Изобретение относится к биофармакологии, а именно к способам промышленного получения рекомбинантного гранулоцит-колонийстимулирующего фактора человека медицинского назначения (далее Г-КСФ), а также к рекомбинантным штаммам Escherichia coli (E.coli) и плазмидам для его получения.
Гранулоцит-колонийстимулирующий фактор человека (Г-КСФ) представляет собой гликопротеид с молекулярной массой 19000 Да. Г-КСФ, продуцируемый моноцитами-макрофагами, фибробластами и эндотелиальными клетками, индуцирует пролиферацию колоний нейтралов и дифференциацию клеток-предшественников в нейтрофилы, а также стимулирует активность зрелых нейтрофилов [1-4].
Лекарственные формы Г-КСФ (синонимы - Нейпоген (Ф.Хоффманн-Ля Рош Лтд.) и Граноцит (Авентис)) широко используются в медицине для терапии нейтропении у больных, получающих цитостатические средства для лечения немиелоидных злокачественных заболеваний; для купирования побочных эффектов миелосупрессивной химиотерапии; для терапии наследственной, периодической и злокачественной нейтропении; для сокращения продолжительности периода нейтропении и ее клинических последствий у больных, готовящихся к трансплантации костного мозга.
Известны способы получения Г-КСФ, основанные на использовании культуральной жидкости клеточных линий, продуцирующих Г-КСФ [5-7], а также на экспрессии гена Г-КСФ в трансформированных вектором клетках высших эукариот COS, CHO, С-127 [1, 7, 8]. Недостатком этих способов является чрезвычайно низкий выход целевого продукта и, как следствие, высокая стоимость препаратов Г-КСФ. Поэтому значительно более перспективным является способ получения Г-КСФ микробиологическим синтезом, который обеспечивает возможность получения продукта с более высоким выходом из сравнительно недорогого исходного сырья. Использование при этом химического подхода позволяет создать оптимальные для бактериальной экспрессии варианты структурного гена, а также регуляторных элементов, контролирующих его экспрессию. К тому же известно, что полисахаридная цепь гликопротеида не является необходимой для его биологической активности [9].
Известен способ получения рекомбинантного Г-КСФ, основанный на получении рекомбинантного Г-КСФ в трансформированных дрожжевых клетках Saccharomyces cerevisiae [10]. При таком подходе удается получить Г-КСФ человека в правильной конформации с хорошей физиологической активностью. Недостатком этого способа является снижение выхода за счет деградации целевого белка внутриклеточными протеазами.
Известен также способ получения Г-КСФ, включающий экспрессию в клетках Escherichia coli, заключающийся в секреции клетками бактерий рекомбинантного белка Г-КСФ в виде растворимого белка в периплазматическое пространство [заявка РСТ WO 01/00549, МКИ С12N 15/70, опубл. 2001]. Способ обеспечивает возможность получения правильной конформации белка без рефолдинга. Недостатком является экспрессия Г-КСФ с дополнительным пептидом на N-конце для устранения токсического действия белка, что делает необходимым отрезание специфической протеазой этого пептида.
Известны наиболее близкие к заявленному способы получения Г-КСФ, включающие экспрессию в клетках Escherichia coli, заключающиеся в быстром биосинтезе клетками бактерий рекомбинантного Г-КСФ в виде нерастворимых «телец включения» [патент РФ №2113483, МКИ С12N 15/27, опубл. 1/21 и патент РФ №2260049, МКИ С12N 15/27, опубл. 1/21]. В этих способах проводят конструирование рекомбинантных плазмидных ДНК, кодирующих конститутивный синтез полипептида со свойствами Г-КСФ человека и штаммы Escherichia coli, обеспечивающие синтез этого пептида с уровнем экспрессии 10-30% суммарного клеточного белка.
Наряду с особенностями используемых штаммов эффективность процесса во многом зависит от используемой технологии выделения и очистки Г-КСФ.
Задачей данного изобретения является конструирование рекомбинантного промышленного штамма продуцента E.coli, обладающего высоким уровнем биосинтеза Г-КСФ и создание препарата Г-КСФ в форме водного раствора для инъекций.
Указанная задача решалась созданием рекомбинантной плазмидной ДНК рSХ70 и штамма Escherichia coli SX70, депонированных во Всероссийской коллекции Промышленных Микроорганизмов (ВКПМ), ФГУП Гос. НИИ генетика, коллекционный номер ВКПМ В-8842.
