Данная заявка является частичным продолжением находящейся в процессе одновременного рассмотрения заявки на патент США с порядковым номером 08/503226, поданной 17 июля 1995 года, которая, в свою очередь, является частичным продолжением заявки с номером 08/404731, поданной 15 марта 1995 года, которая, в свою очередь, является частичным продолжением находящейся в процессе одновременного рассмотрения заявки на патент США с номером 08/344227, поданной 23 ноября 1994 года.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится в общем к регуляции фосфатазной ферментативной активности кальцинейрина и модуляции экспрессии интерлейкина 2 Т-клетками. Более конкретно, данное изобретение относится к ингибированию фосфатазной активности кальцинейрина некоторыми пептидами и усилению экспрессии Т-клетками интерлейкина 2 посредством обработки этих клеток некоторыми другими пептидами.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Кальцинейрин является Са2+/кальмодулин-зависимой протеинфосфатазой и участвует во многих внутриклеточных путях передачи сигналов. Guerini and Klee, Proc. Natl. Acad. Sci. USA 86:9183-9187 (1989). Этот фермент был идентифицирован в эукариотических клетках в диапазоне от дрожжей до млекопитающих. Cyert and Thorner, J. Cell. Biol., 107:841a (1989) и Klee et al., Adv. Enzymol. , 61: 149-200 (1984). Поскольку кальцинейрин может участвовать во многих путях передачи сигналов в одной и той же клетке, должны существовать некоторые средства специфического нацеливания активности кальцинейрина. Одним из клеточных средств для специфического нацеливания активности фермента является нацеливание посредством компартментализации. Компартментализация разделяет пути передачи сигналов и способствует специфичности клеточных ответов на различные стимулы. Компартментализация определенных ферментов осуществляется посредством взаимодействия этих ферментов со специфическими якорными белками. Например, цАМФ-зависимая протеинкиназа (ПКА) закрепляется в специфических внутриклеточных сайтах посредством связывания с якорными белками А-киназы (АКАР). Поскольку было показано, что АКАР связывает и белки, иные, чем ПКА, это семейство белков называется здесь обычно якорными белками. Hirsch et al., J. Biol. Chem., 267:2131-2134 (1992). цАМФ активирует ПКА связыванием с регуляторными субъединицами (Р) покоящегося голофермента ПКА и вызывает высвобождение активной каталитической субъединицы (К). Существуют два класса субъединицы Р; PI и PII, которые образуют голоферменты ПКА типа I и типа II. Субклеточные распределения этих изоформ ПКА, по-видимому, являются различными. Имеется сообщение, что изоформы PI (PIα или PIβ) являются преимущественно цитоплазматическими и не присутствуют в ядерном компартменте, тогда как изоформы PII (PIIα и PIIβ) являются частицами и связаны либо с плазматической мембраной, компонентами цитоскелета, секреторными гранулами, аппаратом Гольджи, центросомами, либо, возможно, с ядрами.
Якорные белки были обнаружены в разнообразных организмах. Были идентифицированы по меньшей мере семь белков, которые связывают регуляторную субъединицу ПКА в Aplysia californica, морском беспозвоночном животном. Cheley et al., J. Biol. Chem., 269:2911-2920 (1994). Один из таких белков присутствует в большом количестве в неочищенных мембранных фракциях и стабилизированных таксолом микротрубочках и может, следовательно, прикреплять микротрубочки к клеточной мембране, а также связывать ПКА. Был идентифицирован якорный белок млекопитающих, который связан с микротрубочками; ассоциированный с микротрубочками белок 2 (МАР2) присоединяет ПКА к цитоскелету. Threurkauf and Valee, J. Biol. Chem., 257:3284-32-90 (1982) и DeCamilli et al. , J. Cell. Biol. 103:189-203 (1986). ПКА-связывающий сайт на МАР2 представляет собой пептид из 31 остатка в аминоконцевом районе этой молекулы. Rubino et al. , Neuron, 3: 631-638 (1989) и Obar et al., Neuron, 3:639-645 (1989).
Другой якорный белок, который связан с микротрубочками, АКАР 150, накапливается в дендритах в тесной связи с микротрубочками. Glantz et al., Mol. Biol. Cell, 3:1215-1228 (1992). АКАР 150 присутствует в нескольких типах нервных клеток и является членом семейства якорных белков, которые являются основными якорными белками в мозге млекопитающих. Другие члены этого семейства включают в себя АКАР 75, обнаруженный в бычьем мозге, и АКАР 79, обнаруженный в мозге человека. Glantz et al., J. Biol. Chem., 268:12796-12804 (1993). По-видимому, АКАР 75 связывает элементы цитоскелета через два несмежных района вблизи N-конца АКАР 75. АКАР 79 присутствует преимущественно в постсинаптических плотностях (PSD) в переднем мозге человека. Carr et al., J. Biol. Chem., 267:16816-16823 (1992).
Были также охарактеризованы другие якорные белки. Было показано, что выдерживание зернистых клеток яичника с фолликулостимулирующим гормоном и эстрадиолом положительно регулирует экспрессиию АКАР 80 кДа. Carr et al., J. Biol. Chem. , 268:20729-20732 (1993). Другой АКАР, Ht31, был клонирован из библиотеки кДНК щитовидной железы человека. Carr et al., J. Biol. Chem., 267: 13376-13382 (1992). Другой якорный белок, АКАР 95, меняет свою внутриклеточную локализацию во время клеточного цикла. АКАР 95 является интегральным ядерным белком во время интерфазы, но становится связанным с цитоплазматической ПКА при разрушении ядерной мембраны во время митоза. Это предполагает, что АКАР 95 мог бы играть роль в нацеливании активности определенных изоформ ПКА во время отвечающих на цАМФ событий, связанных с клеточным циклом. Coghlan et al., J. Biol. Chem., 269:7658-7665 (1994). Другие известные якорные белки включают в себя АКАР 85 кДа, который связывает ПКА с аппаратом Гольджи (Rios et al., EMBO J., 11:1723-1731 (1992)), и АКАР 350 кДа, который связывает ПКА с центромерами (Keryer et al., Exp. Cell Res., 204:230-240 (1993)).
Известные якорные белки связывают ПКА по обычному механизму. Хотя первичная структура якорных белков не является консервативной, каждый имеет мотив вторичной структуры, который включает в себя амфипатический спиральный район. Scott and McCartney, Mol. Endo., 8:5-11 (1994). Связывание якорных белков с регуляторной субъединицей ПКА блокируется пептидом, который имитирует эту спиральную структуру ПКА-связывающего района якорных белков. Разрушение спиральной структуры этого пептида аминокислотной заменой устраняет блокирование связывания ПКА с якорным белком (Carr et al., J. Biol. Chem., 266: 14188-14192 (1991)), показывая, что связывание ПКА происходит в амфипатической спирали якорных белков и управляется вторичной структурой молекул якорных белков. Это внутриклеточное связывание и локализация ПКА якорными белками обеспечивает средство для расхождения киназы, которая, подобно кальцинейрину, является общей для многих путей передачи сигналов, но все же может действовать специфическим в отношении конкретного пути образом.
ПКА функционирует во многих внутриклеточных путях. Например, было показано, что ингибирование связывания между АКАР 79 и ПКА в нейронах гиппокампа ингибирует рецепторы альфа-амино-3-гидрокси-5-метил-4-изксазолпропионовой кислоты/каината-глутамата. Rosenmund et al., Nature, 368:853-856 (1994). Это свидетельствует о том, что ПКА регулирует эти рецепторы. ПКА регулирует также активность гликогенфосфорилазы обратимым фосфорилированием фермента в ответ на гормонально индуцируемые увеличения внутриклеточного цАМФ. Walsh et al., J. Biol. Chem., 243:3763-3765 (1969). Было также показано, что цАМФ ингибирует передачу сигнала через пути МАР-киназы. Wu et al., Science, 262: 1065-1072 (1993). Это ингибирование опосредовано активацией ПКА, которая ингибирует активацию Raf-1 посредством Ras, блокируя тем самым путь МАР-киназы. Vojtek et al., Cell, 74:205-214 (1993) и Hafner et al., Mol. Cell Biol., 14: 6696-6703 (1994). Эти пути являются важными во многих типах клеток, и предполагалось их участие во многих клеточных функциях, таких как транскрипционная активация гена интерлейкина 2, которая является важной в активации Т-клеток. Weiss and Littman, Cell, 76:263-274 (1994); Owaki et al., EMBO J., 12:4367-4373 (1993).
Подобно ПКА, кальцинейрин связан с активацией Т-клеток. Clipstone and Crabtree, Nature, 357: 695-697 (1992); O'Keefe et al., Nature, 357:692-694 (1992). В Т-клетках кальцинейрин участвует в регуляции экспрессии IL-2 после стимуляции Т-клеток. Weiss and Littman, supra. Было показано, что ядерный фактор активированных Т-клеток (NFATp) является субстратом фосфатазной активности кальцинейрина. Было сделано предположение, что, после стимуляции Т-клеток, опосредованное кальцинейрином дефосфорилирование NFATp делает возможной транслокацию NFATp из цитоплазмы в ядро, где NFATp взаимодействует с Fos и Jun с индукцией экспрессии гена IL-2. Jain et al., Nature, 365:352-355 (1993).
Роль кальцинейрина в активации Т-клеток обеспечивает мишень для терапевтического вмешательства в Т-клеточно-опосредованные нарушения, и были разработаны многочисленные лекарственные средства для ингибирования кальцинейрина. Два кальцинейрин-ингибирующих лекарственных средства, циклоспорин А (циклоспорин) и FK506, использовали в клинических условиях. Thomson and Starzl, Immunol. Rev., 136:71-98 (1992). Как циклоспорин, так и FK506 ингибируют кальцинейрин только после связывания с другими внутриклеточными белками, известными как иммунофилины (циклофилин и FKBP 12 соответственно). Schreiber and Crabtree, Immunology Today, 13:136-142 (1992). Таким образом, циклоспорин и FK506 действуют как пролекарства. После связывания с их соответствующими иммунофилинами комплексы лекарственное средство/иммунофилин связывают кальцинейрин, ингибируя посредством этого фосфатазную активность.
Ингибирование кальцинейрина наиболее эффективно использовали в лечении отторжения трансплантата после трансплантации органов. Циклоспорин и FK506 использовали после пересадки почек, печени, сердца, легкого и костного мозга. Canadian Multicentre Transplant Study Group, N. Engl. J. Med., 314: 1219-1225 (1986); Oyer et al. , Transplant Proc., 15:Suppl 1:2546-2552 (1983); Starzl et al. , N. Engl. J. Med., 305:266-269 (1981); The Toronto Lung Transplant Group, JAMA, 259:2258-2262 (1988); и Deeg et al., Blood, 65: 1325-1334 (1985). Применение этих лекарственных средств значимо удлиняло выживание трансплантата и уменьшало смертность после трансплантации. Najarian et al. , Ann. Surg., 201:142-157 (1985) и Showstack et al., N. Engl. J. Med., 321:1081-1092 (1989).
Циклоспорин использовали также в различных связанных с аутоиммунитетом заболеваниях. Увеит обычно дает улучшение в пределах нескольких недель терапии, но быстро дает рецидив после прекращения введения циклоспорина. Nussenblatt et al., Am. J. Ophthalmol., 96:275-282 (1983). Подобным образом, псориаз обычно обнаруживает улучшение при терапии циклоспорином, но быстро дает рецидив после лечения. Ellis et al., JAMA, 256:3110-3116 (1986). Периоды "медового месяца" инсулиннезависимости могут быть индуцированы и пролонгированы при возникновении сахарного диабета как типа I, так и типа II при введении циклоспорина в пределах двух месяцев инсулиновой терапии. Feutren et al., Lancet, 2:119-124 (1986) и Bougneres et al., N. Engl. J. Med., 318: 663-670 (1988). Различные нефропатии, в том числе очаговые и сегментальные с минимальными изменениями, мембранные и IgM-опосредованные нефропатии, также могут быть чувствительными к циклоспорину, хотя наблюдаемые уменьшения в протеинурии могут быть обусловлены уменьшением скорости гломерулярной фильтрации, а не заживалением базальной мембраны. Tejani et al., Kidney Intl. , 29: 206 (1986). Введение циклоспорина оказывает также зависимое от дозы действие на ревматоидный артрит, хотя такое лечение связано с высокой частотой нефротоксичности. Forre et al., Arthritis Rheum., 30:88-92 (1987).
Как упоминалось выше, циклоспорин был связан с нефротоксичностью. Mason, Pharmacol. Rev., 42:423-434 (1989). Пониженная почечная функция встречается фактически у всех пациентов, которых лечат циклоспорином. Обычно это может быть устранено прекращением циклоспориновой терапии. К сожалению, у реципиентов органов-трансплантатов замена циклоспорина другими обычно используемыми иммуносупрессорами несет большой риск отторжения трансплантата. У пациентов с трансплантированной почкой это может потребовать повторного установления диализа. В случае пациентов, подвергшихся пересадке сердца, легкого или печени, отторжение трансплантата может быть фатальным. Хотя и менее обычно, чем нефротоксичность, нейротоксичность и гепатотоксичность также связаны с циклоспориновой терапией. De Groen et al., N. Engl. J. Med., 317:861-866 (1987) и Kahan et al., Transplantation, 43:197-204 (1987).
Значительная токсичность стала также очевидной в использовании FK506. Подобно циклоспорину, FK506 связан с нефротоксичностью. Peters et al., Drugs, 4: 746-794 (1993). Клиническая картина, морфология повреждений и частота встречаемости приблизительно эквиваленты этим параметрам циклоспорина. McCauley, Curr. Op. Nephrol. Hyperten., 2:662-669 (1993). Нейротоксичность была также связана с FK506. Eidelman et al., Transplant. Proc., 23: 3175-3178 (1991) и Fung et al., Transplant. Proc., 23:3105-3108 (1991). В противоположность циклоспорину, FK506 обладает скорее гепатотрофическим, чем гепатотоксическим действием. Peters et al., supra.
Ввиду значительной потенциальной токсичности иммуносупрессивных агентов, таких как циклоспорин и FK506, ясно, что существует потребность в данной области в отношении дополнительных агентов, ингибирующих кальцинейрин. Эти агенты предпочтительно должны быть ассоциированы с меньшими токсичными побочными действиями, чем доступные в настоящее время агенты, и, следовательно, могли бы обеспечить прогресс в иммуносупрессивной терапии. Кроме того, существует потребность в агентах, которые ингибируют ПКА в Т-клетках, делая возможной повышенную экспрессию интерлейкина 2 этими клетками.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение основывается, отчасти, на открытии того, что кальцинейрин связывает АКАР 79. Связыванием как ПКА, так и кальцинейрина АКАР 79 ко-локализует киназу и фосфатазу, что может регулировать поток сигналов через специфический путь передачи сигналов. Таким образом, данное изобретение обеспечивает композиции и способы для выделения кальцинейрина, а также для ингибирования активности кальцинейрина в клетке. Способы выделения предусматривают контактирование клеточной фракции с АКАР 79 или его кальцинейринсвязывающим фрагментом, который был иммобилизован на твердом субстрате, и затем элюцию из него кальцинейрина. Способы ингибирования кальцинейрина предусматривают контактирование клетки с АКАР 79 или его кальцинейринсвязывающим фрагментом. Предпочтительно, кальцинейринсвязывающий пептид не связывает также и ПКА. Предпочтительные пептиды содержат следующую аминокислотную последовательность:
Arg-Arg-Lys-Arg-Ser-Gln-Ser-Ser-Lys-Glu-Glu-Lys-Pro
(SEQ ID NO:1)
Альтернативные пептиды, используемые в применении на практике способов ингибирования кальцинейрина данного изобретения, включают в себя:
Arg-Arg-Lys-Arg-Ser-Gln-Ser-Ser-Lys-Glu-Glu-Lys-Pro-Leu-Gln
(SEQ ID NO:2)
и
Arg-Arg-Lys-Arg-Ser-Gln-Ser-Ser-Lys-Glu-Glu-Lys-Pro-Phe-Lys
(SEQ ID NO:3).
