Изобретение относится к области биотехнологии, генной инженерии, сельского хозяйства и ветеринарии и представляет собой плазмиду pART27INT6 и способ создания с ее помощью инбредной линии растений Nicotiana tabacum L. - продуцента иммунного гамма-интерферона быка, содержащих вставку гена sIFNG под контролем конститутивного 35S CaMV промотора, обеспечивающую синтез внутриклеточного гамма-интерферона быка.
Гамма-интерферон - сигнальный гликопротеид семейства цитокинов, являющийся продуктом экспрессии активированных Т-лимфоцитов и натуральных киллеров под воздействием соответственно вирусов и маркеров опухолевых клеток, и играющий важную роль в организации иммунного ответа. Воздействие гамма-интерферона на клетки эукариот индуцирует их переход в устойчивое к вирусам состояние, а также активирует системы неспецифичного иммунитета. В качестве фармакологического препарата гамма-интерферон повышает естественную резистентность организма к вирусным инфекциям, а также усиливает иммунный ответ на вакцины. Показано, что гамма-интерферон повышает бактерицидную и фунгицидную активность макрофагов и, таким образом, является эффективным препаратом для лечения различных заболеваний, в том числе смешанной этиологии (вирусной на фоне бактериальной и т.п.).
В настоящее время рекомбинантный гамма-интерферон успешно введен в ветеринарную практику. В животноводстве препарат бычьего гамма-интерферона используют для профилактики туберкулеза и лейкемии крупного рогатого скота. Различают два вида интерферонизации: введение готового интерферона либо веществ-индукторов, стимулирующих процесс его синтеза клетками организма. Интерфероновые препараты имеют широкий спектр действия, обусловленный активацией естественных механизмов защиты организма на ранних стадиях инфекционного процесса. Немаловажно и то, что эффект проявляется сразу же после введения препарата. К сожалению, сохраняется он очень недолгое время (до 12 ч). Долговременное же введение интерферона в больших дозах может приводить к угнетению защитных функций иммунной системы, а также вызывать аллергию. Интерфероновые препараты в терапевтических дозах хорошо переносятся животными, не оказывают местно-раздражающего, а также эмбриотоксического, тератогенного и мутагенного действия. Согласно мнению экспертов Всемирной Организации Здравоохранения цитокиновая терапия поможет решить проблему появления патогенных микроорганизмов, устойчивых к антибиотикам, при терапии заболеваний различной этиологии у сельскохозяйственных животных [1] (Lowenthal J.W. et al. Vet. Immun. Immunopathol. 1999. V. 72. P. 183-188).
Долгое время применение интерфероновых препаратов в ветеринарии ограничивалось дороговизной и низкой эффективностью получения действующего вещества. Наработка интерфероновых препаратов в количествах, достаточных для широкомасштабного применения, стало возможно на базе трансгенных организмов-продуцентов, в геном которых биотехнологическим путем перенесены несвойственные для них гены. Большинство присутствующих на рынке препаратов относятся к этой категории (напр. «Интерферон бычий рекомбинантный» ТУ BY 790624352.002-2010, «Гентаферон-С» ТУ BY 790624352.003-2010). Действующим веществом является рекомбинантный интерферон, выработанный в генетически модифицированных микроорганизмах.
Известны способы получения бычьего гамма-интерферона именно в бактериях [2] (Kashima Т. et al., J. Vet. Med. Sci. 1999, V.61, Р.171-173.) и дрожжах [3-4] (патенты RU 1660388 C, кл. C12N 15/23, 1/19, опубл. 20.05.1995 и RU 2231545 C1, кл. C12N 1/19, 15/23, 15/81 // (C12N 1/19, C12R 1:84), опубл. 27.06.2004). Однако промышленное выращивание микроорганизмов требует достаточно больших затрат на приобретение культиваторов, соблюдение условий стерильности, приготовление питательных сред и т.п. Этого можно избежать, выращивая растения-продуценты. Применение растительных систем в качестве биореакторов для производства рекомбинантных белков в промышленных масштабах имеет существенные перспективы, поскольку являются удобной, безопасной и экономически выгодной альтернативой для продукции различных белков, вакцин и антител по сравнению с системами экспрессии на основе микроорганизмов, культур животных клеток или трансгенных животных [5] (Рукавцева Е.Б. и др. Вопросы биологической, медицинской и фармацевтической химии. 2006. №2. С. 3-12). Растительные организмы автотрофны не требуют дорогостоящих культиваторов для своего выращивания. В отличие от микроорганизмов (бактерий и грибов), а также культур клеток животных, растительные продуценты лишены целого ряда токсинов, антигенов, прионов и вирусов, опасных для потребителей целевых белков - животных и человека. Разработаны различные методы переноса гетерологичных генов в геном растений, наиболее распространенным из которых является метод агробактериальной трансформации. К настоящему времени ведущими биотехнологическими центрами мира модифицировано более 150 видов растений [6] (Дейнеко Е.В. Вестник Томского государственного университета. Биология. 2012, №2(18). С. 41-51). Все это обуславливает актуальность получения растений-продуцентов.
Описанные в заявленном изобретении растения-продуценты табака вырабатывают бычий гамма-интерферон, который можно использовать для производства препаратов в ветеринарии и сельском хозяйстве. Этим заявленное изобретение отличается от других растительных систем экспрессии, в которых осуществляется продукция цитокинов человека: интерферонов [7] (патент RU 2103361 С1, кл. C12N 15/00, 15/22, 15/82, опубл. 27.01.1998) и интерлейкинов [8-9] (патенты RU 2354692 C2, кл. C12N 5/04, опубл. 10.05.2009 и RU 2374321 C2, кл. C12N 15/82, 15/24, опубл. 27.11.2009).
К заявляемому изобретению наиболее близким по решению технической задачи по первому объекту (рекомбинантной плазмиде ДНК) является описанное в [10] изобретение (Патент RU 2231545 С1, МКИ C12N 1/19, 15/23, 15/81; 2002; авторы - Градобоева А.Е., Падкина М.В. и др.); по второму объекту (способу получения на основе инбредной линии растений табака) описанное в [11] изобретение (Патент RU 2302460 С1, МКИ C12N 15/09; C12N 15/24; 2007; авторы - Турчинович А.А. и др.).