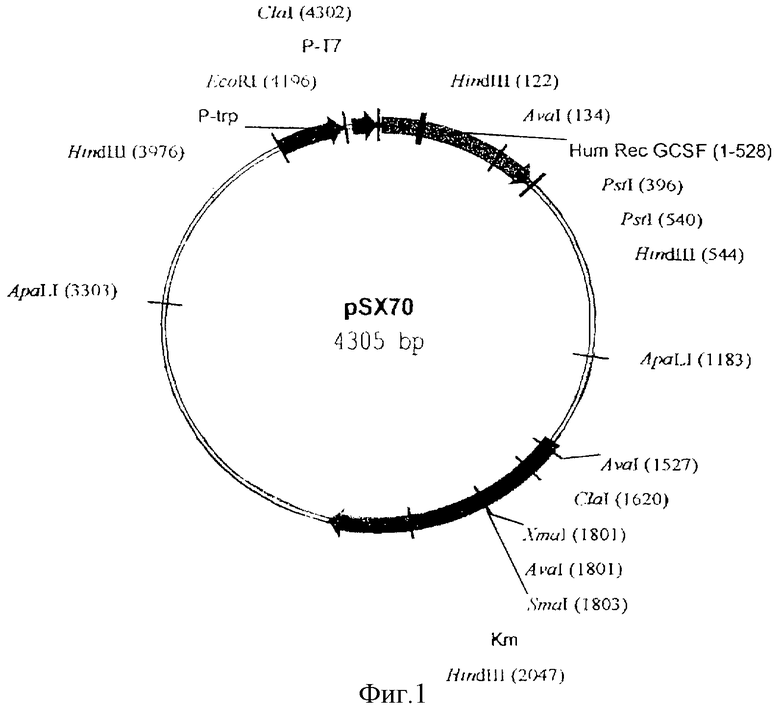
Плазмида pSX70 имеет 4305 пар оснований (п.о.) и характеризуется наличием следующих фрагментов:
- Последовательность с 1 по 528 н. включает фрагмент ДНК размером 529 п.о., содержащий ген Г-КСФ со следующими нуклеотидными заменами: 6 (С⇒А), 9 (С⇒А), 10 (С⇒Т), 11 (G⇒A), 15 (C⇒T), 21 (C⇒T), 22-24 (AGC⇒TCG), 25-27 (ТСС⇒AGT), 189 (С⇒Т), 198 (С⇒А), 210 (G⇒С), 270 (A⇒C), 291 (С⇒Т), 294 (С⇒A), 321 (G⇒T), 327 (G⇒C), 336 (C⇒T), 385 (С⇒A), 391 (C⇒A), 442 (С⇒Т), 454 (G⇒Т), 457 (С⇒Т), 460 (G⇒A).
- Последовательность с 540 по 550 н. включает синтетический фрагмент ДНК размером 11 п.о., содержащий синтетический полилинкер;
- Последовательность с 551 по 1070 включает в себя фрагмент плазмиды pSX50, содержащий последовательность строгого терминатора транскрипции rrnBT1T2.
- Последовательность с 1071 по 2802 н. включает фрагмент ДНК плазмиды pUC19 с 885 по 1735 н. (ген  -лактамазы, придающей устойчивость к ампициллину - AmpR), в который по сайтам ScaI - BglI вставлен ген устойчивости к канамицину KanR, фрагмент плазмиды pSX50 размером 816 п.о..
-лактамазы, придающей устойчивость к ампициллину - AmpR), в который по сайтам ScaI - BglI вставлен ген устойчивости к канамицину KanR, фрагмент плазмиды pSX50 размером 816 п.о..
- Последовательность с 2803 по 4200 включает в себя фрагмент плазмиды pSX50, содержащий последовательность, ответственную за репликацию плазмиды (ori) и lac промотора (Plac), а также последовательность, содержащую триптофановый промотор (Ptrp);
- Последовательность с 4201 по 4305 н. содержит промотор фага Т7 (РТ7), а также синтетический полилинкер.
На фиг.1 изображена физическая карта плазмиды pSX70, на фиг.2 - последовательность нуклеотидов гена Г-КСФ.
Штамм Escherichia coli SX70 получен трансформацией клеток Escherichia coli BL21 плазмидой pSX70 с использованием традиционной генно-инженерной технологии.
