Эта заявка частично продолжает патентную заявку США N 08/481130, поданную 7 июня 1995 г., которая частично продолжает заявку США N 08/245295, поданную 18 мая 1994 г., которая, в свою очередь, является частичным продолжением заявки США N 08/102852, поданной 5 августа 1993 г. как частичное продолжение заявки США N 08/009266, поданной 22 января 1993 г. как частичное продолжение заявки США N 07/894061, поданной 5 июня 1992 г. как частичное продолжение заявки США N 07/889724, поданной 26 мая 1992 г. как частичное продолжение одновременно рассматриваемой заявки США N 07/827689, поданной 27 января 1992 г.
Настоящее изобретение относится, в основном, к факторам клеточной адгезии, а более конкретно к клонированию и экспрессии ДНК, кодирующей ранее неизвестный полипептид, обозначенный как "ICAM-4", который обладает структурным родством с факторами межклеточной адгезии ICAM-1, ICAM-2 и ICAM-R.
Исследования последнего десятилетия в значительной степени пролили свет на молекулярные явления, сопутствующие межклеточным взаимодействиям в теле, в особенности на явления, имеющие место при перемещении и активации клеток в иммунной системе, и на явления, сопутствующие развитию и нормальной физиологической функции клеток в нервной системе. См., в основном, Springer, Nature, 346: 425-434(1990) - относительно клеток иммунной системы; Yochihara et al., Neurosci. Res., 10: 83-105 (1991), а также Sonderegger and Rathjen, J. Cell Biol., 119: 1387-1394 (1992) - относительно клеток нервной системы. Белки поверхности клеток и, особенно, так называемые факторы клеточной адгезии (CAM) соответственно были предметом фармацевтических исследований и разработок, имеющих своей целью вмешательство в процессы проникновения лейкоцитов в области воспаления и перемещения лейкоцитов в точно определенные ткани, а также дифференциация и образование сложных нейронных цепей. Выделение и исследование факторов клеточной адгезии, клонирование и экспрессия последовательностей ДНК, кодирующих такие факторы, и получение терапевтических и диагностических средств для воспалительных процессов, а также развития и деятельности нервной системы также было предметом изобретения многочисленных американских и зарубежных патентных заявок. См. Edwards, Current, Opinion in Therapeutic Patents, 1(11): 1617-1630 (1991) и особенно приведенные здесь "ссылки на патентную литературу".
Основной интерес с точки зрения предпосылок настоящего изобретения представляет известная идентификация и исследование некоторых медиаторов явлений клеточной адгезии, "лейкоинтегринов", LFA-1, MAC-1 и gp 150.95 (известных в номенклатуре ВОЗ как CD18/CD11a, CD18/CD11b и CD18/CD11c соответственно), которые образуют подсемейство белков поверхности клеток гетеродимерных "интегринов", присутствующих на B-лимфоцитах, T-лимфоцитах, моноцитах и гранулоцитах. См., например, Springer, табл. 1 на стр. 429. Также представляют интерес другие адгезивные молекулы простой цепи (CAM), которые участвовали в активации лейкоцитов, адгезии, подвижности и т.п., то-есть явлениях, присущих воспалительному процессу. Например, в настоящее время считается, что перед выделением лейкоцитов, что характерно для воспалительных процессов, происходит активация интегринов, существенно выраженных на лейкоцитах, за которой следует тесное лиганд/рецепторное взаимодействие между интегринами (например, LFA-1) и одним из двух или обоими отдельными факторами межклеточной адгезии (ICAM), обозначенными ICAM-1 и ICAM-2, которые выражены на поверхностях эндотелиальных клеток кровеносных сосудов и на других лейкоцитах.
Как другие CAM, исследованные в настоящее время [например, васкулярная адгезивная молекула (VCAM-1), описанная в заявке PCT WO 90/13300, опубликованной 15 ноября 1990 г., и адгезивная молекула эндотелиальных клеток тромбоцита (PECAM-1), как описано в работе Newman et al., Science, 247: 1219-1222 (1990) и заявке PCT WO 91/10683, опубликованной 25 июля 1991 г.], ICAM-1 и ICAM-2 являются структурно гомологическими по отношению к другим элементам суперсемейства генов иммуноглобулина в том, что внеклеточная часть каждой из них содержит ряд доменов, разделяющих подобный карбоксиконцевой рисунок. "Типичный" иммуноглобулинподобный домен содержит петельную структуру, обычно прикрепленную посредством дисульфидной связи между двумя цистеинами на конце каждой петли. ICAM-1 включает пять иммуноглобулинподобных доменов; ICAM-2, который отличается от ICAM-1 распределением клеток, включает два таких домена; PECAM-1 - шесть; VCAM - шесть или семь, в зависимости от вариантов соединения, и т. д. Более того, CAM обычно включают гидрофобную "трансмембранную" область, которая, как считают, участвует в ориентации молекулы у поверхности клетки, и карбоксиконцевую "цитоплазматическую" область. Графические модели расположения CAM в действии обычно показывают молекулу, прикрепленную в клеточной мембране у трансмембранной области к цитоплазматическому концевому сегменту, проходящему в клеточную цитоплазму, и одной или более иммуноглобулиноподобных петель, выходящих за пределы поверхности клетки.
Количество нейронных клеток выражает поверхностные рецепторы с внеклеточными Ig-подобными доменами, структурно схожими с ICAM. См., например, Yoshihara et al. В дополнение к Ig-подобным доменам многие адгезивные молекулы нервной системы также содержат тандемно повторяющиеся фибронектинподобные последовательности во внеклеточном домене.
Использование в терапевтических целях факторов межклеточной адгезии было расширено вследствие способности ICAM-1 связывать "H"-риновирусы. Например, в европейской патентной заявке N 468257 A, опубликованной 29 января 1992 г., описывается развитие многочисленных конфигураций и форм ICAM-1 (включая полномерные и процессированные формы молекул), предложенных для достижения улучшенной лиганд/рецепторной связывающей активности, особенно при связывании с вирусами, лимфоцит-ассоциированными антигенами и патогенами, такими как Plasmodium falciparum.
Подобным образом было расширено использование протеинов, иммунологически связанных с факторами межклеточной адгезии. Например, в заявке WO 91/16928, опубликованной 14 ноября 1991 г., описаны гуманизированные химерные антитела анти-ICAM-1 и их использование при лечении специфического и неспецифического воспаления, вирусной инфекции и астмы. Антитела анти-ICAM-1 и их фрагменты описаны как полезные при лечении эндотоксинового бактериально-токсического шока в заявке WO 92/04034, опубликованной 19 марта 1992 г. Ингибирование ICAM-1-эависимых воспалительных реакций антиидиотипическими антителами анти-ICAM-1 и фрагментами антител описано в заявке WO 92/06119, опубликованной 16 апреля 1992 г.
Несмотря на значительные успехи, достигнутые при проникновении в сущность явлений клеточной адгезии в результате идентификации и исследования белков межклеточной адгезии, таких как ICAM-1, и взаимодействующих с лимфоцитами интегринов, таких как LFA-1, общая картина далека от завершенности. Обычно считают, что множество других белков участвует в воспалительных процессах и в направленном движении лимфоцитов в теле. Например, в патентных заявках США NN 07/827689, 07/889724, 07/894061 и 08/009266 и соответствующей опубликованной заявке PCT WO 93/14776 (опубликованной 5 августа 1993 г.) описано клонирование и экспрессия ICAM-ассоциированного белка, ICAM-R. Описания изобретений этих заявок частично включены как ссылочный материал в данную заявку, и ДНК и аминокислотные последовательности ICAM-R представлены в ней в SEQ ID N 4. Было обнаружено, что этот новый лиганд был выражен на человеческих лимфоцитах, моноцитах и гранулоцитах.
Особый интерес для настоящей заявки представляет другой идентифицированный ICAM-подобный поверхностный фактор, тканевая специфическая экспрессия которого отличает его от других известных факторов ICAM. В работе Mori et al. [Proc. Natl. Acad. Sci. (USA), 84: 3921-3925 (1987)] приводится идентификация антигена конечного мозга кролика, обладающего специфической иммунореактивностью с моноклональным антителом 271A6. Этот поверхностный антиген получил название теленцефалин. В работе Imamura et al. [Neurosci. Letts., 119: 118-121 (1990)] при использовании поликлонального антитела для определения локализованной экспрессии утверждалось, что экспрессия теленцефалина в зрительной зоне коры головного мозга кошек показывает изменение в слоях ткани, а также сообщалось, что экспрессия теленцефалина изменяется как функция развития. Затем в работе Oka et al. [Neuroscience, 35: 93-103 (1990)] сообщалось о выделении теленцефалина при использовании моноклонального антитела 271A6. В публикации сообщается, что молекулярная масса поверхностной молекулы около 500 kD и что молекула состоит из четырех субъединиц, каждая с нативной молекулярной массой 130 kD, и примерно 100 kD - последующая обработка N-глюканазой. В работе Yoshihiro et al. [Neuroscience, Research Supplement, 18, p. S83 (1994)] сообщается о кДНК и аминокислотных последовательностях для теленцефалина кролика на 17-й ежегодной встрече Японского неврологического общества в Нагойе, Япония, 7-9 декабря 1993 г., и на 23-й ежегодной встрече неврологического общества в Вашингтоне, 9 ноября 1993 г. [Society for Neuroscience Abstracts 19 (1-3), p.646 (1993)]. Установленная аминокислотная последовательность позволяла предположить, что теленцефалин с молекулярной массой 130 kD является интегральным белком мембраны с девятью внеклеточными иммуноглобулин(Ig)подобными доменами. Из них восемь периферических доменов показали гомологию к другим Ig-подобным доменам ICAM. Такие же данные содержатся в работе Yoshihara et al., Neuron 12: 543-553 (1994).
Таким образом, возникла необходимость в выявлении дополнительных белков, участвующих в межклеточных взаимодействиях человеческого организма, и, в особенности, необходимость в информации, служащей для специфической идентификации и исследования таких белков в том, что касается их аминокислотной последовательности. Более того, исходя из факта, что такие молекулы могут образовать основу для развития терапевтических и диагностических средств, важно, чтобы получила разъяснение кодирующая их ДНК. Такая конструктивная информация обеспечила бы, между прочим, крупномасштабное производство белков, позволила бы идентифицировать клетки, продуцирующие их естественным путем, и подготовить антитела или другие новые связывающие белки, специфически реактивные с ними и/или тормозящие лиганд/рецепторные связывающие реакции, в которых они задействованы.
В одном из этих аспектов настоящее изобретение направлено на получение очищенных и выделенных полинуклеотидов (например, последовательностей ДНК, транскриптов РНК и их десенсибилизирующих олигонуклеотидов), кодирующих новый полипептид, ICAM-4, а также его полипептидные варианты (включая фрагменты и аналоги делеции, замены и присоединения), которые воспроизводят одну или более лиганд/рецепторные связывающие биологические активности и/или иммунологические свойства, характерные для ICAM-4. Присущие ICAM-4 лиганд/рецепторные связывающие биологические активности включают в себя взаимодействия как внеклеточных, так и цитоплазматических доменов ICAM-4 с другими молекулами (например, в процессе межклеточной адгезии и/или сигнальной трансдукции). Предпочтительные последовательности ДНК по настоящему изобретению включают геномные и кДНК-последовательности, а также последовательности полностью или частично химически синтезированных ДНК. Предпочтительный в настоящее время полинуклеотид представлен в SEQ ID N 1 и кодирует крысиный видовой ICAM-4. Рассматриваются биологические реплики (т.е. копии выделенных последовательностей ДНК, осуществленных in vivo или in vitro) последовательностей ДНК по данному изобретению. Также рассматриваются автономно реплицирующиеся рекомбинантные конструкции, такие как векторы плазмиды и вирусной ДНК, включающие последовательности ICAM-4 и, особенно, векторы, в которых ДНК, кодирующая ICAM-4 или вариант ICAM-4, соединена в действии с ДНК-последовательностью контроля эндогенной или экзогенной экспрессии.
В соответствии с другим аспектом изобретения клетки-хозяева, особенно одноклеточные клетки-хозяева, такие как прокариотические или эукариотические клетки, устойчиво трансформируются последовательностями ДНК по данному изобретению таким образом, чтобы в них были выражены нужные полипептиды. Клетки-хозяева, выражающие такие продукты ICAM-4 и вариантов ICAM-4, могут использоваться в самых различных целях. В той степени, в какой выраженные продукты "отображены" на поверхностях клеток, клетки могут образовывать полезный иммуноген для развития антител, специфически иммунореактивных с ICAM-4 и вариантами ICAM-4. Клетки-хозяева по данному изобретению очень полезны в способах крупномасштабного получения ICAM-4 и вариантов ICAM-4, где клетки выращиваются в соответствующей культурной среде и нужные продукты полипептида выделяются из клеток или из среды, в которой выращиваются клетки.
Новые ICAM-4 по данному изобретению могут быть получены как изоляты из источников естественных клеток, но, наряду с продуктами вариантов ICAM-4, их предпочтительно получают посредством рекомбинантных методов с участием клеток-хозяев по данному изобретению. Предпочтительная в настоящее время аминокислотная последовательность для полипептида IСAM-4 представлена в SEQ ID N 2. Продукты могут быть получены в полностью или частично гликозилированной, частично или полностью дегликозилированной или негликозилированной форме, в зависимости от клетки-хозяина, выбранной для рекомбинантного получения и/или процессирования после выделения. Варианты ICAM-4 по данному изобретению могут содержать водорастворимые или нерастворимые мономерные, мультимерные или циклические фрагменты ICAM-4, которые включают все или часть одной или более областей доменов, описанных выше и имеющих биологическое или иммунологическое свойство ICAM-4, включая, например, способность связываться со связывающим партнером ICAM-4 и/или ингибировать связывание ICAM-4 с естественным связывающим партнером. Варианты ICAM-4 по данному изобретению могут также содержать аналоги полипептида, в которых одна или более определенных аминокислот исключается или заменяется: (1) без утраты и преимущественно с улучшением одной или более биологических активностей или иммунологических характеристик, специфических для ICAM-4, или (2) с конкретной утратой определенной функции лиганд/рецепторного связывания. Рассматриваются аналогичные полипептиды, включая дополнительные аминокислотные (например, лизина или цистеина) остатки, которые облегчают мультимерное образование.
В настоящем изобретении рассматриваются также антитела (например, моноклональные и поликлональные антитела, фрагменты антител, антитела простой цепи, химерные антитела, антитела с транслантированным CDR и т.п.) и другие связывающие белки (например, полипептиды и пептиды), которые являются специфическими (т.е. нереактивными с факторами межклеточной адгезии ICAM-1, ICAM-2 и ICAM-R, которым ICAM-4 структурно близок) для ICAM-4 или вариантов ICAM-4. Изобретение затрагивает также клеточные линии гибридомы, которые выделяют моноклональные антитела в соответствии с данным изобретением. Предпочтительными гибридомами по данному изобретению являются 127A, 127H, 173E, 179I и 179H. Антитела могут быть развиты при использовании выделенных естественных или рекомбинантных ICAM-4 или вариантов ICAM-4, или клеток, выражающих такие продукты на их поверхностях. Связывающие белки по данному изобретению дополнительно используются для исследования структур(ы) связывающего сайта (например, эпитопов и/или восприимчивости связывающих свойств к модификациям в аминокислотной последовательности ICAM-4).
Связывающие белки используются, в свою очередь, в составах для иммунизации, а также для очистки полипептидов по данному изобретению и идентификации клеток, воспроизводящих полипептиды на их поверхностях. Они также используются при модулировании (в т.ч. блокировании, торможении или стимулировании) лиганд/рецепторных связывающих биологических активностей с участием ICAM-4, в особенности тех эффекторных функций ICAM-4, которые связаны со специфическими и неспецифическими ответами иммунной системы. Рассматриваются также антиидиотипические антитела, специфические для антител анти-ICAM-4, и применение таких антиидиотипических антител в модулировании иммунных ответов. При анализе для обнаружения и количественного определения ICAM-4 на поверхностях клеток и в жидкостях организма, например в серозной или цереброспинальной жидкостях, может быть использовано, например, единственное антитело или множество антител в форме многослойного иммуносэндвича. При обнаружении ICAM-4 в жидкости организма антитела по данному изобретению также используются для анализа проявления невропатологий, которые могут соотноситься с повышенными уровнями циркулирующего ICAM-4. Такие нейропатологии включают (но не ограничиваются) церебральную ишемию (т.е. приступ), являющуюся результатом различных нарушений, включающих, например, тромбоз, эмболию и т.п.
