Предпосылки изобретения
Область техники, к которой относится изобретение
Настоящее изобретение в целом касается области травматологии и лечения ран. Более конкретно настоящее изобретение касается способов улучшения заживления ран и усовершенствованных материалов для раневых покрытий.
Описание области техники
Ожоговые повреждения представляют собой одну из наиболее тяжелых форм травм. Чем сильнее повреждение от ожога, тем более тяжелыми являются последствия и выше вероятность неблагоприятных исходов и смерти. Ежегодно число пациентов с ожогами превышает 2 миллиона человек, а стоимость лечения превосходит один миллиард долларов в год. Огневые и ожоговые повреждения являются третьими в списке причин повреждений и смерти детей в возрасте от 1 до 18 лет. В последнее десятилетие смертность от ожогов снижалась в основном благодаря ранней и адекватной реанимационной инфузионной терапии, раннему и насильному искусственному питанию, улучшенной борьбе с инфекциями, улучшенным обработке и заживлению ран и гормональной модуляции.
Заживление ран является наиболее важным в излечении ожоговых больных и, следовательно, для результатов лечения. Было показано, что раннее иссечение раны и пересадка тканей улучшают гиперметаболический ответ и повышают выживаемость после ожоговых повреждений. Для пересадки на место иссеченной раны может быть использована собственная кожа (донорский сайт), однако, такой путь лечения неэффективен у больных с большой поверхностью ожога. В таких случаях используют синтетические кожные материалы или трупную кожу.
Покрытие раны необходимо отличать от закрытия раны. Материалы для закрытия ран являются биологически приемлемыми для раневого ложа и постепенно включаются в заживающую раневую область. С другой стороны, материалы для раневого покрытия основаны на включении в раневой коагулят и врастании гранулезных тканей с целью прикрепления: такое явление типично для многих материалов для покрытия ран. Материал для покрытия ран обычно не является биологически разрушаемым и, следовательно, может являться лишь временным замещением эпидермиса. Материалы для покрытия ран, следовательно, должны быть заменены на кожу пациента в результате либо реэпителизации, либо пересадки кожи. В случае использования временного покрытия рана не должна быть обсеменена бактериями и должна быть достаточно неглубокой, чтобы можно было ожидать ее полного заживления в течение 3 недель. Эпителиальные клетки из эпидермальных дериватов растут и замещают разрушенный эпидермис, и постепенно материал раневого покрытия вытесняется. Следовательно, исходной целью использования материалов для покрытия ран в случае неглубоких ожогов второй степени является сокращение микробного обсеменения раневого ложа (создание микробного барьера), обеспечивающее предотвращение инфекции, а также сокращение доступа воздуха, что тем самым ослабляет боль.
Материалы для покрытия ран также используют для глубоких повреждений второй степени или третьей степени перед окончательным закрытием раны собственной кожей больных с обширными ожоговыми повреждениями. Оптимальный материал раневого покрытия пока не установлен. Однако требования к материалам для закрытия ран связаны с имитацией нормальных дермы и эпидермиса. В частности, требования к современному материалу для закрытия ран таковы: (а) создать нетоксичный, антисептический, невоспалительный и неантигенный барьер для бактерий и других микробов; (b) обеспечить нормальную передачу тепла и воды; (с) обеспечить непосредственное, однообразное и плотное прикрепление к раневому ложу; (d) обеспечить поддержание механизмов нормального местного иммунитета и заживления раны; (e) поддержать эластичность и долговременную прочность; (f) сохранить возможность к росту; и (g) выполнить долговременные механические и косметические функции наряду с контрактурными свойствами раневой области, сравнимые с таковыми у полнослойного трансплантата.
Очень хорошие параметры биологической совместимости и заживления демонстрируют INTEGRA™, ALLODERM™ или BIOBRANE™. Однако указанные материалы очень дороги, что ограничивает их широкое применение. Трупная кожа является относительно эффективным и дешевым материалом для покрытия ран. Однако существенным оказывается риск переноса ВИЧ, ЦМВ, вируса простого герпеса и гепатита: следовательно, это ограничивает использование трупной кожи.
Плодные оболочки характеризуются многими преимущественными свойствами, которые делают такой материал применимым в качестве материала для раневого покрытия, включая: (а) низкую иммуногенность; (b) нетоксичность, антисептичность и невоспалительность; (с) отсутствие инфицированности ВИЧ, вирусом простого герпеса или ЦМВ; (d) неограниченное количество (что, следовательно, делает плодную оболочечную ткань недорогой альтернативой для существующих коже заменителей); (е) варьирование длины, диаметра и толщины; и (f) присутствие эндогенных механических компонентов, таких как коллаген, ламинин и фибронектин, что гарантирует механическую стабильность, поддержку роста и потенциальное сходство с таковыми признаками нормальной человеческой кожи.
С биохимической точки зрения тепловые повреждения представляют собой особенно тяжелую форму травмы, сопровождающуюся гиперметаболическим ответом, отличающимся высоким минутным сердечным выбросом, повышенным потреблением кислорода, нарушенным иммунным ответом и белковым и жировым катаболизмом [34]. Ожоговая рана поддерживает такое уязвимое гиперметаболическое состояние за счет выработки и секреции тромбоксана и провоспалительных цитокинов [35-37]. Следовательно, заживление раны важно для выживаемости и выздоровления ожоговых больных [22, 38-39]. Как было показано, анаболические средства, такие как гормон роста и инсулиноподобный фактор-1 роста, ослабляют гиперметаболический ответ и улучшают заживление раны [35, 39-41].
Инсулиноподобный фактор-1 роста (IGF-I) - небольшой полипептид с молекулярной массой примерно 7,5 кД - является анаболическим агентом, для которого было продемонстрировано улучшение метаболизма [35], функции слизистой кишечника [42] и белковых потерь [43] после теплового повреждения. IGF-I опосредует активности гормона роста при гиперметаболическом состоянии за счет ослабления потерь безжировой массы тела, нарушенного иммунного ответа, острой фазы ответа и за счет улучшения заживления раны [35, 38, 44-47]. Обработка IGF-I улучшает заживление раны за счет стимуляции образования коллагена и митогенной активности фибробластов и кератиноцитов [40, 41, 48]. Имеются неблагоприятные побочные эффекты, такие как гипогликемия, изменения психического состояния, отек, утомление и головные боли, которые ограничивают терапевтическое применение IGF-I в лечении ожогов [49, 50]. Указанные неблагоприятные побочные проявления наиболее вероятно обусловливаются сверхфизиологическими дозами свободного IGF-I, которые необходимы для достижения биологической эффективности [49, 50].
Первостепенным является выбор подходящей системы для доставки гена [1, 2]. В качестве ген-доставочных систем были использованы вирусы, в частности, аденовирусы благодаря их способности к специфичной трансфекции [1-3]. Однако вирусы проявляют связанные с вирусной инфекцией токсичность, нарушение иммунитета и вероятные мутагенные или канцерогенные эффекты, что делает данный подход потенциально опасным [1]. Таким образом, использование липосом в качестве доставочной системы становится привлекательной моделью благодаря их невирусному составу, стабильности и способности взаимодействовать с клеточной мембраной [4]. Добавление катионных свойств к стандартной липосомной структуре и включение холестерина наряду с использованием цитомегаловирусных (ЦМВ) промоторов в кДНК-конструкциях, используемых для переноса генов, повышает эффективность и уровни экспрессии трансгенов равным образом с теми, которые достигаются с использованием аденовирусных конструкций [4, 5].
В известном уровне техники имеется недостаток способов улучшения заживления ран и усовершенствованных материалов для раневых покрытий. Настоящее изобретение восполняет указанную, существовавшую в течение длительного времени потребность и необходимость в данной области техники.
Краткое содержание изобретения
Настоящее изобретение описывает способ улучшения заживления ран с использованием липосом, несущих гены, которые кодируют факторы усиления роста. Кроме того, настоящее изобретение описывает усовершенствованный материал для покрытия ран, пропитанный липосомами, несущими гены, которые экспрессируют факторы роста, для целей улучшения заживления ран. Объектом настоящего изобретения является снижение гиперметаболического ответа и, соответственно, улучшение результатов лечения и повышение выживаемости после травм, в частности, тепловых повреждений.
Настоящее изобретение описывает внесение липосомных генных конструкций непосредственно в рану и/или в материал покрытия для улучшения заживления раны и усиления функциональности материала для раневого покрытия. Также настоящее изобретение описывает применение усовершенствованной плодной человеческой амниотической оболочки в сочетании с липосомно-генными конструкциями, экспрессирующими факторы роста в качестве непостоянного материала для раневого покрытия при полнослойном заживлении ран. Плодная оболочка обладает преимуществами по сравнению с используемыми в настоящее время материалами, такими как Integra™, Biobrane™, Alloderm™, и определяет эффективный и безопасный способ для улучшения результатов лечения.
Одним из объектов настоящего изобретения является представление способов улучшения заживления ран, способов усовершенствования материала для раневого покрытия и усовершенствованного материала для раневого покрытия.
В варианте настоящего изобретения предусматривается способ улучшения заживления ран, включающий стадию инъецирования в рану липосомы, причем липосома включает по крайней мере один ген, кодирующий фактор усиления роста.
В другом варианте настоящего изобретения предусматривается способ улучшения заживления ран, включающий стадии: покрытия раны материалом для раневого покрытия, причем материал для раневого покрытия пропитан липосомами, причем липосомы включают по крайней мере один ген, который кодирует фактор усиления роста.
Еще в одном варианте настоящего изобретения предусматривается способ улучшения заживления ран, включающий стадии: покрытия раны материалом для закрытия ран, причем материал для закрытия ран пропитан липосомами, причем липосомы включают по крайней мере один ген, который кодирует фактор усиления роста.
В следующем варианте настоящего изобретения предусматривается усовершенствованный материал для перевязывания ран, включающий: материал для раневого покрытия и липосому, содержащую по крайней мере один ген, который кодирует фактор усиления роста.
В другом варианте настоящего изобретения предусматривается композиция для улучшения заживления ран, содержащая: липосому, причем липосома содержит по крайней мере один ген, который кодирует фактор усиления роста, и фармацевтически приемлемый носитель.
Другие и последующие аспекты, свойства и преимущества настоящего изобретения будут ясны из нижеследующего описания предпочтительных путей воплощения настоящего изобретения. Указанные варианты воплощения приведены для целей описания.
Краткое описание чертежей
Прилагающиеся чертежи включены здесь для того, чтобы указывавшиеся выше свойства, преимущества и объекты изобретения стали бы ясными и могли быть поняты в деталях. Указанные чертежи образуют часть настоящей заявки. Необходимо отметить, однако, что прилагающиеся чертежи иллюстрируют предпочтительные варианты воплощения изобретения и не должны рассматриваться как ограничивающие объем настоящего изобретения.
На фигуре 1 схематически показан эскиз мест проведения инъекций. Животные с одним местом для инъекций получали инъекцию только по сайту I, в то время как животные с двумя инъекциями получали инъекции по инъекционным сайгам I и II. Кожные биопсии I, II и III были взяты для анализа через 33 дня после ожога.
На фигуре 2 схематически показана плазмида с кДНК IGF-I (pcDNA3) под контролем промотора CMV. Структурно сходную плазмиду для β-галактозидазы (ген lacZ) упаковывали в ту же липосому.
На фигуре 3 отображены процентные изменения массы тела, определенные в течение 7-дневного анализа. *- Статистически достоверное различие между группами при р<0,05. Данные выражены как среднее ± s.e.m.
На фигуре 4 изображены гистологические срезы кожи после гистохимической реакции на активность β-галактозидазы с усиливающим окрашиванием эозином. (4А). Тонкогранулярный зелено-голубой продукт реакции присутствует внутри многих миофибробластов и гистиоцитозных клеток в гранулярной ткани, подстилающей ожоговую рану. Увеличение 380х. (4В). В инъецированной физиологическим раствором (контрольной) кожной ткани, подстилающей поврежденную кожу рядом с ожоговой раной, продукта реакции не выявлено. Увеличение 380х.
На фигуре 5 показано присутствие мРНК IGF-I в коже после трансфекции, липокоплексами, содержащими кДНК, которая кодирует IGF-I. Не было выявлено мРНК IGF-I в кожных биопсийных пробах, взятых у крыс, трансфицированных “чистыми” липосомами или физиологическим раствором (дорожки А и В). Имелось существенное количество мРНК IGF-I в кожных биопсиях от крыс, трансфицированных кДНК IGF-I (дорожка С). Показан репрезентативный образец.