Штамм E.coli SX70 характеризуется следующими признаками:
Культурально-морфологические признаки
Клетки мелкие, прямые, утолщенной палочковидной формы, грамотрицательные, неспороносные.
Клетки хорошо растут на простых питательных средах. При росте на агаре "Дифко" образуются круглые, гладкие, выпуклые, мутные, блестящие, серые колонии, с ровными краями. При росте в жидких средах (в минимальной среде с глюкозой или в LB-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки
Аэроб. Температурный диапазон роста 4-42°С при оптимуме рН 6,5-7,5.
В качестве источника азота используют как минеральные соли в аммонийной и нитратной формах, так и органические соединения в виде аминокислот, пептона, триптона, дрожжевого экстракта и т.д.
В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к канамицину (до 100 мкг/мл). Штамм Escherichia coli SX70 - продуцент Г-КСФ.
Способ, условия и состав среды для хранения штамма
В L-агаре с добавлением канамицина до концентрации 20 мкг/мл под маслом, в L-бульоне, содержащем 15% глицерина и соответствующими антибиотиками в ампулах при температуре минус 70°С, в лиофилизированном состоянии в ампулах при температуре 4°С.
Штамм Escherichia coli SX70 идентифицирован по Определителю Берги (1974) как штамм вида Escherichia coli.
Особенностью заявляемого способа является разработка технологии, позволяющая выделять Г-КСФ человеческий рекомбинантный из нерастворимой формы, накапливающейся в течение ферментации, что позволяет существенно упростить технологическую схему процесса выделения и повысить выход целевого продукта.
Способ заключается в культивировании в питательной среде штамма Escherichia coli SX70, разрушении клеток микроорганизма в проточном дезинтеграторе при давлении 700 bar, выделении и очистке нерастворимой формы Г-КСФ с использованием детергентов и последующим центрифугированием, растворением осадка в растворе гуанидин гидрохлорида и последующей ренатурации Г-КСФ.
Полученный ренатурированный Г-КСФ подвергают очистке с помощью обращенно-фазной хроматографии на фенил-сефарозе FF High Sub, ионообменной хроматографии на ионообменных смолах типа CM Sepharose Fast Flow и гель фильтрационной хроматографии на смолах типа Superdex 75.
Оптимальными условиями проведения отдельных стадий получения Г-КСФ являются следующие:
- ферментацию проводят при непрерывном добавлении субстратов в течение всего процесса, что обуславливает высокий уровень экспрессии Г-КСФ;
- разрушение клеток осуществляют в проточном дезинтеграторе при давлении не менее 700 bar;
- удаление растворимых клеточных компонентов (ДНК, РНК, белков, липополисахаридов и т.д.) производят промыванием нерастворимой формы Г-КСФ буферными растворами, содержащими детергенты (CHAPS, мочевина и т.д.);
- образовавшийся осадок, содержащий Г-КСФ, растворяют в буферном растворе 6 М гуанидин гидрохлорида;
- ренатурацию Г-КСФ проводят в физиологическом буферном растворе, содержащем хаотропные агенты;
- трехстадийную хроматографическую очистку Г-КСФ проводят на фенил-сефарозе FF High Sub, на катионообменной смоле CM Sephsrose Fast Flow и гель-фильтрационную хроматографию на смоле типа Superdex 75;
- после каждой хроматографической очистки проводят стерилизующую фильтрацию через апирогенные фильтры с размерами пор 0.22 микрон.
Выход Г-КСФ в результате применения описанного способа составляет не менее 200 мг чистого белка с 1 л культуральной среды.
Существенными отличиями заявляемого способа от прототипа являются:
- использование конструкции штамма с более высокой производительностью, что позволяет получать при биосинтезе большее количество Г-КСФ с 1 л культуральной среды;
- использование эффективного механического разрушения клеточной биомассы, что позволяет получать более чистый экстракт нерастворимой формы Г-КСФ за более короткое время, с меньшими потерями;
- использование физиологических буферных растворов при ренатурации в присутствии хаотропных агентов позволяет повысить выход корректно ренатурированной формы Г-КСФ;
- трехстадийная хроматографическая очистка Г-КСФ позволяет получать субстанцию Г-КСФ более 98% чистоты по данным изоэлектрического фокусирования и RF HPLC, и практически не содержащую пирогенов (LAL-тест).
Сущность и преимущества заявляемой группы изобретений иллюстрируются следующими примерами.
