Настоящее изобретение конкретно относится к новым рекомбинантным обратным транскриптазам теломеразам, молекулам нуклеиновых кислот, кодирующим их, к клеткам, содержащим указанную молекулу нуклеиновой кислоты, и к применению этих клеток для получения целевого вещества.
В 1965 L.Hayflick сделал открытие, что клетки обладают программируемым моментом гибели. В качестве одного из объяснений старения он предположил, что число раз, которое человеческая клетка может делиться, ограничено (Exp Cell Res. 1965 Mar; 37: 614-36). Теперь известно, что это вызвано укорочением теломеров, когда клетка делится. Хромосомы кэппированы теломерами, состоящими из консервативной, тандемно повторяющейся, некодирующей гексамерной последовательности ДНК, ассоциированной с одно- и двунитевыми связывающими белками. Теломеры ответственны за функции стабильности генома и, в частности, за репликацию концов хромосом. Для успешной концевой репликации хромосом требуется как уникальная структура теломера, так и специализированный фермент обратная транскриптаза теломераза, который представляет собой нуклеопротеин, обладающий ферментативной активностью обратной транскриптазы. Обратная транскриптаза теломераза способна к удлинению ряда повторов теломера, что дает возможность протяженной репликации комплементарной дочерней нити. В клетках, в которых отсутствует обратная транскриптаза теломераза, теломерная ДНК укорачивается при последовательных делениях, поскольку ферменты синтеза ДНК неспособны полностью реплицировать концы хромосом, как только удалена инициирующая РНК затравка. В различных работах сообщены данные о том, что так называемые "теломерные часы" являются важным признаком продолжительности жизни человеческой клетки. Теломерная гипотеза клеточного старения предполагает, что укорочение теломера связано с полным отсутствием активности обратной транскриптазы теломеразы и является пусковым механизмом нестабильности хромосом, приводящей к старению, апоптозу. Теломеразная обратно-транскриптазная активность подлежит понижающей регуляции в соматических клеточных линиях в процессе развития in vivo и в первичных клетках in vitro, что коррелирует с укорочением теломеров. Напротив, повышающая регуляция или нарушение регуляции теломеразной обратно-транскриптазной активности происходит в трансформированных клетках и опухолях. кДНК обратной транскриптазы теломеразы (TERT) из нескольких млекопитающих и одной амфибии клонирована и изучена (Nakamura et al., 1997, Science 277, 955-959; Greenberg et al., 1998, Oncogene 16, 1723-1730).
Поскольку гиперэкспрессия TERT в клетке приводит к иммортализации указанной клетки, предложено использование TERT для получения клеточных линий (McSharry et al., 2001 J Gen Virol. 82, 855-63). Однако активность TERT ограничена видом. Например, человеческая TERT несовместима с птичьим аппаратом сохранения теломеров (Michailidis et al., 2005, Biochem Biophys Res Commun. 335 (1), 240-6). Следовательно, для разработки птичьих клеточных линий и более конкретно клеточных линий семейства утиных существует необходимость в TERT, которая работает в этих конкретных клетках.
Эукариотические клеточные линии являются фундаментальными для изготовления вирусных вакцин и многих продуктов биотехнологии. Биологические вещества, продуцируемые в клеточных культурах, включают ферменты, гормоны, иммунобиологические агенты (моноклональные антитела, интерлейкины, лимфокины) и противораковые агенты. Хотя многие более простые белки можно продуцировать, используя бактериальные клетки, более сложные белки, которые являются гликозилированными, в настоящее время должны быть получены в эукариотических клетках.
Птичьи клеточные линии особенно полезны, поскольку многие вирусы, используемые в фармацевтической композиции, способны реплицироваться на них. Более конкретно, различные вирусы способны расти только на птичьих клетках. Это верно, например, в случае модифицированного вируса вакцинии Анкара (MVA), который неспособен расти на большинстве клеток млекопитающих. Этот поксвирус, который образован из вируса вакцинии в результате более чем 500 пассажей на CEF, использовали в ранних семидесятых годах для вакцинации людей с иммунодефицитом против оспы. В настоящее время MVA, главным образом, используют в качестве вектора для целей генотерапии и иммунотерапии. Например, MVA используют в качестве вектора для гена MUC1 для вакцинации пациентов против опухоли, экспрессирующей данный антиген (Scholl et al., 2003, J Biomed Biotechnol., 2003, 3, 194-201). MVA, несущий ген, кодирующий антигены HPV, также используют в качестве вектора для терапевтического лечения поражений шейки матки высокой степени. В более недавнее время MVA был вектором выбора для изготовления профилактической терапии против вновь возникающих заболеваний или вероятного биологического оружия, такого как вирус лихорадки Западного Нила и сибирской язвы.
Следовательно, существует необходимость в новой обратной транскриптазе теломеразе, способной к иммортализации птичьих клеток и более конкретно клеток птиц семейства утиных.
Как используют на протяжении всей заявки, единственное число существительного используют в том смысле, что оно означает "по меньшей мере один", "по меньшей мере первый", "один или более чем один" или "множество" упоминаемых компонентов или стадий, если контекст явно не диктует противоположное. Например, термин "клетка" включает множество клеток, включая их смеси.
Термин "и/или", где его используют здесь, включает значение "и", "или" и "все комбинации или любая комбинация элементов, объединенных указанным термином".
Как используют здесь, термины "содержащий" и "содержать" предназначены для обозначения того, что продукты, композиции и способы включают упоминаемые компоненты или стадии, но не исключают другие. "Состоящий по существу из" при использовании для определения продуктов, композиций и способов должен означать исключение других компонентов или стадий любой существенной значимости. Таким образом, композиция, состоящая по существу из перечисленных компонентов, не должна включать следовые примеси и фармацевтически приемлемые носители. "Состоящий из" должен означать исключение более чем следовых элементов других компонентов или стадий.
Настоящее изобретение относится к изолированному и/или рекомбинантному полипептиду, содержащему аминокислотную последовательность, которая имеет по меньшей мере 60% идентичности аминокислотной последовательности SEQ ID №:1. В более предпочтительном воплощении изобретения полипептид по изобретению содержит аминокислотную последовательность, которая имеет по меньшей мере 70%, предпочтительно по меньшей мере 80% и даже более предпочтительно по меньшей мере 90% идентичности аминокислотной последовательности SEQ ID №: 1. В более предпочтительном воплощении изобретения полипептид по изобретению содержит аминокислотную последовательность, представленную в SEQ ID №: 1.
В предпочтительном воплощении полипептид по изобретению обладает активностью TERT, и в более предпочтительном воплощении изобретения экспрессия полипептида по изобретению дает возможность иммортализации клетки, принадлежащей к семейству утиных.
Как используют здесь, термин "изолированный" и/или "рекомбинантный" означает, что молекула нуклеиновой кислоты, ДНК, РНК, полипептиды или белки, обозначенные таким образом, получены в такой форме руками человека и, таким образом, выделены из их нативного клеточного окружения in vivo. В результате этого вмешательства человека рекомбинантные ДНК, РНК, полипептиды и белки по изобретению применимы в способах, описанных здесь, тогда как эти ДНК, РНК, полипептиды и белки, как они встречаются в природе, неприменимы.
В другом воплощении настоящее изобретение относится к изолированной молекуле нуклеиновой кислоты, которая кодирует полипептид по изобретению.
В предпочтительном воплощении настоящего изобретения молекула нуклеиновой кислоты, кодирующая полипептид по изобретению, содержит по существу такую же нуклеотидную последовательность, как представлена в SEQ ID №:2. Предпочтительные молекулы нуклеиновой кислоты, кодирующие полипептид по изобретению, содержат такую же нуклеотидную последовательность, как представлена в SEQ ID №:2.
Как используют здесь, термин "по существу такая же нуклеотидная последовательность" относится к молекуле нуклеиновой кислоты, обладающей достаточной идентичностью с упомянутым полинуклеотидом, чтобы она могла гибридизоваться с упомянутым полинуклеотидом в умеренно жестких условиях гибридизации. В одном воплощении молекула нуклеиновой кислоты, имеющая по существу такую же нуклеотидную последовательность, как упомянутая нуклеотидная последовательность, кодирует по существу аминокислотную последовательность, представленную в SEQ ID №:1. В другом воплощении молекула нуклеиновой кислоты, имеющая по существу такую же нуклеотидную последовательность, как упомянутая нуклеотидная последовательность, имеет по меньшей мере 70%, более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95% идентичностью с нуклеиново-кислотной последовательностью, представленной в SEQ ID №:2.
Гибридизация относится к связыванию комплементарных нитей нуклеиновой кислоты (то есть смысловой и антисмысловой нити или зонда и ДНК-мишени) друг с другом посредством водородных связей, подобных связям, которые встречаются в природе в хромосомной ДНК. Уровни жесткости, используемые для гибридизации данного зонда с ДНК-мишенью, может легко варьировать специалист в данной области техники.
Выражение "жесткая гибридизация" используют здесь как относящееся к условиям, при которых полинуклеиново-кислотные гибриды являются стабильными. Как известно специалистам в данной области техники, стабильность гибридов отражена в температуре плавления (Tm) гибридов. Как правило, стабильность гибрида является функцией концентрации ионов натрия и температуры. Типично реакцию гибридизации проводят в условиях более низкой жесткости с последующими отмывками варьирующей, но более высокой жесткости. Ссылка на жесткость гибридизации относится к таким условиям отмывки.
Как используют здесь, выражение "умеренно жесткая гибридизация" относится к условиям, которые позволяют ДНК-мишени связывать комплементарную нуклеиновую кислоту, которая имеет примерно 60% идентичности, предпочтительно примерно 75% идентичности, более предпочтительно примерно 85% идентичности с ДНК-мишенью; причем особенно предпочтительно более 90% идентичности с ДНК-мишенью. Предпочтительно умеренно жесткими условиями являются условия, эквивалентные гибридизации в 50% формамиде, 5*растворе Денхардта, 5*SSPE, 0,2% ДСН при 42°С с последующей отмывкой в 0,2*SSPE, 0,2% ДСН при 65°С.
Молекула нуклеиновой кислоты по изобретению может представлять собой РНК, кДНК или геномную последовательность или иметь смешанный тип. Она может, где пригодно, содержать один или более чем один интрон, где эти интроны имеют нативное, гетерологичное (например, интроны β-глобиновых генов кролика и т.д.) или синтетическое происхождение, в целях повышения экспрессии в клетках-хозяевах.
Настоящее изобретение также относится к вектору, который несет молекулу нуклеиновой кислоты согласно изобретению.
Как используют здесь, термин "вектор" понимают как означающий вектор плазмидного или вирусного происхождения, и возможно такой вектор комбинируют с одним или более чем одним веществом, улучшающим эффективность трансфекции и/или стабильность указанного вектора и/или защиту указанного вектора in vivo против иммунной системы организма-хозяина. Эти вещества широко документированы в литературе, которая доступна специалистам в данной области техники (см., например, Feigner et al., 1987, Proc. West. Pharmacol. Soc. 32, 115-121; Hodgson and Solaiman, 1996, Nature Biotechnology 14, 339-342; Remy et al., 1994, Bioconjugate Chemistry 5, 647-654). Для иллюстрации, но без ограничения, они могут представлять собой полимеры, липиды, в частности катионные липиды, липосомы, ядерные или вирусные белки или нейтральные липиды. Эти вещества можно использовать отдельно или в комбинации. Примеры таких соединений, в частности, доступны в патентных заявках WO 98/08489, WO 98/17693 WO 98/34910, WO 98/37916, WO 98/53853, EP 890362 или WO 99/05183. Комбинация, которая может быть рассмотрена, представляет собой плазмидный рекомбинантный вектор, комбинированный с катионными липидами (DOGS, DC-Chol (3-[N-(N',N'-диметиламиноэтан)карбамоил]-холестерин), спермин-холестерин, спермидин-холестерин и тому подобное) и нейтральными липидами (DOPE).
