Область техники
Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии. То есть, созданный генотерапевтический ДНК-вектор с целевым геном может быть использован для введения в клетки организма животных и человека, характеризующихся сниженной или недостаточной экспрессией белка, кодируемого данным геном, обеспечивая, таким образом, достижение терапевтического эффекта.
Уровень техники
Генная терапия - это современный медицинский подход, направленный на лечение наследственных и приобретенных заболеваний путем введения нового генетического материала в клетки пациента с целью компенсации или подавления функции мутантного гена и/или исправления генетического дефекта. Конечным продуктом экспрессии гена может являться молекула РНК или белка. Однако осуществление большей части физиологических процессов в организме связано с функциональной активностью белковых молекул, тогда как молекулы РНК являются либо промежуточным продуктом в синтезе белков, либо осуществляют регуляторные функции. Таким образом, целью генной терапии является, в большинстве случаев, введение в организм генов, обеспечивающих транскрипцию и последующую трансляцию белковых молекул, кодируемых этими генами. В рамках описания настоящего изобретения под экспрессией гена подразумевается продукция белковой молекулы, аминокислотная последовательность которой кодируется этим геном.
Гены SKI, TGFB3, FMOD, TIMP2, входящие в группу генов, играют ключевую роль в ряде процессов в организме человека и животных. Эти гены участвуют в сигнальном каскаде TGF-beta (или TGFB), который в той или иной мере задействован в большинстве биологических процессов в организме. Показана связь низких/недостаточных концентраций этих белков с различными неблагоприятными состояниями человека, которая, в ряде случаев, подтверждена нарушениями в нормальной экспрессии генов, кодирующих эти белки. Таким образом, генотерапевтическое повышение экспрессии гена, выбранного из группы генов SKI, TGFB3, FMOD, TIMP2, обладает потенциалом для коррекции различных состояний человека и животных.
Ген SKI кодирует белок-кофактор транскрипции, в частности он выступает в роли корепрессора транскрипции и регулирует TGFB/Smad сигнальный каскад в ходе эмбриогенеза и поддержания гомеостаза тканей. Также показана его коактиваторная роль в случае NF1 семейства транскрипционных факторов, а также транскрипционного фактора MyoD, главной функцией которого является экспрессия генов, участвующих в миогенезе. Помимо миогенеза, SKI участвует в регуляции таких процессов как нейрогенез, гематопоэз, метаболизм жирных кислот, апоптоз и пролиферация клеток, а также в патогенезе различных заболеваний. Ряд неблагоприятных состояний организма сопровождается недостаточной экспрессий SKI. Например, при фиброзе печени, индуцированном метаболитами этилового спирта, наблюдается подавление экспрессии SKI. При системном склерозе экспрессия белка SKI повышена, однако нарушена его функциональная активность. В ходе ремоделлинга тканей при заживлении повреждений печени SKI подавляет ряд антипролиферативных сигналов, тогда как в случае регенерации сосудов, SKI ингибирует пролиферацию клеток. Эти наблюдения свидетельствуют о разносторонних функциях белка SKI в зависимости от условий и контекстного окружения (Tecalco-Cruz et al. 2018).
Негативная регуляция TGFB сигнального каскада за счет кофактора SKI может потенциально предотвращать фиброз тканей. Известно, что недостаточная экспрессия SKI ассоциирована с фиброзом почек, легких и печени (Li X, et al. 2016; Tang H, et al. 2016; Chen J, et al. 2013; Tan R, et al. 2006). На экспериментальной модели фиброза почки было показано, что повышение экспрессии SKI препятствует развитию патологических изменений, связанных с фиброзом тканей (Yang J, Zhang X, Li Y, Liu Y. 2003). Также при раневых повреждениях кожи, у животных с гиперэкспрессией белка SKI, полученной за счет введения ДНК-вектора с трансгеном SKI, наблюдалось ускорение заживления ран и ингибирование формирования рубцов в месте повреждений (Liu X, et al. 2006; Peng Y, et al. 2016). Увеличение локальной экспрессии SKI путем введения экспрессирующего ген SKI плазмидного вектора приводило к ускоренному заживлению ран за счет снятия воспаления, ускорения реэпителизации и увеличения образования грануляционной ткани. Было продемонстрировано уменьшение формирования рубцов за счет уменьшения образования коллагена в области повреждения кожи у крыс (Li, P., et al. // J Pathol, 2011. 223(5): p. 659-71).
Роль SKI в канцерогенезе недостаточно изучена, имеются данные как о протоонкогенной, так и противоонкогенной роли SKI. Так, например, у мышей гомозиготных по одной из мутаций гена SKI наблюдается повышенная частота спонтанных лимфом и восприимчивость к канцерогенам (Shinagawa Т, et al. 2001). У пациентов с некоторыми типами дисплазии наблюдается сниженная экспрессия SKI в зоне поражения по сравнению с участками, не затронутыми дисплазией (Villanacci V, et al. 2008). Некоторые типы метастазирующих раков легких характеризуются сниженной экспрессией SKI (Yang Н, et al. 2015), тогда как повышенная экспрессия SKI при раке груди, напротив, потенциирует метастазирование опухоли (Ritter М, et al. 2006).
Также описан наследственный синдром Шпринтцена-Гольдберга, вызванный делеционной мутацией в гене SKI, который проявляется скелетными, неврологическими, сердечно-сосудистыми и другими аномалиями (Zhu X, et al. 2013).
Ген TGFB3 кодирует белок TGFB3, который является плейотропным цитокином и представляет собой одну из трех изоформ TGFB белков, участвующих в одноименном сигнальном каскаде. Наряду с другими изоформами TGFB1 и TGFB2, TGFB3 играет ключевую роль в иммунной системе и обладает как про-, так и противовоспалительными функциями, участвует в регуляции продукции антител и иммунитете слизистых оболочек. Показана связь мутаций TGFB3 с рядом аутоиммунных заболеваний, например, с HLA-B7+ спондилоартропатиями. Также было показано, что мыши с недостаточной экспрессией TGFB3 развивали аутоиммунные нарушения с волчаночно-подобными проявлениями (Komai Т et al. 2018).
В экспериментальной работе было показано, что мезенхимальные стволовые клетки, гиперэкспрессирующие TGFB3, значительно улучшали заживление ран у лабораторных кроликов. Этот подход также позволил изменить баланс различных типов коллагенов, что способствовало уменьшению формирования рубцов (Li М et al. 2018).
В другой работе было показано, что мезенхимальные стволовые клетки, трансдуцированные рекомбинантным аденовирусным вектором, экспрессирующим TGFB3 и рекомбинантным аденовирусным вектором, экспрессирующим ВМР-2, способствовали регенерации поврежденного хряща в экспериментальной модели на поросятах (Wang X et al. 2015).
С использованием модели дегенеративных изменений позвоночных дисков у кроликов, показали, что введение рекомбинантного лентивирусного вектора, экспрессирующего белки сурвивин, TGFB3 и TIMP2, способствовало замедлению дегенеративных изменений позвоночных дисков (Yue В et al. 2016).
В отличии от других изоформ TGFB, TGFB3 обладает противофиброзным действием и ингибирует образование рубцовой ткани. Авотермин (препарат Juvista®, Renovo; выпускается в 2-х формах - для внутрикожного введения и для топических апликаций), который представляет собой рекомбинантный белок TGFB3, ускорял заживление ран и способствовал профилактике образования рубцов в ходе I/II фазы клинических испытаний (McCollum РТ et al. 2011).
Принимая во внимание короткое время полужизни активной формы белка TGFB3 (2-3 минуты), генотерапевтический подход является наиболее перспективным направлением для повышения концентрации белка TGFB3 в тканях человека и животных.
Ген TIMP2 кодирует белок TIMP2, который является ингибитором матриксных металлопротеиназ (ММР) и участвует в поддержании гомеостаза тканей. ММР играют важную роль в физиологических и патологических процессах, включая эмбриогенез, тканевое ремоделирование, заживление ран, воспаление, артрит и рак. Клетки некоторых опухолей сами образуют ММР. Способствуя инвазии опухоли и метастазированию, ММР в то же время являются мощными стимуляторами неоангиогенеза. Таким образом, TIMP опосредованное ингибирование ММР может являться одной из стратегий терапии опухолей. Концентрация TIMP-2 в сыворотке крови коррелирует как с длительностью ремиссии, так и выживаемостью у пациенток с раком молочной железы. При раке яичников было показано, что рекомбинантный аденовирусный вектор, экспрессирующий ген TIMP2, оказывал противоопухолевое действие как за счет ингибирования матриксных металлопротеиназ, так и за счет ММР-независимого механизма действия (Yang SW et al. 2011).
В отличии от других ингибиторов ММР, TIMP2 обладает прямым ингибирующим действием на пролиферацию клеток эндотелия, что потенциально свидетельствует о возможности его применения для коррекции состояний, связанных с патологическим ангиогенезом (Kim HJ et al. // Cancer Lett. 2014 Feb 28; 343(2):210-6).
Некоторые аллели гена TIMP2 ассоциированы с повышенным риском развития остеоартроза коленного сустава (Xu Р et al. // Oncotarget. 2017 Jan 3; 8(1):1166-1176), количеством активных сперматозоидов (Kurzawski М et al. // Andrologia. 2017 Jun; 49(5)) и депрессивными расстройствами ( K et al. // J Affect Disord. 2016 Nov 15; 205:119-129).
K et al. // J Affect Disord. 2016 Nov 15; 205:119-129).
Недостаточная экспрессия TIMP2 ассоциирована с развитием миопии, при этом субконьюктивальное введение белка TIMP2 предотвращало прогрессию этого заболевания (Liu НН et al. 2017).
Было продемонстрировано усиление миграции эпидермальных кератиноцитов и ускоренное заживление раны у мыши при введении рекомбинантного белка TIMP2. Также показано, что TIMP2 подавляет синтез коллагена в культуре фибробластов (Dohi, Т., et al. // Plast Reconstr Surg Glob Open, 2015. 3(9): p. e520.).
Как уже отмечалось выше, с использованием модели дегенеративных изменений позвоночных дисков у кроликов, показали, что введение рекомбинантного лентивирусного вектора, экспрессирующего белки сурвивин, TGFB3 и TIMP2, способствовало замедлению дегенеративных изменений позвоночных дисков (Yue В et al. 2016).
Ген FMOD кодирует белок FMOD (фибромодулин), играющий важную роль в ряде нормальных и патологических процессов. Белок фибромодулин относится к семейству протеогликанов богатых лейцином, участвующих в безрубцовых заживлениях ран у плода (Soo, С., et al. // Am J Pathol, 2000. 157(2): p. 423-33). FMOD взаимодействует с ферментами, осуществляющими формирование структуры коллагеновых волокон, тем самым участвуя в процессе формирования внеклеточного матрикса. Также показано, что FMOD задействован в формировании мышечных клеток, репрограммировании клеток и ангиогенезе (Jan AT, Lee EJ, Choi I. // Int J Biochem Cell Biol. 2016 Nov; 80:66-70).
Было показано, что уровень экспрессии фибромодуллина выше у плода, чем у взрослой крысы. Использование рекомбинантного аденовирусного вектора, экспрессирующего FMOD, приводит к понижению экспрессии TGFB1 и TGFB2 и увеличению экспрессии TGFB3 в фибробластах. Также, гиперэкспрессия фибромодуллина приводит к увеличению экспрессии TIMP2, что приводит к уменьшению формирования рубцов на модели раны у кроликов (Stoff, A., et al. // J Mol Med (Berl), 2007. 85(5): p. 481-96).
Таким образом предшествующий уровень техники свидетельствует о том, что мутации в генах SKI, TGFB3, TIMP2, FMOD или недостаточная экспрессия белков, кодируемых этими генами, связаны с развитием спектра заболеваний, включающих в себя, но не ограничивающихся, аутоиммунными заболеваниями, раком, наследственными и приобретенными патологическими состояниями, такими как повреждения соединительной ткани, и другими процессами. Этим обусловлено объединение генов SKI, TGFB3, TIMP2, FMOD в рамках данного патента в группу генов. Генетические конструкции, обеспечивающие экспрессию белков, кодируемых генами из группы SKI, TGFB3, TIMP2, FMOD, могут быть использованы для разработки лекарственных препаратов для предотвращения и терапии различных заболеваний и патологических состояний.
Более того, приведенные данные свидетельствуют о том, что недостаточная экспрессия белков, кодируемых генами SKI, TGFB3, TIMP2, FMOD, входящими в группу генов, связана не только с патологическими состояниями, но и с предрасположенность к их развитию. Также приведенные данные свидетельствуют о том, что недостаточная экспрессия данных белков может не проявляться в явном виде в форме патологии, которая может быть однозначно описана в рамках существующих стандартов клинической практики (например, с применением кода МКБ), однако при этом вызывать состояния, которые неблагоприятны для человека и животных и связанны с ухудшением качества жизни.
Анализ подходов для повышения экспрессии целевых генов подразумевает возможность использования различных генотерапевтических векторов.
Генотерапевтические векторы разделяют на вирусные, клеточные и ДНК-векторы (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal Products
EMA/CAT/80183/2014). В последнее время в генной терапии все большее внимание уделяется разработке невирусных систем доставки генетического материала, среди которых лидируют плазмидные векторы. Плазмидные векторы лишены недостатков, присущих клеточным и вирусным векторам. В клетке-мишени они существуют в эписомальной форме, не интегрируют в геном, производство их достаточно дешево, отсутствие иммунного ответа и побочных реакций на введение плазмидного вектора делают их удобным инструментом генной терапии и генетической профилактики (ДНК-вакцины) (Li L, Petrovsky N. // Expert Rev Vaccines. 2016; 15(3):313-29).
Тем не менее, ограничениями для использования плазмидных векторов для генной терапии являются: 1) наличие генов устойчивости к антибиотикам для наработки в бактериальных штаммах, 2) наличие различных регуляторных элементов, представленных последовательностями вирусных геномов 3) размер терапевтического плазмидного вектора, определяющий эффективность проникновения вектора в клетку-мишень.
Известно, что Европейское агентство по лекарственным средствам считает необходимым избегать введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies). Данная рекомендация связана, в первую очередь, с потенциальной опасностью проникновения ДНК-вектора или горизонтального переноса генов антибиотикорезистентности в клетки бактерий, представленных в организме в составе нормальной или оппортунистической микрофлоры. Помимо этого, наличие генов антибиотикорезистентности значительно увеличивает размер ДНК-вектора, что приводит к снижению эффективности его проникновения в эукариотические клетки.
Необходимо отметить, что гены антибиотикорезистентности также вносят принципиальный вклад в способ получения ДНК-векторов. В случае наличия генов антибиотикорезистентности штаммы для наработки ДНК-векторов обычно культивируются в среде, содержащей селективный антибиотик, что создает риск наличия следовых количеств антибиотика в недостаточно очищенных препаратах ДНК-векторов. Таким образом, получение ДНК-векторов для генной терапии, в которых отсутствуют гены антибиотикорезистентности, связано с получением штаммов, обладающих такой отличительной особенностью как способность к стабильной амплификации целевых ДНК-векторов в среде без содержания антибиотиков.
Кроме того, Европейское Медицинское Агентство рекомендует избегать наличия в составе терапевтических плазмидных векторов регуляторных элементов для повышения экспрессии целевых генов (промоторов, энхансеров, посттрансляционных регуляторных элементов), являющихся нуклеотидными последовательностями геномов различных вирусов (Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products, http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf). Данные последовательности, хотя и могут увеличивать уровень экспрессии целевого трансгена, однако создают риск рекомбинации с генетическим материалом вирусов дикого типа и интеграции в геном эукариотической клетки. Более того, целесообразность гиперэкспрессии того или иного гена в целях терапии остается нерешенным вопросом.
Также, существенным моментом является размер терапевтического вектора. Известно, что современные плазмидные векторы зачастую перегружены нефункциональными участками, серьезно увеличивающими размер вектора (Mairhofer J, Grabherr R. // Mol Biotechnol. 2008. 39(2):97-104). Например, ген устойчивости к ампициллину в векторах серии pBR322, как правило, состоит из не менее чем 1000 п.н., что составляет более 20% от размера самого вектора. При этом наблюдается обратная зависимость между размером вектора и его способностью проникать в эукариотические клетки - ДНК-векторы с небольшим размером эффективней проникаю в клетки человека и животных. Так, например, в серии экспериментов по трансфекции клеток HELA ДНК-векторами с размером от 383 до 4548 п.н. было показано, что разница в эффективности проникновения может достигать двух порядков (отличаться в 100 раз) (Hornstein BD et al. // PLoS ONE. 2016; 11(12): e0167537.).
Таким образом, при выборе ДНК-вектора в целях безопасности и наибольшей эффективности следует отдавать предпочтение тем конструкциям, в которых не содержатся гены устойчивости к антибиотикам, последовательности вирусного происхождения и размер которых позволяет эффективно проникать в эукариотические клетки. Штамм для получения такого ДНК-вектора в количествах, достаточных для целей генной терапии, должен обеспечивать возможность стабильной амплификации ДНК-вектора с использованием питательных сред, не содержащих антибиотики.
Примером использования рекомбинантных ДНК-векторов для генной терапии является способ получения рекомбинантного вектора для генетической иммунизации по патенту US 9550998 В2. Плазмидный вектор представляет собой суперскученный плазмидный ДНК-вектор и предназначен для экспрессии клонированных генов в клетках животных и человека. Вектор состоит из ориджина репликации, регуляторных элементов, включающих промотор и энхансер цитомегаловируса человека, регуляторные элементы из Т-лимфотропного вируса человека.
Накопление вектора проводят в специальном штамме Е. coli без использования антибиотиков за счет антисенс-комплементации гена sacB, введенного в штамм посредством бактериофага. Недостатком данного изобретения является наличие в составе ДНК-вектора регуляторных элементов, представляющих собой последовательности вирусных геномов.
Известна заявка CN 101818171 A, в которой описан способ повышения экспрессии гена SKI с использованием генотерапевтического ДНК-вектора pUC118-Ski для заживления ран человека. Недостатком данного изобретения является использование ДНК-вектора, содержащего последовательности вирусного происхождения (CMV промотор) и ген устойчивости к ампициллину, а также ограниченность области применения.
Известна заявка WO 1996039196 А1, в которой описан способ модулирования соединительной ткани путем введения ДНК-вектора или аутологичных хондроцитов, трансформированных ДНК-вектором, экспрессирующим ген, выбранный из группы генов, содержащей, в том числе, гены TGFB3 и TIMP2. Недостатком данного изобретения является ограниченность использования, неопределенные требования к безопасности используемых ДНК-векторов.
Известна заявка CN 102448983 A, в которой предлагается использовать рекомбинантный белок FMOD для коррекции состояний, связанных с недостаточной функцией FMOD. Недостатком данного изобретения является использование рекомбинантного белка, а не генотерапевтического подхода для повышения экспрессии гена FMOD.
Прототипом настоящего изобретения в части использования генотерапевтических подходов для повышения уровня экспрессии генов из группы SKI, TGFB3, TIMP2, FMOD является заявка JP 2018512876 A, в которой описано использование олигонуклеотидов, в частности некодирующих РНК, для повышения экспрессии генов, в число которых входят гены из группы SKI, TGFB3, TIMP2 и FMOD. Одним из предлагаемых в данной заявке способов доставки некодирующих РНК является использование векторов, в том числе ДНК-векторов в виде плазмид. Недостатком данного прототипа является механизм повышения экспрессии генов, заключающийся в опосредованном действии некодирующих РНК.
Раскрытие изобретения
Задачей изобретения является конструирование генотерапевтических ДНК-векторов для повышения уровня экспрессии группы генов SKI, TGFB3, TIMP2, FMOD в организме человека и животных, сочетающих в себе следующие свойства:
I) Эффективность генотерапевтического ДНК-вектора для повышения уровня экспрессии целевых генов в эукариотических клетках.
II) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов.
III) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора генов антибиотикорезистентности.
IV) Технологичность получения и возможность наработки генотерапевтического ДНК-вектора в промышленных масштабах.