Пример 1. Конструирование рекомбинантной плазмиды pSX70
Способ конструирования плазмиды pSX70 включает следующие этапы:
- конструирование промежуточной рекомбинантной плазмиды рХХ1;
- конструирование рекомбинантной плазмиды pSX53 (4068 п.о.);
- конструирование рекомбинантной плазмиды pSX70 (4305 п.о.).
Конструирование промежуточной рекомбинантной плазмиды рХХ1
Рекомбинантная плазмида pXX1 представляет собой вектор pCHEreP, в который по сайтам ClaI и SalI клонирован ген gcsf.
10 мкг плазмидной ДНК pGSF(b) [11] обрабатывают последовательно рестриктазами ClaI и SalI в соответствии с методикой, описанной в работе [12], и из полученного гидролизата выделяют в 1% геле легкоплавкой агарозы фрагмент длиной 530 п.о., содержащий ген gcsf.
10 мкг плазмидной ДНК pCHFreP обрабатывают последовательно рестриктазами ClaI и SalI и из полученного гидролизата выделяют векторную часть плазмиды.
Полученный фрагмент и векторную часть плазмиды pCHEreP сшивают при помощи лигазной реакции в 50 мкл буфера для лигирования [12]. 10 мкг реакционной смеси используют для трансформации компетентных клеток E.coli DH5a. Трансформанты высевают на LB-агар, содержащий 75 мкг/мл ампицилина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК и анализируют рестрикционным анализом. Отбирают ДНК, содержащую нужный набор рестрикционных фрагментов. В результате получают плазмиду pXX1.
Конструирование рекомбинантной плазмиды pSX53
Плазмида pSX53 представляет собой плазмиду pSX50, последовательность которой с 1050 по 1861 н. заменена на фрагмент ДНК плазмиды pUC19 с 885 по 1735 н. (ген -лактамазы, придающей устойчивость к ампициллину AmpR), в который затем по сайтам ScaI - BglI вставлен ген устойчивости к канамицину - KanR.
Плазмиду pSX53 получают из промежуточной плазмиды pSX50-b введением гена устойчивости к канамицину по сайтам ScaI - BglI.
Для получения плазмиды pSX50-b проводят два раунда амплификации ДНК методом ПЦР (полимеразная цепная реакция). В ходе первого раунда, используя ДНК плазмиды pUC19 к качестве матрицы, проводят амплификацию фрагмента ДНК размером 850 п.о., гена -лактамазы, придающей устойчивость к ампициллину - AmpR) с использованием праймеров Amp1 и Amp2 (фрагмент 1):
Amp1 5` - ATG AGT ATT CAA С-3'
Аmр2 5` - TTA CCA ATG CTT AAT С-3'
Данную и последующие ПЦР реакции проводят в следующих условиях: 20 mM Tris-HCl, pH 8,8, 10 mM (NH4)2SO4, 10 mM KCl, 2 mM MgCl2, 0,1% Triton X100, 0,1 mg/ml BSA, 0,2 mM каждого dNTP, 1,25 ед. Pfu ДНК полимеразы, 100 нг ДНК.
Процесс амплификации состоит из следующих стадий: прогревание при 95°С 5 мин, 35 циклов ПЦР (30 сек 95°С, 30 сек 56°С, 2 мин 72°С) и инкубация 10 мин при 72°С. После амплификации (и после последующих амплификаций) фрагмент ДНК очищают электрофоретически в 1% агарозном геле.
В ходе второго раунда, используя ДНК плазмиды pSX50 в качестве матрицы, проводят амплификацию фрагмента ДНК размером 2402 п.о. (фрагмент 2) с помощью праймеров S1 и S2:
S1 5`- CTG TCA GAC CAA G-3'
S2 5`- ACT CTT CCT TTT TCA AT-3'
Полученные фрагменты (фрагмент 1 и фрагмент 2) сшивают при помощи лигазной реакции в 50 мкл буфера для лигирования. 10 мкг реакционной смеси используют для трансформации компетентных клеток E.coli DH5a. Трансформанты высевают на LB-агар, содержащий 75 мкг/мл ампицилина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК и анализируют рестрикционным анализом. Отбирают ДНК, содержащую нужный набор рестрикционных фрагментов. В результате получают плазмиду рХХ1.
10 мкг плазмидной ДНК pSX50 обрабатывают последовательно рестриктазами ScaI - BglI и из полученного гидролизата выделяют в 1% геле легкоплавкой агарозы фрагмент длиной 816 п.о., содержащий ген устойчивости к Kan.