Техническим результатом заявленного изобретения является возможность использования на основе полученной рекомбинантной плазмидной ДНК pART27INT6, несущей ген бычьего гамма-интерферона и с ее помощью полученной инбредной линии растений Nicotiana tabacum L., которые синтезируют в тканях иммунный гамма-интерферон быка, в качестве лекарственного средства в ветеринарии и сельском хозяйстве для профилактики и лечения различных заболеваний крупного рогатого скота.
Указанный технический результат в заявленном изобретении достигается тем, что в заявленной рекомбинантной плазмиде ген интерферона быка фланкирован геном октопинсинтазы, а также тем, что ген NPTII фланкирован промотором и терминатором нопалинсинтазы и кроме этого размер плазмиды, что изложено в приведенных ниже примерах апробации.
Трансгенные растения табака используются для синтеза фармакологических препаратов белковой природы [12] (Sharma А.К., Sharma М.К. Biotechnol. Adv. 2009. V. 27. Р. 811-832), в том числе человеческого гамма-интерферона [13] (Leelavathi S., Reddy V.S. Mol. Breeding. 2003. V. 11. P. 49-58).
Выбор табака как объекта трансформации был обусловлен следующими особыми свойствами данного растения: высокая эффективность агробактериальной трансформации; легкая регенерация из каллусов; короткий вегетационный период и в то же время возможность многолетнего выращивания и вегетативного размножения; самоопыление и высокий репродуктивный потенциал (с одного растения можно получить до 10000 семян); большая биомасса растения, высокая скорость роста.
Недостатком табака является высокое содержание токсичных веществ, прежде всего алкалоидов. Это заставляет прибегать к процедуре очистки искомого соединения.
Поставленная техническая задача достигается следующим способом.
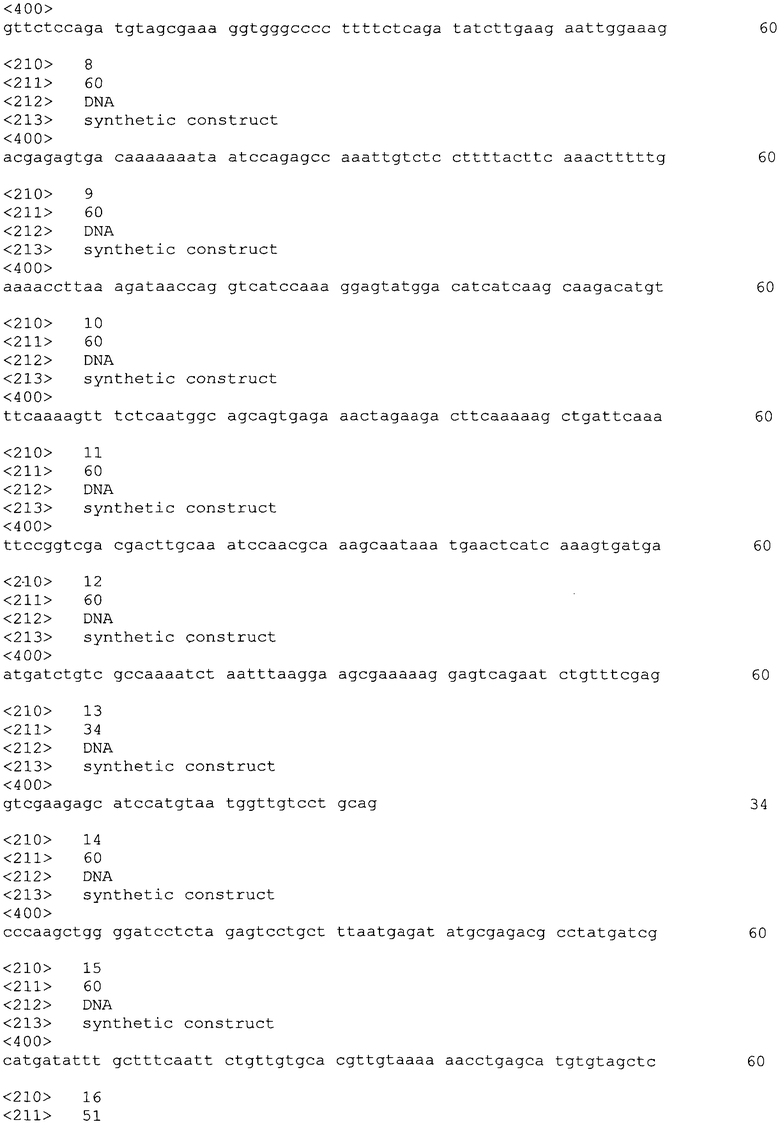
Ген бычьего гамма-интерферона был внедрен в растения методом агробактериальной трансформации с помощью штамма Agrobacterium tumefaciens ЕНА105, содержащего бинарный вектор, состоящий из хелперной (vir-регион) и векторной (pART27) плазмид. Векторная плазмида pART27INT6 получена на основе первичного клонирующего вектора pART7. Общий размер плазмиды pART27INT6 - 13.3 т.п.о. Физическая карта плазмидной ДНК pART27INT6 с указанием генетических маркеров изображена на фиг. 1, где RB и LB - правая и левая границы Т-ДНК области; p35S - промотор гена 35S РНК вируса мозаики цветной капусты; sINFG - гетерологичный ген γ-интерферона быка; OCST - терминатор гена октопинсинтазы из Ti-плазмиды A. tumefaciens; pNOS - промотор гена нопалинсинтазы из Ti-плазмиды Agrobacterium tumefaciens; NPTII - ген устойчивости к канамицину, кодирующий неомицинфосфотрансферазу II; NOST - терминатор гена нопалинсинтазы из Ti-плазмиды A. tumefaciens; OriRK2 - репликон, ответственный за поддержание плазмиды в A. tumefaciens, OriColE1 - репликон; ответственный за обеспечение высококопийности в Escherichia coli; SpR/StrR - ген устойчивости к спектиномицину/стрептомицину в качестве бактериального селективного маркера; SalI, NotI и BamHI - сайты действия соответствующих рестриктаз.
