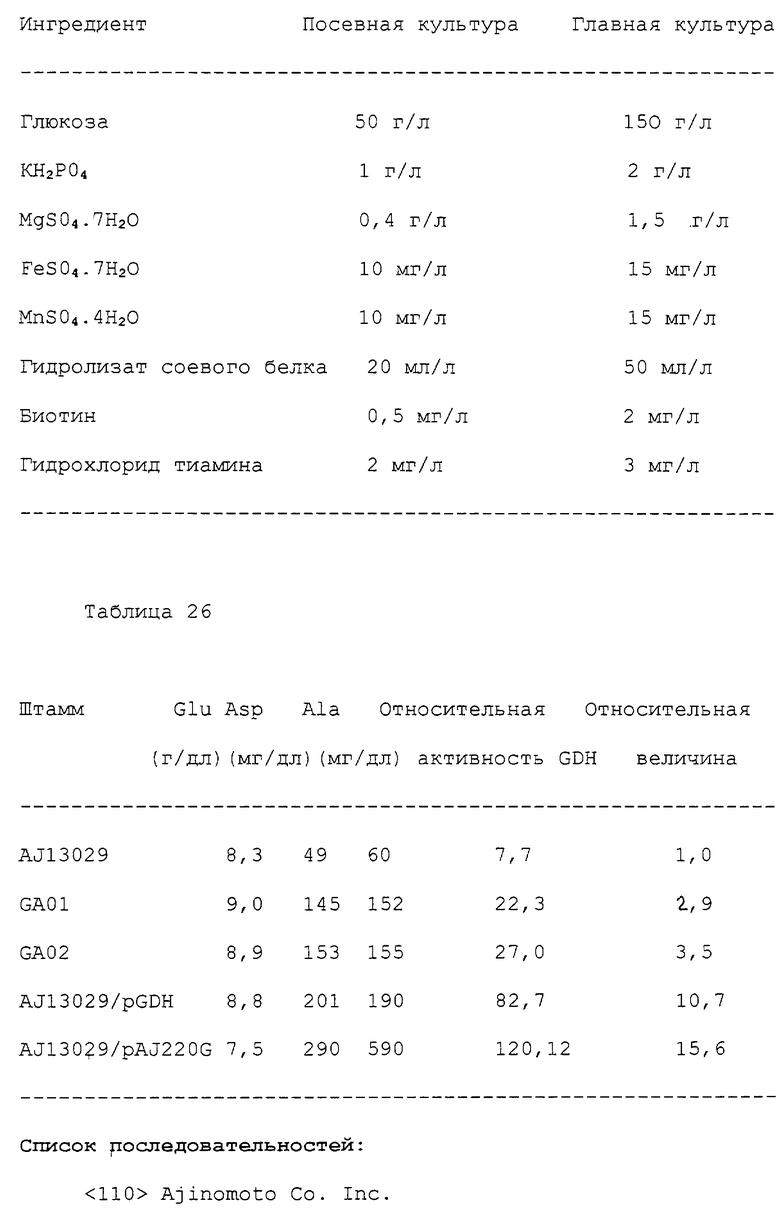
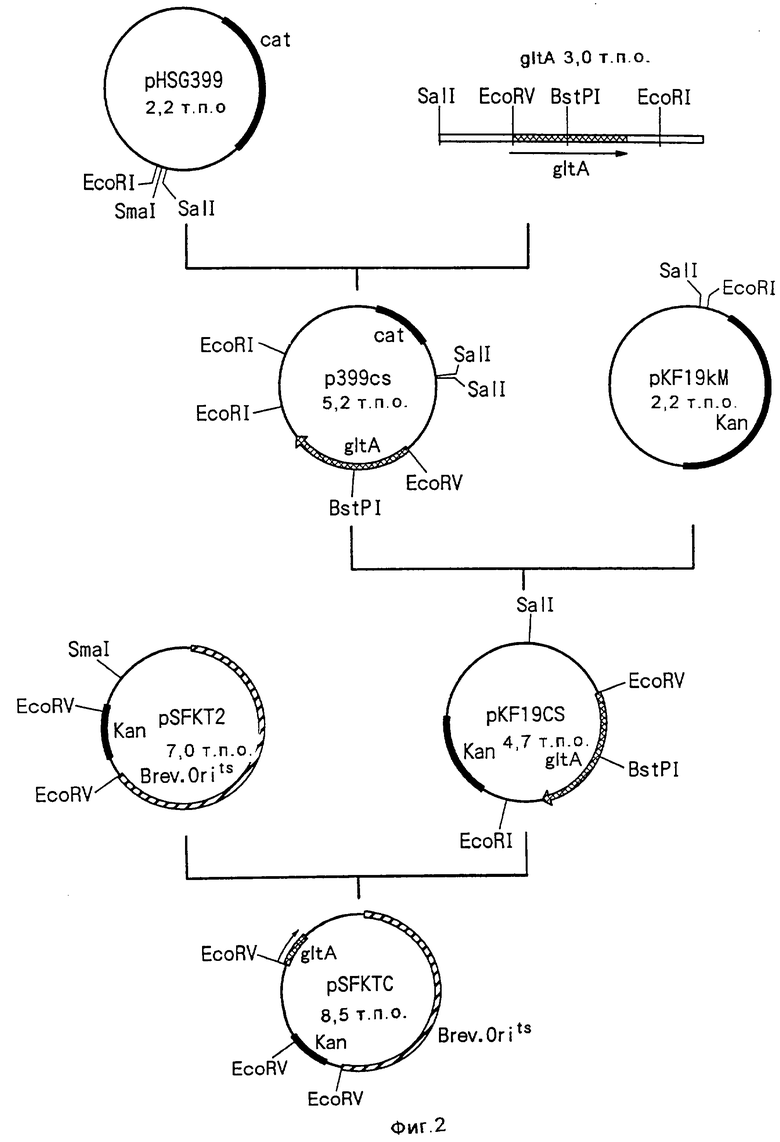
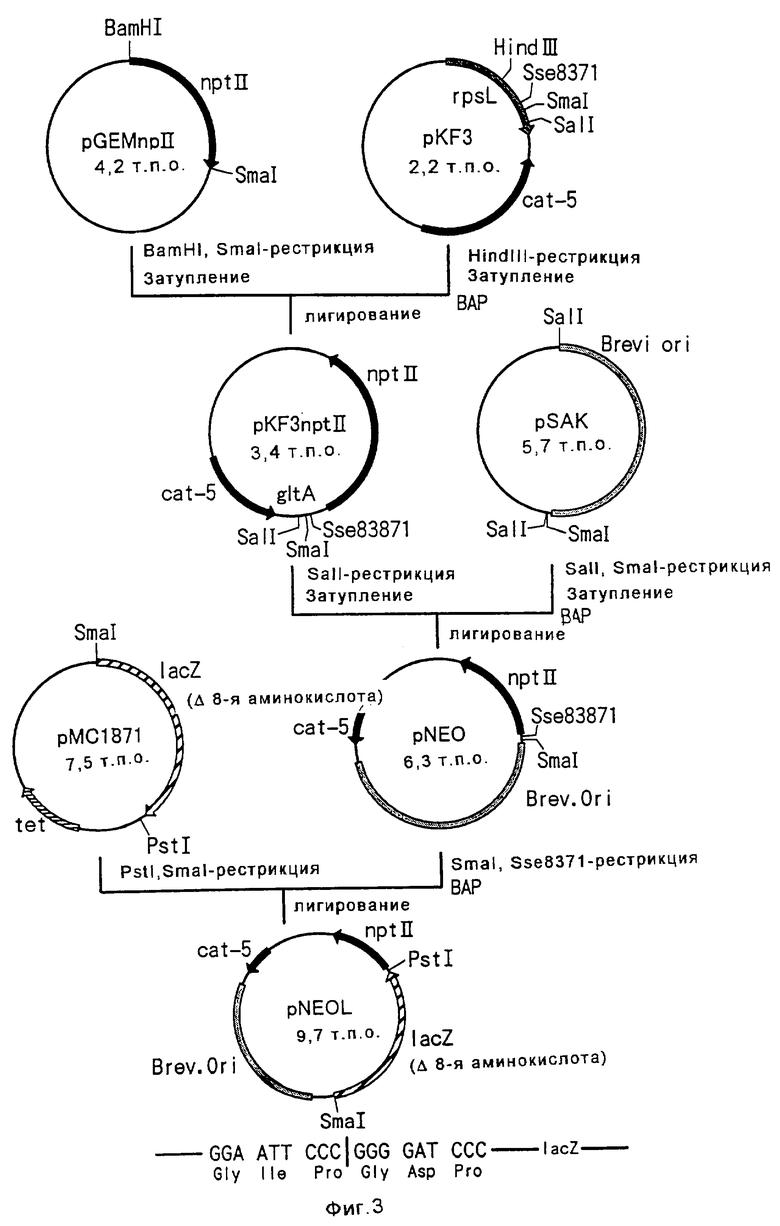
Изобретение относится к биотехнологии и касается способа конструирования штаммов коринеформных бактерий, обладающих повышенной продуктивностью аминокислот, а также способа получения аминокислоты путем ферментации с использованием сконструированных штаммов коринеформных бактерий. Способ конструирования щтаммов коринеформных бактерий включает введение мутации в промоторную последовательность генов биосинтеза аминокислот на хромосоме коринеформной бактерии с получением мутанта коринеформного микроорганизма. Культивирование полученного мутанта. Отбор мутанта, способного продуцировать нужную аминокислоту в больших количествах. Изобретение обеспечивает получение высокого выхода аминокислот посредством усиления и регуляции экспрессии нужного гена без использования плазмиды. 3 с. и 20 з.п. ф-лы, 26 табл., 3 ил.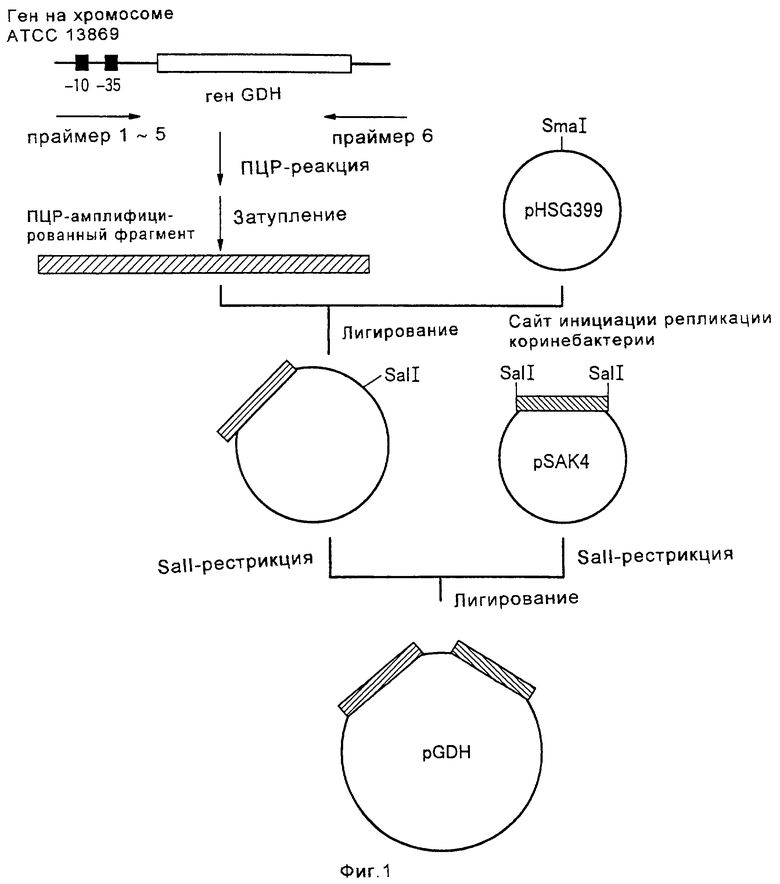
1. Способ конструирования коринеформных бактерий, обладающих повышенной продуктивностью аминокислот, включающий стадии: введение мутации в промоторную последовательность генов биосинтеза аминокислот на хромосоме коринеформной бактерии в целях генерирования последовательности, подобной консенсусной последовательности с получением мутанта коринеформного микроорганизма, продуцирующего аминокислоту; культивирование этого мутанта; и отбор мутанта, способного продуцировать нужную аминокислоту в больших количествах. 2. Способ по п.1, отличающийся тем, что аминокислота выбрана из группы, состоящей из глутаминовой кислоты, лизина, аргинина, серина, фенилаланина, пролина и глутамина. 