На фигуре 6 показаны концентрации сывороточного трансферрина в ходе 7-дневного периода исследований. После ожогового повреждения сывороточный трансферрин снижался. Липосомы ослабляли это уменьшение на 2-й и 5-й дни после ожога. * - Статистически значимое различие между группами: р<0,05. Данные приведены как среднее ± s.e.m. (Нормальный уровень сывороточного трансферрина: >72 мг/дл).
На фигуре 7 показан сывороточный кислый α1-гликопротеин с 1-го по 7-й день после теплового повреждения. * - Статистически значимое различие между вариантами физиологического раствора и липосом при р<0,05. Данные приведены как среднее ± s.e.m. (Нормальный уровень сывороточного кислого α1-гликопротеина: 55-70 пкг/мл).
На фигуре 8 показан сывороточный TNF-α с 1-го по 7-й день после теплового повреждения. * - Статистически значимое различие между вариантами физиологического раствора и липосом при р<0,05. Данные приведены как среднее ± s.e.m. (Нормальный уровень сывороточного TNF-α: 1-10 пкг/мл).
На фигуре 9 показан сывороточный IL-1β с 1-го по 7-й день после теплового повреждения. *- Статистически значимое различие между вариантами физиологического раствора и липосом при р<0,05. Данные приведены как среднее ± s.e.m. (Нормальный уровень сывороточного IL-1β: 4-20 пкг/мл).
На фигуре 10 показаны результаты планиметрического измерения области реэпителизации раны. Крысы, получавшие упакованные конструкции кДНК IGF-I, проявили наивысший процент реэпителизации за время анализа по сравнению с группами липосом или физиологического раствора. *- Липосомы-кДНК против липосомы и физиологического раствора: р<0,05. Данные приведены как среднее ± s.e.m.
На фигуре 11 показаны результаты планиметрического измерения области реэпителизации раны. Крысы, получавшие множественные инъекции упакованных конструкций кДНК IGF-I, проявили наивысший процент реэпителизации за время анализа по сравнению с однократными инъекциями. * - Множественные инъекции кДНК IGF-I против однократных инъекций: р<0,05. Данные приведены как среднее ± s.e.m.
На фигуре 12 показано, что присутствие β-галактозидазного белка было выявлено с помощью теста на хемолюминесценцию репортерного гена в кожных биопсиях I, II и III. (12A). Крысы, получавшие однократную инъекцию кДНК-конструкции, проявили существенное снижение экспрессии β-галактозидазы вдоль края раны. *- Статистически значимое различие между кожными биопсиями I и III: p<0,05. (12В). Крысы, получавшие множественные инъекции, показали существенно повышенные уровни экспрессии β-галактозидазы. Различия между кожными биопсиями I, II или III отсутствовали. Данные приведены как среднее ± s.e.m.
На фигуре 13 показана концентрация белка IGF-I в кожных биопсиях I, II и III, которая была измерена с помощью РИА. (13А). Крысы, получавшие однократную инъекцию, показали снижение концентрации IGF-I от I-й к III-й биопсии. * - Статистически значимое различие между кожными биопсиями I и III: p<0,05. (13В). Животные, получавшие множественные инъекции, показали стабильно высокие уровни IGF-I. Данные приведены как среднее ± s.e.m.
Подробное описание изобретения
Настоящее изобретение демонстрирует преимущества использования доставочных на основе липосом векторов, экспрессирующих факторы усиления роста, в отношении травматических ран, в частности, тепловых повреждений, у крыс. Кроме того, в настоящем изобретении сравнивается функциональность различных материалов для раневого покрытия, таких как плодная оболочка, человеческая кожа и некоторые имеющиеся в продаже синтетические материалы, у свиней миниатюрной йоркширской породы.
Более конкретно, настоящее изобретение описывает внесение липосомных генных конструкций, кодирующих факторы усиления роста, например, инсулиноподобный фактор-1 роста (IGF-I) или фактор роста кератиноцитов (KGF), непосредственно в раневое ложе или в материал для покрытия раны, и анализ эффективности по сравнению с ранами, не обрабатывавшимися липосомами. Эффективность липосомной генотерапии оценивают путем измерения скорости восприятия и времени заживления, анализа сужения раны гистологическими методами и иммунных маркеров отторжения покрывающего материала и степени гиперметаболического ответа с недельными интервалами в течение 4-месячного периода.
Основываясь на описанных здесь данных, генотерапия, в которой один или большее число генов, экспрессирующих фактор(ы) роста, доставляют с помощью липосомных систем, инъецированных непосредственно в рану или внесенных в материал для покрытия ран, который после этого применяют на рану, сможет реформировать и улучшить текущее лечение тепловых повреждений, кожных язв и процедур кожной трансплантации.
Настоящее изобретение направлено на способы улучшения заживления ран и усовершенствованные материалы для раневого покрытия.
Также настоящее изобретение нацелено на способ улучшения заживления ран, включающий стадию инъецирования в рану липосомы, причем липосома включает по крайней мере один ген, кодирующий фактор усиления роста.
Кроме того, настоящее изобретение направлено на способ улучшения заживления ран, включающий стадии: покрытие раны материалом для раневого покрытия, причем материал для раневого покрытия пропитан липосомами, причем липосома включает по крайней мере один ген, кодирующий фактор усиления роста.
Еще в одном аспекте настоящее изобретение также нацелено на способ улучшения заживления ран, включающий стадии: покрытие раны материалом для закрытия ран, причем материал для закрытия ран пропитан липосомами, причем липосома включает по крайней мере один ген, кодирующий фактор усиления роста.
В следующем аспекте настоящее изобретение также направлено на усовершенствованный материал для перевязывания ран, содержащий: материал для раневого покрытия и липосому, включающую по крайней мере один ген, который кодирует фактор усиления роста. Обычно липосому, включающую фактор усиления роста, вносят в материал для покрытия ран с помощью инъекции, что может быть сделано до или после применения на рану материала для раневого покрытия.
Еще в одном аспекте настоящее изобретение направлено на композицию для улучшения заживления ран, содержащую: липосому, причем липосома включает по крайней мере один ген, кодирующий фактор усиления роста, и фармацевтически приемлемый носитель. Композиция по данному варианту воплощения может быть упакована таким образом, чтобы композицию можно было легко набрать в шприц, или, как альтернатива, она может быть упакована непосредственно в шприц.
В указанных вариантах воплощения характерными ранами, которые можно лечить с использованием композиций и способов по настоящему изобретению, являются тепловая травма, химическая травма, резаная рана, хирургическая травма или ссадина. Предпочтительными липосомами являются холестерин-содержащие катионные липосомы. В целом, факторами усиления роста являются гормон роста, инсулиноподобный фактор-1 роста, фактор роста кератиноцитов, фактор роста фибробластов, эпидермальный фактор роста, тромбоцитарный фактор роста или трансформирующий β-фактор роста. Предпочтительно фактором усиления роста является инсулиноподобный фактор-1 роста (IGF-I), а концентрация гена, кодирующего IGF-I в липосоме, составляет примерно 2,2 мкг на 1 мл липосом.
Характерными материалами для раневого покрытия являются человеческая плодная амниотическая оболочка, человеческий плодный хорион, трупная человеческая кожа, синтетическая кожа и другие материалы, известные специалистам в данной области техники. Характерными материалами для закрытия ран являются человеческая плодная амниотическая оболочка, человеческий плодный хорион, сингенная кожа человека, аллогенная человеческая кожа и другие материалы, хорошо известные специалистам в данной области техники.
В соответствии с настоящим изобретением могут быть использованы стандартные методы молекулярной биологии, микробиологии и рекомбинантных ДНК, известные специалистам в данной области техники. Такие методы полно представлены в научной литературе: см., например, Sambrook, Fritsch & Maniatis, 1982, "Molecular Cloning: A Laboratory Manual"; "DNA Cloning: A Practical Approach", Volumes I & II (ed. D.N.Glover), 1985; "Oligonucleotide Synthesis" (ed. M.J.Gait), 1984; "Nucleic Acid Hybridization" (eds. B.D.Hames & S.J.Higgins), 1985; "Transcription and Translation" (eds. B.D.Hames & S.J.Higgins), 1984; "Animal Cell Culture" (ed. R.I.Freshney), 1986; "Immobilized Cells and Enzymes" (IRL Press), 1986; B. Perbal, 1984, "A Practical Guide to Molecular Cloning". Следовательно, следующие термины, используемые в настоящем описании, должны быть определены в соответствии с приведенным далее.
“Молекула ДНК” обозначает полимерную форму дезоксирибонуклеотидов (аденина, гуанина, тимина или цитозина) либо в одноцепочечной форме, либо в форме двухцепочечной спирали. Данный термин относится исключительно к первичной и вторичной структуре молекулы и не ограничивает ее какими-либо конкретными пространственными формами. Таким образом, данный термин охватывает двухцепочечную ДНК, выявляемую, помимо прочего, в линейных молекулах ДНК (например, рестрикционных фрагментах), вирусах, плазмидах и хромосомах. При обсуждении структуры в данном тексте в соответствии с принятым стандартом указывается только последовательность в направлении 5’→3’ по не транскрибируемой цепи ДНК (т.е. цепи, имеющей последовательность, гомологичную мРНК).
“Вектор” представляет собой репликон, такой как плазмида, фаг или космида, к которому может быть присоединен другой сегмент ДНК таким образом, чтобы осуществлялась репликация присоединенного сегмента. “Репликоном” является любой генетический элемент (например, плазмида, хромосома, вирус), который функционирует в качестве автономной единицы репликации ДНК in vivo, т.е. он способен реплицироваться под своим собственным контролем. “Сайт (точка) начала репликации” относится к тем последовательностям ДНК, которые участвуют в синтезе ДНК. “Последовательностью-регулятором экспрессии” является последовательность ДНК, которая контролирует и регулирует транскрипцию и трансляцию другой последовательности ДНК. Кодирующая последовательность “функционально присоединена” и “находится под контролем” последовательностей транскрипционных и трансляционных регуляторов в клетке тогда, когда РНК-полимераза транскрибирует кодирующую последовательность с образованием мРНК, которая затем транслируется в белок, кодируемый данной кодирующей последовательностью.
В целом, экспрессирующие векторы, включающие промоторные последовательности, которые обеспечивают эффективную транскрипцию и трансляцию встроенного ДНК-фрагмента, используют в сочетании с организмом-хозяином. Обычно экспрессирующий вектор включает сайт начала репликации, промотор(ы), терминатор(ы), а также конкретные гены, которые способны обеспечивать отбор трансформированных клеток по фенотипу. Трансформированные организмы-хозяева могут быть подвергнуты ферментированию и культивированию с применением методов, известных в данной области техники, с достижением оптимального роста клеток.
“Кодирующей последовательностью” ДНК является двухцепочечная последовательность ДНК, которая транскрибируется и транслируется в полипептид in vivo тогда, когда она помещена под контроль соответствующих регуляторных последовательностей. Границы кодирующей последовательности определяются старт-кодоном с 5’-(N-) конца и стоп-кодоном трансляции с 3’-(С-) конца. Кодирующая последовательность может включать, тем самым не ограничиваясь, прокариотические последовательности, кДНК на матрице эукариотической мРНК, последовательности геномной ДНК эукариот (например, млекопитающие) и даже синтетические последовательности ДНК. Сигнал полиаденилирования и транскрипционный терминатор обычно должны быть расположены с 3’-конца кодирующей последовательности. Термин “кДНК” определяет ДНК-копию или комплементарную ДНК, которая является продуктом реакции обратной транскрипции на матрице мРНК-транскрипта. “Экзон” обозначает экспрессированную последовательность, транскрибированную с генного локуса, в то время как “интрон” является неэкспрессированной последовательностью из состава генного локуса.
Последовательностями транскрипционных и трансляционных регуляторов являются регуляторные ДНК-последовательности, такие как промоторы, энхансеры, сигналы полиаденилирования, терминаторы и подобное, которые обеспечивают экспрессию кодирующей последовательности в клетке-хозяине. “Цис-элементом” является нуклеотидная последовательность, также обозначаемая как “консенсусная последовательность” или “мотив”, которая взаимодействует с другими белками, способными позитивно регулировать или негативно регулировать (усиливать или ослаблять) экспрессию конкретного генного локуса. Также в состав кодирующей последовательности может быть включена “сигнальная последовательность”. Такая последовательность кодирует сигнальный пептид, находящийся с N-конца полипептида, который сигнализирует клетке-хозяину и направляет полипептид в соответствующий клеточный сайт. Сигнальные последовательности можно найти в связи с различными белками, нативными для прокариот и эукариот.