Очевидна научная ценность информации, полученной при раскрытии сущности ДНК- и аминокислотных последовательностей по настоящему изобретению. Как показано на ряде примеров, знание последовательности кДНК для ICAM-4 делает возможным выделение, посредством гибридизации ДНК/ДНК, последовательностей геномных ДНК, кодирующих ICAM-4, и определение регуляторных последовательностей контроля экспрессии ICAM-4, таких как промоторы, операторы и т.п. Ожидается, что методы гибридизации ДНК/ДНК, осуществляемые при помощи последовательностей ДНК по данному изобретению и при соблюдении жестких условий, позволят выделять ДНК, кодирующие аллельные варианты ICAM-4, другие структурно родственные белки, обладающие одним или более биологическими и/или иммунологическими свойствами, специфическими для ICAM-4, и белки, гомологичные ICAM-4, из других видов. ДНК по данному изобретению используются при анализе гибридизации ДНК/РНК для определения способности клеток к синтезированию ICAM-4. Данным изобретением обеспечивается также наличие десенсибилизирующих полинуклеотидов, релевантных регулированию экспрессии ICAM-4 такими клетками, которые первоначально выражали то же самое. Как видно из другого ряда примеров, знание ДНК- и аминокислотных последовательностей ICAM-4 делает возможным образование посредством рекомбинантных средств вариантов ICAM-4, таких как гибридные слитые белки (которые иногда относят к "иммуноадгезивам"), характеризующиеся наличием последовательностей белков ICAM-4 и константных областей тяжелых цепей иммуноглобулина и/или шарнирных областей. [См. Capon tn al. , Nature, 337: 525-531 (1989); Ashkenazi et al., P.N.A.S.(USA), 88: 10535-10539 (1991) и заявка PCT WO 89/02922, опубликованная 6 апреля 1989 г. ] Слитые белки могут также включать, например, выбранные внеклеточные домены ICAM-4 и части других факторов клеточной адгезии.
ДНК по настоящему изобретению также позволяет осуществлять идентификацию нетранслированных последовательностей ДНК, которые специфически стимулируют экспрессию полинуклеотидов, соединенных в действии с промоторными областями. Идентификация и использование таких промоторных последовательностей особенно желательны в случаях, например, переноса генов, который может требовать конкретно гетерогенной экспрессии в ограниченной нейронной среде. В изобретении также рассматриваются векторы, содержащие промоторы по данному изобретению, а также конструкции рекомбинантных генов, в которых промотор по данному изобретению связан в действии с гетерогенной полинуклеотидной последовательностью и сигналом терминации транскрипции.
Информация о ДНК- и аминокислотных последовательностях, полученная в соответствии с данным изобретением, также дает возможность систематического анализа структуры и функции ICAM-4 и определения тех молекул, с которыми он будет взаимодействовать на внеклеточном и внутриклеточном уровнях. Идиотипы моноклональных антител ICAM-4 по настоящему изобретению характерны для таких молекул и могут имитировать естественные связывающие белки (пептиды и полипептиды), посредством которых модулируются межклеточные и внутриклеточные активности ICAM-4, или при помощи которых ICAM-4 модулирует межклеточные и внутриклеточные явления. В альтернативном варианте они могут представлять новые классы модуляторов активностей ICAM-4. Антиидиотипические антитела, в свою очередь, могут представлять новые классы биологически активных эквивалентов ICAM-4. Анализ in vivo для идентификации антител или других соединений, которые модулируют активность ICAM-4, может включать, например, иммобилизацию ICAM-4 или естественного лиганда, с которым связан ICAM-4, маркирование при выявлении неиммобилизированного связывающего партнера, совместное инкубирование связывающих партнеров и определение влияния соединения, полученного при исследовании, на величину границы метки, когда уменьшение границы метки в присутствии полученного соединения, по сравнению с величиной границы метки в отсутствие полученного соединения, указывает на то, что испытываемое вещество является ингибитором связывания ICAM-4.
Информация о последовательностях ДНК, полученная в соответствии с данным изобретением, делает также возможным развитие посредством гомологической рекомбинации или принципа ударной сепарации частиц [см. Kapecchi, Science, 244: 1288-1292 (1989)] родентов, которые не могут выражать белок функционального ICAM-4 или которые выражают белок вариантного ICAM-4. Такие роденты используются как модели для изучения активностей ICAM-4 или модуляторов ICAM-4 in vivo.
В описание включены ссылки на родовую патентную заявку США N 08/102852, поданную 5 августа 1993 г. Примеры этой заявки касаются, в числе прочего, следующего: разработка и конструкция олигонуклеотидных зондов для PCR-амплификации ICAM-ассоциированных ДНК; использование зондов для амплификации геномного фрагмента человека, гомологичного, но отличающегося от ДНК, кодирующих ICAM-1 и ICAM-2; скрининг библиотек кДНК с геномными фрагментами для выделения дополнительных кодирующих ICAM-R последовательностей; скрининг библиотек кДНК для выделения полномерной последовательности человеческой кДНК, кодирующей ICAM-R; исследование информации о ДНК- и аминокислотных последовательностях для ICAM-R, особенно в отношении ICAM-1 и ICAM-2; развитие клеток-хозяев млекопитающего, выражающих ICAM-R; оценка указаний об участии ICAM-R в явлениях адгезии с участием CD18-зависимых и CD18-независимых путей; подавление клеточной адгезии с ICAM-R посредством извлеченных из ICAM-R пептидов; экспрессия вариантов ICAM-R; подготовка и исследование антител анти-ICAM-R и их фрагментов; картирование эпитопов ICAM-R, распознанных моноклональными антителами анти-ICAM-R; оценка распределения и биологическое исследование ICAM-R и РНК, кодирующих одно и то же; оценка ICAM-R в гомотипической межклеточной адгезии и иммунной клеточной активации/пролиферации; исследование моноклональных антител ICAM-R; оценка различных фосфориляционных и цитоскелетных ассоциаций цитоплазматического домена ICAM-R. Также была описана идентификация кодирующей родентный ICAM ДНК, которая оказалась на этот раз крысиным гомологом человеческого ICAM-R, и использование этой ДНК для конструирования и выражения ДНК, кодирующих слитые белки глутатион-S-трансферазы. Подробное описание идентификации этой родентной ДНК можно найти в родовой заявке (U. S.S. N 08/102852) в примере 6, и оно воспроизводится здесь в примере 1. Когда были идентифицированы другие последовательности, кодирующие родентный ICAM, стало очевидно, что ДНК родентного ICAM не кодирует крысиный гомолог человеческого ICAM-R, но, фактически, кодирует новый полипептид ICAM, названный ICAM-4. Чтобы оценить явления, которые привели к идентификации ICAM-4, соблюдается хронология, сопровождаемая детальным описанием изобретения.
Была идентифицирована первая последовательность родентного геномного ICAM-4, которая кодировала область, гомологичную домену 2 (здесь SEQ ID N 3 и SEQ ID N 23 в U.S.S. N 08/102852) человеческого ICAM-R (здесь SEQ ID N 4). Была также идентифицирована вторая, перекрывающая геномная ДНК (здесь SEQ ID N 5 и SEQ ID N 26 в U.S.S. N 08/102852), которая кодировала как область домена 2 в соответствии с SEQ ID N 3, так и последовательности для ICAM-1. Используя SEQ ID N 3 как зонд, была идентифицирована родентная кДНК селезенки (здесь SEQ ID N 6, SEQ ID N 25 U.S.S. N 08/102852), которая кодировала домены 2-5, а также пятый домен, ранее не наблюдавшийся как домен ICAM. На этот раз вновь идентифицированные родентные ДНК кодировали родентный гомолог человеческого ICAM-R, однако сравнительный анализ 3' областей этих ДНК с другими ICAM оказался затруднительным.
Последующее выделение клона ДНК в 1 kb из библиотеки крысиной селезенки и амплификация фрагмента RT-PCR показали, что часть как клонов кДНК, так и геномных клонов не были секвенированы. Другой продукт амплификации RT-PCR (SEQ ID N 7) подтвердил это. Было установлено, что фрагмент из 177 bp был удален из геномных клонов и клонов кДНК посредством EcoRI-гидролиза клонов, чтобы выделить эти последовательности из λ- фага для изучения секвенирования ДНК. Повторный анализ SEQ ID N 5 и 6 в свете этих других последовательностей позволил идентифицировать более точные и полные последовательности для первоначально выделенных геномных клонов и клонов кДНК, представленных здесь в исправленном виде в SEQ ID N 8 и 9.
Чтобы идентифицировать полную кодирующую последовательность для ICAM-4, была выделена кДНК головного мозга крысы (SEQ ID N 10) и определена 5' концевая последовательность посредством 5' быстрой амплификации концов кДНК (5' RACE); продукт амплификации представлен в SEQ ID N 11. Объединение информации клона RT-PCR (SEQ ID N 7), кДНК головного мозга (SEQ ID N 10) и продукта амплификации RACE (SEQ ID N 11) позволило идентифицировать полную кодирующую последовательность для ICAM-4 (SEQ ID N 1).
Таким образом, настоящее изобретение проиллюстрировано следующими примерами. Более конкретно. Пример 1 относится к клонированию ДНК частично родентного ICAM-4. В примере 2 представлен анализ методом назерн-блоттинга транскрипции родентного ICAM-4. В примере 3 показано выделение кДНК полномерного родентного ICAM-4. Пример 4 относится к гибридизации in situ родентного ICAM-4 в мозговой ткани. Пример 5 относится к образованию слитых белков ICAM-4 в прокариотах. В примере 6 описано получение моноклональных антител, специфических для крысиных слитых белков ICAM-4/GST. В примере 7 описана экспрессия растворимых крысиных белков ICAM-4 методом бакуловирусной экспрессии. Пример 8 относится к получению моноклональных антител, специфических для крысиных ICAM-4, выраженных бакуловирусным методом. В примере 9 описан иммуноцитохимический анализ экспрессии крысиного ICAM-4. Пример 10 относится к клонированию человеческой геномной ДНК, кодирующей ICAM-4. Пример 11 относится к клонированию человеческой кДНК, клонирующей ICAM-4. В примере 12 описан назерн-анализ экспрессии человеческого ICAM-4. В примере 13 описано образование человеческих слитых белков ICAM-4/GST. Пример 14 относится к получению моноклональных антител, иммунологически специфических для человеческого ICAM-4. В примере 15 описано развитие анализа захватом для определения концентрации растворимого ICAM-4 в конкретной жидкости. В примере 16 показано применение метода анализа захватом для определения концентрации ICAM-4 в серозной жидкости больных припадками. Пример 17 относится к анализу транскрипции ICAM-4 в модели крысиной эпилепсии. Пример 18 относится к клонированию области промотора для человеческого ICAM-4.
ПРИМЕР 1. Клонирование крысиной ICAM-ассоциированной ДНК.
A. Выделение крысиной геномной ICAM-ассоциированной ДНК домена 2.
Крысиная геномная библиотека, сконструированная в λEMBL3, была проверена при помощи [32P]-меченого зонда, полученного посредством PCR из ДНК, кодирующей домен 2 человеческого ICAM-3. Последовательность зонда представлена в SEQ ID N 12. Пятна библиотеки были перенесены на Hybond N+ найлоновые мембраны (Amercham, Arlington Heights, IL). Скрининг всех кДНК и геномных библиотек был проведен в соответствии со стандартными протоколами. Предгибридизация и гибридизации были проведены в растворе 40-50% формамида, 5X Denhardt's, 5X SSPE и 1,0% SDS при 42oC. Зонды ([32P] -меченые) были добавлены при концентрации раствора гибридизации 105-106 cpm/ml. После этого в течение 16-18 часов гибридизации найлоновые мембраны подвергались экстенсивному промыванию при комнатной температуре в растворе 2X SSPE с 0,1% SDS, а затем подвергнуты воздействию рентгеновской пленки при -80oC в течение ночи. Положительные пятна были подвергнуты одному или более циклам гибридизации, чтобы получить клональный фаг. ДНК, приготовленная из лизата позитивных клонов, была субклонирована в pBS+ и секвенирована.
Был идентифицирован первый геномный клон, кодирующий крысиный ICAM-ассоциированный домен 2, который был определен как гомологичный областям домена 2 в других членах семейства ICAM (см., например, табл. 1 патентной заявки США N 08/102852), но отличался от ранее известных нуклеотидных последовательностей для крысиного ICAМ-1 [Kita et al., Biochem. Biophys. Acta, 1131: 108-110 (1992)] или мышиного ICAM-2 [Xu et al., J.Immunol., 149: 2560-2565 (1992)] . Нуклеиновокислотные и установленные аминокислотные зависимости для этого клона были описаны в одновременно поданных родовых заявках как вариантные формы крысиного ICAM-R и были представлены как SEQ ID N 23 и 24 соответственно в U. S.S. N 08/102852. В данной заявке эти же последовательности представлены в SEQ ID N 3 и 13 соответственно.
Второй, перекрывающий клон был также идентифицирован с теми же зондами и был определен как содержащий последовательность домена 2 ICAM (SEQ ID N 3) и 5' ДНК, кодирующую, по крайней мере, часть крысиного ICAM-1. Нуклеиновокислотная последовательность для этого клона была представлена в одновременно поданной родовой заявке как SEQ ID N 26, а в данной заявке - как SEQ ID N 5. Этот второй клон показал, что фрагмент ICAM-ассоциированного гена первого клона и ген, кодирующий крысиный ICAM-1, расположены на одной и той же хромосоме крысы в пределах 5 kb друг от друга.
B. Выделение крысиной, ICAM-ассоциированной кДНК.
Чтобы идентифицировать более полную, кодирующую белок последовательность для ICAM-ассоциированного полипептида, [32P]-меченая ДНК, кодирующая последовательность домена 2 из крысиного геномного клона, идентифицированного в Секции A (SEQ ID N 3), как указано выше, была использована для скрининга части библиотек кДНК из различных типов клеток крыс и мышей, включая крысиный макрофаг (Clontech. Palo Alto, CA), лимфоцит периферической крови (PBL) (Clontech), T-клетку (сконструированную внутри) и B-клетку (сконструированную внутри).
Был идентифицирован единичный клон в библиотеке кДНК крысиной селезенки (Clontech), который содержал пять Ig-подобных доменов, четыре из которых были гомологичными доменам 2-5 как в ICAM-1, так и в ICAM-R. Более того, этот клон включал 3' ДНК, кодирующую явный пятый Ig-подобный домен, который не был ранее идентифицирован в каком-либо другом полипептиде ICAM. Кроме того, клон содержал необычную 3' последовательность, определенную впоследствии как частичный интрон (описан ниже), расположенный между доменами 4 и 5, что дает возможность предположить, что клон являлся продуктом неразвитого транскрипта или транскрипта, полученного при отклоняющемся от нормы сплайсинге. Присутствие уникального домена и определение того факта, что 3' область не сравнивалась должным образом с другими известными ICAM, дало возможность предположить, что ICAM-ассоциированная ДНК может кодировать новый крысиный полипептид ICAM. Нуклеиновокислотная последовательность для этого клона была представлена в родственной заявке как SEQ ID N 25; в данной заявке нуклеиновокислотная последовательность для этого клона кДНК селезенки представлена в SEQ ID N 6.