На Фиг. 1 представлена карта плазмидной ДНК pART27INT6, использованной для трансформации растений табака.
После трансформации поколение T0 включало несколько независимых растений-трансформантов, которые подвергли самоопылению. Для получения инбредной линии растений-продуцентов, характеризующейся стабильным наследованием и экспрессией гетерологичного гена sIFNG и синтезирующей бычий гамма-интерферон, растения каждого поколения были подвергнуты тестированию на наличие и активность трансгенной вставки, а также на присутствие в тканях белка бычьего гамма-интерферона. Наличие в геноме растений трансгенной вставки проверялось методом ПНР-анализа. Анализ уровня и стабильности экспрессии гена гамма-интерферона у трансгенных растений табака в ряду поколений был выполнен с помощью двух методов: ОТ-ПЦР-анализа и количественного ПЦР-анализа в реальном времени. С отобранных растений, демонстрировавших положительный результат во всех тестах, в результате самоопыления получали семена, из которых выращивали in vitro no 25 растений следующего поколения. Эти растения снова подвергали анализу и отбору наиболее продуктивных форм, которые пересаживали в почву и путем самоопыления получали семена. В результате были отобраны растения Nicotiana tabacum L. инбредной гомозиготной линии InterS 11.2., которые начиная с поколения T2 стабильно наследуют активную вставку гена sIFNG, не демонстрируя расщепления. Растения с активной трансгенной вставкой исследовали на наличие в тканях белка бычьего гамма-интерферона и его биологическую активность.
В результате пяти поколений селекции была получена гомозиготная инбредная линия растений Nicotiana tabacum L., стабильно наследующая активную вставку гена sIFNG и аккумулирующая в тканях белок бычьего гамма-интерферона. Линия растений-продуцентов характеризуется следующими признаками.
Морфологические признаки
Инбредная линия продуцентов гетерологичного бычьего гамма-интерферона получена на основе растений табака обыкновенного или виргинского (Nicotiana tabacum L.) сорта Трапезунд. Это однолетнее растение высотой до 2,5 м. Листья очередные, широкояйцевидные, цветки в верхушечных соцветиях розовые с белой сердцевиной. Трансгенные растения имеют нормальный фенотип, не отличающийся от растений дикого типа, и размножаются генеративным способом, образуя жизнеспособные семена.
Культуральные признаки
Семена могут прорастать и расти на средах с добавлением антибиотика канамицина в концентрации 100 мг/л.
Физиолого-биохимические признаки
Трансгенные растения несут вставку гена sIFNG, кодирующего бычий гамма-интерферон, под контролем конститутивного промотора 35S CMV вируса мозаики цветной капусты и растительный селективный маркер - ген устойчивости к канамицину, кодирующий неомицинфосфотрансферазу II, находящийся под контролем промотора нопалинсинтазы. Трансформация растительного материала была осуществлена с помощью штамма Agrobacterium tumefaciens EHA105. Штамм ЕНА105 содержит бинарный вектор, состоящий из хелперной (vir-регион) и векторной (pART27INT6) плазмид. Для растений линии Inter311.2 - продуцента гамма-интерферона показан максимальный уровень экспрессии гетерологичного гена и содержание целевого белка в пределах 1-1,5 мкг на 1 г сырой биомассы листьев. Инбредная линия Inter311.2 получена в результате самоопыления трансгенного растения Inter311 из поколения T0. В пределах линии отсутствует расщепление по наследованию гена sIFNG и изменчивость уровня экспрессии, т.е. растения, принадлежащие к семье Inter311.2, гомозиготны и экспрессируют ген sIFNG примерно на одном и том же уровне. Полученная линия растений-продуцентов может использоваться в качестве биореактора для наработки бычьего гамма-интерферона с последующей очисткой для нужд ветеринарии.
Основные отличительные признаки заявляемого способа получения бычьего гамма-интерферона:
- в качестве вектора используется рекомбинантная плазмидная ДНК pART27INT6, в которой целевым геном sIFNG служит нуклеотидная последовательность гамма-интерферона быка. Применение такой последовательности ДНК в качестве целевого гена позволило получить растения - продуценты биологически активного гамма-интерферона с дальнейшим выделением рекомбинантного интерферона в чистом виде из растений методами высаливания или металл-хелатной аффинной хроматографии.
- в качестве растительного материала для трансформации используется табак, обладающий высокой скоростью роста и высоким регенеративным потенциалом, а также значительной биомассой зеленых тканей, что позволяет значительно упростить и удешевить процедуру получения большого количества рекомбинантного белка для использования в ветеринарии.
Способы получения плазмиды pART27INT6 и линии трансгенных растений табака - продуцентов бычьего гамма-интерферона, выделения гетерологичного белка бычьего гамма-интерферона и определение его биологической активности проиллюстрированы следующими примерами.
Пример 1
Ген иммунного интерферона быка получали методом ПЦР-реакции, матрицей для которой служила плазмида pBIG, обеспечивающая продукцию иммунного интерферона быка в клетках дрожжей [10] (Градобоева А.Е., Падкина М.В., Парфенова Л.В. и др. Патент RU 2231545 C1. - МКИ C12N 1/19, 15/23, 15/81. - 2002). Клетки бактерий Escherichia coli, содержащие плазмиду pBIG, выращивали при 37°C в течение ночи в 1 л питательной среды Луриа-Бертани (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия), содержащей ампициллин в концентрации 50 мг/л. Клетки собирали центрифугированием при 5000 об/мин в течение 10 минут при 4°C, суспендировали в 20 мл 25 мМ трис-хлоридного буфера (pH 8,0), содержащего 10 мМ ЭДТА и 50 мМ глюкозы. Плазмидную ДНК из Е. coli выделяли при помощи набора Fermentas GeneJet™ Plasmid DNA isolation kit (Fermentas) согласно инструкциям производителя.
В качестве прямого праймера для амплификации гена sIFNG был использован олигонуклеотид 5′-CCGCTCGAGAAAAGACAGGGCCAATTTT, содержащий сайт для рестриктазы Xho1. В качестве обратного праймера использовали олигонуклеотид 5′-GGAATTCATTACATGGATGCAGAACCCTTCCT, который содержит сайт для рестриктазы EcoR1. ПЦР проводили 60 сек при 94°C, 60 сек при 41°C и 60 сек при 60°C, повторяя этот цикл 30 раз, после чего инкубировали пробу при 72°C 5 мин. Синтезированный ген гамма-интерферона быка размером 0,42 т.п.о. выделяли при помощи электрофореза в 1% агарозном геле. Полосу, соответствующую по размеру продукту интереса, вырезали из геля и выделяли при помощи набора Евроген Cleanup Mini™ (Евроген). Все процедуры проводили в соответствии с приложенными инструкциями.
Полученный ПЦР-продукт обрабатывали рестриктазами Xho1 и EcoRI (Fermentas) в 10 мМ трис-хлоридном буфере (pH 7,5), содержавшем 50 мМ хлористого натрия, 10 мМ хлористого магния и 1 мМ дитиотреитол. К 5 мкг ДНК в объеме 20 мкл добавляли по 5 ед. каждой рестриктазы, после чего пробу инкубировали в течение 25 часов при 37°. Далее проводили лигирование ПЦР-продукта, гидролизованного рестриктазами XhoI/EcoRI с первичным вектором для клонирования pART7 [14] (Gleave A.P. Plant Mol. Biol. 1992. V.20. P.1203-1207), предварительно обработанным этими же рестриктазами. Для этого смешивали ДНК ПЦР-продукта и ДНК вектора в соотношении 1:5 в 10 мкл 70 мМ трис-хлоридного буфера (pH 7,6), содержащего 5 мМ дитиотреитола, 5 мМ хлористого магния, 1 мМ АТФ, с добавлением 10 ед. ДНК-лигазы фага Т4 (Fermentas) и инкубировали при 14°C в течение ночи.
Полученной лигазной смесью трансформировали компетентные клетки Е. coli штамма DH5α (генотип F′/endAl hsdR17 (
Из выросших на ампициллине отдельных клонов трансформантов выделяли плазмидную ДНК при помощи описанной ранее методики. Далее проводили рестрикционный анализ с помощью эндонуклеаз рестрикции XhoI и EcoRI. При рестрикции плазмиды pART7(γ-IFN) и последующем электрофорезе в 1% агарозном геле обнаруживаются фрагменты 4,9 и 0,42 т.п.о. Из выявленного подобным образом клона препаративно выделяли плазмиду pART7(γ-IFN), в которой экспрессионная кассета, содержащая 35S CaMV промотор, ген бычьего гамма-интерферона и терминатор гена октопинсинтазы из Ti-плазмиды A. tumefaciens, фланкирована сайтами рестрикции NotI. С помощью этой рестриктазы (Fermentas) экспрессионная кассета была вырезана из первичного экспрессионного вектора pART7(γ-IFN) и перенесена в бинарный вектор pART27 [14] (Gleave A.P. Plant Mol. Biol. 1992. V. 20. P. 1203-1207). Реакционные смеси помещали в одну пробирку и осуществляли лигирование Т4 лигазой (Fermentas). Все манипуляции проводили согласно прилагающейся инструкции. Обработанную эндонуклеазами рестрикции плазмиду pART27 и фрагмент добавляли в реакционную смесь в молярном соотношении 1:3. Лигазной смесью трансформировали компетентные клетки Е. coli. Трансформантов отбирали на селективной среде с содержанием 50 мг/л стрептомицина.
Из селектированных на канамицине клонов трансформантов выделяли плазмидную ДНК при помощи описанной ранее методики и проводили рестрикционный анализ с помощью эндонуклеазы рестрикции SalI. Клон, у которого электрофорез в 1% агарозном геле выявил фрагменты 9,0 и 4,3 т.п.о., использовали для дальнейшей работы. Плазмидную ДНК pART27IFT6 выделяли из клеток Е. coli при помощи набора Fermentas GeneJet™ Plasmid DNA isolation kit (Fermentas) согласно инструкциям производителя и трансформировали ею компетентные клетки Agrobacterium tumefaciens штамма ЕНА105 (генотип С58 pTiBo542; T-region::aph, Km(S); A281 pEHA101; NPTII).
Пример 2
Выращивание агробактерий и трансформация. Для выращивания агробактерии использовали среды Луриа-Бертани, жидкую и твердую, с добавлением агара 20 г/л, а также селективных агентов - канамицина (50 мг/л), стрептомицина (50 мг/л) и рифампицина (100 мг/л). Культивирование агробактерий осуществляли на чашках Петри, содержащих твердую среду Луриа-Бертани с добавлением необходимых антибиотиков, в темноте при температуре 25°C. Для приготовления ночной культуры агробактерий использовали жидкую среду Луриа-Бертани с добавлением селективных агентов. Культуру выращивали на шейкере в течение 12-15 часов в темноте при температуре 25°C.
Агробактериальную трансформацию табака проводили по стандартной методике трансформации с использованием листовых дисков [15] (Дрейпер Дж. и др. Генная инженерия растений. Мир, 1991. 344 с.). Для проведения трансформации на листовые диски табака наносили надрезы (15-20 на листовой диск), а затем помещали их в ночную культуру агробактерий и культивировали в течение 1-2 часов на шейкере при температуре 18-20°C. После этого листовые диски подсушивали в ламинаре и переносили на среду Мурасиге-Скуга [16] (Murashige Т., Skoog F. Physiol. Plant. 1962. V. 15(3). P. 473-497), покрытую фильтром и содержащую гормоны для индукции каллусообразования (0,5 мг/л НУК и 0,5 мг/л БАП) и селективный агент канамицин (100 мг/л). Через 2 дня экспланты переносили на свежую среду Мурасиге-Скуга (0,5 мг/л НУК + 0,5 мг/л БАП + канамицин 100 мг/л), также содержащую клафоран (300 мг/л) - антибиотик, необходимый для элиминации агробактерий. Каллусообразование наблюдали через 2-3 недели. После получения предположительно трансгенных каллусов их переносили на среду Мурасиге-Скуга с антибиотиками (канамицин 100 мг/л + клафоран 300 мг/л) для индукции регенерации. После того как регенеранты достигали 1,5-2 см, их отделяли от материнского каллуса и помещали в отдельные чашки, содержащие среду Мурасиге-Скуга с антибиотиками (канамицин 100 мг/л + клафоран 300 мг/л). Через 5-7 дней регенеранты укоренялись.
Пример 3
После первичного отбора трансформантов по способности расти в присутствии канамицина, в них проверяли наличие вставки гена sIFNG. Выделение ДНК из растений проводили по стандартной методике с применением цетилтриметиламмоний бромида (ЦТАБ) [17] (Rogers S.O., Bendich A.J. Plant Mol. Biol. 1985. V.5. P.69-76). ПЦР реакцию проводили в объеме 25 мкл с использованием Taq полимеразы и праймеров к гену sIFNG прямой 5′-TAATGCAAGTTCTTCAGATGTAGC и обратный 5′-ACATGGATGCTCTTCGACCT. В результате были получены 6 независимых трансформантов, для которых было показано наличие фрагмента, соответствующего гену γ-интерферона. Возможны ложноположительные результаты, обусловленные тем, что Т-ДНК фрагмент, проникший в клетки, может не выстроиться в геномную ДНК, что приведет к его утрате в ходе полового размножения. Растения-трансформанты, в геном которых произошла успешная интеграция Т-ДНК, с точки зрения классической генетики являются гетерозиготами, т.е. в их потомстве в результате рекомбинации при половом размножении (в случае самоопыления) проявляются три генотипических класса, в том числе утратившие трансгенную вставку. Также возможно встраивание в геном растения нескольких копий вставки, что может привести к активации защитных механизмов растения (сайленсинг), выражающихся в утрате активности гетерологичного гена. Все это приводит к необходимости проведения селекционных мероприятий на протяжении нескольких поколений, заключающихся в анализе потомков растений-трансформантов, полученных в результате самоопыления, методом ПЦР на наличие в гена sIFNG, и отборе особей с положительным результатом для дальнейшего размножения с целью получить линию растений, гомозиготных по гетерологичному гену.
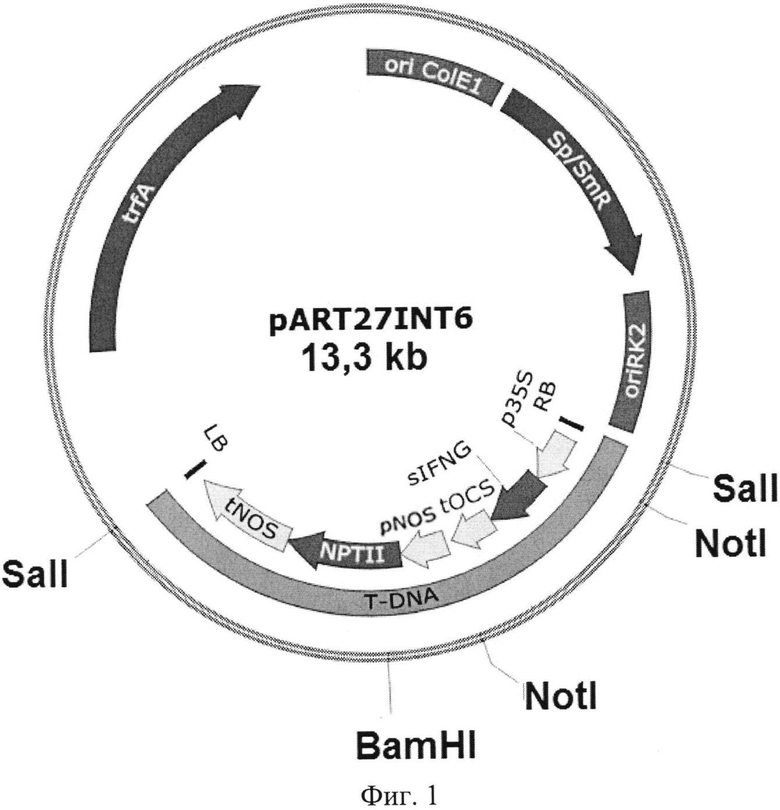
Растения поколения T0 пересаживали в почву, выращивали до зацветания, изолировали соцветия и путем самоопыления получали семена T1. Их проращивали и в свою очередь проверяли на наличие вставки гетерологичного гена. На фиг. 2 представлены результаты ПЦР-анализа тотальной ДНК растений поколения T1 на наличие последовательности sIFNG (электрофореграмма, 1% агарозный гель), где К- - ДНК нетрансгенного растения табака; К+ - положительный контроль (pART27IFNG); 311.2, 311.5, 605.1 - ДНК отобранных трансформантов поколения T1; цифрами подписаны маркеры длины фрагментов ДНК. Наличие фрагментов ДНК, соответствующих гену sIFNG, длиной 360 п.о., подтверждает присутствие в растительном геноме вставки целевого гена.
На Фиг. 2 представлены результаты ПЦР-анализа тотальной ДНК из растений поколения T1 на наличие последовательности sIFNG.
Экспрессию гена sIFNG у трансгенных растений табака выявляли методом обратной транскрипции-ПНР (ОТ-ПЦР) с последующим электрофорезом в агарозном геле. Выделение растительной РНК проводили по стандартной методике с применением реагента PUREZOL. Полученные пробы подвергали ДНКазной обработке с использованием 5 ед. акт. ДНКазы на пробу. После завершения реакции РНК переосаждали 70% этанолом для удаления остатков ДНКазы. Измеряли концентрацию мРНК и ставили реакцию обратной транскрипции с использованием олиго-dT праймера и 1000 ед. акт. M-MuLV обратной транскриптазы на пробу. Для синтеза кДНК брали не менее 2 мкг РНК. Пробы кДНК хранили при -80°C. кДНК анализировали методом ПЦР (как изложено выше) на наличие последовательности, соответствующей гену sIFNG.