Пункты II и III предусмотрены в данном техническом решении в соответствии с рекомендациями государственных регуляторов к лекарственным средствам для генной терапии, в частности, Европейского Агентства по лекарственным средствам касательно отказа от введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011, EMA/CAT/GTWP/44236/2009 Committee for advanced therapies) и касательно отказа от введения в разрабатываемые плазмидные векторы для генной терапии элементов вирусных геномов (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products / 23 March 2015, EMA/CAT/80183/2014, Committee for Advanced Therapies).
Задачей изобретения также является конструирование штаммов, несущих эти генотерапевтические ДНК-вектора, для наработки и производства в промышленных масштабах генотерапевтических ДНК-векторов.
Поставленная задача решается за счет того, что создан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген, выбранный из группы генов SKI, TGFB3, TIMP2, FMOD, для повышения уровня экспрессии этого целевого гена в организме человека и животных, при этом генотерапевтический ДНК-вектор VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD имеет нуклеотидную последовательность SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 соответственно. Каждый из созданных генотерапевтических ДНК-векторов: VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD за счет ограниченного размера векторной части VTvaf17, не превышающей 3200 п.н., обладает способностью эффективно проникать в клетки и экспрессировать клонированный в него целевой ген, выбранный из группы генов SKI, TGFB3, TIMP2, FMOD соответственно. В составе генотерапевтического ДНК-вектора отсутствуют нуклеотидные последовательности вирусного происхождения и отсутствуют гены антибиотикорезистентности, обеспечивая возможность его безопасного применения для генетической терапии человека и животных.
Создан также способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: ген SKI, ген TGFB3, ген TIMP2, ген FMOD, который заключается в том, что каждый из генотерапевтических ДНК-векторов: VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD получают следующим образом: кодирующую часть целевого гена из группы SKI, или TGFB3, или TIMP2, или FMOD клонируют в ДНК-вектор VTvaf17 и получают генотерапевтический ДНК-вектор VTvaf17-SKI, SEQ ID №1, или VTvaf17-TGFB3, SEQ ID №2 или VTvaf17-TIMP2, SEQ ID №3, или VTvaf17-FMOD, SEQ ID №4 соответственно.
Способ применения созданного генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: ген SKI, ген TGFB3, ген TIMP2, ген FMOD для повышения уровня экспрессии этих целевых генов, заключается во введении выбранного генотерапевтического ДНК-вектора или нескольких выбранных генотерапевтических ДНК-векторов в клетки, органы и ткани человека или животного, и/или во введении в органы и ткани человека или животного аутологичных клеток человека или животного, трансфицированных выбранным генотерапевтическим ДНК-вектором или несколькими выбранными генотерапевтическими ДНК-векторами, или в сочетании обозначенных способов.
Способ получения штамма Escherichia coli SCS110-AF/VTvaf17-SKI, или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AF/VTvaf17-FMOD заключается в электропорации компетентных клеток штамма Escherichia coli SCS110-AF созданным генотерапевтическим ДНК-вектором и последующей селекцией стабильных клонов штамма с использованием селективной среды.
Заявлен штамм Escherichia coli SCS110-AF/VTvaf17-SKI, или штамм Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамм Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамм Escherichia coli SCS110-AF/VTvaf17-FMOD, несущий генотерапевтический ДНК-вектор для его наработки с возможностью культивирования штамма без использования антибиотиков.
Способ производства в промышленных масштабах генотерапевтического ДНК-вектора заключается в масштабировании бактериальной культуры штамма до количеств, необходимых для наращивания бактериальной биомассы в промышленном ферментере, после чего биомассу используют для выделения фракции, содержащей целевой ДНК-продукт - генотерапевтический ДНК-вектор VTvaf17-SKI или VTvaf17-TGFB3 или VTvaf17-TIMP2 или VTvaf17-FMOD многостадийно фильтруют и очищают хроматографическими методами.
Изобретение поясняется чертежами, где:
На фиг. 1
приведена схема генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SKI, TGFB3, TIMP2, FMOD, который представляет собой кольцевую двуцепочечную молекулу ДНК, способную к автономной репликации в клетках бактерии Escherichia coli.
На фиг. 1 приведены схемы, соответствующие:
А - генотерапевтический ДНК-вектор VTvaf17-SKI,
В - генотерапевтический ДНК-вектор VTvaf17-TGFB3,
С - генотерапевтический ДНК-вектор VTvaf17-TIMP2,
D - генотерапевтический ДНК-вектор VTvaf17-FMOD.
На схемах отмечены следующие структурные элементы вектора:
EF1aprom - промоторная область гена человеческого фактора элонгации EF1A с собственным энхансером, содержащимся в первом интроне гена. Служит для обеспечения высокого уровня транскрипции рекомбинантного гена в большинстве тканей человека;
Рамка считывания целевого гена, соответствующая кодирующей части гена SKI (фиг. 1А), или TGFB3 (фиг. 1В), или TIMP2 (фиг. 1С), или FMOD (фиг. 1D) соответственно;
hGH-TAterm - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
pBR322 ori - ориджин репликации, служащий для автономной репликации с однонуклеотидной заменой для повышения копийности плазмиды в клетках большинства штаммов Escherichia coli;
Отмечены уникальные сайты рестрикции.
На фиг. 2
показаны графики накопления ампликонов кДНК целевого гена, а именно, гена SKI, в первичной культуре клеток фибробластов кожи человека HDFa (АТСС PCS-201-01) до их трансфекции и через 48 часов после трансфекции этих клеток генотерапевтическим ДНК-вектором VTvaf17-SKI с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг. 2 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена SKI в первичной культуре клеток фибробластов кожи человека HDFa до трансфекции ДНК-вектором VTvaf17-SKI
2 - кДНК гена SKI в первичной культуре клеток фибробластов кожи человека HDFa после трансфекции ДНК-вектором VTvaf17-SKI
3 - кДНК гена В2М в первичной культуре клеток фибробластов кожи человека HDFa до трансфекции ДНК-вектором VTvaf17-SKI
4 - кДНК гена В2М в первичной культуре клеток фибробластов кожи человека HDFa после трансфекции ДНК-вектором VTvaf17-SKI
В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг. 3
показаны графики накопления ампликонов кДНК целевого гена, а именно гена TGFB3, в первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa (АТСС PCS-200-01) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-TGFB3 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг. 3 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена TGFB3 в первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa до трансфекции ДНК-вектором VTvaf17-TGFB3
2 - кДНК гена TGFB3 первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa после трансфекции ДНК-вектором VTvaf17-TGFB3
3 - кДНК гена В2М в первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa до трансфекции ДНК-вектором VTvaf17-TGFB3
4 - кДНК гена В2М первичной культуре первичной культуры кератиноцитов эпидермиса человека HEKa после трансфекции ДНК-вектором VTvaf17-TGFB3
В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг. 4
показаны графики накопления ампликонов кДНК целевого гена, а именно гена TIMP2 в клетках фибробластов крайней плоти человека линии Hs27 (АТСС CRL-1634) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-TIMP2 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг. 4 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена TIMP2 в клетках фибробластов крайней плоти человека линии Hs27 до трансфекции ДНК-вектором VTvaf17-TIMP2
2 - кДНК гена TIMP2 клетках фибробластов крайней плоти человека линии Hs27 после трансфекции ДНК-вектором VTvaf17-TIMP2
3 - кДНК гена В2М клетках фибробластов крайней плоти человека линии Hs27 до трансфекции ДНК-вектором VTvaf17-TIMP2
4 - кДНК гена В2М клетках фибробластов крайней плоти человека линии Hs27 после трансфекции ДНК-вектором VTvaf17-TIMP2
В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг. 5
показаны графики накопления ампликонов кДНК целевого гена, а именно гена FMOD, в клетках эндотелия капилляров кожи человека линии НМЕС-1 (АТСС CRL-3243) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-FMOD с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг. 5 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена FMOD в клетках эндотелия капилляров кожи человека линии НМЕС-1 до трансфекции ДНК-вектором VTvaf17-FMOD
2 - кДНК гена FMOD в клетках эндотелия капилляров кожи человека линии НМЕС-1 после трансфекции ДНК-вектором VTvaf17-FMOD
3 - кДНК гена В2М в клетках эндотелия капилляров кожи человека линии НМЕС-1 до трансфекции ДНК-вектором VTvaf17-FMOD
4 - кДНК гена В2М в клетках эндотелия капилляров кожи человека линии НМЕС-1 после трансфекции ДНК-вектором VTvaf17-FMOD
В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг. 6
показана диаграмма концентрации белка SKI в клеточном лизате первичной культуры фибробластов кожи человека HDFa (АТСС PCS-201-01) после трансфекции этих клеток ДНК-вектором VTvaf17- SKI с целью оценки функциональной активности, то есть экспрессии на уровне белка, по изменению количества белка SKI в лизате клеток.
На фиг. 6 отмечены следующие элементы:
культура А - культура клеток первичных фибробластов кожи человека HDFa, трансфицированная водным раствором дендримеров без плазмидной ДНК (контроль)
культура В - культура клеток первичных фибробластов кожи человека HDFa, трансфицированных ДНК-вектором VTvaf17
культура С - культура клеток первичных фибробластов кожи человека HDFa, трансфицированных ДНК-вектором VTvaf17-SKI
На фиг. 7
показана диаграмма концентрации белка TGFB3 в лизате клеток первичной культуры кератиноцитов эпидермиса человека HEKa (АТСС PCS-200-01), после трансфекции этих клеток ДНК-вектором VTvaf17-TGFB3 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген TGFB3.
На фиг. 7 отмечены следующие элементы:
культура А - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированная водным раствором дендримеров без плазмидной ДНК (контроль)
культура В - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированная ДНК-вектором VTvaf17
культура С - первичная культура клеток кератиноцитов эпидермиса человека HEKa, трансфицированных ДНК-вектором VTvaf17-TGFB3
На фиг. 8
показана диаграмма концентрации белка TIMP2 в лизате клеток фибробластов крайней плоти человека линии Hs27 (АТСС CRL-1634) после трансфекции этих клеток ДНК-вектором VTvaf17-TIMP2 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген TIMP2.
На фиг. 8 отмечены следующие элементы:
культура А - культура клеток фибробластов крайней плоти человека линии Hs27, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль)
культура В - культура клеток фибробластов крайней плоти человека линии Hs27, трансфицированных ДНК-вектором VTvaf17
культура С - культура клеток фибробластов крайней плоти человека линии Hs27, трансфицированных ДНК-вектором VTvaf17-TIMP2.
На фиг. 9
показана диаграмма концентрации белка FMOD в лизате клеток эндотелия капилляров кожи человека линии НМЕС-1 (АТСС CRL-3243) после трансфекции этих клеток ДНК-вектором VTvaf17-FMOD с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген FMOD.
На фиг. 9 отмечены следующие элементы:
культура А - культура клеток эндотелия капилляров кожи человека линии НМЕС-1, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль)
культура В - культура клеток эндотелия капилляров кожи человека линии НМЕС-1, трансфицированных ДНК-вектором VTvaf17
культура С - культура клеток эндотелия капилляров кожи человека линии НМЕС-1, трансфицированных ДНК-вектором VTvaf17-FMOD
На фиг. 10
показана диаграмма концентрации белка FMOD в биоптатах кожи трех пациентов после введения в кожу этих пациентов генотерапевтического ДНК-вектора VTvaf17-FMOD с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген FMOD.
На фиг. 10 отмечены следующие элементы:
П1I - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17-FMOD
П1II - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П1III - биоптат кожи пациента П1 из интактного участка,
П2I - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17-FMOD
П2II - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П2III - биоптат кожи пациента П2 из интактного участка,
П3I - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17-FMOD
П3II - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П3III - биоптат кожи пациента П3 из интактного участка
На фиг. 11
показана диаграмма концентрации белка TIMP2 в биоптатах икроножной мышцы трех пациентов после введения в икроножную мышцу этих пациентов генотерапевтического ДНК-вектора VTvaf17-TIMP2, с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген TIMP2.
На фиг. 11 отмечены следующие элементы:
П1I - биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17- TIMP2
П1II - биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П1III - биоптат интактного участка икроножной мышцы пациента П1
П2I - биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17- TIMP2
П2II - биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П2III - биоптат интактного участка икроножной мышцы пациента П2
П3I - биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17- TIMP2
П3II - биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П3III - биоптат интактного участка икроножной мышцы пациента П3
На фиг. 12
показана диаграмма концентрации белка TGFB3 в биоптатах кожи трех пациентов после введения в кожу этих пациентов генотерапевтического ДНК-вектора VTvaf17-TGFB3 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген TGFB3.
На фиг. 12 отмечены следующие элементы:
П1I - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17-TGFB3
П1II - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П1III - биоптат кожи пациента П1 из интактного участка,
П2I - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17-TGFB3
П2II - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П2III - биоптат кожи пациента П2 из интактного участка,
П3I - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17-TGFB3
П3II - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
П3III - биоптат кожи пациента П3 из интактного участка
На фиг. 13
показана диаграмма концентрации белка TGFB3 в биоптатах кожи человека после введения в кожу культуры аутогенных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3 с целью демонстрации способа применения путем введения аутологичных клеток, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3
На фиг. 13 отмечены следующие элементы:
П1А - биоптат кожи пациента П1 в зоне введения культуры аутогенных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3
П1В - биоптат кожи пациента П1 в зоне введения аутогенных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17
П1С - биоптат кожи пациента П1 из интактного участка.
На фиг. 14
показана диаграмма концентраций белков: белка SKI человека, белка TGFB3 человека, белка TIMP2 человека, белка FMOD человека в биоптатах трех крыс в зоне предварительно хирургически-смоделированного рубца плоской раны кожи у крыс линии Wistar после инъекционного введения в зону рубца смеси генотерапевтических векторов: генотерапевтического ДНК-вектора VTvaf17-SKI, генотерапевтического ДНК-вектора VTvaf17-TGFB3, генотерапевтического ДНК-вектора VTvaf17-TIMP2, генотерапевтического ДНК-вектора VTvaf17-FMOD с целью демонстрации способа применения смеси генотерапевтических ДНК-векторов.
На фиг. 14 отмечены следующие элементы:
К1I - биоптат кожи крысы К1 в зоне введения смеси генотерапевтических ДНК векторов: VTvaf17- SKI, VTvaf17- TGFB3, VTvaf17- TIMP2 и VTvaf17- FMOD
К1II - биоптат кожи крысы К1 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
К1III - биоптат контрольного интактного участка кожи крысы К1,
К2I - биоптат кожи крысы К2 в зоне введения смеси генотерапевтических ДНК векторов: VTvaf17-SKI, VTvaf17-TGFB3, VTvaf17-TIMP2 и VTvaf17-FMOD
К2II - биоптат кожи крысы К2 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
K2III - биоптат контрольного интактного участка кожи крысы К2,
К3I - биоптат кожи крысы К3 в зоне введения смеси генотерапевтических ДНК векторов: VTvaf17- SKI, VTvaf17- TGFB3, VTvaf17- TIMP2 и VTvaf17- FMOD
K3II - биоптат кожи крысы К3 в зоне введения генотерапевтического ДНК вектора VTvaf17 (плацебо)
К3III - биоптат контрольного интактного участка кожи крысы К3.
На фиг. 15
показаны графики накопления ампликонов кДНК целевого гена TIMP2 в клетках фибробластов кожи быка (ScienCell, Кат. #В2300) до и через 48 часов после трансфекции этих клеток ДНК-вектором VTvaf17-TIMP2 с целью демонстрации способа применения путем введения генотерапевтического ДНК-вектора животным
На фиг. 15 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена TIMP2 в клетках фибробластов кожи быка до трансфекции генотерапевтическим ДНК-вектором VTvaf17- TIMP2
2 - кДНК гена TIMP2 в клетках фибробластов кожи быка после трансфекции генотерапевтическим ДНК-вектором VTvaf17- TIMP2
3 - кДНК гена ACT в клетках фибробластов кожи быка до трансфекции генотерапевтическим ДНК-вектором VTvaf17- TIMP2
4 - кДНК гена ACT в клетках фибробластов кожи быка после трансфекции генотерапевтическим ДНК-вектором VTvaf17- TIMP2
В качестве референтного гена использовали ген актина быка/ коровы (ACT), приведенного в базе данных GenBank под номером АН001130.2.
Реализация изобретения
На основе ДНК-вектора VTvaf17 размером 3165 п.н. созданы генотерапевтические ДНК-векторы, несущие целевые гены человека, предназначенные для повышения уровня экспрессии этих целевых генов в тканях человека и животных. При этом способ получения каждого генотерапевтического ДНК-вектора, несущего целевые гены заключается в том, что в полилинкер генотерапевтического ДНК-вектора VTvaf17 клонируют белок-кодирующую последовательность целевого гена, выбранного из группы генов: ген SKI (кодирует белок SKI), ген TGFB3 (кодирует белок TGFB3), ген TIMP2 (кодирует белок TIMP2), ген FMOD (кодирует белок FMOD) человека. Известно, что способность ДНК-векторов проникать в эукариотические клетки обусловлена, главным образом, размером вектора. При этом ДНК-вектора с наименьшим размером обладают более высокой проникающей способностью. Таким образом, предпочтительным является отсутствие в составе вектора элементов, которые не несут функциональной нагрузки, но при этом увеличивают размер ДНК-вектора. Данные особенности ДНК-векторов были учтены при получении генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SKI, TGFB3, TIMP2, FMOD путем отсутствия в составе вектора крупных нефункциональных последовательностей и генов антибиотикорезистентности, что позволило, помимо технологических преимуществ и преимуществ в плане безопасности применения, значительно уменьшить размер полученного генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SKI, TGFB3, TIMP2, FMOD. Таким образом, способность проникать в эукариотические клетки полученного генотерапевтического ДНК-вектора обусловлена его небольшими размерами.
Каждый из генотерапевтических ДНК-векторов: ДНК-вектор VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD получали следующим образом: кодирующую часть целевого гена SKI, или TGFB3, или TIMP2, или FMOD клонировали в генотерапевтический ДНК-вектор VTvaf17 и получали генотерапевтический ДНК-вектор VTvaf17-SKI, SEQ ID №1, или VTvaf17-TGFB3, SEQ ID №2 или VTvaf17-TIMP2, SEQ ID №3, или VTvaf17-FMOD, SEQ ID №4 соответственно. Кодирующую часть гена SKI размером 2228 п.н., или гена TGFB3 размером 1252 п.н., или гена TIMP2 размером 704 п.н., или гена FMOD размером 1146 п.н. получали путем выделения суммарной РНК из биологического образца ткани здорового человека. Для получения первой цепи кДНК генов SKI, TGFB3, TIMP2, FMOD человека использовали реакцию обратной транскрипции. Амплификацию проводили с использованием созданных для этого методом химического синтеза олигонуклеотидов. Расщепление продукта амплификации специфическими эндонуклеазами рестрикции проводили с учетом оптимальной процедуры дальнейшего клонирования, причем клонирование в генотерапевтический ДНК-вектор VTvaf17 проводили по сайтам рестрикции BamHI, EcoRI, HindIII расположенными в полилинкере вектора VTvaf17. Выбор сайтов рестрикции проводили таким образом, чтобы клонированный фрагмент попадал в рамку считывания экспрессионной кассеты вектора VTvaf17, при этом белок-кодирующая последовательность не содержала сайты рестрикции для выбранных эндонуклеаз. При этом специалистам в данной области техники понятно, что методическая реализация получения генотерапевтического ДНК-вектора VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD может варьировать в рамках выбора известных методов молекулярного клонирования генов, при этом эти способы подпадают под объем настоящего изобретения. Так, например, могут быть использованы различные последовательности олигонуклеотидов для амплификации гена SKI, или TGFB3, или TIMP2, или FMOD различные эндонуклеазы рестрикции или такие лабораторные техники как безлигазное клонирование генов.
