млекопитающих с использованием набора для трансфекции клеток млекопитающих (Transfection MBS Mammalian Transfection Kit, Stratagene, La Jolla, CA) в соответствии с инструкцией производителя. Каждую плазмиду вводили в две различные клеточные линии, клеток-предшественников COS-7 и NT2 (Ntera2/D1) от Stratagene. Клетки COS 7 представляют собой широко используемую обезьянью фибробластподобную клеточную линию, трансформированную вирусом SV40, вследствие чего эта линия стала пригодной для регулирования экспрессии гена под контролем промотора SV40 в клетках, трансфецированных промоторным вектором позитивного контроля pGL3. Клетки-предшественники NT2 представляют собой коммитированную линию нервных клеток-предшественников и, хотя они не экспрессируют ICAM-4, однако они могут быть более характерными для типов клеток, которые экспрессируют ICAM-4.
Каждую лунку 6-луночного плоскодонного планшета для культивирования тканей (Falkon) засевали 2,5•105 клетками. Трансфекции клеток COS 7 и NT2 были проведены последовательно в дубликате с использованием 5 мкг плазмидной ДНК для каждой лунки. Эти клетки культивировали в течение 48 часов при 37oС, лизировали и анализировали на активность люциферазы с использованием системы для анализа люциферазы (Promega).