“Промоторной последовательностью” является ДНК-регуляторный сегмент, способный связывать РНК-полимеразу в клетке и инициировать транскрипцию расположенной далее (в направлении 3’) кодирующей последовательности. Для целей определения настоящего изобретения промоторная последовательность ограничена по своему 3’-концу сайтом инициации транскрипции и продолжается вверх (в направлении 5’) так, чтобы включить минимальное количество нуклеотидов или элементов, необходимых для инициации транскрипции на уровнях, выявляемо превышающих фоновый уровень. В составе промоторной последовательности должен находиться сайт инициации транскрипции, а также домены связывания белков (консенсусные последовательности), ответственные за связывание РНК-полимеразы. Часто, но не всегда, эукариотические промоторы включают боксы ТАТА и боксы CAT. Прокариотические промоторы включают последовательности Шайна-Дальгарно в дополнение к консенсусным последовательностям в положениях [-10] и [-35].
В целом, термин “ген” призван охватить промотор и регуляторные элементы, функционально присоединенные к кодирующей последовательности. Такие промоторные последовательности и регуляторные элементы могут являться нативными для гена промотором и/или регуляторными элементами или могут быть гетерологичными.
“Гетерологичным” участком является идентифицируемый сегмент ДНК в пределах более крупной молекулы ДНК, который не обнаруживается в связи с такой более крупной молекулой в природе. Так, когда гетерологичный участок кодирует ген млекопитающего, ген обычно должен быть фланкирован (например, промоторными последовательностями и/или регуляторными элементами) такой ДНК, которая не фланкирует геномную ДНК млекопитающего в геноме организма-источника. В другом примере кодирующей последовательностью является конструкция, в которой кодирующая последовательность сама по себе в природе не обнаруживается (например, кДНК, в которой геномная кодирующая последовательность включает интроны, или синтетические последовательности, включающие кодоны, которые отличаются от таковых в составе нативного гена). В соответствии с определенным здесь аллельные варианты или встречающиеся в естественных условиях мутационные события не приводят к образованию гетерологичного участка ДНК.
“Технология рекомбинантных ДНК” указывает на методы объединения двух гетерологичных молекул ДНК, обычно в результате лигирования in vitro ДНК от разных организмов. Молекулы рекомбинантной ДНК обычно получают экспериментально методами генетической инженерии. Терминами-синонимами являются “сплайсинг генов”, “молекулярное клонирование” и “генетическая инженерия”. Продуктом таких манипуляций является “рекомбинант” или “рекомбинантная молекула”.
Клетка считается “трансформированной” или “трансфицированной” экзогенной или гетерологичной ДНК тогда, когда такая ДНК была внесена внутрь клетки. Трансформирующая ДНК может быть или может не быть встроена (путем ковалентного присоединения) в геном клетки. У прокариот, дрожжей и клеток млекопитающих, например, трансформирующую ДНК можно поддерживать в виде эписомного элемента, такого как вектор или плазмида. В отношении эукариотических клеток стабильно трансформированной клеткой является клетка, в которой трансформирующая ДНК была встроена в хромосому таким образом, что она наследуется дочерними клетками в результате репликации хромосом. Такая стабильность демонстрируется способностью эукариотической клетки формировать клеточные линии или клоны, составляющие популяцию дочерних клеток, несущих трансформирующую ДНК. “Клоном” является популяция клеток производных от единственной клетки или предка в результате митозов. “Клеточной линией” является клон первичной клетки, который способен стабильно расти in vitro в течение многих поколений. Организм, такой как растение или животное, который был трансформирован экзогенной ДНК, называют “трансгенным”.
По использованию в данном тексте термин “амниотическая оболочка” обозначает тонкую экстраэмбриональную оболочку, окружающую эмбрион и производную от эктодермальных и мезодермальных тканей.
По использованию в данном тексте термин “хорион” обозначает наиболее внешнюю экстраэмбриональную оболочку, которая в конечном счете является частью плаценты, в которой она в дополнение к дыхательным функциям доставляет пищевые компоненты и выводит шлаки.
По использованию в данном тексте термин “липосома” обозначает маленький пузырек, окруженный двухслойной липидной мембраной и искусственно полученный из фосфолипидов. ДНК, белки и другие материалы могут быть упакованы внутрь липосомы и внесены в клетки животного путем слияния с плазматической мембраной. По использованию в данном тексте термин “холестериновая катионная липосома” обозначает непосредственно холестеринсодержащую липосому.
По использованию в данном тексте “термин фактор усиления роста” обозначает соединения, которые стимулируют рост.
По использованию в данном тексте термин “скорость восприятия” обозначает “скорость заживления” или “скорость принятия” донорской ткани на ткани реципиента.
В частности, предусматривается, что фармацевтические композиции могут быть приготовлены с использованием липосом, описанных в настоящем изобретении. В таком случае фармацевтическая композиция содержит ген, кодирующий фактор роста, липосому по настоящему изобретению и фармацевтически приемлемый носитель. Для специалиста в данной области техники не составит труда без дополнительных экспериментов определить соответствующие дозировки и пути введения липосомы по настоящему изобретению. При использовании для лечения больного липосомный носитель, описанный в настоящем изобретении, вводят больному или животному в терапевтически эффективных количествах, т.е. количествах, которые эффективно доставляют соответствующие количества ДНК, кодирующей фактор усиления роста. Обычным будет введение инъецируемой формы в рану или в материал для раневого покрытия, хотя и другие пути введения можно использовать в качестве подходящих. Доза и режим введения будут зависеть от природы раны (тяжести повреждения тканей) и ее размера, анамнеза больного и других факторов. Количество вводимых липосом будет в основном определяться площадью раны при введении липосом примерно 4 см/часть. Противодействуя отрицательным последствиям лечения, лечение может быть продолжено для достижения оптимальной эффективности: см. Remington’s Pharmaceutical Science, 17th ed., 1990, Mark Publ. Co., Easton, PA; и Goodman & Gilman’s "The Pharmacological Basis of Therapeutics" 8th ed., 1990, Pergamon Press: включены здесь для сведения в виде библиографических ссылок. Для местного применения липосомы наиболее обычно приготавливают в виде стандартной дозы инъецируемой формы (раствора, суспензии, эмульсии) в сочетании с фармацевтически приемлемым носителем. Такой носитель предпочтительно нетоксичен и не обладает лечебными свойствами. Примерами таких носителей являются вода, физиологический раствор, раствор Рингера, раствор декстрозы и 5%-ный человеческий сывороточный альбумин. Также можно использовать неводные носители, такие как фиксированные масла и этилолеат. Носитель может включать небольшие количества добавок, таких как вещества, которые усиливают изотоничность и химическую стабильность, например, буферы и консерванты. Обычно липосомы нужно приготавливать в таких носителях в концентрации примерно 1-10 мкг.
Нижеследующие примеры даны для цели иллюстрирования различных вариантов воплощения изобретения и не призваны в чем бы то ни было ограничить настоящее изобретение.
Пример 1
Экспериментальные животные – крысы
Взрослых самцов крыс линии Sprague-Dawley (350-375 г) помещали в садки с проволочным дном и выдерживали в помещении с контролируемой температурой при 12-часовом световом дне. Животных акклимировали к соответствующей среде в течение 7 дней перед началом слепого анализа. Все они получали равные количества жидкого рациона от Sustacal (Mead Johnson Nutritionals, Evansville, IN, США) и воду без ограничений на протяжении всего эксперимента. Каждая крыса получала полнослойный паровой ожог 60% от общей площади поверхности тела (ОППТ). Затем крыс с тепловым повреждением случайным образом разделяли на:
(a) 2 группы получали инъекции холестерин-содержащих катионных липосом (20 мкл липосом в 180 мкл физиологического раствора: n=28) или физиологического раствора (контроль, 200 мкл: n=28);
(b) 2 группы получали еженедельные подкожные инъекции липосом (10 мкл липосом в 180 мкл физиологического раствора), включающих 2,2 мкг конструкции кДНК IGF-I, и 0,2 мкг конструкции кДНК репортерного гена β-галактозидазы lacZ под контролем промотора CMV (n=12) в одном сайте инъекции по краю ожоговой раны (фиг.1), или еженедельные подкожные инъекции липосом (10 мкл липосом в 180 мкл физиологического раствора), содержащих 2,2 мкг конструкции кДНК IGF-I под контролем цитомегаловируса и 0,2 мкг репортерного гена β-галактозидазы, кДНК lacZ (n=12), в двух сайтах инъекции по краю ожоговой раны (фиг.1); или
(с) 3 группы получали еженедельные подкожные инъекции физиологического раствора (200 мкл нормального физиологического раствора: n=10); еженедельные подкожные инъекции липосом (10 мкл липосом в 180 мкл физиологического раствора), включающих 0,2 мкг конструкции кДНК репортерного гена β-галактозидазы lacZ (n=10); или еженедельные подкожные инъекции липосом (10 мкл липосом в 180 мкл физиологического раствора), содержащих 2,2 мкг конструкции кДНК IGF-I и 0,2 мкг конструкции кДНК репортерного гена β-галактозидазы lacZ (n=10).
Пример 2
Экспериментальные животные - свиньи
Механизмы заживления ран у миниатюрных йоркширских свиней близки к таким механизмам у людей. Поэтому миниатюрные йоркширские свиньи являются хорошо исследованной моделью: их использовали в ряде экспериментальных исследований травм. Каждая из 10 йоркширских свиней получала 4 полнослойные раны (стандартная модель) под наркозом и с обезболиванием. Каждый раневой участок имел квадратную или прямоугольную форму при приблизительных размерах 8×8 см, при расстоянии между участками приблизительно 5 см, и находился в области верхней части боков и на спине. Раны должны были располагаться и размещаться в соответствии с конкретной схемой таким образом, чтобы животное могло нормально ложиться после высвобождения из ограничивающей подвесной повязки.
Сразу после индукции раны ее накрывают препаратом человеческого плодного амниона/хориона, INTEGRA™ (Life Science), AL-LODERM™ (Life-Cell) или BIOBRANE™ (Dow-Hickhan). Покрытие прикрепляют к необожженному участку раны и покрывают марлей, пропитанной мазью с тремя антибиотиками, и плотной повязкой. Скрепки и/или швы используют по необходимости.
После хирургического вмешательства анализируемых животных удерживали в подвесной повязке до выхода из-под наркоза так, чтобы защитить трансплантаты от повреждения или смещения. Животные оставались в указанной подвесной повязке не более 24 часов.
Пример 3
Липосомы
Использованные липосомы являлись холестеринсодержащими катионными липосомами, реагент DMRIE-C (1,2-димиристилокси-пропил-3-диметилгидроксилэтиламмонийбромид) приготавливали с отфильтрованной на холестериновой мембране водой (Life Technologies, Rockville, MD). Конструкцию кДНК IGF-I (фиг.2), включавшую плазмиду с кДНК IGF-I под контролем цитомегаловируса, получали в UTMB Sealy Center при Molecular Science Recombinant DNA Core Facility (кДНК IGF-I была любезно предоставлена G.Rotwein, NIH, Bethesda, MD). Использованные дозы составили 10% липосом (наивысшая концентрация, использованная в экспериментах по переносу ДНК, которая не приводит к неблагоприятным последствиям в отношении растворимости ДНК и совместима с принципами генной доставки). С целью определения периода действия липосом липосомы инъецировали внутривенно в хвостовую вену крысам группы (а) через полчаса после теплового повреждения, а изменения отслеживали в течение 7-дневного периода. Объем 200 мкл составляет количество, которое можно вводить в хвостовую вену крысы, не вызывая неблагоприятных побочных действий, таких как остановка сердца. Непосредственно после теплового повреждения каждая крыса в группе (b) или (с) получала 0,2 мл растворов, инъецированных по одному из сайтов в 1 см от края раны или в двух сайтах в противоположных друг от друга участках раны (фиг.1). Указанную пропись повторяли один раз в неделю в течение 4 недель. Липокомплексы приготавливали свежими каждую неделю перед проведением инъекций.
Пример 4
Методика
Массу тела определяли в одно и то же время ежедневно. Процентное изменение общей массы тела в целом увеличивалась в первые 2 дня с последующим снижением массы тела через 2 дня после ожога. Крысы, получавшие липосомы, сохраняли свою массу тела лучше по сравнению с контролями, которым вводили физиологический раствор: р<0,05 (фиг.3). Крыс умерщвляли с помощью декапитации через 1, 2, 5 или 7 дней после ожога для особей группы (а), или, как альтернатива, через 5 дней после последней инъекции (33 дня после ожога) для крыс групп (b) или (с). Кровь собирали в сывороточные и плазматические сепараторы, центрифугировали при 1000 g в течение 15 минут, декантировали и замораживали при -73°С. Печень и икроножную мышцу выделяли, взвешивали, разрезали, и каждый из образцов высушивали при 60°С до достижения постоянной массы. Для оценки содержания белка использовали отношение сухого веса к весу сырой ткани. Три образца кожи спины, определенные как биопсические пробы I, II или III, собирали, быстро замораживали в жидком азоте и хранили при -73°С для анализа (Фиг.1). Между животными, которым вводили липосомы, и контрольными особями отсутствовали различия в отношениях сухого веса и веса сырой ткани для икроножной мышцы и печени.