C. Повторный анализ крысиной кДНК и геномных ДНК.
После подачи 5 августа 1993 г. патентной заявки США N 08/102852 было определено, что в частичном клоне кДНК крысиной селезенки (SEQ ID N 25 в родовой заявке и SEQ ID N 6 в данной заявке) и геномном клоне крысиной печени (SEQ ID N 26 в родовой заявке и SEQ ID N 5 в данной заявке) был пропущен внутренний фрагмент EcoRI в 177 bp, который являлся частью каждого из этих клонов, но был потерян на стадии субклонирования, когда вставки библиотеки были удалены из α-вектора гидролизом EcoRI и лигированы в секвенирующий вектор. Предположение о том, что в клоне ДНК и геномном клоне мог быть пропущен кодирующий фрагмент, стало очевидно при сравнении крысиных геномной и ДНК-последовательностей с различными продуктами амплификации RT-PCR, включая SEQ ID N 7, которая обнаружила разрыв в крысиной последовательности.
Последующее выделение и сравнительный анализ последовательностей кДНК из библиотеки селезенки с использованием клона кДНК селезенки (SEQ ID N 6) как зонда дали возможность впервые определить, что часть клонов кДНК селезенки и геномного не была секвенирована. Дальнейшее подтверждение этого предположения стало очевидным при амплификации фрагмента RT-PCR, охватывающего домены 3-5, с использованием 5' праймера (RRD3 5' Xho, содержащего 5' XhoI сайт рестрикции для облегчения клонирования), представленного в SEQ ID N 14, и 3' праймера (RRD5 3' Hind, включающего сайт HindIII для облегчения клонирования), представленного в SEQ ID N 15.
GAACTCGAGGCCATGCCTCCACTTCC (SEQ ID N 14)
CCATAAGCTTTATTCCACCGTGACAGCCAC (SEQ ID N 15)
Сравнительный анализ этих двух ДНК ясно показал, что в клонах ДНК и геномном был утерян фрагмент перед секвенированием; это предположение нашло дальнейшее подтверждение в ходе секвенирования ДНК RT-PCR (описано ниже). Был сделан вывод, что гидролиз с EcoRI для удаления кДНК- и геномного фрагментов перед секвенированием имел своим результатом удаление фрагмента из 177 bp, который не был определен визуально при выделении клонов агарозным гелем из последовательностей λ- фага. Последующий анализ последовательностей подтвердил расположение двух сайтов EcoRI, примыкающих к фрагменту из 177 bp в обоих первоначальных клонах.
Фрагмент EcoRI из 177 bp расположен между нуклеотидами 719 и 896 в крысином частичном клоне кДНК, как представлено в SEQ ID N 9, и между нуклеотидами 2812 и 2989 в частичном геномном клоне, как представлено в SEQ ID N 8.
D. ДНК, выявленная клоном RT-PCR.
RT-PCR использовалась для получения более полной информации последовательности для крысиного, ICAM-ассоциированного гена. Информация последовательности из геномного клона (SEQ ID N 3) была использована для конструирования сенсибилизирующих праймеров, комплементарных 5' области из кодирующей белок области, как определено из клона кДНК, и десенсибилизирующих праймеров, комплементарных кодирующим последовательностям и 3' областям, к кодирующей последовательности в клоне кДНК (SEQ ID N 6).
кДНК темплаты для реакций PCR была приготовлена следующим образом. Приблизительно 2 мкг поли-A+ РНК, выделенной из клеток крысиной селезенки, было денатурировано нагреванием при 65oC в объеме 10 мкл. После денатурации было добавлено 0,1 мкл RNasin (Invitrogen, San Diego, CA), 5 мкл 5X RTase Buffer (BRL, Bethesda, MD), 2 мкл произвольного гексамера pd(N)6 при 100 мкг/мл (Pharmacia, Piscataway, NJ), 6 мкл dNTPs (каждого по 2 мМ) и 2 мкл AМV RTase (BRL), и реакция была инкубирована при 42oC в течение 60-90 минут. Реакции хранились при 20oC до нужного времени.
Первоначальная серия экспериментов проводилась для идентификации пар праймеров олигонуклеотидов, которые вырабатывали продукт амплификации в реакциях PCR с использовавшем кДНК крысиной селезенки как темплаты. Различные 5' сенсибилизирующие праймеры подбирались в PCR в пару с 3' праймером, который был сконструирован комплементарным внутренней, кодирующей последовательности; 3' праймер был обозначен RRD2 3-1 и представлен в SEQ ID N 16.
AACGTGCGGAGCTGTCTG (SEQ ID N 16)
(В полностью выделенном продукте RT-PCR (SEQ ID N 7, приведенная ниже) праймер RRD2 3-1 соответствовал нуклеотидам 719-736). Подобным образом, различные 3' десенсибилизирующие праймеры спаривались с 5' праймером, комплементарным другой внутренней кодирующей последовательности; 5' праймер в этих реакциях был обозначен RGen3900S и представлен в SEQ ID N 17.
ACGGAATTCGAAGCCATCAACGCCAGG (SEQ ID N 17)
(В SEQ ID N 7 праймер RGen3900S соответствовал нуклеотидам 1719-1736.) На основе размера продуктов амплификации и способности этих продуктов к гибридизации с частичным клоном кДНК, одна пара праймеров была определена как наиболее эффективная, и она была использована в последующих амплификациях PCR. Праймер 5' был обозначен RGen 780S (SEQ ID N 18) и праймер 3' был обозначен RGen 4550AS (SEQ ID N 19).
CATGAATTCCGAATCTTGAGTGGGATG (SEQ ID N 18)
ATAGAATTCCTCGGGACACCTGTAGCC (SEQ ID N 19)
(В SEQ ID N 7, приведенной ниже, праймер RGen780S соответствовал нуклеотидам 1-18, а праймер RGen4550AS соответствовал нуклеотидам 2197-2214.)
Эта пара праймеров была использована в PCR при различных условиях для оптимизации амплификации. Все 15 различных буферов PCR, которые варьировались в концентрации pH и Mg++, были использованы при двух различных температурах отжига, и образец продукта каждой реакции был разделен на 1% агарозном геле. Так как никакой продукт амплификации не мог быть определен при визуальном осмотре окрашенного этидным бромидом геля при любых условиях реакции, была применена более чувствительная саузерн-гибридизация для определения продуктов PCR.
Слои амплифицированной ДНК были разделены электрофорезом, перенесены на Hybond+ найлоновую мембрану при использовании обычных методов саузерн-блоттинга и гибридизированы с полной крысиной кДНК, которая была [32P]-меченой. Условия гибридизации были, в основном, такими, как описанные выше для метода скрининга библиотеки в Секции A. Авторадиография показала, что небольшое количество ДНК (приблизительно 2,2 kb) было получено в двух из этих реакций, а остальной продукт амплификации от этих двух реакций был разделен на агарозном геле. Область из 2,2 kb была выделена из геля, хотя при визуальном осмотре никакого бэнда не было заметно, и использована как темплат в другой реакции PCR с применением тех же праймеров (SEQ ID N 18 и 19), буфера Tris-HCl, pH 8,0, с содержанием 1 mM Mg++, и при температуре отжига 55oC. Продукт амплификации после вторичной PCR был видимым в геле, и он был выделен и клонирован в плазмиду pBS+ (Stratagene, La Jolla, CA) для анализа последовательности.
Было определено, что полученный в результате PT-PCR клон содержал 2214 bp, как представлено в SEQ ID N 7. Клон кодировал домены 2-6, обнаруженные в клоне кДНК крысиной селезенки, дополнительный аминоконцевой домен 1, дополнительный карбоксиконцевой домен 7 и 164 bp, которые оказались еще одним карбоксиконцевым доменом 8. Непосредственно 5' к домену 1 была дополнительная последовательность из 144 bp, предположительно, извлеченная из интрона между лидерной последовательностью и первым доменом. Этот клон не содержал 5' лидерную последовательность или 3' трансмембранную и цитоплазматическую области. В дополнение к предварительно идентифицированному домену 6 в клоне кДНК селезенки 7-й и 8-й домены в клоне RT-PCR подтвердили гипотезу о том, что этот клон был новым родентным ICAM.
ПРИМЕР 2. Анализ методом назерн-блоттинга.
Для дальнейшего исследования возможности того факта, что ICAM-ассоциированные клоны, идентифицированные в примере 1, кодировали новый полипептид ICAM, как предполагалось по уникальным Ig-подобным доменам, тканевая специфическая экспрессия была обследована при помощи метода назерн-блоттинга для возможности сравнения с ранее известными типами экспрессии человеческих ICAM. [ICAM-1, Dustin et al. , J.Immunol., 137: 245-254 (1986); ICAM-2, Staunton et al. , Nature, 339: 61-64 (1989); ICAM-R, de Fourgerolles and Springer, J.Exp.Med., 174: 185-190(1992)].
Полная клеточная РНК из крысиных легкого, головного мозга, спинного мозга, печени, пищеварительного тракта, вилочковой железы, лимфатических узлов и селезенки была приготовлена с использованием реагентов выделения РНК STAT60 (Tel-test "B", Inc. Friendswood, Texas) в соответствии с протоколом, предложенным предводителем. Поли- A+ РНК была очищена от полной РНК с использованием колонок с олиго-dT-целлюлозой. Приблизительно 5 мкг РНК, извлеченной из каждой ткани, было разделено на 1% формальдегидном агарозном геле и перенесено на hybond-C нитроцеллюлозные мембраны (Amersham).
Фрагмент кДНК крысиной селезенки из примера 1, соответствующий доменам 2-4 (нуклеотиды 1-724 в SEQ ID N 6), был субклонирован в pBluescript SK+ (Strategene), и десенсибилизирующий рибозонд был образован посредством транскрипции in vitro с использованием [32P]-меченого UTP и приблизительно 500 ng линеаризированного темплата в соответствии с протоколом, предложенным производителем (Boehringer Mannheim, Indianapolis, IN). Связанная с мембраной РНК была предварительно гибридизирована в растворе, содержащем 50% формамида, 5X SSC, 1X PE (50 mM Tris-HCl, pH 7,5, 0,1% пирофосфата натрия, 0,2% поливинилпирролидона, 0,2% фиколла, 5 mM EDTA, 1% SDS) и 150 мкг/мл денатурированной ДНК спермы лосося. Меченный радиоактивным изотопом зонд был денатурирован при кипении и добавлен к раствору предгибридизации до окончательной концентрации 1•106 cpm/ml. Гибридизация могла продолжаться в течение 16-18 часов при температуре 65oC. Затем мембраны были промыты при 65oC в 2X SSC, содержащем 0,1% SDS, и затем подвергались воздействию рентгеновской пленки в течение 3-16 часов.
Анализ методом назерн-блоттинга показал, что ICAM-ассоциированная кДНК, идентифицированная в примере 1, была выражена только в головном мозге крысы; эта тканевая специфичность не была ранее известна для каких-либо других полипептидов ICAM. Этот тип экспрессии, в сочетании с уникальными Ig-подобными доменами, неизвестными в других полипептидах ICAM, показал, что ICAM-ассоциированный клон был новым членом ICAM-семейства протеинов, и он был назван ICAM-4.
Тот факт, что первоначально идентифицированные клоны кДНК были выявлены в библиотеке крысиной селезенки, позволил предположить, что субпопуляция клеток в селезенке может выражать ICAM-4 на низких уровнях. Однако клоны, должным образом подвергнутые сплайсингу, не могли быть выявлены в многочисленных библиотеках гемопоэтических кДНК, что привело к сомнениям по поводу того, выражен ли действительно белок ICAM-4 в какой-либо ткани кроме головного мозга. Одним объяснением выявления кДНК ICAM-4 в селезенке является то, что чувствительность PCR могла амплифицировать незначительное количество транскрипта, даже несмотря на то, что эти ткани не выражают кодированный белок.
ПРИМЕР 3. Выделение полномерной кДНК крысиного ICAM-4.
А. Идентификация клона кДНК крысиного головного мозга.
Принимая во внимание специфическую экспрессию ICAM-4 в ткани, мРНК ткани головного мозга была использована при попытке выделить полномерную кДНК, кодирующую ICAM-4. Два зонда - один, комплементарный доменам 1-2, и другой, комплементарный доменам 3-5 клона кДНК селезенки, идентифицированного в примере 1 (SEQ ID N 7),- были мечены радиоактивным изотопом и использованы для скрининга библиотеки кДНК крысиного головного мозга в λgt10, сконструированного предварительно внутри. Условия гибридизации были такими, как описанные в примере 1, и положительные пятна были подвергнуты одному или более циклов скрининга для получения клонального фага.
Было идентифицировано девять положительных клонов, два из которых были гибридизированы к обоим зондам. Самый длинный из двух клонов, обозначенный как клон 7, содержал 2550 bp, кодирующих четыре из пяти Ig-подобных доменов, обнаруженных в кДНК зонда. Кроме того, клон 7 кодировал четыре других Ig-подобных домена, не обнаруженных в зонде. Были идентифицированы предполагаемые трансмембранные и цитоплазматические домены, которые сопровождались стоп-кодоном, сигналом полиаденилирования и поли-A хвостом. В клоне 7 не хватало, по крайней мере, одного 5' Ig-подобного домена, как было определено при сравнении с клоном RT-PCR (SEQ ID N 7), а также не хватало лидерной последовательности; повторный скрининг библиотеки не дал больше клонов, которые содержали бы эти последовательности. Последовательность нуклеиновой кислоты для клона 7 представлена в SEQ ID N 10.
B. Определение 5' конца.
Чтобы выделить домен 1 и другие 5' последовательности, метод PCR, названный быстрой амплификацией концов кДНК (RACE) [Протоколы PCR: Руководство по методам и применению, Innis et al. , (eds) Academic Press: New York (1990), pp, 28-38], был применен с использованием набора 5' RACE (Clontech). В этом методе используется внутренний праймер, спаренный со вторым праймером, комплементарным адаптерной последовательности, лигированной к 5' концу молекул библиотеки кДНК. PCR с этой парой праймеров будет поэтому амплифицировать и облегчать идентификацию интронов. Информация о перекрывающей последовательности может тогда бить использована для получения полной последовательности гена.
Готовая к RACE кДНК из головного мозга крысы (снабженная набором) была использована в PCR с олигонуклеотидом набора и десенсибилизирующим праймером на основе внутренней последовательности ICAM-4. 3' десенсибилизирующий праймер, обозначенный Spot714AS, был сконструирован в соответствии с последовательностью домена 4 ICAM- 4 и представлен в SEQ ID N 20.
CARGGTGACAAGGGCTCG (SEQ ID N 20)
Продукт амплификации, полученный от этой пары праймеров, был затем подвергнут вторичной PCR с использованием того же праймера 5' набора, спаренного с 3' праймером, комплементарным области в домене 1 ICAM-4. Второй 3' праймер был обозначен RRACE2 и представлен в SEQ ID N 21.
TATGAATTCAGTTGAGCCACCAGCGAGC (SEQ ID N 21)
Каждый праймер, использованный во вторичной PCR, содержал сайт EcoRI для облегчения клонирования полученных продуктов амплификации в pBS (Stratagene). Полученная плазмидная ДНК, которая содержала 5' конец гена, была идентифицирована гибридизацией к зонду доменов 1 и 2 крысиного ICAM-4, соответствующему нуклеотидам 1-736 в SEQ ID N 7. Информация о частичной последовательности для домена 1 и гидрофобной лидерной последовательности была определена из полученного продукта амплификации.
Продукт, полученный методом 5' RACE, был фрагментом ДНК длиной 222 bp, содержащим 60 bp в обратном направлении от инициирующей метиониновой группы, лидерную последовательность из 82 bp и последовательность из 80 bp из домена 1. Продукт амплификации представлен в SEQ ID N 11.