Эти пептиды являются гомологичными аминокислотным последовательностям АКАР 79, связывающим кальцинейрин. Хотя эти пептиды подобны кальцинейринсвязывающему району FKBP12, в отличие от ингибирования кальцинейрина комплексом FK506/FKBP12, эти пептиды ингибируют активность кальцинейрина без необходимости взаимодействия с другой молекулой.
Эти пептиды могут быть модифицированы для облегчения поступления в клетку, например, сопряжением с растворимой в липидах частью молекулы. Например, пептиды могут быть конъюгированы с миристиновой кислотой. Альтернативно эти пептиды могут быть упакованы в липосомы, которые могут сливаться с клеточными мембранами и высвобождать эти пептиды в клетки.
Другим аспектом данного изобретения являются способы определения, содержит ли клетка кальцинейринсвязывающий и ПКА-связывающий якорный белок. Эти способы обычно предусматривают лизис клетки с образованием лизата; инкубирование лизата с твердым носителем, имеющим иммобилизованные на нем молекулы кальцинейрина; вымывание лизата из твердого носителя; контактирование твердого носителя с меченой регуляторной субъединицей ПКА, вымывание несвязавшейся регуляторной субъединицы из твердого носителя; детектирование метки, остающейся на твердом носителе; и определение на основе этого детектирования присутствия кальцинейринсвязывающего и ПКА-связывающего якорного белка в клетке. Альтернативно регуляторная субъединица ПКА может быть иммобилизована на твердом носителе, а кальцинейрин может быть меченой молекулой. Обычно регуляторная субъединица ПКА будет субъединицей РII.
Эти способы применимы для идентификации дополнительных белков, связывающих как ПКА, так и кальцинейрин. Идентификация других подобных белков может обеспечить тканеспецифические мишени для терапевтического вмешательства.
Данным изобретением рассматриваются также способы для идентификации соединений, которые модулируют связывание между кальцинейрином и якорным белком кальцинейрина. Либо кальцинейрин, либо якорный белок может быть связан с твердым носителем. Несвязанный патнер связывания является детектируемо меченным. Партнеры связывания инкубируют в присутствии тест-соединения. Действие тест-соединения на связывание между кальцинейрином и якорным белком кальцинейрина определяют мониторингом количества метки, связанной с иммобилизованным партнером связывания. Уменьшение количества метки, связанной в присутствии тест-соединения, в сравнении с количеством метки, связанной в отсутствие тест-соединения, указывает на то, что тест-соединение является ингибитором связывания между кальцинейрином и якорным белком кальцинейрина. Могут быть также использованы другие анализы, такие как анализы сцинтилляционной близости.
Дополнительный аспект данного изобретения включает в себя способы усиления экспрессии интерлейкина 2 Т-клетками. Ингибирование киназной активности ПКА или локализации ПКА в Т-клетках усиливает экспрессию белков под контролем промоторных элементов, регулирующих транскрипцию гена интерлейкина 2. Эти способы обычно включают в себя контактирование Т-лимфоцита с одной из следующих аминокислотных последовательностей:
Gly-Arg-Arg-Asn-Ala-Ile-His-Asp-Ile
(SEQ ID NO:5)
или
Asp-Leu-Ile-Glu-Glu-Ala-Ala-Ser-Arg-Ile-Val-Asp-Ala-Val-Ile- Glu-Gln-Val-Lys-Ala-Ala-Gly-Ala-Tyr
(SEQ ID NO:9).
Пептид SEQ ID NO:5 является пептидом, ингибирующим киназную активность ПКА. Пептид SEQ ID NO:9 является пептидом, который гомологичен ПКА-связывающему району якорного белка НТ31. Эти пептиды могут быть модифицированы для облегчения прохождения в клетки или упакованы в липосомы, как описано выше. Данное изобретение рассматривает различные применения способов с использованием этих пептидов. Например, эти способы могут быть использованы для стимуляции иммунного ответа, для стимуляции активированных Т-клеток для выбранного клонального размножения или для усиления Т-клеточных ответов на экспериментальные стимулы для оценки ранних событий в биологии Т-клеток и активации иммунного ответа.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг. 1А-1В иллюстрируют ингибирование фосфатазной активности кальцинейрина полноразмерным АКАР 79 и кальцинейринсвязывающим фрагментом АКАР 79.
Фиг.2А-2С иллюстрируют субклеточную локализацию ПКА типа II и кальцинейрина, а также ко-локализацию (совместную локализацию) ПКА типа II и кальцинейрина.
Фиг. 3 иллюстрирует гомологию между клоном 11.1 и изоформой 11.1 кальцинейрина человека.
Фиг. 4 иллюстрирует увеличение внутриклеточной концентрации цАМФ, индуцируемое обработкой клеток Jurkat форсколином и IBMX.
Фиг. 5А-5Н иллюстрируют FACS-графики, демонстрирующие действие ингибирования и делокализации ПКА на транскрипцию белков под контролем промотора интерлейкина 2.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Пептиды, применяемые в способах данного изобретения, могут быть синтезированы в растворе или на твердом носителе в соответствии с общепринятыми способами, как описано в Stewart and Young, Solid Phase Peptide Synthesis, 2nd ed. Pierce Chemical Company (1984) или Tam et al., J. Am. Chem. Soc., 105: 6442 (1983). Обе статьи включены здесь в качестве ссылок. Пептиды могут быть миристоилированы стандартными способами, как описано в работе Eichholtz et al. , J. Biol. Chem., 268:1982-1986 (1993), включенной здесь в качестве ссылки. Инкапсулирование пептидов в липосомы может быть также выполнено стандартными способами, как описано в общем виде в патентах США с номерами 4766046, 5169637; 5180713; 5185154; 5204112 и 5252263 и Патентной заявке РСТ с номером 92/02244, все включены здесь в качестве ссылок.
Нижеследующие примеры приведены в качестве иллюстрации, а не для ограничения. Пример 1 описывает связь кальцинейрина с АКАР 79 и ПКА. Пример 2 относится к ингибированию активности кальцинейрина с использованием пептидов, полученных из аминокислотных последовательностей АКАР 79. Пример 3 относится к субклеточному распределению ПКА типа II и кальцинейрина. Пример 4 описывает дигибридный анализ, демонстрирующий физиологическое связывание между АКАР 79 и кальцинейрином. Пример 5 относится к анализу связывания АКАР 79 и кальцинейрина. Пример 6 описывает применение мутантов кальцинейрина для определения сайта связывания АКАР 79. Пример 7 относится к взаимодействию между АКАР 79 и субъединицей PI ПКА. Пример 8 описывает способ скрининга на ингибиторы компартментализации ПКА. Пример 9 описывает участие якорного белка в модулировании экспрессии IL-2. Пример 10 относится к идентификации других АКАР 79-связывающих белков. Пример 11 описывает взаимодействие между АКАР 79 и ПКС. Пример 12 относится к потенциальному терапевтическому применению якорных белков.
Пример 1
В этом примере продемонстрирована природно встречающаяся связь кальцинейрина с АКАР 79 и ПКА. Таким образом, АКАР 79 функционирует для ко-локализации (совместной локализации) как встречающейся повсюду киназы, так и встречающейся повсюду фосфатазы. Эта совместная локализация может обеспечивать специфическую регуляцию ферментов в путях передачи сигналов через фосфорилирование или дефосфорилирование, осуществляемое этими ферментами.
Иммунопреципитацию кальцинейрина (CaN) из очищенного кальмодулин-агарозой экстракта бычьего мозга получали с использованием аффинно-очищенных антител, специфических либо для CaN А, либо для CaN В, как описано в общих чертах в Harlowe and Lane, Antibodies: A Laboratoty Manual, Cold Spring Harbor Press, Cold Spring Harbor, NY (1988), за исключением того, что была включена конечная очистка с использованием буфера А (10 мМ HEPES рН 7,9, 1,5 мМ MgCl2, 10 мМ KCl, 1 мМ ПМСФ и 10 мкМ IBMX)+0,4 М NaCl. ПКА-активность измеряли, как описано в Scott et al., Proc. Natl. Acad. Sci. USA, 82:4379-4383 (1985), включенной здесь в качестве ссылки, после элюции иммунопреципитата 0,1 мМ цАМФ. Фосфорилирование иммунопреципитированных белков инициировали добавлением 0,1 мМ 32Р-АТФ (1,5•105 имп/мин/нмоль) и, после 30 мин при 30oС, реакции останавливали добавлением буфера для нанесения с ДСН и подвергали электрофорезу на ДСН-ПААГ. Р-субъединицу ПКА очищали из 30-60% (NH4)2SO4-фракции экстракта мозга с использованием цАМФ-агарозы способами, описанными в работе Coghlan et al., J. Biol. Chem., 269:7658-7665 (1994) (включенной здесь в качестве ссылки), за исключением того, что белок элюировали 0,5 мМ пептидом Ht31 (SEQ ID NO:4). Вестерн-блоты и наложения PII ПКА выполняли, как описано в Coghlan et al., supra.
Киназную активность детектировали в очищенном кальмодулином экстракте, и она была увеличенной в 123±3,6 раза (± стандартное отклонение; n=3) в СаN-иммунопреципитате и специфически ингибировалась пептидом, ингибирующим киназную активность ПКА, пептидом ПКИ (SEQ ID NO:5), что указывало на то, что каталитическая (К) субъединица ПКА была компонентом выделенного комплекса. Бычий гомолог АКАР 79 (АКАР 75) и PII, оба субстрата для К-субъединицы, также присутствовали в этом иммунопреципитате и фосфорилировались при добавлении цАМФ и 32Р-АТФ. В дополняющих экспериментах Р-субъединицы ПКА выделяли из неочищенных экстрактов бычьего мозга аффинной хроматографией на цАМФ-агарозе. Обработка аффинной колонки пептидом Ht31 специфически элюировала АКАР 75, отделяя его от цАМФ-связанной PII, и также высвобождала субъединицы как CaN А, так и CaN В. Было обнаружено, что приблизительно 5% тотального CaN, присутствующего в лизате, были связаны с АКАР 75 и PII, как было детектировано на Вестерн-блотах. В целом, эти результаты предполагают одновременную связь ПКА и CaN с якорным белком.
Пример 2
Данный пример демонстрирует ингибирование фосфатазной активности кальцинейрина пептидами из АКАР 79.
Для определения, является ли связывание пептида АКАР 79 ингибирующим, активность кальцинейрина (CaN) анализировали в присутствии рекомбинантного АКАР 79. Вкратце, рекомбинантный АКАР 79 экспрессировали в Е. coli, как описано в Carr et al., J. Biol. Chem., 267:16816-16823 (1992), включенной здесь в качестве ссылки. CaN и конститутивно активный укороченный мутант CaN420 (укороченную Са2+/кальмодулин-независимую конститутивно активную форму CaN (Perrino et al., J. Biol. Chem., in press) экспрессировали в клетках Sf9 и очищали на кальмодулин-Сефарозе, как описано в работе Perrino et al., J. Biol. Chem. , 267: 15965-15969 (1992), включенной здесь в качестве ссылки. Фосфатазную активность в отношении 32Р-PII-пептидного субстрата измеряли, как описано в Perrino et al., supra. CaN (30 нМ), кальмодулин (100 нМ) и 32Р-РII-пептид (22 мкМ) инкубировали с белком АКАР 79 и пептидом АКАР 79 (SEQ ID NO:1 - аминокислоты 81-102) в диапазоне концентраций, указанном на фиг.1В. Кальмодулин опускали в анализах CaN420. 32Р, высвобожденный из субстрата, измеряли в трех повторностях проб в трех отдельных экспериментах с использованием сцинтилляционного счета. Константу ингибирования (Кi) рекомбинантного АКАР 79 в отношении CaN определяли посредством линейного регрессионного анализа данных. Величины К, для пептида АКАР 79 оценивали путем определения IC50 с использованием фиксированной концентрации субстрата при Km (42 мкМ).
Фиг. 1А иллюстрирует график Lineweaver-Burk АКАР 79-ингибирования как полноразмерного CaN (Са2+/кальмодулин-зависимого) (кружки), так и CaN420 (квадраты) неконкурентным образом в отношении фосфорилированного РII-пептидного субстрата. Белые символы представляют фосфатазную активность в отсутствие АКАР 79, а черные символы представляют фосфатазную активность в присутствии АКАР 79. Синтетический пептид, соответствующий пептиду АКАР 79, ингибировал как полноразмерный CaN (черные кружки), так и CaN420, тогда как пептид Ht31 не был ингибитором CaN (фиг.1В). Наблюдаемое ингибирование было специфическим для кальцинейрина; пептид АКАР 79 не оказывал значимого влияния на активность протеинфосфатаз 1 (белые ромбы) или 2А (кресты) при таких высоких концентрациях пептида, как 0,4 мМ. Хотя сайты связывания CaN на АКАР 79 и FKBP-12 являются сходными, их различия могут иметь функциональное значение: FK506 (2 мкМ) не влиял на силу ингибирования и рекомбинантный АКАР 79 не проявлял пептидилпролилизомеразную активность в отношении флуоресцентного пептидного субстрата. Кроме того, субъединица CaN В, которая необходима для взаимодействия FK506/FKBP с субъединицей CaN А, не требуется для взаимодействия АКАР 79 с субъединицей CaN А. Кроме того, хотя взаимодействие FK506/FKBP с CaN А является кальций/кальмодулинзависимым, АКАР 79-ингибирование активности кальцинейрина является кальций/кальмодулиннезависимым. В целом, эти открытия предполагают, что CaN в его неактивном состоянии локализуется (якорным) белком АКАР 79 способом, аналогичным прикреплению связанной с белком ПКА.
Пример 3
Данный пример демонстрирует субклеточное распределение ПКА типа II и кальцинейрина в различных тканях.
Субклеточная локализация многих протеинкиназ и протеинфосфатаз определяется связью с нацеливающими субъединицами. АКАР 79 является новым членом этого класса регуляторных белков, так как он выполняет бифункциональную роль в локализации как ПКА, так и СаN.
Клетки культивировали, фиксировали формалином и иммуноокрашивали, как описано в Rosenmund et al., Nature, 368:853-856 (1994). ФИТЦ-конъюгированные антикозьи вторичные антисыворотки использовали для окрашивания РII. Биотинилированные антикроличьи вторичные антитела и стрептавидин-Техасский красный (Jackson) использовали в окрашивании на СаN. Изображения получали с использованием конфокальной лазерной сканирующей системы Biorad MRC-600 (фильтры А1 и А2) с микроскопом Nikon optiphot 2, снабженным масляно-иммерсионной линзой 60х planappo chromat (1,6 NA). Конфокальные срезы имели абсолютную толщину между 1,5 и 2 мкм.