Выбор плазмид, которые можно использовать в контексте настоящего изобретения, огромен. Они могут представлять собой клонирующие и/или экспрессионные векторы. Как правило, они известны специалистам в данной области техники, и ряд их имеется в продаже, но также возможно сконструировать или модифицировать их методами генной инженерии. Могут быть упомянуты в качестве примеров плазмиды, имеющие происхождение от pBR322 (Gibco BRL), pUC (Gibco BRL), pBluescript (Stratagene), pREP4, pCEP4 (Invitrogene) или p Poly (Lathe et al., 1987, Gene 57, 193-201). Предпочтительно плазмида, используемая в контексте настоящего изобретения, содержит точку начала репликации, обеспечивающую инициацию репликации в продуцирующей клетке и/или клетке-хозяине (например, точка начала репликации ColE1 может быть выбрана для плазмиды, предназначенной для продуцирования в E. coli, а система oriP/EBNA1 может быть выбрана, если для нее желательно самореплицироваться в клетке-хозяине млекопитающего (Lupton and Levine, 1985, Mol. Cell. Biol. 5, 2533-2542; Yates et al., Nature 313, 812-815). Она может содержать дополнительные элементы, улучшающие ее сохранность и/или ее стабильность в данной клетке (последовательность cer, которая стимулирует мономерное сохранение плазмиды (Summers and Sherrat, 1984, Cell 36, 1097-1103), последовательности для интеграции в клеточный геном).
Что касается вирусного вектора, возможно рассмотреть вектор, имеющий происхождение от поксвируса (вируса вакцинии, в частности, MVA, вируса оспы канареек и тому подобного), от аденовируса, от ретровируса, от вируса герпеса, от альфавируса, от обезьяньего пенистого вируса или от аденоассоциированного вируса. Предпочтительно следует использовать нерепликативный вектор. В этом отношении аденовирусные векторы наиболее конкретно пригодны для осуществления настоящего изобретения.
Согласно предпочтительному воплощению изобретения вектор согласно изобретению дополнительно содержит элементы, необходимые для экспрессии молекулы нуклеиновой кислоты по изобретению в клетке-хозяине.
Элементы, необходимые для экспрессии, состоят из группы элементов, дающих возможность транскрипции нуклеотидной последовательности на РНК и трансляции РНК на полипептид, в частности, промоторных последовательностей и/или регуляторных последовательностей, которые эффективны в указанной клетке, и возможно последовательностей, которые необходимы, чтобы дать возможность выделения или экспрессии на поверхности клеток-мишеней для указанного полипептида. Эти элементы могут быть регулируемыми или конститутивными. Конечно, промотор адаптирован к выбранному вектору и к клетке-хозяину. Для примера можно упомянуть эукариотические промоторы генов PGK (фосфоглицераткиназы), МТ (металлотионеина; Mclvor et al., 1987, Mol. Cell Biol. 7, 838-848), oc-1 антитрипсина, CFTR, промоторы гена, кодирующего мышечную креатинкиназу, легочный поверхностно-активный актин, иммуноглобулин или β-актин (Tabin et al., 1982, Mol. Cell Biol. 2, 416-436), SRα (Takebe et al., 1988, Mol. Cell. 8, 466-472), ранний промотор вируса SV40 (обезьяньего вируса), ДКП (длинный концевой повтор) RSV (вируса саркомы Рауса), промотор MPSV, промотор TK-HSV-1, ранний промотор вируса CMV (цитомегаловируса), промоторы вируса вакцинии р7,5K pH5R, pKIL, p28, pi1 и аденовирусные промоторы Е1А и MLP, либо комбинацию указанных промоторов. Ранний промотор цитомегаловируса (CMV) наиболее конкретно предпочтителен.
Согласно предпочтительному воплощению вектор по изобретению дополнительно содержит две последовательности клеточного генома, которые гомологичны участкам последовательности, содержащимся внутри области последовательности ДНК-мишени, нативной для генома. Присутствие указанных гомологичных последовательностей дает возможность сайт-специфической инсерции молекулы нуклеиновой кислоты по изобретению в последовательность ДНК-мишени в результате гомологичной рекомбинации.
Термин "гомологичная рекомбинация" относится к обмену фрагментами ДНК между двумя молекулами ДНК в сайте по существу идентичных нуклеотидных последовательностей. Предпочтительно гомологичные последовательности в векторе на сто процентов гомологичны области последовательности-мишени. Однако можно использовать более низкую гомологию последовательности. Таким образом, можно использовать столь низкую гомологию последовательности, как 80%.
Гомологичные последовательности в векторе включают по меньшей мере 25 п.о., предпочтительны более длинные области, по меньшей мере 500 п.о. и более предпочтительно по меньшей мере 5000 п.о.
Согласно более предпочтительному воплощению изобретения молекула нуклеиновой кислоты окружена гомологичными последовательностями в векторе.
Как используют здесь, "окружена" означает, что одна из гомологичных последовательностей расположена до молекулы нуклеиновой кислоты по изобретению и что одна из гомологичных последовательностей расположена после молекулы нуклеиновой кислоты по изобретению. Как используют здесь, "окружена" не обязательно означает, что две гомологичные последовательности непосредственно сшиты с 3' или с 5' концом молекулы нуклеиновой кислоты по изобретению, и гомологичные последовательности могут быть разделены неограниченным числом нуклеотидов.
Как используют здесь, "последовательность ДНК-мишени" представляет собой область внутри генома клетки, которая является мишенью для модификации путем гомологичной рекомбинации с вектором. Последовательность ДНК-мишени включает структурные гены (то есть последовательности ДНК, кодирующие полипептиды, включающие в случае эукариот интроны и экзоны), регуляторные последовательности, такие как энхансерные последовательности, промоторы и тому подобное, а также другие области внутри интересующего генома. Последовательность ДНК-мишени может также представлять собой последовательность, которая, когда она направлена вектором, не обладает эффектом на функцию генома хозяина.
Как используют здесь, "встроенный в последовательность ДНК-мишени" в широком смысле означает, что процесс гомологичной рекомбинации, который приводит к инсерции молекулы нуклеиновой кислоты по изобретению, вводит делецию или перерыв в последовательность ДНК-мишени.
Специалист в данной области техники способен выбрать подходящие гомологичные последовательности с целью направления специфичной последовательности ДНК в геном клетки. Например, одна гомологичная последовательность может быть гомологична части последовательности ДНК-мишени, тогда как другая гомологичная последовательность гомологична последовательности ДНК, расположенной до или после последовательности-мишени. В соответствии с другим примером одна из гомологичных последовательностей может быть гомологична последовательности ДНК, расположенной до последовательности ДНК-мишени, тогда как другая гомологичная последовательность гомологична последовательности ДНК, расположенной после последовательности ДНК-мишени. В другом примере обе гомологичные последовательности гомологичны последовательностям, расположенным внутри последовательности ДНК-мишени.
Согласно предпочтительному воплощению изобретения последовательность ДНК-мишени представляет собой ген HPRT (гипоксантинфосфорибозилтрансферазы).
Геномная последовательность, содержащая промотор HPRT и ген HPRT Cairina moschata, представлена в SEQ ID №:3. Последовательность, кодирующая старт HPRT при кодоне ATG в положении 8695 нуклеиново-кислотной последовательности, представлена в SEQ ID №:3, последовательность выше кодона ATG представляет собой промоторную последовательность HPRT.
Специалист в данной области техники способен выбрать гомологичные последовательности, необходимые для интеграции молекулы нуклеиновой кислоты по изобретению в ген HPRT. Поскольку между различными членами семейства геномные последовательности, кодирующие HPRT, высокогомологичны среди птиц, специалист в данной области техники, таким образом, способен сконструировать гомологичные последовательности, необходимые для направления на ген HPRT других птичьих клеток.
Согласно более предпочтительному воплощению изобретения гомологичные последовательности адаптированы с целью инсерции молекулы нуклеиновой кислоты по изобретению после промотора HPRT. В данном конкретном воплощении молекула нуклеиновой кислоты по изобретению оперативно сцеплена с эндогенным промотором HPRT клетки. В контексте настоящего изобретения "оперативно сцепленный" следует понимать так, что молекула нуклеиновой кислоты сцеплена с промотором таким образом, чтобы дать возможность ее экспрессии в клетке.
Согласно данному конкретному воплощению гомологичная последовательность выше молекулы нуклеиновой кислоты предпочтительно имеет нуклеиново-кислотную последовательность, которая гомологична по меньшей мере с 500 смежных п.о. и более предпочтительно по меньшей мере с 5000 смежных п.о. нуклеиново-кислотной последовательности, начиная от нуклеотида в положении 1 и заканчивая нуклеотидом в положении 8694 нуклеиново-кислотной последовательности, представленной в SEQ ID №:3, при условии, что указанная гомологичная последовательность не гомологична с нуклеиново-кислотной последовательностью, начиная с нуклеотида в положении 8695 и заканчивая нуклеотидом в положении 26916 нуклеиново-кислотной последовательности, представленной в SEQ ID №:3. Кроме того, эта верхняя (левая) нуклеотидная последовательность предпочтительно непосредственно сшита со стартовым кодоном молекулы нуклеиновой кислоты согласно данному изобретению. Согласно даже более предпочтительному воплощению изобретения гомологичная последовательность выше молекулы нуклеиновой кислоты по изобретению состоит из нуклеиново-кислотной последовательности, начиная от нуклеотида в положении 1383 и заканчивая нуклеотидом в положении 8694 нуклеиново-кислотной последовательности, представленной в SEQ ID №:3. Например, вектор согласно изобретению содержит нуклеиново-кислотную последовательность, начиная от нуклеотида в положении 1 и заканчивая нуклеотидом в положении 11227 нуклеиново-кислотной последовательности, представленной в SEQ ID №:4. Гомологичная последовательность, расположенная после молекулы нуклеиновой кислоты по изобретению, предпочтительно имеет нуклеиново-кислотную последовательность, которая гомологична по меньшей мере с 500 смежных п.о. и более предпочтительно по меньшей мере с 5000 смежных п.о. нуклеиново-кислотной последовательности, начиная от нуклеотида в положении 10581 и заканчивая нуклеотидом в положении 17800 нуклеиново-кислотной последовательности, представленной в SEQ ID №:3. И более предпочтительно указанная гомологичная последовательность, расположенная после молекулы нуклеиновой кислоты по изобретению, состоит из нуклеиново-кислотной последовательности, начиная от нуклеотида в положении 10581 и заканчивая нуклеотидом в положении 17800 нуклеиново-кислотной последовательности, представленной в SEQ ID №:3.
Согласно предпочтительному воплощению вектор по изобретению содержит первый селективный маркер, где этот первый селективный маркер представляет собой маркер положительной селекции, и где указанный первый селективный маркер и молекула нуклеиновой кислоты по изобретению расположены в одном и том же участке вектора, где границы указанного участка определены гомологичными последовательностями.
Как используют здесь, термин "маркер положительной селекции" конкретно относится к гену, кодирующему продукт, который обеспечивает выживание и/или рост только клеток, несущих этот ген, в определенных условиях. Типичные селективные маркеры кодируют белки, которые придают устойчивость к антибиотикам или другим токсинам, например к ампициллину, неомицину, метотрексату или тетрациклину, комплементируют ауксотрофные дефициты или обеспечивают критические питательные вещества, недоступные из комплексной среды. В предпочтительном воплощении согласно изобретению первый селективный маркер кодирует белок, который придает устойчивость к антибиотикам.
Согласно более предпочтительному воплощению изобретения первый селективный маркер в векторе окружен последовательностями, дающими возможность его супрессии. Указанные последовательности, дающие возможность супрессии первого селективного маркера, не окружают молекулу нуклеиновой кислоты по изобретению. Когда вектор является кольцевым, последовательности, дающие возможность супрессии первого селективного маркера, первый селективный маркер и молекула нуклеиновой кислоты по изобретению расположены в одном и том же участке вектора переноса, где границы указанного участка определены гомологичными последовательностями.
Последовательности, дающие возможность супрессии фрагмента нуклеиновой кислоты, хорошо известны специалистам в данной области техники (Nunes-Duby, S. et al. (1998) Nucleic Acids Res. 26:391-406). Эти последовательности могут распознаваться одним или более чем одним специфичным ферментом, который индуцирует супрессию нуклеиновой кислоты, содержащейся между указанными последовательностями, где эти ферменты называют "рекомбиназами". Например, три хорошо известные рекомбиназы, дающие возможность супрессии фрагмента нуклеиновой кислоты, представляют собой рекомбиназы FLP, ISCEI и Cre.
Типичной сайт-специфической рекомбиназой является рекомбиназа Cre. Cre представляет собой 38 кДа продукт гена cre (рекомбинации циклизации) бактериофага Р1 и является сайт-специфической ДНК рекомбиназой семейства Int. Sternberg, N. et al. (1986) J. Mol. Biol. 187: 197-212. Cre распознает 34-п.о. сайт на геноме Р1, называемый loxP (локус X-over P1) и эффективно катализирует реципрокную консервативную рекомбинацию ДНК между парами сайтов loxP. Сайт loxP состоит из двух 13-п.о. инвертированных повторов, фланкирующих 8-п.о. непалиндромный коровый участок. Рекомбинация, опосредованная Cre, между двумя прямыми повторяющимися сайтами loxP приводит в результате к эксцизии ДНК между ними в виде ковалентно замкнутого кольца. Рекомбинация, опосредованная Cre, между парами сайтов loxP в обратной ориентации приведет в результате вероятнее к инверсии расположенной между ними ДНК, чем к эксцизии. Разрыв и соединение ДНК ограничено дискретными положениями внутри корового участка и иногда протекает на нити путем образования временной фосфотирозиновой связи ДНК-белок с ферментом.
