Изобретение относится к способам исследования бактериальных клеток методами полярографии, в частности к способу определения дыхательного контроля (ДК) популяции бактерий, и может быть использовано для оценки качества вакцин, их полуфабрикатов и других бактерийных препаратов на основе вегетативных форм микробов по степени энергозависимой регуляции дыхательной активности бактериальных культур.
Известен способ определения дыхательного контроля у пробы митохондрий, выделенных из клеток эукариот (способ определения ДК по Чансу-Уильямсу), состоящий в одномоментном добавлении сукцината в среду инкубации деэнергизованных митохондрий, содержащей ортофосфат и АДФ, либо в одномоментном добавлении АДФ в среду инкубации энергизованных митохондрий, окисляющих сукцинат в присутствии ортофосфата, АДФ и измерении отношения увеличившейся при этом добавлении скорости потребления кислорода митохондриями (быстрая фаза активного дыхания - V3) к уменьшившейся через несколько минут скорости потребления кислорода (медленная фаза контролируемого дыхания - V4), то есть ДК=V3/V4 [Chance В., Williams G.R. The respiratory chain and oxidative phosphorylation / Adv. Enzymol., 1956. - No 17. - P.65-134; Патологическая физиология. Учебник для мед. вузов /Под ред. А.Д.Адо, М.А.Адо, В.И.Пыцкого и др. - М., 2000, Триада-Х. - С.23-25]. По величине ДК судят об уровне сохранения физиологической работоспособности митохондрий, нарушении их биоэнергетических функций как наиболее ранних признаках патологических изменений в клетках эукариот.
Общим с заявленным способом является методический прием формирования in vitro последовательности метаболических состояний изучаемого биологического объекта, а именно:
1) активация дыхательной активности (V3), вызванная окислением одного из метаболитов цикла Кребса (сукцината) при добавлении АДФ, как результат низкоэнергетического сдвига;
2) регулируемое (контролируемое) торможение дыхательной активности (V4), вызванное превращением добавленной АДФ в АТФ в реакции окислительного фосфорилирования при утилизации одного из метаболитов цикла Кребса (сукцината), как результат накопления макроэргического продукта и увеличения мембранного потенциала митохондрий.
Недостатком данного способа является неоднократно экспериментально подтвержденная невозможность моделирования in vitro метаболических состояний популяции бактерий при добавлении к ней экзогенной АДФ и, как следствие, невозможность определения у бактериальной популяции дыхательного контроля по Чансу-Уильямсу, поскольку интактные бактерии непроницаемы для экзогенной АДФ, и они не содержат транслоказы адениннуклеотидов [Лукоянова М.А., Тихонова Г.В. Организация дыхательной цепи и аккумуляция энергии у бактерий. Итоги науки и техники //Сер. Биол. химия. - ВИНИТИ. - М., 1982. - №17. - Р.33-74.]. Создание специальных условий в опытах с интактными бактериями, например применение креатинкиназной фосфатакцепторной системы и ингибитора гликолиза в системе "гексокиназной ловушки" [Голубинский Е.П., Рублев Б.Д., Кирдеев В. К. Окислительное фосфорилирование у чумного микроба / Вопросы мед. химии, - 1971. - No 16(5). - С.512-515; Канчух А.А., Голубинский Е.П., Иванова B.C. Использование гексокиназной и креатинкиназной систем для измерения окислительного фосфорилирования у чумного микроба / Биохимия. - 1973. - №38(2). - С.246-249], а также опыты с изолированными мембранными препаратами бактерий [Burstein С., Tiankova L., Kepes A. Respiratory control in Escherichia coli К 12 / Eur. J.Biochem. - 1979. - No 94. - P.387-392.; Haddock B.A., lones C.W. Bacterial respiration /J. Bacteriol.- 1977. - No 41(1). - P.47-99] позволяли регистрировать лишь незначительную активацию дыхания экзогенным АДФ (V3), но не последующее ауторегулируемое торможение активного дыхания популяции бактерий (V4).
Известен способ определения дыхательного контроля у популяции бактерий по величине стимуляции дыхания покоящихся клеток Escherichia coli разобщителем карбонилцианид-р-трихлорметоксифенилгидразоном [Burstein С., Tiankova L., Kepes A. Respiratory control in Escherichia coli К 12 / Eur. J.Biochem. - 1979. - No 94. - P.387-392].