10 мкг плазмидной ДНК pSX50 обрабатывают последовательно рестриктазами ScaI - BglI и из полученного гидролизата выделяют векторную часть плазмиды.
Полученные фрагменты (фрагмент 1 и фрагмент 2) сшивают при помощи лигазной реакции в 50 мкл буфера для лигирования. 10 мкг реакционной смеси используют для трансформации компетентных клеток E.coli DH5a. Трансформанты высевают на LB-агар, содержащий 20 мкг/мл канамицина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК и анализируют рестрикционным анализом. Отбирают ДНК, содержащую нужный набор рестрикционных фрагментов. В результате получают плазмиду pSX53.
Конструирование векторной плазмиды pSX70
10 мкг плазмидной ДНК рХХ1 обрабатывают последовательно рестриктазами EcoRI и HindIII, и из полученного гидролизата выделяют в 1% геле легкоплавкой агарозы фрагмент длиной 547 п.о., содержащий промотор фага Т7 (PТ7), последовательность Шайно-Дальгарно и ген gcsf, кодирующий Г-КСФ.
10 мкг плазмидной ДНК pSX53 обрабатывают последовательно рестриктазами EcoRI и PstI и из полученного гидролизата выделяют векторную часть плазмиды.
Далее электрофоретически очищенные фрагменты объединяют, лигируют ферментом лигаза фага T4, ДНК трансформируют в клетки штамма E.coli DH5 и высевают на среду LA, содержащую 20 мкг/мл канамицина. После инкубирования в течение 12 час при 37°С клоны отсевают, выделяют плазмидную ДНК, проводят рестрикционный анализ и определяют первичную структуру ДНК. В результате получают плазмиду рSХ70 размером 4305 п.о.
Целевая рекомбинантная плазмида pSX70 содержит уникальные сайты узнавания рестрикционными эндонуклеазами, имеющие следующие координаты.
Пример 2. Получение штамма E.coli SX70 - продуцента Г-КСФ
Штамм - продуцент интерферона E.coli SX70 получают путем трансформации клеток штамма E.coli BL21 рекомбинантной плазмидой pSX70. Штамм - продуцент Г-КСФ выращивают в 30 л ферментере до оптической плотности 25,0-30,0 о.е. в среде М9, содержащей 1% кислотного гидролизата казеина, 1% глюкозы, 40 мкг/мл канамицина, при температуре 38°С. В процессе ферментации проводят непрерывное добавление питательного субстрата, используя гравитометрический контроллер.
Содержание Г-КСФ в биомассе клеток, получаемые с 1 л культуры, составляет в зависимости от серии 0,9-1,0 г Г-КСФ.
Пример 3. Способ выделения Г-КСФ из штамма Е.coli SX70
Получение Г-КСФ проводили в четыре этапа:
1 этап. Культивирование штамма Е.coli SX70
2 этап. Выделение и очистка нерастворимой формы Г-КСФ
3 этап. Растворение и ренатурация Г-КСФ
4 этап. Хроматографическая очистка Г-КСФ
1 этап. Культивирование штамма E.coli SX50
Выращенный посевной материал штамма E.coli SX70 в объеме 3 л в богатой среде LB в течение 12 ч при 28°С асептически вносят в ферментер, содержащий 27 л стерильной среды, содержащей М9, 1% кислотного гидролизата казеина, 1% глюкозы, 1 мМ MgCl2, 0,1 mM CaCl2, 40 мг/мл канамицина. Культивирование в ферментере проводят при температуре 38-39°С, поддерживая pH 7±0,15 путем автоматической подтитровки 12,5%-ным раствором аммиака. Концентрацию растворенного кислорода в диапазоне (50±10)% от насыщения поддерживают путем изменения скорости оборотов мешалки от 100 до 800 об/мин и подачи воздуха от 1 до 15 л/мин. Концентрацию субстратов, в частности глюкозы, измеряют в течение ферментации и поддерживают их концентрацию путем изменения скорости подачи концентрированных растворов через перистальтические насосы, используя гравиметрический контроллер.
Накопление Г-КСФ в виде нерастворимой формы - "телец включений" контролируют с помощью фазово-контрастной микроскопии. Ферментацию останавливают по достижении максимальной оптической плотности (˜25-30 о.е.) и появлении зрелых "телец включений". По окончании ферментации культуральную жидкость отделяют центрифугированием в проточном роторе при скорости вращения 5000-10000 об/мин. Биомассу фасуют в полиэтиленовые пакеты и замораживают при температуре минус 70°С.