Другой сайт-специфической рекомбиназой является I-Scel. Другая эндонуклеаза «хоминга» интронов, например, I-Tlil, I-Ceul, I-Crel, I-Ppol и P1-Pspl, может быть также заменена на l-Scel. Многие перечислены Belfort and Roberts ((1997) Nucleic Acids Research 25:3379-3388). Многие из этих эндонуклеаз имеют происхождение от интронов органелл, в которых использование кодонов отличается от стандартного ядерного использования кодонов. Для использования таких генов для ядерной экспрессии этих эндонуклеаз может быть необходимо изменить кодирующую последовательность, чтобы она совпадала с таковой ядерных генов. I-Scel является двунитевой эндонуклеазой, которая расщепляет ДНК внутри ее сайта распознавания. I-Scel образует 4 п.о. зигзагообразный разрыв с нависающими 3'ОН.
Фермент I-Scel имеет известный сайт распознавания. Сайт распознавания I-Scel представляет собой несимметричную последовательность, которая имеет протяженность 18 п.о.
5' TAGGGATAACAGGGTAAT 3'
3' ATCCCTATTGTCCCATTA 5'
Следовательно, в предпочтительном воплощении изобретения последовательности, дающие возможность супрессии первого селективного маркера, содержат сайт распознавания I-Scel.
Другой сайт-специфической рекомбиназой является рекомбиназа FLP. Рекомбиназа FLP распознает отдельный 34-п.о. минимальный сайт, который выдерживает только ограниченную вырожденность его последовательности распознавания (Jayaram, 1985; Senecoff et al., 1988). Взаимодействие между рекомбиназой FLP и последовательностью FRT изучено (Panigrahi et al, 1992). Примеры вариантов последовательностей FRT приведены Jayaram (1985) и Senecoff et al. (1988), и анализ на FLP-опосредованную рекомбинацию на различных субстратах описан Snaith et al. (1996).
В конкретном воплощении, где вектор по изобретению содержит последовательности, дающие возможность супрессии первого селективного маркера, указанный вектор может предпочтительно содержать первую гомологичную последовательность А и вторую гомологичную последовательность В, где гомологичные последовательности А и В имеют достаточную длину и достаточную гомологию, которые дают возможность гомологичной рекомбинации между ними. В отношении гомологичных последовательностей А и В "достаточная гомология" предпочтительно относится к последовательностям по меньшей мере с 70%, предпочтительно 80%, предпочтительно по меньшей мере 90%, особенно предпочтительно по меньшей мере 95%, весьма предпочтительно по меньшей мере 99%, наиболее предпочтительно со 100% гомологии внутри этих гомологичных последовательностей на протяжении длины по меньшей мере 20 пар оснований, предпочтительно по меньшей мере 50 пар оснований, особенно предпочтительно по меньшей мере 100 пар оснований, весьма предпочтительно по меньшей мере 250 пар оснований, наиболее предпочтительно по меньшей мере 500 пар оснований. В данном воплощении вектор по изобретению содержит в 5'-3'-ориентации по отношению к молекуле нуклеиновой кислоты по изобретению первую гомологичную последовательность А, последовательность, дающую возможность супрессии первого селективного маркера, первый селективный маркер, последовательность, дающую возможность супрессии первого селективного маркера, и гомологичную последовательность В.
Согласно предпочтительному воплощению вектор по изобретению содержит второй селективный маркер, который не окружен указанными гомологичными последовательностями, где указанный второй селективный маркер представляет собой маркер отрицательной селекции. Указанный второй селективный маркер особенно полезен, когда вектор по изобретению является кольцевым. Когда вектор является кольцевым, тот факт, что второй селективный маркер не окружен указанными гомологичными последовательностями, означает, что второй селективный маркер и молекула нуклеиновой кислоты по изобретению не расположены в одном и том же участке вектора переноса, где границы указанного участка определены гомологичными последовательностями.
Согласно предпочтительному воплощению изобретения вектор по изобретению содержит третий селективный маркер, где указанный третий селективный маркер представляет собой маркер отрицательной селекции, и где указанный третий селективный маркер расположен между последовательностями, дающими возможность супрессии первого селективного маркера. Когда вектор является кольцевым, тот факт, что третий селективный маркер расположен между последовательностями, дающими возможность супрессии первого селективного маркера, означает, что третий селективный маркер и первый селективный маркер расположены в одном и том же участке вектора переноса, где границы указанного участка определены последовательностями, дающими возможность супрессии первого селективного маркера.
Как используют здесь, термин "маркер отрицательной селекции" конкретно относится к гену, кодирующему продукт, который уничтожает клетки, которые несут ген, в определенных условиях. Эти гены конкретно включают "суицидный ген". Продукты, кодируемые этими генами, способны трансформировать пролекарство в цитотоксическое соединение. Различные пары суицидный ген/пролекарство в настоящее время доступны. Можно более конкретно упомянуть пары:
- тимидинкиназа вируса простого герпеса типа I (HSV-1 ТК) и ацикловир или ганцикловир (GCV) (Caruso et al., 1993, Proc. Natl. Acad. Sci. USA 90, 7024-7028; Culver et al., 1992, Science 256, 1550-1552; Ram et al., 1997, Nat. Med. 3, 1354-1361);
- цитохром р450 и циклофосфамид (Wei et al., 1994, Human Gene Therapy 5, 969-978);
- нуклеозидфосфорилаза пуринов из Escherichia coli (E.coli) и дезоксирибонуклеозид 6-метилпурин (Sorscher et al., 1994, Gene Therapy 1, 233-238);
- гуанинфосфорибозилтрансфераза из E.coli и 6-тиоксантин (Mzoz and Moolten, 1993, Human Gene Therapy 4, 589-595) и
- цитозиндезаминаза (ЦДаза) и 5-фторцитозин (5FC);
- FCUI и 5-фторцитозин (5FC) (WO 9954481);
- FCUI-8 и 5-фторцитозин (5FC) (WO 2005007857).
Первый, второй и третий селективный маркер можно использовать по отдельности. Например, вектор по изобретению может содержать первый и третий селективный маркер, но не содержать второй, или второй и третий селективный маркер, но не первый.
Согласно предпочтительному воплощению изобретения первый, второй и/или третий селективный маркер помещают под контролем элементов, необходимых для их экспрессии в клетке-хозяине.
Элементы, необходимые для экспрессии, состоят из группы элементов, дающих возможность транскрипции нуклеотидной последовательности на РНК и трансляции РНК на полипептид, в частности, промоторных последовательностей и/или регуляторных последовательностей, которые эффективны в указанной клетке, и возможно последовательностей, которые необходимы, чтобы дать возможность выделения или экспрессии на поверхности клеток-мишеней для указанного полипептида. Эти элементы могут быть регулируемыми или конститутивными. Конечно, промотор адаптирован к выбранному вектору и к клетке-хозяину. Для примера можно упомянуть эукариотические промоторы генов PGK (фосфоглицераткиназы), МТ (металлотионеина; Mclvor et al., 1987, Mol. Cell Biol. 7, 838-848), oc-1 антитрипсина, CFTR, промоторы гена, кодирующего мышечную креатинкиназу, легочный поверхностно-активный актин, иммуноглобулин или β-актин (Tabin et al., 1982, Mol. Cell Biol. 2, 416-436), SRa (Takebe et al., 1988, Mol. Cell. 8, 466-472), ранний промотор вируса SV40 (обезьяньего вируса), ДКП (длинный концевой повтор) RSV (вируса саркомы Рауса), промотор MPSV, промотор TK-HSV-1, ранний промотор вируса CMV (цитомегаловируса), промоторы вируса вакцинии р7,5К pH5R, pKIL, p28, pi1 и аденовирусные промоторы Е1А и MLP, либо комбинацию указанных промоторов. Ранний промотор цитомегаловируса (CMV) наиболее конкретно предпочтителен.
Настоящее изобретение также относится к клетке, трансфицированной молекулой нуклеиновой кислоты или вектором согласно изобретению, и к клеткам, имеющим происхождение от нее. Как используют здесь, термин "имеющие происхождение" относится к клеткам, которые развиваются или дифференцируют из клетки, либо имеют в качестве предка клетку, трансформированную молекулой нуклеиновой кислоты согласно изобретению.
Настоящее изобретение также относится к применению полипептидов, молекул нуклеиновой кислоты и векторов согласно изобретению для иммортализации клетки.
Настоящее изобретение также относится к клетке, содержащей молекулу нуклеиновой кислоты по изобретению, где указанная молекула нуклеиновой кислоты оперативно сцеплена с эндогенным промотором HPRT клетки. "Оперативно сцепленный" следует понимать так, что молекула нуклеиновой кислоты сцеплена с промотором таким образом, чтобы дать возможность ее экспрессии в клетке. В предпочтительном воплощении клетка согласно изобретению содержит нуклеиново-кислотную последовательность, представленную в SEQ ID №:4.
Настоящее изобретение также относится к способу иммортализации клетки, включающему стадию трансфекции вектора согласно изобретению в указанную клетку.
Иммортализованная клетка, как используют здесь, относится к клетке, способной расти в культуре в течение более чем 35 пассажей.
Термин "число пассажей" относится к числу раз, которое клеточная популяция извлечена из культурального сосуда и претерпела процесс субкультивирования (пассажа) с целью поддержания клеток при достаточно низкой плотности, чтобы стимулировать дальнейший рост.
Как используют здесь, термин "трансфицированный" относится к стабильной трансфекции или транзитной трансфекции клетки по изобретению.
Термин "стабильная трансфекция" или "стабильно трансфицированный" относится к введению и интеграции чужеродной ДНК в геном трансфицированной клетки. Термин "стабильный трансфектант" относится к клетке, которая имеет стабильно интегрированную чужеродную ДНК в геномной ДНК.
Термин "транзитная трансфекция" или "транзитно трансфицированный" относится к введению чужеродной ДНК в клетку, где чужеродная ДНК неспособна интегрировать в геном трансфецированной клетки. Чужеродная ДНК пребывает в ядре трансфецированной клетки в течение нескольких суток. Термин "транзитный трансфектант" относится к клеткам, которые захватили чужеродную ДНК, но неспособны интегрировать эту ДНК.
Согласно предпочтительному воплощению изобретения клетка по изобретению имеет происхождение от птичьей клетки и более предпочтительно от клетки семейства Anatidae или семейства Phasianidae. Среди Anatidae особенно предпочтительны клетки, принадлежащие к родам Cairina или Anas. Даже более предпочтительно клетки по изобретению принадлежат виду Cairina moschata или Anas platyrhynchos.
Предпочтительно клетки по изобретению имеют происхождение от эмбрионального организма. Способы, дающие возможность выделения клеток из живого организма, хорошо известны специалистам в данной области техники. Например, можно использовать способы, раскрытые в примере 2. Согласно предпочтительному воплощению изобретения первичную клетку изолируют из эмбриона, принадлежащего к семейству Anatidae, который находится в возрасте между 0 и 20 сутками, более предпочтительно между 5 и 15 сутками и даже более предпочтительно между 11 и 14 сутками.
Когда вектор, используемый в способе по изобретению, содержит первый селективный маркер, интеграция первого селективного маркера дает возможность селекции клеток, в которые включена молекула нуклеиновой кислоты по изобретению. Соответственно, способ согласно изобретению может дополнительно включать стадию, где указанные клетки культивируют в среде, которая позволяет расти только клеткам, которые включили первый селективный маркер, например, в среде, которая содержит антибиотик.
Когда вектор, используемый в способе по изобретению, содержит последовательности, дающие возможность супрессии первого селективного маркера, способ согласно изобретению может дополнительно включать стадию, состоящую в супрессии первого селективного маркера из генома указанной первичной клетки. В целях супрессии указанного первого селективного маркера клетку трансфецируют геном, кодирующим рекомбиназу, специфичную для последовательностей, дающих возможность супрессии первого селективного маркера. Способы и вектор, способные переносить указанный ген в клетку, хорошо известны специалистам в данной области техники, например, можно использовать способ, раскрытый в примере 4 настоящей заявки.