Сывороточный холестерин, свободные жирные кислоты, сывороточные белки острой фазы (гаптоглобин и α2-макроглобулин), конститутивные печеночные белки (общий белок, трансферрин и альбумин) и глюкозу определяли с помощью нефелометра Behring (Behring, Dearfield, IL). Сывороточный кислый α1-гликопротеин определяли методом ТИФА (Wako Chemicals Inc., Richmond, VA). Калибровочная кривая для концентраций крысиного кислого α1-гликопротеина была прямой от 0 до 1500 пкг/мл на логарифмической шкале. Уровни плазменного TNF-α определяли методом ТИФА (Endogen, Woburn, MA). Калибровочная кривая, использованная для количественного определения TNF-α крысы, была прямой от 0 до 833 пкг/мл в логарифмической шкале. Уровни сывороточного IL-1β определяли методом ТИФА (Biosource Int., Camarillo, CA), и калибровочная кривая, использованная для количественного определения крысиного IL-1β, была прямой от 0 до 1500 пкг/мл в логарифмической шкале. Сывороточный IL-6 определяли в биотесте с использованием клеток В9 в логарифмической фазе роста (линия мышиной гибридомы), обработанных повышающимися концентрациями сыворотки. Пролиферацию клеток в ответ на добавляемую сыворотку измеряли методом количественной восстановительной MTS-спектрофотометрии.
Пример 5
Трансфекция
Трансфекцию контролировали путем измерения присутствия β-галактозидазы. Белок β-галактозидазы выявляли с помощью гистохимического окрашивания Bluo-gal в трех кожных биопсиях. Образцы кожи фиксировали в течение ночи при 4°С в фиксаторе, содержащем 4% параформальдегида в забуференном HEPES растворе Хенкса при рН 7,6. После промывки в буфере и фосфатно-солевом буфере (ФСБ) образцы инкубировали в течение ночи при 37°С в 0,1%-ном растворе субстрата Bluo-Gal (галогенированная индолил-β-D-галактозидаза) (Life Technologies, Gaithersburg, MD, США), забуференном до рН 7,6. После экстенсивной промывки ткани заливали парафином, приготавливали гистологические срезы, которые окрашивали гематоксилин-эозином или только эозином.
Присутствие белка β-галактозидазы также выявляли с помощью теста на хемолюминесценцию репортерного гена (GALACTO-LIGHT PLUS™ Tropix Inc., Bedford, MA, США) в коже. Образцы были приготовлены в соответствии с [55]: 100 мг ткани гомогенизировали в 200 мкл буфера для лизиса (40 мМ Трис (рН 7,5), 1 мМ ЭДТА, 150 мМ NaCl) приблизительно в течение 30 секунд. Образцы центрифугировали (12000 g) в течение 3 минут. Надосадочную фракцию отбирали, измеряли объем и хранили на льду. Оставшийся сгусток споласкивали в 200 мкл буфера для лизиса и микроцентрифугировали. Тест проводили в 96-луночных планшетах в соответствии с инструкциями производителя.
Мелкозернистый сине-зеленый продукт β-галактозидазной реакции в основном присутствовал в гранулированной ткани, составленной веретенообразными миофибробластами, макрофагами и растущими небольшими сосудами, расположенными под раной. Клетками, интенсивно окрашенными на β-галактозидазу, являлись миофибробласты, эндотелиальные клетки и макрофаги в областях воспаления, а также многоядерные гигантские клетки, что указывает на предпочтительную трансфекцию клеток, характеризующихся более высокой скоростью пролиферации (фиг.4А). Хотя наибольшее прокрашивание на β-галактозидазу характеризовало цитоплазму, некоторое количество продуктов реакции было выявлено за пределами границ клеток, что, по-видимому, обусловлено выходом ферментов из погибших клеток или простейшей диффузией продуктов реакции. Небольшое количество продукта реакции также было выявлено в матриксе волосяных фолликулов рядом с сайгами инъекций. Не было выявлено β-галактозидазы у животных, которым вводили физиологический раствор (фиг.4В).
Экспрессия белка β-галактозидазы усиливалась по периметру раны у крыс, получавших упакованные в липосому конструкции кДНК lacZ вместе с кДНК IGF-I по сравнению с крысами, которым вводили физиологический раствор (р<0,05: табл.1). Не было различий между группами по концентрации β-галактозидазы в клетках крови; печени, селезенки или почек. Такие данные согласуются с заключением, согласно которому системные клетки не подвержены трансфекции после подкожных инъекций кДНК (табл.2).
Экспрессия β-галактозидазы в коже
Экспрессия β-галактозидазы в крови, печени, почках и селезенке
Пример 6
IGF-I
У крыс, получавших конструкцию кДНК IGF-I, имелось подтверждение того, что мРНК IGF-I находится в коже поблизости от сайтов трансфекции, но только не в контрольных или имитационно обработанных тканях (фиг.5).
Концентрации белка IGF-I определяли с помощью РИА в трех кожных биопсиях (фиг.1). Белки экстрагировали путем пульверизации приблизительно 40 мг ткани под жидким азотом, добавления экстракционного буфера (ФСБ, 0,25 мл PMSF, 50 мг лейпептина, 100 мг апротинина и 50 мг антипаина) в объеме 1:7 (7 мл буфера на 1 г ткани) и гомогенизации смеси. Для достижения выделения белка образцы замораживали в течение ночи при -80°С. После оттаивания 50 мкл гомогената добавляли к 150 мкл экстракционного раствора и центрифугировали при 13500 об/мин в течение 5 минут. 100 мкл надосадочной фракции добавляли к 400 мкл нейтрализующего раствора, и РИА проводили в соответствии с описанным в руководствах к набору реактивов (Diagnostic System Laboratories, Webster, TX, США).
Во всех взятых кожных биопсиях у животных, которым вводили конструкции кДНК IGF-I, имелись более высокие концентрации белка IGF-I по периметру раны по сравнению с биопсиями, взятыми у животных, которым вводили пустые липосомы или физиологический раствор (р<0,05: табл.3). Отсутствовали различия между группами по концентрации белка IGF-I в сыворотке, печени, селезенке или почках (табл.4).
Концентрация белка IGF-I в коже
Концентрация белка IGF-I в крови, печени, почках и селезенке
Пример 7
Конститутивные печеночные белки, белки острой фазы и цитокины
Концентрация общего сывороточного белка снижалась после ожогового повреждения. Крысы, получавшие липосомы, характеризовались повышением общего сывороточного белка через 5 дней после ожога (липосомы: 5,6±0,1 г/дл, против контроля: 5,2±0,1 г/дл): р<0,05. Сывороточный трансферрин снижался после ожогового повреждения примерно на 30% ниже нормы. У крыс, которым вводили липосомы, показано повышение сывороточного трансферрина на 2-й и 5-й дни после ожога по сравнению с контролем (р<0,05; фиг.6). Отсутствовали достоверные различия по уровню сывороточного альбумина между обработанными и контрольными группами. Более того, отсутствие достоверных различий между обработанными и контрольными животными по уровням сывороточного холестерина, свободных жирных кислот и глюкозы можно было видеть в течение всего периода эксперимента.
Содержание белков острой фазы I типа - сывороточного гаптоглобина и кислого α1-гликопротеина - повышалось после теплового повреждения. Липосомы приводили к снижению уровня сывороточного кислого α1-гликопротеина через 1 и 5 дней после ожога (р<0,05; фиг.7). Не было выявлено достоверных различий в уровне сывороточного гаптоглобина у животных, которым вводили липосомы, и контроля. Количество белков острой фазы II типа возрастало после ожога почти на 50%. Не было обнаружено различий в уровне α2-макроглобулина между животными, которым вводили липосомы, и контрольными животными.
Количество всех исследованных цитокинов повышалось после теплового повреждения. Сывороточный IL-1β снижался в течение первого дня после ожога, а сывороточный TNF-α - на 1-й и 2-й дни после ожога у крыс, получавших липосомы, по сравнению с контролем (р<0,05; фиг. 8). Не было выявлено изменений в уровне сывороточного IL-6 у крыс, которым вводили липосомы, по сравнению с контрольными особями.
Пример 8
Биологическая эффективность
Все крысы выживали при паровом ожоге 60% ОППТ и инъекциях лекарственного средства при отсутствии неблагоприятных побочных эффектов. Заживление ран определяли следующим образом: раневой струп оставляли незатронутым в первые 28 дней и затем удаляли путем аккуратной вытяжки с условием, чтобы не повредить или не разрушить заживающий край по периферии раны. После удаления струпа животных помещали на размеченную поверхность, и область раны отпечатывали на ацетатные пластины вдоль отчетливо различимой реэпителизованной и необожженной границы и передним краем нового эпителия. Площади таких отпечатков подсчитывали с помощью компьютерного планиметра (Sigma Scan, программа Sigma Plot). Кроме того, кожные биопсии отбирали от края раны через 33 дня после ожога, и анализ с помощью светового микроскопа проводили с помощью известных методик. В гистологических измерениях линейной кожной реэпителизации использовали метод окрашивания гематоксилином-эозином.
Через 28 и 33 дня после теплового повреждения у крыс, получавших конструкцию кДНК IGF-I, была показана существенная интенсификация реэпителизации по сравнению с теми животными, которые получали пустые липосомы или физиологический раствор (р<0,05; фиг.10). Такое улучшение в заживлении раны наиболее вероятно обусловливается митогенной стимуляцией кератиноцитов и фибробластов под влиянием IGF-I, поскольку повышенная митогенная активность была обнаружена у крыс, которым вводили конструкцию кДНК IGF-I, по сравнению с теми крысами, которые получали пустые липосомы или физиологический раствор. Крысы, получавшие конструкцию кДНК IGF-I, характеризовались более высокими уровнями сывороточных белков и уровнями общего белка печени по сравнению с животными, получавшими липосомы или физиологический раствор (р<0,05; табл.5).
Концентрация общего белка в сыворотке и печени
Пример 9
Заготовка донорской ткани плодной амниотической оболочки и приготовление амниона и хориона
После информированного согласия, взятия истории болезни и оценки вероятных факторов риска, таких как раковые и инфекционные заболевания, злоупотребление лекарственными средствами и половое поведение, предполагаемого донора анализировали на гепатит В и С, RPR, ВИЧ-1, ВИЧ-2 (что стандартным способом выполняется с помощью кожного банка SBI и в соответствии с определенным в ААТВ). Указанный анализ выполняют во время родов и во время запланированного амбулаторного обследования через 60-90 дней после родов. Сразу после родов плаценты со сращенными плодными оболочками отбирают от подходящих доноров (в соответствии с руководством ААТВ). После предварительной промывки раствором Рингера в родовом помещении плаценту переносят в среду для тканевых культур (например, среду RPMI-1640), содержащую противобактериальные и противогрибковые средства, и переносят при 4°С в место обработки.
Стадии заготовки осуществляют с использованием стерильной методики таким образом, чтобы не происходило бактериального и грибкового загрязнения продукта. На каждом этапе обработки проводят микробиологический анализ так, чтобы контролировать загрязнение. На образцах тканей также могут быть проведены дополнительные анализы методом ПЦР, чтобы исключить вирусную инфекцию (например, ВИЧ-1 или ВИЧ-2).
В месте обработки (отделение тканевого банка SBI) проводят дополнительные промывку и ополаскивание плаценты раствором Рингера через пупочные сосуды. Амниотический и хорионический слои затем механическим путем отделяют друг от друга и от плаценты и покрывающий клеточный материал удаляют путем ферментной обработки трипсином (Boehringer Mannheim, Indianapolis, IN) (разведение 1:1 дистиллированной воды и трипсина) в течение 2 часов при 20°С с последующим повторным ополаскиванием фосфатно-солевым буфером (в соответствии с международной патентной заявкой WO 93/10722). Для сохранения структуры амнион сохраняют с использованием стандартных методов контролируемого по скорости замораживания. Для достижения различных параметров и характеристик ткани структуры плодных оболочек перекрестно сшивают действием 1,5%-ного раствора глутарового альдегида (Sigma, St.Louis, МО) в течение 20 минут при 20°С с последующей трехкратной промывкой в 1,5%-ном глицине (Sigma, St.Louis, МО) в течение 15 минут в соответствии с описанным для консервации тканей для Европейского кожного банка [61].