C. Полномерная последовательность крысиного ICAM-4.
Композиционный клон полномерного ICAM-4 был сконструирован при помощи информации последовательности, полученной из метода 5' RACE (SEQ ID N 11), клона RT-PCR (SEQ ID N 7) и клона 7 кДНК головного мозга (SEQ ID N 10). Было определено, что полномерный ген крысиного ICAM-4 содержит 2985 bp с единственной открытой рамкой считывания, кодирующей установленный белок аминокислоты 917. Предполагаемая последовательность Козака расположена в обратном направлении от метиониновой группы в лидерной последовательности. Гидрофобная лидерная последовательность аминокислоты 27 сопровождается девятью Ig-подобными доменами, трансмембранной областью и цитоплазматическим концевым сегментом аминокислоты 58. кДНК композиционного ICAM-4 представлена в SEQ ID N 1, а установленная аминокислотная последовательность представлена в SEQ ID N 2.
Как другие полипептиды ICAM, ICAM-4 содержит внеклеточные, трансмембранные и цитоплазматические домены. Во внеклеточном домене амино-конец ICAM-4 является лидерной последовательностью, содержащей аминокислоты 1-27, которая сопровождается девятью иммуноглобулин(Ig)подобными доменами; это признак, свойственный только ICAM-4, так как ICAM-1, ICAM-2 и ICAM-R содержат пять, два и пять внеклеточных Ig-подобных доменов соответственно. В ICAM-4 домен 1 содержит аминокислоты 28-118; домен 2 содержит аминокислоты 119- 224; домен 3 содержит аминокислоты 225-321; домен 4 содержит аминокислоты 322-405; домен 5 содержит аминокислоты 406-488; домен 6 содержит аминокислоты 489-569; домен 7 содержит аминокислоты 570-662; домен 8 содержит аминокислоты 663-742 и домен 9 содержит аминокислоты 743-830. В каждом домене характерная петельная структура сформирована посредством дисульфидной связи между цистеиновыми группами, расположенными обычно на противоположных концах аминокислотной последовательности домена. Другие структурные признаки ICAM-4 включают трансмембранную область, содержащую аминокислоты 831-859, и цитоплазматическую область, содержащую аминокислоты 860-917.
Сравнение гомологии аминокислотной последовательности каждого домена в крысином ICAM-4 с другими членами семейства ICAM было ограничено соответствующими последовательностями человеческого ICAM-1, ICAM-2 и ICAM-R, так как информация последовательности для всех трех родентных гомологов не была ранее известна. В первом домене родентный ICAM-4 показывает 21-, 30- и 28-процентную идентичность с человеческим ICAM-1, ICAM-2 и ICAM-R соответственно. Второй домен более консервативен и имеет 60-, 42- и 62-процентную идентичность с ICAM-1, -2 и -3 соответственно. Домены 3-5 показывают процентную идентичность 48, 49 и 50 с ICAM-1 и 60, 59 и 29 соответственно для ICAM-R. Интересно, что домены 6-8 крысиного ICAM-4 являются более гомологичными с доменом 5 (идентичность которого находится в диапазоне 29-42%), что, возможно, является следствием явления дупликации сегмента гена. Девятый и последний внеклеточный домен слабо сравним с другими доменами ICAM, но имеет 22-процентную идентичность с 3-м и 6-м доменами человеческого VCAM-1, другого члена Ig-семейства белка, который участвует в клеточной адгезии. Цитоплазматический концевой сегмент имеет длину 58 аминокислот. Он длиннее, чем другие члены семейства ICAM, в котором человеческие ICAM-1, -2 и -3 содержат 28, 26 и 37 аминокислот соответственно. Что касается девятого домена, цитоплазматический концевой сегмент крысиного ICAM-4 является наиболее гомологичным с цитоплазматическим концевым сегментом человеческого VCAM-1, который содержит только 19 аминокислот. Расположенные ближе к центру мембраны 19 аминокислот крысиного ICAM-4 разделяют 7 аминокислотных остатков с VCAM-1 (37%).
Наконец, функциональное связывание карт LFA1 (CD11a/CD18) с первым доменом в ICAM. В работе Vonderheide et al. [J.Cell Biol., 125: 215-222 (1994)] идентифицирован рисунок последовательности, предположительно имеющий место в связывании интегрина. Несмотря на относительно низкую гомологию между крысиным ICAM-4 и другими ICAM в домене 1, этот рисунок последовательности связывания сохранился, что дает возможность предположить, что крысиный ICAM-4 может быть лигандом для LFA-1 и, возможно, других интегринов.
ПРИМЕР 4. Гибридизация in situ в ткани головного мозга.
Чтобы локализировать специфическую ткань головного мозга, которая выразила ICAM-4, была применена гибридизация in situ с десенсибилизирующими рибозондами домена 1 ICAM-4 и доменов 3-4 ICAM-4. Зонды были мечены посредством транскрипции in vitro с использованием 35S-меченого UTP.
Замороженные срезы ткани обычного крысиного мозга были зафиксированы в 4% параформальдегиде в течение 20 минут, промыты и дегидрированы, и фиксированная РНК была денатурирована в течение двух минут в 2X SSC, 70% формамиде при 70oC перед гибридизацией. Срезы ткани были гибридизированы на протяжении ночи при 50oC в растворе, содержащем 50% формамид, 0,3 М NaCl, 20 mM Tris-HCl, pH 7,4, 5 mM EDTA, 10% декстрансульфат, IX Denhardt, 0,5 mg/ml РНК дрожжей, 100 mM DTT и концентрацию зонда 50000 cpm/μl. Предметные стекла были промыты один раз в 4X SSC, 10 mM DTT при комнатной температуре в течение 60 минут, один раз в 50% формамиде, 2X SSC, 10 mM DTT при 60oC в течение 40 минут и по разу в 2X SSC и IX SSC в течение 30 минут при комнатной температуре. Специфичность гибридизации была определена в параллельных экспериментах, выполненных по такому же протоколу, но включающих также более тщательную промывку в 50% формамиде, IX SSC, 10 mM DTT при 60oC в течение 40 минут. После промывки предметные стекла были погружены в эмульсию NTB2 (Kodak, Rochester, NY) и находились там от 2 до 21 дня перед развитием и контрастным окрашиванием. Средства отрицательного контроля включали сенсибилизирующие зонды, полученные из сенсибилизирующих рибозондов домена 1 ICAM-4 и доменов 3-4 ICAM-4, в дополнение к рибозонду вируса иммунодефицита человека (HIV-1).
Сигнал, определенный в ткани головного мозга, был первоначально локализирован в сером веществе при помощи наиболее сильного сигнала в коре головного мозга и гиппокампе. Профиль гибридизации был согласован с экспрессией ICAM-4 прежде всего в нервных клетках головного мозга.
ПРИМЕР 5. Получение слитых белков ICAM-4.
Слитые белки ICAM-4/глутатион-S-трансферазы (GST) были получены с использованием вектора pGEX экспрессии прокариота (Pharmacia, Alameda, CA), чтобы получить моноклональные антитела против специфических фрагментов полипептида ICAM-4.
Праймеры PCR, соответствующие 5' и 3' концам домена 1 и 5' и 3' концам домена 2, были использованы для амплификации фрагментов ДНК, кодирующих отдельные домены. Полученные фрагменты были раздельно клонированы в EcoRI-сайт pGEX-2T; анализ последовательности ДНК подтвердил правильную ориентацию и рамку считывания. Трансформанты были последовательно подвергнуты скринингу для выявления их способности продуцировать слитые белки соответствующей молекулярной массы.
Слитые белки ICAM-4 как домена 1/GST, так и домена 2/GST остались в нерастворимой фракции после того, как бактерии были разрушены ультразвуком в PBS (забуференный фосфатом физиологический раствор), содержащем 1% SDS (додецилсульфат натрия). Нерастворимая фракция белка из 100 ml культур была подвергнута кипячению в SDS, содержащем краситель, и выделена на 10-процентном SDS-полиакриламидном геле. Гель был окрашен в ледяном KCl в количестве 0,4 М, и бэнды слитого белка были удалены. Слитые белки были подвержены электроэлюции из срезов геля в диализной трубке в буфере, содержащем 25 мМ Tris-HCl и 192 мМ глицина. Соответствующая концентрация белка была определена посредством OD280, чистота препарата была определена на SDS-PAGE, окрашенном в синий цвет.
ПРИМЕР 6. Получение моноклональных антител против слитых белков крысиного ICAM-4/GST.
Мыши Balb/c были иммунизированы посредством подкожной инъекции 40-50 мкг домен-2/GST слитого белка ICAM-4 (описанного в примере 5), эмульгированного в полном адъюванте Фрейнда (FCA). Через две недели мыши были снова иммунизированы тем же белком, эмульгированным, однако, в неполном адъюванте Фрейнда. Две окончательные интраперитональные иммунизации, проведенные через две недели после второй иммунизации, включали растворимый антиген при отсутствии адъюванта. Сыворотка каждой иммунизированной мыши была проверена посредством ELISA (твердофазный иммуноферментный анализ) на предмет способности к специфической реакции с крысиным ICAM-4, полученным вышеописанным методом бакуловирусной экспрессии.
Селезенка из мыши #1654 была стерильно удалена и помещена в 10 ml RPMI 1640 без сыворотки. Одноклеточная суспензия была образована посредством измельчения ткани селезенки между матовыми концами двух предметных стекол, погруженных в RPMP 1640 без сыворотки (Gibco, Burlington, Ottawa, Canada) с добавлением 2 мМ L-глутамина, 1 мМ пирувата натрия, 100 ед./мл пенициллина и 100 мкг/мл стрептомицина. Клеточная суспензия была профильтрована через стерильный 70-ячеистый сетчатый клеточный фильтр Нитекса (Becton Dickinson, Parcippany, NJ) и дважды промыта при помощи RPMI, а затем подвержена центрифугированию при 200 x g в течение 5 минут. Полученный после окончательной промывки дебрис был ресуспендирован в 20 мл RPMI без сыворотки. Тимоциты, взятые от трех мышей Balb/c, были приготовлены аналогичным способом.
Перед слиянием клетки NS-1 миеломы культивировались в лог-фазе в RPMI с 11% феталклональной сыворотки (FBS) (Hyclone laboratories, Logan, Utah) в течение трех дней. После харвестирования клетки были центрифугированы при 200 х g в течение 5 минут, и дебрис был промыт дважды, как это описано выше. После промывки суспензия клеток была помещена в окончательный объем 10 мл в RPMI без сыворотки. Слой в 20 мкл этого раствора был удален, разбавлен 1:50 RPMI без сыворотки, и слой в 20 мкл этого раствора был удален, смешан с 20 мкл 0,4% трипанового синего красителя в 0,85% солевом растворе (Gibco), помещен на гемоцитометр (Baxter Healthcare, Deerfield, IL), и клетки были подсчитаны. Приблизительно 2,425•108 клеток селезенки были объединены с 4,85•107 клеток NS-1, смесь центрифугирована и супернатант удален. Полученный дебрис был извлечен посредством постукивания по пробирке и 2 мл 50-процентного PEG 1500 в 75 mM Hepes, pH 8,0 (Boehringer Mannheim, Indianapolis, IN), было добавлено при помешивании в течение 1 минуты. Затем было добавлено еще 14 мл RPMI без сыворотки в течение 7 минут. Суспензия клеток была центрифугирована при 200 х g в течение 10 минут и супернатант удален. Дебрис был ресуспендирован в 200 мл RPMI, содержащем 15% FBS, 100 мкM гипоксантина натрия, 0,4 мкM аминоптерина, 16 мкM тимидина (HAT) (Gibco), 25 ед./мл IL-6 (Boehringer Mannheim) и 1,5•106 тимоцитов/мл. Суспензия была сначала помещена в колбу 225 см2 (Corning, Essex, United Kingdom) при 37oC на четыре часа, а затем распределена в десять плоскодонных планшетов для культивирования тканей с 96 лунками (Corning) по 200 мкл в лунку. Клетки в планшетах подпитывали на 3-й, 4-й, 5-й и 6-й день после слияния путем аспирации приблизительно 100 мкл из каждой лунки иглой 20 G (Becton Dickinson) и добавления в каждую лунку 100 мкл вышеописанной планшетной среды с той разницей, что она содержала 10 ед./мл IL-6 и была лишена тимоцитов.
Планшеты были подвергнуты скринингу первоначально методом ELISA с захватом антигена, как описано ниже. Планшеты с Immulon 4 (Dynatech, Cambridge, MA) были покрыты на ночь при 4oC, по 100 ng на каждую лунку, слитым белком домена 1-GST или домена 2-GST в 50 mM карбонатного буфера. Планшеты были блокированы, по 100 мкл на каждую лунку, 0,5-процентным желатином из рыбьей кожи (Sigma, St. Louis, MO) в PBS в течение 30 минут при 37oC. После блокирования планшеты были промыты 3X при помощи PBS, содержащего 0,05% Tween 20 (PBST), и было добавлено 50 мкл в лунку супернатанта гибридомы из каждого слияния. После инкубации при 37oC в течение 30 минут планшеты были промыты, как описано выше, и было добавлено 50 мкл раствора 1:3500 конъюгированного пероксидазой козьего антимышиного IgG (Fc) (Jackson Immuno-Research, West Grove, Pennsylvania). Планшеты были снова подвергнуты инкубации в течение 30 минут и промыты 4X при помощи PBST. Был добавлен субстрат, по 100 мкл в лунку, состоящий из 1 мг/мл о-фенилендиамина (Sigma) и 0,1 мкл/мл 30% H2O2 в 100 mM цитрата, pH 4,.5. Цветная реакция проходила в течение 10 минут и была прекращена путем добавления по 50 мкл в лунку 15% H2SO4. Затем оптическая плотность при 490 nm была определена на автоматизированном планшет-ридере (Dynatech).
Лунки, которые были положительными для белка домена 2-GST, но не для белка домена 1-GST, были затем подвергнуты скринингу посредством ELISA против бакуловирусного супернатанта (описано ниже). ELISA проводился, как это описано выше, за исключением того, что планшеты с Immulon 4 были первоначально покрыты на ночь бакуловирусным супернатантом, растворенным в соотношении 1:4 в 50 mM карбонатного буфера. Три лунки (103A, 103B и 103F) были клонированы от двух до трех раз последовательно путем повторного разбавления в RPMI, 15% FBS, 100 мкM гипоксантина натрия, 16 мкM тимидина и 10 ед./мл IL-6. Лунки планшетов с клонами были визуально подсчитаны через 4 дня, и было записано число колоний в лунках с наименьшей плотностью. Выбранные лунки каждого клонирования были вновь подвергнуты анализу посредством ELISA после 7-10 дней против либо белка домена 1-GST и белка домена 2-GST, либо бакуловирусного супернатанта.
Моноклональные антитела, продуцированные гибридомами, были изотипизированы посредством ELISA. Планшеты с Immulon 4 (Dynatech) были покрыты при 4oC, по 50 мкл на каждую лунку, козьим антимышиным IgA, IgG или IgM (Organon Teknika, Durham, NC), растворенным 1:5000 в 50 mM карбонатного буфера, pH 9,6. Лунки были блокированы на 30 минут при 37oC посредством 1% BSA в PBS, промыты 3X посредством PBST. Раствор 1:10 супернатанта культуры гибридомы (50 мкл) был добавлен к каждому планшету, подвергнут инкубации и промыт, как описано выше. После последней промывки было добавлено 50 мкл конъюгированного пероксидазой кроличьего антимышиного IgG1, G2a, G2b или G3 (Zymed, San Francisco, CA) (разбавленного 1:1000 в PBST с 1% нормальной козьей сывороткой. Планшеты были инкубированы, как описано выше, промыты 4X посредством PBST и было добавлено 100 мкл субстрата. Цветная реакция была прекращена через пять минут путем добавления 50 мкл 15% H2SO4 и определена оптическая плотность при 490 nm на планшет-ридере (Dynatech).
Результаты показали, что антитела 103A, 103B и 103F были все изотипами IgG1. Эти антитела были впоследствии использованы при иммуноцитохимическом анализе, вестерн-блоттинге и для очистки белка, выраженного в бакуловирусе.
ПРИМЕР 7. Бакуловирусная экспрессия крысиного ICAM-4.