3. Способ по п.1, отличающийся тем, что аминокислота является глутаминовой кислотой, а промотор для гена биосинтеза выбран из группы, состоящей из промотора гена глутаматдегидрогеназы (GDH), промотора гена цитратсинтазы (CS), промотора гена изоцитратсинтазы (ICDH), промотора гена пируватдегидрогеназы (PDH) и промотора гена аконитазы (АСО). 4. Способ по п.3, отличающийся тем, что промотор гена глутаматдегидрогеназы (GDH) имеет ДНК-последовательность, выбранную из группы, состоящей из (i) по крайней мере, одной ДНК-последовательности, выбранной из группы, состоящей из CGGTCA, TTGTCA, TTGACA и TTGCCA в области -35, (ii) последовательности ТАТААТ или той же самой последовательности ТАТААТ, но в которой основание АТААТ заменено другим основанием, в области -10, и (iii) комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 5. Способ по п.4, отличающийся тем, что промотор для GDH имеет TGGTCA в области -35, и ТАТААТ в области -10, или TTGTCA в области -35 и ТАТААТ в области -10. 6. Способ по п.3, отличающийся тем, что промотор для CS имеет (i) последовательность TTGACA в области -35, (ii) последовательность ТАТААТ в области -10, или (iii) последовательность комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 7. Способ по п.3, отличающийся тем, что по крайней мере один из первого и второго промоторов для ICDH имеет (i) последовательность TTGCCA или TTGACA в области -35, (ii) последовательность ТАТААТ в области -10, или (iii) последовательность комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 8. Способ по п.3, отличающийся тем, что промотор для PDH имеет (i) последовательность TTGCCA в области -35, (ii) последовательность ТАТААТ в области -10, или (iii) последовательность комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 9. Способ по п.1, отличающийся тем, что аминокислота является аргинином, а промотор для гена биосинтеза является промотором для аргининосукцинатсинтазы. 10. Способ по п.9, отличающийся тем, что промотор для аргининосукцинатсинтазы имеет ДНК-последовательность, выбранную из группы, состоящей из (i) по крайней мере, одной ДНК-последовательности, выбранной из группы, состоящей из TTGCCA, TTGCTA и TTGTCA в области -35, (ii) последовательности ТАТААТ или ТАТААТ, но в которой основание АТААС заменено другим основанием, в области -10, и (iii) комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 11. Способ по п.10, отличающийся тем, что промотор для аргининосукцинатсинтазы имеет ДНК-последовательность, выбранную из группы, состоящей из (i) TTGTCA в области -35, (ii) последовательности ТАТААТ в области -10, и (iii) комбинации (i) и (ii). 12. Способ конструирования коринеформных бактерий, обладающих повышенной продуктивностью аминокислот, включающий стадии: введение замены в промоторную последовательность генов биосинтеза аминокислот на хромосоме коринеформной бактерии посредством рекомбинации генов в целях создания последовательности, подобной консенсусной последовательности с получением мутанта коринеформного микроорганизма, продуцирующего аминокислоту; культивирование этого мутанта; и отбор мутанта, способного продуцировать нужную аминокислоту в больших количествах. 13. Способ по п. 12, отличающийся тем, что аминокислота выбрана из группы, состоящей из глутаминовой кислоты, лизина, аргинина, серина, фенилаланина, пролина и глутамина. 14. Способ по п.12, отличающийся тем, что аминокислота является глутаминовой кислотой, а промотор для гена биосинтеза выбран из группы, состоящей из промотора гена глутаматдегидрогеназы (GDH), промотора гена цитратсинтазы (CS), промотора гена изоцитратсинтазы (ICDH), промотора гена пируватдегидрогеназы (PDH) и промотора гена аконитазы (АСО). 15. Способ по п.14, отличающийся тем, что промотор гена глутаматдегидрогеназы (GDH) имеет ДНК-последовательность, выбранную из группы, состоящей из (i) по крайней мере, одной ДНК-последовательности, выбранной из группы, состоящей из CGGTCA, TTGTCA, TTGACA и TTGCCA в области -35, (ii) последовательности ТАТААТ или той же самой последовательности ТАТААТ, но в которой основание АТААТ заменено другим основанием, в области -10, и (iii) комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 16. Способ по п.15, отличающийся тем, что промотор для GDH имеет TGGTCA в области -35, и ТАТААТ в области -10, или TTGTCA в области -35 и ТАТААТ в области -10. 17. Способ по п.14, отличающийся тем, что промотор для CS имеет (i) последовательность TTGACA в области -35, (ii) последовательность ТАТААТ в области -10, или (iii) последовательность комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 18. Способ по п.14, отличающийся тем, что по крайней мере один из первого и второго промоторов для ICDH имеет (i) последовательность TTGCCA или TTGACA в области -35, (ii) последовательность ТАТААТ в области -10, или (iii) последовательность комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 19. Способ по п. 14, отличающийся тем, что промотор для PDH имеет (i) последовательность TTGCCA в области -35, (ii) последовательность ТАТААТ в области -10, или (iii) последовательность комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 20. Способ по п.12, отличающийся тем, что аминокислота является аргинином, а промотор для гена биосинтеза является промотором для аргининосукцинатсинтазы. 21. Способ по п.20, отличающийся тем, что промотор для аргининосукцинатсинтазы имеет ДНК-последовательность, выбранную из группы, состоящей из (i) по крайней мере, одной ДНК-последовательности, выбранной из группы, состоящей из TTGCCA, TTGCTA и TTGTCA в области -35, (ii) последовательности ТАТААТ или ТАТААТ, но в которой основание АТААС заменено другим основанием, в области -10, и (iii) комбинации (i) и (ii), где указанная последовательность не ингибирует функцию промотора. 22. Способ по п.21, отличающийся тем, что промотор для аргининосукцинатсинтазы имеет ДНК-последовательность, выбранную из группы, состоящей из (i) TTGTCA в области -35, (ii) последовательности ТАТААТ в области -10, и (iii) комбинации (i) и (ii). 23. Способ получения аминокислоты путем ферментации, включающий стадии: культивирование коринеформной бактерии, сконструированной способом по любому из пп.1-22 и имеющей повышенную продуктивность аминокислоты или коринеформной бактерии, имеющей ген глутаматсинтазы, имеющий промотор по любому из пп. 4-8 и 15-19, или коринеформной бактерии, имеющей ген аргининсинтазы, имеющий промотор по пп.10 или 21, в культуральной среде с образованием и накоплением нужной аминокислоты в культуральной среде; и сбор этой аминокислоты из культуральной среды.