Общим с заявленным способом является объект исследования - популяция бактерий, а также методический прием полярографической регистрации скорости дыхания популяции бактерий с использованием платинового электрода.
Недостаток способа заключается в использовании разобщителя-протонофора для определения дыхательного контроля, что позволяет оценить лишь влияние величины протонного заряда цитоплазматической мембраны бактерий на регуляцию их дыхательной активности и не учитывает вклад других регуляторов, а именно: АТФ, АДФ, АМФ, НАД· Н, НАД+ и др. Следствием этого является сложность физиологической интерпретации получаемых результатов определения дыхательного контроля, который, например, не выявляется у растущей популяции Escherichia coli. Кроме того, способ требует применения дополнительных приемов истощения бактерий по эндогенным субстратам, для чего проводится предварительная интенсивная аэрация клеточной суспензии при 37° С до 30 минут, что неизбежно влияет на метаболический статус исследуемой популяции и искажает результаты опытов, свидетельством чему является то обстоятельство, что полученные авторами экспериментальные данные статистически не подтверждены, в то время как определяемые этим способом значения дыхательного контроля в разных условиях варьируют от 1,0 до 9,0.
Наиболее близким к заявляемому техническому решению является способ [Tsuchiya Т., Rosen В.Р. Respiratory control in Escherichia coli / FEBS Lett. - 1980. - No.120(1). - Р.128-130], основанный на одномоментном добавлении в инкубационную среду с дышащей популяцией бактерий углеводов - олигосахаридов, являющихся субстратами Δ μ
Общим с заявляемым способом является методический прием полярографического измерения дыхательной активности популяции бактерий с помощью платинового электрода в ячейке объемом 2 см3, заполненной средой инкубации, который включает предварительное инкубирование микробной популяции для получения стабильного уровня эндогенного дыхания клеток, одномоментное введение в среду инкубации субстрата углеводной природы и количественный анализ динамики убыли кислорода из среды инкубации, а также то, что активация дыхания при добавлении углеводов вызвана инициацией окисления эндогенных субстратов в исследуемой популяции бактерий.
К недостаткам этого способа следует отнести применение и качестве одномоментно добавляемых в среду инкубации популяции бактерий неметаболизируемых веществ олигосахаридной природы - субстратов лактозотранспортной системы в заведомо высокой конечной концентрации 10 мМ (10000 нмоль/см3), что не позволяет регистрировать такую форму дыхательного контроля, как ауторегулируемое торможение активного дыхания популяции бактерий. Кроме того, по условиям способа бактериальные клетки дважды отмываются в буферном растворе и подвергаются дополнительному стрессовому воздействию интенсивного аэрирования и голодания, что, безусловно, отрицательно влияет на их исходный физиологический и энергетический статус. Изложенные недостатки ограничивают области применения способа. Сравнительный анализ величин дыхательного контроля возможен только у популяций, состоящих из родительских штаммов и штаммов-мутантов, несущих конститутивные признаки дефектности по ферментным системам транспорта, дыхания и окислительного фосфорилирования.
Задачей изобретения является разработка способа определения дыхательного контроля у популяции бактерий по степени ауторегулируемого торможения активного дыхания клеток, которое непосредственно связано с биоэнергетической эффективностью метаболических процессов в микробной популяции.
Поставленная задача решается благодаря тому, что в заявляемом способе определения дыхательного контроля у популяции бактерий предусмотрены следующие отличия:
для определения дыхательного контроля не требуется отмывка и истощение по эндогенным субстратам исследуемой бактериальной популяции. В качестве анализируемой пробы используется бактериальная популяция in situ (в составе жидких культур любого происхождения, а также в составе сухих культур после их регидратации в щадящих условиях);
в качестве одномоментно добавляемого в среду инкубации со стабильно дышащей на эндогенных субстратах популяции бактерий используется хорошо утилизируемый этими микроорганизмами углевод моносахаридной природы - Д(+)глюкоза в микроконцентрациях (10... .25 нмоль/см3);
по полярограмме дыхания популяции бактерий регистрируется не только увеличение скорости убыли кислорода из среды инкубации, но и последующее ее торможение через 2... 5 минут либо отсутствие этого торможения в течение всего времени дыхания, вплоть до исчерпания кислорода в среде инкубации;
количественное определение дыхательного контроля у популяции бактерий производят по соотношению скоростей до и после торможения активного дыхания бактерий, утилизирующих глюкозу.