Генотерапевтический ДНК-вектор VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD обладает нуклеотидной последовательностью SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 соответственно. При этом специалистам в данной области техники известно свойство вырожденности генетического кода, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей, отличающихся инсерцией, делецией или заменой нуклеотидов, которые не приводят к изменению полипептидной последовательности, кодируемой целевым геном, и/или не приводят к потере функциональной активности регуляторных элементов вектора VTvaf17. При этом специалистам в данной области техники известно явление генетического полиморфизма, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей генов из группы генов SKI, TGFB3, TIMP2, FMOD, которые при этом кодируют различные варианты аминокислотных последовательностей белков SKI, TGFB3, TIMP2, FMOD, не отличающихся от приведенных по своей функциональной активности при физиологических условиях.
Способность проникать в эукариотические клетки и функциональную активность, то есть способность экспрессировать целевой ген, полученного генотерапевтического ДНК-вектора VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD подтверждают путем введения в эукариотические клетки полученного вектора и последующим анализом экспрессии специфической мРНК и/или белкового продукта целевого гена. Наличие специфической мРНК в клетках, в которые был введен генотерапевтический ДНК-вектор VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD свидетельствует как о способности полученного вектора проникать в эукариотические клетки, так и о его способности экспрессировать мРНК целевого гена. При этом, как известно специалистам в данной области техники, наличие мРНК гена является обязательным условием, но не доказательством трансляции белка, кодируемого целевым геном. Поэтому для подтверждения свойства генотерапевтического ДНК-вектора VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD экспрессировать целевой ген на уровне белка в эукариотических клетках, в которые был введен генотерапевтический ДНК-вектор, проводят анализ концентрации белков, кодируемых целевыми генами, с использованием иммунологических методов. Наличие белка SKI, или TGFB3, или TIMP2, или FMOD подтверждает эффективность экспрессии целевых генов в эукариотических клетках и возможность повышения уровня концентрации белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SKI, TGFB3, TIMP2, FMOD. Таким образом для подтверждения эффективности экспрессии созданного генотерапевтического ДНК-вектора VTvaf17-SKI, несущего целевой ген, а именно, ген SKI, генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего целевой ген, а именно, ген TGFB3, генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего целевой ген, а именно, ген TIMP2, генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего целевой ген, а именно, ген FMOD использовали следующие методы:
A) ПЦР в реальном времени - изменение накопления ампликонов кДНК целевых генов в лизате клеток человека и животного, после трансфекции различных клеточных линий человека и животного генотерапевтическим ДНК-векторами;
B) Иммуноферментный анализ - изменение количественного уровня целевых белков в лизате клеток человека, после трансфекции различных клеточных линий человека генотерапевтическими ДНК-векторами;
C) Иммуноферментный анализ - изменение количественного уровня целевых белков в супернатанте биоптатов тканей человека и животного, после введения в эти ткани генотерапевтических ДНК-векторов;
D) Иммуноферментный анализ - изменение количественного уровня целевых белков в супернатанте биоптатов тканей человека, после введения в эти ткани аутологичных клеток этого человека, трансфицированных генотерапевтическими ДНК-векторами.
Для подтверждения реализуемости способа применения созданного генотерапевтического ДНК-вектора VTvaf17-SKI, несущего целевой ген, а именно, ген SKI, генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего целевой ген, а именно, ген TGFB3, генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего целевой ген, а именно, ген TIMP2, генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего целевой ген, а именно, ген FMOD выполняли:
A) трансфекцию генотерапевтическими ДНК-векторами различных клеточных линий человека и животного;
B) введение генотерапевтических ДНК-векторов в различные ткани человека и животного;
C) введение в ткани животного смеси генотерапевтических ДНК-векторов;
D) введение в ткани человека аутологичных клеток, трансфицированных генотерапевтическими ДНК-векторами.
Указанные способы применения характеризуются отсутствием потенциальных рисков для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов, и за счет отсутствия в составе генотерапевтического ДНК-вектора генов устойчивости к антибиотикам, что подтверждается отсутствием участков, гомологичных вирусным геномам и генам антибиотикорезистентности в нуклеотидных последовательностях генотерапевтического ДНК-вектора VTvaf17-SKI, или генотерапевтического ДНК-вектора VTvaf17-TGFB3, или генотерапевтического ДНК-вектора VTvaf17-TIMP2, или генотерапевтического ДНК-вектора VTvaf17-FMOD (SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 соответственно).
Как известно специалистам в данной области техники, гены антибиотикорезистентности в составе генотерапевтических ДНК-векторов используются с целью получения этих векторов в препаративных количествах путем наращивания бактериальной биомассы в питательной среде, содержащей селективный антибиотик. В рамках настоящего изобретения в целях возможности безопасного применения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SKI или TGFB3, или TIMP2 или FMOD, использование селективных питательных сред, содержащих антибиотик, не представляется возможным. В качестве технологического решения для получения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SKI, TGFB3, TIMP2, FMOD для возможности масштабирования до промышленных масштабов получения генотерапевтических векторов предлагается способ получения штаммов для наработки указанных генотерапевтических векторов на основе бактерии Escherichia coli SCS110-AF. Способ получения штамма Escherichia coli SCS110-AF/VTvaf17-SKI или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AF/VTvaf17-FMOD заключается в получении компетентных клеток штамма Escherichia coli SCS110-AF с введением в эти клетки генотерапевтического ДНК-вектора VTvaf17-SKI, или ДНК-вектора VTvaf17-TGFB3, или ДНК-вектора VTvaf17-TIMP2, или ДНК-вектора VTvaf17-FMOD соответственно с помощью методов трансформации (электропорации), общеизвестных специалистам в данной области техники. Полученный штамм Escherichia coli SCS110-AF/VTvaf17-SKI, или штамм Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамм Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамм Escherichia coli SCS110-AF/VTvaf17-FMOD используется для наработки генотерапевтического ДНК-вектора VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD соответственно с возможностью использования сред без содержания антибиотиков.
Для подтверждения получения штамма Escherichia coli SCS110-AF/VTvaf17-SKI, или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AF/VTvaf17-FMOD проводили трансформацию, селекцию и последующее наращивание с выделением плазмидной ДНК.
Для подтверждения технологичности получения и возможности масштабирования до промышленного производства генотерапевтического ДНК-вектора VTvaf17-SKI, несущего целевой ген, а именно, ген SKI, генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего целевой ген, а именно, ген TGFB3, генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего целевой ген, а именно, ген TIMP2 генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего целевой ген, а именно, ген FMOD, выполняли ферментацию в промышленном масштабе штамма Escherichia coli SCS110-AF/VTvaf17-SKI или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3 или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AF/VTvaf17-FMOD, каждый из которых содержит генотерапевтический ДНК-вектор VTvaf17, несущий целевой ген, а именно SKI, или TGFB3, или TIMP2, или FMOD.
Способ масштабирования получения бактериальной массы до промышленных масштабов для выделения генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов SKI, TGFB3, TIMP2, FMOD, заключается в том, что затравочную культуру штамма Escherichia coli SCS110-AF/VTvaf17-SKI, или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AF/VTvaf17-FMOD инкубируют в объеме питательной среды без содержания антибиотика обеспечивающим подходящую динамику накопления биомассы, по достижению достаточного количества биомассы в логарифмической фазе роста, бактериальную культуру переносят в промышленный ферментер, после чего растят до достижения стационарной фазы роста, затем выделяют фракцию, содержащую целевой ДНК-продукт - генотерапевтический ДНК-вектор VTvaf17-SKI, или генотерапевтический ДНК-вектор VTvaf17-TGFB3, или генотерапевтический ДНК-вектор VTvaf17-TIMP2, или генотерапевтический ДНК-вектор VTvaf17-FMOD многостадийно фильтруют и очищают хроматографическими методами. При этом специалистам в данной области техники понятно, что условия культивирования штаммов, состав питательных сред (за исключением содержания антибиотиков), используемое оборудование, методы очистки ДНК могут варьировать в рамках стандартных операционных процедур в зависимости от отдельно взятой производственной линии, но известные подходы к масштабированию, промышленному получению и очистке ДНК-векторов с использованием штамма Escherichia coli SCS110-AF/VTvaf17-SKI, или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AF/VTvaf17-FMOD, или подпадают под объем настоящего изобретения.
Описанное раскрытие изобретения подтверждается примерами реализации настоящего изобретения.
Изобретение поясняется следующими примерами.
Пример 1.
Получение генотерапевтического ДНК-вектора VTvaf17-SKI, несущего целевой ген, а именно, гена SKI.
Генотерапевтический ДНК-вектор VTvaf17-SKI конструировали клонированием кодирующей части гена SKI размером 2228 п. н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции BamHI и HindIII. Кодирующую часть гена SKI размером 2228 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
SKI_F CAGGATCCGCGGGAGCGGCCGGGGGAG
SKI_R TATAAGCTTACGGCTCCAGCTCCGCAG
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США).
Генотерапевтический ДНК-вектор VTvaf17 конструировали объединением шести фрагментов ДНК, полученных из разных источников:
(а) ориджин репликации получали путем ПЦР-амплификации участка коммерческой плазмиды pBR322 с внесением точечной мутации;
(б) промоторный регион EF1a получали путем ПЦР-амплификации участка геномной ДНК человека;
(в) терминатор транскрипции hGH-TA получали путем ПЦР-амплификации участка геномной ДНК человека;
(г) регуляторный участок транспозона Tn10 PHK-out получали путем синтеза из олигонуклеотидов;
(д) ген устойчивости к канамицину получали путем ПЦР-амплификации участка коммерческой плазмиды рЕТ-28 человека;
(е) полилинкер получали отжигом двух синтетических олигонуклеотидов.
ПЦР-амплификацию проводили с использованием коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США) в соответствии с инструкцией производителя. Фрагменты имеют перекрывающиеся области для возможности их объединения с последующей ПЦР-амплификацией. Объединяли фрагменты (а) и (б) с использованием олигонуклеотидов Ori-F и EF1-R, а также фрагменты (в), (г) и (д) с использованием олигонуклеотидов hGH-F и Kan-R. Далее, полученные участки объединяли путем рестрикции с последующим лигированием по сайтам BamHI и Ncol. В результате получали плазмиду, пока еще не содержащую полилинкер. Для его введения проводили расщепление плазмиды по сайтам BamHI и EcoRI, и лигирование с фрагментом (е). Таким образом, получали вектор размером 3165 п.н., несущий ген устойчивости к канамицину, который фланкирован сайтами рестрикции Spel. Далее этот участок выщепляли по сайтам рестрикции Spel, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор VTvaf17 размером 3165 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков.
Расщепление продукта амплификации кодирующей части гена SKI и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции BamHI и HindIII (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-SKI размером 5369 п.н. с нуклеотидной последовательностью SEQ ID №1 и общей структурой изображенной на фиг. 1А.
Пример 2.
Получение генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего целевой ген, а именно, гена TGFB3.
Генотерапевтический ДНК-вектор VTvaf17-TGFB3 конструировали клонированием кодирующей части гена TGFB3 размером 1252 п.н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции BamHI и HindIII. Кодирующую часть гена TGFB3 размером 1252 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
TGFB3_F GGATCCACCATGAAGATGCACTTGCAAAGG
TGFB3_R TATAAGCTTAGCTACATTTACAAGACTTCACCA
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции BamHI и HindIII (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-TGFB3 размером 4393 п.н. с нуклеотидной последовательностью SEQ ID №2 и общей структурой изображенной на фиг. 1В.
При этом генотерапевтический ДНК-вектор VTvaf17 конструировали по Примеру 1.
Пример 3
Получение ДНК-вектора VTvaf17-TIMP2, несущего целевой ген, а именно, гена TIMP2 человека.
Генотерапевтический ДНК-вектор VTvaf17-TIMP2 конструировали клонированием кодирующей части гена TIMP2 размером 704 п.н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции BamHI и HindIII. Кодирующую часть гена TIMP2 размером 704 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
TIMP2_F CAGGATCCGGCCCCCGCCCGCCCAGC
TIMP2_R TATAAGCTTATGGGTCCTCGATGTCGAGA
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции BamHI и HindIII (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-TIMP2 размером 3845 п.н. с нуклеотидной последовательностью SEQ ID №3 и общей структурой изображенной на фиг. 1С.
При этом генотерапевтический ДНК-вектор VTvaf17 конструировали по Примеру 1.
Пример 4.
Получение генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего целевой ген, а именно, гена FMOD.
Генотерапевтический ДНК-вектор VTvaf17-FMOD конструировали клонированием кодирующей части гена FMOD размером 1146 п.н. в ДНК-вектор VTvaf17 размером 3165 п.н. по сайтам рестрикции BamHII и EcoRI. Кодирующую часть гена FMOD размером 1146 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
FMOD_F GGATCCACCATGCAGTGGACCTCCCTCCT
FMOD_R TATGAATTCTTAGATCTCGATGAGGCTGGCA
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора VTvaf17 проводили эндонуклеазами рестрикции BamHII и EcoRI (New England Biolabs, США).
В результате получали ДНК-вектор VTvaf17-FMOD размером 4275 п.н. с нуклеотидной последовательностью SEQ ID №4 и общей структурой изображенной на фиг. 1D.
При этом генотерапевтический ДНК-вектор VTvaf17 конструировали по Примеру 1.
Пример 5.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-SKI, несущего целевой ген, а именно, ген SKI, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена SKI, в первичной культуре фибробластов кожи человека HDFa (АТСС PCS-201-01) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-SKI, несущим ген SKI человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Для оценки изменения накопления мРНК целевого гена SKI, использовалась первичная культура фибробластов кожи человека HDFa. Клеточную культуру HDFa выращивали в стандартных условиях (37°С, 5% CO2) с использованием питательной среды Fibroblast Growth Kit-Serum-Free (АТСС® PCS-201-040). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-SKI, экспрессирующим ген SKI человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) согласно рекомендациям производителя. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора ДНК-вектора VTvaf17-SKI (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивали легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора Lipofectamin 3000. Аккуратно перемешивали легким встряхиванием. Добавляли содержимое пробирки №1 к содержимому пробирки №2, инкубировали 5 мин при комнатной температуре. Полученный раствор по каплям добавляли к клеткам в объеме 40 мкл.
В качестве контроля использовали клетки HDFa, трансфицированные генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена (кДНК гена SKI до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена на фигурах не показано) Подготовку контрольного вектора VTvaf17 для трансфекции проводили как описано выше.
Суммарную РНК из клеток HDFa выделяли с использованием Trizol Reagent (Invitrogen, США) согласно рекомендациям производителя. В лунку с клетками добавляли 1 мл Trizol Reagent и гомогенизировали с последующим прогреванием в течении 5 мин при 65°С. Далее образец центрифугировали при 14000 g в течении 10 мин и снова прогревали в течении 10 мин при 65°С. Далее добавляли 200 мкл хлороформа, плавно перемешивали и центрифугировали при 14000 g в течении 10 мин. Затем отбирали водную фазу, добавляли к ней 1/10 объема 3М ацетата натрия, рН 5.2 и равный объем изопропилового спирта. Инкубировали образец при -20°С в течении 10 мин с последующим центрифугированием при 14000 g в течении 10 мин. Осадок промывали 1 мл 70% этилового спирта, высушивали на воздухе и растворяли в 10 мкл воды, свободной от РНКаз. Определение уровня экспрессии мРНК гена SKI после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени. Для получения и амплификации кДНК, специфичной для гена SKI человека, использовали олигонуклеотиды SKI_SF и SKI_SR:
SKI_SF TGGACACCAAGGAAGCCAAAG
SKI_SR GCTGTTCCTGCAGCTCCTTCAC.
Длина продукта амплификации - 468 п.н.
Реакцию обратной транскрипции и ПЦР-амплификацию проводили с помощью набора реагентов SYBR GreenQuantitect RT-PCR Kit (Qiagen, США) для ПЦР в режиме реального времени. Реакцию проводили в объеме 20 мкл, содержащих: 25 мкл QuantiTect SYBR Green RT-PCR MasterMix, 2,5 мМ хлорида магния, по 0,5 мкМ каждого праймера, 5 мкл РНК. Реакцию осуществляли на амплификаторе CFX96 (Bio-Rad, США) при следующих условиях: 1 цикл обратной транскрипции при 42°С - 30 минут, денатурация 98°С - 15 мин, затем 40 циклов, включающих денатурацию 94°С - 15 сек, отжиг праймеров 60°С - 30 сек и элонгацию 72°С - 30 сек. В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов SKI и В2М. В качестве отрицательного контроля использовали деионизированную воду. Количество динамику накопления ампликонов кДНК генов SKI и В2М оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 2.
Из фигуры 2 следует, что в результате трансфекции первичной культуры клеток фибробластов человека HDFa генотерапевтическим ДНК-вектором VTvaf17-SKI, уровень специфической мРНК гена SKI человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SKI на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-SKI для повышения уровня экспрессии гена SKI в эукариотических клетках.
Пример 6.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего целевой ген, а именно, ген TGFB3, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена TGFB3, в первичной культуре кератиноцитов эпидермиса человека НЕКа (АТСС PCS-200-011) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-TGFB3, несущим ген TGFB3 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Первичную культуру клеток кератиноцитов эпидермиса человека НЕКа выращивали в среде Keratinocyte Growth Kit (АТСС® PCS-200-040™) в стандартных условиях (37°С, 5% CO2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-TGFB3, экспрессирующим ген TGFB3 человека, проводили как описано в примере 5. В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток НЕКа, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген (кДНК гена TGFB3 до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 5, за исключением олигонуклеотидов с отличающимися от примера 5 последовательностями. Для амплификации кДНК, специфичной для гена TGFB3 человека, использовали олигонуклеотиды TGFB3_SF и TGFB3_SR:
TGFB3_SF TGAGCACATTGCCAAACAGC
TGFB3_SR GAGGCAGATGCTTCAGGGTT.
Длина продукта амплификации - 594 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов TGFB3 и В2М. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов - кДНК генов TGFB3 и В2М, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 3.
Из фигуры 3 следует, что в результате трансфекции культуры клеток НЕКа генотерапевтическим ДНК-вектором VTvaf17-TGFB3, уровень специфической мРНК гена TGFB3 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TGFB3 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-TGFB3 для повышения уровня экспрессии гена TGFB3 в эукариотических клетках.
Пример 7.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего целевой ген, а именно, ген TIMP2, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена TIMP2 в культуре клеток фибробластов крайней плоти человека линии Hs27 (АТСС CRL-1634) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-TIMP2, несущим ген TIMP2 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Культуру клеток фибробластов крайней плоти человека линии Hs27 выращивали в среде Dulbecco's Modified Eagle's Medium (DMEM) (АТСС® 30-2002™ с добавлением 10% сыворотки крупного рогатого скота (АТСС® 30-2020™) в стандартных условиях (37°С, 5% CO2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-TIMP2, экспрессирующим ген TIMP2 человека, проводили как описано в примере 5. В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток линии Hs27, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген (кДНК гена TIMP2 до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 5, за исключением олигонуклеотидов с отличающимися от примера 5 последовательностями. Для амплификации кДНК, специфичной для гена TIMP2 человека, использовали олигонуклеотиды TIMP2_SF и TIMP2_SR:
TIMP2_SF GCAACAGGCGTTTTGCAATG
TIMP2_SR AGGGCACGATGAAGTCACAG.
Длина продукта амплификации - 296 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов TIMP2 и В2М. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов - кДНК генов TIMP2 и В2М, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 4.
Из фигуры 4 следует, что в результате трансфекции культуры клеток фибробластов крайней плоти человека линии Hs27 генотерапевтическим ДНК-вектором VTvaf17-TIMP2 уровень специфической мРНК гена TIMP2 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TIMP2 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-TIMP2 для повышения уровня экспрессии гена TIMP2 в эукариотических клетках.
Пример 8.