Метод бакуловирусной экспрессии (Invitrogen) был использован для получения растворимого белка, соответствующего доменам 1-6 ICAM-4. Поскольку лидерная последовательность ICAM-4 не была известна, конструкция экспрессии содержала кодирующую последовательность для ICAM-4, слитого 3' с лидерной последовательностью ICAM-1 в соответствующей рамке считывания. Далее изложены специфические подробности, касающиеся конструкции плазмиды экспрессии ICAM-1/ICAM-4.
ДНК крысиного IGAM-1, копирующая пять Ig-подобных доменов, была амплифицирована посредством PCR с использованием праймеров, которые включали несколько признаков для облегчения конструирования слитой плазмиды. 5' олигонуклеотидный праймер включал сайты HindIII и BglII в дополнение к консенсусной последовательности Козака в обратном направлении от первого метионина в лидерной последовательности. 3' олигонуклеотидный праймер включал кодирующую последовательность для шести гистидинов, сопровождаемых стоп-кодоном и клонирующим сайтом HindIII. Продукт амплификации PCR был клонирован в HindIII-гидролизированный вектор pES+, и анализ последовательности подтвердил соответствующую конструкцию. Внутренний сайт SmaI в лидерной последовательности ICAM-1 и другой сайт SmaI в области множественного клонирования вектора (от 3' до Ig-подобного домена 5 ICAM-1) были гидролизированы, что удалило большую часть кодирующей последовательности ICAM-1. После этих операций линеаризованный вектор с тупым концом содержал часть области множественного клонирования обратного направления (сайты рестрикции 5' первоначального сайта HindIII в области множественного клонирования), последовательность Козака и большую часть лидерной последовательности ICAM-1.
Кодирующая последовательность для доменов 1-6 крысиного ICAM-4 была амплифицирована посредством PCR с использованием праймеров, позволяющих кодировать эту последовательность в линеаризованный вектор, описанный выше. 5' олигонуклеотидный праймер включал сайт EcoRV и кодоны, необходимые для завершения лидерной последовательности ICAM-1. 3' олигонуклеотидный праймер включал кодоны для шести групп гистидина, стоп-кодон и сайты рестрикции HindIII и EcoRV. Продукт амплификации от этой PCR был гидролизирован EcoRV для получения последовательности с тупым концом, которая была затем лигирована в линеаризованный SmaI-гидролизированный вектор pBS+ с тупым концом. Вся последовательность, содержащая лидерную последовательность ICAM-1 5' до доменов 1-6 ICAM-4, была удалена из конструкции гидролизом BglII и HindIII, и последовательность слияния очищенного ICAM-1/ICAM-4 клонирована непосредственно в BelII/HindIII-гидролизированный вектор pBluesac III (Invitrogen).
Продуцирование белка рекомбинантным вирусом было проанализировано посредством ELISA, первоначально с использованием иммунных сывороток от мышей, иммунизированных белком слияния домена-2 крысиного ICAM-4 и GST, описанным в примере 5. В более поздней работе моноклональные антитела, полученные от этих мышей, были использованы для очистки белка ICAM-4, продуцированного рекомбинантными бакуловирусами в клетках SF9.
ПРИМЕР 8. Производство моноклональных антител против выраженного бакуловирусом крысиного ICAM-4.
Домены 1-6 крысиного ICAM-4 были выражены методом бакуловирусной экспрессии, как описано в примере 7. Рекомбинантный белок был очищен с использованием моноклонального антитела 103A (как описано в примере 6).
Излагая кратко, 30 мг очищенного моноклонального 103A (в 100 mM бората натрия, 500 mM хлорида натрия) были соединены с тремя граммами активированной цианогенной бромидной сефарозы 4B (Pharmacia, Piscataway, NJ). Бакуловирусный супернатант, содержащий рекомбинантный крысиный ICAM-4 (домены 1-6) был помещен на колонку с сефарозой на ночь при 4oC. Колонка была промыта в физиологическом растворе, забуференном фосфатом без магния и кальция (CMF-PBS), и связанный материал был элюирован в 50 mM лимонной кислоты, 500 mM NaCl, pH 4,0. Образец был нейтрализован посредством 1/10 объема Tris, pH 10, и хранился при -20oC. Очищенный белок, отделенный на SDS-PAGE, обладал чистотой более чем на 90% и мигрировал приблизительно при 80 kD.
Мыши были иммунизированы очищенным рекомбинантным белком доменов 1-6 крысиного ICAM-4 подобным образом, как описано в примере 6. Селезенка мыши # 1945 была использована для слияния #127. Процедура слияния была такой, как описано в примере 6. Лунки слияния были подвергнуты скринингу посредством ELISA на рекомбинантном белке ICAM-4. Повторный скрининг включал иммуноцитохимический анализ на срезах крысиного головного мозга (как описано ниже в примере 9). Четыре дополнительных антитела, специфических для крысиного ICAM-4, были клонированы из этого слияния: 127A, 127B, 127E и 127H. Иммуноцитохимическая картина окрашивания каждого антитела на срезах головного мозга крысы была такой же, что наблюдавшаяся с моноклональным антителом 103A (см. пример 9). Моноклональные антитела были исследованы на их способность связывать слитые белки D1/GST и D2/GST (описано в примере 5). Моноклональное антитело 127A распознало слитый белок D1/GST и 127H распознало слитый белок D2/GST. Эти две определенные связывающие специфичности, наряду с другими, которые не связывали ни один из белков GST, дают возможность предполагать, что, по крайней мере, 3 различных эпитопа распознавались панелью антител. Гибридомы 127A и 127H были депонированы 31 мая 1995 г. и 1 июня 1995 г. соответственно в Американской коллекции типовых культур, 12301 Parklawn Drive, Rockville, Maryland 20852, и им были присвоены каталожные номера HB11905 и HB11911 соответственно.
ПРИМЕР 9. Иммуноцитохимический анализ экспрессии крысиного ICAM-4.
Иммуноцитохимический анализ при помощи моноклонального антитела 103A был проведен для локализации продуцирования белка в головном мозге крысы.
Был взят головной мозг у здоровой взрослой женской особи мыши Льюиса, сагиттально рассечен и промыт в 1X PBS без RNase на льду в течение 30 минут. Срезы мозга были затем помещены в тканевые криоформы Tek II (Miles Laboratories, Inc., Naperville, IL) с небольшим количеством состава ОСТ (Miles, Inc. , Elkhart, IN). Мозг был размещен в центре криоформы, которая была затем заполнена составом ОСТ и помещена в контейнер с 2-метилбутаном (Aldruich Chemical Company, Inc., Milwaukee, WI), который был помещен в жидкий азот. Когда ткань и состав ОСТ в криоформе были заморожены, блоки хранились при -80oC до того, как потребовалось делать срезы.
Были сделаны срезы толщиной 6 мкм, прикреплены на предметных стеклах с пленкой Vectabond (Vector Laboratories, Inc., Burlingame, CA) и оставлены сушиться при комнатной температуре на ночь перед использованием. Срезы были зафиксированы в простом этиле в течение 5 минут при комнатной температуре. Когда срезы были удалены из этила, реагенту дали возможность испариться. Каждый срез ткани был блокирован при помощи 150 мкл 50% сыворотки здоровой крысы (Sigma) и 2% альбумина бычьей сыворотки (BSA) (Sigma) в 1X PBS (приготовленном только с фосфатами натрия) в течение 30 минут при комнатной температуре. После блокирования раствор был осторожно удален из срезов промоканием, и очищенное антитело 103A (1,65 мг/мл) было разбавлено 1:10 в блокирующем растворе и 150 мкл внесено в каждый срез ткани. Предметные стекла были помещены во влажную камеру и инкубированы при 4oC в течение ночи.
На следующий день раствор антитела был осторожно удален промоканием из среза, и предметные стекла промыты три раза в 1X PBS, каждый раз по четыре минуты. Излишек PBS был удален со стекла, и 100 мкл вторичного, крысиного, антимышиного, биотинилированного антитела (Jackson Immuno-Research Laboratories), разбавленного 1:100 в растворе 10% сыворотки здоровой мыши и 2% BSA в 1X PBS, было внесено в ткани. Инкубирование проходило в течение одного часа при комнатной температуре. Срезы были промыты два раза в 1X PBS, каждый раз по четыре минуты, затем 100 мкл реагента ABC из набора Vectastain ABC иммуноглобулина G элитной крысы (Vector Laboratories, Inc., Burlingame, CA), приготовленного в соответствии с вставкой продукта, было внесено в каждый срез. Инкубирование проходило в течение 30 минут при комнатной температуре. После инкубирования срезы были промыты два раза в 1X PBS (каждый раз по четыре минуты), и 150 мкл раствора субстрата пероксидазы вектора VIP (Vector Laboratories, Inc. , Burlingame, CA) было внесено в каждый срез приблизительно на 10 минут. После проявления цвета срезы были промыты в проточной водопроводной воде в течение пяти минут, подвергнуты контрастному окрашиванию гематоксилином Майера (Sigma) в течение 20 секунд и вновь промыты под слабой струей водопроводной воды в течение пяти минут. Предметные стекла были обезвожены посредством ряда этанолов, пропущены через ксилол и установлены при помощи Accumount 60 (Stephens Scientific, Riverdale, NJ).
Иммуногистохимический анализ срезов головного мозга крысы, окрашенных mAb 103A, показал, что крысиный ICAM-4 выражен в нейронных клетках гиппокампа. Картина окрашивания дала основания предполагать, что белок мог быть ограничен нейронными образованиями (дендритами). Срезы головного мозга, окрашенные подобным образом при помощи нерелевантного антитела или одного реагента на второй стадии, не дают четкой картины экспрессии, полученной с MAb 103A.
ПРИМЕР 10. Клонирование геномной ДНК человеческого ICAM-4.
При клонировании крысиного ICAM-4 из геномной ДНК было обнаружено, что ICAM-4 и ICAM-1 были расположены в пределах 5 kb друг от друга, и эта информация была использована при попытке клонирования человеческого гомолога ICAM-4.
В Genom Systems Inc. (St. Louis, MO) амплифицировали фрагменты в библиотеке человеческого P1 посредством PCR с использованием праймеров домена 3 человеческого ICAM-1, сенсибилизирующего праймера, комплементарного домену 3 человеческого ICAM-1 (H-1/D3 S) и десенсибилизирующего праймера, комплементарного домену 3 человеческого ICAM-1 (H-1/D3 AS). Эти праймеры представлены в SEQ ID N 22 и 23 соответственно.
CCGGGTCCTAGAGGTGGACACGCA (SEQ ID N 22)
TGCAGTGTCTCCTGGCTCTGGTTC (SEQ ID N 23)
Два клона, обозначенные 1566 и 1567, были идентифицированы и подвергнуты дальнейшему анализу. Оба клона P1 содержали приблизительно 75-95 kb вставок геномной ДНК. Клоны были гидролизированы BamH1, отделены электрофорезом агарозного геля и блотированы на найлоновые мембраны. Саузерн-блоттинг был выполнен либо с низкой степенью точности (30% формамид), либо с высокой степенью точности (60% формамид) при 42oC с меченными радиоактивными изотопами зондами человеческого ICAM-1, ICAM-3 или крысиного ICAM-4; другие компоненты раствора гибридизации были теми же, что описаны в примере 1. При гибридизации с низкой степенью точности была осуществлена промывка при комнатной температуре в 2X SSPE, содержащем 0,1% SDS. При гибридизации с высокой степенью точности была осуществлена промывка при 65oC в 0,2X SSPE, содержащем 0,1% SDS. Промытые мембраны были подвергнуты воздействию рентгеновской пленки в течение 3,5 часов.
Различные виды гибридизации показали, что человеческий ICAM-1 содержался на фрагменте BamH1 в 5,5 kb, тогда как человеческий ICAM-3 был расположен на фрагментах BamH1 в 4,0 kb и 1,5 kb. Фрагменты человеческого ICAM-1 и ICAM-R были субклонированы в pBS+, и их идентичность подтвердилась при анализе ограниченной последовательности.
Фрагмент BamH1 в 7,0 kb, который гибридизировал с крысиным ICAM-4 в условиях высокой степени точности, был субклонирован и подвержен дальнейшему фрагментированию гидролизом рестрикции RsaI. Три фрагмента RsaI, которые гибридизировали с крысиным ICAM-4, были идентифицированы, и их последовательности определены. Основанные на гомологии крысиному ICAM-4 эти фрагменты содержали домены 2, 3, 4, 5 и часть домена 6.
ПРИМЕР 11. Клонирование кДНК человеческого ICAM-4.
Фрагменты геномной ДНК, соответствующие доменам 2-5 человеческого ICAM-4 (описанного в примере 10), были использованы в качестве зондов для скрининга библиотеки кДНК человеческого гиппокампа μgt10 (Clontech, Palo Alto, CA). Процедура скрининга библиотеки была, в основном, такой же, как описано в примере 1.
Самый длинный клон человеческого ICAM-4 (#18), который был обнаружен в этой библиотеке, имел только 992 bp (SEQ ID N 24) и соответствовал примерно середине прогнозированного гена в 3 kb. ДНК-вставка из 992 bp из клона 18 (SEQ ID N 24) была использована в качестве зонда для скрининга библиотеки кДНК человеческого гиппокампа ZAPII (Stratagene, La Jolla, CA). В этой библиотеке было обнаружено несколько положительных клонов. Самый длинный клон, #34, имел 2775 bp (SEQ ID N 25). На основе сравнительного анализа полномерного крысиного ICAM-4 предположили, что и в этом клоне были пропущены лидерная последовательность и приблизительно 30 bp на 5' конце домена 1. Поли-A+ хвост на 3' конце был пропущен, но стоп-кодон трансляции присутствовал.
Фрагмент ДНК, соответствующий первым трем доменам (нуклеотиды 1-840 в клоне # 34), был использован в качестве зонда для скрининга библиотеки кДНКλgt10, извлеченной из коры головного мозга человека (Clontech, Palo Alto, CA). Один клон, 16-1 (SEQ ID N 26) был идентифицирован как имеющий 1557 bp и включал 39 bp 5' нетранслированной ДНК, лидерную последовательность и информацию последовательности до пятого домена. Перекрывающие клоны # 34 (SEQ ID N 25) и 16-1 (SEQ ID N 26) были использованы для получения композита полномерной последовательности человеческого ICAM-4 (SEQ ID N 27).
Полномерный ген имеет длину 2927 bp и кодирует белок аминокислоты 924. Нуклеотидная последовательность ICAM-4 представлена в SEQ ID N 27, и аминокислотная последовательность представлена в SEQ ID N 28. Сравнительный анализ последовательности с полномерным геном крысиного ICAM-4 (SEQ ID N 11) показал 82% идентичность всей последовательности ДНК и 85% идентичность на аминокислотном уровне. Явная структура белка с 9 Ig-подобными внеклеточными доменами является консервативной между крысиной и человеческой. Лидерная последовательность простирается от аминокислоты 1 до 28; домен 1 - от аминокислоты 29 до 117; домен 2 - от аминокислоты 118 до 224; домен 3 - от аминокислоты 225 до 320; домен 4 - от аминокислоты 321 до 405; домен 5 - от аминокислоты 406 до 488; домен 6 - от аминокислоты 489 до 570; домен 7 - от аминокислоты 571 до 663; домен 8 - от аминокислоты 664 до 743; домен 9 от аминокислоты 744 до 837; трансмембранная область - от аминокислоты 838 до 857 и цитоплазматический концевой сегмент - от аминокислоты 858 до 924.