Указанные отличия обусловлены следующими причинами:
исключение этапа предварительной подготовки бактериальной популяции, связанной со стрессовыми воздействиями на клетки (отмывка и голодание), обеспечивает проведение анализа в физиологических условиях, то есть позволяет регистрировать величину дыхательного контроля, близкую к условиям in situ;
глюкоза является универсальным энергетическим субстратом питания бактерий и часто используется в качестве основного источника углерода и энергии при выращивании микробных культур, в связи с чем бактериальные клетки всегда содержат конститутивные системы транспорта и катаболизма этого моносахарида в отличие от олигосахаридов, использующих индуцибельную лактозопермеазную систему транспорта, а также менее эффективные и ограниченные по составу ферментные системы внутриклеточного расщепления этих веществ [Готтшалк Г. Метаболизм бактерий / Пер. с англ. под ред. Е.Н.Кондратьевой. - М., Мир, 1982. - С.19-42, 93-95; Степанов А.С., Наумов А.В., Кондрашин Ю.И. Система транспорта Д-глюкозы и метил-α -Д-глюкопиранозида у чумного микроба // В кн.: Патологическая физиология особо опасных инфекций. Труды противочумного учреждения. - Саратов, 1984. - С.16-21]; использование глюкозы в микроколичествах (10... 25 нмоль) позволяет регистрировать эффект ауторегулируемого торможения активного дыхания популяции бактерий, утилизирующих этот углевод, либо отсутствие эффекта торможения дыхания даже после потребления бактериями кислорода в количествах, превышающих стехиометрически рассчитанное (60... 150 нмоль) на полное окисление одномоментно введенного в среду инкубации моносахарида. Последнее обусловлено тем обстоятельством, что у популяции бактерий под влиянием, например, стрессовых воздействий ("замораживание-оттаивание", длительное хранение при околонулевых и отрицательных температурах и др.) происходит нарушение регуляции дыхательной активности, опосредованное структурно-функциональными повреждениями цитоплазматической мембраны, в которой интегрированы системы транспорта, окислительного фосфорилирования и комплексы дыхательных ферментов (дегидрогеназы, цитохромы, убихиноны и др.).
Способ включает следующие этапы:
приготовление среды инкубации и раствора Д(+)глюкозы;
подготовка к работе полярографической установки;
подготовка к анализу исследуемой пробы;
проведение анализа (с записью динамики дыхательной активности исследуемой популяции бактерий на диаграммную ленту самописца);
расчет коэффициента дыхательного контроля (ДК).
Приготовление среды инкубации и раствора Д(+)глюкозы:
для приготовления среды инкубации в мерную колбу (V=1000,0 мл) помещают 4,0 г КН2РО4, 32,2 г Na2HPO4·12H2O и 0,8 г МgСl2·6Н2О, добавлением дистиллированной воды объем раствора доводят до метки;
для приготовления раствора Д(+)глюкозы в мерную колбу (V=10,0 мл) помещают 18 мг углевода, добавлением среды инкубации объем раствора доводят до метки, последующим разведением концентрацию полученного 0,01 М раствора глюкозы доводят до любой в диапазоне 0,001... 0,0025 М (по выбору исследователя).