2 этап. Выделение и очистка нерастворимой формы Г-КСФ
100 г замороженной биомассы штамма E.coli SX70 суспедируют в 0,8 л буфера 1 (20 мМ Tris-HCl, pH 8,0, 10 мМ ЭДТА, 0,1% Triton X100). Суспензию пропускают через проточный дезинтегратор, поддерживают давление 700 бар и центрифугируют к проточном роторе при 15000 об/мин. Полученный осадок промывают в аналогичных условиях последовательно буферами 2 (20 мМ Tris-HCl, рН 8,0, 1 мМ ЭДТА, 2 М мочевины), буфером 3 (20 мМ Tris-HCl pH 8,0, 1 мМ ЭДТА, 3 М мочевины) и буфером 4 (20 мМ Tris-HCl рН 8,0, 1 мМ ЭДТА, 0,1% Тритон Х-100) и окончательно осадок Г-КСФ ресуспендируют в 150 мл буфера 5 (20 мМ Tris-HCl рН 8,0, 1 мМ ЭДТА, 0,001 М фенилметилсульфонилфторида). При этом время выделения и очистки нерастворимой формы Г-КСФ составляет не более 5 час.
3 этап. Растворение и ренатурация Г-КСФ
К полученной на предыдущем этапе суспензии нерастворимой формы Г-КСФ добавляют сухой гуанидин гидрохлорид до концентрации 6 М, добавляют раствор дитиотреитола до концентрации 50 мМ и инкубируют при комнатной температуре в течение 2 ч. Нерастворившийся материал отделяют при стерилизующей фильтрации через мембраны с диаметрами пор 0,22 микрона. Добавляют 1 мл концентрированного раствора 10% CHAPS.
Ренатурацию Г-КСФ проводят путем медленного разбавления полученного раствора в 100 раз буфером 6 (20 мМ Tris-HCl рН 8,0, 20 мМ NaCl, 0,1 мМ ЭДТА, 1 М мочевина, 0,1% CHAPS, 0,5 мМ глутатиона окисленного и 0,5 мМ глутатиона восстановленного). После чего ренатурационную смесь инкубируют при постоянном перемешивании в течение 12-15 час при температуре 4-8°С. Далее агрегированный материал удаляют стерилизующей фильтрацией через мембранный фильтр с диаметром пор 0,22 микрона.
4 этап. Хроматографическая очистка Г-КСФ
Хроматографическую очистку Г-КСФ осуществляют в три стадии.
1. Полученный ренатурированный Г-КСФ на первом этапе подвергают очистке с помощью обращенно-фазной хроматографии на фенил-сефарозе FF High Sub (GE Healthcare). Для этого раствор Г-КСФ наносят на колонну с фенил-сефарозой FF High Sub и связанный материал элюируют буфером 0,025 М ацетата натрия рН 4,5 и 0,001 М ЭДТА.
2. На второй стадии хроматографической очистки раствор Г-КСФ наносят на катионообменную смолу типа CM Sepharose Fast Flow (GE Healtheare) и связанный материал элюируют градиентом растворов (0,0-0,5 М NaCl) в буфере 50 мМ Na(CH3COO) pH 4,5.
3. Очистка мономерной формы Г-КСФ от остатков полимерных форм проводят на третьей стадии очистки Г-КСФ - гель фильтрации на смоле типа Superdex 75 (GE Healthcare). Хроматографию проводят в буфере 10 мМ Na(CH3COO) pH 5,0, содержащим 5% сорбитола и 0,004% полисорбата 80.
Описанный способ выделения и очистки Г-КСФ позволяет получить 5-6 г высокоочищенного Г-КСФ за один цикл выделения в течение 7-10 дней из биомассы, полученной с 30 л культуральной среды. Качество получаемого Г-КСФ в полной мере соответствует следующим требованиям:
- Содержание правильно ренатурированной формы Г-КСФ не менее 95% по данным RF HPLC;
- Изоэлектрическая точка выделенного Г-КСФ находится в районе рН 5,8-6,3;
- Содержание бактериальных эндотоксинов не более 100 ME на 1 мг интерферона.
На основе высокоочищенного Г-КСФ, полученного по описанным условиям, готовится композиция состава препарата, содержащая стабилизирующее вещество сорбитол, неионогенное поверхностно-активное вещество полисорбат 80, буферную систему, включающую натрия ацетат для обеспечения pH раствора 3,8-4,2, и воду для инъекций.
Способ приготовления
Для приготовления указанного раствора смешивают 1400 мл воды для инъекций, 500 мл концентрированного раствора 20% сорбитола и вносят 1,2 мл ледяной уксусной кислоты. Доводят pH до 4,0 с помощью 2 М раствора гидроксида натрия и вносят 0,8 мл концентрированного раствора 10% Полисорбата 80 при непрерывном перемешивании. Доводят объем до 2 л водой для инъекций и перемешивают. Раствор охлаждают до 4°С, после чего к нему добавляют раствор субстанции Г-КСФ до концентрации 30 мл ЕД/мл в 2 л конечного раствора. Полученный раствор доводят до 2 л водой для инъекций, подвергают стерилизующей фильтрации на мембранном фильтре 0,22 мкм и разливают по 1 мл во флаконы емкостью 2 мл. Полученный раствор стабилен при хранении при 2-8°С в течение 24 месяцев.
Как следует из приведенных примеров, заявляемая группа изобретений позволяет получать Г-КСФ с высоким выходом при относительно простой и надежной технологии и готовить на его основе препарат в форме водного раствора для инъекций.
Источники информации
1. Nagata S., Tsuchiya M., Asano S., Kaziro Y., Yamazaki Т., Yamamoto O., Hirata Y., Kubota N., Oheda M., Nomura H., Ono M. // Nature, 1986, v.319, No.6052, p.415-418.
2. Nagata S., Tsuhiya M., Asano S., Yamamoto O., Hirata Y., Jubota N., Oheda M., Nomura H., Yamazaki T. // EMBO J., 1986, v.5, p.575-581.
3. Souza L.M., Boone Т.C., Gubrilove J., Lai P.H., Zsebo K.M., Mudrock D.C., Chazin V.R., Bruszewski J., Kenneth H.L., Chen K.K., Barendt J., Platzer E., Moore M.A.S. Mertelsmann R., Welle K. // Science, 1986, v.232, No.4746, p.61-65.
4. Cohen A.M., Zsebo K.M., Inoue H., Hines D., Boone T.C., Chazin V.R., Tsai L., Ritch T., Soyza L.M. // Proc. Nail. Acad. Sci, USA, 1987, v.87. No.8, p.2484-2288.
5. Патент США N4833127, кл. Ф61К 37/02, 1989.
6. Nomura H., Imazeki I., Oheda M., Kubota N., el al. // EMBO J., 1986, v.5, p.871-876.
7. Европейский патент N215126, кл. С12N 15/00, 1987.
8. Европейский патент N220520, кл. С12N 15/00, 1987.
9. Oheda M., Hasegawa M., Hattori K., Kuboniwa H., Kojima Т., Orita Т., Tomonou K., Yamazaki T., Ochi N. // J. Biol. Chem., 1990, v.265, No.20, p.11432-11435.
10. Gillis S., Urdal D.L., Clevenger W., Klinke R., Sassenfeld H., Price V., Cosman D. // Behring Inst. Mitt., 1988, p.1-7.
11. Кашьян С.П., Быстров Н.C., Болдырева Е.Ф., Полякова И.А., Северцова И.В., Коробко В.Г. // Биоорган. химия, 1992, т.18, N1, с.71-77.
12. Sambrook J., Fritsch E.F., Maniatis T. // Molecular Cloning. A. Laboratory Manual. 2bd ed. Cold Spring Harbor, NY, 1989.

Изобретение относится к биофармакологии, а именно к способам промышленного получения рекомбинантного гранулоцит-колонийстимулирующего фактора человека медицинского назначения (далее Г-КСФ), а также к рекомбинантным штаммам Escherichia coli (E.coli) и плазмидам для его получения. Группа изобретений включает создание рекомбинантной плазмидной ДНК pSX70, конструирование рекомбинантного промышленного штамма продуцента Escherichia coli pSX70, депонированных во Всероссийской коллекции Промышленных Микроорганизмов (ВКПМ), ФГУП Гос. НИИ генетика, коллекционный номер ВКПМ В-8842. Изобретение позволяет получать Г-КСФ с высоким выходом при относительно простой и надежной технологии и готовить на его основе композицию в форме водного раствора для инъекций. 4 н. и 5 з.п. ф-лы, 2 ил.