Результаты эксперимента, систематизированные в Таблице 5, указывают на высокий уровень промоторной активности, присутствующей на участках от -408 до -19 и от -480 до -19 вышерасположенной регуляторной области ICAM-4 в клетках NT2. Поскольку клетки NT2 являются нервными клетками, то они могут экспрессировать некоторые транскрипционные факторы, распознающие промотор ICAM-4, которые не были обнаружены в клетках другого типа. Наиболее высокий уровень промоторной активности в клетках COS был получен с плазмидой, содержащей участок от нуклеотида -560 до нуклеотида -19. Хотя промоторный вектор позитивного контроля pGL3 хорошо "работает" в клетках NT2, однако он обнаруживает очень низкую активность в клетках NT2, что свидетельствует о клеткоспецифической предпопочтительности некоторых промоторных последовательностей.
Поскольку ни клетки COS 7, ни клетки NT2 обычно не экспрессируют IСАМ-4, то тот же самый эксперимент был повторен с использованием первичных культивированных нейронов гиппокампа крысы, которые экспрессируют IСАМ-4 и должны обязательно экспрессировать механизм транскрипции, необходимый для IСАМ-4-промоторной активности. Путем трансфекции отдельных промоторных конструкций, описанных в настоящей заявке, а также в других заявках, в более естественное окружение можно будет более точно идентифицировать те нуклеотиды в вышерасположенной регуляторной области, которые ответственны за жесткую регуляцию гена IСАМ-4 в головном мозге.
Вышеупомянутые иллюстративные примеры относятся к предпочтительным вариантам осуществления изобретения, в которые, как очевидно для каждого специалиста, могут быть внесены различные модификации и изменения. Таким образом, настоящее изобретение ограничено лишь объемом формулы изобретения, сформулированной ниже.


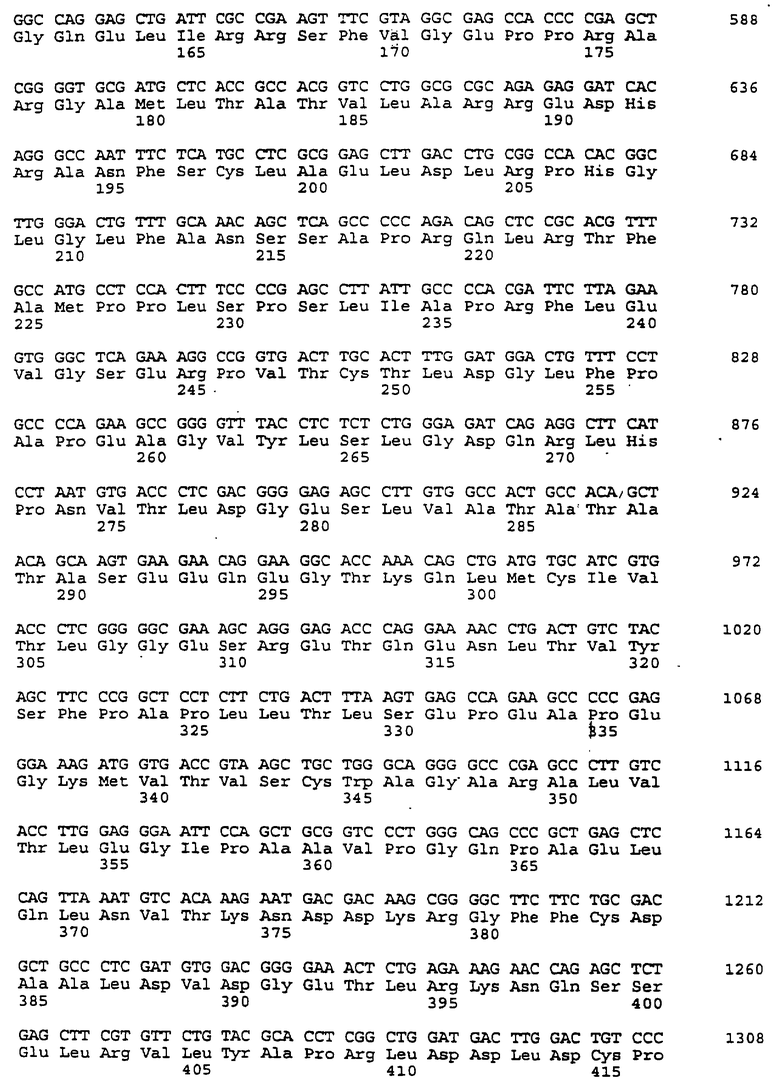