Человеческий ICAM-4 (HuICAM-4), в дополнение к тому, что он генетически связан с ICAM-1 и ICAM-R, показал также некоторые общие структурные признаки, позволяющие группировать их вместе как семейство молекул. Сравнительный анализ каждого из доменов HuICAM-4 с другими членами семейства ICAM показал различные степени гомологии. Аминокислотная последовательность HuICAM-4 домена 1 идентична последовательностям ICAM 1,2 и 3 домена 1 соответственно на 21, 30 и 26%. Домен 2 человеческого ICAM-4 идентичен ICAM 1, 2 и 3 соответственно на 61, 39 и 62%. Домен 3 человеческого ICAM-4 идентичен ICAM 1 и 3 соответственно на 50 и 56%. Домен 4 человеческого ICAM-4 идентичен ICAM 1 и 3 соответственно на 54 и 64%. Домены 5-8 человеческого ICAM-4 являются более гомологичными пятым доменам ICAM-1 и 3, и их идентичность находится в диапазоне 33-37% для домена 5 с ICAM-1 и 21-31% для домена 5 с ICAM-R. Девятый домен агентов HuICAM-4 мало подлежит сравнительному анализу с другими членами семейства ICAM, но является гомологичным доменам 3 (идентичен на 24%) и 6 (идентичен на 23%) человеческого ICAM-1.
ПРИМЕР 12. Назерн-анализ экспрессии человеческого ICAM-4.
Два блота Назерна множества тканей человека (MTN) были получены из Clontech (Palo Alto, CA). Они содержали, по крайней мере, 2 g поли A+-ДНК из 16 различных тканей человека (как показано в табл. 1), нанесенной на денатурирующий формальдегидный 1,2% агарозный гель и перенесенной на нейлоновую мембрану. Блоты были предварительно гибридизированы в течение трех часов при 42oC в 10 мл раствора, содержащего 5X SSPE, 10X раствор Denhardts, 50% формамид, 2% SDS и 100 мкг/мл денатурированной ДНК спермы лосося. Блоты были гибридизированы в указанном растворе с меченным радиоактивным изотопом зондом человеческого ICAM-4 (клон #18, SEQ ID N 24) в течение 16 часов при 42oC. На следующий день блоты были промыты в растворе 0,1X SSC/0,1% SDS при комнатной температуре, а затем при температуре 50oC. Блоты были подвергнуты воздействию рентгеновской пленки при -80oC в течение 24 часов. Результаты анализов даны ниже в табл. 1.
Только полоса, содержащая ДНК из головного мозга, гибридизировала к зонду ICAM-4 и давала при этом единственный бэнд приблизительно в 3 kb. Более длительное экспонирование (пять дней) подтвердило, что только головной мозг имел определяемый уровень переноса. Чтобы определить, содержали ли все полосы сравнимые количества ДНК сравнимого качества, тот же блот был гибридизирован с контрольным β-актиновым зондом. Блоты были удалены с зонда ICAM-4 посредством обработки кипящим раствором 0,1% SDS в течение 15 минут и затем подвергнуты зондированию подобным способом при помощи β-актинового зонда, предоставленного производителем. За исключением небольшой разницы в количествах во всех полосах была обнаружена РНК хорошего качества.
Из Clontech были получены два дополнительных назерн-блота, которые содержали поли-A+ РНК из 16 различных субрайонов головного мозга человека (как показано в табл. 2). Блоты были зондированы способом, подобным тому, что был использован при анализе тканей, и результаты представлены в табл. 2. Качество и количество введенной РНК было проверено зондированием блотов β- актиновым зондом.
Все области, которые показали экспрессию ICAM-4, являются частью конечного мозга, за исключением таламуса, который считается частью промежуточного мозга. Гиппокамп и кора головного мозга имели наивысший уровень экспрессии. Величина транскрипта во всех случаях была одинаковой, 3 kb. Совершенное тканевое распределение экспрессии ICAM-4 предполагает, что область промотора может содержать элементы, которые подтверждают наблюдавшуюся эволюционную и пространственную экспрессию продукта генов. Использование такой информации может дать понимание управления экспрессией нейронных генов.
ПРИМЕР 13. Образование слитых белков человеческого ICAM-4/IgG.
Плазмиды экспрессии слитых белков ICAM-4/IgG были сконструированы для продуцирования белков для получения моноклональных антител и для использования при адгезионном анализе для идентификации потенциальных лигандов ICAM-4. Были сделаны две конструкции: первая включала ДНК, кодирующую домены 1-3 человеческого ICAM-4, а вторая - домены 4-8. Обе были соединены с Fc областью человеческого IgG1 в векторе pDCS1, который использует цитомегаловирусный (CMV) промотор, чтобы дать возможность экспрессии и сигнальной последовательности из IgG4 облегчить секрецию молекул.
Праймеры PCR (показаны ниже как SEQ ID N 29-32) были сконструированы для получения необходимых фрагментов ДНК для субклонирования. "Сенсибилизирующий" праймер для 5' конца домена 1 (H14-D1(s), SEQ ID N: 29) был сконструирован для внесения 30 основных пар домена 1, пропущенных в клоне # 34. Праймеры H14-D1(S) (SEQ ID N 29) и H14-D(AS) (SEQ ID N 30) были использованы для получения фрагмента ДНК, кодирующего домены 1-3 человеческого ICAM-4, соответствующие области в SEQ ID N: 1 от нуклеотида 130 до нуклеотида 996. Праймеры H14-D3(S) (SEQ ID N 31) и H14-D8(AS) (SEQ ID N 32) были использованы для получения фрагмента ДНК, кодирующего домены 4-8 человеческого ICAM-4, соответствующие области в SEQ ID N 30 от нуклеотида 997 до нуклеотида 2268. Каждый 5' праймер кодировал сайт рестрикции BamH1 (GGATCC, выделенный жирным шрифтом в приведенной ниже последовательности), и каждый 3' (десенсибилизирующий) праймер содержал сайт XhoI (CTCGAG, выделенный жирным шрифтом в приведенной ниже последовательности) для облегчения субклонирования 5' к гену IgG1. Все олигонуклеотиды содержат спейсерные нуклеотиды (подчеркнуты в приведенной ниже последовательности) и 5' конец для обеспечения гидролиза рестрикции.
H14-D1(S) (SEQ ID N 29)
GTACTTACAGGATCCGCGGTCTCGCAGGCCCTTCTGGGCGGACCTACAGCCTGCGTGGCGTTC
H14-D3(AS) (SEQ ID N 30)
ATTTCTCTCGAGGATGGTCACGTTCTCCCGG
H14-D4(S) (SEQ ID N 31)
ATTTCTGGATCCTACAGCTTCCCGGCACCACTC
H14-D8(AS) (SEQ ID N 32)
ATTTCTCTCGAGTTCCACGCCCACAGTGACGG
Реакции PCR проводились в объеме 50 мкл с использованием буферов, предоставленных Perkin Elmer с энзимом AmpliTaq. Праймеры были добавлены к окончательной концентрации 10 мкг/мл, и все четыре dNTP были включены при 2 mM. Реакции продолжались на протяжении 30 циклов денатурирования (94oC в течение 4 минут), отжига (50oC в течение 2 минут) и удлинения (72oC в течение одной минуты). Продукты PCR были видимыми на агарозном геле, и слой каждой реакции был использован для субклонирования продуктов PCR в вектор pCRII (Invitrogrn, SanDlego, CA). Был проведен анализ последовательности, чтобы определить возможные погрешности в результате процесса амплификации и подтвердить нужную ориентацию. Соответствующие клоны были гидролизированы с BamHI и XhoI, и фрагменты выделены посредством электрофореза агарозного геля. Очищенные фрагменты были лигитированы в вектор pDCSI, предварительно гидролизированный с BamHI и XhoI, и полученные в результате плазмиды были секвенированы для подтверждения должной ориентации и рамки считывания.
Белки слияния доменов 1-3 и 4-8 человеческого домена и IgG1 были получены после переходной трансфекции плазмидов экспрессии в клетки COS7 и выделения секретированного белка из культурной среды. Трансфекция проводилась следующим образом. Адгезивные клетки COS7 при приблизительно 50-60% слиянии были промыты при помощи CMF-PBS и затем соединены с 10-15 мкг плазмидной ДНК в 7,5 мл не содержащей сыворотки среды MDEM (Gibco, Gaithersburg, MD), включающей 6 мкл 0,25 M хлорохина (Sigma, St. Louis, MO). Было добавлено дополнительно 7,5 мл не содержащей сыворотки среды, включающей 150 мкл DEAE декстрана (50 мг/мл) (Sigma, St. Louis, MO), и планшеты инкубировали 2-3 часа до того, как среда была удалена и замещена на 10% DMSO (Mallinckrodt, McGaw Park, Illinois) в PBS. После одноминутной инкубации раствор DMSO был удален и замещен свежей средой, содержащей 5% FBS. Каждая трансфекция включала множество планшетов, и среды из клеток, выражающих один и тот же белок, были объединены для выделения белка.
Среды собирались каждые три дня на протяжении курса из 3-4 сборов. Белки были очищены с использованием 0,4-0,8 мл Procep A колонки (Bioprocessing Ltd, England), предварительно уравновешенной 35 mM Tris, 150 mM NaCl, pH 7,5. Культурная среда помещалась на колонку два раза при низкой скорости, менее 60 объемов колонки в час. Колонка была промыта один раз при помощи 20 объемов колонки буфера Tris/NaCl, 20 объемов 0,55 М диэтаноламина, pH 8,5, и 20 объемов колонки 50 mМ лимонной кислоты, pH 5,0. Слитые белки были элюированы в 1 мл фракции с использованием 50 mM лимонной кислоты, pH 3,0, и каждая фракция была нейтрализована посредством 1/10 объема 1 М Tris, pH 9,5. Концентрация белков была определена посредством OD280, а чистота была определена при использовании SDS-PAGE.
Было отмечено значительное загрязнение из бычьего IgG (присутствующего в FBS). Несмотря на то, что было прогнозировано меньшее количество слитого белка доменов 1-3, чем доменов 4-8, оба мигрировали при приблизительно 90 kD. Одно возможное объяснение этого наблюдения заключается в том, что меньшее количество слитого белка доменов 1-3 могло быть более гликозилировано, чем большее количество слитого белка доменов 4-8.
В дополнение к использованию очищенных белков для получения моноклональных антител, как описано выше, белки могут быть также использованы при адгезивном анализе для идентификации лигандов ICAM-4.
ПРИМЕР 14. Получение моноклональных антител.
Очищенный белок, описанный в примере 13, был использован для получения моноклональных антител с использованием процедуры иммунизации, описанной в примере 6.
Селезенка мыши #2250 (иммунизированной HuICAM-4 D1-3/IgG1) была использована для слияния 172 и селезенка мыши #2272 (иммунизированной HuICAM-4 D4-8/IgG1) была использована для слияния 173. Была использована процедура слияния, описанная в примере 6. Планшеты слияний были подвергнуты скринингу посредством ELISA (в основном, как описано в примере 6), с использованием каждого слитого белка HuICAM-4/IgG1. Супернатанты лунок слияния, которые распознали иммуногенный белок, и никакие другие, принимались во внимание для клонирования. Иммуноцитохимический анализ на срезах гиппокампа человека был использован как повторный скрининг.
Один первоначальный клон из каждого слияния, был положительным при иммуноцитохимическом анализе и был клонирован. Один из двух клонов не смог развиться при клонировании, оставив только одного кандидата для достижения цели, клон 173E, который был извлечен из мыши, иммунизированной HuICAМ-4 D4-8/IgG1. Гибридома 173E была депонирована 1 июня 1995 г. Американской коллекцией типовых культур, 12301 Parklawn Drive, Rockville, Maryland 20852, и ей был присвоен каталожный номер HB11912.
В другом слиянии, полученном из мыши, иммунизированной растворимым фрагментом ICAM-4, соответствующим доменам 1-3, были обнаружены шесть клонов (179A, 179B, 179D, 179H, 179I и 179K), специфических для доменов от 1 до 3 (D1-3) человеческого ICAM-4. Все шесть антител в серии 179 связаны с дендритными процессами в зубчатой извилине, а также полиморфическими и пирамидальными клеточными слоями. Моноклональное антитело 179A окрасило нейронные клеточные тела из этих областей в дополнение к дендритным процессам.
Дополнительные слияния выполняются подобным образом, чтобы получить другие антитела, специфически иммунореактивные с определенными областями ICAM-4.
ПРИМЕР 15. Проведение анализа методом улавливания.
Шесть моноклональных антител из слияния 179 были исследованы в различных сочетаниях на их способность улавливать и выявлять растворимый ICAM-4 в растворах. Был проведен такой же анализ, как описанный выше, чтобы определить уровни растворимого ICAM-4 в жидкостях человека в здоровом и больном состояниях.
Покрытие из антител 1791 было нанесено на 96 лунок планшетов с Immulon 4 (Dynatech) при 3 мкг/мл, 125 мкл в каждой лунке, на два часа при 37oC. Раствор антител был удален отсасыванием, и лунки были блокированы на 30 минут при комнатной температуре посредством 300 мкл блокирующего раствора, содержащего 5% желатин Teleostean в несодержащем кальция и магния PBS (CMF-PBS). Блокирующий раствор был удален отсасыванием, 100 мкл жидкости образца, разведенной в Omni-разбавителе (CMF-PBS, 1% желатин и 0,05% Tween 20), было добавлено в каждую лунку, и смесь инкубирована при 37oC в течение 30 минут. Планшеты были промыты три раза при помощи PBST (CMF-PBS, 0,05% Tween 20). Антитело 179H было биотинилировано при 1,5 мг/мл с использованием NHS-LC-Биотина (Pierce) в соответствии с протоколом предполагаемого производителя, разведено в соотношении 1:2000 и добавлено в лунки (100 мкл в лунку). Полученная смесь была инкубирована в течение 30 минут при 37oC, и планшеты промыты три раза в PBST. Стрептавидин-HRP (Pierce) был добавлен (100 мкл, 0,25 g/ml) в каждую лунку, и эта смесь инкубирована при 37oC в течение 30 минут. Планшеты были промыты четыре раза в PBST перед добавлением 100 мкл тетраметилбензидина (Sigma) (биомасса 10 мг/мл в DMSO), разбавленного 1:100 в забуференном субстрате (13,6 g/L тригидрат ацетата натрия, pH до 5,5, с 1 М лимонной кислоты, с 150 мкл/L 30% перекиси водорода, добавленной непосредственно перед развитием). Реакция развивалась в течение 30 минут при комнатной температуре в темноте, после чего реакция была остановлена путем добавления 15% H2SO4 по 50 мкл в каждую лунку. Оптическая плотность прочитывалась при 450 nm.
Результаты показали, что при помощи анализа можно определить рекомбинантный белок D1-3 растворимого HuICAM-4 при такой низкой концентрации, как 5-10 ng/ml, с линейной частью кривой в диапазоне 10-100 ng/ml. Не наблюдалось никакой перекрестной реактивности с HuICAM-4 D4-8, когда эта область белка была исследована при 1 и 10 мкг/мл.
ПРИМЕР 16. Оценка растворимого ICAM-4 в сыворотке больных припадками.
Для оценки роли ICAM-4 при неврологических болезнях и состояниях сыворотку двадцати восьми больных, страдающих тяжелыми припадками, и двадцати восьми молодых здоровых добровольцев (не подобранных соответственно по возрасту) подвергали анализу таким образом, как это описано выше, для выявления разницы в концентрации растворимого ICAM-4.
Результаты показали, что сыворотка здоровых людей не содержала определяемого уровня ICAM-4. У двадцати из двадцати восьми больных были выявлены определяемые уровни растворимого ICAM-4. Сигнал от таких больных соответствовал диапазону 5-38 ng/ml норматива (рекомбинантный белок доменов 1-3 растворимого ICAM-4).
ПРИМЕР 17. Уровни мРНК с ICAM-4 в гиппокампе в крысиной модели эпилепсии.
Уровни выраженной мРНК с ICAM-4 были исследованы в гиппокампе крыс, которых обрабатывали таким образом, чтобы создать животную модель активизируемого эпилептогенезиса [Lothman et al., Brain Res., 360: 83-91 (1985)]. В этой модели гиппокамп крысы стимулировали серией субконвульсивных электрических ударов посредством электрода, имплантированного в области головного мозга, что постепенно приводило к сильным эпилептическим припадкам. Процесс активизации включает в себя двадцать стимуляций в день, проводимых через день на протяжении восьми дней. При полной активизации единственный раздражитель может вызывать эпилептические припадки и гистологические изменения, подобные тем, которые имеют место при эпилепсии у человека. Полностью активизированные крысы подвергались двум стимуляциям в день на протяжении двух недель, и после последней стимуляции животных умертвили. Гиппокамп был удален и рассечен для приготовления РНК.