Подготовка к работе полярографической установки:
в заявляемом способе используют полярографическую установку лабораторного изготовления с термостатируемой полярографической ячейкой и электрической схемой питания электродов (делитель напряжения) [Руководство по изучению биологического окисления полярографическим методом / Под ред. Г.М.Франка. - М.: Наука, 1973. - С.8-25, 50-59, 81-85] либо полярограф промышленного изготовления типа ОН-102 (ОН-105); Венгрия [Метаболизм микроорганизмов (практикум) / Под ред. Н.С.Егорова. - М.: Изд-во Моск. ун-та, 1986. - С.34-38];
в качестве электродов применяют платиновый электрод измерения ЭПВ-1 и хлорсеребряный электрод сравнения ЭВЛ-1МЗ из комплектов рН-метра-милливольтметра рН-121, либо универсального иономера-потенциометра ЭВ-74, либо аналогичные;
за 30 мин до начала анализа включают полярограф (делитель напряжения), самописец и термостат;
полярографическую ячейку заполняют средой инкубации, в центральное отверстие плотно вставляют платиновый электрод ЭПВ-1, в одно из боковых отверстий - агар-солевой мостик, обеспечивающий контакт с вспомогательным электродом ЭВЛ-1МЗ;
включают магнитную мешалку и перемешивают среду в ячейке в течение 3... 4 минут, после чего удаляют с помощью салфеток остатки среды с поверхности ячейки и контролируют отсутствие в ячейке пузырьков воздуха: при наличии последних удаляют их, выключив мешалку и добавив в ячейку среду инкубации;
температуру среды инкубации поддерживают в физиологических пределах в зависимости от вида исследуемых бактерий (например, для бактерий Y.pestis и E.coli - 26... 28° С, либо 36... 38° С);
на платиновый и хлорсеребряный электроды с помощью полярографа (делителя напряжения) подают потенциал 0,6 В ("минус" - на платиновом электроде);
устанавливают скорость протяжки диаграммной ленты самописца 20 мм/мин и приступают к выполнению анализа после установления постоянного уровня диффузионного тока (прямая линия по направлению движения диаграммной ленты в крайнем положении шкалы самописца), который соответствует 100% исходного содержания растворенного в среде кислорода (для перевода пера самописца в крайнее положение используют потенциометры настройки полярографа либо делителя напряжения).
Подготовка к анализу исследуемой пробы:
через боковое отверстие полярографической ячейки в среду инкубации вводят 10... 50 мкл исследуемой пробы популяции бактерий из расчета создания в среде общей концентрации, например, (1... 3)· 109 клеток/см3, с тем, чтобы отклонение пера самописца в сторону уменьшения силы тока ("белковый бросок") не превышало 20% ширины диаграммной ленты, а скорость эндогенного дыхания популяции составляла 2... 10 мм/мин.
Проведение анализа:
на фоне стабильного уровня эндогенного дыхания популяции бактерий (через 5... 10 минут после введения пробы) с помощью автоматической пипетки в среду одномоментно вводят 20 мкл раствора глюкозы в выбранной из диапазона 0,001... 0,0025 М концентрации и регистрируют на ленте самописца динамику убыли кислорода из среды инкубации;