Подтверждение способности генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего целевой ген, а именно, ген FMOD, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена FMOD, в культуре клеток эндотелия капилляров кожи человека линии НМЕС-1 (АТСС CRL-3243) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17-FMOD, несущим ген FMOD человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Культуру клеток эндотелия капилляров кожи человека НМЕС-1 выращивали в среде MCDB131 (Gibco™, Кат.10372019) без содержания глутамина и с добавлением 10 нг/мл рекомбинантного EGF (Sigma, Е9644, США), 10 мМ глутамина (Панэко, Россия), 1 мкг/мл гидрокортизона (Sigma Н0888, США), 10% HyClone™ Fetal Bovine Serum (Hyclone Laboratories Inc SH30068.03HI, США) в стандартных условиях (37°С, 5% CO2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17-FMOD, экспрессирующим ген FMOD человека, проводили как описано в примере 5. В качестве контроля использовали культуру клеток НМЕС-1, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим целевой ген (кДНК гена FMOD до и после трансфекции генотерапевтическим ДНК-вектором VTvaf17, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 5, за исключением олигонуклеотидов с отличающимися от примера 5 последовательностями. Для амплификации кДНК, специфичной для гена FMOD человека, использовали олигонуклеотиды FMOD_SF и FMOD_SR:
FMOD_SF CCTACACCTACGGCTCTCCA
FMOD_SR CATCCGGGTCAGGTTGTTGT.
Длина продукта амплификации - 326 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов FMOD и В2М. В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов -кДНК генов FMOD и В2М, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 5.
Из фигуры 5 следует, что в результате трансфекции культуры клеток эндотелия капилляров кожи человека НМЕС-1 генотерапевтическим ДНК-вектором VTvaf17-FMOD, уровень специфической мРНК гена FMOD человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген FMOD на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором VTvaf17-FMOD для повышения уровня экспрессии гена FMOD в эукариотических клетках.
Пример 9.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-SKI, несущего ген SKI, для повышения экспрессии белка SKI в клетках млекопитающих.
Оценивали изменение количества белка SKI в лизате фибробластов кожи человека HDFa (АТСС PCS-201-01) после трансфекции этих клеток ДНК-вектором VTvaf17-SKI, несущим ген SKI человека.
Клеточную культуру фибробластов кожи человека выращивали как это описано в примере 5.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена SKI (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-SKI, несущий ген SKI человека. Приготовление комплекса ДНК/дендример проводили по методике производителя (QIAGEN, SuperFect Transfection Reagent Handbook, 2002) с некоторыми изменениями. Для трансфекции клеток в одной лунке 24-луночного планшета к 1 мкг ДНК-вектора, растворенного в буфере ТЕ, добавляли культуральную среду до конечного объема 60 мкл, затем добавляли 5 мкл SuperFect Transfection Reagent и осторожно перемешивали пятикратным пипетированием. Комплекс инкубировали при комнатной температуре в течении 10-15 мин. Далее отбирали из лунок культуральную среду, лунки промывали 1 мл буфера PBS. К образовавшемуся комплексу добавляли 350 мкл среды, содержащей 10 мкг/мл гентамицина, осторожно перемешивали и добавляли к клеткам. Клетки инкубировали с комплексом 2-3 часа при 37°С в атмосфере, содержащей 5% CO2.
Затем осторожно удаляли среду, промывали клеточный монослой 1 мл буфера PBS. Затем добавляли среду, содержащую 10 мкг/мл гентамицина, и инкубировали 24-48 часов при 37°С в атмосфере 5% CO2.
После трансфекции клетки промывали три раза PBS, далее к клеткам добавляли 1 мл PBS и подвергали трехкратному замораживанию/оттаиванию. Далее суспензию центрифугировали при 15000 об/мин в течение 15 мин, отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка SKI проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор SKI ELISA Kit (Human) (Aviva Systems Biology Кат. OKCA01520, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка SKI, входящим в состав набора. Чувствительность метода составляла не менее 7,8 пг/мл, диапазон измерения - от 31.25 пг/мл до 2,000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 6.
Из фигуры 6 следует, что трансфекция клеток фибробластов кожи человека HDFa генотерапевтическим ДНК-вектором VTvaf17-SKI приводит к увеличению количества белка SKI по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген SKI на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-SKI для повышения уровня экспрессии SKI в эукариотических клетках.
Пример 10.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего ген TGFB3, для повышения экспрессии белка TGFB3 в клетках млекопитающих.
Оценивали изменение количества белка TGFB3 в кондиционированной среде лизате клеток первичной культуры кератиноцитов эпидермиса человека НЕКа (АТСС PCS-200-01) после трансфекции этих клеток ДНК-вектором VTvaf17-TGFB3, несущим ген TGFB3 человека. Клетки выращивали как описано в примере 6.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена TGFB3 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-TGFB3, несущий ген TGFB3 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток НЕКа проводили как описано в примере 9.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (рН 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка TGFB3 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human TGF-β3 (Transforming Growth Factor Beta 3) ELISA Kit (Elabscience Кат. E-EL-H2339, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка TGFB3, входящим в состав набора. Чувствительность метода составляла не менее 9,38 пг/мл, диапазон измерения - от 15.63 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 7.
Из фигуры 7 следует, что трансфекция первичной культуры клеток эндометрия матки человека генотерапевтическим ДНК-вектором VTvaf17-TGFB3 приводит к увеличению количества белка TGFB3 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TGFB3 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-TGFB3 для повышения уровня экспрессии TGFB3 в эукариотических клетках.
Пример 11.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего ген TIMP2, для повышения экспрессии белка TIMP2 в клетках млекопитающих.
Оценивали изменение количества белка TIMP2 в лизате клеток фибробластов крайней плоти человека линии Hs27 (АТСС CRL-1634) после трансфекции этих клеток ДНК-вектором VTvaf17-TIMP2, несущим ген TIMP2 человека. Клетки культивировали как описано в примере 7.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена TIMP2 (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-TIMP2, несущий ген TIMP2 человека. Приготовление комплекса ДНК-дендример и трансфекцию клеток Hs27 проводили как описано в примере 9.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (рН 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка TIMP2 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human TIMP2 ELISA Kit (Sandwich ELISA) (LifeSpan Bio Кат. LS-F196, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка TIMP2, входящим в состав набора. Чувствительность метода составляла не менее 9,6 пг/мл, диапазон измерения - от 9,6 пг/мл до 2333 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 8.
Из фигуры 8 следует, что трансфекция культуры клеток фибробластов крайней плоти человека линии Hs27 генотерапевтическим ДНК-вектором VTvaf17-TIMP2 приводит к увеличению количества белка TIMP2 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TIMP2 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-TIMP2 для повышения уровня экспрессии TIMP2 в эукариотических клетках.
Пример 12.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего ген FMOD, для повышения экспрессии белка FMOD в клетках млекопитающих.
Оценивали изменение количества белка FMOD в лизате культуры клеток эндотелия капилляров кожи человека линии НМЕС-1 (АТСС CRL-3243) после трансфекции этих клеток ДНК-вектором VTvaf17-FMOD, несущим ген FMOD человека. Клетки культивировали как описано в примере 7.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор VTvaf17, не содержащий кДНК гена FMOD (В), в качестве трансфицируемых агентов - ДНК-вектор VTvaf17-FMOD, несущий ген FMOD человека. Приготовление комплекса ДНК-дендримеры и трансфекцию клеток НМЕС-1 проводили как описано в примере 9.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (рН 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка FMOD проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human FMOD / Fibromodulin ELISA Kit (Sandwich ELISA) (LifeSpan Bio Кат. LS-F4730, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка FMOD, входящим в состав набора. Чувствительность метода составляла не менее 14,3 пг/мл, диапазон измерения - от 31,25 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 9.
Из фигуры 9 следует, что трансфекция культуры клеток эндотелия капилляров кожи человека линии НМЕС-1 генотерапевтическим ДНК-вектором VTvaf17-FMOD приводит к увеличению количества белка FMOD по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген FMOD на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора VTvaf17-FMOD для повышения уровня экспрессии FMOD в эукариотических клетках.
Пример 13.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего ген FMOD, для повышения экспрессии белка FMOD в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего целевой ген, а именно, ген FMOD и реализуемости способа его применения оценивали изменения количества белка FMOD в коже человека при введении в кожу человека генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего ген человека FMOD.
С целью анализа изменения количества белка FMOD трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор VTvaf17-FMOD, несущий ген FMOD и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор VTvaf17, не содержащий кДНК гена FMOD.
Пациент 1, женщина 59 лет, (П1); пациент 2, женщина 56 лет, П2); пациент 3, мужчина, 52 года, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор VTvaf17-FMOD, который содержит кДНК гена FMOD и генотерапевтический ДНК-вектор VTvaf17, используемый в качестве плацебо, который не содержит кДНК гена FMOD, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор VTvaf17 (плацебо) и генотерапевтический ДНК вектор VTvaf17-FMOD, несущий ген FMOD вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора VTvaf17 (плацебо) и генотерапевтического ДНК вектора VTvaf17-FMOD, несущего ген FMOD - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора VTvaf17-FMOD, несущего ген FMOD (I), генотерапевтического ДНК-вектора VTvaf17 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000 g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа (ELISA), используя набор Human FMOD / Fibromodulin ELISA Kit (Sandwich ELISA) (LifeSpan Bio Кат. LS-F4730, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка FMOD, входящим в состав набора. Чувствительность метода составляла не менее 14,3 пг/мл, диапазон измерения - от 31,25 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 10.
Из фигуры 10 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора VTvaf17- FMOD, несущего целевой ген FMOD человека, произошло увеличение количества белка FMOD в сравнении с количеством белка FMOD в области введения генотерапевтического ДНК-вектора VTvaf17 (плацебо), не содержащего ген FMOD человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-FMOD и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 14.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-TIMP, несущего ген TIMP, для повышения экспрессии белка TIMP в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-TIMP, несущего целевой ген TIMP и реализуемости способа его применения оценивали изменение количества белка TIMP в мышечной ткани человека при введении генотерапевтического ДНК-вектора VTvaf17-TIMP, несущего целевой ген, а именно, ген TIMP человека.
С целью анализа изменения количества белка TIMP трем пациентам вводили в ткань икроножной мышцы генотерапевтический ДНК-вектор VTvaf17-TIMP, несущий ген TIMP с транспортной молекулой, и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор VTvaf17, не содержащий кДНК гена TIMP с транспортной молекулой.
Пациент 1, женщина 51 год, (П1); пациент 2, женщина 42 года (П2); пациент 3, мужчина, 62 года, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция), пробоподготовку проводили в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор VTvaf17 (плацебо) и генотерапевтический ДНК вектор VTvaf17-TIMP, несущий ген TIMP вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину около 10 мм. Объем вводимого раствора генотерапевтического ДНК-вектора VTvaf17 (плацебо) и генотерапевтического ДНК вектора VTvaf17-TIMP, несущего ген TIMP - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагали медиально на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков мышечной ткани пациентов в зоне введения генотерапевтического ДНК вектора VTvaf17-TIMP, несущего ген TIMP (I), генотерапевтического ДНК-вектора VTvaf17 (плацебо) (II), а также из интактного участка икроножной мышцы (III), используя устройство для взятия биопсии MAGNUM (BARD, США). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 20 куб. мм, масса - около 22 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000 g. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка TIMP проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор используя набор Human TIMP2 ELISA Kit (Sandwich ELISA) (LifeSpan Bio Кат. LS-F196, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка TIMP2, входящим в состав набора. Чувствительность метода составляла не менее 9,6 пг/мл, диапазон измерения - от 9,6 пг/мл до 2333 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 11.
Из фигуры 11 следует, что в икроножной мышце всех трех пациентов в области введения генотерапевтического ДНК вектора VTvaf17-TIMP, несущего целевой ген, а именно, ген TIMP человека, произошло увеличение количества белка TIMP в сравнении с количеством белка TIMP в области введения генотерапевтического ДНК-вектора VTvaf17 (плацебо), не содержащего ген TIMP человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-TIMP и подтверждает реализуемость способа его применения, в частности при внутримышечном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 15.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего ген TGFB3, для повышения экспрессии белка TGFB3 в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего целевой ген, а именно, ген TGFB3 и реализуемости способа его применения оценивали изменения количества белка TGFB3 в коже человека при введении в кожу человека генотерапевтического ДНК-вектора VTvafl 7-TGFB3, несущего ген человека TGFB3.
С целью анализа изменения количества белка TGFB3 трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор VTvaf17-TGFB3, несущий ген TGFB3 и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор VTvaf17, не содержащий кДНК гена TGFB3.
Пациент 1, женщина 48 лет (П1); пациент 2, мужчина 49 лет (П2); пациент 3, мужчина, 57 лет (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор VTvaf17-TGFB3, который содержит кДНК гена TGFB3 и генотерапевтический ДНК-вектор VTvaf17, используемый в качестве плацебо, который не содержит кДНК гена TGFB3, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор VTvaf17 (плацебо) и генотерапевтический ДНК вектор VTvaf17-TGFB3, несущий ген TGFB3 вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора VTvaf17 (плацебо) и генотерапевтического ДНК вектора VTvaf17-TGFB3, несущего ген TGFB3 - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора VTvaf17-TGFB3, несущего ген TGFB3 (I), генотерапевтического ДНК-вектора VTvaf17 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000 g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа (ELISA), используя набор используя набор Human TGF-β3 (Transforming Growth Factor Beta 3) ELISA Kit (Elabscience Кат. E-EL-H2339, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка TGFB3, входящим в состав набора. Чувствительность метода составляла не менее 9,38 пг/мл, диапазон измерения - от 15.63 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 12.
Из фигуры 12 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора VTvaf17- TGFB3, несущего целевой ген TGFB3 человека, произошло увеличение количества белка TGFB3 в сравнении с количеством белка TGFB3 в области введения генотерапевтического ДНК-вектора VTvaf17 (плацебо), не содержащего ген TGFB3 человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-TGFB3 и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 16.
Подтверждение эффективности генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего ген TGFB3 и реализуемости способа его применения для повышения уровня экспрессии белка TGFB3 в тканях человека путем введения аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего ген TGFB3 и реализуемости способа его применения оценивали изменения количества белка TGFB3 в коже человека при введении в кожу пациента культуры аутогенных фибробластов этого пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3.
Пациенту вводили в кожу предплечья соответствующую культуру аутогенных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3, несущим ген TGFB3, и параллельно вводили плацебо, представляющее собой культуру аутогенных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген TGFB3.
Первичную культуру фибробластов человека выделяли из биоптатов кожи пациента. Используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL, Италия) брали образец биоптата кожи из зоны, защищенной от действия ультрафиолета, а именно за ушной раковиной или с боковой внутренней поверхности в зоне локтевого сустава, размером около 10 кв. мм, массой - около 11 мг. Кожу пациента предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Культивирование первичной культуры клеток осуществляли при 37°С в атмосфере, содержащей 5% CO2 в среде DMEM с 10% фетальной телячьей сывороткой и ампициллином 100 Ед/мл. Пересев культуры и смену культуральной среды осуществляли каждые 2 дня. Общая продолжительность роста культуры не превышала 25-30 дней. Из культуры клеток отбирали аликвоту, содержащую 5×104 клеток. Культуру фибробластов пациента трансфицировали генотерапевтическим ДНК-вектором VTvaf17-TGFB3, несущим ген TGFB3 или плацебо - вектор VTvaf17, не несущий целевой ген TGFB3.
Трансфекцию осуществляли с помощью катионного полимера полиэтиленимина JETPEI (Polyplus Transfection, Франция), согласно инструкции фирмы-производителя. Клетки культивировали 72 часа и затем вводили пациенту. Введение пациенту культуры аутогенных фибробластов этого пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3, и плацебо, представляющее собой культуру аутогенных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, осуществляли туннельным методом в область предплечья иглой 30G длиной 13 мм на глубину около 3 мм. Концентрация модифицированных аутогенных фибробластов в вводимой суспензии составляла примерно 5 млн клеток на 1 мл суспензии, количество вводимых клеток не превышало 15 млн. Очаги введения культуры аутогенных фибробластов располагались на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 4-е сутки после введения культуры аутогенных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3, несущим целевой ген, а именно, ген TGFB3 и плацебо. Взятие биопсии осуществляли из участков кожи пациента в зоне введения культуры аутогенных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3, несущим целевой ген, а именно, ген TGFB3 (С), культуры аутогенных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvafl 7, не несущим целевой ген TGFB3 (плацебо) (В), а также из участков интактной кожи (А), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу в зоне отбора биоптата пациентов предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000 g. Отбирали супернатант и использовали его для количественного определения целевого белка как это описано в примере 15.
Графики, полученные в результате анализа, представлены на фиг. 13.
Из фигуры 13 следует, что в коже пациента в области введения культуры аутогенных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3, несущим ген TGFB3, произошло увеличение количества белка TGFB3 в сравнении с количеством белка TGFB3 в области введения культуры аутогенных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17, не несущим ген TGFB3 (плацебо), что свидетельствует об эффективности генотерапевтического ДНК-вектора VTvaf17-TGFB3 и подтверждает реализуемость способа его применения для повышения уровня экспрессии TGFB3 в тканях человека, в частности при введении аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором VTvaf17-TGFB3.
Пример 17.
Подтверждение эффективности и реализуемости способа сочетанного применения генотерапевтического ДНК-вектора VTvaf17-SKI, несущего ген SKI, генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего ген TGFB3, генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего ген TIMP2, генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего ген FMOD, для повышения уровня экспрессии белков SKI, TGFB3, TIMP2 и FMOD в тканях млекопитающих.
Оценивали изменение количества белков SKI, TGFB3, TIMP2 и FMOD в зоне предварительно хирургически-смоделированного рубца плоской раны кожи крысы при введении в эту зону смеси генотерапевтических векторов.
Рубец формировали у крыс линии Вистар под общей анестезией путем хирургического иссечения кожи на дорсальной поверхности с образованием раны округлой формы, диаметром 2 см процедуру проводили согласно известному из литературы описанию (Li, P., et al. //J Pathol, 2011. 223(5): p. 659-71).
В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Эквимолярную смесь генотерапевтических ДНК-векторов растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя. Объем вводимого раствора составлял 0,3 мл с суммарным количеством ДНК 100 мкг. Введение раствора осуществляли тоннельным методом иглой 30G на глубину 2-3 мм в зону предварительно хирургически-смоделированного рубца на коже крысы через 48 часов после формирования рубца.
Биопсийные образцы отбирали на 2 сутки после введения генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков рубца на коже животных в зоне введения смеси четырех генотерапевтических ДНК-векторов, несущих гены SKI, TGFB3, TIMP2, FMOD (зона I), генотерапевтического ДНК-вектора VTvaf17 (плацебо) (зона II), а также из участков аналогичного модельного рубца, не подвергавшегося никаким манипуляциям (зона III), используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL). Зону отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер каждого биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Каждый образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000 g. Отбирали супернатант и использовали его для количественного определения целевых белков как это описано в примере 9 (количественное определение белка SKI), примере 10 (количественное определение белка TGFB3), примере 11 (количественное определение белка TIMP2), примере 12 (количественное определение белка FMOD). Графики, полученные в результате анализа, представлены на фиг. 12.
Из фигур 14А и 14В следует, что в зоне предварительно хирургически-смоделированного рубца плоской раны кожи крыс (зона I), в которую вводилась смесь генотерапевтических векторов: генотерапевтического ДНК-вектора VTvaf17-SKI, несущего целевой ген SKI, генотерапевтического ДНК-вектора VTvaf17-TGFB3, несущего целевой ген TGFB3, генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего целевой ген TIMP2, генотерапевтического ДНК-вектора VTvaf17-FMOD, несущего целевой ген FMOD, произошло увеличение количества белков SKI, TGFB3, TIMP2 и FMOD по сравнению с зоной II (зона плацебо) и зоной III (интактная зона). Полученные результаты свидетельствуют об эффективности сочетанного применения генотерапевтических ДНК-векторов и подтверждает реализуемость способа применения для повышения уровня экспрессии целевых белков в тканях млекопитающих.
Пример 18.
Подтверждение эффективности генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего ген TIMP2 и реализуемости способа его применения для повышения уровня экспрессии белка TIMP2 в клетках млекопитающих.