Гомологи АКАР 79 наблюдались в бычьем, свином, кроличьем и мышином мозге. Это указывает на то, что ко-локализация ПКА и CaN может быть универсальным феноменом, который адаптирует нейроны в отношении специфических событий трансдукции (передачи) сигналов. С использованием иммуноцитохимических способов субклеточное распределение ПКА типа II и CaN исследовали в культивируемых нейронах гиппокампа. Распределения окрашивания для PII (зеленая метка на фиг. 2А) и CaN (красная метка на фиг.2В) были регионально разбросаны и перекрывались в невритах (PII является красным и CaN является зеленым на фиг.2С). Эти открытия согласуются с ко-локализацией ПКА типа PII и CaN якорным белком и предполагают роль этого трехкомпонентного комплекса в регуляции синаптической передачи возбуждения. Это согласуется с экспериментами, демонстрирующими ко-локализацию РII и АКАР 79 в этих клетках, и с исследованиями, показывающими, что АКАР 79, ПКА типа II и CaN являются компонентами постсинаптических плотностей. Потенциальные субстраты для локализованного трехкомпонентного комплекса трансдукции сигналов могут включать в себя АМРА/каинатные рецепторы, которые модулируются нацеленной якорным белком ПКА.
Пример 4
Данный пример демонстрирует взаимодействие между АКАР 79 и кальцинейрином в дрожжевом дигибридном анализе. С использованием АКАР 79 в качестве "приманки" было обнаружено, что кальцинейрин, кодируемый кДНК из библиотеки мышиных Т-клеток, связывается с АКАР 79.
Этот анализ выполняли по существу, как описано в статье Durfee, et al., Genes and Development 7:555-567 (1993), включенной здесь в качестве ссылки. "Мишенью" и "приманкой" были две плазмиды, каждая из которых содержит часть фактора транскрипции Gal-4. Плазмида-"приманка" (pAS1) была плазмидой на основе плазмиды 2 микрон с промотором АДГ, связанным с ДНК-связывающей субъединицей Gal-4 [аминокислоты 1-147, как описано в статье Keegan et al., Science, 231:699-704 (1986), включенной здесь в качестве ссылки], за которым следовали гемагглютининовая (ГА) метка, поликлональный сайт и терминатор АДГ. Отбор поддерживали с использованием среды SC-Trp. Конструкцией-"мишенью" была leu2, плазмида на основе плазмиды 2 микрон, содержащая промотор и терминатор АДГ с доменом II активации транскрипции Gal-4 [аминокислоты 768-881, как описано в статье Ма and Ptashne, Cell, 48:847-853 (1987), включенной здесь в качестве ссылки], за которыми следовал сайт множественного клонирования. Этот вектор, рАСТ, использовали в конструировании библиотеки слитых кДНК мышиных Т-клеток. Saccharomyces cerevisiae y190, используемый в скрининге, конструировали с двумя репортерными генами, интегрированными в его геном. Эти репортерные гены находятся под контролем промотора Gal-4, содержащего сайты связывания Gal-4. Если белки, кодируемые плазмидой-приманкой и плазмидой-мишенью, связываются, субъединицы фактора транскрипции Gal-4 соединяются и функционируют, инициируя транскрипцию репортерных генов.
NcoI/BamHI-фрагмент 1,3 кДа, содержащий кодирующий район АКАР 79, выделяли из каркаса pET11d и лигировали с pAS1 для действия в качестве "приманки" для скрининга. Один мкг этой конструкции трансформировали в y190 МАТа и y190 MATα с использованием стандартного протокола трансформации с ацетатом лития-ПЭГ. Четыре изолята каждого типа спаривания (у190А pAS1 АКАР 79 1-4 и y190α pAS1 АКАР 79 1-4) тестировали на их способность взаимодействовать со слитой (гибридной) конструкцией рАСТ-РII, которая содержит регуляторную субъединицу (аминокислоты 1-89 PII) ПКА. Это достигалось спариванием этих штаммов на YEPD (1% бакто-дрожжевой экстракт, 2% бактопептон, 2% декстроза и 2% бактоагар) в течение ночи при 30oС с последующим отбором на диплоиды на SC-Leu-Trp-чашках. Затем ген lac Z E. coli, действующий в качестве репортера, можно было тестировать на β-галактозидазную активность. Спаренные штаммы переносили методом реплик на SC-Leu-Trp-чашки, на которые накладывали фильтры Hybond-N (Amersham), и выращивали в течение ночи. Фильтры помещали в жидкий азот на одну минуту для раскрытия клеток дрожжей. Диск из бумаги ЗММ насыщали приблизительно 3 мл 0,1% X-gal (5-бром-4-хлориндолил-β-D-галактозидом) в 60 мМ Na2HPO4, 40 мМ NaH2PO4, 10 мМ KCl и 10 мМ MgSO4. Фильтр с лизированными дрожжами помещали на верхнюю сторону диска и давали проявляться при 30oС в течение приблизительно 1-2 ч. Диплоидные штаммы, содержащие слияния как pAS1 AKAP 79, так и рАСТ PII, которые были положительными в отношении β-gal-активности, обнаруживали по приданию дрожжевому "пластырю" синего цвета. В качестве контроля плазмида-приманка AKAP 79 оставалась белой при спаривании с пустой контрольной плазмидой рАСТ.
Обнаружение слитого белка Gal-4 AKAP 79 достигалось выращиванием у190А AKAP 79 (изолятов 1 и 2) и у190а AKAP 79 (изолятов 1 и 2) до плотности клеток 2•107 клеток/мл в 50 мл среды SC-Trp. Клетки осаждали при 3000•g в течение 10 мин и лизировали с 200 мкл стеклянных шариков (бус) (размер 425-600 микрон) в 25 мМ Трисе рН 8, 5 мМ ЭДТА, 5 мМ ЭГТА, 2 мМ о-фенантролине, 1 мМ ДТТ, 25 мкМ 4-(2-аминоэтил)бензолсульфонилфториде-HCl, с молекулярным весом 239,5 (AEBSF), 1 мМ бензанидине, 1 мкг/мл PLACC (пепстатин, лейпептин, апротинин, калпаин I и II) и 20 мкг/мл бестантинового лизисного буфера. Альтернативно клетки встряхивали на вортексе в течение одной минуты и охлаждали на льду в течение одной минуты, всего в течение 24 минут (12 циклов). Определяли концентрацию белка и 30 мкг общего белка наносили на 10% гель для электрофореза в ДСН-ПААГ. Гель переносили во влажном виде на Immobilon-P (Millipore) и детектирование проводили стандартными процедурами с использованием моноклонального антитела 12CF5 против НА (Bab Co., Berkeley, CA) и козьей вторичной антисывороткой против мышиного IgG, конъюгированной со щелочной фосфатазой (Biorad, Hercules, CA). Слитый белок Gal-4 AKAP 79 приблизительно 100 кДа легко детектировался, что указывало на присутствие продукта правильного размера в этих штаммах.
Изолят 1 у190 pAS1 AKAP 79 был выбран для скрининга библиотеки кДНК мышиных Т-клеток в рАСТ. Собирали 500 мл SC-Trp-культуры (OD600 =0,6-0,8), промывали 100 мл дистиллированной воды и повторно осаждали. Осадок помещали в 50 мл LiSORB (100 мМ ацетат лития, 10 мМ Трис рН 8, 1 мМ ЭДТА рН 8 и 1 М сорбит), переносили в колбу на 1 л и встряхивали при 220 об/мин в течение инкубирования 30 мин при 30oС. Затем клетки осаждали и ресуспендировали с 625 мкл LiSORB и держали на льду при получении ДНК.
ДНК получали для трансформации кипячением 400 мкл 10 мг/мл ДНК спермы лосося в течение 10 мин, после чего добавляли 500 мкл LiSORB и давали медленно охлаждаться до комнатной температуры. Добавляли ДНК из Ми-библиотеки Т-клеток (40-50 мкг) из исходного раствора 1 мг/мл. Охлажденную на льду культуру дрожжей разливали в 10 эппендорфовских пробирок с 120 мкл полученной ДНК. Пробирки инкубировали при 30oС при 220 об/мин. Спустя 30 мин 900 мкл 40% ПЭГ3350 в 100 мМ Li-ацетате, 10 мМ Трисе рН 8 и 1 мМ ЭДТА рН 8 смешивали с каждой культурой и возвращали для инкубирования в течение дополнительных 30 мин. Затем эти пробы объединяли и небольшую аликвоту (5 мкл) извлекали для испытания на эффективность трансформации и высевали на чашки с SC-Leu-Trp. Остальные клетки добавляли к 100 мл среды SC-Leu-Trp-His и выращивали в течение 1 ч при 30oС при встряхивании при 220 об/мин. Собранные клетки ресуспендировали в 5,5 мл среды SC-Leu-Trp-His + 50 мМ ЗАТ (3-аминотриазол) и аликвоты по 300 мкл высевали на чашки 150 мм со средой SC-Leu-Trp-His + 50 мМ ЗАТ и оставляли расти в течение 1 недели при 30oС.
Спустя четыре дня титрационные чашки считали и 1,1•105 колоний подвергали скринингу. Крупномасштабные β-gal-анализы проводили на чашках с библиотекой и для отдельных колоний выделяли десять положительных клонов. Одна из этих колоний росла до значительно больших размеров, чем остальные, и она была названа клоном 11.1. Тотальную дрожжевую ДНК получали из этих штаммов и выделяли ДНК плазмиды leu2. Эту "спасенную" плазмиду использовали для повторной трансформации исходного штамма-приманки у190А pAS1 AKAP 79 и у190а. Только клон 11,1 оставался положительным в отношении β-галактозидазной активности в у190А pAS1 AKAP 79, причем у190а, содержащий клон рАСТ 11.1, оставался белым, являясь отрицательным контролем.
Рестрикционное расщепление эндонуклеазой Xhol высвобождало инсерт 2,3 т. п.н., и эту плазмиду секвенировали в прямом и обратном направлениях. Реакции из набора для секвенирования Dye Deoxy Terminator Cycle Sequencing Kit (Applied Biosystems, Inc. Foster City, CA) с применением симметричной полимеразной цепной реакции (ПЦР) на двухцепочечных матрицах анализировали на автоматизированном секвенаторе ABI 373A (Applied Biosystems, Inc.). Последовательность из клона 11.1 обнаружила открытую рамку считывания длиной 487 аминокислот (SEQ ID NO:6), которая была правильно слита с доменом активации Gal-4 рАСТ. База данных последовательностей Национального института здоровья США (NIH) была подвергнута поиску и была обнаружена последовательность, высокогомологичная кальмодулинзависимой протеинфосфатазе человека, кальцинейрину. Компьютерный анализ сопоставления между клоном 11.1 и изоформой А1 человека показал 80%-ную идентичность на уровне нуклеиновых кислот и 93% идентичность на уровне аминокислот (фиг.3). Первые 10 аминокислот и вставка 18 аминокислот в последовательности человека не присутствуют в мышиной последовательности 11.1. Клон 11.1 является близкородственным последовательности β мышиного кальцинейрина А, но явно отличающимся на карбоксиконце. Подобным образом, изоформы кальцинейрина А1 человека и кальцинейрина А2 человека являются высокогомологичными, но отличающимися друг от друга на их 3'-концах.
Специфичность взаимодействия АКАР 79 - кальцинейрин демонстрировали спариванием содержащего кальцинейрин-рАСТ штамма с другими неродственными штаммами-приманками. "Скрещивания" выполняли, как описано выше, со штаммами, содержащими рАS1, слитую с PII (1-89), казеинкиназой 1, фосфодиэстеразой 32 (HDUN2) и АКАР Ht31. β-галактозидазная активность была отрицательной во всех этих диплоидных штаммах.
Пример 5
Для дополнительной оценки природы взаимодействия АКАР 79 с клоном 11.1 конструировали ряд делеционных мутантов кальцинейрина 11.1 и каждую плазмиду тестировали в дигибридной системе.
С использованием одного и того же 5'-олигонуклеотида (олиго) (МН47) и четырех 3'-олигонуклеотидов (МН48, МН49, МН50 и МН51) проводили ПЦР-реакции для амплификации районов кальцинейрина 11.1, кодирующих аминокислоты 1-104, 1-204, 1-312 и 1-400 соответственно. Эти фрагменты расщепляли BglII и клонировали в рАСТ. Ориентацию подтверждали рестрикционным картированием и ПЦР-ошибки определяли автоматическим секвенированием. Плазмиды, которые, как было определено, правильно кодируют желательный делеционный мутант, трансформировали в у190МАТа и y190MATα. Штаммы дрожжей спаривали с y190apAS1 и y190apAS1 АКАР 79 вместе с исходным клоном рАСТ 11.1, кодирующим аминокислоты 1-487 в SEQ ID NO:6. Полученную чашку со спариваниями анализировали на фильтре, как описано выше, и наблюдали, что только слитые белки, кодирующие либо аминокислоты 1-400, либо аминокислоты 1-487, были способны инициировать транскрипцию репортерного гена. Наблюдение, что слитый белок, содержащий аминокислоты 1-312, был неспособен инициировать транскрипцию, указывал на то, что связывание с АКАР 79 требует остатков между аминокислотами 313-400. Ранее было показано, что этот район включает в себя домен связывания FKBP/FK506, а также район связывания кальцинейрина В [Husi, et al., J. Biol. Chem., 269:14199-14204 (1994)].
Для более точного определения аминокислотных последовательностей кальцинейрина, требующихся для связывания АКАР 79, конструировали дополнительные делеционные мутанты и анализировали их на связывание АКАР 79. Экспрессионные конструкции получали с использованием рАСТ, кодирующей домены 332-441, 332-487 и 442-487 кальцинейрина 11.1. Как и прежде, каждую конструкцию секвенировали и определяли экспрессиию правильного мутанта перед трансформацией в дрожжевой штамм pAS1 АКАР 79.
Однако при трансформации не детектировалась экспрессия репортерного гена, что указывало на то, что эти мутанты были неспособны взаимодействовать с АКАР 79. Одним из возможных объяснений отсутствия связывания АКАР 79 является то, что вторичная структура белка, необходимая для связывания, была потеряна этими укороченными клонами или что некоторая часть аминоконцевой последовательности может быть также необходимой для связывания.
Предыдущие наблюдения показывали, что взаимодействие между комплексом иммунофилинов FKBP/FK506 и кальцинейрином А требует кальцинейрина В [Haddy et al. , FEBS 314:37-40 (1992)]. Для определения, участвует ли кальцинейрин В, эндогенно экспрессируемый в штамме дрожжей у190, в наблюдаемом связывании АКАР 79/кальцинейрин А, штамм кальцинейрин В-, названный y153b (Mat а gal14 gal80 his3 trp-901 ade2-101 ura3-52 leu2-3-112+URA::GAL-->lacZ,LYS2::GAL-->HIS3cnb1Δ1::ADE2) использовали для исключения возможности участия кальцинейрина В в связывании кальцинейрин А/АКАР 79. Сначала y153b трансформировали pAS1 и pAS1 AKAP 79 и анализировали на β-gal-активность в отсутствие плазмиды-"жертвы". Экспрессия репортерного гена не детектировалась, что указывало на то, что экспрессия репортерного гена после трансформации клоном 11.1 необязательно является результатом связывания AKAP 79/11.1. Затем плазмиды рАСТ-кальцинейрин 11.1 и рАСТ-кальцинейрин 1-400 отдельно вводили в у153b pAS1 AKAP 79 с использованием стандартных процедур. β-gal-активность наблюдали в штаммах, трансформированных каждой плазмидой, что указывало на то, что взаимодействие между AKAP 79 и кальцинейрином А не требует кальцинейрина В. Этот результат дополнительно предполагает, что связывание комплекса иммунофилинов FKBP/FK506 с кальцинейрином А отличается от связывания AKAP 79.