Полная РНК была приготовлена из каждого образца при использовании метода экстрагирования гуанидин/фенол/хлороформа [Chomezynski and Sacchi, Anal. Biochem. , 162: 156-159 (1987)]. РНК была отделена на денатурирующем формальдегидном агарозном геле, перенесена на найлоновые мембраны и гибридизирована с меченным радиоактивными изотопами зондом крысиной ICAM-4 и зондом глицеральдегид-3-фосфатдегидрогеназной (GAPDH) специфичной ДНК. GAPDH является основным выраженным геном, который общеизвестен как контроль для выявления изменений от полосы к полосе в количестве РНК, помещенной на гель. Колебания в соотношении ICAM-4/GAPDH рассматривались как изменения в уровне экспрессии ICAM-4. Гибридизирующие бэнды для ICAM-4 и GAPDH были подсчитаны при помощи фосфорного формирователя изображения, и определено соотношение ICAM-4/GAPDH.
Соотношение ICAM-4/GAPDH было значительно выше у контрольных животных, которые не подвергались стимуляции (n=5), по сравнению с группой животных, прошедших испытание (n=5), что позволяет предположить, что ICAM-4 снижался вследствие процесса активизации. Нужно, однако, отметить, что контрольная группа не подвергалась никакому стимулирующему воздействию, поэтому существует такая вероятность, что уровни мРНК с ICAM-4 были модулированы в ответ на хирургическое вмешательство в соединении с воздействием током.
ПРИМЕР 18. Клонирование и анализ регуляторной ДНК обратного направления с человеческим ICAM-4.
Экспрессия ICAM-4-гена является регулируемой в пространстве и времени, причем экспрессия ограничена самой передней или вентральной областью головного мозга, конечным мозгом. При попытке идентифицировать последовательности гена, ответственные за ограниченную транскриптивную регуляцию ICAM-4, нуклеотидная область от 5' до кодирующих последовательностей человеческого ICAM-4 была исследована.
Фрагмент BamHI/PstI основной пары 2607, извлеченный из фрагмента геномного BamHI в 7,0 kb (описан в примере 10), был секвенирован и обнаружил содержание 1684 нуклеотидов в обратном направлении от старт-кодона ATG. Полная последовательность для этой области обратного направления представлена в SEQ ID N 33. Что касается положения кодирующей области ICAM-4, "A" в старт-кодоне ATG (пронумеровано в SEQ ID N 33 как нуклеотиды 1685-1687) обозначен +1 нуклеотид, а нуклеотид непосредственно 5' к A+1 нуклеотиду обозначен -1. Таким образом, вся последовательность показана простирающейся от нуклеотида -1684 до нуклеотида +3, соответствующих нумерации в Перечне последовательностей от нуклеотида 1 до нуклеотида 1687.
На основе геномной последовательности HuICAM-4 олигонуклеотиды были синтезированы и использованы в PCR для получения молекул ДНК различной длины в пределах регуляторной области обратного направления. Каждый олигонуклеотид, представленный в табл. 3, содержал спейсерную область (выделено курсивом), приблизительно 6-10 bp, чтобы обеспечить ферментативный гидролиз продукта PCR, сайт рестрикции NheI или HindIII (выделено жирным шрифтом) и последовательность праймера специфичной гибридизации (подчеркнуто). Названия олигонуклеотидов содержат номера, которые обозначают их расположение в пределах регуляторной области обратного направления. В амплификациях PCR олигонуклеотиды были спарены, как показано в табл. 4, для получения фрагментов ДНК, содержащих специфичные области регуляторной области обратного направления.
Сайты рестрикции и спейсерная область, образованная в каждом олигонуклеотиде, обеспечили ферментативный гидролиз и последующее прямое клонирование продуктов отдельной PCR в Основной Вектор pGL3 (Promega, Madison, WI), который содержит ген регистратора люциферазы непосредственно в прямом направлении сайта множественного клонирования (MCS). Промоторная активность, клонированная в область MCS вектора, приводит в действие экспрессию гена регистратора люциферазы в трансфецированных клеточных линиях, и легкая продукция из выраженной люциферазы может быть определена как индикатор промоторной активности. Основной вектор pGL3 не имеет промотора и поэтому служит в качестве отрицательного контроля, тогда как вектор pGL3, содержащий промотор SV40, служит в качестве положительного контроля. Последовательность конструкции каждой экспрессии была проверена посредством рестрикционного анализа и секвенирования ДНК.