Для подтверждения эффективности генотерапевтического ДНК-вектора VTvaf17-TIMP2, несущего ген TIMP2 оценивали изменение накопления мРНК целевого гена TIMP2 в клетках фибробластов кожи быка (ScienCell, Кат. #В2300) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором VTvaf17- TIMP2, несущим ген TIMP2 человека.
Клетки фибробластов кожи быка BDF (ScienCell, Кат. #В2300) выращивали в среде FM-2 (ScienCell, Кат. #2331). Трансфекцию генотерапевтическим ДНК-вектором VTvaf17- TIMP2, несущим ген TIMP2 человека и ДНК-вектором VTvaf17, не несущим ген TIMP2 человека (контороль), выделение РНК, реакцию обратной транскрипции, ПЦР амплификации и анализ данных проводили как описано в примере 7. В качестве референтного гена использовали ген актина быка/ коровы (ACT), приведенного в базе данных GenBank под номером АН001130.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности генов TIMP2 и ACT. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов - кДНК генов TIMP2 и ACT, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager2.1. (Bio-Rad, USA).
Графики, полученные в результате анализа представлены на фиг. 15.
Из фигуры 15 следует, что в результате трансфекции клеток фибробластов кожи быка BDF генотерапевтическим ДНК-вектором VTvaf17-TIMP2, уровень специфической мРНК гена TIMP2 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TIMP2 на уровне мРНК. Представленные результаты подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором VTvaf17-TIMP2 для повышения уровня экспрессии гена TIMP2 в клетках млекопитающих.
Пример 19.
Штамм Escherichia coli SCS110-AF/VTvaf17-SKI или штамм Escherichia coli SCS110-AF/VTvaf17-TGFB3 или штамм Escherichia coli SCS110-AF/VTvaf17-TIMP2 или штамм Escherichia coli SCS110-AF/VTvaf17-FMOD, несущий генотерапевтический ДНК-вектор, и способ его получения.
Конструирование штамма для наработки в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: SKI, TGFB3, TIMP2, FMOD, а именно штамма Escherichia coli SCS110-AF/VTvaf17-SKI, или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AF/VTvaf17-FMOD, несущего генотерапевтический ДНК-вектор VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD соответственно для его наработки с возможностью селекции без использования антибиотиков заключается в получении электрокомпетентных клеток штамма Escherichia coli SCS110-AF с проведением электропорации этих клеток генотерапевтическим ДНК-вектором VTvaf17-SKI, или ДНК-вектором VTvaf17-TGFB3, или ДНК-вектором VTvaf17-TIMP2, или ДНК-вектором VTvaf17-FMOD, после чего клетки высеваются на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола. При этом штамм Escherichia coli SCS110-AF для наработки генотерапевтического ДНК-вектора VTvaf17 или генотерапевтических ДНК-векторов на его основе с возможностью положительной селекции без использования антибиотиков получали путем конструирования линейного фрагмента ДНК, содержащего регуляторный элемент RNA-in транспозона Tn10 для селекции без применения антибиотиков размером 64 п.н., ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде размером 1422 п.н., ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация размером 763 п.н. и две гомологичные последовательности, обеспечивающие процесс гомологичной рекомбинации в области гена recA с одновременной его инактивацией размером 329 п.н. и 233 п.н., после чего проводили трансформацию клеток Escherichia coli путем электропорации и отбирали клоны, выжившие на среде, содержащей 10 мкг/мл хлорамфеникола.
Полученные штаммы для наработки были внесены в коллекции Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и NCIMB Patent Deposit Service, UK с присвоением регистрационных номеров:
штамм Escherichia coli SCS110-AF/VTvaf17-SKI - регистрационный № ВКПМ В-13198, INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43083,
штамм Escherichia coli SCS110-AF/VTvaf17-TGFB3 регистрационный № ВКПМ B-13197, INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43084,
штамм Escherichia coli SCS110-AF/VTvaf17-TIMP2 регистрационный № ВКПМ B-13196, INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43085,
штамм Escherichia coli SCS110-AF/VTvaf17-FMOD регистрационный № ВКПМ B-13199, INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43082.
Пример 20.
Способ масштабирования получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: SKI, TGFB3, TIMP2, FMOD до промышленного масштаба.
Для подтверждения технологичности получения и возможности производства в промышленных масштабах генотерапевтического ДНК-вектора VTvaf17-SKI (SEQ ID №1), или VTvaf17-TGFB3 (SEQ ID №2), или VTvaf17-TIMP2 (SEQ ID №3), или VTvaf17-FMOD (SEQ ID №4), или проводили масштабную ферментацию штамма Escherichia coli SCS110-AF/VTvaf17-SKI, или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AF/VTvaf17-FMOD, каждый из которых содержит генотерапевтический ДНК-вектор VTvaf17, несущий целевой ген, а именно SKI, или TGFB3, или TIMP2, или FMOD. Каждый штамм Escherichia coli SCS110-AF/VTvaf17-SKI, или Escherichia coli SCS110-AF/VTvaf17-TGFB3, или Escherichia coli SCS110-AF/VTvaf17-TIMP2, или Escherichia coli SCS110-AF/VTvaf17-FMOD получали на базе штамма Escherichia coli SCS110-AF (Cell and Gene Therapy LLC, Великобритания) по примеру 21 путем электропорации компетентных клеток этого штамма генотерапевтическим ДНК-вектором VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD, несущим целевой ген, а именно, SKI, или TGFB3, или TIMP2, или FMOD с последующим высевом трансформированных клеток на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы и отбором отдельных клонов.
Ферментацию полученного штамма Escherichia coli SCS110-AF/VTvaf17-SKI, несущего генотерапевтический ДНК-вектор VTvaf17-SKI проводили в ферментере объемом 10 л с последующим выделением генотерапевтического ДНК-вектора VTvaf17-SKI.
Для ферментации штамма Escherichia coli SCS110-AF/VTvaf17-SKI готовили среду, содержащую на 10 л: 100 г триптон, 50 г дрожжевой экстракт (Becton Dickinson, США), доводили водой до 8800 мл и автоклавировали при 121°С 20 мин, затем добавляли 1200 мл 50% (вес/объем) сахарозы. Далее засевали в колбу затравочную культуру штамма Escherichia coli SCS110-AF/VTvaf17-SKI в объеме 100 мл. Инкубировали в шейкере-инкубаторе 16 ч при 30°С. Переносили затравочную культуру в ферментер Techfors S (Infors НТ, Швейцария), растили до достижения стационарной фазы. Контроль осуществляли измерением оптической плотности культуры при длине волны 600 нм. Клетки осаждали центрифугированием 30 мин при 5000-10000 g. Супернатант удаляли, клеточную массу ресуспендировали в 10% по объему фосфатно-солевого буфера. Повторно центрифугировали 30 мин при 5000-10000 g. Супернатант удаляли, к клеточной массе добавляли раствор 20 мМ TrisCl, 1 мМ ЭДТА, 200 г/л сахароза, рН 8,0 в объеме 1000 мл, тщательно перемешивали до образования гомогенной суспензии. Добавляли раствор яичного лизоцима до конечной концентрации 100 мкг/мл. Инкубировали 20 мин на льду при бережном перемешивании. Далее, добавляли 2500 мл раствора 0,2 М NaOH, 10 г/л додецилсульфат натрия, инкубировали 10 мин на льду при бережном перемешивании, затем добавляли 3500 мл раствора 3М ацетат натрия, 2М уксусная кислота, рН 5-5,5, инкубировали 10 мин на льду при бережном перемешивании. Полученный образец центрифугировали 20-30 мин при 15000 g или более. Раствор аккуратно декантировали, от остатков осадка избавлялись фильтрацией через грубый фильтр (фильтровальную бумагу). Добавляли РНКазу A (Sigma, США) до конечной концентрации 20 нг/мл, инкубировали ночь 16 ч при комнатной температуре. Раствор центрифугировали 20-30 мин при 15000 g, затем фильтровали через мембранный фильтр с порами 0,45 мкм (Millipore, США). Далее проводили ультрафильтрацию через мембрану размером отсечения 100 кДа (Millipore, США) и разбавляли до начального объема буфером 25 мМ TrisCl, рН 7.0. Операцию повторяли три - четыре раза. Наносили раствор, на колонку с 250 мл сорбента DEAE sepharose HP (GE, США), уравновешенную раствором 25 мМ TrisCl, рН 7.0. После нанесения образца колонку промывали тремя объемами этого же раствора, а затем проводили элюцию генотерапевтического ДНК-вектора VTvaf17-SKI линейным градиентом от раствора 25 мМ TrisCl, рН 7.0 до раствора 25 мМ TrisCl, рН 7.0, 1М NaCl в объеме пяти объемов колонки. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм. Хроматографические фракции, содержащие генотерапевтический ДНК-вектор VTvaf17-SKI, объединяли и проводили гель-фильтрацию на сорбенте Superdex 200 (GE, США). Колонку уравновешивали фосфатно-солевым буфером. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм, фракции анализировали электрофорезом в агарозном геле. Фракции, содержащие генотерапевтический ДНК-вектор VTvaf17-SKI объединяли и хранили при -20°С. Для оценки воспроизводимости техпроцесса обозначенные технологические операции повторяли пятикратно. Все технологические операции для штаммов Escherichia coli SCS110-AF/VTvaf17-TGFB3, или Escherichia coli SCS110-AF/VTvaf17-TIMP2, или Escherichia coli SCS110-AF/VTvaf17-FMOD проводили аналогичным образом.
Воспроизводимость техпроцесса и количественные характеристики выхода конечного продукта подтверждают технологичность получения и возможность производства в промышленных масштабах генотерапевтического ДНК-вектора VTvaf17-SKI, или VTvaf17-TGFB3, или VTvaf17-TIMP2, или VTvaf17-FMOD.
Таким образом, задача, поставленная в данном изобретении, а именно: конструирование генотерапевтических ДНК-векторов для повышения уровня экспрессии генов SKI, TGFB3, TIMP2, FMOD, сочетающих в себе следующие свойства:
I) Эффективность повышения экспрессии целевых генов в эукариотических клетках за счет полученных генотерапевтических векторов с минимальным размером;
II) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов и генов антибиотикорезистентности;
III) Технологичность получения и возможность наработки в штаммах в промышленных масштабах;
IV) также задача по конструированию штаммов, несущих эти генотерапевтические ДНК-вектора для производства этих генотерапевтических ДНК-векторов решена, что подтверждается примерами:
для - пример 1, 2, 3, 4, 5; 6; 7; 8; 9; 10; 11; 12; 13; 14; 15; 16; 17; 18
для II - пример 1, 2, 3, 4
для III и п. IV - пример 19, 20.
Промышленная применимость:
Все представленные выше примеры подтверждают промышленную применимость предлагаемого генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген, выбранный из группы генов: ген SKI, ген TGFB3, ген TIMP2, ген FMOD для повышения уровня экспрессии этих целевых генов, штамма Escherichia coli SCS110-AF/VTvaf17-SKI или Escherichia coli SCS110-AF/VTvaf17-TGFB3 или Escherichia coli SCS110-AF/VTvaf17-TIMP2 или Escherichia coli SCS110-AF/VTvaf17-FMOD, несущего генотерапевтический ДНК-вектор, способа получения генотерапевтического ДНК-вектора, способа производства в промышленных масштабах генотерапевтического ДНК-вектора.
Перечень последовательностей олигонуклеотидов
SEQ ID NO: 5 SKI_F
CAGGATCCGCGGGAGCGGCCGGGGGAG
SEQ ID NO: 6 SKI_R
TATAAGCTTACGGCTCCAGCTCCGCAG
SEQ ID NO: 7 SKI _SF
TGGACACCAAGGAAGCCAAAG
SEQ ID NO: 8 SKI _SR
GCTGTTCCTGCAGCTCCTTCAC
SEQ ID NO: 9 TGFB3_F
GGATCCACCATGAAGATGCACTTGCAAAGG
SEQ ID NO: 10 TGFB3_R
TATAAGCTTAGCTACATTTACAAGACTTCACCA
SEQ ID NO: 11 TGFB3_SF
TGAGCACATTGCCAAACAGC
SEQ ID NO: 12 TGFB3_SR
GAGGCAGATGCTTCAGGGTT
SEQ ID NO: 13 TIMP2_F
CAGGATCCGGCCCCCGCCCGCCCAGC
SEQ ID NO: 14 TIMP2_R
TATAAGCTTATGGGTCCTCGATGTCGAGA
SEQ IDNO: 15 TIMP2_SF
GCAACAGGCGTTTTGCAATG
SEQ IDNO: 16 TIMP2_SR
AGGGCACGATGAAGTCACAG
SEQ ID NO: 17 FMOD_F
GGATCCACCATGCAGTGGACCTCCCTCCT
SEQ ID NO: 18 FMOD_R
TATGAATTCTTAGATCTCGATGAGGCTGGCA
SEQ ID NO: 19 FMOD_SF
CCTACACCTACGGCTCTCCA
SEQ ID NO: 20 FMOD_SR
CATCCGGGTCAGGTTGTTGT
Перечень сокращений
VTvaf17 - вектор генотерапевтический, не содержащий последовательностей вирусных геномов и маркеров антибиотикорезистентности (vector therapeutic virus-antibiotic-free)
ДНК - дезоксирибонуклеиновая кислота
кДНК - комплементарная дезоксирибонуклеиновая кислота
РНК - рибонуклеиновая кислота
мРНК - матричная рибонуклеиновая кислота
п.н. - пар нуклеотидов
ПЦР - полимеразная цепная реакция
мл - миллилитр, мкл - микролитр
куб. мм - кубический миллиметр
л - литр
мкг - микрограмм
мг - миллиграмм
г - грамм
мкМ - микромоль
мМ - миллимоль
мин - минута
сек - секунда
об/мин - обороты в минуту
нм - нанометр
см - сантиметр
мВт - милливатт
о.е. ф-относительная единица флуоресценции
РВС - фосфатно-солевой буфер
Список литературы
1. Молекулярная биология, 2011, том 45, №1, с. 44-55
2. Заявка CN 101818171 A
3. Заявка CN 102448983 A
4. Заявка JP 2018512876 А
5. Заявка WO 1996039196 А1
6. Патент US 9550998 B2
7.  Szemraj J, Czarny Р,
Szemraj J, Czarny Р,  Role of MMP-2, MMP-7, MMP-9 and TIMP-2 in the development of recurrent depressive disorder. J Affect Disord. 2016 Nov 15; 205:119-129.
Role of MMP-2, MMP-7, MMP-9 and TIMP-2 in the development of recurrent depressive disorder. J Affect Disord. 2016 Nov 15; 205:119-129.
8. Chen J, et al. Docosahexaenoic acid (DHA) ameliorates paraquat-induced pulmonary fibrosis in rats possibly through up-regulation of SMAD 7 and SnoN. Food Chem. Toxicol. 2013; 57:330-337.
9. Dohi, Т., et al., Tissue Inhibitor of Metalloproteinase-2 Suppresses Collagen Synthesis in Cultured Keloid Fibroblasts. Plast Reconstr Surg Glob Open, 2015. 3(9): p. e520.
10. Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products, http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf
11. Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal Products EMA/CAT/80183/2014
12. Hornstein BD, Roman D,  Engevik MA, Zechiedrich L. Effects of Circular DNA Length on Transfection Efficiency by Electroporation into HeLa Cells.
Engevik MA, Zechiedrich L. Effects of Circular DNA Length on Transfection Efficiency by Electroporation into HeLa Cells.  ed. PLoS ONE. 2016; 11(12):e0167537. doi:10.1371/journal.pone.0167537.
ed. PLoS ONE. 2016; 11(12):e0167537. doi:10.1371/journal.pone.0167537.
13. Jan AT, Lee EJ, Choi I. Fibromodulin: A regulatory molecule maintaining cellular architecture for normal cellular function. Int J Biochem Cell Biol. 2016 Nov; 80:66-70.
14. Kim HJ, Cho YR, Kim SH, Seo DW. TIMP-2-derived 18-mer peptide inhibits endothelial cell proliferation and migration through cAMP/PKA-dependent mechanism. Cancer Lett. 2014 Feb 28; 343(2):210-6.
15. Komai T, Okamura T, Inoue M, Yamamoto K, Fujio K. Reevaluation of Pluripotent Cytokine TGF-β3 in Immunity. Int J Mol Sci. 2018 Aug 1; 19(8).
16. Kurzawski M, Kaczmarek M,  Malinowski D, Kazienko A, Kurzawa R,
Malinowski D, Kazienko A, Kurzawa R,  MMP2, MMP9 and TIMP2 polymorphisms affect sperm parameters but not fertility in Polish males. Andrologia. 2017 Jun; 49(5).
MMP2, MMP9 and TIMP2 polymorphisms affect sperm parameters but not fertility in Polish males. Andrologia. 2017 Jun; 49(5).
17. Li L, Petrovsky N. Molecular mechanisms for enhanced DNA vaccine immunogenicity. Expert Rev Vaccines. 2016; 15(3):313-29
18. Li M, Qiu L, Hu W, Deng X, Xu H, Cao Y, Xiao Z, Peng L, Johnson S, Alexey L, Kingston PA, Li Q, Zhang Y. Genetically-modified bone mesenchymal stem cells with TGF-β(3) improve wound healing and reduce scar tissue formation in a rabbit model. Exp Cell Res. 2018 Jun 1; 367(1):24-29.
19. Li X, et al. The downregulation of SnoN expression in human renal proximal tubule epithelial cells under high-glucose conditions is mediated by an increase in Smurf2 expression through TGF-β1 signaling. Int. J. Mol. Med. 2016; 37:415-422.
20. Li, P., et al., SKI, a modulator of wound healing and scar formation in the rat skin and rabbit ear. J Pathol, 2011. 223(5): p. 659-71.
21. Liu HH, Kenning MS, Jobling AI, McBrien NA, Gentle A. Reduced Scleral TIMP-2 Expression Is Associated With Myopia Development: TIMP-2 Supplementation Stabilizes Scleral Biomarkers of Myopia and Limits Myopia Development. Invest Ophthalmol Vis Sci. 2017 Apr 1; 58(4): 1971-1981.
22. Liu X, et al. Expression and possible mechanism of c-ski, a novel tissue repair-related gene during normal and radiation-impaired wound healing. Wound Repair Regen. 2006; 14:162-171.
23. Mairhofer J, Grabherr R. Rational vector design for efficient non-viral gene delivery: challenges facing the use of plasmid DNA. Mol Biotechnol. 2008. 39(2):97-104
24. McCollum PT, Bush JA, James G, Mason T, O'Kane S, McCollum C, Krievins D, Shiralkar S, Ferguson MW. Randomized phase II clinical trial of avotermin versus placebo for scar improvement. Br J Surg. 2011 Jul; 98(7):925-34.
25. Peng Y, et al. Comparative evaluation of the wound-healing potency of recombinant bFGF and ski gene therapy in rats. Growth Factors. 2016; 34:119-127.
26. Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies
27. Ritter M, et al. A. Inhibition of retinoic acid receptor signaling by Ski in acute myeloid leukemia. Leukemia. 2006; 20:437-443.
28. Shinagawa T, et al. Increased susceptibility to tumorigenesis of ski-deficient heterozygous mice. Oncogene. 2001; 20:8100-8108.
29. Soo, С., et al., Differential expression of fibromodulin, a transforming growth factor-beta modulator, in fetal skin development and scarless repair. Am J Pathol, 2000. 157(2): p. 423-33
30. Stoff, A., et al., Effect of adenoviral mediated overexpression of fibromodulin on human dermal fibroblasts and scar formation in full-thickness incisional wounds. J Mol Med (Berl), 2007. 85(5): p. 481-96
31. Tan R, et al. Downregulation of SnoN expression in obstructive nephropathy is mediated by an enhanced ubiquitin-dependent degradation. J. Am. Soc. Nephrol. 2006; 17:2781-2791.