Приготовление для окончательного хранения включает лиофилизацию с использованием системы постепенной лиофилизации с двойной лиофилизационной процедурой, в результате применения которой содержание влаги составляет 6% воды или меньше, или консервацию путем смешивания плодных оболочек в 85%-ном глицерине при 20°С в течение 3 часов. Применение последней процедуры приводит к остаточной влажности 15% воды и обеспечивает инактивацию ВИЧ и других вирусов [62]. В результате последней обработки не остается жизнеспособных клеток и снижается антигенность, что выражается в уменьшенной иммунологической активности [63, 64]. Законсервированную в глицерине ткань хранят в глицерине при 4°С. Обе процедуры завершаются переносом ткани в емкость из фольги, что обеспечивает дальнейшую стерилизацию γ-облучением или газообразным этиленом. Перед использованием ткань реконституируют погружением в физиологический раствор (Рингера). Дополнительные материалы для раневого покрытия, такие как синтетическая ткань, содержащая коллагены I и II типов, и человеческая кожа, приготавливают для проведения сравнительного анализа (патент США №5002071; Walther et al., 1998).
Пример 10
Холестерин-содержащие катионные липосомы в качестве доставочной системы для генотерапии травмы ослабляют ответ острой фазы у крыс с тепловыми повреждениями
Одной из основных составляющих гиперметаболизма, ассоциированного с тепловым повреждением, является повышение содержания белков острой фазы и цитокинов [9, 10, 11]. В данном исследовании было показано, что введение холестерин-содержащих катионных липосом улучшает показатели массы тела и экспрессию конститутивных печеночных белков, трансферрина и общего белка после ожога по сравнению с физиологическим раствором. Дополнительно было показано, что липосомы снижают уровни сывороточных белков острой фазы I типа и связанных с белками острой фазы I типа сывороточных провоспалительных цитокинов TNF-α и IL-1β. По-видимому, снижение уровней TNF-γ и IL-1β приводит к снижению содержания указанных белков острой фазы.
Причинами благоприятных проявлений, связанных с липосомами, по-видимому, являются непосредственное влияние липидных составляющих липосом на мембраны поврежденных клеток или усиление поглощения внеклеточных питательных веществ с инкапсуляцией in situ и защита эндогенных факторов роста и цитокинов, выработанных в конкретном участке, что является частью гиперметаболического процесса, который опосредуется белками острой фазы и цитокинами. Провоспалительные цитокины, в частности, TNF-α, подавляют синтез белка и индуцируют потери веса [16-18]. После теплового повреждения, равно как и после сепсиса, сывороточные уровни TNF-α повышаются наряду с интенсификацией вызванного сепсисом мышечного протеолиза [19-21]. Сниженные уровни TNF-α в сыворотке связаны с улучшением суммарного белкового баланса и со снижением потерь массы тела у детей с тепловыми повреждениями [22, 23]. Таким образом, как показано в данном тексте, снижение уровня сывороточного TNF-α может сохранять массу тела и повышать содержание трансферрина в сыворотке.
В ряде исследований была предпринята попытка установления механизмов, действие которых обеспечивает проявление катионными липосомами ингибирующего влияния на экспрессию цитокинов in vitro, однако точные механизмы к настоящему времени не определены [25-30]. Представляется вероятным, что важную роль играет транскрипционный фактор кВ (NF-кВ) [27]. NF-кВ является транскрипционным сигналом и ключевым фактором в формировании клеточного иммунитета и воспалительного ответа [27]. Вследствие электростатических взаимодействий между клетками и катионными липосомами липосомы связываются с рецептором окисленных низкомолекулярных липопротеинов (OxLDL) [28, 29]. Было установлено (Sambrand et al.), что такое конкурентное связывание и последующая активация рецептора OxLDL косвенно подавляют активацию и/или связывание NF-кВ с соответствующим ему сайтом ДНК [28]. Более того, было показано (Aramaki et al.), что катионные липосомы подавляют фосфорилирование остатков тирозина в составе р41 - белка из семейства МАР-киназных транскрипционных факторов, - что в результате приводит к негативной регуляции NF-кВ [25-27]. Ингибирование р41 и NF-кВ подавляет индукцию индуцибельной синтетазы окиси азота (iNOS), что снижает экспрессию окиси азота (NO) [25, 26]. Данные изменения в сигнальной цепочке, как предполагалось, ответственны за избирательное подавление экспрессии TNF-γ на посттранкрипционном уровне [25, 26, 31]. С учетом того, что NO и iNOS стимулируют экспрессию IL-1β, представляется вероятным, что подавление активности NO или iNOS с участием катионных липосом приводит к снижению экспрессии IL-1β [32, 33]. Следовательно, введение катионных липосом очевидно является благоприятным в лечении воспалительных заболеваний, в частности, благодаря подавлению ими секреции провоспалительных цитокинов [31, 34].
Введение холестерин-содержащих катионных липосом модулирует гиперметаболический ответ за счет нарушения экспрессии цитокинов, хотя эффект инъекции скорее всего не продолжается более 5 дней. Отсутствуют различия в содержании сывороточных цитокинов и белков через 7 дней после ожога между животными, которым вводили липосомы, и контрольными животными. Более того, сывороточный трансферрин снижался с 5-го по 7-й дни после ожога у крыс, получавших липосомы, в то время как контрольные особи характеризовались повышением сывороточного трансферрина с 5-го по 7-й дни после ожога. Кроме того, содержание гаптоглобина в сыворотке возрастало с 5-го по 7-й дни в группе, получавшей липосомы, в то время как уровень сывороточного гаптоглобина снижался за тот же период времени у контрольных животных. Следовательно, холестерин-содержащие липосомы повышали содержание конститутивных печеночных белков и снижали количество белков острой фазы I типа со взаимосвязанным снижением уровней IL-1β и TNF-α. Таким образом, холестерин-содержащие липосомы, по-видимому, являются подходящей доставочной системой для генотерапии травм, поскольку липосомы благоприятно модулируют индуцированный травмой гиперметаболический ответ и не проявляют цитотоксичности, обычно связанной с применением других катионных липосом in vivo [4, 7].
Пример 11
Клиническое значение множественных инъекций конструкции гена IGF-I после теплового повреждения
Все крысы в каждой группе выживали при ожоге паром 60% ОППТ и инъекциях лекарственного средства при отсутствии каких-либо неблагоприятных побочных действий. Общая масса тела возрастала примерно на 2% в неделю в первые 4 недели после ожога у животных, трансфицированных однократной и многократными инъекциями конструкций кДНК IGF-I в липосомах, при отсутствии различий между двумя группами. Крысы, получавшие множественные инъекции конструкции кДНК IGF-I, характеризовались более высокими уровнями сывороточных белков (5,2±0,05 г/дл при однократной инъекции против 5,5±0,06 г/дл при множественных инъекциях) и общего печеночного белка (0,64±0,002 мг/мл при однократной инъекции против 0,71±0,03 мг/дл при множественных инъекциях) по сравнению с однократной инъекцией (р<0,05). После удаления струпа процент реэпителизуемой области ожоговой раны был существенно выше через 33 дня после ожога у крыс, получавших двойную инъекцию кДНК IGF-I, по сравнению с однократной инъекцией (38±2% против 31±%, соответственно: р<0,05). Указанные результаты были подтверждены гистологическими измерениями размерных параметров реэпителизации. Крысы, получавшие двойные инъекции кДНК IGF-I, показали существенно более интенсивную реэпителизацию по сравнению с однократными инъекциями (р<0,05; фиг.11).
Уровень трансфекции, определяемой с помощью теста на хемолюминесценцию репортерного гена β-галактозидазы, возрастал по периметру раны у животных, получавших множественные инъекции упакованных в липосомы конструкций кДНК lacZ и кДНК IGF-I, по сравнению с однократными инъекциями (р<0,05; фиг.12А, В). Концентрации белка IGF-I в коже снижались от кожной биопсии в сайте I к кожной биопсии в сайте III у крыс, получавших однократную инъекцию конструкции кДНК IGF-I, вдоль края раны (фиг.13А). Животные, получавшие двойные инъекции конструкции кДНК, характеризовались стабильно повышенными концентрациями белка IGF-I вдоль всего раневого края (фиг.13В). Кроме того, трансфекция выявлялась практически уже через день после подкожной инъекции гена lacZ. Уровень трансфекции повышался и достигал пика на 5-й день после инъекции. В противоположность данным экспериментов in vitro, в которых трансфекция не выявлялась в течение 7 дней после введения, трансфекция кожных клеток сохранялась выявляемой через 7 и 14 дней после инъекции in vivo.
После подкожной инъекции кДНК IGF-I и репортерной конструкции lacZ идентифицировали трансфицированные кожные клетки, миофибробласты, эндотелиальные клетки и макрофаги, включая многоядерные гигантские клетки, для которых известна способность к пролиферации. Уровни мРНК IGF-I возрастали в коже у крыс, трансфицированных конструкцией IGF-I. То, что мРНК транслировалась в белок, согласуется с выявленным увеличением концентраций белка IGF-I в коже. Такое перемежающееся повышение местной экспрессии белка IGF-I способно обусловливать соответствующую стимуляцию синтеза белка IGFBP-3 и повышенные уровни биологически активного комплекса IGF-I/IGFBP-3 локально без каких-либо сопутствующих нефизиологических повышений уровней свободного белка IGF-I в циркуляции и, следовательно, без нежелательных побочных эффектов [49, 50]. Небольшие количества белка IGF-I, экспрессированного после липосомной трансфекции, эффективны в паракринном механизме без неблагоприятных действий более крупных количеств, необходимых при проведении системного введения.
Повышение концентрации белка IGF-I в коже улучшало заживление раны с точки зрения реэпителизации и восстановления кожных клеток в результате митозов таких клеток. Крысы, получавшие кДНК IGF-I, также характеризовались увеличением массы тела и концентраций общего белка в сыворотке и печени. Поскольку ни трансфекция, ни усиление экспрессии IGF-I не были обнаружены в крови, печени, селезенке или почках, благоприятные проявления (например, сохранение массы тела, повышение концентрации сывороточных и печеночных белков) обусловливаются улучшенным заживлением раны и улучшенным восстановлением кожных клеток после повреждения, причиной чего являются паракринные механизмы, связанные с более высокими локальными уровнями белка IGF-I, что противоположно изменениям в уровне белка IGF-I в циркуляции, вызываемым системной трансфекцией. Обусловленное IGF-I митогенное влияние на кератиноциты и фибробласты со стимуляцией синтеза коллагена, а также улучшенная восстанавливаемость клеток после повреждения являются причинами улучшенного заживления раны [40, 41, 48].
Преимущества раннего закрытия раны, продемонстрированные в некоторых клинических исследованиях [36, 37], включают сведенный к минимуму гиперметаболический ответ на ожог и снижение уровней воспалительных медиаторов, таких как IL-1, IL-6, IL-8 и TNF-α [36, 37]. Более того, белок IGF-I снижает уровни экспрессии провоспалительных цитокинов IL-1β и TNF-α после теплового повреждения [57]. Следовательно, IGF-I может проявлять свое благоприятное действие за счет усиления реэпителизации и/или снижения провоспалительного ответа в коже, что является одним из основных источников синтеза и секреции цитокинов после ожога [58, 59].
В данном исследовании было показано, что трансфекция ограничена кожей в пределах небольшого периметра сайтов инъекций. Природа экспрессии IGF-I наиболее вероятно обусловливалась взаимодействиями между положительными поверхностными зарядами на катионных липосомах и отрицательно заряженными внешними клеточными мембранами, что ограничивало перемещение липосом [54]. Однако трансфекция и экспрессия IGF-I в существенной степени возрастали тогда, когда использовались множественные инъекционные сайты по сравнению с единственным сайтом инъекции. Множественные инъекции кДНК IGF-I показали согласованное усиление экспрессии белка IGF-I наряду с улучшенным заживлением раны, в то время как при однократной инъекции выявлялось меньше белка IGF-I вокруг раны. Такой результат имеет клиническое значение, поскольку липосомы, в которые упакован ген, должны быть применены на точно определенном расстоянии от травмированной области с целью обеспечения оптимальных трансфекции и экспрессии белка.
Проведенные здесь эксперименты показывают, что подкожное введение упакованных в липосомы конструкций кДНК IGF-I приводит к успешной трансфекции кожных клеток. Также было продемонстрировано, что кДНК способна транскрибироваться в мРНК и транслироваться в белок IGF-I. Множественные инъекции кДНК IGF-I повышали количество трансфицированных клеток и усиливали экспрессию белка, что улучшало заживление раны по сравнению с однократной инъекцией. Процессы трансфекции, транскрипции и трансляции были ограничены кожей, в то время как системная трансфекция или усиление экспрессии белка IGF-I в организме в целом выявлены не были. Биологическими ответами на повышение IGF-I в коже явились улучшение заживления раны с последующим системным улучшением гиперметаболического ответа.