Пример 6
В попытке более точного определения района связывания AKAP 79 на кальцинейрине 11.1 конструировали дополнительную серию плазмид, кодирующих делеционные мутанты, отличающиеся от описанных выше, или точковые мутации.
А. Концевые делеции
Данный пример демонстрирует, что взаимодействие между AKAP 79 и кальцинейрином 11,1 требует остатков 30-336 кальцинейрина. Вкратце, конструировали праймеры для различных районов кальцинейрина 11.1 для использования в ПЦР-реакциях для создания N-концевых и С-концевых делеций, описанных в таблице. ПЦР-продукты генерировали смешиванием 1 мкг каждого из 3'- и 5'-праймеров с 200 мкг каждого из dNTP и 1 нг плазмидной матрицы с ПЦР-буфером 2 (содержащим 20 мМ Трис-HCl, рН 8,75, 10 мМ KCl, 10 мМ (NH4)2SO4, 2 мМ МgSО4, 0,1% Тритон Х-100 и 100 мкг/мл БСА) (Stratagene) и 2,5 единиц ДНК-полимеразы Perococus furiosus (Pfu) (Stratagene) в реакционном объеме 100 мкл. Проводили тридцать циклов, каждый состоял из одной минуты при 95oС, двух минут при 50oС и четырех минут при 72oС. Продукты амплификации очищали и клонировали в BglII-сайт рАСТ. Полученные конструкции анализировали на ПЦР-ошибки и ориентацию секвенированием, как описано выше.
Каждую конструкцию индивидуально трансформировали в дрожжевые штаммы у190α, у190а pASI AKAP 79 и y153b pASI AKAP 79, каждый из которых описан выше в примере 4А, и тесты на β-галактозидазу на фильтрах выполняли, как описано ранее. Результаты с использованием первого набора векторов, кодирующих С-концевые делеции, определили зону между аминокислотами 312-400, требуемую для связывания AKAP 79. Положительные тесты на фильтрах из трансформантов y153b pASI AKAP 79 также подтвердили, что кальцинейрин В не требовался для связывания AKAP 79.
Предыдущие исследования показали, что связывание кальцинейрина В требует аминокислот 348, 349, 355 и 356 [Watanabe, et al., J. Biol. Chem., 270: 456-460 (1995)], аутоингибирующий домен кальцинейрина включает в себя аминокислоты 442-487 и связывание FKBP/FK506 требует аминокислот 350, 353 и 359 [Kawamura and Su, J. Biol. Chem., 270:15463-15466 (1995)]. Дополнительные конструкции кальцинейрина 11.1, кодирующие дополнительные С-концевые делеции, показали, что связывание комплекса кальцинейрин 11.1/АКАР 79 требовало аминокислот 1-336. Эти делеции демонстрируют, что кальмодулинсвязывающий домен [ГДЕ НАХОДИТСЯ ЭТОТ ДОМЕН?], аутоингибирующий домен и кальцинейрин В-связывающий домен не требуются для AKAP 79 и кальцинейрина А для образования комплекса.
Результаты связывания для всех делеций представлены в таблице. Аминоконцевые делеции показали, что по меньшей мере одна зона, требуемая для связывания АКАР 79, лежит между остатками 30-99. Как и прежде, трансформанты y153b pAS1 АКАР 79, экспрессирующие N-концевые делеции, не требуют кальцинейрина В для связывания.
В. Точковые мутации
Для точной оценки, какие аминокислоты участвуют в связывании АКАР 79, точковые мутации кальцинейрина 11.1 создавали с использованием стратегии на основе ПЦР. Три аланиновых мутанта, Cys335-->Ala, Ser336-->Ala и Pro339-->Ala генерировали и анализировали на модуляцию связывания АКАР 79 в дигибридной системе. Ни один из этих мутантов не предотвращал связывание АКАР 79 с кальцинейрином, свидетельствуя о том, что только модификация этих остатков является недостаточной для нарушения связывания АКАР 79.
Пример 7
Дополнительный скрининг с использованием рАСТ-Mu-библиотеки кДНК мышиных Т-клеток и штамма-приманки pASI АКАР 79 проводили для идентификации других АКАР 79-связывающих белков согласно описанному выше протоколу. Результаты скрининга приблизительно 211000 колоний дали один положительный клон, названный рАСТ 2-1, который оставался положительным после спасения и повторной трансформации. Последовательность библиотеки извлекали из этой плазмиды расщеплением XhoI, и было показано, что она представляла собой инсерт 1200 п.н. Секвенирование и последующий поиск в базе данных показали, что этот клон имел 91%-ную идентичность с регуляторной субъединицей типа 1α крысиной протеинкиназы A (PI).
Библиотеку повторно подвергали скринингу с использованием той же самой АКАР 79-приманки, и были детектированы пятнадцать положительных изолятов из приблизительно 5200000 трансформантов. Было обнаружено, что из этих пятнадцати, одиннадцать были гомологичными регуляторной субъединице типа I крысиной ПКА. Каждый из этих изолятов сливался с 5'-нетранслируемым районом PI и оставался открытым через инициирующий метионин. На основании рестрикционного анализа и данных секвенирования были выделены девять индивидуальных клонов, в том числе первоначальный изолят рАСТ 2-1.
Эти результаты являются первой демонстрацией якорного белка, который связывается с регуляторными субъединицами как PII, так и PI ПКА, что является неожиданным ввиду структурно несходных первичных структур между этими двумя субъединицами.
В попытке дополнительного определения последовательности взаимодействия между PI и АКАР 79 и для определения, является ли это взаимодействие уникальным в отношении АКАР 79, получали новые штаммы дрожжей. С использованием ВglII-сайта в первых 400 п.н. PI выделяли фрагмент, кодирующий аминокислоты 1-80, из плазмиды рАСТ72 и лигировали его с pAS1 и рАСТ. Ориентацию подтвержали рестрикционным анализом. С использованием стандартных процедур трансформации дрожжей ДНК плазмиды вводили в у190 MAT и трансформированные дрожжи анализировали на β-gal-активность. Было определено, что укороченный слитый продукт РI неспособен стимулировать экспрессию репортерного гена. Затем эти трансформированные штаммы использовали в ряде экспериментов для определения, взаимодействует ли укороченная форма РI с АКАР 79.
Экспрессию репортерного гена наблюдали в дважды трансформированном штамме дрожжей, что указывает на то, что для связывания РI/АКАР 79 важны первые 80 аминокислот РI.
Наконец, в попытке определения, является ли способность связывания как субъединицы РI, так и субъединицы РII уникальной в отношении АКАР 79, АКАР щитовидной железы человека [Carr, et al., J. Biol. Chem., 267:133376-133382 (1992)], генный продукт рАСТ Ht31, анализировали при помощи дигибридного теста с вышеописанным укороченным пептидом PI, содержащим аминокислоты 1-80 и кодируемым на плазмиде pAS1 (1-80). Наблюдаемое связывание Ht31/RI, в сочетании с прежним наблюдением, что Ht31 связывает PII, показало, что связывание якорного белка как с PI, так и с PII не является уникальным в отношении АКАР 79.
Пример 8
Ввиду того что было показано, что АКАР 79 связывается с субъединицами как PI, так и PII ПКА, авторы разработали способ скрининга с использованием сцинтилляционной близости для идентификации специфических ингибиторов, которые нарушают локализацию ПКА вмешательством в связывание АКАР 79 с ПКА.
Сначала конструировали экспрессионную плазмиду слитого белка тиоредоксин (TRX)-AKAP 79. См. , в общих чертах, LaVallie, et al., BIO/TECHNOLOGY 11: 187-193 (1993). Вкратце, XbaI/HindIII-фрагмент тиоредоксина субклонировали в pUC19, содержащую ген lac Z и промотор tacZ. Полученная плазмида была названа TRX F/S pUC19. Для встраивания кодирующей последовательности АКАР 79 в TRX F/S pUC19 создавали
NcoI-сайт с олигонуклеотидом (SEQ ID NO:32), имеющим концевые SpeI и HindIII последовательности. После расщепления SpeI/HindIII этот олигонуклеотид встраивали в вектор и NcoI/XhoI-фрагмент, кодирующий АКАР 79, лигировали в рамке считывания с геном тиоредоксина. Слитый белок экспрессировали в Е. coli и иммобилизовали на 96-луночных ScintiStrip-планшетах (Wallac, Tyrbu, Finland), которые содержали сцинтиллятор, заделанный в твердый носитель. Планшеты предварительно покрывали кроличьими антимышиными антителами, которые использовали для иммобилизации мышиного моноклонального антитела, иммуноспецифического для TRX. Затем слитый белок TRX-AKAP 79 прикрепляли на планшетах через антитело против TRX и к планшетам добавляли 3H-РII в присутствии или в отсутствие ссылочного ингибитора, например немеченого РII. При связывании 3H-РII с АКАР 79 метка подводилась достаточно близко к заделанному в носитель сцинтиллятору, приводя к излучению, детектируемому в сцинтилляционном счетчике MicroBeta.
Результаты из этого анализа показали, что немеченый РII и пептид Ht31, описанный выше, были способны ингибировать связывание АКАР 79/PII с IC50 1 мМ и 50 нМ соответственно. Эти результаты являются сходными с сообщенными величинами других якорных белков [Care, et al., J. Biol. Chem., 267:13376-13382 (1992)] . Пролинзамещенный пептид Ht31, также описанный выше, не ингибировал связывание АКАР 79/РII. Поскольку эти результаты согласуются с результатами, наблюдаемыми в предыдущих Вестерн-блоттинге и анализе с наложением фильтров, предполагается, что этот способ сделает возможным быстрый скрининг потенциальных ингибиторов связывания АКАР 79/PII, а также ингибиторов связывания АКАР 79 с другими известными физиологическими партнерами, например кальцинейрином и протеинкиназой С.
Пример 9
Данный пример демонстрирует, что связывание ПКА с якорным белком в Т-клетках модулирует активность ПКА в отношении активации NFAT, модулируя тем самым продуцирование интерлейкина 2.
Экспрессия гена IL-2 тесно связана с активацией Т-клеток. Исследовали транскрипцию IL-2 после активации ФМА и иономицином. Известно, что эти два агента усиливают соответственно реакции вторичных мессенджеров, протеинкиназы С и кальция (в том числе активацию CaN). Протеинкиназа С активирует путь Ras-Raf-1-Mek-MAP-киназы, которая участвует в индукции ядерного компонента NFAT. Увеличенная концентрация кальция активирует кальцинейрин, который, в свою очередь, активирует цитоплазматический компонент NFAT и делает возможным транслокацию в ядро. Эта активация компонентов NFAT индуцирует экспрессию гена IL-2. Для количественного определения транскрипции линию Т-клеток Jurkat (NFATZ) стабильно трансфицировали вектором, содержащим 3 тандемные копии NFAT-связывающего сайта и минимальный промотор IL-2, слитый с геном lacZ, кодирующим β-галактозидазу (β-gal). Количественное определение транскрипции IL-2 достигалось посредством анализа β-gal-активности с использованием клеточного сортера с возбуждением флуоресценции (FASC).
Обычно 1•106 клеток NFATZ в 1 мл культуральной среды предынкубировали в течение 60 мин при 37oС с варьирующими концентрациями циклоспорина и миристилированными пептидами, включающими в себя аминокислоты 81-108 АКАР 79 (SEQ ID NO:8, описанная в статье Glantz et al., J. Biol. Chem., 268:12796-12804 (1993), включенной здесь в качестве ссылки), ПКИ (ингибиторный пептид ПКА (GRRNAIHDI-SEQ ID NO:5)) и пептид Ht31 (SEQ ID NO:9; аминокислоты 493-515 полноразмерного белка Ht31, описанная в Carr et al., J. Biol. Chem., 267: 13376-13382 (1992), включенной здесь в качестве ссылки, которая ингибирует взаимодействие якорного белка с субъединицей PII ПКА). Каждый из пептидов миристилировали, как описано в Eichholtz et al., J. Biol. Chem., 268: 1982-1986 (1993).
В экспериментах с циклоспорином ПКИ (SEQ ID NO:5) и пептидом Ht31 (SEQ ID NO:9) инкубирование с циклоспорином или соответствующими пептидами сопровождали дополнительным 30-минутным инкубированием с форсколином (25 мкМ) и изобутилметилксантином (IBMX; 0,1 мМ). Инкубирование со смесью форсколин/IBMX повышает внутриклеточные концентрации цАМФ (фиг.4), активируя посредством этого ПКА. Наконец, добавляли форбол-12-миристат-13-ацетат (ФМА) (10 нг/мл) и иономицин (2 мкМ) и инкубирования продолжали в течение 4 ч. Контроли инкубировали только с ФМА/иономицином или с форсколином/IBMX и ФМА/иономицином в условиях, описанных выше. Во время последних 20 мин инкубирования с ФМА/иономицином добавляли хлорохин (300 мкМ) для ингибирования эндогенной лизосомной β-gal-активности. Клетки откручивали и ресуспендировали в 50 мкл культуральной среды, к которой добавляли 50 мкл флуоресцеин-ди-β-D-галактопиранозида (ФДГ) (конечная концентрация 0,1 мМ; Molecular Probes). Эту процедуру осмотического шока продолжали в течение 75 с до возвращения клеток в изотонические условия путем добавления 1 мл холодного FACS-буфера (содержащего хлорохин). β-gal-активность lacZ измеряли проточной цитометрией, приспособленной для анализа флуоресцеина.
Фиг. 5А-5Н иллюстрируют результаты данного эксперимента. Фиг.5А и 5В являются FACS-графиками, показывающими флуоресценцию фона данного анализа с добавленным красителем или без него. Фиг. 5С показывает, что обработка ФМА/иономицином клеток Jurkat NFATZ индуцировала увеличение в 6-7 раз β-gal-активности. Циклоспорин (CsA) полностью устранял эту активность, как и ожидалось в случае важной роли передачи сигнала CaN в транскрипции IL-2 (фиг. 5D). Было обнаружено, что миристилированный пептид АКАР 79 (SEQ ID NO:8) при использовании при 10 мкМ в среде уменьшал индуцированную ФМА/иономицином β-gal-активность на 40-50%.
Фиг. 5Е показывает, что форсколин и IBMX уменьшали индуцированную ФМА/иономицином β-gal-активность на 50%. Это ингибирование полностью снималось как 100 мкМ миристилированным пептидом ПКИ (SEQ ID NO:5), так и 100 мкМ миристилированным пептидом Ht31 (SEQ ID NO:9) (фиг.5F и 5G). Фиг.5Н показывает, что миристилированный пептид Ht31 с пролиновой заменой, которая, как известно, делает этот пептид неактивным в ингибировании прикрепления ПКА, не влиял на ингибирование форсколином/IBMX. Эти результаты показывают значение ПКА и ее локализации через якорный белок в регуляции экспрессии гена IL-2. Как описано выше, вмешательство в активность ПКА или ее локализацию может быть использовано для усиления иммунного ответа, активации Т-клеток для селективного клонального размножения или исследования ранних событий активации Т-клеток.
Пример 10
Были идентифицированы два дополнительных уникальных изолята, рАСТ 59 и рАСТ 74, которые кодировали тот же самый район другого белка. Последовательности для этих клонов изображены в SEQ ID NO:33 и 34 соответственно. Результаты BLAST-поиска показали значимую гомологию аминокислот с продуктами трех генов неизвестной функции: С. elegans (белком из 319 аминокислот, названным No. U00032 в списке последовательностей), маркерной экспрессирующейся последовательностью мозга плода человека (белком из 97 аминокислот, названным Т08697) и маркерной экспрессирующейся последовательностью HL60 (белком из 90 аминокислот, названным D2073). Была также обнаружена гомология с продуктом гена S. pombe, названным PAD 1+ (белком из 308 аминокислот, названным D31731), который, как было показано, является положительным регулятором РАР 1+, АР-1-подобного фактора транскрипции.
Кроме того, два других положительных клона были детектированы в этом скрининге; рАСТ 36, которая кодирует открытую рамку считывания 143 аминокислот, правильно слитую с Gal4, и рАСТ 69, которая кодирует немного более короткий район, образованный вследствие явной делеции. Последовательности для этих клонов представлены в SEQ ID NO:35 и 36 соответственно. Эти два изолята отличались друг от друга и не обнаружили идентичности с какой-либо известной последовательностью в базе данных NIH.
Пример 11
Предыдущая работа предполагает, что АКАР 79 представляет собой полифункциональный якорный белок, который способен связываться с по меньшей мере двумя ферментами передачи сигналов, ПКА и Са2+/кальмодулинзависимой фосфатазой, кальцинейрином (CaN). Каждый передающий сигнал фермент связывается с отличающимся районом якорного белка и каждый фермент является ингибированным при этом прикреплении. Кроме того, было показано, что Са2+/фосфолипидзависимая протеинкиназа С (ПКС) также связывается с АКАР 79 в районе, отличающемся от района ПКА и CaN. Подобно ПКА и CaN, активность ПКС ингибируется ее связыванием с якорным белком. ПКС-связывающий сайт содержится в первых 75 остатках якорного белка, и исследования пептидов показали, что фрагмент, содержащий остатки 31-52 АКАР 79, ингибирует активность ПКС. Кроме того, имеющиеся доказательства предполагают, что связывание кальмодулина (СаМ) с якорным белком может высвобождать активность ПКС, что предполагает конкуренцию за последовательность АКАР 79. Для более полной характеристики взаимодействия ПКС с АКАР 79 были предприняты эксперименты для характеристики ПКС-связывающего сайта, выделения комплекса ПКС/АКАР из бычьего мозга и определения, является ли СаМ физиологическим регулятором взаимодействия ПКС/АКАР 79.
Наложение (overlay) ПКС сначала выполняли на лизатах бычьего мозга с использованием ПКС мозга кролика в качестве зонда. Связывание ПКС детектировали моноклональным антителом (М7), которое узнает α- и β-изоформы ПКС. Детектировали несколько ПКС-связывающих белков в диапазоне размеров 50-300 кДа, которые включали в себя белок, мигрирующий с подвижностью, одинаковой с подвижностью четко выделяющегося РII-связывающего белка 75 кДа. Контрольные эксперименты подтвердили, что ПКС-связывание было специфическим и могло быть детектировано только в присутствии 1,2 мМ CaCl2 и 20 мкг/мл фосфатидилсерина и при добавлении ПКС к реакционной смеси.
Для определения, может ли идентифицированный белок 75 кДа быть бычьим гомологом АКАР 79, тест наложения ПКС использовали для зондирования АКАР 79 и родственных фрагментов. Вкратце, белки разделяли электрофорезом в ДСН-ПААГ и блоттировали ("промакали") на нитроцеллюлозу согласно стандартным протоколам. Пробы блокировали в Blotto [1 мг/мл бычьего сывороточного альбумина (БСА), 5% сухого молока в забуференном Трисом солевом растворе (TBS)] и инкубировали в течение одного часа при комнатной температуре в тест-буфере [TBS, содержащем 1 мг/мл БСА, 1,2 мМ кальций, 1 мМ ЭГТА, 20 мкг/мл фосфатидилсерина (ФС), 2 мкг/мл лейпептина, 2 мкг/мл пепстатина и 3 мкг/мл частично очищенной ПКС мозга кролика]. Связавшуюся ПКС детектировали моноклональным антителом М7, которое узнает как ПКСα, так и ПКСβ, согласно стандартным способам детектирования хемилюминесценции.
ПКС связывалась с полноразмерным рекомбинантным белком АКАР 79, и рекомбинантные фрагменты, включающие в себя первые 75 остатков этого белка, связывали ПКС, но С-концевые фрагменты, включающие в себя СаN- и РII-связывающие районы, не связывали ПКС. Контрольные эксперименты показали, что 32Р-радиоактивно меченная РII связывалась как с полноразмерным АКАР 79, так и с С-концевыми фрагментами. Эти результаты показали, что АКАР 79 является ПКС-связывающим белком и что основные сайты связывания находятся в пределах первых 75 аминокислот этого белка.
Предыдущие исследования по ПКС-связывающим белкам предполагали, что основные и гидрофобные районы из ПКС-связывающих сайтов участвуют в образовании фосфолипидного мостика с этим ферментом. Первые 75 остатков АКАР 79 содержат основные и гиброфобные районы между положениями 31-52, и несколько линий доказательств предполагают, что этот район является основным сайтом контакта с ПКС. Синтетический пептид к остаткам 31-52 блокировал взаимодействие ПКС/АКАР 79, как было определено в тесте наложения.
Для оценки способности этих пептидов модулировать активность ПКС следующий анализ проводили в присутствии и в отсутствие пептидных фрагментов АКАР 79. ПКС [50 нМ, растворенную в 50 мМ Трис-НСl (рН 7,4), 5 мМ MgCl2, 1,2 мМ CaCl2, 1 мМ ДТТ, 1 мМ ЭГТА и 100 мкг/мл ФС] инкубировали с пептидным субстратом, являющимся рецептором EGF (5 мкМ), при 30oС в течение пяти минут. Реакцию фосфорилирования инициировали добавлением 100 мкМ 32Р-АТФ (500 имп/мин/пмоль) и реакции давали протекать в течение десяти минут при 30oС. Брали аликвоты реакционной смеси и наносили в виде пятен на фильтровальную бумагу Р81 и реакцию останавливали промыванием фильтровальной бумаги избытком 75 мМ фосфорной кислоты (три промывки по три минуты каждая). После конечной промывки в этаноле фильтры Р81 сушили и радиоактивность измеряли жидкостным сцинтилляционным счетом.
Пептид, содержащий остатки 31-52, а также рекомбинантный фрагмент к первым 75 аминокислотам АКАР 79 были сильными ингибиторами активности ПКС с IC50 2 мкМ и 25 нМ соответственно. Более подробный кинетический анализ показал, что пептид 31-52 АКАР 79 проявлял смешанное ингибирование активности ПКС с К, 1,411±0,28 мкМ с использованием рецепторного пептида эпидермального фактора роста (EGF) в качестве субстрата. Кроме того, этот район напоминает также СаМ-связывающий домен, и инкубирование рекомбинантного фрагмента 1-75 или пептида 31-52 с СаМ (15 мкМ) предотвращало ингибирование ПКС в присутствии избытка Са2+. Поскольку АКАР 79 является СаМ-связывающим белком, эти открытия предполагают, что Са2+/СаМ может регулировать связывание ПКС с якорным белком.
В сочетании, эти результаты предполагают, что ПКС связывает АКАР 79 in vitro, ПКС-связывающий сайт находится в первых 75 остатках АКАР 79 и пептиды, включающие в себя остатки 31-52, ингибируют активность ПКС. Эти результаты предполагают также, что взаимодействие ПКС/АКАР 79 может регулироваться СаМ, так как инкубирование с избытком Са2+/СаМ предотвращает ингибирование ПКС пептидом 31-52 (фиг.3). Для более полного понимания характера взаимодействия АКАР 79/ПКС были спланированы эксперименты для 1) идентификации остатков, важных для связывания ПКС с АКАР 79, 2) выделения комплекса ПКС/АКАР 79 из клеток и 3) определения, регулирует ли СаМ взаимодействие ПКС/АКАР 79.
Анализ последовательности нескольких ПКС-связывающих белков показал, что высокий положительный заряд поверхности может быть необходим для связи с ПКС. В соответствии с этой гипотезой находятся предыдущие результаты, где пептидный фрагмент из 31-52 аминокислот АКАР 79, который включает в себя кластер основных и гидрофобных остатков, ингибирует активность ПКС (Ki 1,4±0,28 мкМ), а рекомбинантный фрагмент к этому району является даже еще более сильным ингибитором этой киназы (IC50=25±5 нМ). Для оценки роли основных боковых цепей, локализованных между остатками 31-52 АКАР 70, в качестве детерминант ингибирования ПКС, генерировали семейство мутантов АКАР 79 в рекомбинантном полипептиде АКАР 79, содержащем аминокислоты 1-75, и ПКС-связывающие свойства каждого мутанта анализировали в способе наложения и в отношении изменений ингибирующей силы против ПКС β1.
Конструируют пять мутантов АКАР 79, в которых кластеры основных остатков заменяют аланином. При условии высокой плотности положительного заряда, вероятным является то, что одновременная замена нескольких основных боковых цепей будет необходима перед регистрацией значимых изменений в ПКС-связывающей аффинности. Таким образом, заменяют множественные основные остатки. Точковые мутанты в последовательности АКАР 79 создают при помощи аланин-сканирующего мутагенеза с использованием способов, описанных Hausken, et al. , [J. Biol. Chem., 269:24245-24251 (1994)]. Каждый белок АКАР 79 экспрессируют в виде слитого белка с His-меткой и очищают до гомогенности аффинной хроматографией с никелем. Аланиновые мутантные пептиды показаны ниже. SEQ ID NО:37 представляет последовательность нативного АКАР 79.
АКАР 79 (37-50) FXRRKKAAKALAPK (SEQ ID NO:37)
АКАР 79 AA38, 39 FAARKKAAKALAPK (SEQ ID NO:38)
AKAP 79 AAA40-42 FKRAAAAAKALAPK (SEQ ID NO:39)
AKAP 79 4A38-42 FAAAAAAAKALAPK (SEQ ID NO:40)
AKAP 79 AA45, 50 FKRRKKAAAALAPA (SEQ ID NO:41)
AKAP 79 A37-50 FAAAAAAAAALAPA (SEQ ID NO:42)
Белок ПКС β1 экспрессируют в бакуловирусе и моноклональные антитела М4 и М7 используют для детектирования изоформ ПКС α и β нижеследующим способом.
Кроме того, каждый мутантный фрагмент АКАР 79 тестируют на его способность ингибировать ПКС описанным выше способом.
Поскольку предварительные данные предполагают, что ПКС и АКАР 79 связываются in vitro, должно быть возможным выделение комплекса АКАР 79/ПКС из клеток, если такое же или сходное связывание происходит in vivo. В попытке выделения бинарного комплекса ПКС/АКАР 79 или трехкомпонентного комплекса ПСК/АКАР 79/CaN из бычьего мозга используют два независимых подхода, которые ранее были успешными для выделения комплекса АКАР 79/CaN in vivo. Эти способы кратко описаны ниже.
Начальные исследования включают в себя иммунопреципитацию гомолога АКАР 79, АКАР 75 из бычьего мозга с использованием моноклонального антитела МС16, индуцированного против АКАР 79. Совместную очистку ПКС в этих иммунопреципитатах детектируют Вестерн-блот-анализом с кроличьими поликлональными антисыворотками, которые узнают преобладающие изоформы ПКС мозга α, βI, βIII и γ. Альтернативно, ПКС иммунопреципитируют из экстрактов бычьего мозга моноклональным антителом М7, которое узнает изоформы ПКСα и β мозга, и совместно очищающийся АКАР 79 детектируют наложением PII или Вестерн-блоттингом. Наконец, идентичные пробы, иммунопреципитированные антителами против ПКС, зондируют на CaN моноклональным антителом С24, которое узнает бычью субъединицу CaN А. Эти эксперименты могут определить, образуется ли трехкомпонентный комплекс АКАР 79/ПКС и CaN.
Альтернативно выполняют аффинную очистку для выделения трехкомпонентного комплекса PII, АКАР 79 и ПКС из бычьего мозга. Р-субъединицу ПКА очищают аффинной хроматографией на цАМФ-агарозе и элюат подвергают скринингу на присутствие ПКС и АКАР с использованием Вестерн-блотов с моноклональными антителами М7 и МС16 соответственно. Поскольку рекомбинантные АКАР 79 и ПКС не связываются с цАМФ-агарозой, детектирование любого белка в цАМФ-элюате подтверждает образование комплекса как киназ, так и якорного белка. Подтверждение трехкомпонентного комплекса достигается элюцией ПКС и АКАР 79 из цАМФ-агарозы избытком якорного ингибиторного пептида. Ранее было показано, что этот пептид вытесняет комплекс AKAP/CaN из РII, иммобилизованного на цАМФ-агарозе.
Пример 12
Предыдущая демонстрация, что АКАР 79 связывает кальцинейрин, является важной ввиду того факта, что кальцинейрин является мишенью двух сильнодействующих и клинически применимых иммуносупрессивных агентов, циклоспорина и FK506, оба из которых ингибируют активность кальцинейрина. Как описано ниже, как циклоспорин, так и FK506 применимы в лечении различных заболеваний, но имеют значительные ограничивающие побочные эффекты. Предположительно, факторы, которые модулируют связывание якорный белок/кальцинейрин, могут в конечном счете модулировать активность кальцинейрина аналогичным активностям циклоспорина или FK506 образом. Идентификация такого модулятора, в частности, с меньшими побочными действиями, чем побочные действия, наблюдаемые с другими иммуносупрессорами, имела бы, возможно, широко распространенное терапевтическое применение в лечении многочисленных заболеваний, которые в настоящее время лечат циклоспорином или FK506.
Сообщались многочисленные клинические показания циклоспорина и FK506. Например, циклоспорин определил стандарт для иммуносупрессии после трансплантации, делая возможными трансплантации печени, легкого, кишечника и поджелудочной железы, даже хотя обычно считается, что FK506 является более сильным иммуносупрессором. Пациенты с пересаженными органами, которые не переносят циклоспорин или FK506 или не обнаруживают успеха в случае их применения, иногда с успехом переводятся на другое лекарственное средство.
В качестве другого примера, воспалительное заболевание пищеварительного тракта (IBD) является общим названием для двух заболеваний, имеющих различные клинические проявления, болезни Крона и язвенного колита (UC). Циклоспорин успешно использовали для лечения болезни Крона, причем статистически значимые результаты лечения были продемонстрированы в по меньшей мере одном показателе активности заболевания [Brynskov, Dan. Med. Bull. 41:332-344 (1994)] . Однако другие показатели, которые лучше всего коррелируют со снятием сильных обострений, показали незначимые тенденции в направлении улучшения. Циклоспорин обнаружил также активность в тяжелом остром стероидорезистентном язвенном колите (эти результаты не являются значимыми, так как испытание было приостановлено по этическим причинам). Другое испытание на пациентах со склерозирующим холангитом и UC продемонстрировало действие на грани значимости в направлении более мягкого протекания UC. Рецидив был обычным после отмены лечения и лечение ограничивалось опасением в отношении токсичности [Choi and Targan, Dig. Dis. And Sci. 39:1885-1892 (1994)]. Кроме того, другие иммуносупрессоры использовали успешно в IBD, такие как метотрексат, азатиоприн и 6-МР.