Когда вектор, используемый в способе по изобретению, содержит второй селективный маркер, способ согласно изобретению может дополнительно включать стадию, где клетки культивируют в среде, которая позволяет расти только клеткам, которые не включили второй селективный маркер. Указанную стадию можно проводить одновременно или отдельно от стадии, где указанные клетки культивируют в среде, которая позволяет расти только клеткам, которые включили первый селективный маркер.
Указанный второй селективный маркер особенно полезен, когда вектор, используемый в способе согласно изобретению, является кольцевым. Присутствие указанного второго селективного маркера позволяет разрушить клетки, в которых процесс гомологичной рекомбинации привел к введению участка вектора переноса, который не содержит молекулу нуклеиновой кислоты по изобретению.
Когда вектор, используемый в способе по изобретению, содержит третий селективный маркер, способ согласно изобретению может дополнительно включать стадию, где клетки культивируют в среде, которая не позволяет расти клеткам, содержащим третий селективный маркер. Например, среда, которая не позволяет расти клеткам, содержащим FCU1 в качестве третьего селективного маркера, содержит 5-фторцитозин.
Эта стадия дает возможность селекции клеток, в которых произошла супрессия первого селективного маркера. Это означает, что стадия, состоящая в супрессии первого селективного маркера, также приведет к супрессии третьего селективного маркера. Присутствие третьего селективного маркера позволяет разрушить клетки, в которых присутствует первый селективный маркер.
Настоящее изобретение более конкретно относится, но не ограничено им, к способу иммортализации клетки, включающему стадии:
- переноса в клетку вектора, содержащего:
- - молекулу нуклеиновой кислоты согласно изобретению, окруженную гомологичными последовательностями,
- - первый селективный маркер, где указанный первый селективный маркер представляет собой маркер положительной селекции, и где указанный первый селективный маркер окружен указанными гомологичными последовательностями,
- - последовательности, дающие возможность супрессии первого селективного маркера,
- - второй селективный маркер, который не окружен указанными гомологичными последовательностями, где указанный селективный маркер представляет собой маркер отрицательной селекции,
- - третий селективный маркер, где указанный третий селективный маркер представляет собой маркер отрицательной селекции, и где указанный третий селективный маркер расположен между последовательностями, дающими возможность супрессии первого селективного маркера;
- культивирования указанных клеток в среде, которая позволяет расти только клеткам, которые включили первый селективный маркер,
- культивирования указанных клеток в среде, которая не позволяет расти клеткам, которые включили второй селективный маркер,
- исключения первого селективного маркера из генома указанной клетки,
- культивирования указанных клеток в среде, которая не позволяет расти клеткам, которые содержат третий селективный маркер.
В особенно предпочтительном воплощении изобретение относится к иммортализованной клетке, которая имеет происхождение от клетки животного, принадлежащего к виду Cairina moschata, и которая содержит обратную транскриптазу теломеразу Cairina moschata под контролем промотора HPRT Cairina moschata, встроенную в ген HPRT клетки.
Клетка согласно изобретению может дополнительно содержать одну или более чем одну нуклеиново-кислотную последовательность, дающую возможность размножения дефектного вируса. "Дефектный вирус" относится к вирусу, в котором один или более чем один вирусный ген, необходимый для его репликации, делегирован или сделан нефункциональным. Термин "нуклеиново-кислотная последовательность, дающая возможность размножения дефектного вируса" относится к нуклеиново-кислотной последовательности, обеспечивающей в транс-положении функцию(и), которая позволяет реплицироваться дефектному вирусу. Иными словами, указанная нуклеиново-кислотная последовательность(и) кодирует белок(белки), необходимый для репликации и капсидирования указанного дефектного вируса. В качестве иллюстрации, для получения аденовирусного вектора, в котором отсутствует большая часть области Е1, клетку согласно изобретению можно трансфецировать транзитно или перманентно нуклеиново-кислотной последовательностью, кодирующей область Е1.
Клетка согласно изобретению может также содержать нуклеиново-кислотную последовательность, кодирующую целевое вещество. Как используют здесь, целевое вещество может включать, но не ограничено ими, фармацевтически активный белок, например факторы роста, регуляторы роста, антитела, антигены, их производные, полезные для иммунизации или вакцинации, и тому подобное, интерлейкины, инсулин, G-CSF, GM-CSF, hPG-CSF, M-CSF или их комбинации, интерфероны, например интерферон-α, интерферон-β, интерферон-γ, факторы свертывания крови, например фактор VIII, фактор IX или tPA или их комбинации. "Целевое вещество" также относится к промышленным ферментам, например, для применения при модификации целлюлозно-бумажных, текстильных изделий или при производстве этанола. Наконец, "целевое вещество" также относится к белковой добавке или к продукту с расширенными функциями для корма животных.
Клетки, полученные способом согласно изобретению, клетка по изобретению и клетки, имеющие происхождение от нее, особенно полезны для репликации вируса. Указанные вирусы могут быть живыми, аттенуированными, рекомбинантными или нерекомбинантными. Более предпочтительно указанные клетки особенно полезны для репликации поксвируса (вируса вакцинии, в частности MVA, вируса оспы канареек и т.д.), аденовируса, ретровируса, вируса герпеса, альфавируса, пенистого вируса или аденоассоциированного вируса.
Ретровирусы обладают свойством инфицирования, и в большинстве случаев интеграции в делящиеся клетки, и в этом отношении они особенно пригодны для применения в связи с раком. Рекомбинантный ретровирус согласно изобретению, как правило, содержит последовательности ДКП, область капсидирования и нуклеотидную последовательность согласно изобретению, которая помещена под контроль ретровирусного ДКП или внутреннего промотора, такого как описано ниже. Ретровирусный вектор может содержать модификации, в частности, в ДКП (замещение промоторной области эукариотическим промотором) или в области капсидирования (замещение гетерологичной областью капсидирования, например, типа VL30) (см. французские заявки №№9408300 и 9705203).
В аденовирусном векторе может полностью или частично отсутствовать по меньшей мере одна область, которая существенна для репликации и которая выбрана из областей Е1, Е2, Е4 и L1 L5. Предпочтительна делеция области Е1. Однако ее можно комбинировать с другой (другими) модификацией(ями)/делецией(ями), захватывающей, в частности, полностью или частично области Е2, Е4 и/или L1 L5. В качестве иллюстрации делеция большей части области Е1 и транскрипционной единицы Е4 обладает особыми преимуществами. В целях повышения емкости клонирования в аденовирусе может дополнительно полностью или частично отсутствовать несущественная область Е3. В соответствии с другой альтернативой возможно найти применение минимального аденовирусного вектора, в котором сохранены последовательности, существенные для капсидирования, а именно 5' и 3' ИКП (инвертированный концевой повтор) и область капсидирования. Различные аденовирусные векторы и методики их получения известны (см., например, Graham and Prevect, 1991, в Methods in Molecular Biology, Vol 7, p 109 128; Ed: E.J.Murey, The Human Press Inc.).
Семейство поксвирусов включает подсемейства вирусов хондропоксвирусы и энтомопоксвирусы. Среди них поксвирус по изобретению предпочтительно выбран из группы, включающей ортопоксвирусы, парапоксвирусы, авипоксвирусы, каприпоксвирусы, лепорипоксвирусы, вирусы оспы свиней, вирусы контагиозного моллюска, вирусы опухоли обезьян. Согласно более предпочтительному воплощению поксвирус по изобретению представляет собой ортопоксвирус.
Ортопоксвирус предпочтительно представляет собой вирус вакцинии и более предпочтительно модифицированный вирус вакцинии Анкара (MVA), в частности MVA 575 (ЕСАСС V00120707) и MVA-BN (ECACC V00083008).
Термин "рекомбинантный вирус" относится к вирусу, содержащему эндогенную последовательность, встроенную в его геном. Как используют здесь, экзогенная последовательность относится к нуклеиновой кислоте, которая не присутствует в природе в исходном вирусе.
В одном воплощении экзогенная последовательность кодирует молекулу, обладающую прямой или косвенной цитотоксической функцией. Под "прямой или косвенной" цитотоксической авторы изобретения подразумевают, что молекула, кодируемая экзогенной последовательностью, сама может быть токсичной (например, рицин, фактор некроза опухоли, интерлейкин-2, интерферон-гамма, рибонуклеаза, дезоксирибонуклеаза, экзотоксин A Pseudomonas), либо может претерпевать метаболизм с образованием токсичного продукта, либо может действовать на что-либо еще с образованием токсичного продукта. Последовательность кДНК рицина раскрыта в статье Lamb et al. (Eur. J. Biochem., 1985, 148, 265-270), включенной здесь путем ссылки.
В предпочтительном воплощении изобретения экзогенная последовательность представляет собой суицидный ген. Суицидный ген кодирует белок, способный превращать относительно нетоксичное пролекарство в токсичное лекарство. Например, фермент цитозиндезаминаза превращает 5-фторцитозин (5FC) в 5-фторурацил (5FU) (Mullen et al. (1922) PNAS 89, 33); фермент тимидинкиназа вируса герпеса сенсибилизирует клетки к обработке антивирусным агентом ганцикловиром (GCV) или ацикловиром (Moolten (1986) Cancer Res. 46, 5276; Ezzedine et al. (1991) New Biol 3, 608). Можно использовать цитозиндезаминазу любого организма, например Е. coli или Saccharomyces cerevisiae.
Таким образом, в более предпочтительном воплощении изобретения ген кодирует белок, обладающий активностью цитозиндезаминазы, и даже более предпочтительно белок, который описан в патентных заявках WO 2005007857 и WO 9954481.
В следующем воплощении экзогенный ген кодирует рибозим, способный расщеплять РНК- или ДНК-мишень. Расщепляемая РНК- или ДНК-мишень может представлять собой РНК или ДНК, которая существенна для функции клетки, и ее расщепление приводит в результате к клеточной гибели, либо расщепляемая РНК или ДНК может представлять собой РНК или ДНК, которая кодирует нежелательный белок, например онкогенный продукт, и расщепление этой РНК или ДНК может предотвратить становление клетки канцерогенной.
Еще в одном следующем воплощении экзогенный ген кодирует антисмысловую РНК.
Под "антисмысловой РНК" авторы изобретения подразумевают молекулу РНК, которая гибридизуется с молекулой мРНК, кодирующей белок, или с другой молекулой РНК внутри клетки, такой как пре-мРНК, либо тРНК, либо рРНК, и препятствует ее экспрессии, либо гибридизуется с геном и препятствует экспрессии гена.
В другом воплощении изобретения экзогенная последовательность замещает функцию дефектного гена в клетке-мишени. Существует несколько тысяч наследственных генетических заболеваний млекопитающих, включая людей, которые вызваны дефектными генами. Примеры таких генетических заболеваний включают муковисцидоз, где известно, что он вызван мутацией в гене CFTR; мышечную дистрофию Дюшенна, где известно, что она вызвана мутацией в гене дистрофина; серповидно-клеточную анемию, где известно, что она вызвана мутацией в гене HbA. Многие типы рака вызваны дефектными генами, в частности протоонкогенами, и генами опухолевых супрессоров, которые претерпели мутацию.
Примерами протоонкогенов являются ras, src, bcl и т.д.; примерами генов опухолевых супрессоров являются р53 и Rb.
В следующем воплощении изобретения экзогенная последовательность кодирует опухолевый антиген (ТАА, tumor associated antigen). TAA относится к молекуле, которая обнаруживается с более высокой частотой или плотностью в опухолевых клетках, чем в неопухолевых клетках того же типа ткани. Примеры ТАА включают, но не ограничены ими, СЕА, MART-1, MAGE-1, MAGE-3, GP-100, MUC-1, MUC-2, онкоген ras с точечной мутацией, нормальный р53 или с точечной мутацией, гиперэкспрессируемый р53, СА-125, PSA, C-erb/B2, BRCA I, BRCA II, PSMA, тирозиназу, TRP-1, TRP-2, NY-ESO-1, TAG72, KSA, HER-2/neu, bcr-abl, рах3-fkhr, ews-fii-1, выживаемость и LRP. Согласно более предпочтительному воплощению ТАА представляет собой MUC1.
Рекомбинантный вирус может содержать более чем одну экзогенную последовательность, и каждая экзогенная последовательность может кодировать более чем одну молекулу. Например, может быть полезным сочетание более чем в одном рекомбинантном поксвирусе экзогенной последовательности, кодирующей ТАА, с экзогенной последовательностью, кодирующей цитокин.
В другом воплощении изобретения экзогенный ген кодирует антиген. Как используют здесь, "антиген" относится к лиганду, который может связываться антителом; сам антиген необязательно может быть иммуногенным.
Предпочтительно антиген имеет происхождение от вируса, такого как, например, HIV-1 (такой как gp 120 или gp 160), любой вирус иммунодефицита кошачьих, вирусы герпеса человека и животных, такие как gD или его производные, или немедленно-ранний белок, такой как ICP27, от HSV1 или HSV2, цитомегаловирус (такой как gB или его производные), вирус ветряной оспы, вирус опоясывающего лишая (такой как gpI, II или III), либо от вируса гепатита, такого как вирус гепатита В, например, поверхностный антиген вируса гепатита В или его производное, вирус гепатита А, вирус гепатита С (предпочтительно не структурный белок от штамма ja генотипа Ib) и вирус гепатита Е, либо от других вирусных патогенов, таких как респираторно-синцитиальный вирус, вирус папилломы человека (предпочтительно белок Е6 и Е7 от штамма HPV16) или вирус гриппа, либо имеет происхождение от бактериальных патогенов, таких как Salmonella, Neisseria, Borrelia (например, OspA или OspB или их производные), либо Chlamydia, либо Bordetella, например, Р.69, РТ и FHA, либо имеет происхождение от паразитов, таких как Plasmodium или Toxoplasma.
Фиг.1: Вектор, содержащий ген, кодирующий обратную транскриптазу теломеразу.
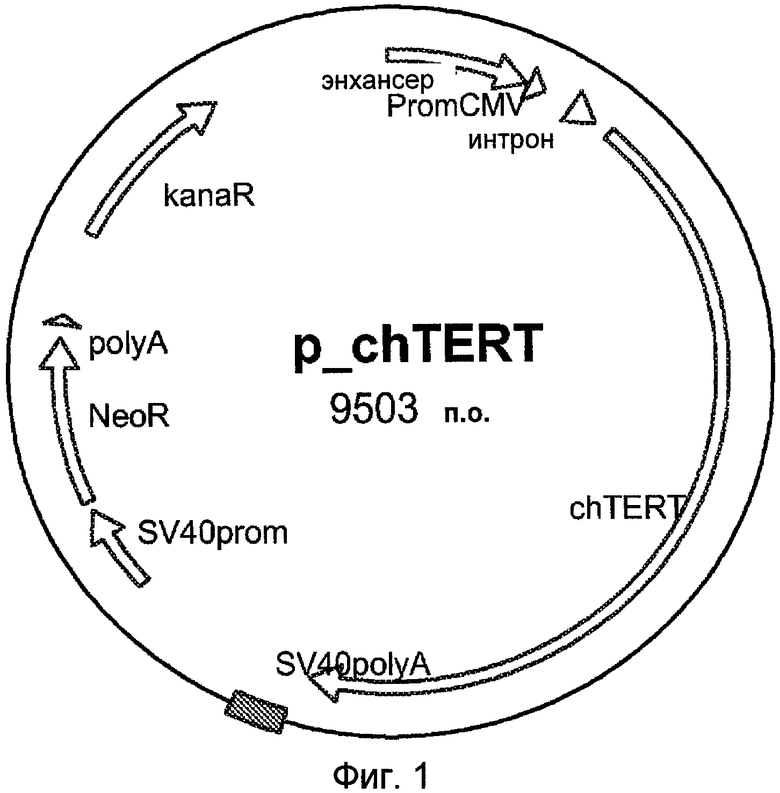
Фиг.2: Схематическое изображение сайт-специфической инсерции гена, кодирующего обратную транскриптазу теломеразу, в ген HPRT.
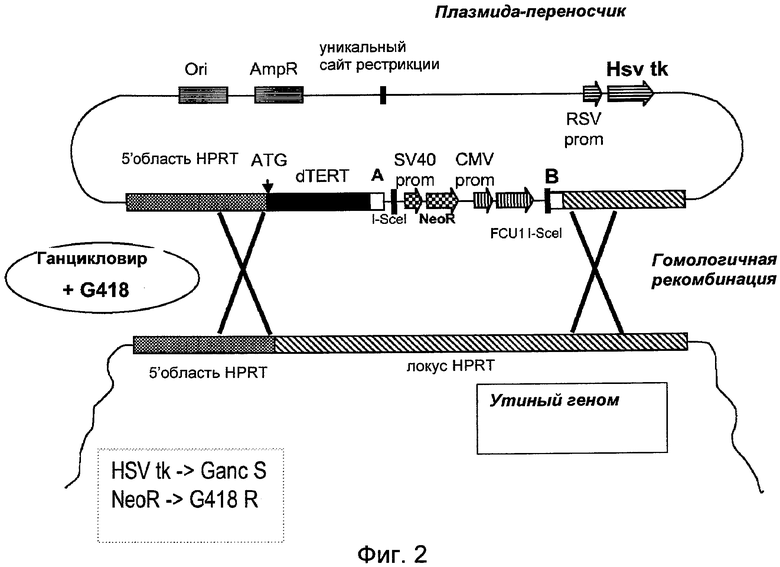
Фиг.3: Схематическое изображение элиминации первого и третьего селективного маркера из генома иммортализованной клетки, полученной способом по изобретению.
Фиг.4: Морфология иммортализованных клеток Т3-17490 (визуализация под световым микроскопом).
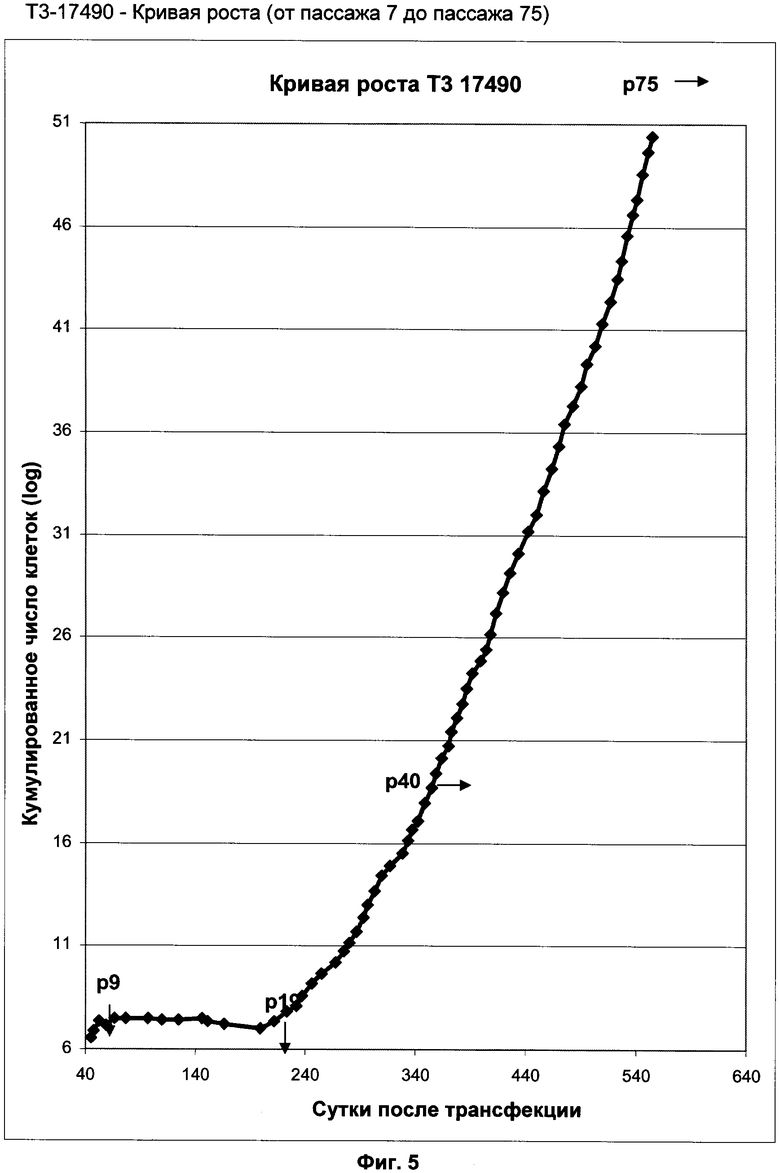
Фиг.5: Кривая роста клеточной линии Т3-17490 (от пассажа 7 до пассажа 75).
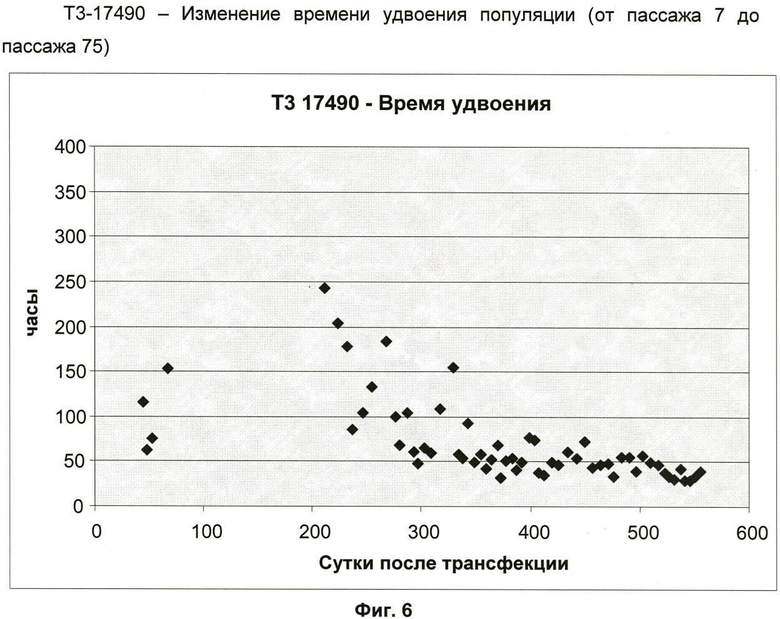
Фиг.6: Изменение времени удвоения популяции клеточной линии Т3-17490 (от пассажа 7 до пассажа 75).
Фиг.7: Морфология иммортализованных клеток Т6-17490 (визуализация под световым микроскопом).
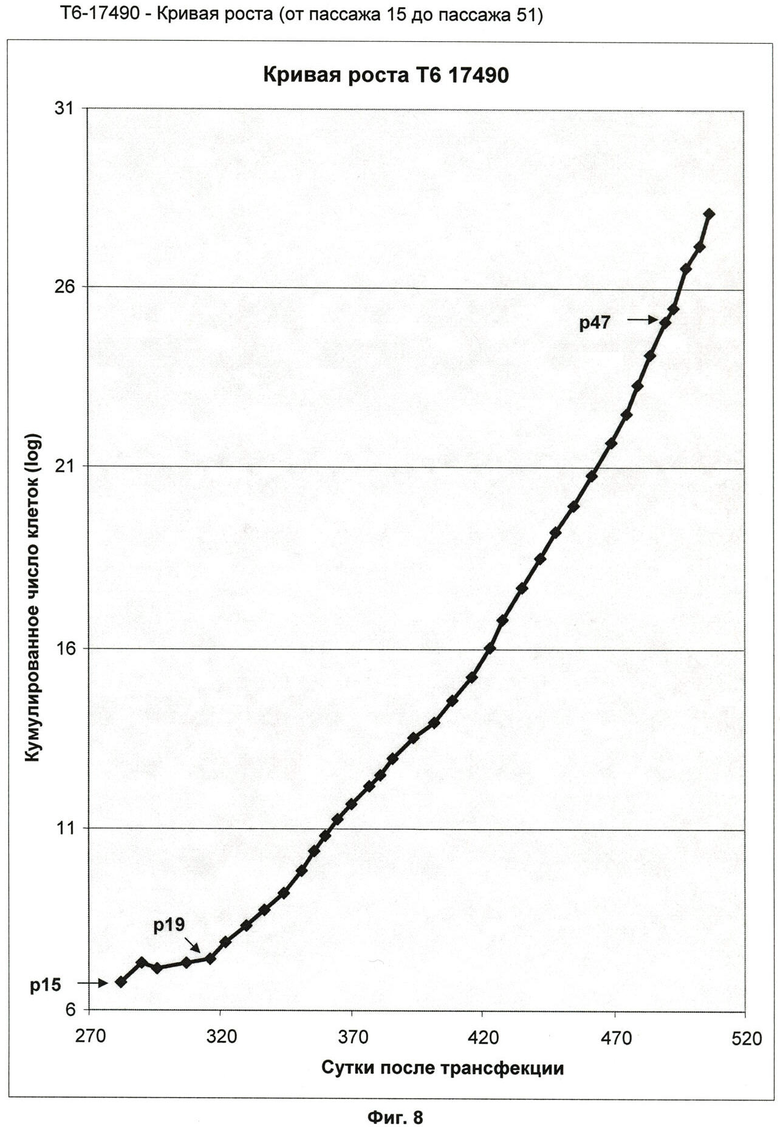
Фиг.8: Кривая роста клеточной линии Т6-17490 (от пассажа 15 до пассажа 51).