Пример 12
Повышение уровня белка IGF-I в клетках кожи в результате переноса упакованного в липосомы гена кДНК IGF-I для стимуляции заживления раны
Применение генотерапии перемежающегося типа после травмы является новым подходом в улучшении результатов лечения и снижении смертности. Одним из наиболее важных факторов является выбор подходящего вектора для доставки гена [1, 2]. Отсутствие вируса в составе, отсутствие цитотоксичности, повышенная инфекционность и противовоспалительная активность делают холестерин-содержащие катионные липосомы перспективным средством для улучшения индуцированного ожогом гиперметаболического ответа [4, 51-53]. В настоящем исследовании было показано, что холестерин-содержащие катионные липосомы представляют собой эффективную доставочную систему in vivo после теплового повреждения. Кроме того, в данном тексте описаны механизмы, в соответствии с которыми трансфекция ДНК и последующая индуцированная генная экспрессия изменяют реакции на тепловое повреждение, равно как и влияние переноса гена IGF-I в модели теплового повреждения. В данном тексте было показано, что подкожное введение упакованных в липосомы конструкций кДНК IGF-I приводит к успешной трансфекции кожных клеток и что кДНК транскрибируется в мРНК и транслируется в белок IGF-I. Процессы трансфекции, транскрипции и трансляции были ограничены кожей. Биологическими ответами на повышение IGF-I в коже явились улучшение заживления раны с последующим системным улучшением гиперметаболического ответа. С учетом указанных результатов делается заключение, что холестерин-содержащие катионные липосомы, в которые упакован экспрессирующий плазмидный вектор для кДНК IGF-I, в случае введения их крысам с тепловым повреждением 60% ОППТ, эффективны в повышении концентраций белка IGF-I в коже и тем самым в улучшении заживления ран.
Пример 13
Сравнение оболочек в качестве материала для покрытия
Для определения биологической эффективности различных материалов для раневого покрытия еженедельно в течение 4-месячного периода проводят результирующие измерения, которые включают: (а) время заживления раны; (b) реэпителизацию по параметрам компьютерной планиметрии и гистологических электронномикроскопических измерений; (с) целостность по времени восприятия и гистологию и иммунологические маркеры отторжения; и (d) параметры эластичности и сократимости. Основываясь на полученных данных, функциональность и эффективность различных материалов для покрытий оценивают с определением наиболее подходящих из них.
Кроме того, для оценки переноса гена используют: (а) гистохимические и хемолюминесцентные тесты с экспрессией репортерного гена (β-галактозидаза) в качестве отражения трансфекции; (b) анализ методом Нозерн-блоттинга мРНК IGF-I в коже в качестве маркера транскрипции; и (с) радиоиммунологические тесты с целью выявления белка IGF-I в коже.
Помимо того, ответ острой фазы определяют по следующим показателям: (а) масса тела, усвоение пищи, азотный баланс и концентрация белка в мышцах и печени; (b) выработка конститутивных печеночных белков (альбумин, трансферрин, преальбумин и ретинолсвязывающий белок), определяемые с помощью нефелометра; (с) белки острой фазы (гаптоглобин, кислый α1-гликопротеин, α1-макроглобулин и фибриноген), определяемые с помощью твердофазного иммуноферментного анализа (ТИФА) и нефелометра; и (е) цитокины (интерлейкин-1β, интерлейкин-4, интерлейкин-6, интерлейкин-8, интерлейкин-10, фактор-α некроза опухолей и γ-интерферон), определяемые методом ТИФА.
Пример 14
Раневое покрытие при ожоговом повреждении
Для более точной симуляции клинической обработки каждая из 15 йоркширских миниатюрных свиней под наркозом и с обезболиванием получает полнослойный огневой ожог 35% от общей площади поверхности тела (ОППТ) (стандартная модель). Животных оставляют зафиксированными в подвесной повязке в течение 24 часов, и они получают тщательный уход, включая соответствующие реанимационные мероприятия и обезболивание. Для имитации клинических условий через двадцать четыре часа после ожога животным проводят иссечение раны и трансплантацию. Указанное приблизительно соответствует временным рамкам, принятым для проведения иссечения и трансплантации в отношении тепловых повреждений у людей. Под общим наркозом и с обезболиванием полную ожоговую рану иссекают и немедленно покрывают либо человеческой амниотической оболочкой, либо человеческим амнионом, пропитанным липосомно-генными комплексами. Для одного животного использовали только один тип покрывающего материала в противопоставление различным типам материала для раневого покрытия на спине одного и того же животного. Покрытие прикрепляли к необожженному участку раны и покрывали марлей, содержащей мазь с тремя антибиотиками, и плотной повязкой. Скрепки и/или швы использовали по необходимости. После хирургической обработки анализируемых животных оставляли в удерживающей подвесной повязке до момента выхода из-под наркоза в соответствии с описанным выше.
Описанные эксперименты показывают влияние различных покрывающих материалов в присутствии липосомных экспрессирующих векторов и без них на печеночный ответ острой фазы и корреляцию такого влияния с заживлением раны. Так, непосредственно после ожога, хирургической обработки и через 1, 4, 6, 8, 10, 12, 14 и 16 дней после операции у животных брали венозную кровь и анализировали ее на конститутивные белки печени, белки острой фазы и цитокины. Резервуаром для липосомно-генных конструкций являются человеческая амниотическая оболочка или INTEGRA™. Благодаря электростатическим взаимодействиям между клетками раневой поверхности липосомы должны перемещаться из пропитанного покрывающего материала и трансфицировать кожу и поврежденные клетки, что тем самым приведет к повышению локальной концентрации IGF-I. Повышение концентрации IGF-I будет иметь своим следствием улучшение заживления раны.
Пример 15
Дополнительные факторы роста во взаимодействии с IGF-I
Сочетание плодной оболочки или человеческой трупной кожи, пропитанной липосомными генными конструкциями, ускоряет и улучшает восприятие трансплантата, функциональность, заживление раны и гиперметаболический ответ после повреждения. Липосомная генная конструкция(и) может кодировать инсулино-подобный фактор-1 роста (IGF-I), фактор роста кератиноцитов (KGF), гормон роста (GH), фактор роста фибробластов (FGF), эпидермальный фактор роста (EGF), тромбоцитарный фактор роста (PDGF), трансформирующий β-фактор роста (TGF-β) или любое сочетание упомянутых выше факторов.
Заключение
Как было показано, липосомы, содержащие кДНК, которая кодирует IGF-I, сохраняют массу тела после повреждения 60% ОППТ, предотвращают потерю мышечного белка, улучшают заживление раны и повышают концентрацию белка IGF-I в коже. Более того, трансфекция клеток и последующая экспрессия гена IGF-I являются локальным событием, ограниченным местом инъекции. По причине отсутствия существенных различий в концентрациях сывороточных IGF-I и IGFBP-3 благоприятные эффекты липосомной доставки гена обусловливаются местным эффектом улучшения заживления раны, а не изменениями циркулирующего IGF-I во всем организме.
Настоящее изобретение демонстрирует, что кДНК, которые кодируют интенсифицирующие рост факторы, в предпочтительном плане модулируют гиперметаболический ответ после теплового повреждения, и что липосомные конструкции могут быть эффективно использованы в качестве доставочной системы для генной трансфекции. Положительный терапевтический эффект липосомной трансфекции генов был дополнительно усилен путем увеличения числа сайтов инъекции вокруг краев раны или за счет “накладывания повязки на рану”, что повышает число трансфицированных клеток и соответствующие уровни генной экспрессии. Следовательно, плодная оболочка или иные материалы для раневого покрытия, пропитанные липосомами, которые содержат генную конструкцию с IGF-I, улучшают заживление раны и представляют собой оптимальное лечение ожоговой раны. Такое применение липосомной генотерапии используется для пропитывания человеческой трупной кожи, амниотической оболочки или других типов материалов для покрытия и/или закрытия ран и может обусловить революционное преобразование пластической, восстановительной, травматологической и ожоговой хирургии и улучшить клинические результаты у подобных больных.
Любые патенты или публикации, упомянутые в настоящей заявке, по уровню соответствуют кругозору специалистов в данной области техники, к которым адресовано настоящее изобретение. Кроме того, указанные патенты и публикации включены здесь для сведения в виде библиографических ссылок в той же степени, что и каждая публикация по отдельности определена, как включаемая здесь для сведения в виде библиографической ссылки.
Для специалиста в данной области техники будет легко понятно, что настоящее изобретение хорошо адаптировано для представления его объектов и достижения результатов и преимуществ, указывавшихся выше, а также тех объектов, результатов и преимуществ, которые с ними связаны. Представленные примеры, а также методы, процедуры, способы обработки, молекулы и конкретные соединения, описанные в данном тексте, относятся к предпочтительным вариантам, служат примером и не призваны в чем-либо ограничить объем настоящего изобретения. Изменения в нем и другие пути применения, которые будут ясны специалистам в данной области техники, соответствуют сущности настоящего изобретения в соответствии с тем, что определяется объемом формулы изобретения.
Источники информации
1. Firedmann T. Scientific American. 1997. 6: 96-101.
2. Felgner P.L. Scientific American. 1997. 6: 102-106.
3. Felgner, P.L. et al. Annals of the New York Academy of Sciences. 1995. 772: 126-139.
4. Felgner, P.L. Human Gene Therapy. 1996. 7: 1791-1793.
5. Wheeler, C.J., et al. Proc Nati Acad Sciec 1996. 93 (21): 11454-11459.
6. Sharata, et al. Intern J Dermatalogy. 1996. 35(11):761-769.
7. Fey G. & Gauldie J. The acute phase response of the liver in inflammation. In Popper, H., Schaffner, F. (Eds.). 89-116.
8. Rotheschild M.A., et al. Hepatology. 1988. 8: 385-401.
9. Moshage H.J. Pathol. 1997. 181: 257-266.
10. Seizman C.H., et al. Shock 1998; 10: 309-318.
11. Gilpin D.A., et al. Surgery. 1996. 119 (6): 664-673.
12. Jarrar D., et al. Arch. Surg 1997. 132: 1171-1176.
13. Xia Z.F., et al. Surgery. 1996. 119: 664-673.
15. Herndon D.M., et al. J. Surg. Res. 1978. 25: 394-403.
16. Beutler В. & Cerami A. Adv Immunol. 1988. 42: 213-231.
17. Moldawer L.L., et al. Ann Rev Nutr. 1988. 8: 585-609.
18. Frost R.A., et al. Endo. 1997. 138 (10): 4153-4159.
19. Marano M. et al. Am Burn Assoc Ann Proc. 1988. 20: 18.
20. Yamada Y. et al. Burns. 1996. 22 (8): 587-593,
21. Balteskard L. et al. Scand J Infect Dis. 1997. 29: 393-399.
22. Gore D.C., et al. Arch Surg. 1991. 126: 38-43.
23. Chrysopoulo M.T., et al. Arch Surg. 1998. (in press).
24. Pennanen N. et al. Pharm Res. 1995. 12: 916-922.
25. Aramaki Y. et al. Biochem Biophys Res Corn. 1996. 220: 1-6.
26. Aramaki Y. et al. Biochem Biophys Res Corn. 1997. 231: 827-830.
27. Mulsch A. et al. Biochem Biophys Res Corn. 1993. 191:1301-1308.
28. Sambrand et al. Proc Nati Acad Sci USA. 1995. 92: 1396-1400.
29. Shackelford R.E., et al. J Biol Chem. 1995. 270: 3745-3478.
30. Mulsch A. et al. FEBS Lett. 1993. 321: 215-218.
31. Brisseau G.F., et al. Antimicrobial Agents and Chemotherapy. 1994. 38: 2671-2675.
32. Xiao B.G., et al. Neurosci Lett. 1998. 249: 17-20.
33. Arnush M. et al. J Clin Invest. 1998. 102: 516-526.
34. Pierre, E.; et al. 1996. Growth hormone therapy in the treatment of burns. In: M.H. Torosian, ed. Growth hormone in critical illness: research and clinical studies. R.G. Lan-des Co, Texas. Pp 105-116.
35. Meyer, N.A., et al. 1996. J Trauma 31 (6): 1008-1012.
36. Herndon, D.N., et al. 1989. Ann. Surg. 209 (5): 547-553.
37. Rodriguez, J.L., et al. 1993. J. Trauma 34: 684-694.
38. Zaizen, Y., et al. 1990. J Ped Surg (25): 70.
39. Herndon, D.N., et al. 1990. Ann. Surg. 212: 424.
40. Martin, P. 1997. Science 276: 75-81. 1997.
41. Steenfos, H. 1994. Scand J Plast Reconstr Hand Surg 28: 95-105.
42. Huang, К.F., et al. 1993. Arch Surg 128: 47-54.
43. Strock, L.L., et al. 1990. Surgery 108 (2): 161-164.
44. Clemmons, D.R. 1994. Ann Intern Med 120: 596-597.
45. Lo, H.C., et al. 1995. Am J Physiol 269: E368-E376.
46. Guler, et al. 1988. Proc-Nati Acad Sci USA 85: 4889-4893.
47. Walker, J.L., et al. 1991. N Engi J Med 324: 1483-1488.
48. Pierre, et al. 1997. J Burn Care Rehab 18 (4): 287-291.
49. Jabri, N. et al. 1994. Diabetes 43: 369-374.
50. Bondy, С.A., et al. 1994. Ann Int Med 120: 593-601.
51. Filion, & Philips. Br J Pharmacol 122: 551-557. 1997.
52. Noguchi A. et al. FEBS Lett; 433: 169-173. 1998.
53. Caplen N.J., et al. Nat Med; 1: 39-46. 1995.
54. Alexander, & Akhurst. 1995. Human Molecular Genetics 4 (12): 2279-2285.
55. O’Connor, & Culp. Biotechniques; 17: 502-509. 1994.
56. Lasic D.D. Liposomes in gene delivery. CRC Press, New York.
57. IGF-I APR
58. Kupper T.S., et al. Surgery 100: 409-414. 1986.
59. Garner W.L., et al. Surgery; 116: 42-48. 1994.
60. Herndon, D.N. et al. 1978. J. Surg. Res.; 25:394-403.
61. Hoekstra M.J. et al., 1994. Burns 20:S43-47.
62. van Baare J. et al., 1994. Burns 20:S77-80.
63. Hettich R. et al., 1994. Burns 20:871-76.
64. Richters C.D. et al., 1997. J. Burn Care Rehabil. 18:228-233.
65. Патент США №4361552, выданный Baur Jr.
66. Патент США №5002071, выданный Harrell.
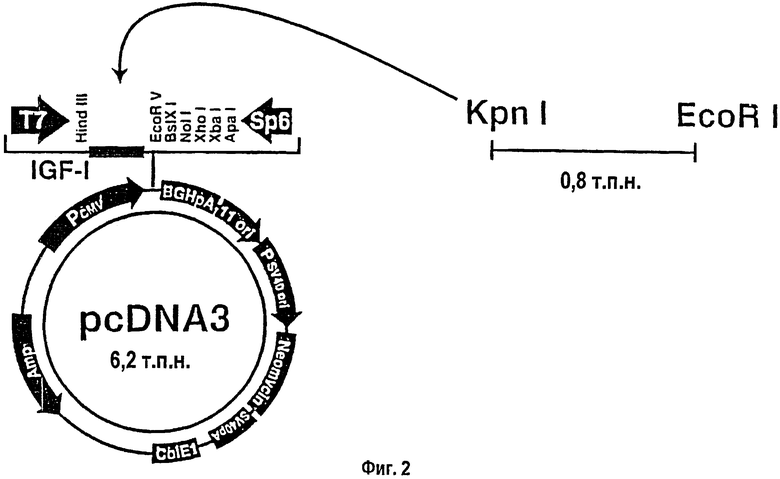
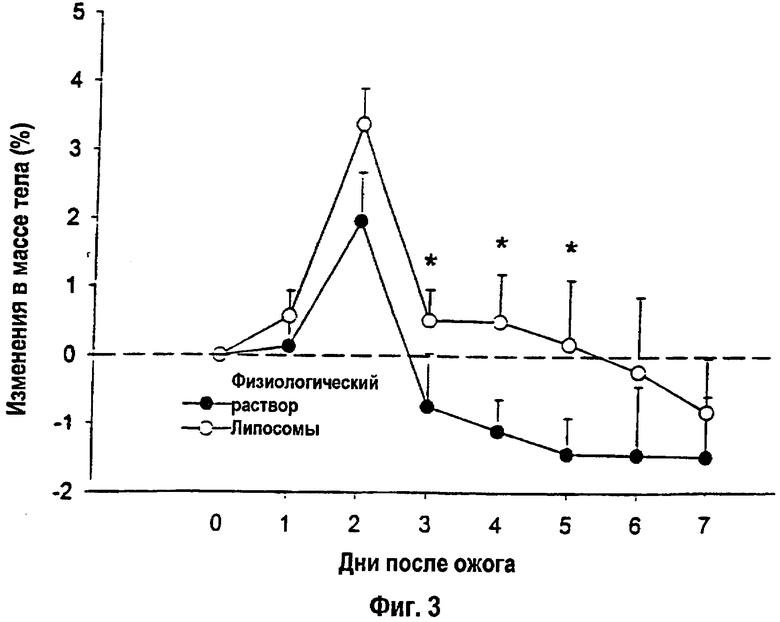


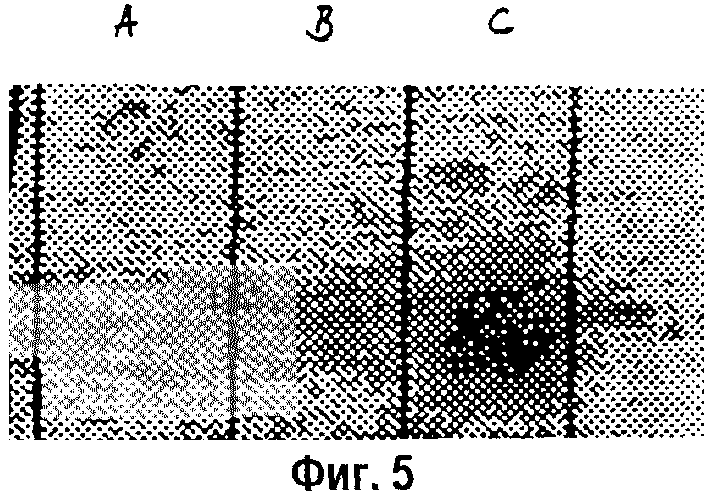
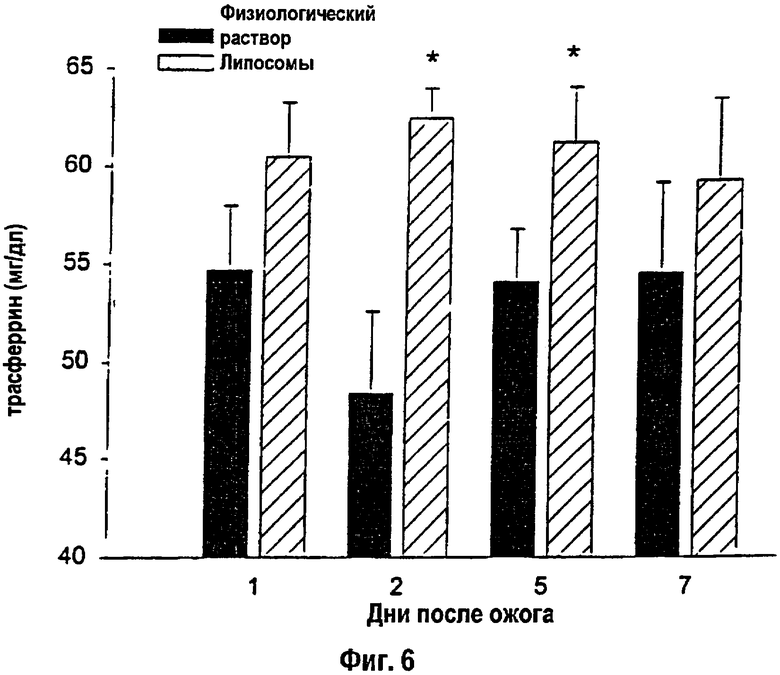
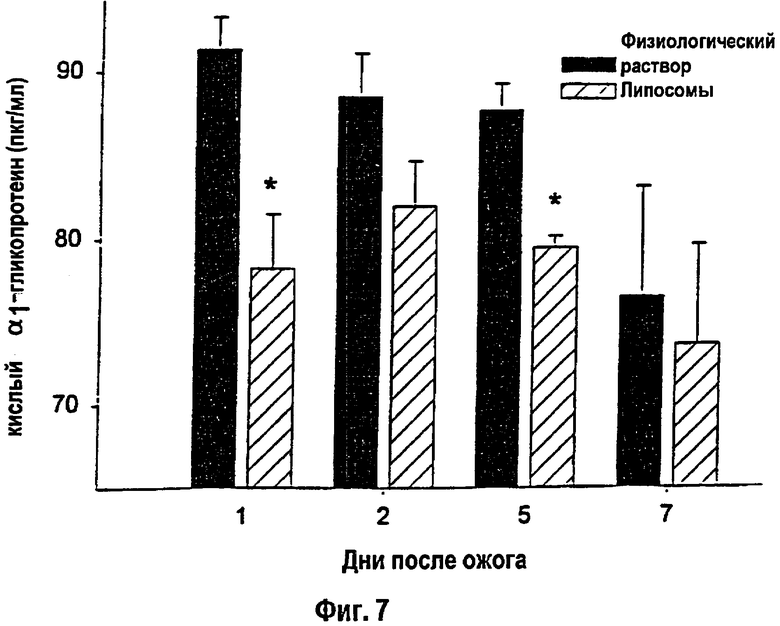