В качестве другого примера, было показано, что циклоспорин был эффективным в лечении ревматоидного артрита в нескольких испытаниях при применении в качестве терапии второй или третьей очереди, т.е. в случае пациентов, которые не имели улучшения с другими установленными способами терапии и имеют тяжелое заболевание. В этих испытаниях было обнаружено, что циклоспорин является обычно таким же эффективным и токсичным, как и другие агенты второй очереди, такие как золото, противомалярийные агенты, азатиоприн, D-пеницилламин и метотрексат [Wells and Tugwell, Br. J. Rheum., 32 (suppl 1):51-56 (1993); Forre et al., Arth. Rheum., 30:88-92 (1987)]. Эти испытания сообщают только о лечении "очень тяжелого резистентного (трудноизлечимого) ревматоидного артрита (RA)" вследствие "потенциально необратимой токсичности" циклоспорина [Dougados and Torley, Br. J. Rheum., 32 (suppl 1): 57-59 (1993)]. Считается, что почечная токсичность первично опосредуется почечной вазоконстрикцией (сужением кровеносных сосудов), которая обостряет нефротоксичность NSAID и болезнь почек, свойственную пациентам с ревматоидным артритом [Leaker and Cairns, Br. J. Hosp. Med., 52:520-534 (1994); Sturrock et al., Nephrol. Dial. Transplant, 9:1149-1156 (1996); Ludwin and Alexopolulou, Br. J. Rheum. , 32 (suppl 1):60-64 (1993)]. Около 10% почечных биопсий из пациентов с RA, получавших циклоспорин, обнаружили морфологические признаки токсичности циклоспорина [International Kidney Biopsy Registry of Cyclosporin in Autoimmune Diseases, Br. J. Rheum., 32(suppl 1): 65-71 (1993)].
В качестве еще одного примера, сообщалось, что циклоспорин является эффективным для лечения стероидзависимой астмы. В одном испытании небольшое количество пациентов случайно выбирали для введения циклоспорина или плацебо, и группа циклоспорина обнаружила увеличенные потоки (вдыхаемого и выдыхаемого) воздуха и форсированную жизненную емкость легких (ФЖЕЛ), а также меньше курсов спасения с использованием преднизолона.
В другом примере было показано, что циклоспорин является эффективным в лечении стероидзависимого нефротического синдрома с заболеванием с минимальными изменениями. Было показано, что пациенты в этом испытании имели меньшую потребность в стероидах при низкой дозе циклоспорина, но все давали рецидивы при прекращении введения циклоспорина. Стероидорезистентные формы нефротического синдрома имеют лишь 20-30% уровни ответа на циклоспорин [Meyrier, Nephrol. Dial. Transplant, 9:596-598 (1994); Hulon et al., Pediatr. Nephrol., 8:401-403(1994)].
Что касается лечения системной красной волчанки (SLE), одно исследование сообщало о значимом снижении показателей активности SLE в перспективном, нерандомизированном, не имеющем достаточных контролей исследовании [Tokuda et al. , Arthr. Rheumat., 37:551-558 (1994)]. Однако другие исследования не показали эффективности в случае SLE.
В качестве другого примера, было показано, что циклоспорин индуцирует ремиссию в инсулинзависимом сахарном диабете при ранней госпитализации после начального проявления. Ремиссии длились в среднем около одного года, хотя некоторые ремиссии пролонгировались до 850 дней [Jenner et al., Diabetologia, 35:884-888 (1992); Bougneres et al., Diabetes, 39:1264-1272 (1990)]. He отмечалось долгосрочного действия циклоспорина в последующем длительном наблюдении после одного исследования [Martin et al., Diabetologia, 34:429-434 (1991)]. Однако в другом исследовании функция почек ухудшалась во время лечения в течение 12-18 месяцев и она не возвращалась полностью к уровню плацебо, что указывает на то, что могло произойти некоторое хроническое повреждение почек [Feldt-Rasmussen et al., Diabetes Medicine, 7:429-433 (1990)]. Более раннее вмешательство требовалось бы для усиления действия иммуносупрессивной терапии на протекание инсулинзависимого сахарного диабета. Некоторые исследователи подвергают скринингу родственников первого порядка и успешно лечат тех из них, которые имеют диабетические маркеры [Elliot and Chase, Diabetologia, 34:362-365 (1991)].
Еще в одном примере псориаз эффективно лечили циклоспорином [Cuellar et al. , Balliere's Clin. Rheum. , 8:483-498 (1994); Ellis et al., JAMA 256: 3110-3116 (1986)] . Терапия высокими дозами была эффективной для лечения псориатического артрита, особенно тяжелой формы деформирующего артрита, и прекращение терапии обычно сопровождалось обострением поражения кожи и суставов. Ввиду потенциальных побочных действий и необходимости непрерывного долгосрочного лечения циклоспорин показан только для резистентного (трудноизлечимого) псориатического артрита, который не лечится адекватно другими средствами.
Кроме того, было показано, что циклоспорин является эффективным для лечения тяжелого атопического дерматита, в двоякослепых исследованиях с плацебо в качестве контроля [Van Joost et al., Br. J. Derm., 130:634-640 (1994); Cooper, J. Invest. Derm., 102:128-137 (1994)]. Пациенты предпочитали не получающей лечения болезни побочные эффекты от этого лекарственного средства в виде тошноты, желудочно-кишечного дискомфорта, парестезии, холестаза и почечной недостаточности. Другое рандомизированное двоякослепое, контролируемое плацебо исследование показало, что лечение циклоспорином значимо повышало качество жизни пациентов с тяжелым атопическим дерматитом [Saiek et al. , Br. J. Derm., 129:422-430 (1993)]. Кожные поражения быстро возобновлялись после прекращения лечения циклоспорином, но качество жизни оставалось улучшенным.
Еще в одном примере, циклоспорин использовали в лечении хронического дерматита рук, заболевания с сообщенной распространенностью 4-22% и обычно подвергаемого лечению наносимыми местно стероидами, на которые, однако, многие пациенты не дают положительной реакции. Было показано, что циклоспорин в низких дозах эффективно излечивал 6 из 7 пациентов в открытом исследовании [Reitamo and Granlund, Br. J. Derm., 130:75-78 (1994)]. Приблизительно половина пациентов имела рецидивы после прекращения лечения циклоспорином.
Еще в одном примере, циклоспорин использовали в лечении крапивницы и гигантской крапивницы (отека Квинке), идиопатических кожных заболеваний, которые присутствуют в виде сыпей (крапивницы) и подкожных опуханий. Патология связана с тучными клетками (мастоцитами) и лечение часто является неэффективным. В одном испытании трех пациентов с резистентными крапивницей и отеком Квинке лечили циклоспорином, и все симптомы устранялись в пределах одной недели [Fradin et al., J. Am. Acad. Derm., 25:1065-1067 (1991)]. Все пациенты должны были приостановить терапию из-за побочных действий, и симптомы вновь появились после прекращения терапии.
Что касается других ревматологических заболеваний, исследования сообщают эффективное лечение циклоспорином других, менее обычных аутоиммунных заболеваний, в том числе болезни Бехчета [Pacor et al., Clin. Rheum. 13:224-227 (1994)] , некротического гранулематоза Вегенера [Allen et al., Cyclosporin A Therapy for Wegener's Granulomatosis in ANCA-Associated Vasculitidis: Immunological and Clinical Aspects, Gross ed. Plenum Press (1993)] и иммуноопосредованной тромбоцитопении [Schultz et al., Blood 85:1406-1408 (1995)].
Во многих вышеописанных испытаниях применение циклоспорина или FK506 было связано с многочисленными нежелательными побочными действиями. Обычно увеличенный риск инфекции и злокачественности связан с общей иммуносупрессией, и маловероятно, что связанные с якорным белком иммуносупрессивные агенты не будут иметь подобного риска. Однако других побочных эффектов можно избежать или они могут быть уменьшены тканеспецифичностью якорных белков. Наиболее обычным серьезным побочным эффектом как циклоспорина, так и FK506 является нефротоксичность, которая по меньшей мере в некоторой степени связана с дозой и встречается в большинстве пациентов, обычно в форме уменьшения скорости гломерулярной фильтрации во время лечения. Однако этот побочный эффект является по меньшей мере частично обратимым при прекращении введения лекарственного средства [Leaker and Cairns, supra]. Обычно прогрессирующая почечная недостаточность не развивается, хотя необходима еще проверка для окончательной оценки. Хроническое повреждение наблюдали также у пациентов, получающих циклоспорин в низких дозах (3-4 мг/кг/сутки), приблизительно 40% биопсий этих пациентов показали изменения интерстициального фиброза, атрофию канальцев и артериолопатию [Svarstad et al., Nephrol. Dial. Transplant, 9: 1462-1467 (1994); Young et al., Kidney International, 46:1216-1222 (1994)]. Изменения в эндотелиальных клетках были также видны в гистологических срезах [Kahan, N. Engl. J. Med. 321:1725-1748 (1989)]. Был сделан вывод, что нефротоксичность первично обусловлена вазоконстрикцией артериол и хронической ишемией низкого уровня [Leaker and Cairns, supra], хотя было показано, что эти лекарственные средства являются непосредственно токсичными для клеток почечных канальцев и сосудистых интерстициальных клеток [Platz et al., Transplantation, 58: 170-178 (1994)]. Некоторые сообщения указывают на то, что встречаемость и тяжесть нефротоксичности может быть слегка более высокой с FK506 [Platz et al., supra].
Другой сообщенной значимой токсичностью как циклоспорина, так и FK506 была нефротоксичность с клиническими проявлениями, в том числе припадками, спутанностью сознания, слепотой, комой, головной болью, атаксией, синдромом Паркинсона, парестезиями, психозом, фокальными дефицитами, акинетическим мутизмом, тремором, невропатией и нарушениями сна [Shimizu et al., Pediatr. Nephrol. , 8: 483-485 (1994); Wilson et al., Muscle and Nerve, 17:528-532 (1994); Reece et al., Bone Marrow Transpl., 8:393-401 (1991); Eidelman et al., Transpl. Proc., 23:3175-3178 (1991); de Groen et al., N. Engl. J. Med., 317: 861-566 (1987)] . Было показано, что после трансплантации печени умеренная-тяжелая нейротоксичность имела место в 10-20% пациентов, получающих FK506, и 3-12% пациентов, получающих циклоспорин. Нейротоксичность была также связана с отклонениями от нормы липидов и дисфукцией печени.
Другие побочные эффекты циклоспорина и/или FK506 включают в себя гепатотоксичность, непереносимость глюкозы, гирсутизм, желудочно-кишечные симптомы, тромбоз вен, панкреатит и гипертрофический гингивит [Morris, J. Heart Lung Transplant, 12: S275-S286 (1993); Fung et al., Transpl. Proc., 23: 3105-3108 (1991); Mason, Pharmacol. Rev., 42:423-434 (1989); Kahan, N. Engl. J. Med. , 321: 1725-1738 (1989); Thomason et al., Renal Failure, 16:731-745 (1994)] . Таким образом, ввиду широко распространенного использования циклоспорина и FK506 и свойственных им побочных эффектов их использования исключительно полезным могло бы быть развитие альтернативных иммуносупрессоров.
Например, возможно, что делокализация кальцинейрина из предполагаемого Т-клеточного якорного белка могла бы ингибировать активность кальцинейрина в активации Т-клеток, обеспечивая тем самым Т-клетко-специфический иммуносупрессор, имеющий полезность циклоспорина или FK506, но с меньшими побочными эффектами. Предыдущее наблюдение, что делокализация ПКА из Т-клеточного якорного белка усиливала экспрессию IL-2 в стимулированных клетках, показало, что локализованная ядерным белком ПКА каким-то образом способствует регуляторной роли в экспрессии IL-2 во время активации Т-клеток. Таким образом, Т-клетко-специфическая делокализация ПКА может обеспечивать средство для усиления секреции IL-2 in vivo, имитируя тем самым введение рекомбинантного IL-2, и, возможно, уменьшения ранее сообщенной токсичности лечения с использованием IL-2, как описано ниже.
IL-2 был одобрен для лечения метастатического рака почек, и приблизительно 15-20% пациентов с метастатическим раком почечных клеток или злокачественной меланомой реагируют на IL-2-терапию. Некоторые из этих ответных реакций являются продолжительными, длящимися более 66 месяцев [Dillman, Cancer Biotherapy, 9:183-209 (1994); Whittington and Faulds, Drugs 46:446-514 (1993)] . Хотя болюсная терапия с высокими дозами была связана с несколькими тяжелыми побочными эффектами (как описано ниже), подкожная или непрерывная инфузионная терапии низкими дозами давала умеренный коэффициент ответа (12%) с уменьшением токсичности [Vogelzang et al., J. Clin, Oncol., 11:1809-1816 (1993)].
Терапия с использованием IL-2 (с интерфероном-α и другими агентами или без интерферона-α и других агентов) исследовалась в лечении других злокачественных заболеваний. Например, стойкие клинические ответные реакции, но без излечиваний, были получены при прямом нанесении IL-2 на ложе опухоли после резекции глиомы [Merchant et al., J. Neuro., 8:173-188 (1990)]. В других испытаниях, ограниченная эффективность была сообщена в лимфоме [Dillman, supra] , колоректальном раке [Whittington and Faulds, supra], ограниченном AML [Bruton and Koeller, Pharmacotherapy, 14:635-656 (1994)], раке яичника и раннем раке мочевого пузыря [Whittington and Faulds, supra]. Однако число участников в каждом из этих исследований было слишком малым, чтобы позволить значимые выводы в отношении эффективности.
IL-2 использовали также в комбинации с адоптивной иммунотерапией, и было показано, что он является эффективным для лечения рака почки [Pierce et al., Sem. Oncol. , 22: 74-80 (1995); Belldegrun et al., J. Urol., 150:1384-1390 (1993)] . Кроме того, IL-2 может быть эффективным для лечения некоторых инфекционных заболеваний путем уменьшения бактериальной нагрузки на кожу и уровней антигена в пациентах с проказой после интрадермальной инъекции [Kaplan, J. Infect. Dis., 167 (suppl 1): S18-22 (1993)]. Наблюдали также, что, в сравнении с ТБ-тест-положительными здоровыми контролями, лимфоциты из пациентов с туберкулезом продуцируют более низкие уровни IL-2 [Sanchez et al. , Int. Immun., 62:5673-5678 (1994)], что свидетельствует о том, что IL-2-терапия может быть ценной в лечении микобактериальных инфекций.
Несмотря на потенциальную терапевтическую ценность IL-2, этот цитокин также связан со значимой токсичностью [если нет других указаний, источниками являются Whittington and Faulds, Dillman and Bruton и Koeller, supra]. Главным лимитирующим лечение побочным эффектом является синдром утечки капилляров. Введение IL-2 увеличивает проницаемость сосудов, вызывая интерстициальный и легочный отек, причем пациенты развивают гипотензию и значительное количество пациентов требует введения вазопрессорных (повышающих кровяное давление) средств. Интенсивная газовая реанимация может вызывать опасный для жизни отек легких. До 20% пациентов могут требовать интубации и механической вентиляции. Введение болюсов с высокими дозами вызывает более тяжелую утечку, чем низкие дозы или медленные непрерывные инфузии, и в некоторых схемах лечения 100% пациентов требуют помещения в отделение интенсивной терапии во время лечения IL-2. Наблюдали также миокардит, кардиомипатии и сердечные аритмии. Острая почечная недостаточность может встречаться как результат индуцированной синдромом утечки капилляров гипотензии.
IL-2 может также вызывать тяжелую диарею с нарушением равновесия электролитов, холестаз, отклонения от нормы щитовидной железы и острый панкреатит. Анемия, требующая трансфузий, имеет место в 15-20% проходящих лечение пациентов [MacFarlane et al., Cancer 75:1030-1037 (1995)]. Может встречаться тромбоцитопения с кровотечением и обычными являются дефекты пути свертывания. Более 70% пациентов испытывают изменения в психическом состоянии, в том числе параноидальный бред, галлюцинации, потерю интересов, нарушение сна и гиперсомнию. Сообщались также кома, дефекты зрения, преходящие ишемические приступы и парестезии. Эти недостатки, связанные с экзогенным IL-2, предполагают, что в качестве потенциальных терапевтических веществ должны быть исследованы альтернативы, в которых, например, может модулироваться продуцирование эндогенного IL-2 и, следовательно, устраняется потребность в лечении экзогенным IL-2.
Кроме обеспечения возможных средств для идентификации иммуносупрессивных лекарственных средств и модуляторов продуцирования IL-2, идентификация якорных белков делает возможной регуляцию другой клеточной активности ввиду разнообразных метаболических путей, в которых, как было показано, участвуют якорные белки. Например, АКАР 79 является важным в регуляции регулируемых рецептором глутамата ионных каналов в постсинаптической плотности нейронов, предположительно через связывание ПКА, ПКС и кальцинейрина. ПКА регулирует активность регулируемых рецептором АМРА каналов, а делокализация или ингибирование ПКА аттенуирует активность ионных каналов АМРА. ПКС регулирует активность регулируемых NMDA-рецептором каналов, а кальцинейрин, как было показано, десенсибилизирует NMDA-рецептор в отношении стимулов. Эти наблюдения показывают, что локализованные киназы (ПКА и ПКС) могут регулировать активность рецепторов глутамата в нейронах. Дефосфорилирование кальцинейрина является противорегуляторным механизмом NMDA-рецепторов. Эта модель согласуется физиологически со свидетельствами припадков, индуцируемых циклоспорином или FK506.
Кроме того, предполагалось, что рецепторы глутамата участвуют во многих неврологических заболеваниях. Глутамат и другие и участвующие в передаче возбуждения аминокислоты могут создавать экситотоксичность в нейронах, и было показано, что избыточная стимуляция постсинаптических рецепторов глутамата является токсичной для нейронов, вызывая острую дегенерацию нейронов. Было показано, что гипоксия (например, после удара или остановки сердца) и травма ЦНС вызывают заметный выход глутамата во внеклеточное пространство, где глутамат взаимодействует с рецепторами глутамата и запускает экситотоксический каскад. Было показано, что противовозбуждающие агенты защищают от мозгового повреждения в моделях животных [Olney, Neurobiology of Aging, 15: 259-260 (1994)] . Интересно, что антагонисты NMDA являются токсичными в отношении некоторых типов нейронов, что свидетельствует о том, что глутамат может ингибировать другие пути передачи возбуждения в этих клетках. Было также показано, что макролидные антитела, такие как FK506, защищают против экситотоксичности NMDA, но не каината, в культивируемых нейронах [Manev, et al., Brain Res., 624:331-335 (1993)].
Предполагалось также участие глутамата в болезни Паркинсона. Антагонисты NMDA защищают допаминергические нейроны в черном веществе обезьян, подвергнутых действию МРТР, химического агента, который индуцирует синдром Паркинсона у человека и других приматов. Амантидин и мемантин являются антагонистами NMDA и использовались в Европе для лечения болезни Паркинсона, однако было показано, что они оба вызывают психоз у некоторых пациентов. Имеется некоторое доказательство того, что глутаматергические нейроны могут быть гиперактивными в болезни Паркинсона и ингибирование могло бы уменьшать моторные симптомы этой болезни [Lange and Riederer, Life Sciences, 55:2067-2075 (1994)].
Глутамат играет также роль в нарушениях, сопровождающихся припадками, участвуя в инициировании, распространении и поддержании активности припадков. NMDA- и нe-NMDA-антагонисты являются сильными противосудорожными средствами [Meldrum, Neurology, 44 (suppl 8):S14-S23 (1994)]. Рецепторы АМРА также участвуют в ALS и в настоящее время проводится испытание антагониста рецептора.49
Ввиду всех этих наблюдений неудивительно, что многочисленные другие иммунодепрессанты находятся в клинических испытаниях. Следующая информация относительно таких испытаний была получена из Haydon and Haynes, Balliere's Clin. Gastroentero, 8: 455-464 (1994); Thomason and Starzi, Immunol. Rev. 1993, 71-98 (1993) и Morris J. Heart Lung Transplant., 12:S275-S286 (1993). Например, азаспиран является соединением SKB, которое ингибирует клеточные инфильтраты трансплантатов и индукцию IL-2R и также прекращает продуцирование IL-2 и IFN-γ. По-видимому, азаспиран индуцирует некий тип супрессорной клетки и имеются некоторые доказательства синергических эффектов с циклоспорином.
В качестве другого примера, микофенолят мофетиал является соединением Syntex, которое ингибирует синтез пуринов и обладает селективным в отношении Т- и В-клеток антипролиферативным действием. Он снижает содержание антител. Микофенолят мофетиал может также удалять молекулы адгезии из клеточных поверхностей. Хотя это лекарственное средство имеет низкую токсичность, оно может вызывать лейкопению и использовалось для лечения псориаза в течение 20 лет.
В качестве другого примера, мизорибин является соединением Sumitomo, которое ингибирует синтез ДНК. Механизм действия идентичен механизму действия микофенолята.
В качестве другого примера, бреквинар является соединением DuPont-Merck, которое ингибирует синтез пиримидинов блокированием дигидрооратдегидрогеназы. Ожидаются полные сообщения по клиническим испытаниям. Сообщалось, что данное лекарственное средство действует синергически с циклоспорином, но может вызывать тромбоцитопению, дерматит и воспаление слизистой оболочки.
В качестве еще одного примера, 15-дезоксиспергуалин является соединением Nippon-Kayaku, которое благоприятно влияет на функцию моноцитов/макрофагов, в том числе ингибирование окислительного метаболизма, синтеза ферментов лизосом, продуцирования IL-1 и экспрессии на поверхности клеток антигенов МНС класса II. Он имеет 70-90%-ную эффективность в стойком отторжении почек, но при более высоких дозах может иметь место токсичность в отношении костного мозга.
В качестве другого примера, лефлуномид является соединением Hoechst, которое ингибирует синтез цитокинов, блокирует активацию Т-клеток и синтез антител. Это соединение не является токсичным в отношении почек или костного мозга.
В качестве другого примера, рапамицин является соединением Wyeth-Ayerst, которое родственно FK506. Рапамицин является пролекарством, которое должно связать иммунофилин, чтобы быть активным, и не ингибирует кальцинейрин или не блокирует продуцирование цитокинов Т-клетками. По неизвестному механизму рапамицин блокирует переход G1 в S.
Ожидается, что многочисленные модификации и вариации в данном изобретении, изложенном в приведенных выше иллюстративных примерах, могут придти в голову специалистам в данной области. Поэтому данное изобретение ограничивается только прилагаемой формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГИБРИДОМА, ОБОЗНАЧЕННАЯ 199М, И МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СЕКРЕТИРОВАННОЕ ЭТОЙ ГИБРИДОМОЙ | 1997 |
|
RU2183671C2 |
| ГОМОЛОГИ (ATR) ПОЛИПЕПТИДА rad3, ПОЛИНУКЛЕОТИДЫ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ И МАТЕРИАЛЫ | 1996 |
|
RU2252256C2 |
| СПОСОБ СКРИНИНГА НЕВРОПАТОЛОГИИ | 1996 |
|
RU2155345C2 |
| ПОЛИПЕПТИД, ИМЕЮЩИЙ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ ФОСФОДИЭСТЕРАЗЫ | 1998 |
|
RU2272841C2 |
| ICAM-4 И ЕГО ДИАГНОСТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ | 1998 |
|
RU2208230C2 |
| МОДИФИЦИРОВАННЫЙ БИОТИН-СВЯЗЫВАЮЩИЙ БЕЛОК, СЛИТЫЕ БЕЛКИ НА ЕГО ОСНОВЕ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2632651C2 |
| СПОСОБ ИДЕНТИФИКАЦИИ МОДУЛЯТОРА СВЯЗЫВАНИЯ МЕЖДУ ПОЛИПЕПТИДОМ α И VCAM-1 (ВАРИАНТЫ) | 1998 |
|
RU2215006C2 |
| ВНУТРИКЛЕТОЧНЫЙ ДОМЕН БЕЛКА HER-2/NEU ДЛЯ ПРЕДУПРЕЖДЕНИЯ ИЛИ ЛЕЧЕНИЯ МАЛИГНИЗАЦИЙ | 1996 |
|
RU2236461C2 |
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2012 |
|
RU2605327C2 |
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2016 |
|
RU2650767C2 |



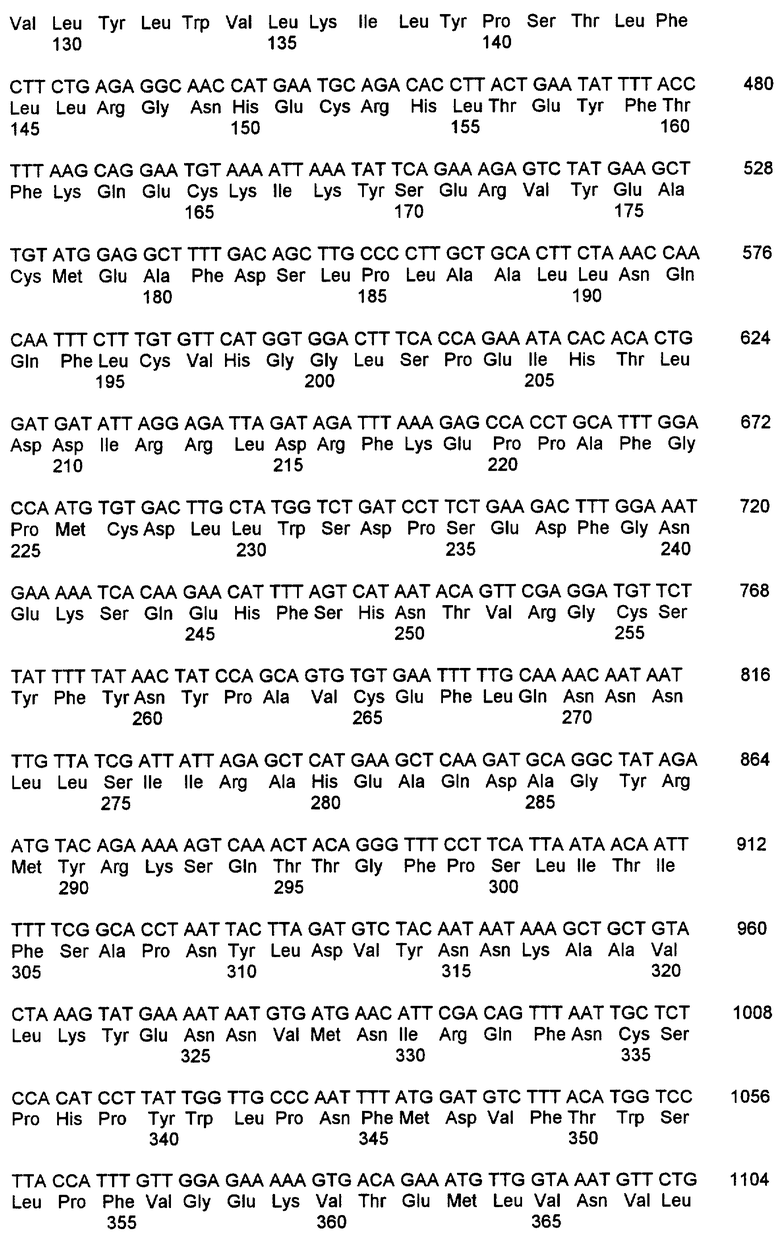












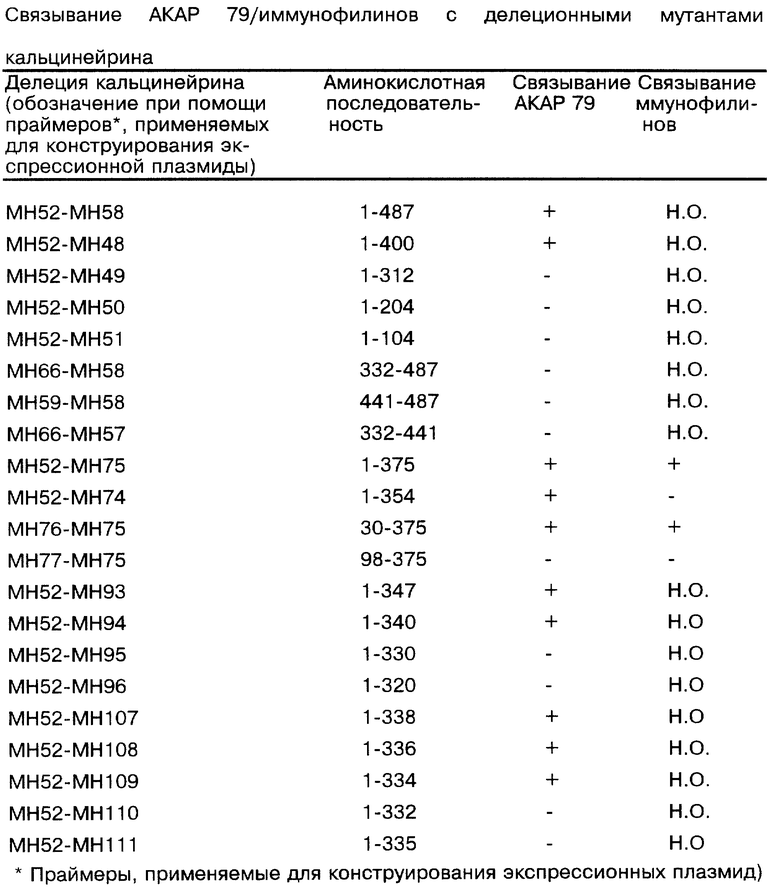

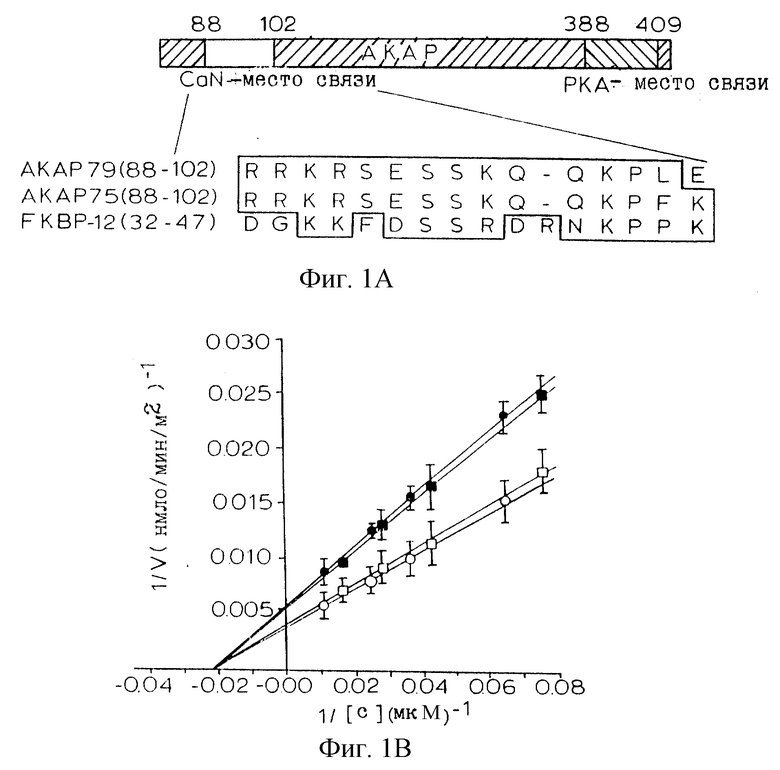
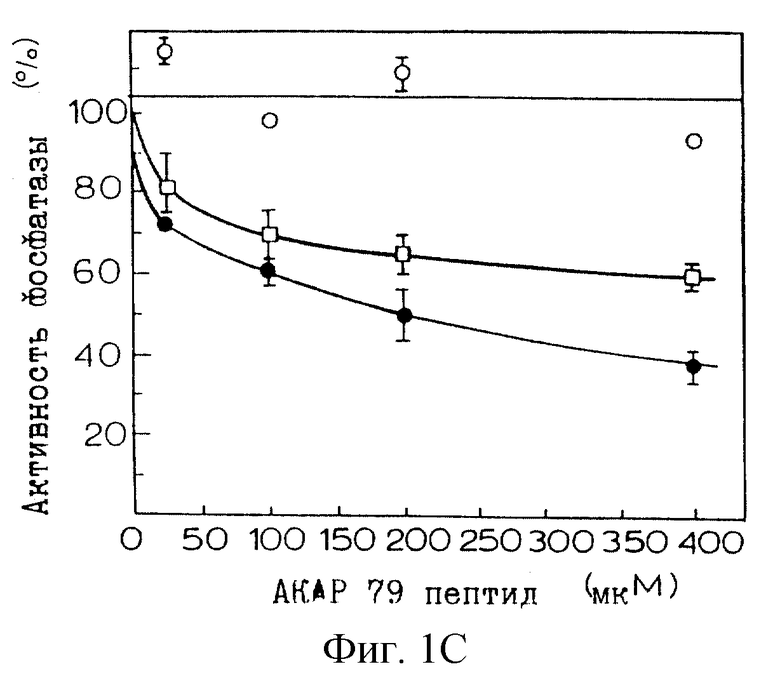


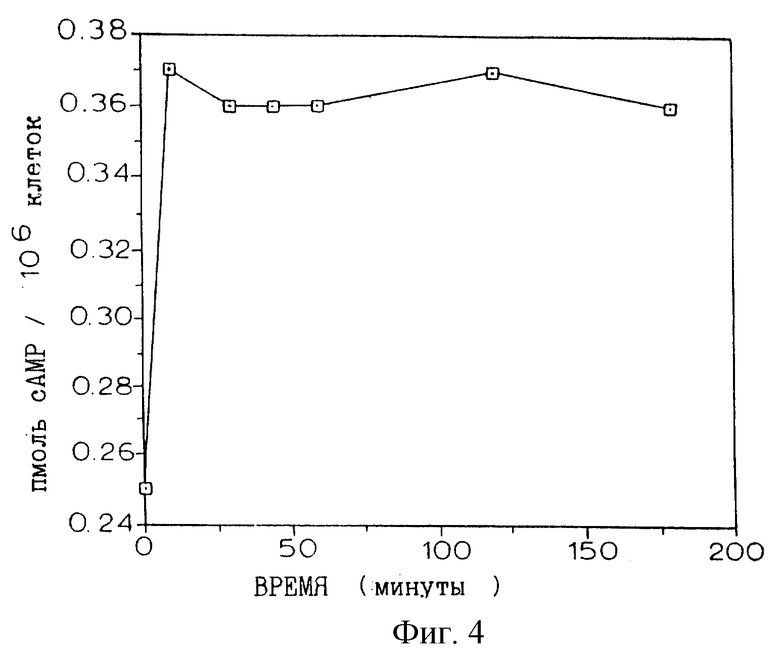
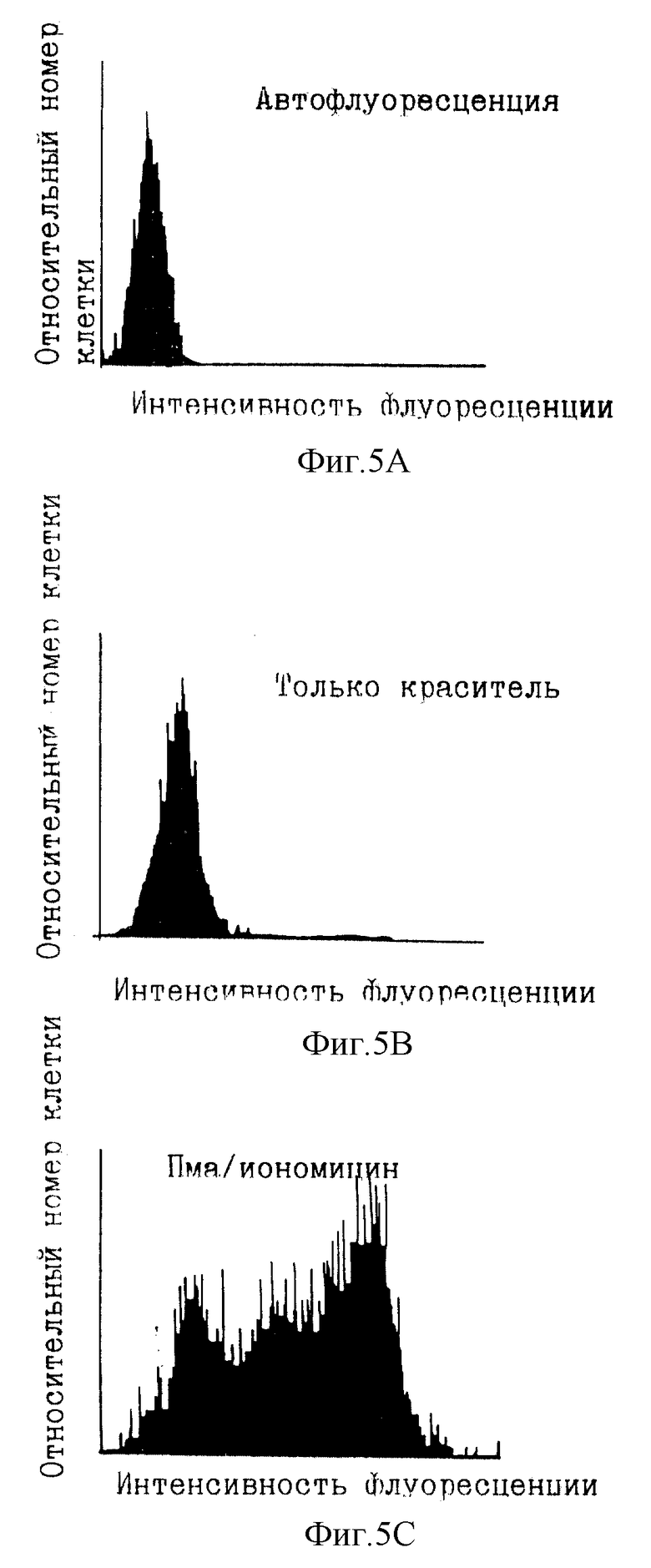
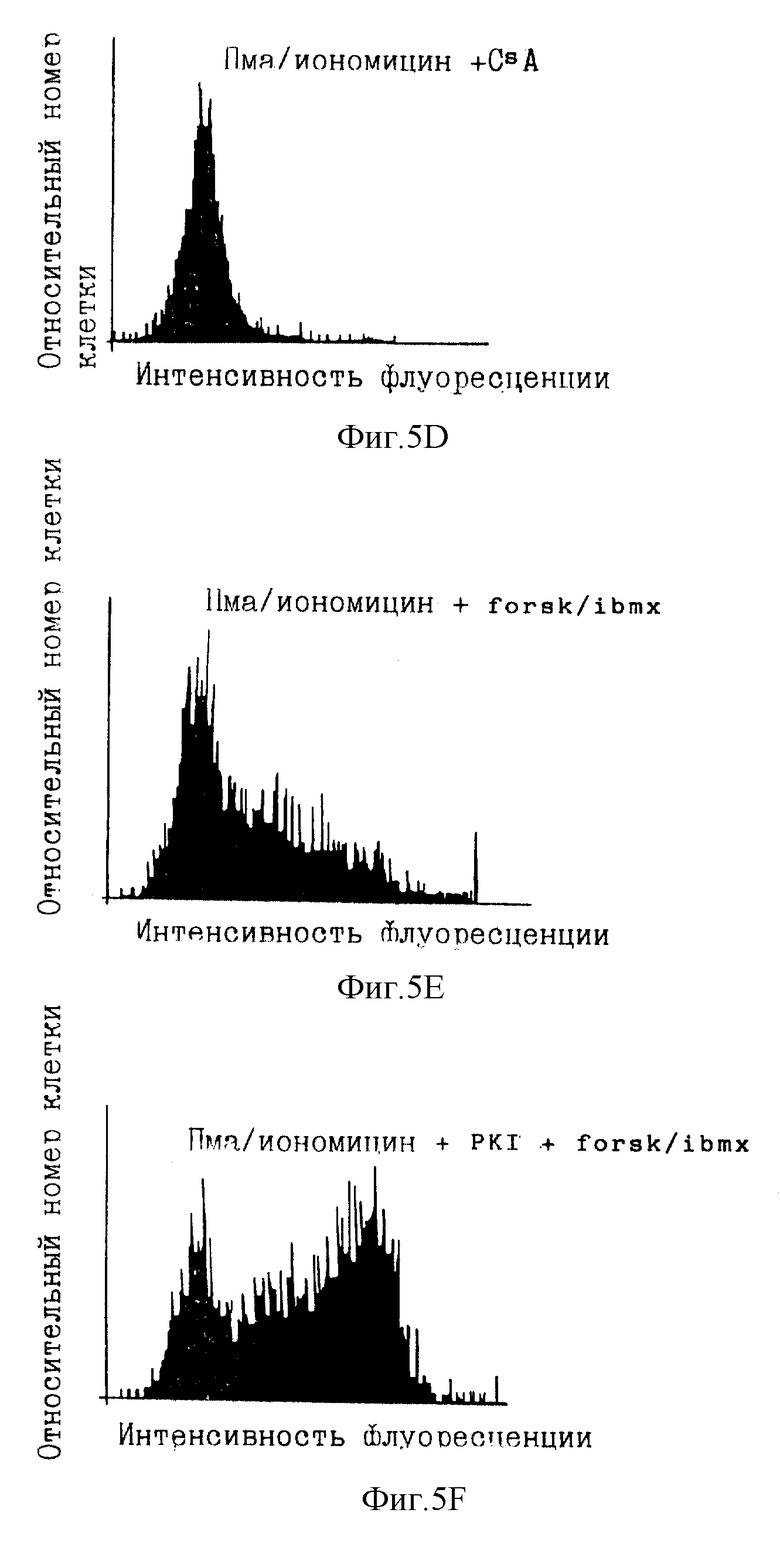
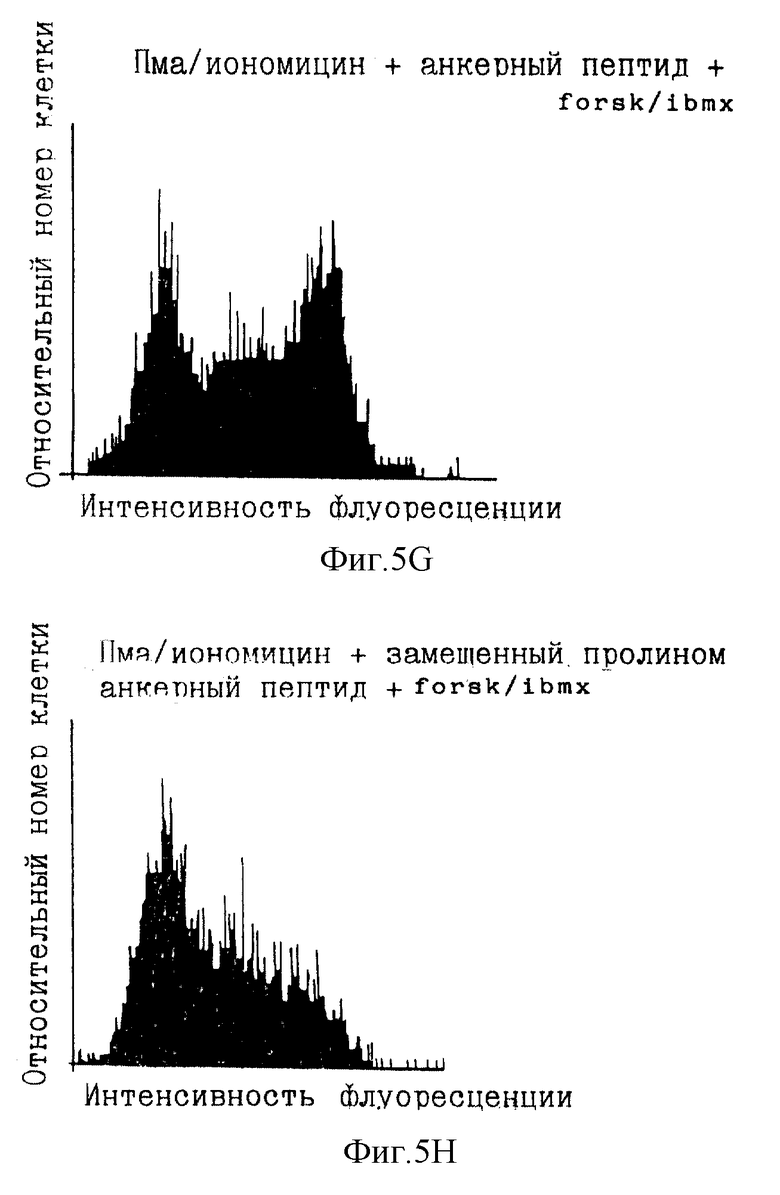
Изобретение относится к области биохимии и биотехнологии и может быть использовано для модуляции якорной функции белка. Ингибитор связывания между якорным белком и регуляторной субъединицей цАМФ-зависимой протеинкиназы (ПКА) типа I или кальцинейрином идентифицируют путем инкубации якорного белка, иммобилизованного на твердом носителе, с меченым партнером связывания в присутствии и в отсутствие предлагаемого ингибитора. Присутствие в клетке кальцинейринсвязывающего и ПКА-связывающего якорного белка проводят инкубированием лизата клеток с иммобилизованным на твердом носителе кальцинейрином. После промывания твердый носитель контактируют с меченой регуляторной субъединицей ПКА. Связавшуюся меченую субъединицу детектируют. Изобретение позволяет определять белки, связывающие и ПКА, и кальцинейрин, а также идентифицировать ингибиторы связывания. 2 с. и 4 з.п.ф-лы, 5 ил., 1 табл.
| CARR, D | |||
| et al., Journal of Biological Chemistry, 1992, v | |||
| Тепловой измеритель силы тока | 1921 |
|
SU267A1 |
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| УСТРОЙСТВО ДЛЯ ПОЛУЧЕНИЯ СУЧЕННЫХ НИТЕЙ ИСКУССТВЕННОГО ШЕЛКА С ПОМОЩЬЮ ВРАЩАЮЩЕЙСЯ НАСАДКИ | 1928 |
|
SU16816A1 |
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |