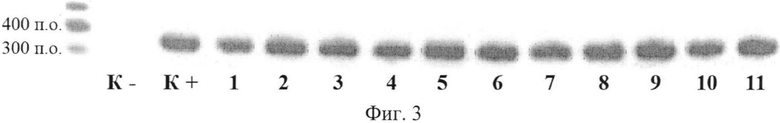
В потомстве 6 первичных трансгенных растений, полученном в результате самоопыления, было обнаружено расщепление по признакам наличия гена sIFNG и его активности. Далее на протяжении четырех поколений в каждой семье проводилась селекция по признаку наличия вставки гена sIFNG и ее активности. В результате селекционных мероприятий удалось получить растения Inter 311.2, гомозиготные по активной форме гена sIFNG, на основе которых создана линия растений-продуцентов гамма-интерферона быка. На фиг. 3 представлены результаты ОТ-ПЦР-анализа кДНК из растений линии Inter 311.2. поколения T2 на наличие последовательности sIFNG (электрофореграмма, 1% агарозный гель), где К- - кДНК из нетрансгенного растения табака; К+ - положительный контроль (pART27IFNG); с 1 по 11 - кДНК растений 311.2.1-311.2.11 - потомков гомозиготного растения 311.2. Наличие фрагментов кДНК, соответствующих гену sIFNG, длиной 360 п.о., подтверждает присутствие в растительном геноме активной вставки целевого гена. Все изученные растения линии 311.2. на протяжении 4 поколений демонстрируют отсутствие расщепления и высокую активность вставки гена sIFNG.
На Фиг. 3 представлены результаты ОТ-ПЦР-анализа кДНК из растений линии Inter 311.2. поколения T2 с праймерами к гену sIFNG.
Пример 4
Растения табака линии Inter 311.2 исследовали на наличие в тканях белка бычьего гамма-интерферона. Белки экстрагировали из ткани молодых листьев, взятых с верхушек растений. Материал гомогенизировали с буфером следующего состава: 0,1 М МЭС, 0,001 М ЭДТА, 1% глицерин, 0,05 М сахароза, 0,01 М дитиотрейтол, 0,011 М аскорбиновая кислота, pH 7,5. Кроме того, в буфер добавляли поперечно-сшитый поливинилпирролидон (0,1% от массы растительной ткани) и ингибитор протеиназ ФМСФ (1 мМ). Полученный гомогенат фильтровали, центрифугировали при 4°C в течение 15 минут при 10000 об/мин и супернатант переносили в новые емкости.
Для выделения из тотального белкового экстракта фракции, содержащей гамма-интерферон, применялся метод высаливания сульфатом аммония. Для получения относительно чистого целевого белка высаливание проводили в два этапа. В предварительных экспериментах установили, что гамма-интерферон осаждается в интервале 40-50% насыщения сульфатом аммония, поэтому вначале концентрацию доводили до 40%, оставляли образец на 1 час на мешалке при 4°C, центрифугировали 15 минут при 10000 об/мин на центрифуге с охлаждением, удаляли осадок, а к супернатанту добавляли навеску сульфата аммония до концентрации 50%, после чего вновь высаливали белки и центрифугировали в тех же условиях и удаляли супернатант.
Содержание гамма-интерферона в тканях оценивали с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия и последующего иммуноблота со специфичными антителами к гамма-интерферону. Для этого осадок, содержащий интерферон, растворяли в 0,0625 М трис-хлоридном буфере (pH 8,6). Разделение белков проводили в 17% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ трис, 192 мМ глицин, 0,1% додецилсульфат натрия, pH 8,6; буфер для геля: 375 мМ трис-хлоридный буфер, pH 8,8). Параллельно проводили разделение белков, полученных из контрольных растений дикого типа, выращенного в идентичных условиях, а также препарата коммерческого рекомбинантного бычьего интерферона бактериального происхождения (R&D systems, Великобритания). По окончании электрофореза белки ренатурировали, выдерживая гели 15 минут в 10 мМ трис-хлоридном буфере (pH 7,5), содержащем 4 М мочевину, 20 мМ ЭДТА, и переносили на нитроцеллюлозную мембрану в 25 мМ трис-190 мМ глициновом буфере (pH 8,3), содержащем 20% метилового спирта, при 10-20 В, в течение 30 мин. Далее мембрану в течение 2 часов выдерживали в 10 мМ трис-хлоридном буфере (TBST, pH 8,0), с добавлением 150 мМ хлористого натрия, 0,05% твин-20 и 1% бычьего сывороточного альбумина. После этого нитроцеллулозную мембрану инкубировали с первичными поликлональными козьими антителами к рекомбинантному γ-интерферону (R&D systems, Великобритания) в течение 2 часов при 37°C. После инкубации проводили три отмывки (по 5 минут каждая) TBST-буфером и инкубировали мембрану с вторичными антителами против иммуноглобулинов козы, меченными пероксидазой хрена (Медгамал, Москва), в течение 1 часа при 37°C. Снова 4 раза промывали TBST-буфером и один раз проявочным буфером (15 мМ фосфатные буфер с добавлением 150 мМ хлористого натрия и 0,05% твин-20, pH 6,0). Разведение первичных антител составило 1:5000, разведение вторичных антител - 1:3000. Проявку мембраны проводили с помощью, добавляя на 50 мл проявочного буфера субстраты для пероксидазы: 0,02% 3,3′-диаминобензидина и 0,006% перекись водорода. Содержание гамма-интерферона быка оценивали, сравнивая интенсивность окрашивания полосы рекомбинантного белка с полосой коммерческого бычьего γ-интерферона бактериального происхождения (R&D systems, Великобритания).
На Фиг. 4 представлены результаты Вестерн-блот гибридизации белков из трансгенных растений линии Inter311.2 (T2) с антителами против бычьего γ-интерферона, где γ-IFN - коммерческий препарат бычьего γ-интерферона. Все трансгенные растения линии синтезируют в среднем от 1 до 1,5 мкг гамма-интерферона быка на 1 г сырого веса. Однако большая биомасса растений табака позволяет получать гетерологичный белок в необходимых количествах.
Пример 5
Противовирусная активность рекомбинантного бычьего гамма-интерферона, синтезированного растениями, была подтверждена в эксперименте на культуре клеток фибробластов из трахеи быка. В результате высаливания белковые экстракты из растений содержали высокую концентрацию сульфата аммония. Для его удаления мы проводили диализ полученной белковой фракции против буфера для экстракции. После диализа полученные пробы использовали для тестирования. Для этого было подготовлено 4 образца: буфер экстракции (супернегативный контроль), экстракт белков из растений табака дикого типа (негативный контроль), экстракт из трансгенных растений линии 311.2.7.2 (поколение T3), а также положительный контроль - рекомбинантный бычий гамма-интерферон бактериального происхождения (R&D systems, Великобритания). Определение специфической активности препаратов интерферона проводили в соответствии с методическими указаниями Государственного научно-исследовательского института стандартизации и контроля медицинских биологических препаратов им. Л.А. Тарасовича, которые устанавливают требования и критерии определения специфической активности препаратов интерферона различного происхождения.
Определение специфической активности препарата интерферона проводили с использованием 2-3 суточного монослоя перевиваемой линии клеток (FBT-фибробласты трахеи плодов коров). Противовирусную активность испытуемого препарата интерферона определяли в сравнении с отраслевым стандартным образцом (ОСО) активности человеческого лейкоцитарного интерферона альфа-2, активность которого составляет 1000 МЕ/мл.
Культуру клеток выращивали на матрацах вместимостью 600 мл (Sarstedt, Германия), используя ростовую питательную среду (среда DMEM с 10% сыворотки крови плодов коров жидкой, с добавлением бензатилбензилпенициллина стерильного 100 ед./мл и стрептомицина сульфата 50 ед./мл). Клетки инкубировали при температуре 37°C в атмосфере, содержащей 5% CO2. После формирования монослоя клетки снимали с подложки смесью р-ров Версена 0,02% и трипсина 0,25% в соотношении 1:1.
Монослой клеток заливали указанной смесью, выдерживали при температуре 37°C 10 мин до набухания и начала отслаивания клеток от подложки, после чего смесь Версена с трипсином сливали, а клетки диспергировали в 10 мл поддерживающей среды (среда DMEM с антибиотиками, без сыворотки плодов коров). В полученной взвеси подсчитывали количество клеток в камере Горяева и доводили концентрацию клеток до 150-300 тысяч в 1 мл путем добавления ростовой питательной среды. Приготовленную таким образом взвесь клеток разливали в 96-луночные культуральные планшеты (Sarstedt, Германия) по 200 мкл в каждую лунку. Инкубировали планшеты с культурой клеток при температуре 37°C в атмосфере, содержащей 5% CO2.
Для определения активности препарата интерферона подготовленные клетки инкубируют с подготовленными пробами испытуемого образца и ОСО активности. Для этого готовили разведения испытуемого образца и ОСО активности в 10 раз с последующим двукратным разведением (титрованием) в поддерживающей среде. Титрование производили с помощью восьмиканальной микропипетки (Fin Pipette, Финляндия) в 96-луночном планшете для иммуноферментного анализа (Медполимер, Россия).
Из лунок культуральных планшетов удаляли ростовую среду и вносили приготовленные разведения испытуемых препаратов и ОСО активности (по 100 мкл), используя на каждое разведение не менее 3 лунок с культурой клеток). Для контроля дозы индикаторного вируса и состояния монослоя клеток оставляли по 3 лунки соответственно. В них вносили по 100 мкл поддерживающей среды, используемой для приготовления разведений испытуемых препаратов. Инокулированные и контрольные культуры клеток инкубировали в течение 24 ч при температуре 37°C в атмосфере, содержащей 5% CO2. Затем в каждую лунку с испытуемым материалом, ОСО и лунки контроля активности индикаторного вируса вносили рассчитанную заранее дозу вируса, соответствующую 100 ТЦД50/0,1 мл (по 100 мкл). В лунки, предназначенные для контроля монослоя, вносили по 100 мкл поддерживающей среды, используемой для приготовления разведении испытуемых препаратов.
В качестве индикатора использовали вирус везикулярного стоматита (ВВС) штамм «Индиана» из коллекции ГИСК им. Л.А. Тарасевича, регистрационный №11/82.
Инокулированные и контрольные культуры клеток инкубировали в течение 18-24 ч при температуре 37°C в атмосфере, содержащей 5% CO2.
Через 18-24 ч инкубации, когда в лунках, содержащих микрокультуры без интерферона (контроль вируса) развивалось тотальное поражение клеточного пласта, среду удаляли, клеточный пласт окрашивали 15 мин 0,1% спиртовым раствором кристаллического фиолетового, после чего планшеты промывали проточной водой и высушивали. Затем проводили учет реакции определения активности. Проверяли качество культуры клеток, просматривая под микроскопом (увеличение 100*) контрольные лунки (К+). Таким же образом проверяли дозу вируса (К-). Лунки должны иметь признаки цитопатического действия (ЦПД) вируса. Титром ИФН считали последнее его разведение, которое вызывало 50% подавление ЦПД ВВС по сравнению с контролем вируса.
Активность испытуемых образцов вычисляли в международных единицах по формуле:
A0=AP*T0/TP
Где A0 - активность исследуемого образца ИФН; AP - активность референс-препарата (ОСО), выраженная в ME (международные единицы); T0 - титр исследуемого ИФН в опыте; TP - титр референс-препарата (ОСО) в опыте.
Интерферон, синтезируемый растениями линии Inter 311.2., обладал активностью против вируса везикулярного стоматита, сопоставимой с таковой у белка, наработанного в трансгенных микроорганизмах (Таблица 1).

Таким образом, как было продемонстрировано в примерах, впервые была получена плазмида pART27INT6, кодирующая внутриклеточный иммунный гамма-интерферон быка в трансгенных растениях, и разработан способ получения инбредной линии растений табака Nicotiana tabacum L., продуцирующего данный иммуномодулятор. Отличительная особенность способа - проведение селекции на протяжении нескольких поколений трансгенных растений, в результате чего была выведена гомозиготная инбредная линия растений Inter 311.2, стабильно наследующая активную вставку гена sIFNG и аккумулирующая в тканях биологически активный белок бычьего гамма-интерферона. Вырабатываемый растениями-продуцентами табака бычий гамма-интерферон можно использовать для профилактики и лечения различных заболеваний крупного рогатого скота в ветеринарии и сельском хозяйстве.
Источники информации
1. Lowenthal J.W. et al. Vet. Immun. Immunopathol. 1999. V. 72. P. 183-188.
2. Kashima T. et al., J. Vet. Med. Sci. 1999, V. 61, P. 171-173.
3. Патент RU №1660388С, кл. C12N 15/23, 1/19, опубл. 20.05.1995.
4. Патент RU №2231545С1, кл. C12N 1/19, 15/23, 15/81, опубл. 27.06.2004.
5. Рукавцева Е.Б. и др. Вопросы биологической, медицинской и фармацевтической химии. 2006. №2. С. 3-12.
6. Дейнеко Е.В. Вестник Томского государственного университета. Биология. 2012, №2 (18). С. 41-51.
7. Патент RU №2103361 С1, кл. C12N 15/00, 15/22, 15/82, опубл. 27.01.1998.
8. Патент RU №2354692С2, кл. C12N 5/04, опубл. 10.05.2009.
9. Патент RU №2374321С2, кл. C12N 15/82, 15/24, опубл. 27.11.2009.
10. Патент RU №2231545 С1, кл. C12N 1/19, 15/23, 15/81; 2002 (прототип для п.1).
11. Патент RU 2302460 С1, МКИ C12N 15/09; C12N 15/24; 2007; авторы - Турчинович А.А. и др. (прототип для п.2).
12. Sharma A.K., Sharma M.K. Biotechnol. Adv. 2009. V. 27. P. 811-832.
13. Leelavathi S., Reddy V.S. Mol. Breeding. 2003. V. 11. P. 49-58.
14. Gleave A.P. Plant Mol. Biol. 1992. V.20. P.1203-1207.
15. Дрейпер Дж. и др. Генная инженерия растений. Мир, 1991. 344 с.
16. Murashige Т., Skoog F. Physiol. Plant. 1962. V. 15 (3). P. 473-497.
17. Rogers S.O., Bendich A.J. Plant Mol. Biol. 1985. V. 5. P. 69-76.



Изобретение относится к области биохимии, в частности к рекомбинантной плазмидной ДНК pART27INT6, которая обеспечивает биосинтез внутриклеточного иммунного гамма-интерферона быка в трансгенных растениях. Также изобретение относится к способу получения на основе вышеуказанной ДНК инбредной линии растений табака, синтезирующей внутриклеточный гамма-интерферон быка. Заявленное изобретение позволит эффективно получать гамма-интерферон быка в трансгенных растениях для применения его в качестве лекарственного препарата в ветеринарии и в сельском хозяйстве. 2 н. и 1 з.п. ф-лы, 4 ил., 1 табл., 5 пр.
1. Рекомбинантная плазмидная ДНК pART27INT6, отличающаяся тем, что она обеспечивает биосинтез внутриклеточного иммунного гамма-интерферона быка в трансгенных растениях, имеет размер 13,3 т.п.о. и состоит из следующих элементов: фрагмент SalI/SalI, соответствующий Т-ДНК области и несущий правую и левую границы, sINFG - гетерологичный ген γ-интерферона быка, фланкированный промотором гена 35S РНК вируса мозаики цветной капусты и терминатором гена октопинсинтазы из Ti-плазмиды Agrobacterium tumefaciens, NPTII - ген устойчивости к канамицину, кодирующий неомицинфосфотрансферазу II, фланкированный промотором и терминатором гена нопалинсинтазы из Ti-плазмиды A. tumefaciens, OriRK2 - репликон, ответственный за поддержание плазмиды в A. tumefaciens, OriColE1 - репликон, ответственный за обеспечение высококопийности в Escherichia coli, и SpR/StrR - ген устойчивости к спектиномицину/стрептомицину в качестве бактериального селективного маркера.
2. Способ получения на основе рекомбинантной плазмидной ДНК pART27INT6 инбредной линии растений табака, синтезирующей внутриклеточный гамма-интерферон быка, включающий конструирование рекомбинантной плазмидной ДНК pART27INT6 по п. 1, перенос полученной плазмидной ДНК в штамм A. tumefaciens, агробактериальную трансформацию растительного материала, отбор трансгенных растений на селективной питательной среде с последующей проверкой отобранных трансформантов на наличие полноразмерной вставки целевого гена sINFG и его экспрессионную активность, селекцию инбредной линии растений табака Nicotiana tabacum L., продуцирующей целевой белок, и проверку биологической активности синтезированного растениями полипептида бычьего гамма-интерферона на культуре клеток фибробластов из трахеи быка, зараженных вирусом везикулярного стоматита.
3. Способ по п. 2, отличающийся тем, что проверку отобранных трансформантов на наследование полноразмерной вставки целевого гена, его экспрессионную активность и биологическую активность продукта осуществляют на протяжении пяти поколений проведением селекционных мероприятий, которые состоят в анализе потомков растений-трансформантов, полученных в результате самоопыления, методом ПЦР на наличие и экспрессию гена sIFNG, и отборе особей с положительным результатом для дальнейшего размножения и получения инбредной линии гомозиготных по целевому гену растений со стабильным наследованием и активностью генной вставки.
| САВЕЛЬЕВА Н.В., Получение трансгенных растений-продуцентов бычьего g-интерферона, Диссертация на соискание ученой степени кандидата биологических наук, Санкт-Петербург, 2009 | |||
| ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS PS105(PBIG)-ПРОДУЦЕНТ ВНУТРИКЛЕТОЧНОГО ИММУННОГО ИНТЕРФЕРОНА БЫКА, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PBIG, ОБЕСПЕЧИВАЮЩАЯ БИОСИНТЕЗ ВНУТРИКЛЕТОЧНОГО ИММУННОГО ИНТЕРФЕРОНА БЫКА И СПОСОБ КОНСТРУИРОВАНИЯ РЕКОМБИНАНТНОЙ ПЛАЗМИДНОЙ ДНК PBIG | 2002 |
|
RU2231545C1 |
| TVOROGOVA V.E | |||
| et al., Creation of adjuvant plants producing bovine gamma-interferon, Adaptation to Climate Change in the Baltic Sea Region: | |||