32. Tang H, et al. MAD2B-mediated SnoN downregulation is implicated in fibroblast activation and tubulointerstitial fibrosis. Am. J. Physiol. Ren. Physiol. 2016;
33. Tecalco-Cruz, A.C, 
 Rosales-Alvarez, R.E., & Macias-Silva, M. (2018). Transcriptional cofactors Ski and SnoN are major regulators of the TGF-β/Smad signaling pathway in health and disease. Signal Transduction and Targeted Therapy, 3, 15.
Rosales-Alvarez, R.E., & Macias-Silva, M. (2018). Transcriptional cofactors Ski and SnoN are major regulators of the TGF-β/Smad signaling pathway in health and disease. Signal Transduction and Targeted Therapy, 3, 15.
34. Villanacci V, et al. Ski/SnoN expression in the sequence metaplasia-dysplasia-adenocarcinoma of Barrett's esophagus. Hum. Pathol. 2008; 39:403-409.
35. Wang X, Li Y, Han R, He C, Wang G, Wang J, Zheng J, Pei M, Wei L. Demineralized bone matrix combined bone marrow mesenchymal stem cells, bone morphogenetic protein-2 and transforming growth factor-β3 gene promoted pig cartilage defect repair. PLoS One. 2014 Dec 29; 9(12):e116061. 2015; 10(5):e0125948.
36. Xu P, Guo W, Jin T, Wang J, Fan D, Hao Z, Jing S, Han C, Du J, Jiang D, Wen S, Wang J. TIMP-2 SNPs rs7342880 and rs4789936 are linked to risk of knee osteoarthritis in the Chinese Han Population. Oncotarget. 2017 Jan 3; 8(1):1166-1176.
37. Yang H, et al. Ski prevents TGF-β-induced EMT and cell invasion by repressing SMAD-dependent signaling in non-small cell lung cancer. Oncol. Rep. 2015; 34:87-94.
38. Yang J, Zhang X, Li Y, Liu Y. Downregulation of SMAD transcriptional corepressors SnoN and Ski in the fibrotic kidney: an amplification mechanism for TGF-β1 signaling. J. Am. Soc. Nephrol. 2003; 14:3167-3177.
39. Yang SW, Cody JJ, Rivera AA, Waehler R, Wang M, Kimball KJ, Alvarez RA, Siegal GP, Douglas JT, Ponnazhagan S. Conditionally replicating adenovirus expressing TIMP2 for ovarian cancer therapy. Clin Cancer Res. 2011 Feb 1; 17(3):538-49
40. Yue B, Lin Y, Ma X, Xiang H, Qiu C, Zhang J, Li L, Chen B. Survivin-TGFB3-TIMP1 Gene Therapy Via Lentivirus Vector Slows the Course of Intervertebral Disc Degeneration in an In Vivo Rabbit Model. Spine (Phila Pa 1976). 2016 Jun; 41(11):926-34.
41. Zhu X, et al. 576 kb deletion in 1p36.33-p36.32 containing SKI is associated with limb malformation, congenital heart disease and epilepsy. Gene. 2013; 528:352-355.








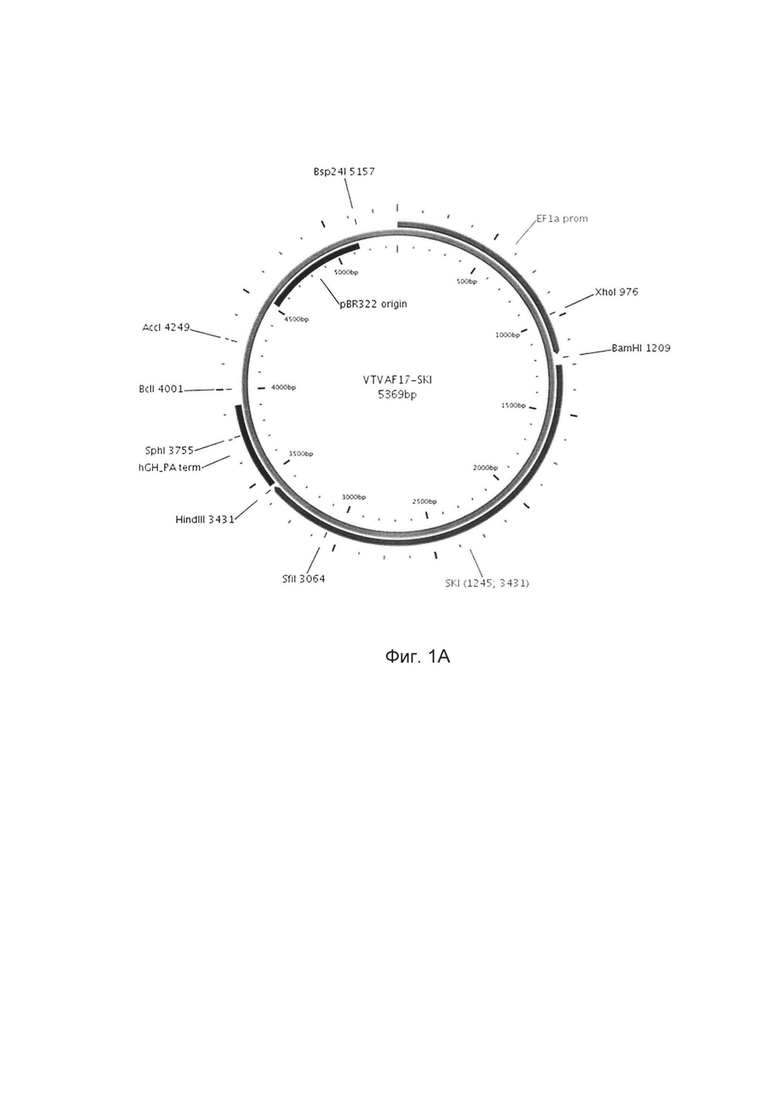
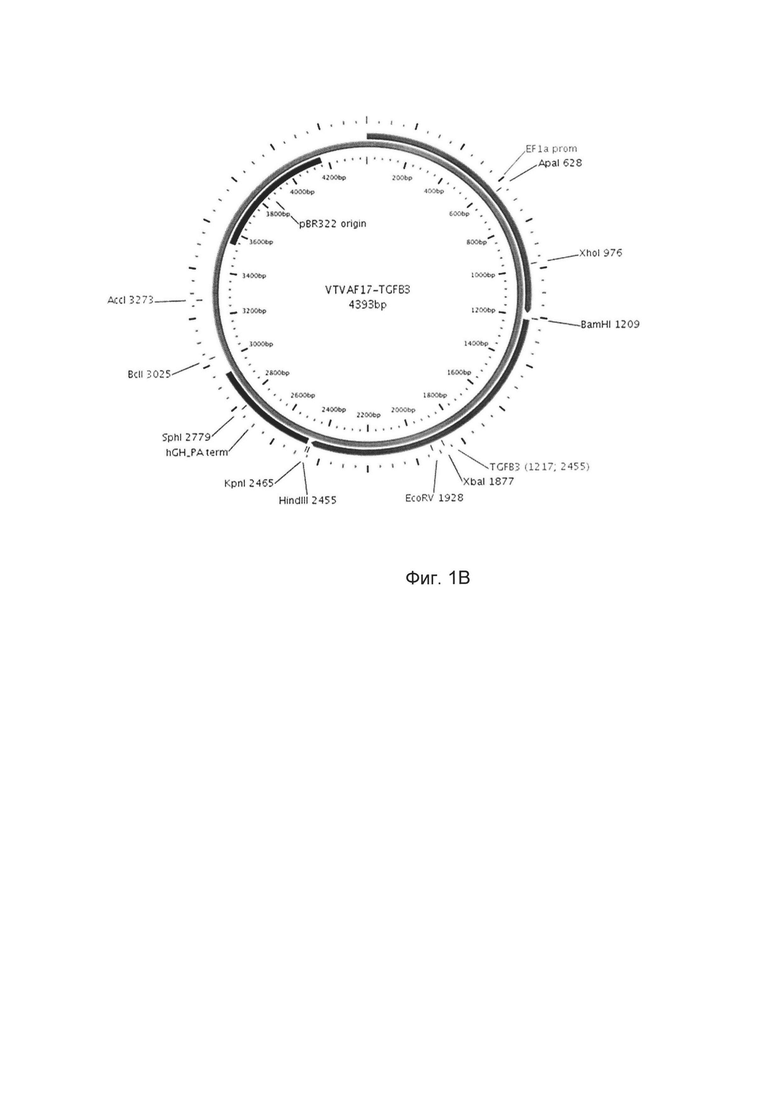

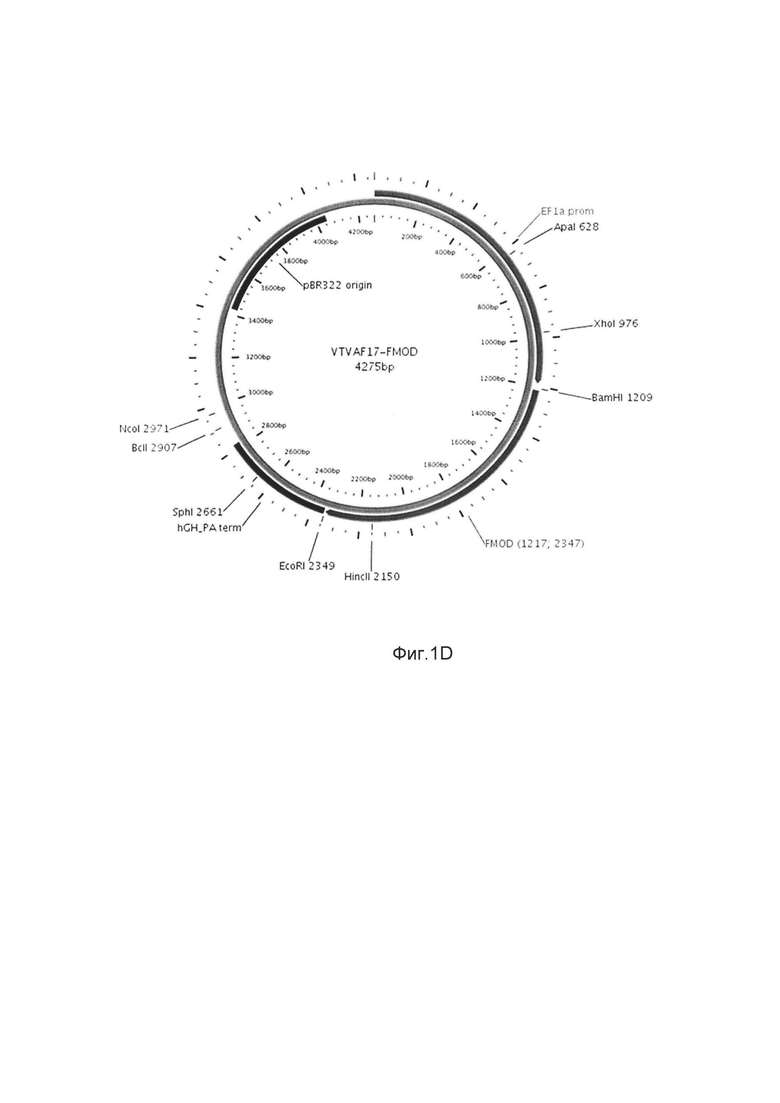
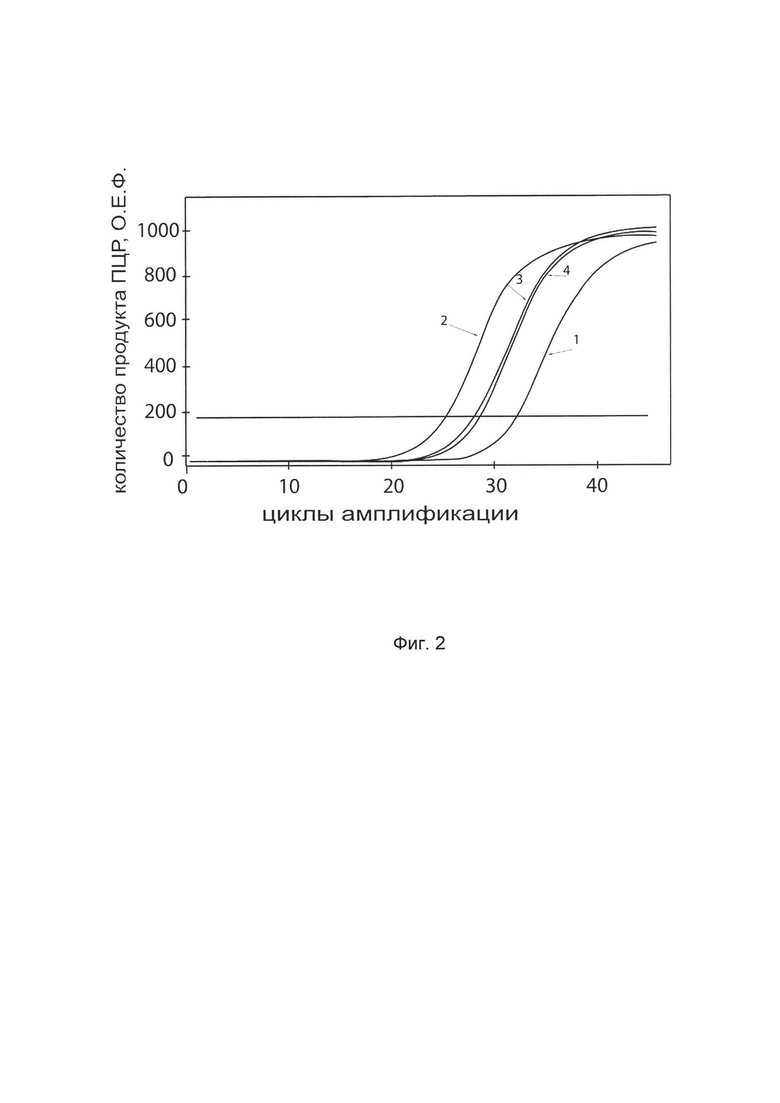
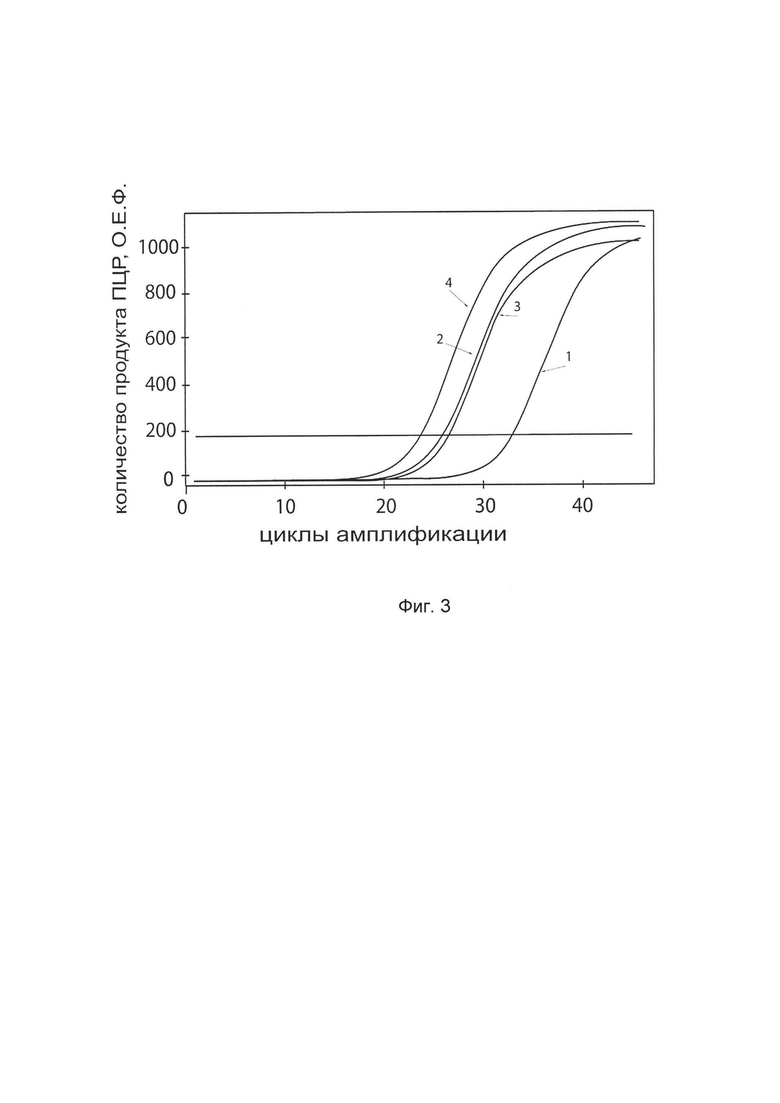
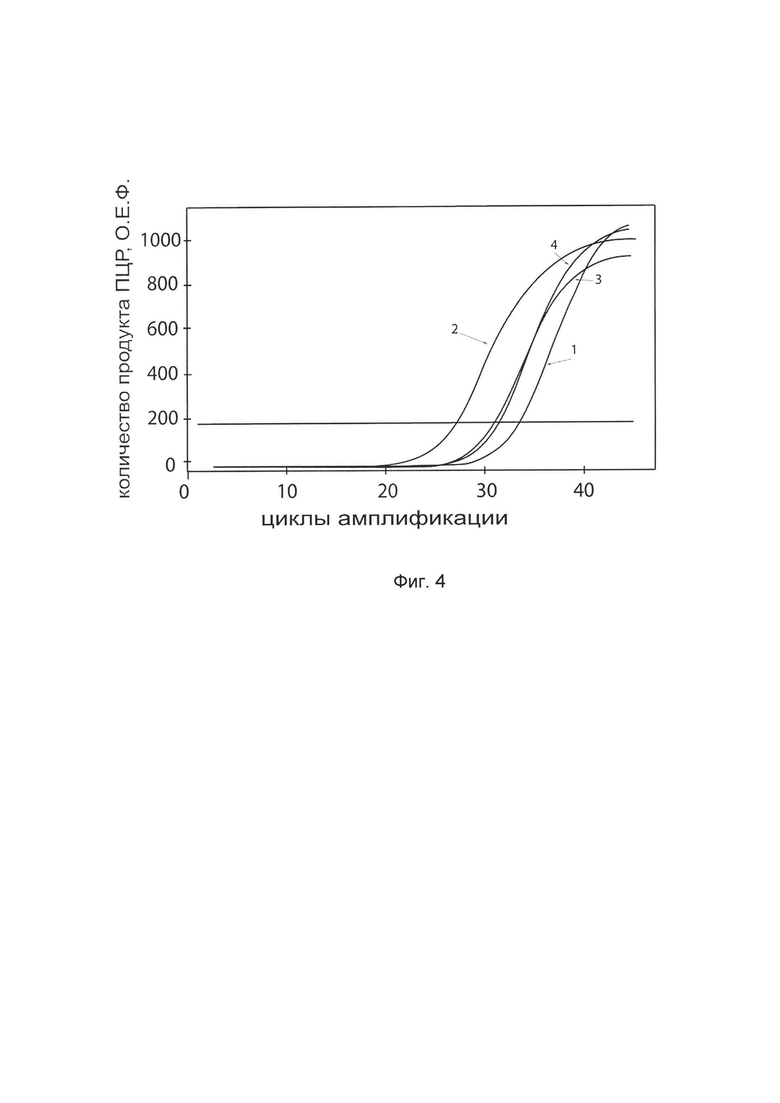
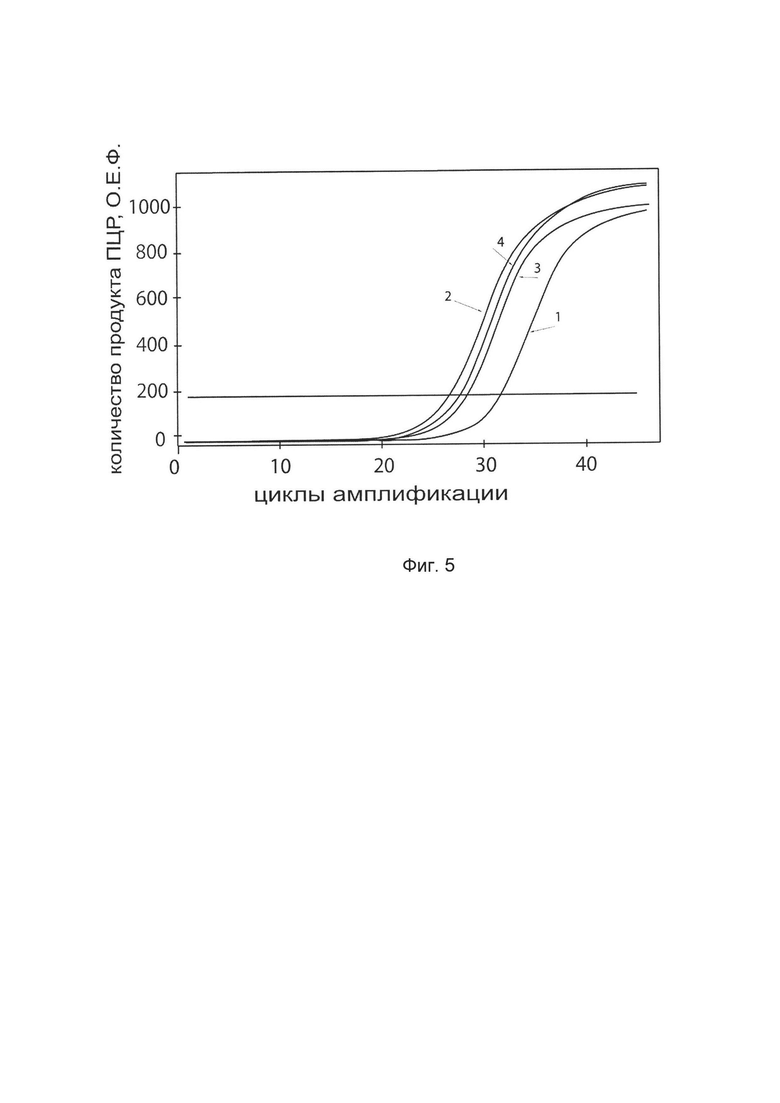

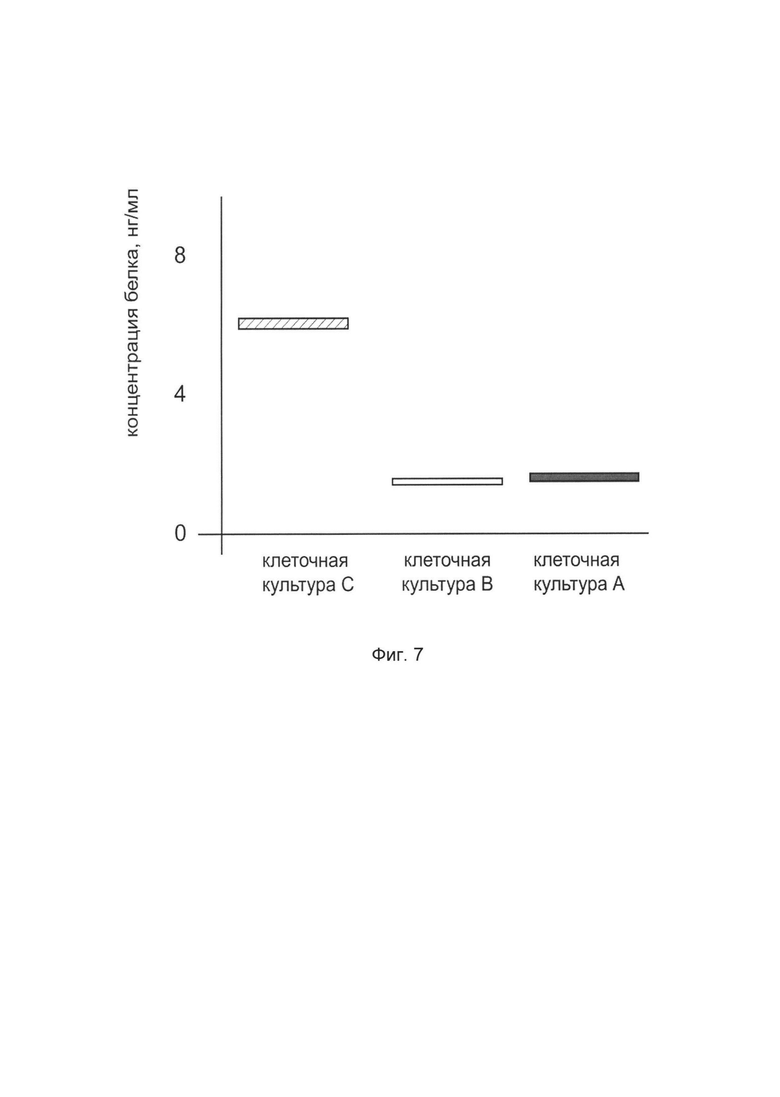
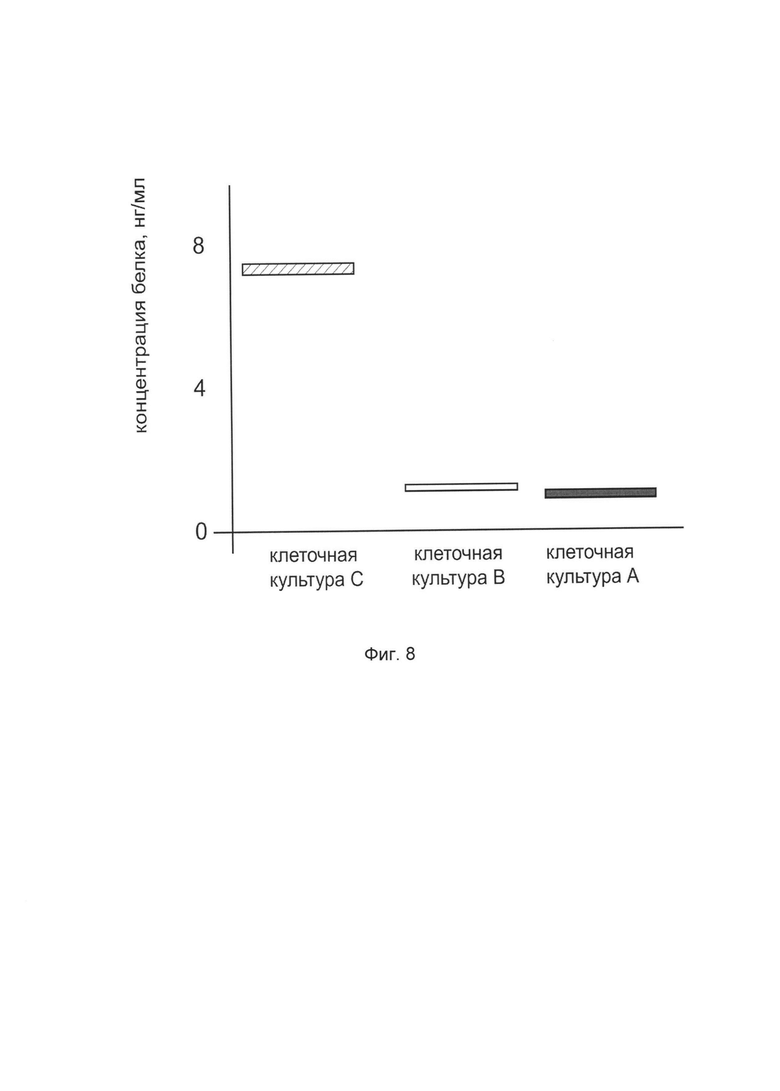
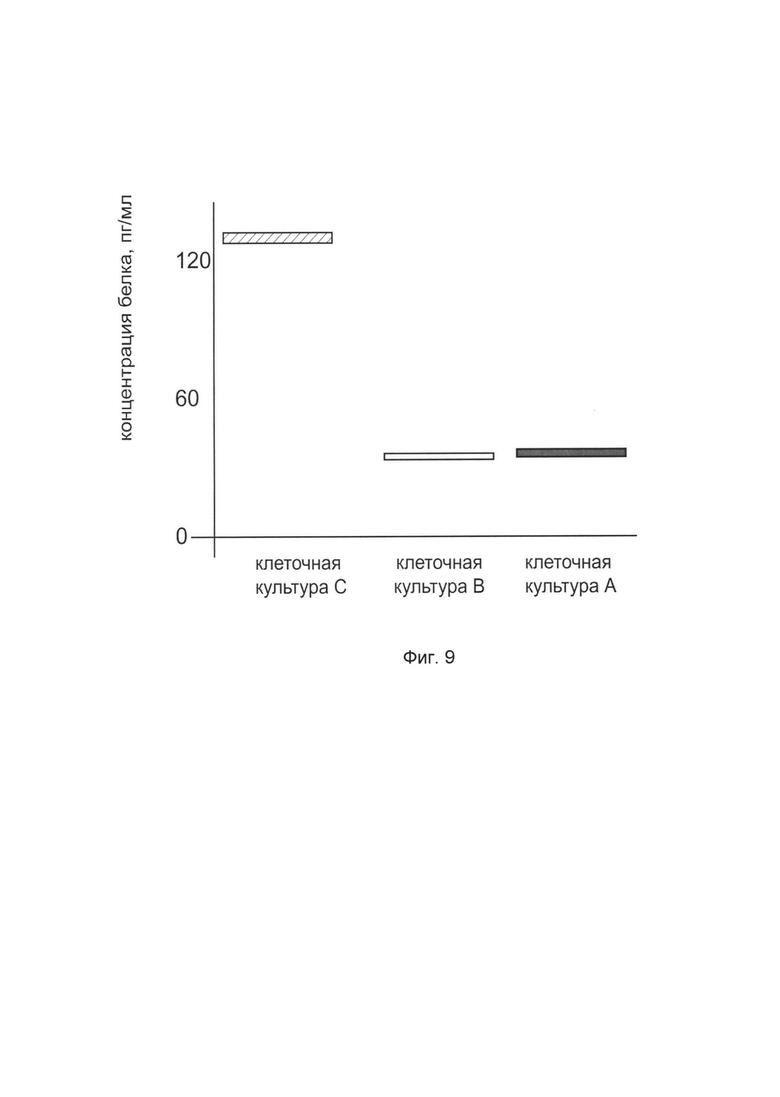
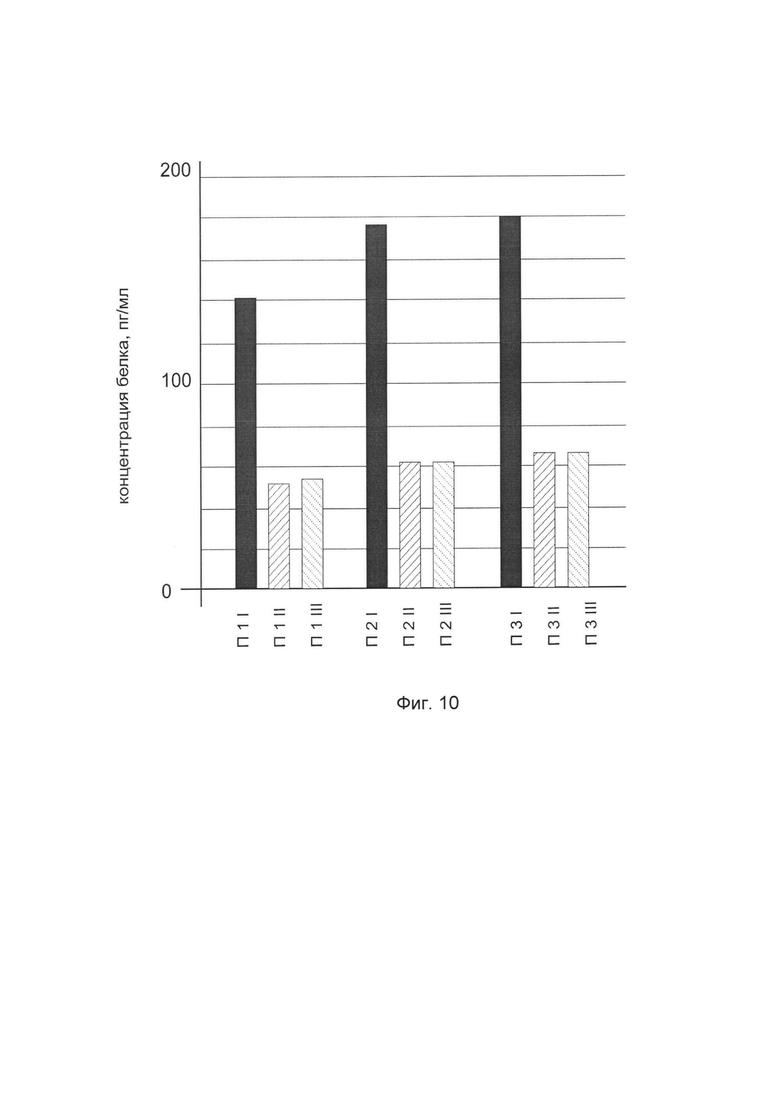
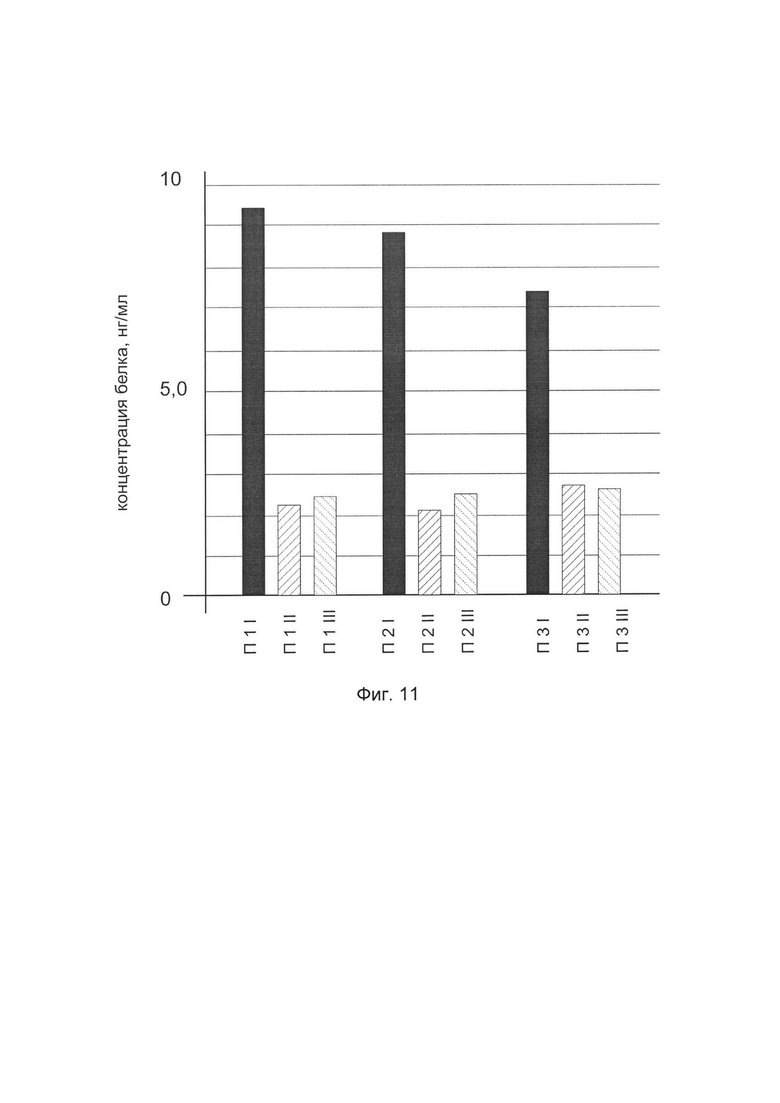
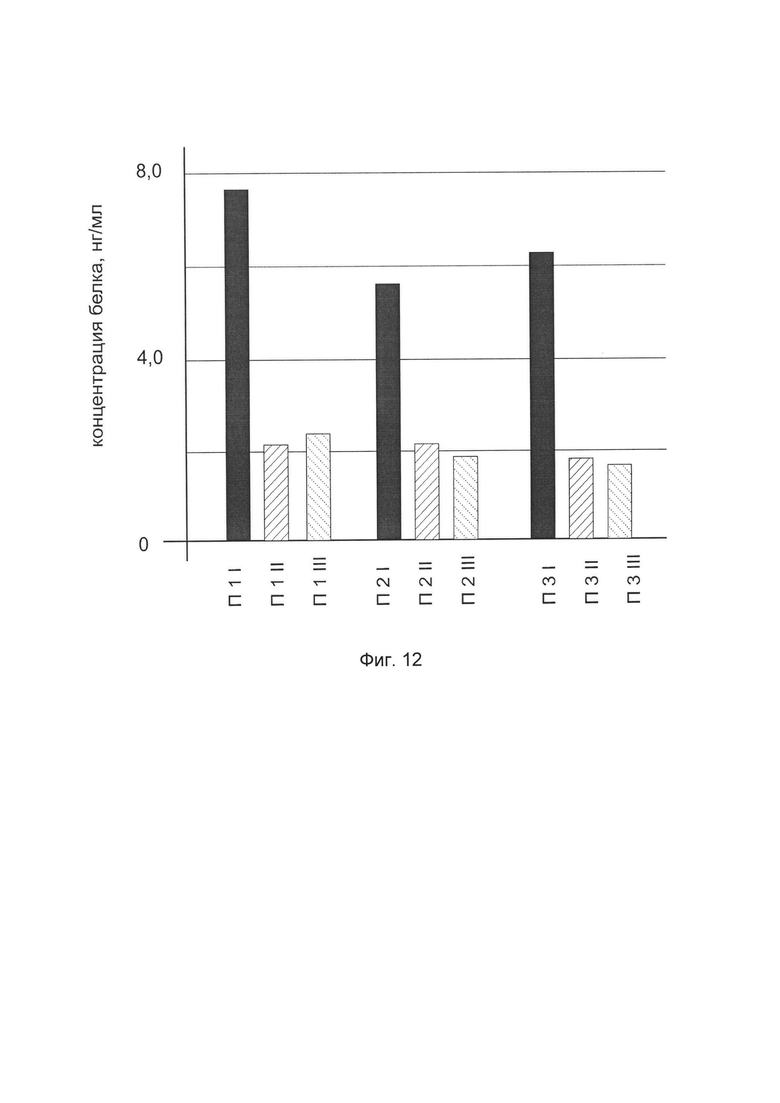
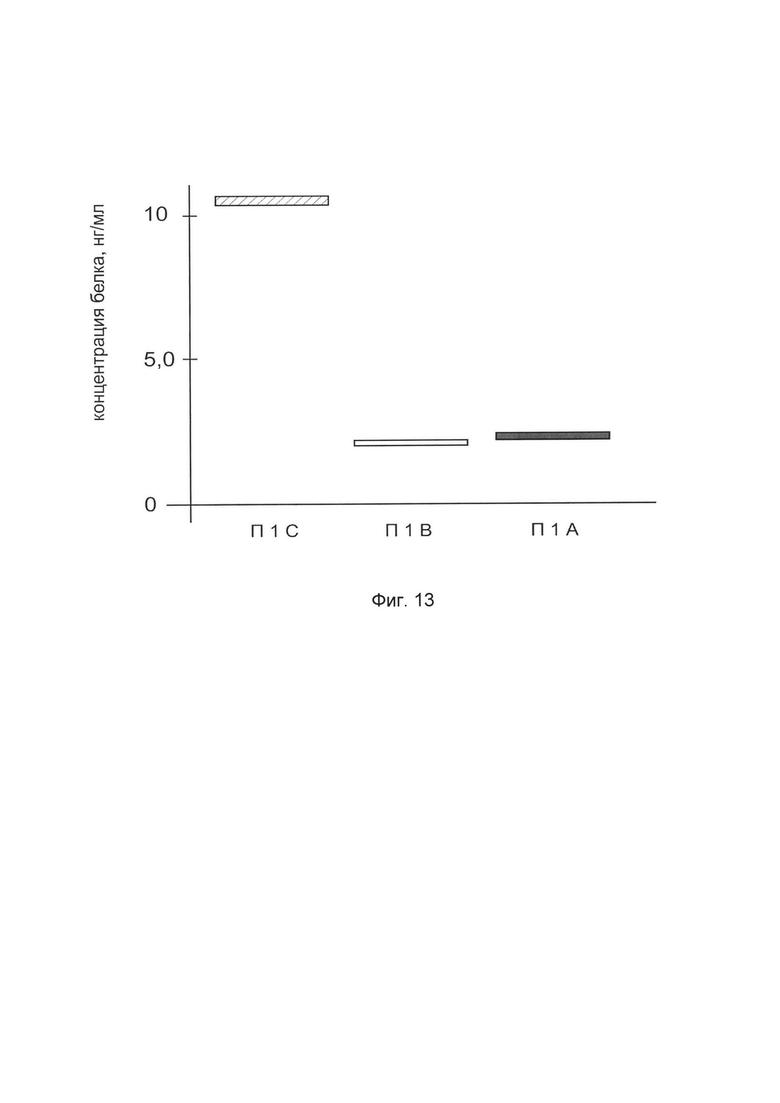
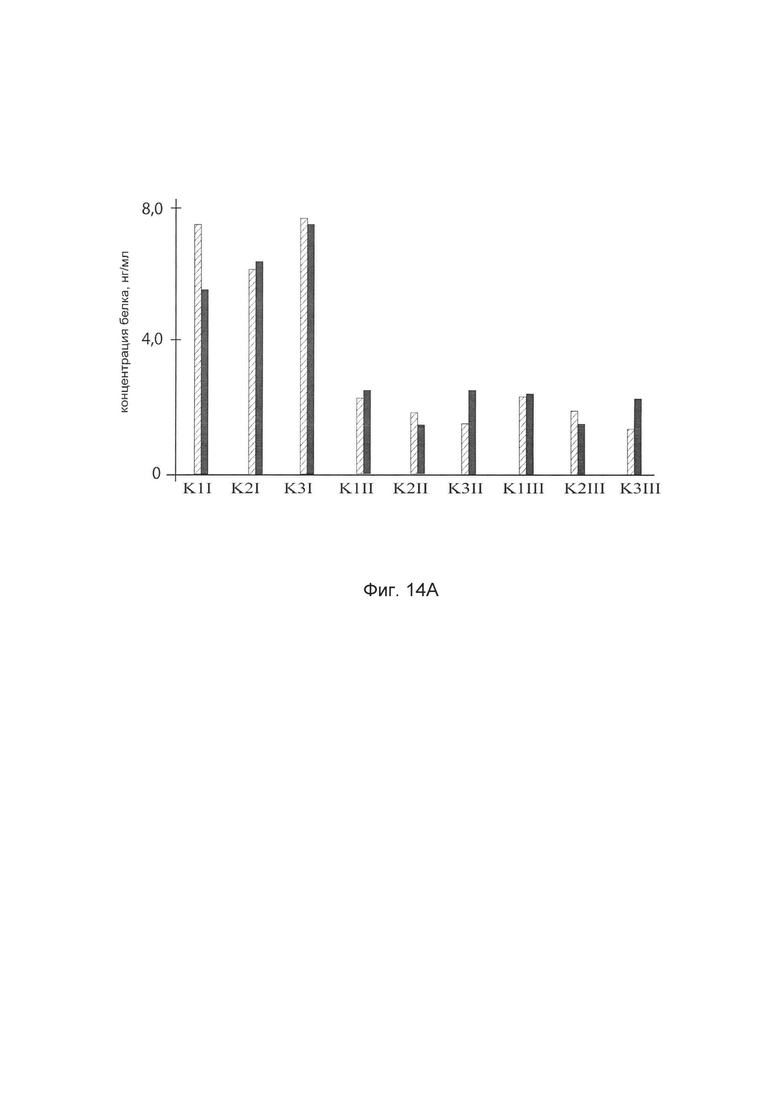
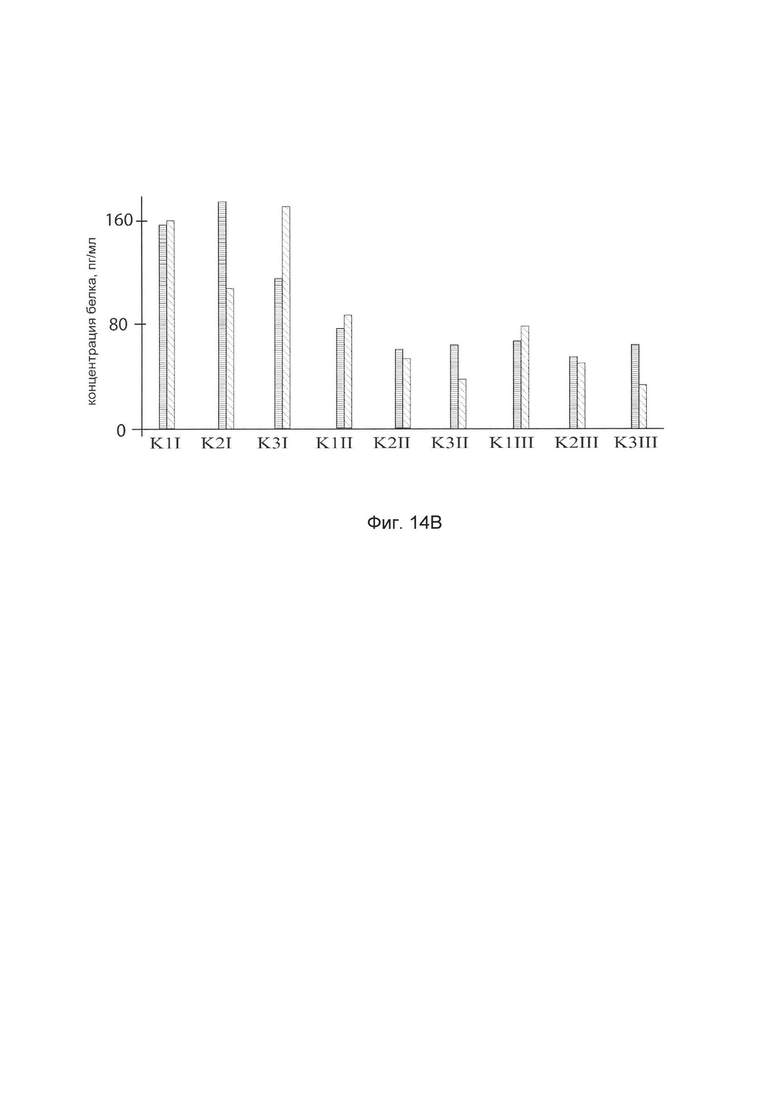
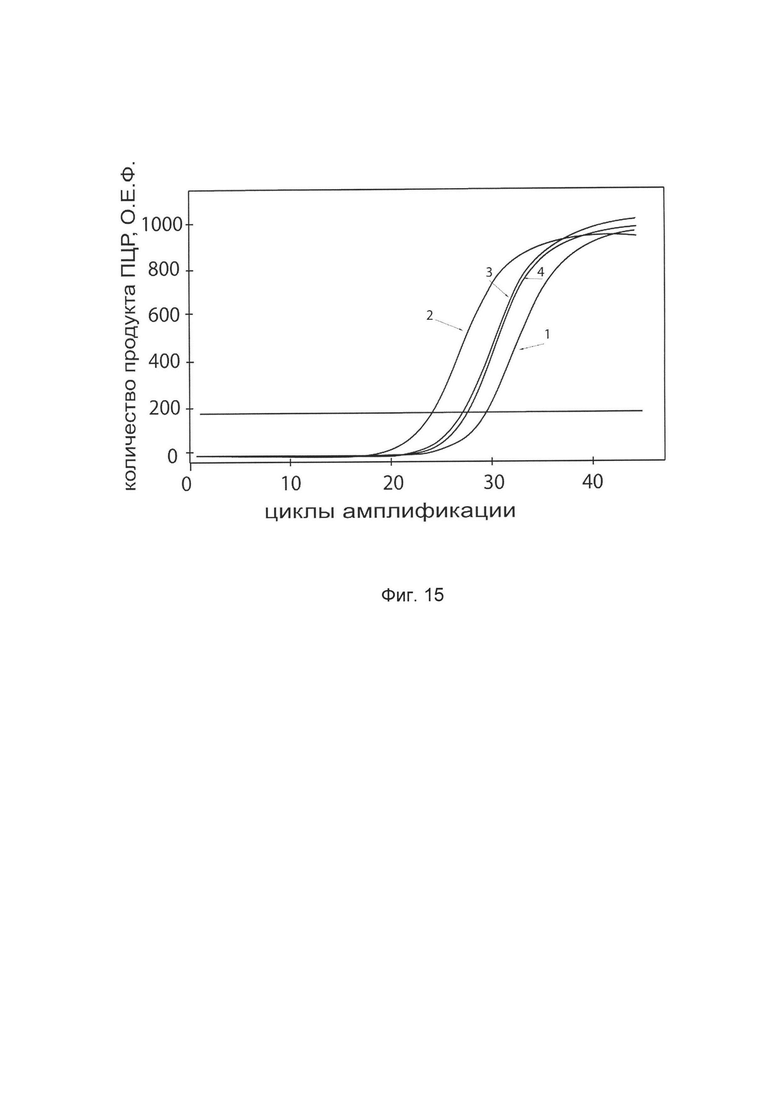
Изобретение относится к генной инженерии. Описан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена SKI в организме человека и животных, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена SKI, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-SKI, с нуклеотидной последовательностью SEQ ID №1. Также описан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена TGFB3, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена TGFB3, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17- TGFB3, с нуклеотидной последовательностью SEQ ID №2. Кроме того, описан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена TIMP2, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена TIMP2, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-TIMP2, с нуклеотидной последовательностью SEQ ID №3. Кроме того, описан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена FMOD, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена FMOD, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-FMOD, с нуклеотидной последовательностью SEQ ID №. 4. Описаны способы получения описанных векторов. Кроме того, описано применение указанных векторов для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена SKI, или TGFB3, или TIMP2, или FMOD. Представлены штаммы: EscherichiacoliSCS110-AF/VTvaf17-SKI, штамм EscherichiacoliSCS110-AF/VTvaf17-TGFB3, штамм EscherichiacoliSCS110-AF/VTvaf17-TIMP2, Штамм EscherichiacoliSCSI 10-AF/VTvaf17-FMOD, и способ их получения. Изобретение расширяет арсенал средств для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани. 12 н. и 2 з. п. ф-лы, 15 ил., 20 пр.
1. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена SKI в организме человека и животных, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена SKI, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-SKI, с нуклеотидной последовательностью SEQ ID №1.
2. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена TGFB3, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена TGFB3, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17- TGFB3, с нуклеотидной последовательностью SEQ ID №2.
3. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена TIMP2, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена TIMP2, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-TIMP2, с нуклеотидной последовательностью SEQ ID №3.
4. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена FMOD, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена FMOD, клонированную в генотерапевтический ДНК-вектор VTvaf17, с получением генотерапевтического ДНК-вектора VTvaf17-FMOD, с нуклеотидной последовательностью SEQ ID №. 4.
5. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген SKI, или TGFB3, или TIMP2, или FMOD по пп. 1-3 или 4, отличающийся тем, что каждый из созданных генотерапевтических ДНК-векторов: VTvaf17- SKI, или VTvaf17- TGFB3, или VTvaf17- TIMP2, или VTvaf17- FMOD по пп. 1-3 или 4, за счет ограниченного размера векторной части VTvaf 17, не превышающей 3200 п.н., обладает способностью эффективно проникать в клетки человека и животных и экспрессировать клонированный в него целевой ген SKI, или TGFB3, или TIMP2, или FMOD.
6. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора VTvaf17, несущий целевой ген SKI, или TGFB3, или TIMP2, или FMOD по пп. 1-3 или 4, отличающийся тем, что в составе каждого из созданных генотерапевтических ДНК-векторов: VTvaf17- SKI, или VTvaf17- TGFB3, или VTvaf17-TIMP2, или VTvaf17- FMOD по пп. 1-3 или 4, в качестве структурных элементов используют нуклеотидные последовательности, которые не являются генами антибиотикорезистентности, вирусными генами и регуляторными элементами вирусных геномов, обеспечивая возможность его безопасного применения для генетической терапии человека и животных.
7. Способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SKI, или TGFB3, или TIMP2, или FMOD по пп. 1-3 или 4, заключающийся в том, что каждый из генотерапевтических ДНК-векторов: VTvaf17- SKI, или VTvaf17- TGFB3, или VTvaf17- TIMP2, или VTvaf17- FMOD получают следующим образом: кодирующую часть целевого гена SKI, или TGFB3, или TIMP2, или FMOD по пп. 1-3 или 4, клонируют в генотерапевтический ДНК-вектор VTvaf17 и получают генотерапевтический ДНК-вектор VTvaf17- SKI, SEQ ID №1, или VTvaf17- TGFB3, SEQ ID №2, или VTvaf17- TIMP2, SEQ ID №3, или VTvaf17- FMOD, SEQ ID №4, соответственно, при этом кодирующую часть целевого гена SKI, или TGFB3, или TIMP2, или FMOD получают путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции и ПЦР-амплификации с использованием созданных олигонуклеотидов и расщеплением продукта амплификации соответствующими эндонуклеазами рестрикции, причем клонирование в генотерапевтический ДНК-вектор VTvaf17 проводят по сайтам рестрикции NheI и HindIII, причем селекцию проводят без антибиотиков,
при этом при получении генотерапевтического ДНК-вектора VTvaf17-SKI, SEQ ID №1 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
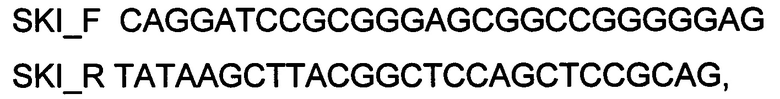
а расщепление продукта амплификации и клонирование кодирующей части гена SKI в генотерапевтический ДНК-вектор VTvaf17 проводят с использованием эндонуклеаз рестрикции BamHI и HindIII,
причем при получении генотерапевтического ДНК-вектора VTvaf17-TGFB3, SEQID №2 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды

а расщепление продукта амплификации и клонирование кодирующей части гена TGFB3 в генотерапевтический ДНК-вектор VTvaf17 проводят с использованием эндонуклеаз рестрикции BamHI и HindIII,
причем при получении генотерапевтического ДНК-вектора VTvaf17-TIMP2, SEQID №3 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
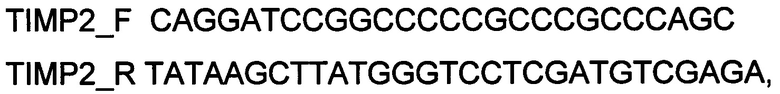
а расщепление продукта амплификации и клонирование кодирующей части гена TIMP2 в генотерапевтический ДНК-вектор VTvaf17 проводят с использованием эндонуклеаз рестрикции BamHI и HindIII,
причем при получении генотерапевтического ДНК-вектора VTvaf17-FMOD, SEQID №4 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
