Область техники, к которой относится изобретение
Изобретение относится к микроорганизму с чужеродными нуклеотидными последовательностями, его применению в качестве лекарственного средства, в особенности вакцины, плазмиде с чужеродными нуклеотидными последовательностями, а также способу получения такого микроорганизма.
Предпосылки создания изобретения и уровень техники
Главной причиной протекания, чаще всего со смертельным исходом, злокачественных опухолевых болезней является неспособность защитной системы организма распознавать и разрушать злокачественные раковые клетки. Онкологические заболевания в промышленных странах относятся к самым частым заболеваниям со смертельным исходом. Так, только в Германии ежегодно умирают свыше 210000 человек со злокачественными новообразованиями (источник: WHO, данные на 1997 г.), что соответствует ежегодному количеству свыше 255 смертельных случаев на 100000 жителей.
Основой настоящего изобретения являются полученные в последнее время новые данные о молекулярных механизмах, которые приводят к злокачественному новообразованию. Уже на очень ранней стадии возникновения онкологического заболевания происходят характерные изменения контроля в отношении роста клеток и/или дифференцировки клеток (Ponten, Cancer Surv., 32, 5-35 (1998)). По существу причастными к этим изменениям являются протеины сигнальной трансдукции и контроля клеточного цикла, которые идентифицированы в последние годы и представляют собой также опухолевые антигены.
Опухолевые антигены грубо подразделяют на три класса (Pardoll, Nat. Med., 4, 525-531 (1998)): i) специфичные к опухоли неоантигены, которые находятся в опухолевой клетке в мутированной и/или суперэксперссируемой форме, как, например, EGF-R, HER-2; ii) специфичные к опухоли эмбриональные антигены, такие как представители протеинового семейства MAGE или СЕА; iii) специфичные к опухолевой ткани дифференцировочные антигены, такие как тирозиназа, Mart-1/melan-A и gp100.
Для эффективности противоопухолевой вакцины решающее значение имеет эффективная индукция CD8+ Т-клеток, так как опухолевые клетки чаще всего не представляют никакие молекулы класса II главного комплекса гистосовместимости, и находящиеся внутри клетки опухолевые антигены чаще всего ограничены классом I главного комплекса гистосовместимости. В случае онкологических больных, естественно имеющихся популяций CD8+, цитотоксических Т-клеток (CTL), по-видимому, недостаточно для распознавания и устранения опухолевых клеток (Jaffee, Ann. N.Y. Acad. Sci., 886, 67-72 (1999)). Сверх того, специфичные к опухоли Т-клетки за счет различных механизмов (анергия, толерантность, нейтрализация) часто не могут эффективно воздействовать на опухолевую ткань (Smyth и др., Nat. Immunol., 2, 293-299 (2001)). Успешно действующая противоопухолевая вакцина поэтому должна нарушать эту анергию или толерантность и индуцировать достаточное количество активированных специфичных CTL, а также специфичных антител. Роль специфичных антител проявляется в успешном использовании моноклональных антител (mAb) против опухолевых антигенов группы (а), как в случае уже коммерчески доступного Herceptin, mAb к HER-2 (Colomber и др., Cancer Invest., 19, 49-56 (2001)).
Уже известно, что аттенуированные внутриклеточные бактерии пригодны в качестве вакциноносителей против определенных бактериальных инфекций, с которыми нужно бороться прежде всего путем так называемого Th1 иммунного ответа (Hess и Kaufmann, FEMS Immunology and Medical Microbiology, 23, 165-173 (1999)). Этот ответ отличается цитотоксическими лимфоцитами (CTL), а также наличием специфичных, секретирующих гамма-интерферон (IFN-g) CD4+ Т-клеток (также Т-клеток-хелперов, Th) (Abbas и др., Nature, 383, 787-793 (1996)). Другие группы исследователей показали, что рекомбинантные бактерии могут защищать против гетерологичной опухоли (Medina и др., Eur. J. Immunol., 29, 693-699 (1999); Pan и др., Cancer Res., 59, 5264-5269 (1999); Woodlock и др., J. Immunother., 22, 251-259 (1999); Paglia и др., Blood, 92, 3172-3176 (1998); Paglia и др., Eur. J. Immunol., 27, 1570-1575 (1997); Pan и др., Nat. Med. 1, 471-477 (1995); Pan и др., Cancer Res., 55, 4776-4779 (1995)). В этих случаях, однако, животных иммунизировали против суррогатного антигена и затем использовали опухолевые клетки, которые экспрессируют этот антиген.
Эти опухолевые системы, однако, нельзя сравнивать с клиническими опухолями, так как в случае этих моделей нет никакой толерантности в отношении опухолевого антигена.
Клинически уже испытано значительное число различных противоопухолевых вакцин. До сих пор, однако, ни с помощью каких-либо противоопухолевых вакцин и ни при использовании какого-либо способа вакцинации не смогли достичь никакого прорыва в лечении опухолевых болезней. Таким образом, существует до сих пор крайне большая потребность в новом способе терапии опухолей.
Известна экспрессия продуктов экспрессии введенных в бактерии нуклеотидных последовательностей в клеточной мембране этих бактерий или возможность секреции этими бактериями. Основой этого способа является гемолизиновая система Escherichia coli HlyAs, которая представляет собой прототип секреторной системы типа I грамотрицательных бактерий. С помощью HlyAs сконструированы векторы секреции, которые делают возможным эффективное выведение антигенов протеинов в случае Salmonellaenterica, Yersinia enterocolitica и Vibrio cholerae. Такого рода векторы секреции содержат кДНК любого антигена протеина, связанную с нуклеотидной последовательностью сигнального пептида HlyA, аппарата секреции гемолизина, hlyB и hlyD и специфичного к hly промотора. С помощью этого вектора секреции протеин можно экспрессировать на поверхности этой бактерии. Такого рода генетически модифицированные бактерии в качестве вакцин индуцируют гораздо более сильную иммунную защиту, чем бактерии, в которых экспрессированный введенной нуклеиновой кислотой протеин остается внутри клетки (Donner и др., заявка на Европейский патент ЕР-1015023-А; Gentschev и др., Gene, 179, 133-140 (1996); Vaccine, 19, 2621-2618 (2001); Hess и др., PNAS, 93, 1458-1463 (1996)). Недостатком этой системы, однако, является то, что за счет применения специфичного к hly промотора количество экспрессированного на наружной поверхности бактерии протеина является очень незначительным.
Разработан способ введения плазмидной ДНК в клетки млекопитающих с помощью бактерий-носителей, таких как Salmonella и Listeria monocytogenes. Содержащиеся в этих плазмидах гены могут экспрессироваться в клетках млекопитающих также в случае, когда они находятся под контролем эукариотического промотора. В микроорганизмы Listeria monocytogenes вводят плазмиды, которые содержат нуклеотидную последовательность любого антигена под контролем любого эукариотического промотора. Введение нуклеотидных последовательностей специфического лизисного гена способствует тому, что микроорганизмы Listeria monocytogenes попадают в цитозоль антигенпредставляющей клетки и высвобождают свои плазмиды, что приводит к последующей экспрессии, процессингу и представлению кодирующих плазмиду протеинов и заметно повышает иммуногенность этих протеинов (Dietrich и др., Nat. Biotechnol., 16, 181-185 (1998); Vaccine, 19, 2506-2512 (2001)).
Получены аттенуированные в отношении вирулентности варианты бактерий, которые попадают внутрь клетки. Например, такого рода варианты Listeria monocytogenes, Salmonella enterica sv. Typhimurium и Typhu, а также Mycobacterium bovis уже используют в качестве приемлемых живых вакцин против тифа и туберкулеза. Эти бактерии, включая их аттенуированные мутанты, в общем, являются иммуностимулирующими и могут вызывать на достаточно хорошем уровне клеточный иммунный ответ. Например, L. monocytogenes за счет активации клеток ТН1 в особенности стимулирует пролиферацию цитотоксических лимфоцитов. Эти бактерии "поставляют" выделенные антигены прямо в цитозоль антигенпредставляющих клеток (АРС; макрофаги и дендритные клетки), которые, со своей стороны, экспрессируют костимулирующие молекулы и вызывают эффективную стимуляцию Т-кеток. Листерии отчасти подвергаются разложению в фагосомных компартментах и продуцированные этими бактериями-носителями антигены поэтому, с одной стороны, могут представляться молекулами класса II главного комплекса гистосовместимости и вместе с тем приводят к индукции Т-клеток-хелперов. С другой стороны, листерии, после высвобождения из фагосомы, реплицируются в цитозоле антигенпредставляющих клеток (APC); продуцированные и выделенные этими бактериями антигены поэтому предпочтительно представляются молекулами класса I главного комплекса гистосовместимости, благодаря чему индуцируются CTL ответы против этих антигенов. Кроме того, показано, что за счет взаимодействия листерий с макрофагами, естественными клетками-киллерами (NK) и нейтрофилами гранулоцитарного ряда индуцируется экспрессия таких цитокинов (альфа-фактор некроза опухоли, гамма-интерферон, интерлейкин-2, интерлейкин-12; Unanue, Curr. Opin. Immunol., 9, 35-43 (1997); Mata и Paterson, J. Immunol., 163, 1449-1456 (1999)), для которых доказана противоопухолевая эффективность. Так, за счет введения L. monocytogenes, которые подвергнуты трансдукции в отношении экспрессии опухолевых антигенов, можно ингибировать антигенспецифическиЙ рост экспериментальных опухолей (Pan и др., Nat. Med., 1, 471-477 (1995); Cancer Res., 59, 5264-5269 (1999); Voest и др., Natl. Cancer Inst., 87, 581-586 (1995); Beatty и Paterson, J. Immunol., 165, 5502-5508 (2000)).
Аттенуированные в отношении вирулентности штаммы Salmonella enterica, в которые введены нуклеотидные последовательности, кодирующие опухолевые антигены, в качестве экспрессирующих опухолевые антигены бактериальных носителей после введения перорально могут вызывать специфическую защиту против различных экспериментальных опухолей (Medina и др., Eur. J. Immunol., 30, 768-777 (2000); Zoller и Christ, J. Immunol., 166, 3440-3450 (2001); Xiang и др., PNAS, 97, 5492-5497 (2000)).
Рекомбинантные штаммы Salmonella также в качестве профилактических вакцин оказались эффективными против вирусных инфекций (вирус папилломы человека (HPV)) (Benyacoub и др., Infect. Immun., 67, 3674-3679 (1999)) и для терапевтической обработки иммортализированной онкогенным вирусом (вирус папилломы человека) мышиной опухоли (Revaz и др., Virology, 279, 354-360 (2001)).
Техническая задача согласно изобретению
В основу изобретения положена техническая задача получения лекарственного средства, которое в особенности в случае профилактики и терапии опухолей представляет собой улучшенную вакцину с преодоленной иммунотолерантностью по отношению к опухолям.
Основная концепция изобретения
В целях решения этой технической задачи изобретение относится к микроорганизму с нуклеотидной последовательностью, кодирующей клеточный антиген, в геном которого вводят и подвергают экспрессии следующие компоненты: I) нуклеотидная последовательность, кодирующая по меньшей мере один эпитоп антигена или нескольких антигенов опухолевой клетки, и/или нуклеотидная последовательность по меньшей мере одного эпитопа антигена или нескольких антигенов, специфичного к тканевой клетке, от которой происходит опухоль; II) необязательно, нуклеотидная последовательность, кодирующая протеин, который стимулирует клетки иммунной системы; IIIA) нуклеотидная последовательность транспортной системы, которая обеспечивает экспрессию продукта экспрессии компонентов I) и, необязательно, II) на наружной поверхности бактерии и/или секрецию продукта экспрессии компонента I) и, необязательно, компонента II); и/или IIIB) нуклеотидная последовательность протеина для лизиса микроорганизмов в цитозоле клеток млекопитающих и для внутриклеточного высвобождения плазмид, содержащихся в лизируемых микроорганизмах; и IV) активирующая последовательность для экспрессии одного или нескольких компонентов от I) до IIIB), выбираемая из группы, состоящей из "активируемой в микроорганизме, специфичной к тканевой клетке и неспецифичной к клетке активирующей последовательности", причем каждый из компонентов от I) до IV) может быть представлен однократно или многократно, соответственно, будучи одинаковыми или разными; а также к применениям такого микроорганизма для получения лекарственного средства.
Объектом изобретения, таким образом, являются микроорганизмы, которые представляют собой носители нуклеотидных последовательностей, кодирующих клеточные антигены, которые, в свою очередь, экспрессируются или секретируются снова в наружной мембране микроорганизмов, и применение этих микроорганизмов для подавления иммунотолерантности в отношении опухолей, и новые противоопухолевые вакцины, которые содержат микроорганизмы в качестве носителей нуклеотидных последовательностей, кодирующих клеточные антигены нормальных клеток и/или опухолевых клеток. Согласно изобретению, в конечном счете, возникает направленная против опухоли иммунная реакция.
В частности, предлагаемые согласно изобретению микроорганизмы содержат следующие компоненты: I) по меньшей мере одна нуклеотидная последовательность, кодирующая по меньшей мере один эпитоп по меньшей мере одного антигена по меньшей мере одного клеточного протеина опухолевой клетки и/или, на выбор, по меньшей мере одна нуклеотидная последовательность по меньшей мере одного эпитопа по меньшей мере одного антигена, специфичного к тканевой клетке, от которой происходит опухоль; II) на выбор, по меньшей мере одна нуклеотидная последовательность по меньшей мере одного протеина, который стимулирует клетки иммунной системы; IIIA) по меньшей мере одна нуклеотидная последовательность транспортной системы для постоянной мембранной экспрессии или для секреции кодированного компонентом I) клеточного антигена и для секреции кодированного компонентом II) иммуностимулирующего протеина; IIIB) на выбор, нуклеотидная последовательность лизина, которая лизирует микроорганизм в цитозоле, так что содержащиеся в микроорганизме плазмиды высвобождаются в цитозоле; IV) по меньшей мере одна нуклеотидная последовательность активирующей последовательности, активируемой в микроорганизме или активируемой неспецифично к клетке, специфично к опухолевой клетке, специфично к тканевой клетке или специфично к функции, для экспрессии компонентов I) и II).
Предпочтительные варианты осуществления
Ниже по отдельности описываются компоненты предлагаемого согласно изобретению микроорганизма.
Компонент I)
Компонент I) представляет собой по меньшей мере одну нуклеотидную последовательность по меньшей мере одного эпитопа по меньшей мере одного антигена по меньшей мере одного клеточного протеина или по меньшей мере одного онкогенно мутированного клеточного протеина опухолевой клетки. Онкогенная мутация клеточного протеина может быть вызвана потерей или усилением его первоначальных клеточных функций. Далее, этот клеточный протеин может быть выбран из группы, состоящей из "рецепторных молекул или их частей, а именно внеклеточных, трансмембранных или внутриклеточных частей рецепторов; адгезионных молекул или их частей, а именно внеклеточной, трансмембранной или внутриклеточной части адгезионных молекул; протеинов сигнальной трансдукции; протеинов контроля клеточного цикла; дифференцировочных протеинов; эмбриональных протеинов и индуцированных вирусом протеинов". Такого рода клеточные антигены осуществляют в клетке регуляцию роста и деления клетки и представляются в клеточной мембране нормальных клеток, например, благодаря молекуле класса I главного комплекса гистосовместимости. В опухолевых клетках эти клеточные антигены часто суперэкспрессируются или специфически подвергаются мутации. Следствием такого рода мутаций могут быть ограничения функции онкогенных супрессоров или активация протоонкогенов до онкогенов, и сами мутации или в совокупности со сверхпродуцированиями в значительной степени могут принимать участие в росте опухоли. Такого рода клеточные антигены представляются в мембране опухолевых клеток и соответственно этому представляют собой антигены опухолевых клеток, однако, не вызывая влияющей на опухолевую болезнь пациента иммунной реакции. Раппом (Rapp; патент США 5156841) уже описано применение онкопротеинов, то есть продуктов экспрессии онкогенов в качестве иммуногенов противоопухолевых вакцин. Этот литературный источник включен в настоящую заявку путем ссылки.
Примерами клеточных антигенов и их онкогенных мутаций согласно изобретению являются: i) рецепторы, такие как, например, Her-2/neu, андрогеновый рецептор, эстрогеновый рецептор, мидкиновый рецептор, EGF-рецептор, ERBB2, ERBB4, TRAIL-рецептор, FAS, рецептор альфа-фактора некроза опухоли; ii) трансдуцирующие сигнал протеины и их онкогенные мутации, такие как, например, с-Raf (Raf-1), A-Raf, B-Raf, Ras, Bcl-2, Bcl-X, Bcl-W, Bfl-1, Brag-1, Mcl-1, A1, Bax, BAD, Bak, Bcl-Xs, Bid, Bik, Hrk, Bcr/abl, Myb, C-Met, IAP1, IAО2, XIAP, ML-IAP LIVIN, Survivin, APAF-1; iii) протеины контроля клеточного цикла и их онкогенные мутации, такие как, например, циклин-D (1-3), циклин-E, циклин-A, циклин-B, циклин-H, Cdk-1, Cdk-2, Cdk-4, Cdk-6, Cdk-7, Cdc25C, P16, p15, p21, p27, p18, pRb, p107, p130, E2F (1-5), GAAD45, MDM2, PCNA, ARF, PTEN, APC, BRCA, P53 и гомологи; iv) факторы транскрипции и их онкогенные мутации, такие как, например, С-Myc, NFkB, c-Jun, ATF-2, Sp1; v) эмбриональные протеины, такие как, например, карциноэмбриональный антиген, альфа-фетопротеин, Mage, PSCA; vi) антигены дифференцировки, такие как, например, Mart, Gp100, тирозиназа, GRP, TCF-4; vii) вирусные антигены, такие как, например, антигены следующих вирусов: вирус папилломы человека, вирус гепатита С, вирус Эпштейна-Барра, вирус огуречной мозаики, вирус простого герпеса.
Альтернативно или дополнительно, компонент I) может представлять собой по меньшей мере одну нуклеотидную последовательность по меньшей мере одного антигена, специфичного к нормальной тканевой клетке, от которой происходит соответствующая опухоль. Такого рода специфичными антигенами являются, например, i) рецепторы, такие как, например, андрогеновые рецепторы, эстрогеновые рецепторы, лактоферриновый рецептор; ii) антигены дифференцировки, такие как, например, основной миелин, альфа-лактальбумин, GFAP, PSA, фибриллярный кислый протеин, тирозиназа, EGR-1, MUC1.
Компонент II)
Компонент II) представляет собой по меньшей мере одну нуклеотидную последовательность по меньшей мере одного протеина, который стимулирует клетки иммунной системы. За счет выбора протеина можно усиливать иммунную реакцию в отношении продукта экспрессии компонента I) и/или более ориентировать на активацию клеток Th1 (в случае клеточной иммунной реакции) или на активацию клеток Th2 (в случае гуморальной иммунной реакции). Иммуностимулирующими протеинами являются, например, i) цитокины, такие как M-CSF, GM-CSF, G-CSF; ii) интерфероны, как альфа-интерферон, бета-интерферон, гамма-интерферон; iii) интерлейкины, такие как интерлейкин-1, -2, -3, -4, -5, -6, -7, -9, -10, -11, -12, -13, -14, -15, -16, фактор ингибирования лейкоза человека (LIF); iv) хемокины, такие как Rantes, хемотактический и активирующий моноциты фактор (MCAF), протеин-1 воспаления макрофагов (MIP-1-a, -Я), протеин-2 активации нейтрофилов (NAP-2), интерлейкин-8.
Компонент IIIA)
Компонентом IIIA) является по меньшей мере одна нуклеотидная последовательность, кодирующая по меньшей мере одну транспортную систему, которая делает возможной экспрессию продуктов экспрессии компонента I) и, необязательно, компонента II) на наружной поверхности микроорганизма. Соответствующий компонент при этом, на выбор, может либо секретироваться, либо экспрессироваться на мембране микроорганизма, то есть подвергаться постоянной мембранной экспрессии. Такого рода транспортными системами являются, например, i) гемолизиновый транспортный сигнал E. coli (нуклеотидные последовательности, содержащие HlyA, HlyB и HlyD при контроле специфичного к hly промотора); можно использовать следующие транспортные сигналы: для секреции - С-концевой транспортный сигнал HlyA, в присутствии протеинов HlyB и HlyD; для постоянной мембранной экспрессии - С-концевой транспортный сигнал HlyA, в присутствии протеина HlyB; ii) гемолизиновый транспортный сигнал E. coli (нуклеотидные последовательности, содержащие HlyA, HlyB и HlyD под контролем неспецифичного к hly бактериального промотора); iii) транспортный сигнал протеина S-ряда (Rsa A) Caulobacter crescentus; можно использовать следующие транспортные сигналы: для секреции и для постоянной мембранной экспрессии - С-концевой транспортный сигнал RsaA; iv) транспортный сигнал TolC-протеина Escherichia coli; можно использовать следующие транспортные сигналы: для постоянной мембранной экспрессии - N-концевой транспортный сигнал протеина TolC (интегральный мембранный протеин TolC из E. coli представляет собой многофункциональный порообразующий протеин наружной мембраны E. coli, который наряду с такими функциями, как, например, поглощение колицина Е1 (Morona и др., J. Bacteriol., 153, 693-699 (1983)) и секреция колицина V (Fath и др., J. Bacteriol., 173, 7549-7556 (1991)), также служит в качестве рецептора U3-фагов (Austin и др., J. Bacteriol., 172, 5312-5325 (1990)); этот протеин находится не только у E. coli, но и также у множества грамотрицательных бактерий (Wiener, Structure Fold. Des., 8, 171-175 (2000)); локализация в наружной мембране и наличие в большом количестве делают TolC идеальным кандидатом для представления гетерологичных антигенов для того, чтобы, например, вызвать иммунный ответ).
Компонент IIIB)
Компонент IIIB) представляет собой нуклеотидную последовательность, кодирующую по меньшей мере один литический протеин, который экспрессируется в цитозоле клетки млекопитающего и лизирует микроорганизм для высвобождения плазмид в цитозоле клетки-хозяина. Такого рода литическими протеинами (эндолизинами) являются, например, специфичные к листериям лизисные протеины, такие как, например, PLY551 (Loessner и др., Mol. Microbiol., 16, 1231-1241 (1995)) и/или специфичный к Listeria холин под контролем листерийного промотора.
Предпочтительным вариантом осуществления настоящего изобретения является комбинация различных компонентов IIIB), например комбинация лизисного протеина с холином.
Компоненты IIIA и/или IIIB могут быть конститутивно активны.
Компонент IV)
Компонент IV) представляет собой по меньшей мере одну нуклеотидную последовательность по меньшей мере одной активирующей последовательности для экспрессии компонента I) и, необязательно, компонента II).
Если имеет место постоянная мембранная экспрессия на наружной поверхности микроорганизма, то активирующую последовательность предпочтительно нужно выбирать так, чтобы она активировалась в микроорганизме. Такого рода активирующими последовательностями, например, являются: i) конститутивно активные участки промотора, такие как участок промотора с "участком связывания рибосомы" (RBS) бета-лактамазного гена E. coli или tetA-гена (Busby и Ebright, Cell, 79, 743-746 (1994)); ii) индуцируемые промоторы, предпочтительно промоторы, которые после попадания в клетку становятся активными. К ним относится actA-промотор L. monocytogenes (Dietrich и др., Nat. Biotechnol., 6, 181-185 (1998)) или pagC-промотор S. typhimurium (Bumann, Infect. Immun., 69, 7493-7500 (2001)).
Если плазмиды высвобождаются из микроорганизма после его лизиса в цитозоле клетки, то активирующая последовательность является неспецифичной к клетке, специфичной к тканевой клетке, специфичной к клеточному циклу или функционально специфичной. Предпочтительно выбирают активирующие последовательности, которые особенно активируются в макрофагах, дендритных клетках и лимфоцитах.
Микроорганизмами в смысле настоящего изобретения являются вирусы, бактерии или одноклеточные паразиты, которые обычно используют для переноса микроорганизмом чужеродных нуклеотидных последовательностей.
Согласно особому варианту настоящего изобретения микроорганизмы представляют собой грамположительные или грамотрицательные бактерии, предпочтительно такие бактерии, как, например, Escherichia coli, Salmonella, Yersinia enterocolitica, Vibrio cholerae, Listeria monocytogenes, Shigella.
Предпочтительно используют такие бактерии, которые аттенуированы в отношении своей вирулентности.
Компоненты согласно изобретению вводят в микроорганизмы известными специалисту способами. Если микроорганизмы представляют собой бактерии, то компоненты вводят в плазмиды и плазмиды переносят в бактерии. Пригодные для этой цели способы и плазмиды в достаточной степени известны специалисту.
Объектом изобретения являются лекарственные композиции, содержащие предлагаемые согласно изобретению микроорганизмы или мембранные оболочки этих микроорганизмов. Эти мембранные оболочки получают, например, по способу, описанному в заявке на Европейский патент ЕР-А-0540525. Такого рода лекарственными композициями являются, например, суспензии предлагаемых согласно изобретению микроорганизмов в обычно используемых специалистом растворах, которые пригодны для инъекции.
Еще одним объектом изобретения является введение лекарственной композиции, содержащей предлагаемые согласно изобретению микроорганизмы. Введение осуществляют локально или системно, например, в эпидермис, подкожно, в мускулатуру, в полость тела, в орган, в опухоль или в систему кровообращения.
Особым объектом настоящего изобретения является пероральное или ректальное введение предлагаемой согласно изобретению лекарственной композиции для профилактики и/или лечения пролиферативного заболевания. Введение можно осуществлять однократно или многократно. В случае любого введения вводят в пределах от 10 до 109 предлагаемых согласно изобретению микроорганизмов. Если введение этого количества предлагаемых согласно изобретению микроорганизмов не вызывает никакой достаточной иммунной реакции, нужно повышать инъецируемое количество.
После введения предлагаемых согласно изобретению микроорганизмов нарушается толерантность к клетке, представляющей компонент I), например к опухолевой клетке или к тканевой клетке, от которой происходит опухоль, и возбуждается цитотоксическая иммунная реакция, направленная против опухоли и/или против ее тканевых клеток.
В зависимости от выбора компонента I) эта цитотоксическая иммунная реакция направлена либо исключительно против опухоли, либо также против опухолевых клеток, включая тканевые клетки, от которых происходят опухолевые клетки.
Объектом изобретения является, таким образом, введение предлагаемой согласно изобретению лекарственной композиции для профилактики или лечения пролиферативного заболевания. К пролиферативным заболеваниям нужно отнести опухолевые заболевания, лейкозы, вызываемые вирусом заболевания, хронические воспаления, отторжения трансплантированных органов и аутоиммунные заболевания.
Согласно особому варианту осуществления настоящего изобретения, в случае которого компонент I) представляет собой по меньшей мере один клеточный антиген, который экспрессируется опухолевой клеткой и тканевыми клетками, от которых происходит опухоль, предлагаемую согласно изобретению лекарственную композицию вводят для профилактики или лечения опухоли щитовидной железы, молочной железы, желудка, почки, яичника, родимых пятен, простаты, шейки (органа) или мочевого пузыря.
Ниже изобретение поясняется подробнее с помощью примеров, представляющих собой только варианты осуществления.
Пример 1
Индукция иммунного ответа у ВхВ-мышей путем иммунизации с помощью экспрессирующих с-Raf сальмонелл
Raf представляет собой обычно цитозольную серин/треонинкиназу (PSK), которая в сочетании с другими протеинами сигнальных каскадов контролирует рост и выживание клеток (Kerkhoff и Rapp, Oncogene, 17, 1457-1462 (1998); Troppmair и Rapp, Recent Results Cancer Res., 143, 245-249 (1997)). Связывание фактора роста с соответствующим рецептором, обычно за счет активации Ras, активации Raf, за счет нескольких стадий фосфорилирования, за счет PSK- и тирозинкиназы МЕК и PSK ERK, приводит к активации механизма репликации в клеточном ядре (Kerkhoff и Rapp, Oncogene, 17, 1457-1462 (1998)). Первый член этой цепи, небольшой G-протеин Ras, бывает измененным примерно в 30% всех человеческих опухолей (Zachos и Spandidos, Crit. Rev. Oncol. Hematol., 26, 65-75 (1997)). Raf представляет собой эффектор Ras и суперэкспрессируется во множестве человеческих опухолей (Naumann и др., Recent Results Cancer Res., 143, 237-244 (1997)).
Для теста на мышиной модели используют трансгенных мышей, которые сверхпродуцируют полную молекулу или конститутивно активный киназный домен (ВхВ) (Kerkhoff и др., Cell Growth Differ, 11, 185-190 (2000)). Тем самым у этих мышей спустя примерно полгода обнаруживаются опухоли легких.
Для создания вакцины человеческую с-Raf кДНК клонировали с помощью полимеразной цепной реакции (ПЦР) "в рамке считывания" с HlyA в плазмиде pMOhly-1 (фиг.1). Затем плазмиду pMO-Raf подвергали трансфекции в аттенуированных сальмонеллах (S. typi murium SL7207), которые обладают дефектом в ароматическом обмене веществ (Hoiseth и Stocker, Nature, 291, 238-239 (1981)). Согласно иммуноблоттингу с помощью антител, направленных к с-Raf, в бактериальном лизате, а также в культуральном супернатанте трансфицированных с помощью pMOhy-Raf бактерий SL7207 можно было обнаружить слитый протеин с-Raf HlyAs.
Теперь трансгенных ВхВ-мышей в возрасте 7-10 недель иммунизировали перорально с помощью сальмонелл (доза 5·109), причем вакцинацию повторяли 2 раза с интервалом 5 дней. Спустя 45 дней после последней иммунизации осуществляли ревакцинацию с помощью 5·105 сальмонелл. Для контроля мышам внутримышечно вводили "голую" ДНК, кодирующую с-Raf.
Спустя 5-7 дней после последней иммунизации отбирали пробы сыворотки и анализировали с помощью вестерн-блотирования антителогенез. Для этого разведенную в соотношении 1:200 сыворотку гибридизировали с мембранами трансфицированных с помощью отделенного протеина и блотированного протеина с-Raf или нетрансфицированных бактерий. Обнаружение связанных сывороточных антител осуществляли с помощью антител, специфичных к мышиному иммуноглобулину-G (IgG). В противоположность контрольным мышам у мышей, иммунизированных бактериями SL7207, трансфицированными с помощью pMohly-Raf, могли индуцироваться специфичные к с-Raf антитела изотипа IgG. Тем самым показано, что за счет иммунизации с помощью описанных сальмонелл можно нарушать самотолерантность, и иммунизация вызывает индукцию CD4+ Т-кеток, которые необходимы для изменения изотипов антител в отношении IgG.
Для анализа CD8+ Т-клеточного ответа, C57BL-6-мышей иммунизировали согласно такому же протоколу. Спустя 7 дней после последней иммунизации выделяли клетки селезенки и стимулировали их с помощью суперэкспрессирующих Raf клеток EL-4. Спустя 1 час после начала стимуляции блокировали везикулярный транспорт за счет брефелдина А и спустя следующие 4 часа клетки окрашивали с помощью специфичных к CD8 и IFN-g антител и анализировали путем проточной цитометрии (Mittrucker и др., Infect. Immun., 70, 199-203 (2002)). Только у иммунизированной с помощью рМО-Raf мыши можно было обнаружить специфичный к Raf антителогенез.
Для обнаружения уничтожающей опухолевые клетки активности иммунизированных и неиммунизированных ВхВ-мышей в возрасте 10, 12 и 14 месяцев умерщвляли и определяли путем взвешивания массу легких. Масса легких является прямым критерием в отношении величины опухоли. В группе, иммунизированной с помощью SL-pMO-Raf, спустя 14 месяцев заметно чаще можно было встретить мышей с уменьшением массы легких, чем в контрольных группах, включая ту группу, которая была иммунизирована "голой" ДНК, кодирующей с-Raf (SL-pCMV-raf). Обычно рост опухоли у необработанных животных является необратимым (Kerkhoff и др., Cell Growth Differ., 11, 185-190 (2000)). Эти данные, таким образом, показывают, что в этом эксперименте вакцинация с помощью SL-pMO-Raf может защищать животных от возникновения опухолей, и описанный согласно настоящему изобретению микроорганизм пригоден в качестве противоопухолевой вакцины.
Эти эксперименты также показывают, что с помощью представленной в настоящем изобретении системы-носителя в принципе можно нарушать самотолерантность и у толерантных в отношении с-Raf животных можно индуцировать специфичный к с-Raf антителогенез и гуморальный иммунный ответ.
С помощью такой экспериментальной системы можно получать в качестве вакцин сальмонеллы, которые экспрессируют изоформы С-Raf (как, например, В-Raf и A-Raf), мутированный С-Raf, B-Raf или A-Raf, эпитопы нормального или мутированного С-Raf, B-Raf или A-Raf, или комбинации эпитопов нормального и/или мутированного C-Raf, B-Raf или A-Raf. Примерами мутации, которая протекает с потерей активности Raf, являются мутации связывающего Ras домена, киназного домена и/или областей фосфорилирования.
Пример 2
Индукция иммунного ответа в случае BALB/c-мышей путем иммунизации с помощью экспрессирующих простатический антиген (PSA) сальмонелл
Наличие специфичных к ткани антигенов, в особенности таких, которые в повышенной мере синтезируются и экспрессируются опухолевыми клетками, является наряду с диагностической пригодностью этих маркеров также возможно применимым фактором для терапевтических целей. В случае карциномы простаты до сих пор идентифицированы три существенных антигена: PSA (специфичный к простате антиген), PSMA (специфичный к простате мембранный антиген) и PSCA (антиген стволовой клетки простаты). В то время как PSA суперэкспрессируется еще в ранних опухолевых формах (Watt и др., Proc. Natl. Acad. Sci. USA, 83, 3166-3170 (1986); Wang и др., Prostate, 2, 89-96 (1981)) и тем самым способствует диагностике карциномы (Labrie и др., J. Urol., 147, 846-851; дискуссия 851-842, (1992)), экспрессия PSCA чаще всего повышена лишь в локально продвинутой, дедифференцированной и метастазированной опухолевой стадии (Gu и др., Oncogene, 19, 1288-1286 (2000); Reiter и др., Proc. Natl. Acad. Sci. USA, 95, 1735-1740 (1998)). Специфичность к органу делает как PSA, так и PSCA потенциальным антигеном-мишенью при разработке иммунных терапий против карциномы простаты (Reiter и др., Proc. Natl. Acad. Sci. USA, 95, 1735-1740 (1998); Hodge и др., Int. J. Cancer, 63, 231-237 (1995); Armbruster, Clin. Chem., 39, 181-195 (1993)).
В этом опыте нужно было показать, могут ли выделяющие PSA сальмонеллы на основе вектора pMOHLY-1 индуцировать у BALB/c-мышей иммунный ответ. Для этого сначала посредством полимеразной цепной реакции (ПЦР) в последовательность кДНК простатического антигена вводили два сайта расщепления NsiI, чтобы сделать возможной вставку "в рамке считывания" амплифицированного фрагмента в вектор-мишень. Для амплификации выбирали фрагмент из 645 пар оснований (bp). В качестве праймеров служили 5'-GTGGATTGGTGATGCATCCCTCATC-3' и 5'-CAGGGCACATGCATCACTGCCCCA-3'. Продукт ПЦР сначала "по тупому концу" клонировали в векторе pUC18 и позднее в сайтах расщепления NsiI лигировали с вектором-мишенью pMOhly-1. Правильность вставки контролировали путем рестрикции и подтверждали путем секвенирования (фиг.2).
С помощью этого штамма сальмонелл иммунизировали BALB/c-мышей назально три раза с интервалом 3 недели с помощью дозы 1·107. Иммунный ответ обнаруживают путем вестерн-блоттинга и внутриклеточного окрашивания цитокинов.
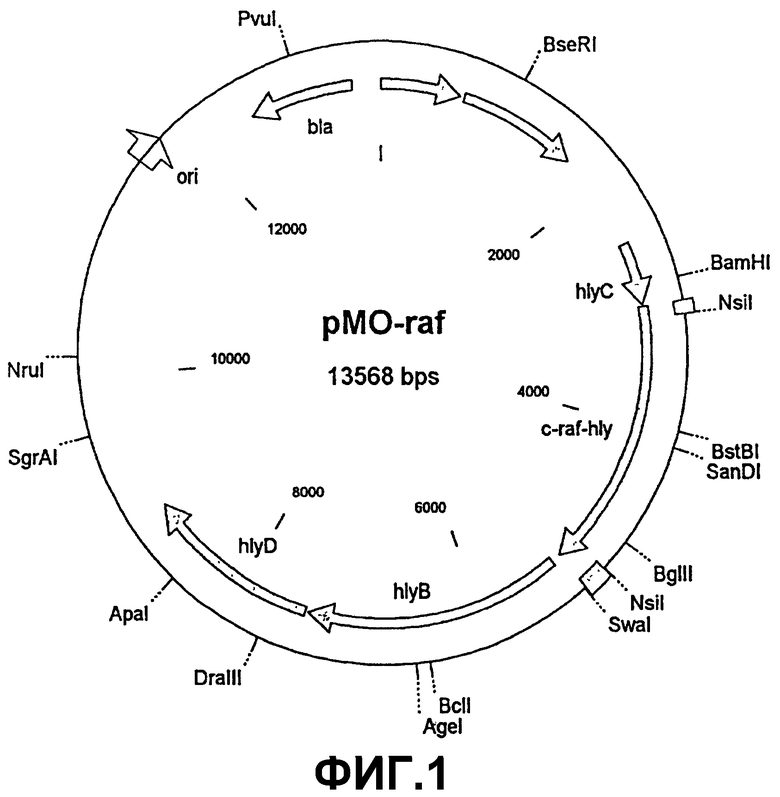
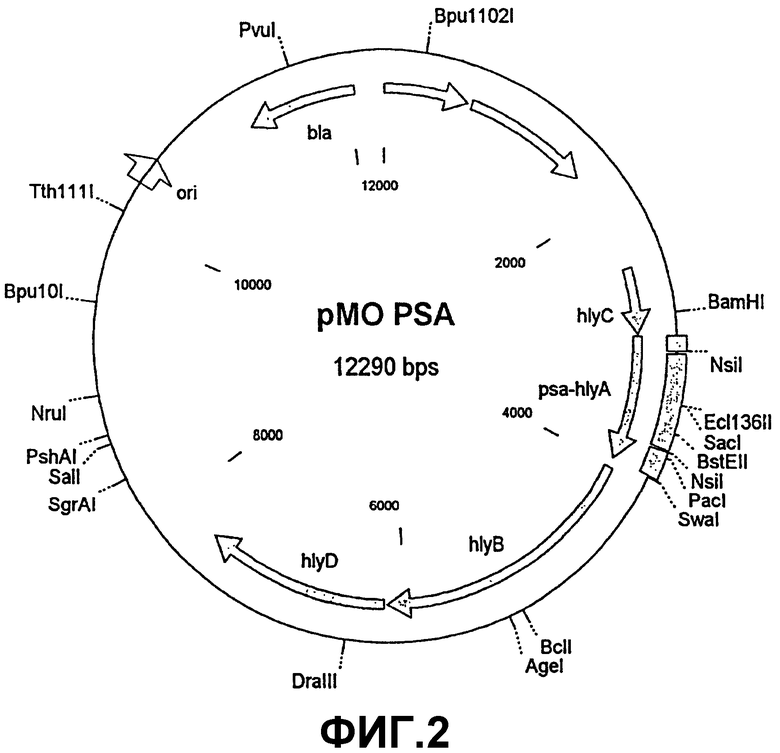
Изобретение относится к генетически модифицированной бактерии, в геном которой введены и экспрессируются следующие компоненты: I) нуклеотидная последовательность, кодирующая по меньшей мере один эпитоп антигена опухолевой клетки, и/или нуклеотидная последовательность по меньшей мере одного эпитопа антигена, специфичного к тканевой клетке, от которой происходит опухоль, II) нуклеотидная последовательность, кодирующая протеин, который стимулирует клетки иммунной системы, IIIA) нуклеотидная последовательность транспортной системы, обеспечивающая экспрессию продукта экспрессии компонентов I) и II) на наружной поверхности бактерии и/или секрецию продукта экспрессии компонента I) и компонента II), и/или IIIB) нуклеотидная последовательность протеина для лизиса бактерии в цитозоле клеток млекопитающих и для внутриклеточного высвобождения плазмид, содержащихся в лизируемых бактериях, и IV) активирующая последовательность, активируемая в бактерии, специфичная к тканевой клетке и неспецифичная к клетке, для экспрессии одного или нескольких компонентов от I) до IIIB), причем каждый из компонентов от I) до IV) может быть представлен однократно или многократно. Данная бактерия пригодна для использования как лекарственное средство в составе противоопухолевой вакцины. Описаны плазмида или вектор экспрессии для получения генетически модифицированной бактерии и способ ее получения. Вакцина на основе генетически модифицированной бактерии является улучшенной в случае профилактики и терапии опухолей с преодоленной иммунотолерантностью по отношению к опухолям. 5 н. и 16 з.п. ф-лы, 2 ил.
I) а) по меньшей мере одну нуклеотидную последовательность, кодирующую по меньшей мере один эпитоп одного антигена клеточного белка или онкогенно мутированного клеточного белка опухолевой клетки и/или
b) по меньшей мере одну нуклеотидную последовательность, кодирующую по меньшей мере один эпитоп одного антигена, специфичного к нормальной тканевой клетке, от которой происходит опухоль,
II) по меньшей мере одну нуклеотидную последовательность, кодирующую белок, стимулирующий клетки иммунной системы и представляющий собой цитокин, интерлейкин, интерферон и/или хемокин,
IIIA) по меньшей мере одну нуклеотидную последовательность, кодирующую транспортную систему, которая обеспечивает экспрессию продукта экспрессии компонентов I) и II) на наружной поверхности бактерии и/или секрецию продукта экспрессии компонентов I) и II) и представляет собой гемолизиновый транспортный сигнал Escherichia coli - нуклеотидные последовательности, содержащие HlyA, HlyB и HlyD под контролем hly-специфичного промотора, гемолизиновый транспортный сигнал Escherichia coli-нуклеотидные последовательности, содержащие HlyA, HlyB и HlyD под контролем не-hly-специфичного бактериального промотора, транспортный сигнал для белка S-ряда (Rsa A) Caulobacter crescentus или транспортный сигнал для TolC-белка Escherichia coli, и/или
IIIB) нуклеотидную последовательность белка для лизиса бактерии в цитозоле клеток млекопитающих и для внутриклеточного высвобождения плазмид, содержащихся в лизируемых бактериях, и
IV) по меньшей мере одну нуклеотидную последовательность активирующей последовательности для экспрессии одного или нескольких компонентов от I) до IIIB), причем указанную активирующую последовательность выбирают из группы, состоящей из активируемой в бактерии, специфичной к тканевой клетке и неспецифичной к клетке активирующей последовательности, причем каждый из компонентов от I) до IV) может быть представлен однократно или многократно, одинаковым или разным.
| WO 9614087 A1, 17.05.1996 | |||
| RU 1438240 C, 20.03.1996 | |||
| RU 1438241 С, 20.03.1996 | |||
| RU 1445193 C, 20.03.1996. |