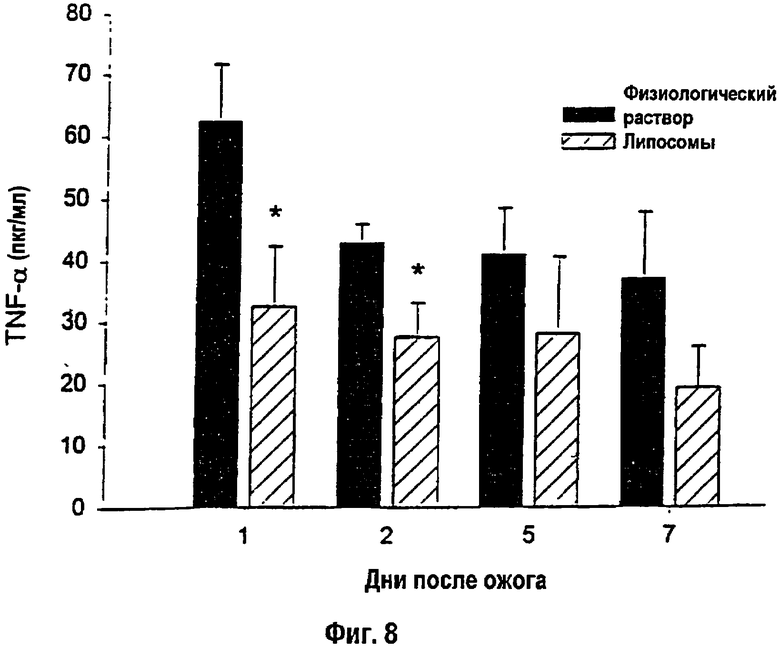
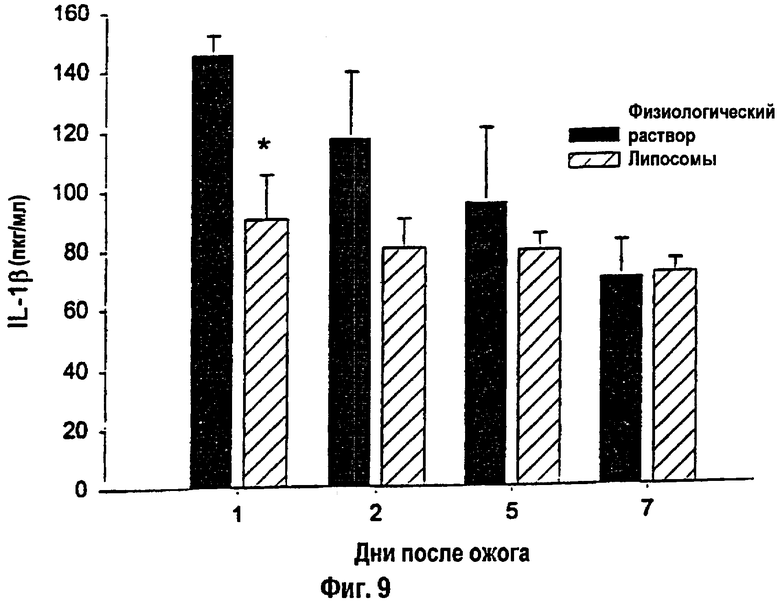
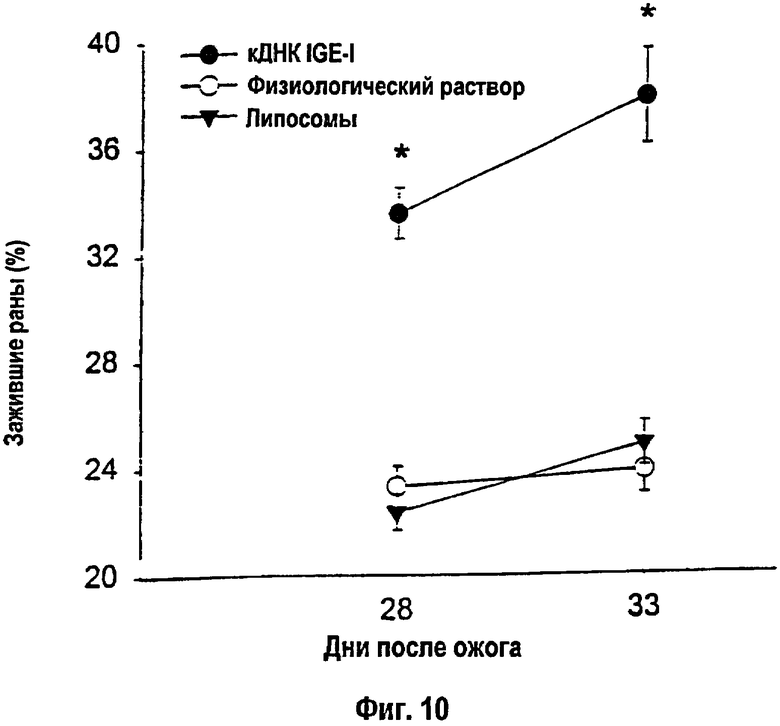
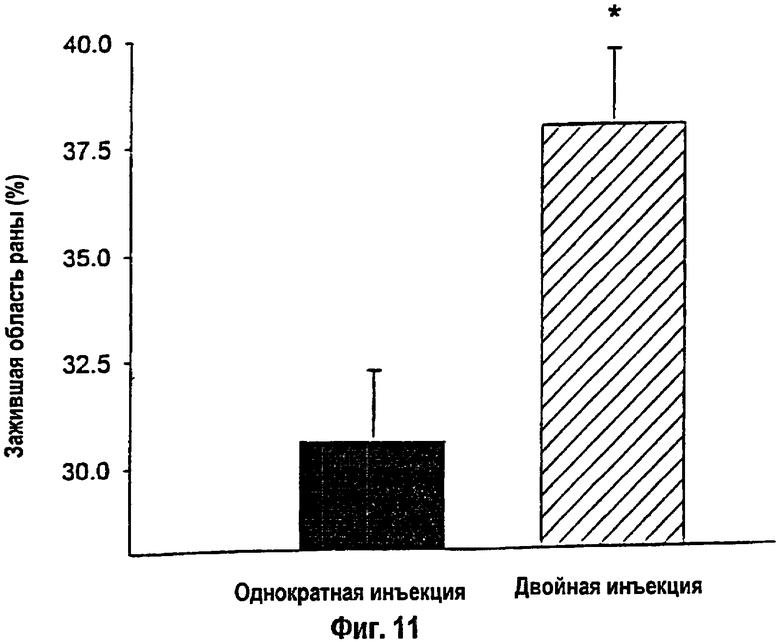
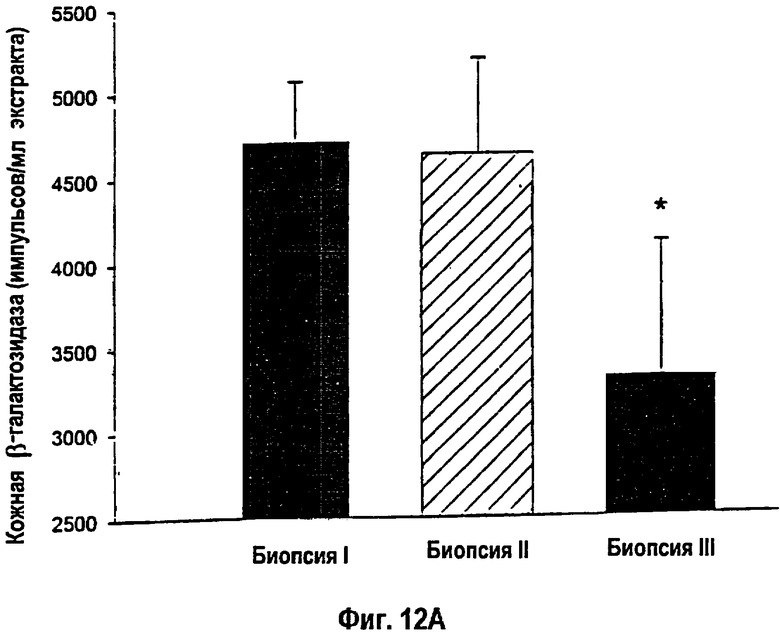
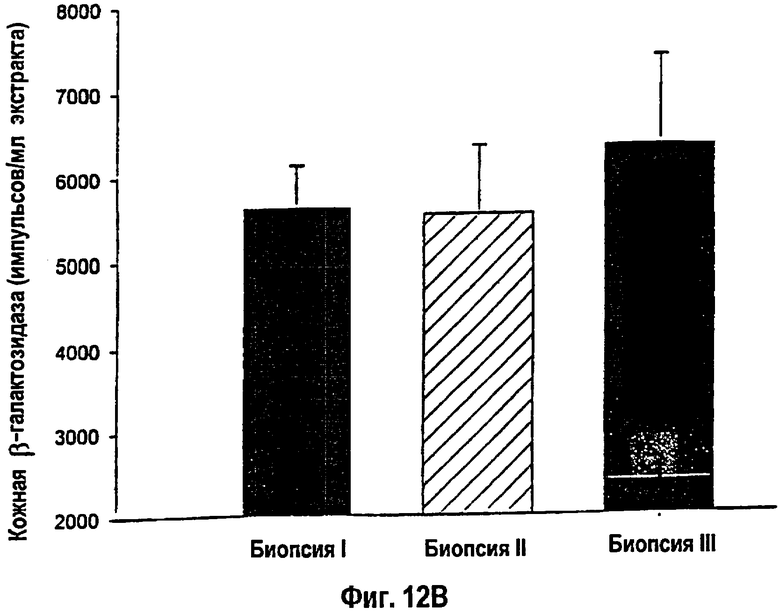
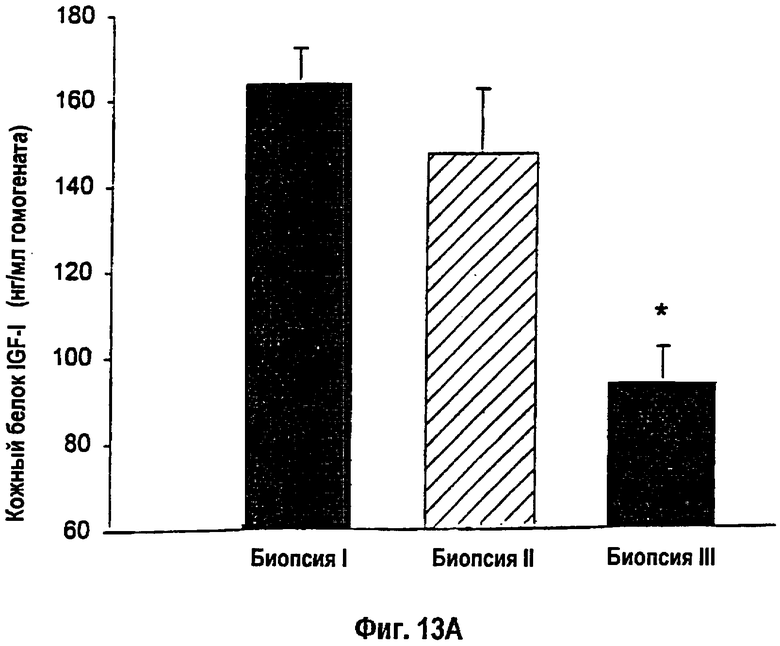
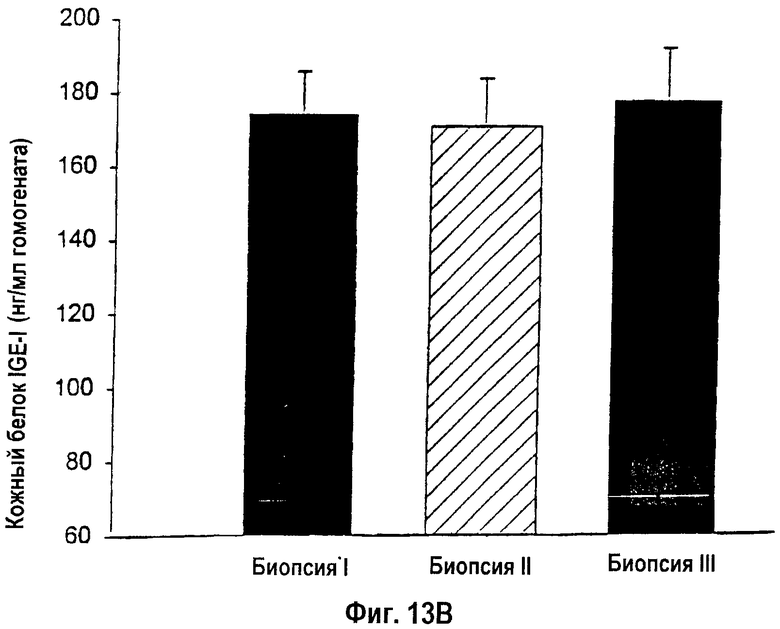
Группа из 5 изобретений относится к медицине, в частности к травматологии, и касается улучшения заживления ран. С этой целью используют композицию (включающую липосомы, содержащие генные конструкции, кодирующие факторы роста), которую используют для инъекций в рану и пропитывания материалов для покрытия или закрытия ран с последующим покрытием или закрытием ран указанными материалами. Предлагают усовершенствованную повязку для ран, включающую материал для покрытия ран и указанные липосомы. Внесение липосомных генных конструкций непосредственно в раны обеспечивает их ускоренное заживление. Усовершенствованные материалы для покрытия и закрытия ран имеют преимущества над материалами, лишенными липосомной генной конструкции, и представляют эффективный и безопасный подход лечению ран. 5 с. и 26 з.п. ф-лы, 4 табл., 13 ил.
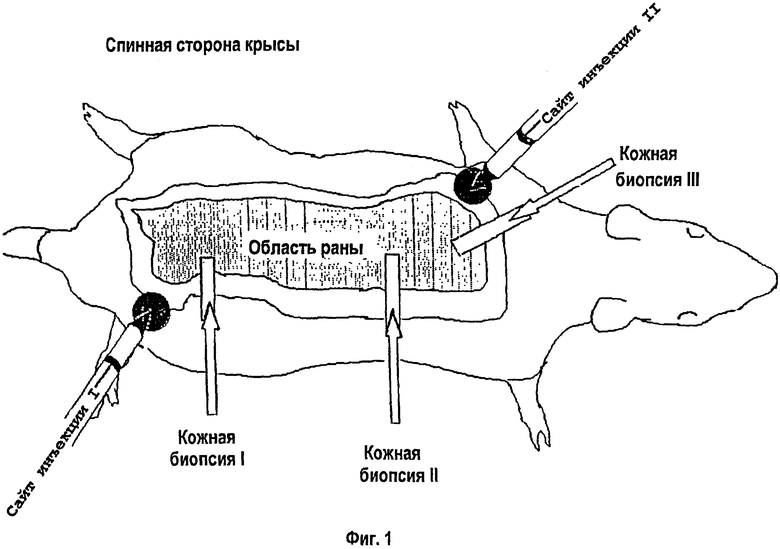
| US 5686116, 11.11.1997 | |||
| РАНОЗАЖИВЛЯЮЩИЙ ПРЕПАРАТ И ХИРУРГИЧЕСКАЯ РАНОЗАЖИВАЮЩАЯ САЛФЕТКА | 1997 |
|
RU2114639C1 |
| ГЛАЗНАЯ МАЗЬ | 1994 |
|
RU2082434C1 |
| RU 95102667 А1, 27.02.1997 | |||
| СПОСОБ ОБРАБОТКИ ВИНОГРАДНЫХ НАТУРАЛЬНЫХ СУХИХ ВИНОМАТЕРИАЛОВ С ПОВЫШЕННОЙ КИСЛОТНОСТЬЮ | 2004 |
|
RU2271388C1 |
| PIERRE E.J | |||
| et al | |||
| Insulin-like growth factor-I liposomal gene transfer and systemic growth hormone stimulate wound healing | |||
| J | |||
| Burn Care Rehabil | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| EPPERLY M.W | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Biol | |||
| Blood Marrow Transplant | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |