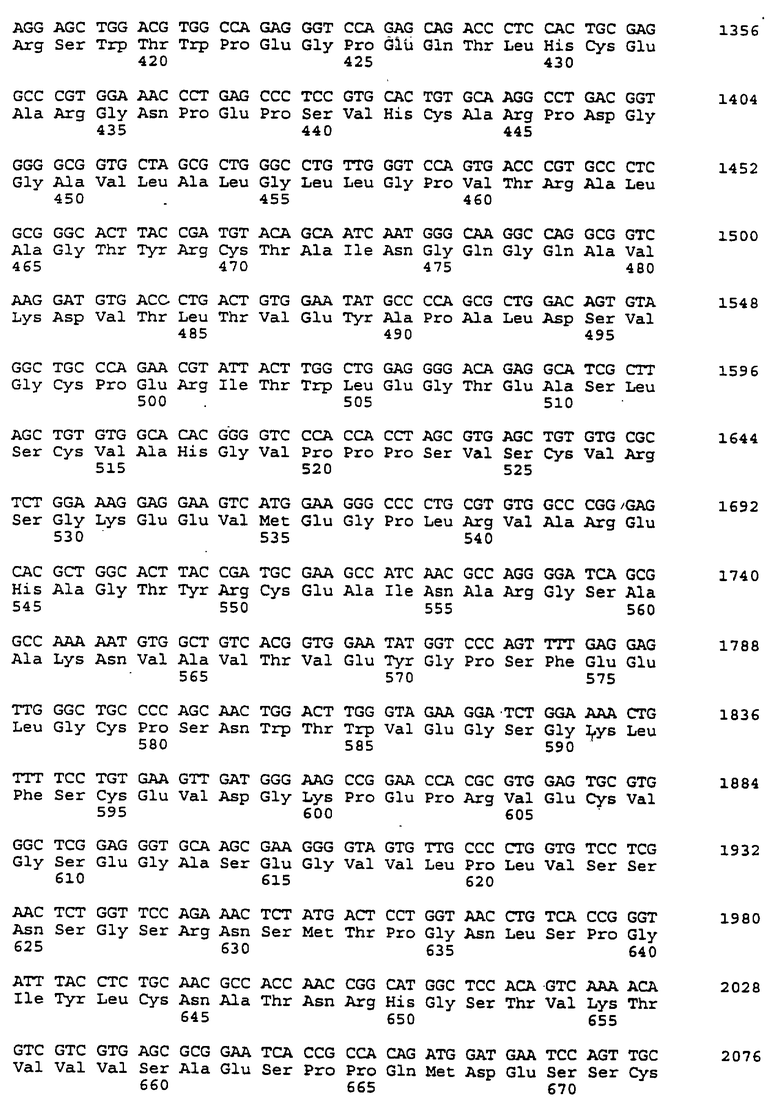
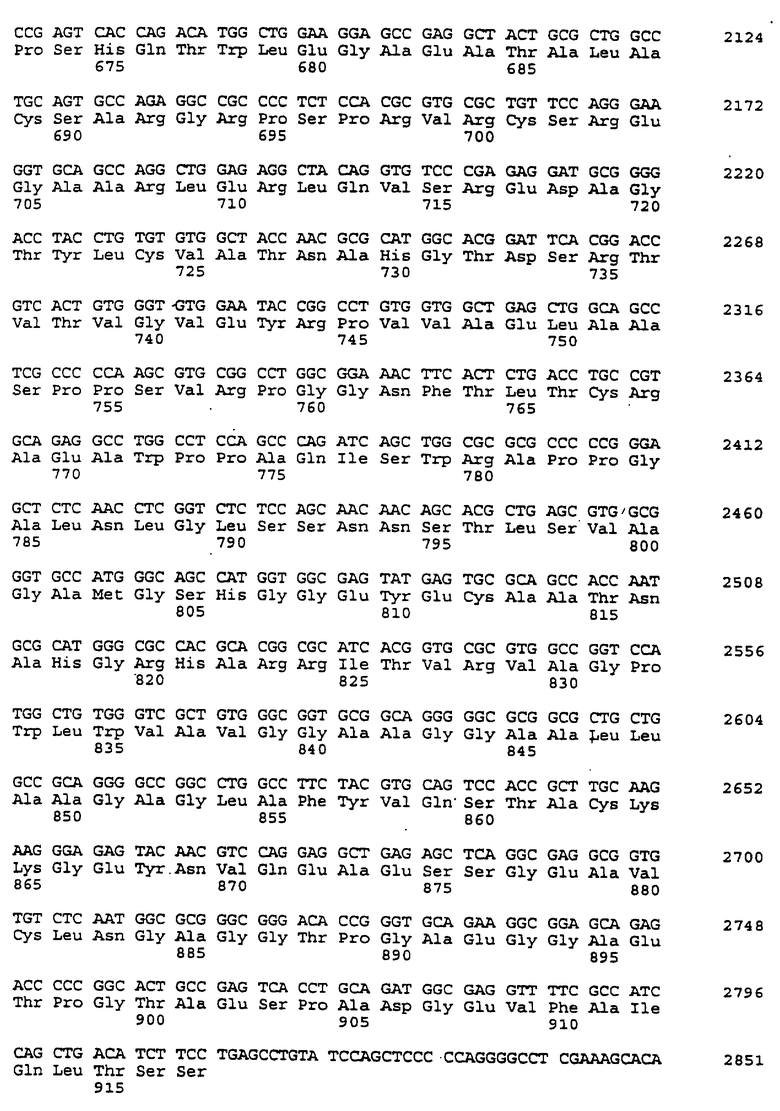




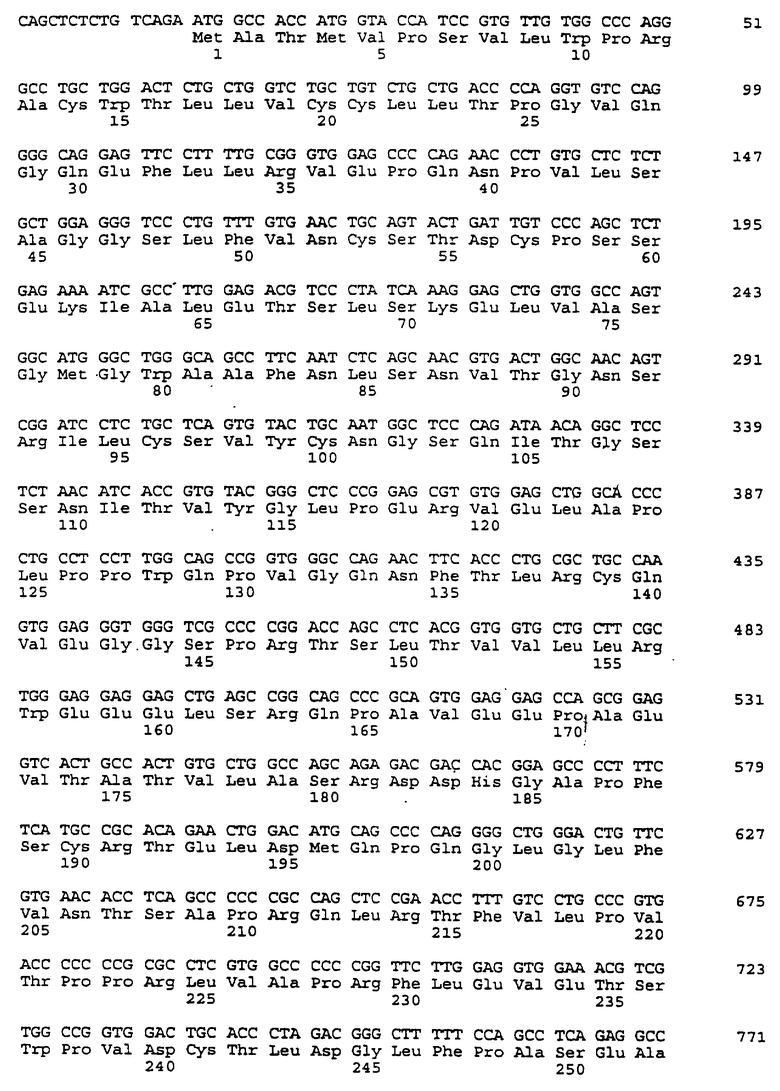
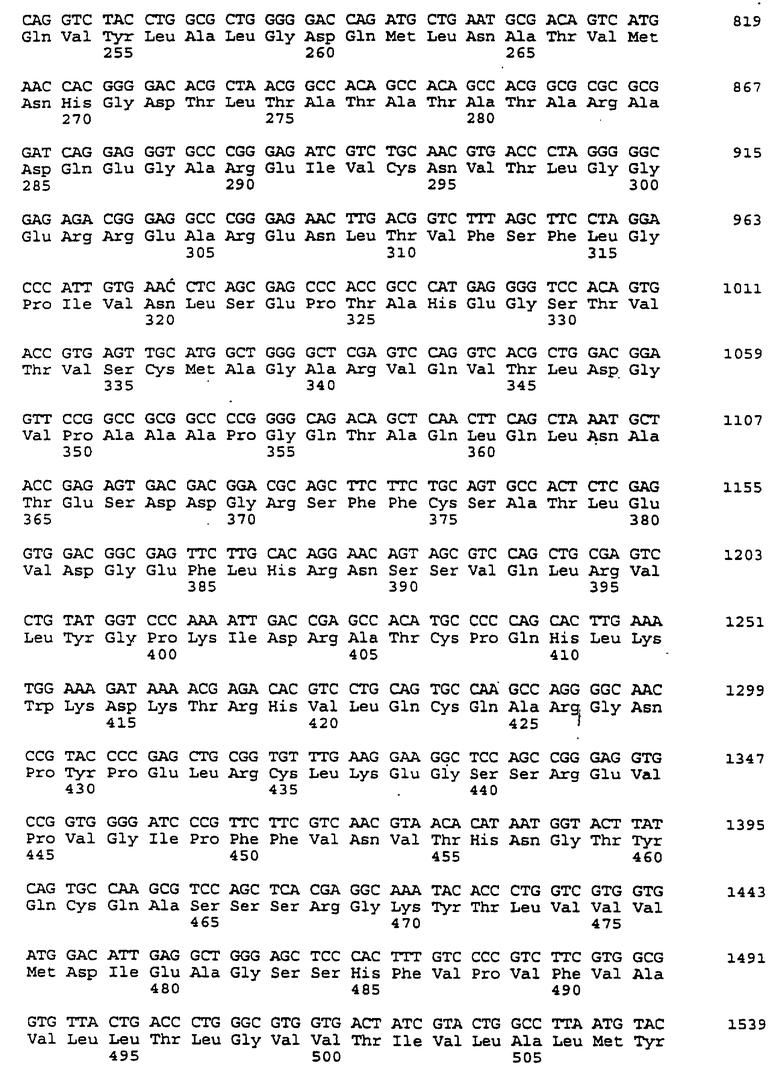




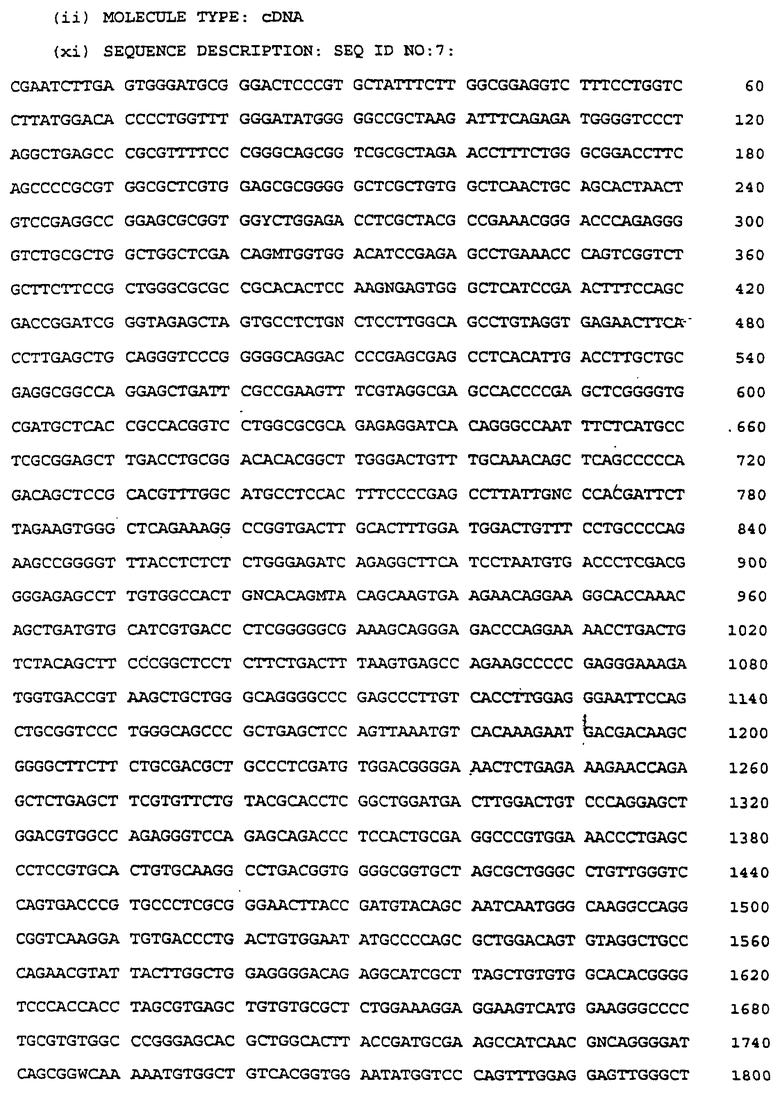










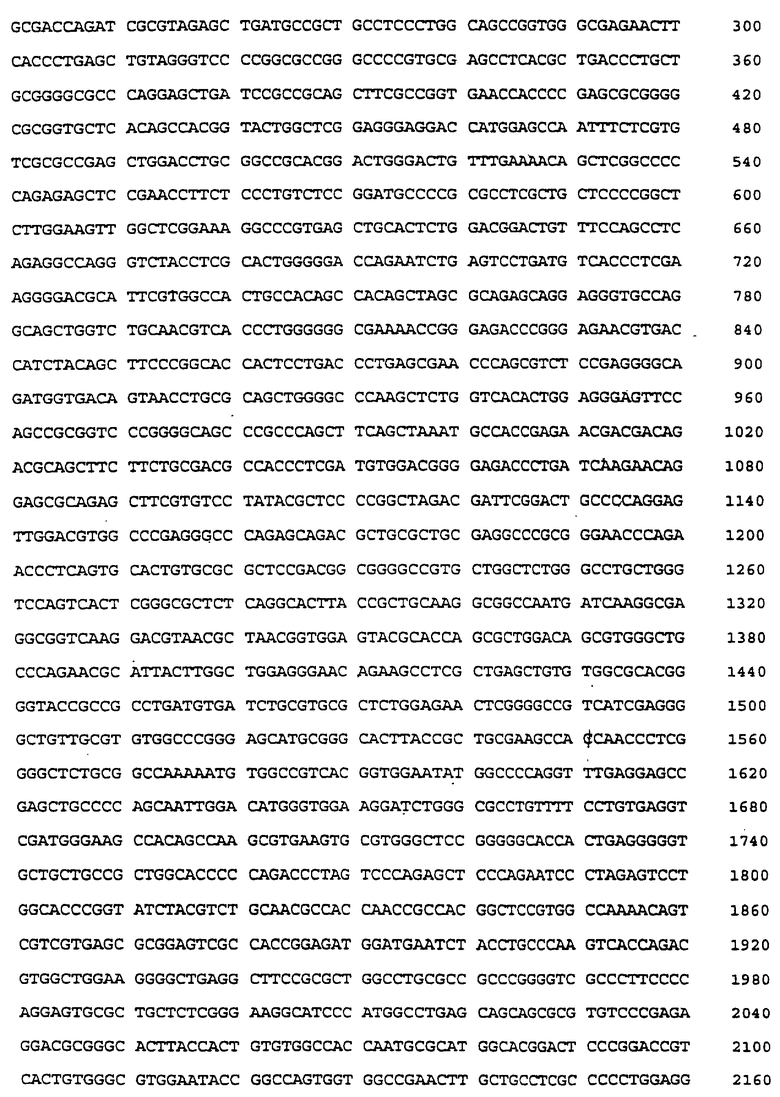


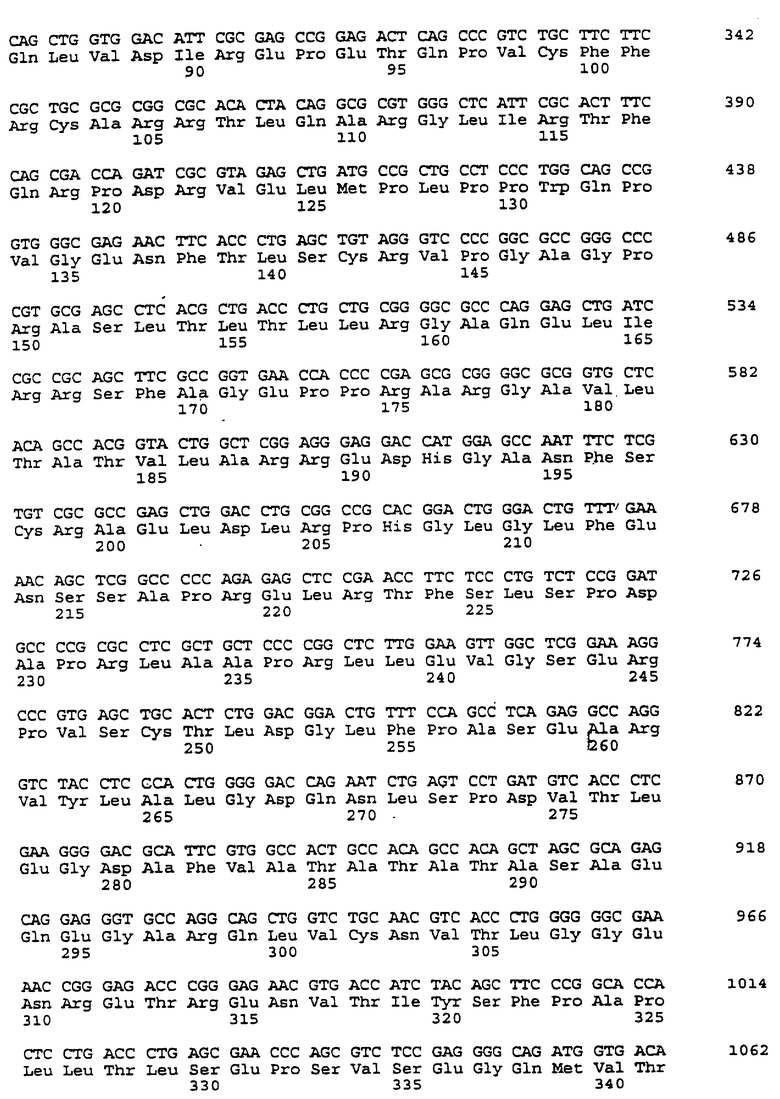
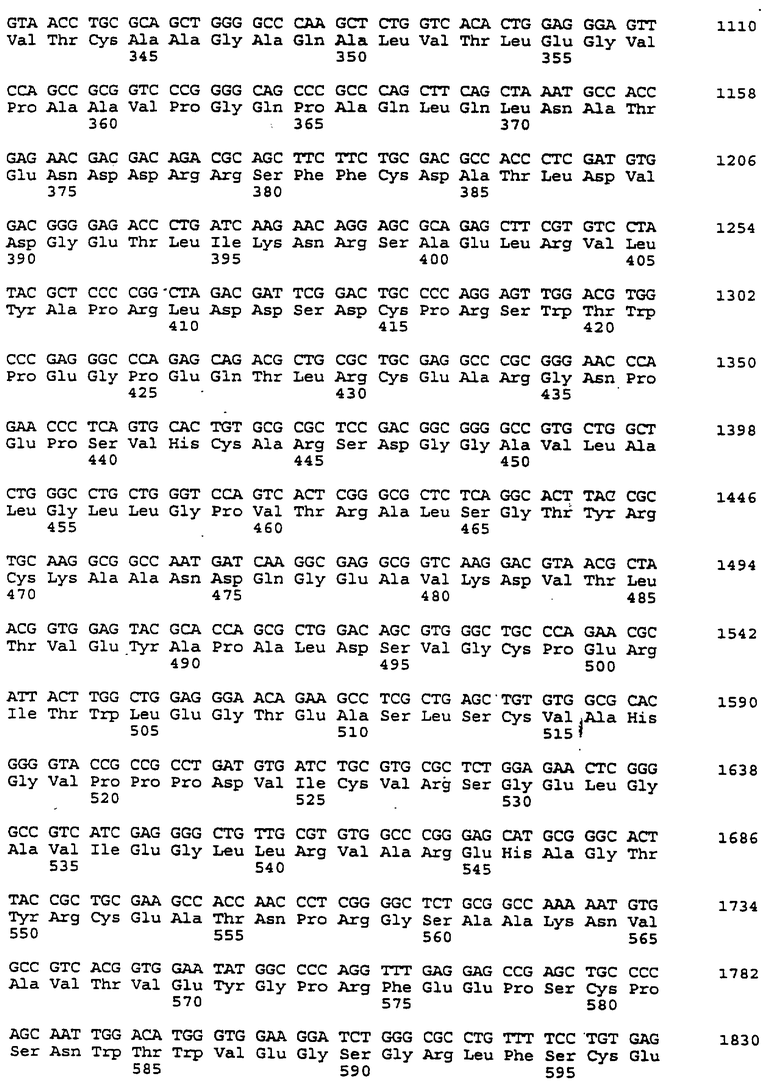













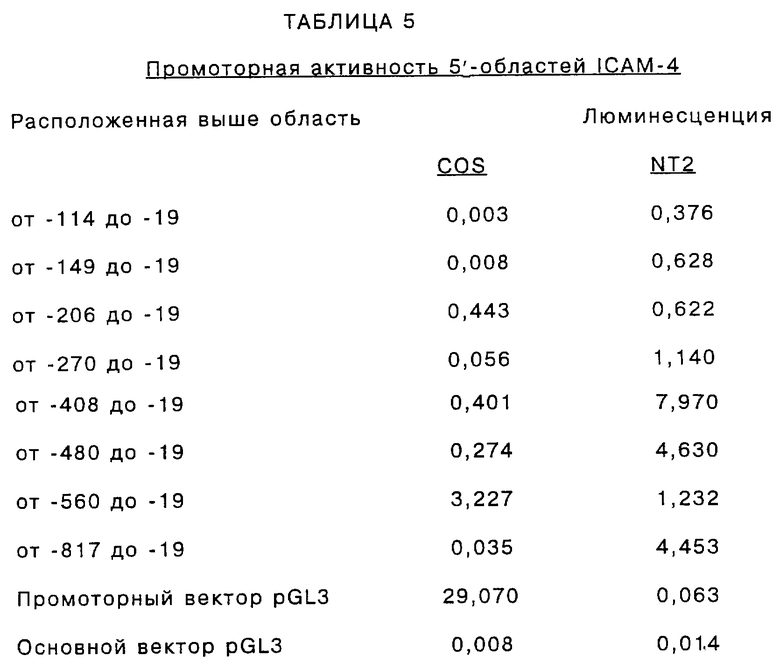
Изобретение относится к медицине, а именно к диагностике нейропатологий с помощью нового полипептида, обозначенного как ICAM-4. Сущность изобретения включает способ диагностики, заключающийся в получении жесткой пробы от индивидуума, контактировании этой пробы с антителом, иммунореактивным к ICAM-4, количественной оценке уровня связывания ICAM-4 антитела и его сравнения по отношению к контролю. Преимущество изобретения заключается в ускорении метода диагностики нейропатологии. 9 з.п.ф-лы, 6 табл.
| АППАРАТУРА ДЛЯ БОКОВОГО КАРОТАЖНОГО ЗОНДИРОВАНИЯ | 0 |
|
SU379904A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| JP 6209788 A, 02.08.1994 | |||
| YOSHIRO et | |||
| al | |||
| An ICAM-related neuronal glycoprotein telencephalin with, brain segment specific expression, Neuron, 1994, v.12, pp.543-553. | |||