Плазмиды, содержащие каждую из амплифицированных последовательностей, описанных в табл. 4, были трансфецированы в клетки млекопитающего при использовании Набора для трансфекции млекопитающего при трансфекции MBS (Stratagene, La Jolla, CA) в соответствии с предложенной производителем схемой. Каждая плазмида была введена в две разные клеточные линии, клетки предшественников COS7 и NT2 (Ntera2/D1 из Stratagene). Клетки COS7 представляют собой обычно используемую линию фибробластподобных клеток обезьяны, трансформированную посредством SV40, что сделало их пригодными для приведения в действие экспрессии гена при контроле промотора SV40 в клетках, трансфецированных посредством промоторного вектора pGL3 положительного контроля. Клетки предшественника NT2 представляют собой клеточную линию коммитированных нейронных клеток-предшественников, и хотя они не выражают ICAM-4, они могут быть более характерными для типа клеток, который выражает ICAM-4.
Каждая лунка 6-луночного планшета культуры ткани с плоским дном (Falcon) была засеяна 2,5•105 клетками. Трансфекции клеток COS7 и NT2 были осуществлены рядом в двух экземплярах с использованием 5 мкг ДНК плазмиды для каждой лунки. Клетки были культивированы при 37oC в течение 48 часов, лизированы и исследованы на люциферазную активность посредством Метода люциферазного анализа (Promega).
Результаты эксперимента, обобщенные в табл. 5, показывают высокий уровень промоторной активности в областях с -408 по -19 и с -480 по -19 регуляторной области обратного направления ICAM-4 в клетках NT2. Так как клетки NT2 нейронного происхождения, они могут выражать определенные факторы транскрипции, распознающие промотор ICAM-4, которые не обнаружены в других типах клеток. Наивысший уровень промоторной активности в трансфектантах клеток COS был достигнут с плазмидой, содержащей нуклеотиды с -560 до -19. Так как Промоторный вектор pGL3 положительного контроля хорошо действовал в клетках COS, он показал очень низкую промоторную активность в клетках NT2, иллюстрируя тем самым специфическое предпочтение типа клеток для некоторых промоторных последовательностей.
Так как ни клетки COS7, ни клетки NT2 не выражали нормально ICAM-4, тот же эксперимент будет повторен с использованием основных культивированных нейронов гиппокампа крысы, которые действительно выражают ICAM-4 и обязательно выражают механизм транскрипции, необходимый для промоторной активности ICAM-4. Посредством трансфецирования конструкций отдельных промоторов, описанных выше, а также других, в более естественную среду, может стать возможным более точное определение того, какие нуклеотиды в регуляторной области обратного направления ответственны за недостаточную регуляцию гена ICAM-4 в головном мозге.
Последующие примеры иллюстративного характера относятся к предпочтительным вариантам осуществления изобретения, и возможны их многочисленные модификации и варианты, очевидные для специалистов в данной области. Таким образом, объем данного изобретения ограничен только прилагаемой формулой изобретения.
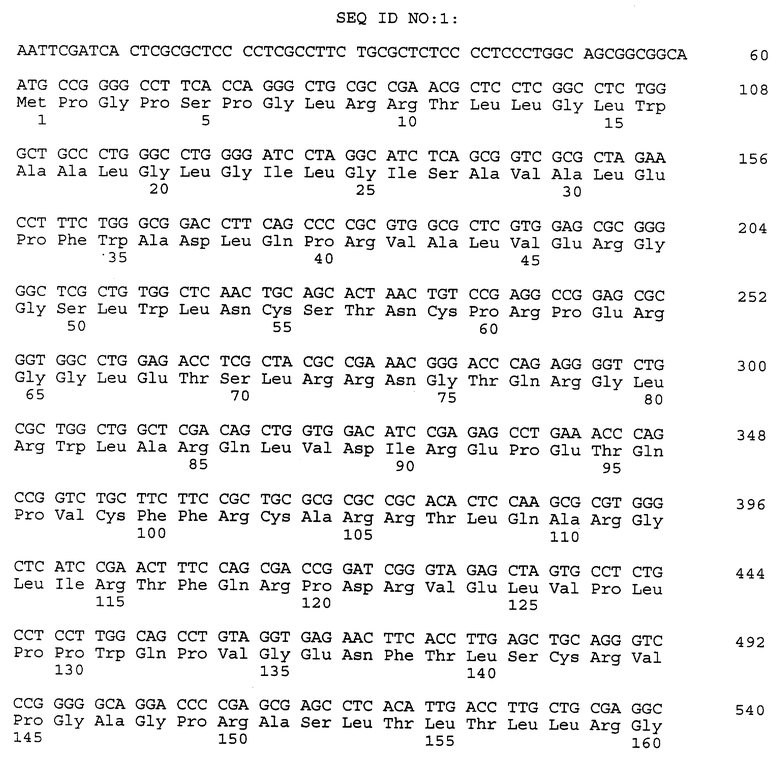
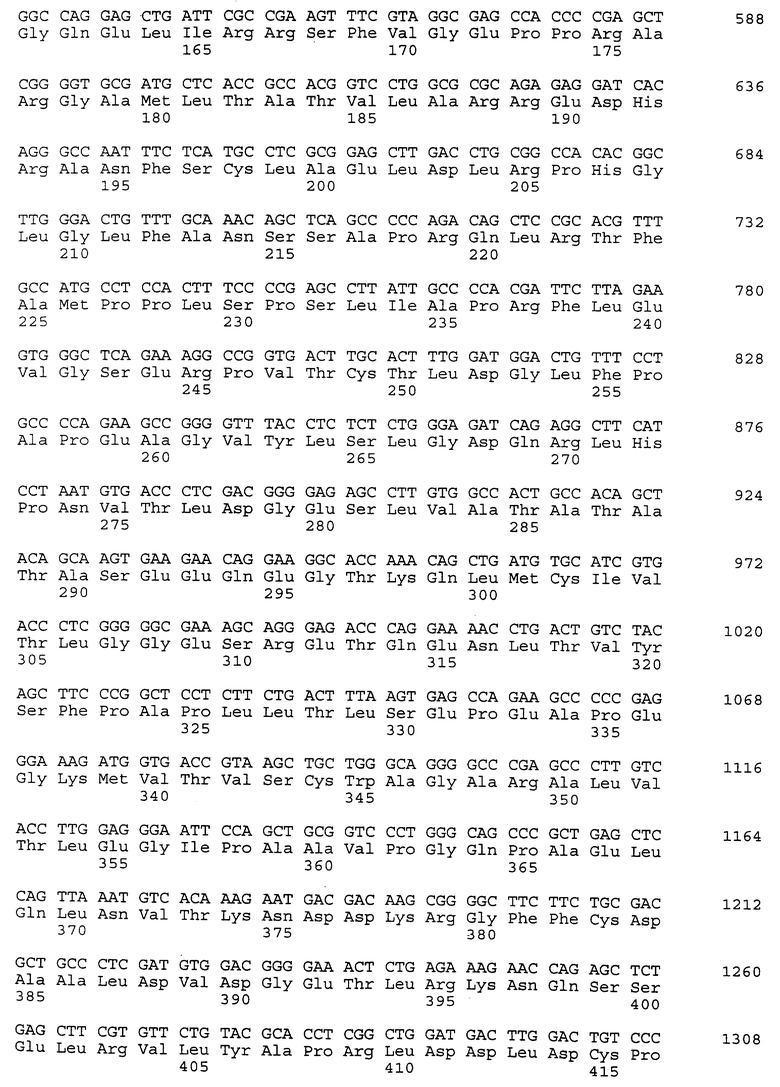
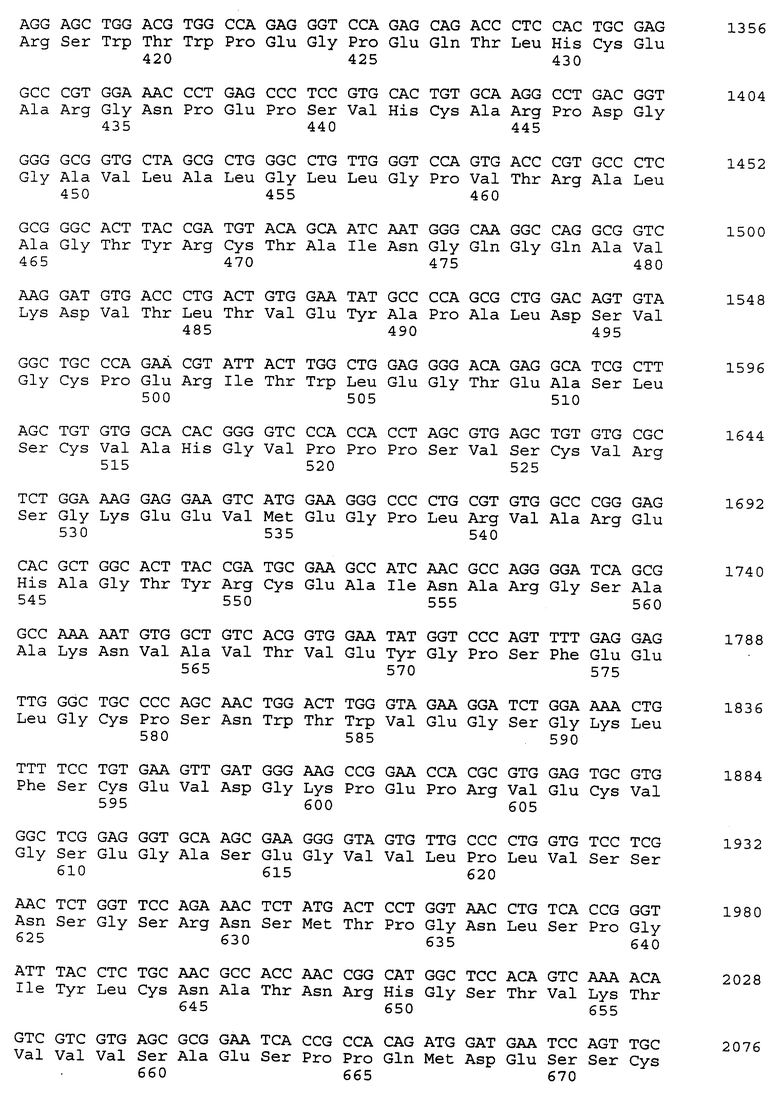
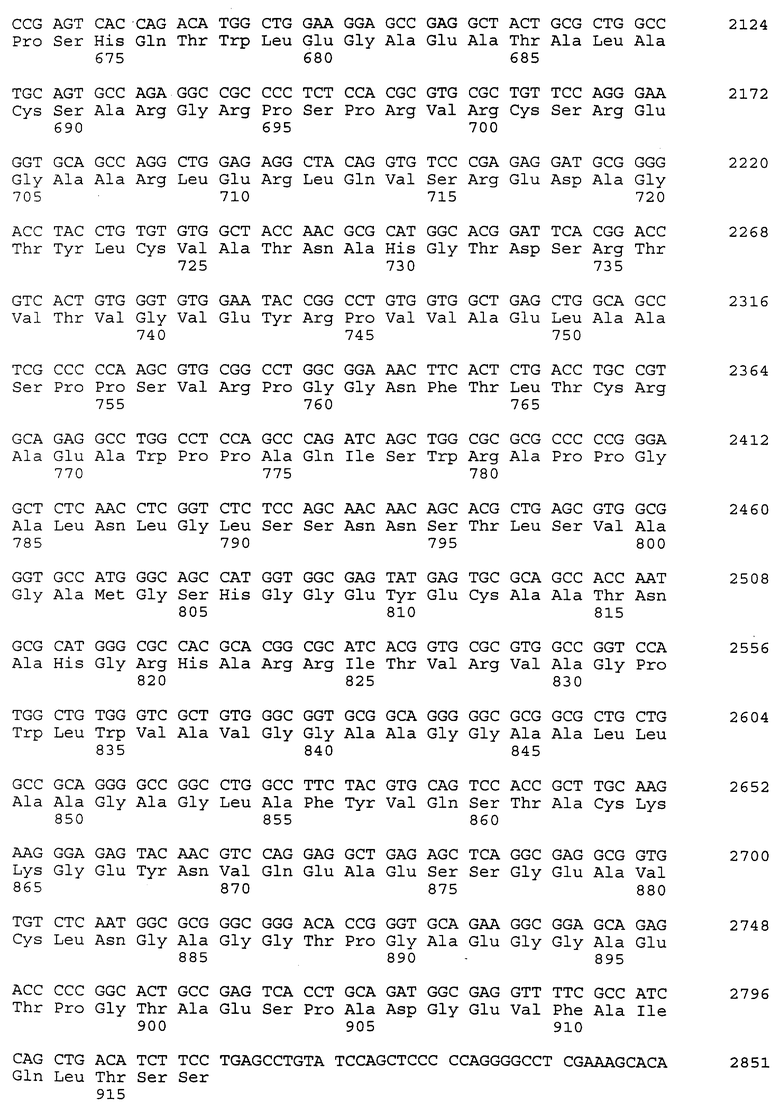

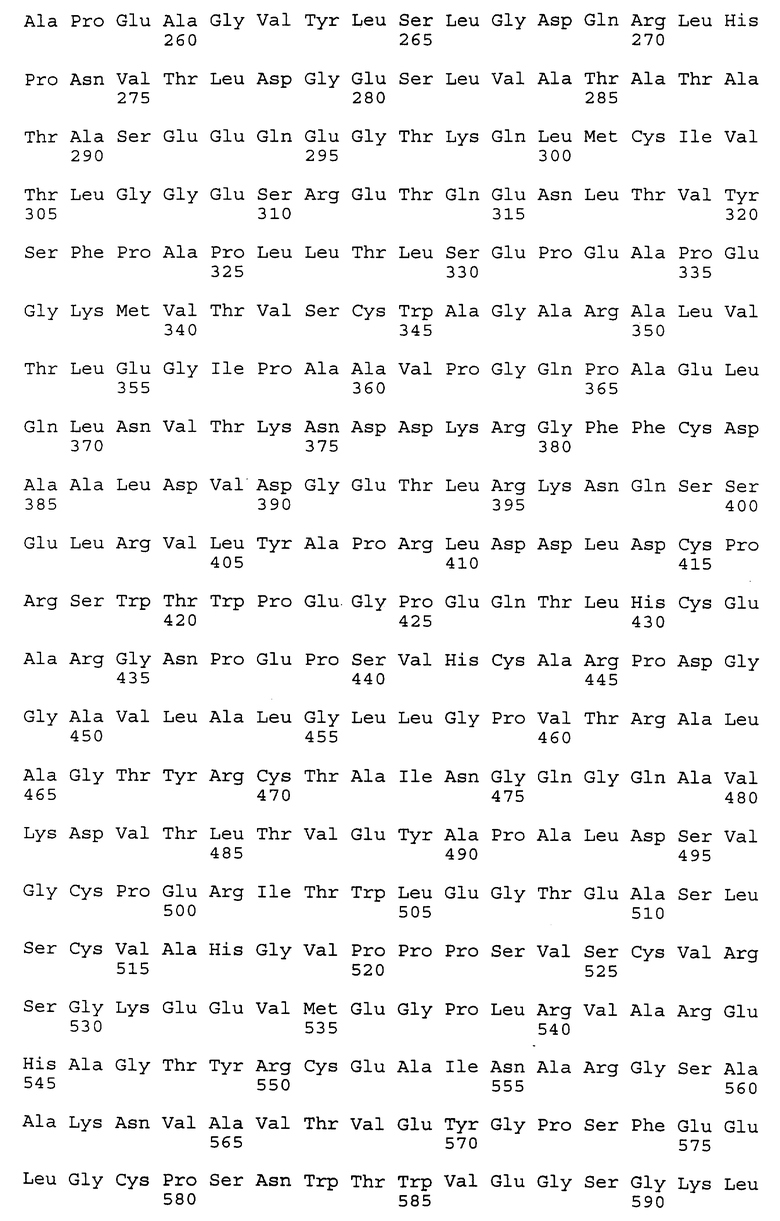



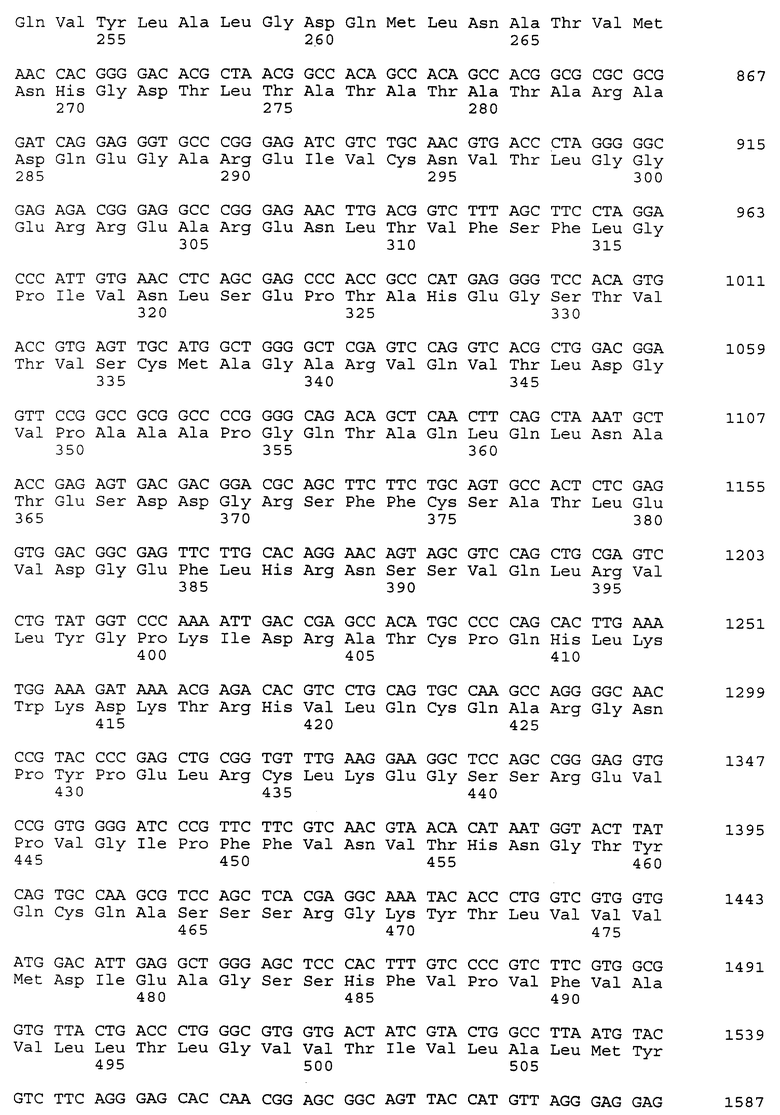


















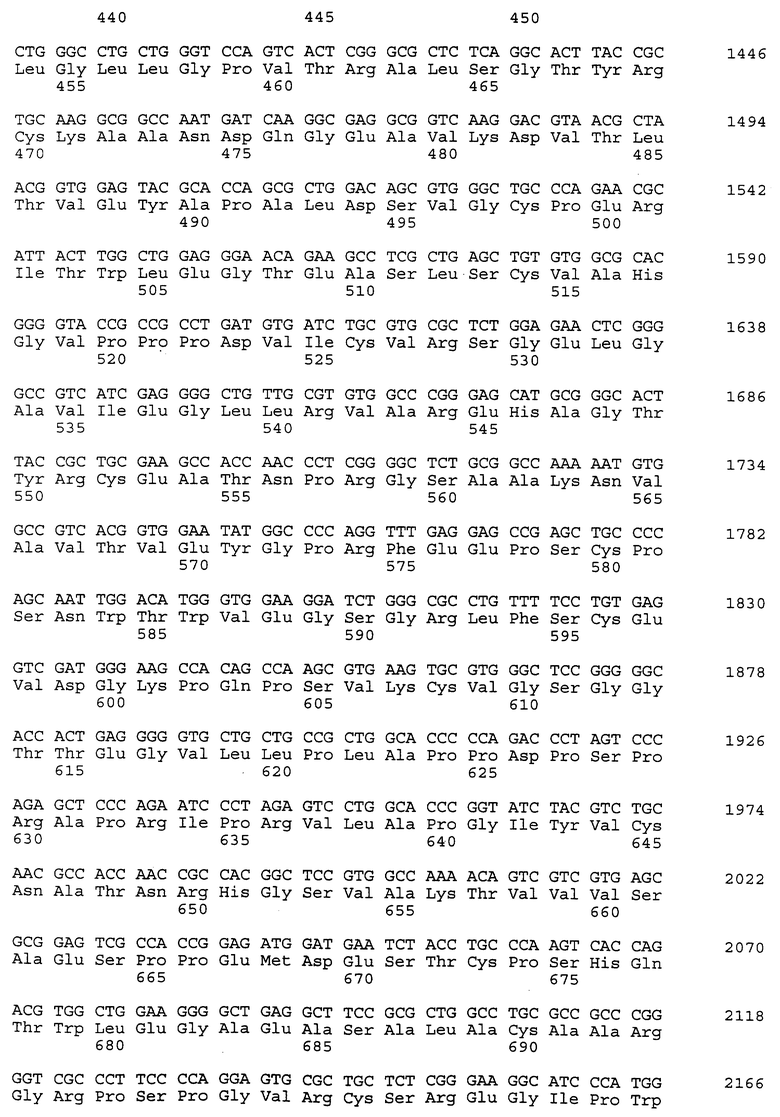
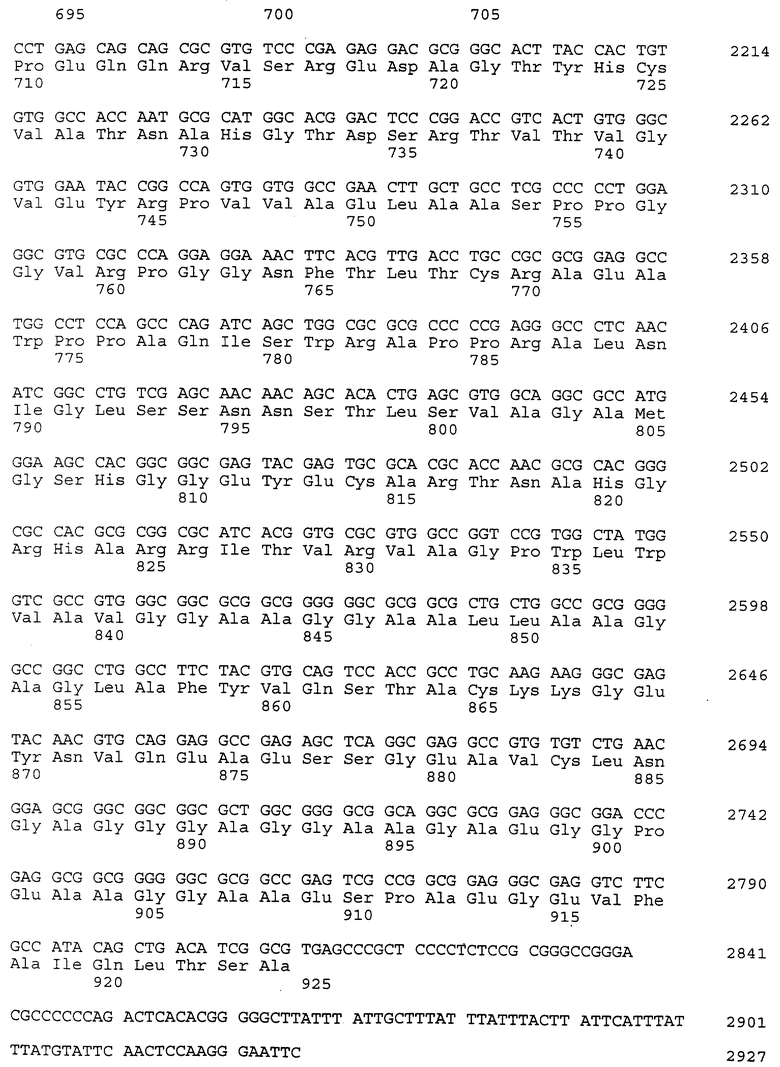












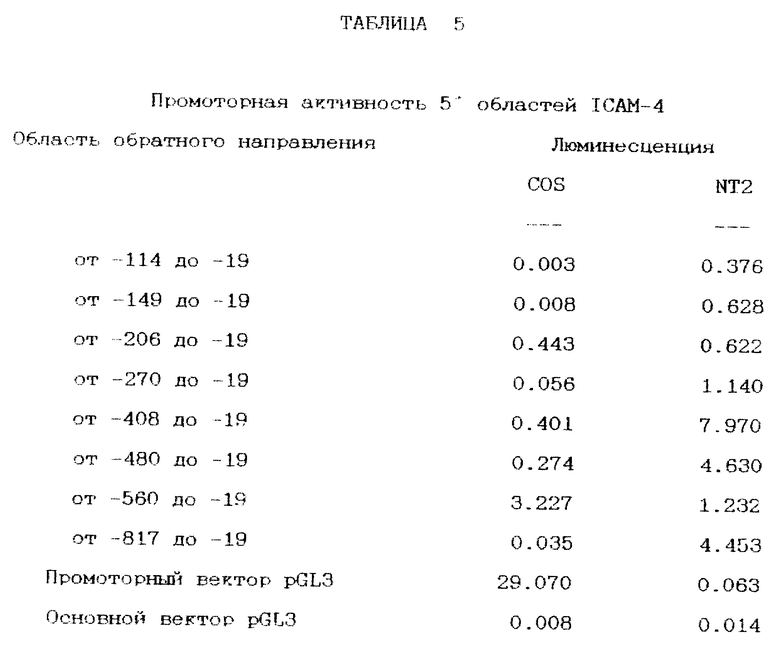
Изобретение предназначено для диагностики невропатологических состояний, например церебральной ишемии, эпилепсии. Способ скрининга для невропатологии включает в себя несколько стадий. Получают образец жидкости от первого индивидуума. Соединяют его с антителом, специфически иммунореактивным с IСАМ-4. Проводят количественный анализ связывания IСАМ-4/антитело в образце. Сравнивают связывание IСАМ/антитело в указанном образце с таким же связыванием в образце от второго индивидуума, не имеющего неврологических заболеваний. Очищенная и выделенная ДНК содержит промотор для IСАМ-4 млекопитающих. Химерный полинуклеотид содержит промотор для гена человеческого IСАМ-4. Вектор экспрессии содержит указанную ДНК. Клетка-хозяин трансформирована указанным вектором экспрессии. Полипептид, обозначенный как "IСАМ-4", относится к факторам клеточной адгезии. Он обладает структурным родством с факторами межклеточной адгезии IСАМ-1, IСАМ-2 и IСАМ-R. Изобретение позволит определить уровни растворимого IСАМ-4 в жидкостях человека в здоровом и больном состоянии. 4 з.п. ф-лы, 5 табл.
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Экономайзер | 0 |
|
SU94A1 |
| Автоматический регулятор усилия подачи бурильных машин | 1972 |
|
SU512301A1 |
| Y | |||
| Yoshihara et al | |||
| An ICAM-related neuronal glycoprotein, teleencephalin, with brain | |||
| Segment-Specific expression | |||
| Neuron | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |