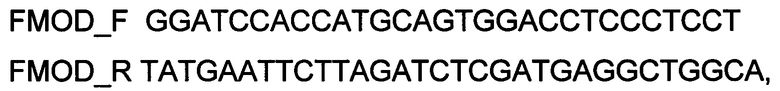
а расщепление продукта амплификации и клонирование кодирующей части гена FMOD в генотерапевтический ДНК-вектор VTvaf17 проводят с использованием эндонуклеаз рестрикции BamHI и EcoRI.
8. Способ применения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SKI, или TGFB3, или TIMP2, или FMOD по пп. 1-3 или 4, для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена SKI, или TGFB3, или TIMP2, или FMOD, заключающийся в трансфекции выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора VTvaf17, или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, а именно VTvaf17- SKI, или VTvaf17- TGFB3, или VTvaf17- TIMP2, или VTvaf17- FMOD, клеток органов и тканей пациента или животного, и/или во введении в органы и ткани пациента или животного аутологичных клеток этого пациента или животного, трансфицированных выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора VTvaf17 или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора VTvaf17 и/или во введении в органы и ткани пациента или животного выбранного генотерапевтического ДНК-вектора, несущего целевой ген на основе генотерапевтического ДНК-вектора VTvaf17 или нескольких выбранных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора VTvaf17, или в сочетании обозначенных способов.
9. Способ получения штамма для производства генотерапевтического ДНК-вектора по пп. 1-3 или 4 для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена SKI, или TGFB3, или TIMP2, или FMOD, заключающийся в получении электрокомпетентных клеток штамма EscherichiacoliSCS110-AF с последующим проведением электропорации этих клеток генотерапевтическим ДНК-вектором VTvaf17SKI, или ДНК-вектором VTvaf17-TGFB3, или ДНК-вектором VTvaf17- TIMP2, или ДНК-вектором VTvaf17- FMOD, после чего клетки высеивают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола с получением в результате штамма Escherichia coli SCS110-AF/VTvaf17-SKI или штамма Escherichia coli SCS110-AF/VTvaf17-TGFB3, или штамма Escherichia coli SCS110-AF/VTvaf17-TIMP2, или штамма Escherichia coli SCS110-AFA/Tvaf17-FMOD.
10. Штамм EscherichiacoliSCS110-AF/VTvaf17-SKI, полученный способом по п. 9, несущий генотерапевтический ДНК-вектор VTvaf17-SKI, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани.
11. Штамм EscherichiacoliSCS110-AF/VTvaf17-TGFB3, полученный способом по п. 9, несущий генотерапевтический ДНК-вектор VTvaf17- TGFB3, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани.
12. Штамм EscherichiacoliSCS110-AF/VTvaf17-TIMP2, полученный способом по п. 9, несущий генотерапевтический ДНК-вектор VTvaf17- TIMP2, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани.
13. Штамм EscherichiacoliSCSI 10-AF/VTvaf17-FMOD, полученный способом по п. 9, несущий генотерапевтический ДНК-вектор VTvaf17- FMOD, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани.
14. Способ производства в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора VTvaf17, несущего целевой ген SKI, TGFB3, TIMP2, FMOD по пп. 1-3 или 4, для лечения заболеваний, характеризующихся развитием фиброза тканей, формированием рубцов, повреждением соединительной ткани и для ускорения заживления ран, реэпителизации, для увеличения образования грануляционной ткани и ингибирования образования рубцовой ткани, путем повышения уровня экспрессии целевого гена SKI, или TGFB3, или TIMP2, или FMOD, заключающийся в том, что генотерапевтический ДНК-вектор VTvaf17-SKI, или генотерапевтический ДНК-вектор VTvaf17-TGFB3, или генотерапевтический ДНК-вектор VTvaf17-TIMP2, или генотерапевтический ДНК-вектор VTvaf17-FMOD, получают путем того, что засевают в колбу с приготовленной средой затравочную культуру, выбранную из штамма EscherichiacoliSCS110-AF/VTvaf17-SKI, или штамма EscherichiacoliSCS110-AF/VTvaf17-TGFB3, или штамма EscherichiacoliSCS110-AF/VTvaf17-TIMP2, или штамма EscherichiacoliSCS110-AF/VTvaf17-FMOD, затем инкубируют затравочную среду в шейкере-инкубаторе и переносят ее в промышленный ферментер, после чего растят до достижения стационарной фазы, затем выделяют фракцию, содержащую целевой ДНК-продукт, многостадийно фильтруют и очищают хроматографическими методами.
| Средство для лечения состояний человеческого организма, связанных с уменьшением уровня экспрессии гена Р4НА1 и/или уменьшением количества белка пролил 4-гидроксилазы альфа 1 на основе генно-терапевтических субстанций с геном Р4НА1, способ получения и использования | 2017 |
|
RU2658428C9 |
| RU 2015140941 A, 30.03.2017 | |||
| RU 2011110736 A, 27.09.2012. | |||