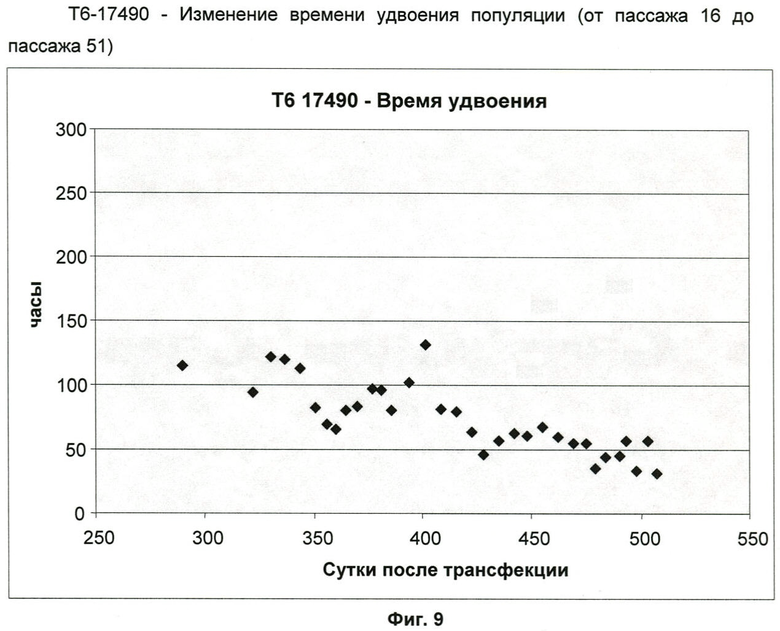
Фиг.9: Изменение времени удвоения популяции клеточной линии Т6-17490 (от пассажа 36 до пассажа 51).
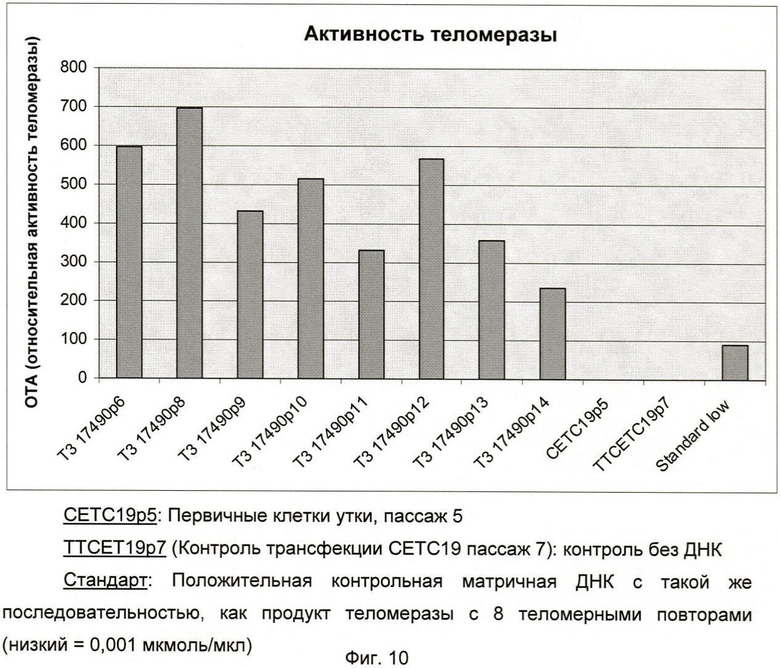
Фиг.10: Активность TERT в различных клеточных линиях.
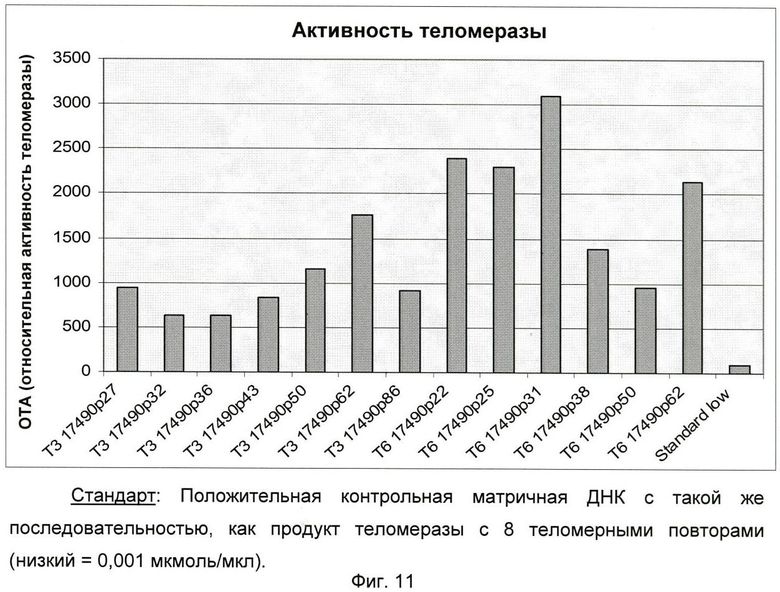
Фиг.11: Активность TERT в различных клеточных линиях.
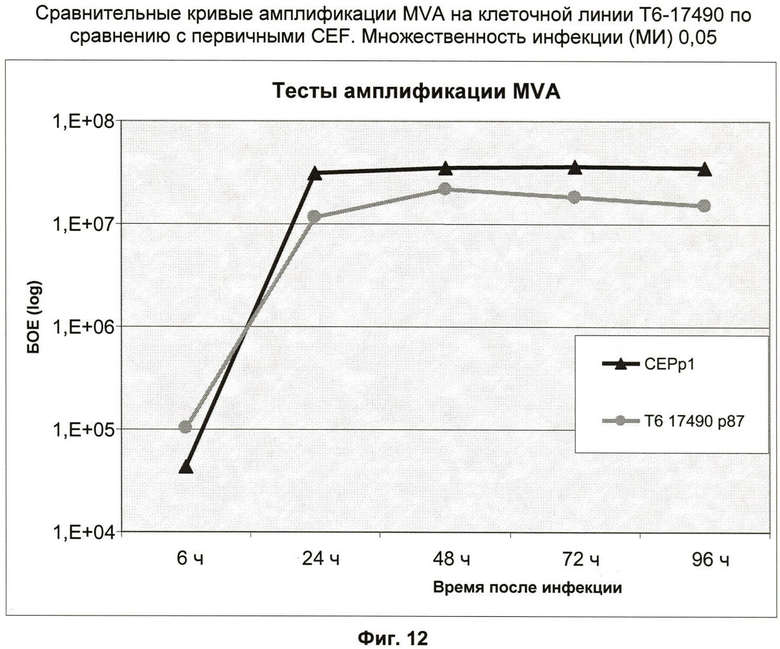
Фиг.12: Амплификации MVA на клеточной линии Т6-17490.
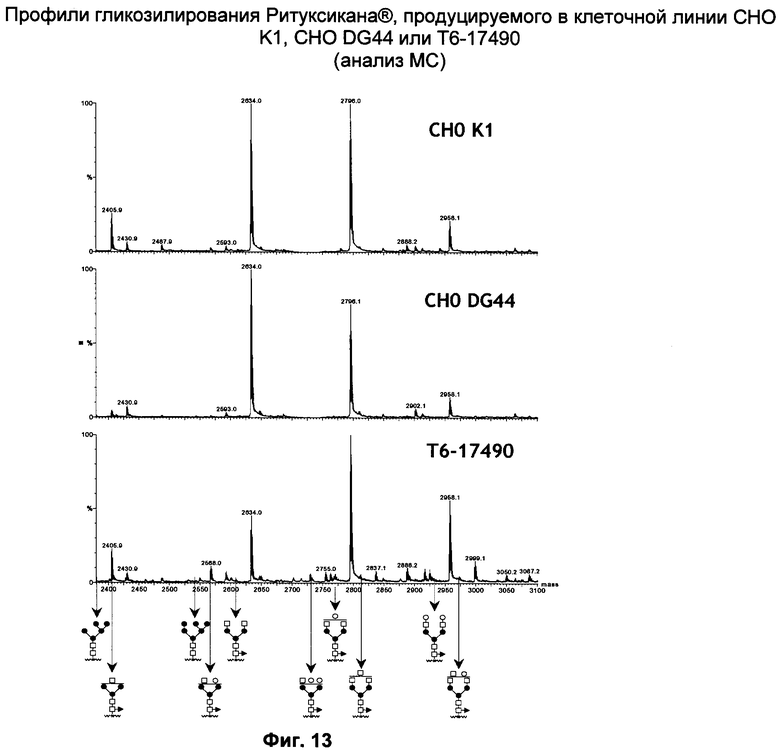
Фиг.13: Профили гликозилирования Ритуксикана®, продуцируемого различными клеточными линиями.
Примеры
Пример 1: Система экспрессии теломеразы
Случайная инсерция
Для данной цели использована плазмида, не имеющая специфичной последовательности гомологии с утиным геномом (фиг.1).
Направленная инсерция
Сконструирована плазмида, содержащая два фрагмента 5 п.о., гомологичных гену HPRT Cairina moschata, окружающие ген обратной транскриптазы теломеразы Cairina moschata, и два селективных маркера. Ген HPRT, кодирующий гипоксантингуанинфосфорибозилтрансферазу, выбран в качестве адекватного сайта для конститутивной экспрессии теломеразы Cairina moschata.
Эти два селективных маркера представляют собой ген FCU1 (Erbs et al. Cancer Res. 2000, 15, 60:3813-22) под контролем промотора CMV (Thomson et al. P.N.A.S. 1984, 81, 3:659-63) и ген устойчивости к неомицину, помещенный под контроль промотора SV40. Экспрессионная кассета устойчивости к неомицину и FCU-1 окружена сайтами расщепления Scel, которые позволяют элиминировать селекционные кассеты из конечной клеточной линии. Вне плеч гена HPRT встроен селективный маркер, кодирующий HSVTK, направляемый промотором RSV (фиг.2).
Пример 2: Система экспрессии теломеразы
Выделение теломеразы утки
РНК выделяли из 7-суточного эмбриона. Два эмбриона ресуспендировали в 2 мл RNA Now (Biogentex Inc, № по каталогу ВХ-101) и гомогенизировали, используя Polytron PT 3100. Затем добавляли 0,2 мл дихлорметана на 1 мл гомогената. Образец помещали на лед на 5 минут. После центрифугирования при 10000 g в течение 10 мин РНК, присутствующую в верхней водной фазе, осаждали изопропанолом и ресуспендировали в воде.
кДНК теломеразы Cairina moschata амплифицировали в 4 частях примерно 1 кб, названных А, В, С и D. Праймеры были сконструированы по гомологии с последовательностью теломеразы кур кДНК TERT. ПЦР амплификацию фрагмента С проводили, используя приведенные ниже праймеры, OTG16896 и OTG16897. Этот фрагмент был секвенирован и показал гомологию 86% с кДНК TERT кур. Фрагмент расщепляли ферментами рестрикции XbaI и SamHI и встраивали в pTG15812 (производный вектор pBS), расщепленный теми же ферментами, с получением pTG-C. Из последовательности фрагмента С были сконструированы праймеры для амплификации фрагментов В и D. ПЦР амплификацию фрагмента В проводили с праймерами OTG16789 и OTG17385. Этот фрагмент был секвенирован и показал гомологию 76% с кДНК TERT кур. Фрагмент расщепляли ферментами рестрикции BamHI и NotI и встраивали в pTG16384 (производный вектор pBS), расщепленный теми же ферментами, с получением pTGB-В. ПЦР амплификацию фрагмента D получили с праймерами OTG17387 и OTG16794. Последовательность этого фрагмента показала гомологию 87% с кДНК TERT кур. Фрагмент D расщепляли ферментами рестрикции SfiI и SacI и встраивали в pTG16384 (производное pBS), расщепленный теми же ферментами, с получением pTG-D. OTG17471 был сконструирован из последовательности фрагмента В для амплификации фрагмента А. ПЦР амплификацию фрагмента А проводили, используя праймеры OTG16894 и OTG17471. Фрагмент расщепляли ферментами рестрикции SalI и NotI и встраивали в pTG16384 (производный вектор pBS), расщепленный теми же ферментами, с получением pTG-A.
Полноразмерную кДНК TERT утки восстанавливали путем повторной сборки фрагментов А, В, С и D. Аминокислотная последовательность, кодируемая фрагментом BCD, раскрыта в SEQ ID №: 1.

Пример 3: Получение партии СЕС из 12-суточных яиц Cairina moschata и описание субпопуляций
25 оплодотворенных яиц SPF инкубируют при 37,5°С. Яйца вскрывают после 12 суток инкубации, следуя доступному протоколу.
23 эмбриона измельчают, промывают один раз фосфатно-солевым буферным раствором - средой Дульбекко (ФСБ) и диссоциируют в TrypLE Select (Invitrogen) 5 часов при комнатной температуре. После низкоскоростного центрифугирования клетки ресуспендируют в базовой среде Игла (МВЕ) с добавлением 10% фетальной сыворотки теленка (ФСТ), гентамицина 0,04 г/л, высевают в тройные флаконы 500 см2 и инкубируют при 37°С 5% CO2.
Через 24 ч конфлюентные клетки извлекают из флаконов, используя TrypLE Select (5 мл/тройной флакон), часть клеток пересевают во флаконы 175 см2 для второго пассажа. Остальные клетки концентрируют при 107 клеток/мл в подходящей среде (60% ВМЕ, 30% ФСТ и 10% ДМСО) и замораживают в контейнере, регулируемом изопропиловым спиртом (контейнер для замораживания NALGENE®, "Mr. Frosty", 1°C), при -80°С, после чего переносят в жидкий азот для длительного хранения, составляя исходный клеточный банк (50×1,5.107 клеток/флакон, 44×1.107 клеток/флакон).
Клетки, оставшиеся в культуре, пересевают классически вплоть до 18 пассажей, во время первых трех пассажей неприкрепленные клетки собирают низкоскоростным центрифугированием кондиционированной среды, пересевают и подвергают следующему пассажу таким же путем, как исходную культуру.
Субпопуляции, проявляющие характеристические различные морфологические признаки, воспроизводимо выделены на протяжении жизни культуры.
Пример 4: Методы трансфекции
В данной области техники известно большое число методов трансфекции для введения вектора, способного к направленной экспрессии интересующей нуклеотидной последовательности. Неограничивающий перечень этих методов представлен здесь ниже: CaPO4 преципитация, электропорация, липофектиновый метод трансфекции. Данный пример основан на методике CaPO4 преципитации.
Конфлюентность клеток должна составлять около 80-50%. Среду заменяют за два часа до добавления СаРО4/ДНК. 30 мкг ДНК ресуспендируют в 31 мкл 2М CaCl2 - 161,3 мМ Трис pH 7,6. Добавляют H2O до конечного объема 0,5 мл.
Для трансфекции 0,5 мл 2Х HEBS распределяют в 15 мл стерильные пробирки Falcon и добавляют по каплям раствор ДНК при осторожном вращении или барботировании раствора ДНК в них. Раствор должен приобрести молочную консистенцию. Смесь оставляют стоять при комнатной температуре на 10-30 мин. Затем пипетируют стерильной пипеткой один раз в боксе для тканевых культур для разрушения хлопьев и наносят по каплям на клетки. Затем клетки инкубируют между 6 часами и ночью при 37°С. Тонкий осадок должен покрывать клеточную поверхность. В целях завершения процедуры трансфекции подогревают до 37°С концентрированный раствор глицерина. Среду отсасывают, добавляют 5 мл ВМЕ для отмывки клеточного слоя, затем среду отсасывают и добавляют 1 мл концентрированного раствора глицерина на 2 мин или менее. Затем осторожно добавляют 10 мл ВМЕ для разведения глицерина, и ВМЕ-глицерин полностью удаляют. Затем добавляют 10 мл желаемой среды, и чашки инкубируют при подходящей температуре.
Пример 5: Методы селекции
Случайная инсерция
Селекционное давление применяют в течение 48 часов после трансфекции: клетки диссоциируют TrypLE Select, центрифугируют при низкой скорости и пересевают в ВМЕ с ФСТ 10% и G418 800 мкг/мл.
Клетки пересевают до тех пор, пока можно будет выделить индивидуальные растущие клоны. Центры размножения выделяют и амплифицируют, после чего количественно определяют теломеразную активность с помощью набора для определения теломеразы TRAPeze® XL (S7707, Chemicon) и проводят саузерн-блот-анализ для установления направленной интеграции в специфичный локус.
Направленная инсерция
Селекционное давление применяют в течение 48 часов после трансфекции: клетки диссоциируют TrypLE Select, центрифугируют при низкой скорости и пересевают в ВМЕ с ФСТ 10%, ганцикловиром 25 мкг/мл и G418 800 мкг/мл.
Клетки серийно пересевают до тех пор, пока можно будет выделить индивидуальные растущие клоны. Центры размножения выделяют и амплифицируют, после чего количественно определяют теломеразную активность с помощью набора для определения теломеразы TRAPeze® XL (S7707, Chemicon) и проводят саузерн-блот-анализ с олигонуклеотидами для установления направленной интеграции в специфичный локус.
Клеточные клоны с обнаруженной восстановленной теломеразной активностью и направленной интеграцией локуса HPRT последовательно трансфецируют экспрессионной плазмидой мегануклеазы I-Scel, следуя описанному ниже методу.
Для отбора элиминации селективных маркеров 5-фторцитозин (5-FC) применяют в течение 48 часов после трансфекции: клетки диссоциируют TrypLE Select, центрифугируют при низкой скорости и пересевают в среду с 5-FC в интервале концентрации от 10-3 до 10-7 М и поддерживают селекцию G418 (ВМЕ с ФСТ 10%, 5-FC и G418 800 мкг/мл).
Пример 6. Полученные иммортализованные клеточные линии Cairina moschata
6-А. Иммортализованная клеточная линия Cairina moschata T3-17490, депонированная в Европейской коллекции клеточных культур (ЕСАСС) под номером по каталогу 08060502
Клетки T3-17490 (пассаж 39) обладают однородной фибробластоподобной морфологией (фиг.4). Статичный монослой стабилен вплоть до 100% конфлюентности и подвержен контактному ингибированию. Клетки проверены как отрицательные на заражение микоплазмой, а также на бактериальное заражение.
Кривая роста клеточной линии T3-17490 (от пассажа 7 до пассажа 75) (фиг.5) показывает фазу непрерывного экспоненциального роста от пассажа 19 до пассажа 75. Фокусируясь на изменении времени удвоения популяции (ВУП), наблюдают его постепенную стабилизацию и уменьшение, в частности, можно отметить, что ВУП стабилизируется на отметке 48 ч во время последних 10 пассажей (фиг.6).
Соответствующее число удвоений популяции (уровень удвоения популяции, УУП) вычислено путем кумуляции 2 фаз экспоненциального роста: в течение 75 пассажей клетки претерпевают по меньшей мере 147 удвоений популяции (УП). Число удвоений популяции, которое может претерпеть первичная клетка до вступления ее в старение, является тканеспецифичным и видоспецифичным. Общеизвестно, что верхний предел расположен между 50 и 60 УП. Клетки Т3-17490, таким образом, далеки от предела Хейфлика и, соответственно, их считают иммортализованной клеточной линией.
6-Б. Иммортализованная клеточная линия Cairina moschata T6-17490, депонированная в Европейской коллекции клеточных культур (ЕСАСС) под номером по каталогу 08060501
Клетки T6-17490 (пассаж 45) обладают однородной фибробластоподобной морфологией (фиг.7). Статичный монослой стабилен вплоть до 100% конфлюентности и подвержен контактному ингибированию. Клетки проверены как отрицательные на заражение микоплазмой, а также на бактериальное заражение.
Кривая роста клеточной линии T6-17490 (от пассажа 15 до пассажа 51) (фиг.8) показывает фазу непрерывного экспоненциального роста от пассажа 19. В течение этого периода измеренное время удвоения популяции (ВУП) постепенно уменьшалось. Среднее ВУП перешло от 94 ч (пассаж 20-35) к 52 ч (пассаж 36-51) (фиг.9).
Число вычисленных удвоений популяции (УУП), соответствующее 51 пассажу, составляет по меньшей мере 71 удвоение популяции. Клетки Т3-17490, таким образом, далеки от предела Хейфлика и, соответственно, их считают иммортализованной клеточной линией.
Примечание
Уровень удвоения популяции (УУП) относится к числу клеточных генераций (2-кратное увеличение биомассы). Вычисление УУП: УУП = Ln(конечное/исходное число клеток)/Ln(2).
Время удвоения популяции (ВУП), также называемое временем генерации, представляет собой время, необходимое для одного удвоения популяции. Вычисление ВУП: PDT=Δt*Ln(2)/Ln(конечное/исходное число клеток).
Пример 7: Экспрессия белка TERT Cairina moschata иммортализованными клеточными линиями птиц Т3-17490 (ЕСАСС 08060502) и Т6-17490 (ЕСАСС 08060501)
I - Материал и способ обнаружения активности TERT
Активность теломеразы анализировали, используя набор Telo TAGGG ПЦР ЭЛАЙЗАплюс на теломеразу (Roche, № по каталогу 12013789001), в соответствии с инструкциями изготовителя. Набор основан на анализе протокола амплификации теломерного повтора (TRAP) с нерадиоактивным обнаружением с помощью ЭЛАЙЗА. Концепция анализа дает возможность полуколичественного определения уровней теломеразы. Предоставленный внутренний стандарт дает возможность обнаружения ингибиторов Taq ДНК полимеразы. Уровень активности теломеразы в данном образце определяют путем сравнения сигнала от образца с сигналом, полученным с использованием известного количества контрольной матрицы (TS8).
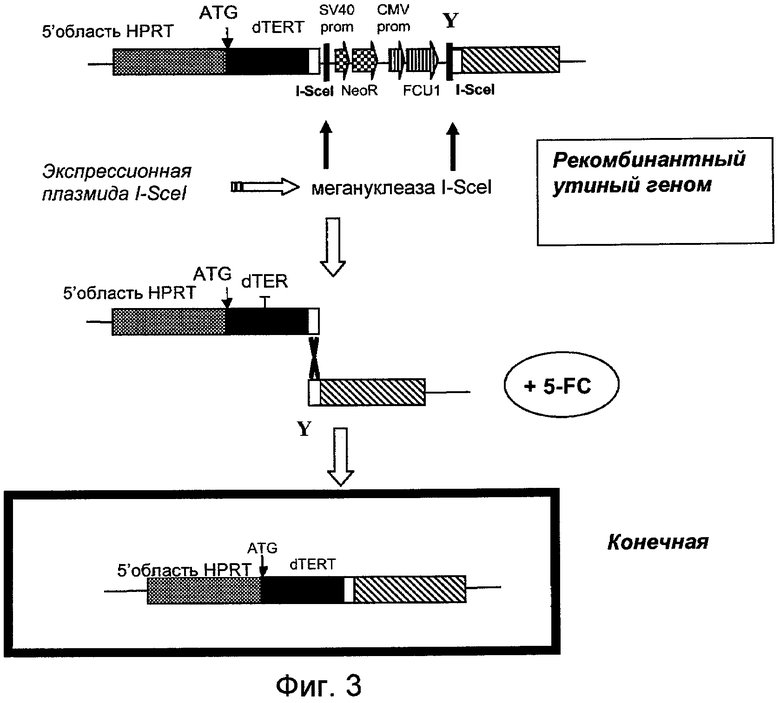
Относительные активности теломеразы (ОТА) внутри различных образцов в эксперименте получены с использованием приведенной ниже формулы:
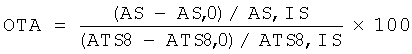
AS: поглощение образца
AS,0: поглощение образца, обработанного нагреванием или РНКазой
AS, IS: поглощение внутреннего стандарта (ВС) образца
ATS8: поглощение контрольной матрицы (TS8)
ATS8,0: поглощение буфера для лизиса
ATS8,IS: поглощение внутреннего стандарта (IS) контрольной матрицы (TS8)
Методику осуществляли на 3 мкл клеточного экстракта, что соответствует 3.103 клеточным эквивалентам. Отрицательный контроль с материалом образца, обработанным нагреванием (10 мин при 85°С), проводили параллельно с каждым неинактивированным образцом для подтверждения специфичности образования продукта анализа TRAP. Было обнаружено, что считывания поглощения для всех отрицательных контролей составляют менее 0,1 единицы отношения А450 нм к А690 нм, что необходимо для проверки правильности данных теста.
II - Результаты
Активность TERT тестировали на первичных клетках утки, в клеточной линии Т3-17490 (ЕСАСС 08060502) и в клеточной линии Т6-17490 (ЕСАСС 08060501). В каждом тесте использовали одно и то же число клеток.
Клеточная линия Т3-17490 (ЕСАСС 08060502): Как показано на фиг.10 и фиг.11, активность TERT была обнаружена для всех протестированных пассажей (от пассажа 6 до пассажа 86).
Примечание; Активность TERT присутствовала, начиная с первой фазы роста (пассажи 6, 8, 9), во время вступления в фазу латентного старения (пассажи 10-14) и во время второй экспоненциальной фазы роста (пассажи 27-86).
Клеточная линия Т6-17490 (ЕСАСС 08060501): Как показано на фиг.11, активность TERT была обнаружена для всех протестированных пассажей (от пассажа 22 до пассажа 62).
Как показано на фиг.10, активности TERT не было обнаружено в:
- первичных клетках уток после 5 пассажей (СЕТС19р5);
- контроле трансфекции Т3-17490 (ЕСАСС 08060502) после 7 пассажей (TTCETC19р7).
Пример 8. Продуцирование вирусов MVA из полученных иммортализованных клеточных линий
Способности клеточной линии Т6-17490 к амплификации MVA оценивали и сравнивали с первичными эмбриональными фибробластами цыпленка (CEF), обычно используемыми в качестве субстрата для продуцирования MVA. MVA, экспрессирующий eGFP, был выбран, чтобы облегчить слежение за размножением вируса и его титрование.
Клеточную линию Т6-17490 и CEF высевали при 2.106 клеток в Т-образный флакон 25 см2 и культивировали в течение 24 часов во влажной атмосфере при 37°С, 5% CO2. Среду удаляли, и клетки инфицировали при МИ (множественности инфекции) 0,05 500 мкл вируса MVA, разведенного в ФСБ с 1% катионами. После 30-минутной стадии адсорбции остаточную суспензию вируса удаляли, клетки промывали один раз ФСБ и добавляли в каждый флакон 5 мл ВМЕ 10% ФСТ. Вирус выделяли с помощью стадии замораживания-оттаивания из клеток и надосадочной жидкости после 0, 24, 48, 72 и 96 часов инфекции при 37°С, 5% CO2.
Титрования проводили в трех повторах на CEF, высеянных в 6-см культуральные чашки, инфицировали логарифмическими разведениями вируса и покрывали верхним агаром. Бляшкообразующие единицы MVA, экспрессирующего eGFP, визуализировали под флуоресцентным бинокуляром и считали после 72 ч. Кривые полученных в результате вирусных титров представлены на фиг.12. Самые высокие вирусные титры достигнуты для Т6-17490 и CEF после 48 ч инфекции. Амплификация вируса кратности 2,32 log достигнута на Т6-17490 по сравнению с 2,75 для CEF.
Пример 9. Продуцирование целевых веществ из полученных иммортализованных клеточных линий
9-А. Продуцирование моноклонального антитела Ритуксикан® из полученных иммортализованных клеточных линий
Ритуксикан® (ритуксимаб) представляет собой химерное моноклональное антитело мыши/человека анти-CD20. CD20 представляет собой гидрофобный трансмембранный белок, который присутствует на В лимфоцитах. Ритуксикан® применяют при лечении неходжкинской лимфомы и ревматоидного артрита.
Участок Fc ритуксимаба опосредует антителозависимую клеточную цитотоксичность (АЗКЦ) и комплементзависимую цитотоксичность (КЗЦ). Обнаружено, что АЗКЦ, литическая атака на клетки, являющиеся мишенью антитела, является одной из критических эффекторных функций, ответственных за клиническую эффективность данного терапевтического антитела. Эта активность зависит от фукозы, присоединенной к наиболее удаленной части GlcNAc N-сшитого олигосахарида Fc посредством α-1,6-связи, и резко усиливается в результате сокращения длины фукозы.
Тяжелая и легкая цепи были синтезированы фирмой GeneArt, и соответствующая экспрессионная плазмида собрана на основе вектора pCI-Neo. Транзитную генную экспрессию осуществляли с помощью специфичной «ФСТ с низким содержанием IgG». Таким образом, клеточную линию Т6-17490 размножали в течение 5 суток в 10% ФСТ с низким содержанием IgG, сравнивая с классической ФСТ, и отрицательного эффекта на клеточный рост не наблюдали. Транзитную генную экспрессию осуществляли в клеточной линии Т6-17490. Надосадочные жидкости собирали, и количество антитела оценивали, используя IgG ЭЛАЙЗА (общий человеческий IgG, Bethyl). В транзитных и неоптимизированных условиях в клеточной линии Т6-17490 было продуцировано 2,5 мкг на мл.
Ритуксикан®, транзитно продуцированный либо в стандартных клеточных линиях СНО (СНО K1 и СНО DG44, клон СНО с дефицитом по дигидрофолатредуктазе) или в клеточной линии Т6-17490, очищали с целью анализа и сравнения соответствующих паттернов гликозилирования. Масс-спектрометрический анализ (фиг.13) показал отсутствие различий в фукозилировании Ритуксикана®, экспрессированного СНО, и Ритуксикана®, экспрессированного клеточной линией Т6-17490.
9-В. Продуцииование интерлейкина-2 (ИЛ-2) из полученных иммортализованных клеточных линий
Интерлейкин-2 (ИЛ-2) представляет собой 153-аминокислотный (17,6 кДа) секретируемый цитокин, продуцируемый Т-клетками в ответ на антигенную или митогенную стимуляцию. Этот белок необходим для Т-клеточной пролиферации и других активностей, критичных для регуляции иммунного ответа. Он стимулирует В-клетки, моноциты, активируемые лимфокином киллерные клетки и естественные киллерные клетки. ИЛ-2 применяют у пациентов с почечно-клеточным раком и метастатической меланомой.
Надосадочные жидкости из клеточных линий Т3-17490 и Т6-17490 подвергали транзитной трансфекции экспрессионной плазмидой ИЛ-2 (промотор CMV). Надосадочные жидкости собирали после 24, 48, 72 и 96 ч. ИЛ-2 количественно определяли с помощью ЭЛАЙЗА (Quantikine, RD Systems), и функциональность определяли в анализе CTLL (клеточной линии, зависимой от ИЛ-2).
Обе клеточные линии Т3-17490 и Т6-17490 продуцировали функциональный ИЛ-2. Количество было эквивалентно между обеими клеточными линиями. Кроме того, уровень продуцирования был независим от использованного способа трансфекции, находясь в интервале от 4865 ед./мл (CTLL) и 211 нг/мл (ЭЛАЙЗА), что соответствует удельной активности 23,1 ед./нг, до 5672 ед./мл (CTLL) и 308 нг/мл, что соответствует удельной активности 18,4 ед./нг. Таким образом, измеренные удельные активности ИЛ-2, продуцируемого в клеточных линиях Т3-17490 и Т6-17490, эквивалентны международному стандарту ВОЗ 13,16 ед./нг для ИЛ-2 (человеческий, полученный из клеточной линии Jurkat). Следовательно, клеточные линии Т3-17490 и Т6-17490 пригодны для продуцирования функциональных цитокинов.




















Настоящее изобретение относится к области биотехнологии и касается полипептида, обладающего активностью теломеразы обратной транскриптазы (TERT), молекулы нуклеиновой кислоты, кодирующей такой полипептид, плазмидного вектора, несущего указанную молекулу нуклеиновой кислоты, иммортализованной клетки, трансфицированной указанной молекулой нуклеиновой кислоты, и применению такой клетки для продуцирования целевого вещества. Представленное изобретение позволяет разрабатывать иммортализованные птичьи клеточные линии с возможностью использования последних для получения какого-либо целевого вещества. 10 н. и 18 з.п. ф-лы, 13 ил.
1. Полипептид, обладающий активностью теломеразы обратной транскриптазы (TERT), содержащий аминокислотную последовательность SEQ ID №:1.
2. Молекула нуклеиновой кислоты, которая кодирует полипептид по п.1.
3. Молекула нуклеиновой кислоты по п.2, где указанная молекула нуклеиновой кислоты содержит нуклеиново-кислотную последовательность SEQ ID №:2.
4. Экспрессирующий плазмидный вектор, несущий молекулу нуклеиновой кислоты по любому из пп.2 или 3.
5. Вектор по п.4, где указанный вектор содержит две последовательности, гомологичные последовательности ДНК-мишени.
6. Вектор по п.5, где указанная молекула нуклеиновой кислоты окружена указанными гомологичными последовательностями.
7. Вектор по п.5, где указанный вектор дополнительно содержит первый селективный маркер, где указанный первый селективный маркер представляет собой маркер положительной селекции и где указанный первый селективный маркер окружен указанными гомологичными последовательностями.
8. Вектор по п.7, где указанный первый селективный маркер окружен последовательностями, дающими возможность его супрессии.
9. Вектор по п.5, где указанный вектор содержит второй селективный маркер, который не окружен указанными гомологичными последовательностями, и где указанный селективный маркер представляет собой маркер отрицательной селекции.
10. Вектор по любому из пп.7-9, где указанный вектор содержит третий селективный маркер, где указанный третий селективный маркер представляет собой маркер отрицательной селекции и где указанный третий селективный маркер расположен между последовательностями, дающими возможность супрессии первого селективного маркера.
11. Иммортализованная клетка птиц, трансфицированная молекулой нуклеиновой кислоты по любому из пп.2 или 3.
12. Клетка по п.11, где указанная молекула нуклеиновой кислоты встроена в ген HPRT указанной первичной клетки.
13. Клетка по п.11, где указанная клетка имеет происхождение от животного, принадлежащего к семейству Anatidae, предпочтительно видам Cairina moschata или Anas platyrhynchos.
14. Клетка по любому из пп.11-13, где она дополнительно содержит кассету комплементации, позволяющую размножаться дефектному вирусу.
15. Применение полипептида по п.1 для иммортализации клетки.
16. Применение молекулы нуклеиновой кислоты по любому из пп.2 или 3 для иммортализации клетки.
17. Применение вектора по любому из пп.4-10 для иммортализации клетки.
18. Применение клетки по любому из пп.11-13 для репликации живого, аттенюированного или рекомбинантного вируса, предпочтительно выбранного из группы, состоящей из поксвирусов.
19. Применение клетки по п.18, где поксвирус представляет собой вирус коровьей оспы, вирус оспы мышей, вирус оспы обезьян, вирус вакцинии, вирус натуральной оспы и MVA.
20. Применение клетки по любому из пп.11-13 для продуцирования целевого вещества.
21. Применение клетки по п.20, где указанное целевое вещество выбрано из группы, состоящей из факторов роста, регуляторов роста, антител, антигенов, интерлейкинов, инсулина, G-CSF, GM-CSF, hPG-CSF, M-CSF, интерферонов, факторов свертывания крови или их комбинации.
22. Способ иммортализации клетки, включающий стадию трансфекции указанной клетки вектором по любому из пп.4-10.
23. Способ по п.22, характеризующийся тем, что он дополнительно включает стадию, где указанные клетки культивируют в среде, которая позволяет расти только клеткам, которые включили первый селективный маркер.
24. Способ по п.22, характеризующийся тем, что он дополнительно включает стадию, где указанные клетки культивируют в среде, которая не позволяет расти клеткам, которые включили второй селективный маркер.
25. Способ по п.22, характеризующийся тем, что он дополнительно включает стадию, состоящую в исключении первого селективного маркера из генома указанной клетки.
26. Способ по п.25, где клетки, полученные после указанной стадии, состоящей в исключении первого селективного маркера из генома указанных клеток, культивируют в среде, которая не позволяет расти клеткам, которые включили третий селективный маркер.
27. Способ по любому из пп.22-26, где указанная клетка взята от организма, принадлежащего к семейству Anatidae, предпочтительно вида Cairina moschata или Anas platyrhynchos.
28. Способ по любому из пп.22-26, где указанная молекула нуклеиновой кислоты встроена в последовательность ДНК-мишени указанной клетки, предпочтительно в ген HPRT.
| MARY E.DELANY et al., The chicken telomerase reverse transcriptase (chTERT): molecular and cytogenetic characterization with a comparative analysis, Gene, 2004, Vol.339, p.61-69 | |||
| SWANBERG SE et al., Telomerase activity and differential expression of telomerase genes and c-myc in chicken cells in vitro, Dev Dyn., 2004 Sep, Vol.231, No1, p.14-21. |