регистрацию динамики убыли кислорода прекращают через 3... 5 минут при отсутствии стимуляции дыхания глюкозой, либо через 5... 7 минут после смены быстрой фазы дыхания медленной, либо до исчерпания кислорода в среде инкубации при отсутствии торможения быстрой фазы дыхания популяции бактерий, после чего регистрацию прекращают, трижды промывают ячейку и платиновый электрод дистиллированной водой.
Расчет коэффициента дыхательного контроля (ДК):
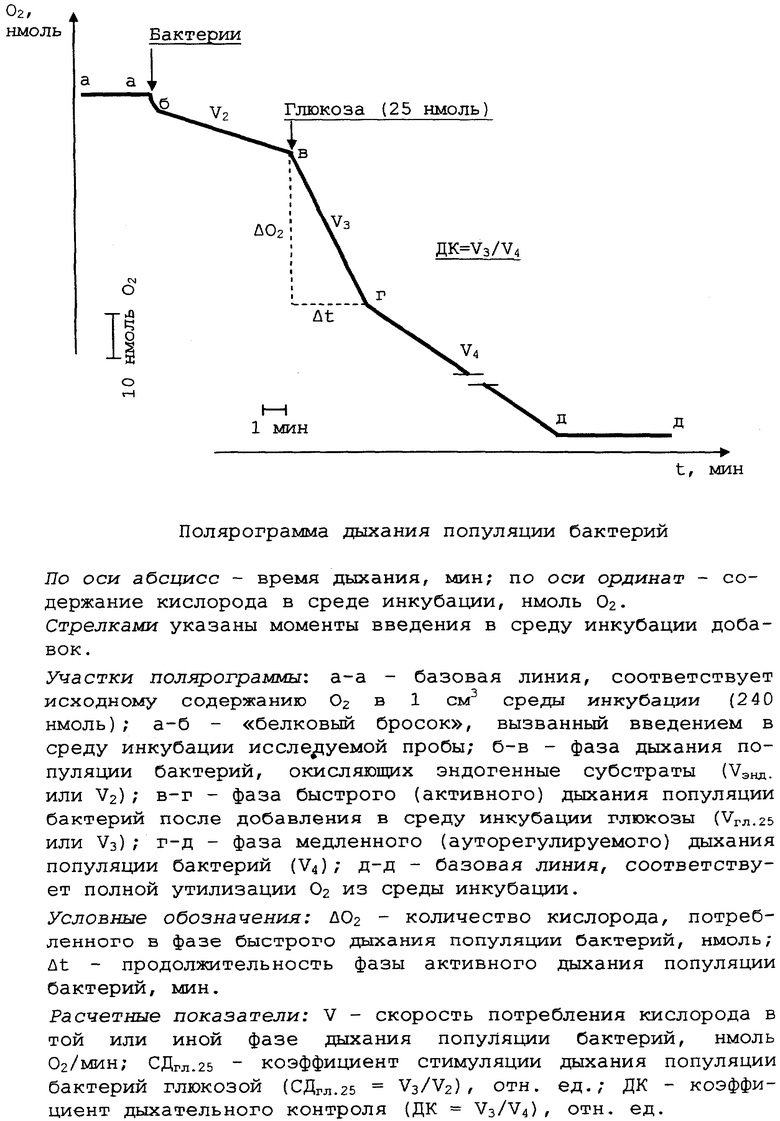
по полярограмме дыхания, типичный вид которой приведен на чертеже, рассчитывают для исследуемой популяции бактерий величину дыхательного контроля как отношение скорости в быстрой фазе окисления (V3) к скорости в медленной фазе окисления (V4);
поскольку показатель ДК является относительным, расчет скоростей дыхания V3 и V4 носит условный характер: каждая из них численно равна длине катета в (миллиметрах), противолежащего углу между вектором дыхания и направлением движения диаграммы, причем длину прилежащего катета выбирают равной 20 мм, и при скорости протяжки ленты 20 мм/мин этот катет на абсциссе времени численно равен 1 минуте;
пример расчета: ДК=V3/V4=20 мм/8 мм=2,50 отн.ед;
окончательный результат анализа оформляется в виде среднего арифметического трех определений.
Значимость заявляемого способа и рассчитываемого параметра дыхательного контроля для физиолого-биохимической оценки эффективности регуляции дыхательной активности популяции бактерий обоснована в экспериментах с культурами чумного микроба вирулентного (1300) и вакцинного (ЕВ) штаммов, также с культурами кишечной палочки (штамм К12), окислительный метаболизм и дыхательный аппарат которых весьма схожи [Голубинский Е.П. Дыхательный аппарат и окислительный метаболизм чумного микроба: Автореферат дис... на соискание ученой степени д-ра мед. наук. - ВНИИПЧИ "Микроб", Саратов, 1974. - 34 с.; Haddock В.A., lones C.W. Bacterial respiration / J. Bacteriol, 1977. - Vol.41. - No 1. - P.47-99; Ingledew W.J., Poole P.K. The respiratory chain of Escherichia coli. / Microbiol. Rev., 1984. - Vol.48. - No.3. - P.222-271]. Статистическая обработка экспериментальных данных произведена в соответствии с рекомендациями, изложенными в учебном пособии [Г.Ф.Лакин. Биометрия / Учебное пособие. - 3-е изд., перераб. и дополн., М., Высшая школа, 1980. - С.83-116, 142-163].
Как следует из полярограммы, приведенной на чертеже, за период фазы быстрого окисления 25 нмоль глюкозы популяция бактерий потребляет, как правило, 30... 40 нмоль кислорода (Δ О2). Однако в некоторых опытах торможения дыхания популяции бактерий не происходит и после потребления 150 нмоль О2, необходимых для полного окисления всей добавленной глюкозы
(ДК=1,00), либо это торможение слабо выражено (ДК=1,10... 1,30).
Регистрируемая в опытах вариабельность значений дыхательного контроля (1,0<ДК<3,0), а также отсутствие в ряде случаев двухфазности процесса дыхания клеток (ДК=1,00) имеют следующее теоретическое и экспериментальное объяснение.
Первичные этапы катаболизма глюкозы у бактерий включают транспорт углевода и превращение его в глюкозо-6-фосфат и фруктозо-1,6-дифосфат. Поскольку эти процессы требуют затраты энергии АТФ, следует предположить, что внутриклеточное отношение [АТФ]/[АДФ] у бактерий уменьшается, а активация дыхания, как и в способе по прототипу при утилизации олигосахаридов, в значительной мере обусловлена окислением эндогенных субстратов в дыхательной цепи для компенсации низкоэнергетического сдвига. Для подтверждения этого предположения в предварительных опытах с культурами чумного микроба и кишечной палочки показано, что продолжительность фазы активного дыхания (Δ t) популяций бактерий, окисляющих глюкозу в микроколичествах, совпадает по времени с продолжительностью ее полного потребления клетками из среды инкубации (таблица 1).
Другими подтверждениями сформулированного предположения явились приведенные в таблице 2 данные, которые показывают, что ингибирование гликолиза моноиодацетатом натрия (МИА) устраняет эффект стимуляции дыхания популяции бактерий глюкозой в микроколичестве (СД=1,00) только после ее повторного добавления. В то же время добавление на этом фоне экзогенного пирувата - одного из С3 - продуктов катаболического расщепления глюкозы и одновременно ключевого субстрата цикла трикарбоновых кислот - приводит к новой стимуляции дыхания популяции бактерий (СД>1,00). Эти экспериментальные данные свидетельствуют в пользу того, что механизм стимулирующего действия глюкозы в микроколичествах не отличается от такового у ее биохимического аналога α -метил-β -D-глюкопиранозида (α -МГ), метаболизм которого ограничивается первичными актами фосфорилирования при его переносе в клетки бактерий [Степанов А.С., Наумов А.В., Кондрашин Ю.И. Система транспорта Д-глюкозы и метил-α -Д-глюкопиранозида у чумного микроба // В кн.: Патологическая физиология особо опасных инфекций. Труды противочумного учреждения. - Саратов, 1984. - С.16-21]. Однако для регистрации стимулирующего действия α -МГ требуются значительно большие его количества, как и в прототипном способе - на примере олигосахаридов.
Таким образом, совпадение продолжительности активного дыхания (Δ t) и продолжительности аккумуляции глюкозы в микроколичествах, а также результаты, представленные в таблице 2, подтверждают, что активация дыхания бактерий при добавлении микроколичеств глюкозы связана с инициацией окисления эндогенных субстратов, компенсирующего затраты энергии на транспорт и первичные реакции фосфорилирования утилизируемого углевода. Фаза активного дыхания продолжается до тех пop, пока происходит энергопотребляющий процесс утилизации глюкозы, т.е. создаются условия для сдвига равновесия реакции "фосфорилирование АДФ  дефосфорилирование АТФ" в правую сторону.
дефосфорилирование АТФ" в правую сторону.
В то же время степень ауторегулируемого торможения дыхания популяции бактерий, окисляющих глюкозу, характеризуемая величиной ДК, то есть переход ее в состояние "отдыха”" (V4) после периода активного дыхания (V3) представляет собой по аналогии с митохондриями наиболее информативный показатель эффективности энергетической регуляции окислительного обмена.
Для подтверждения этого в качестве добавок, снижающих энергизованность клеток в составе популяции бактерий, использовали разобщитель - протонофор 2,4-динитрофенол (2,4-ДНФ) в конечной концентрации 0,1 мМ и неметаболизируемый аналог глюкозы α -МГ (10 мМ), энергозависимый транспорт которого в клетки связан с затратами энергии АТФ. Избирательное ингибирование протонтранслоцирующей аденозинтрифосфатазы (АТФ-азы) дициклогексилкарбодиимидом (ДЦКД) в концентрации 5 мМ, напротив, способствует энергизации клеток в экспоненциальной фазе развития, когда активность данного фермента наиболее высока [Burstein С., Tiankova L., Kepes A. Respiratory control in Escherichia coli К 12 / Eur. J.Biochem,- 1979, No 94. - P.387-392]. Результаты исследования закономерностей ауторегуляции дыхательной активности популяции бактерий под влиянием энергизующих и деэнергизующих добавок представлены в таблице 3.
Из данных таблицы 3 следует, что воздействия, приводящие к разобщению окислительного фосфорилирования и деэнергизации клеток, вызывают ослабление ауторегулируемого торможения окислительной активности, что проявляется в увеличении скорости дыхания популяции бактерий в состоянии "отдыха" (V4) и, как следствие, в снижении величины ДК. Напротив, энергизация клеток, обработанных ДЦКД, приводит к достоверному снижению скорости дыхания V4 и достоверному увеличению значений ДК. Диапазон и направленность изменений других параметров дыхания в этих опытах не адекватна характеру воздействий на клеточный метаболизм и не поддается однозначной трактовке.
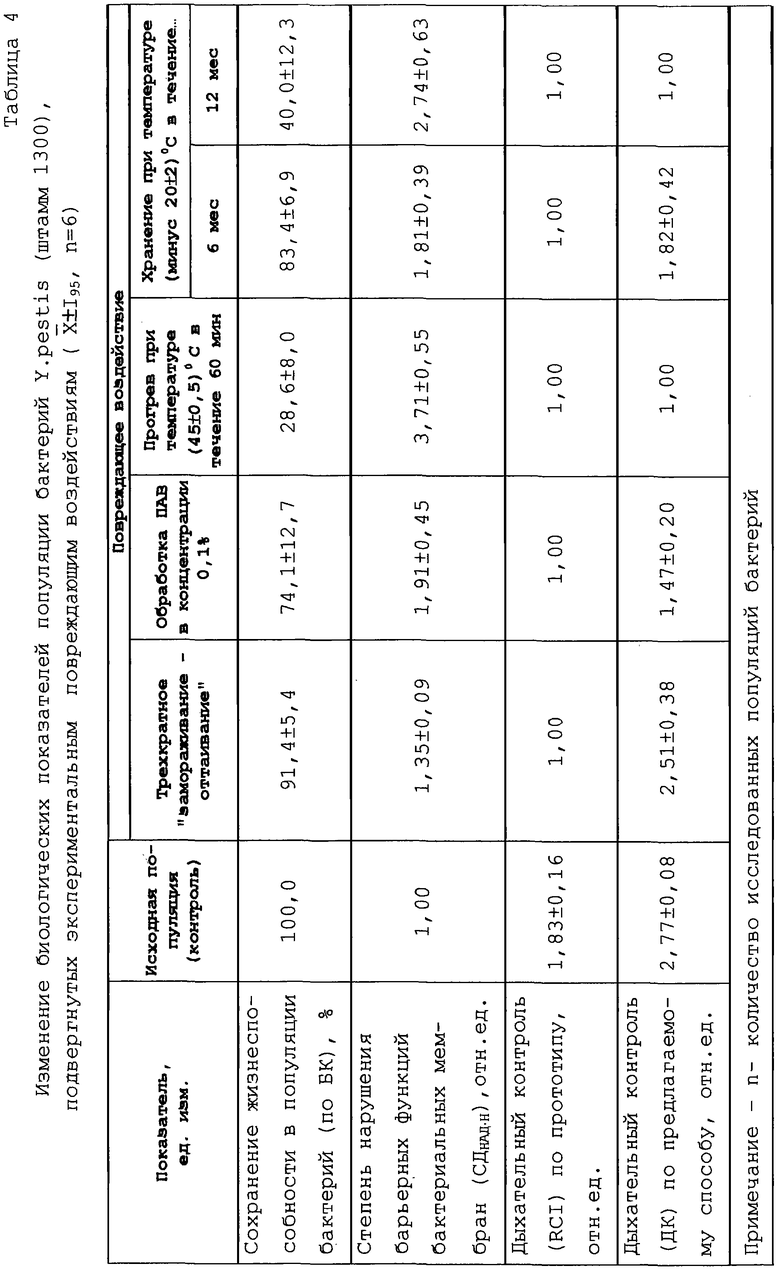
Сравнительную оценку значимости показателей дыхательного контроля по заявляемому (ДК) и прототипному (RCI) способам их определения для характеристики биоэнергетического статуса популяции бактерий осуществили в опытах с культурами чумного микроба (штамм 1300), подвергнутым экспериментальным повреждающим воздействиям: трехкратное "замораживание-оттаивание" в диапазоне температур минус 20° С... плюс 20° С; обработка поверхностно-активным веществом (ПАВ) толуолом в конечной концентрации 0,1%; прогрев культур при температуре (45± 0,5)° С в течение 60 минут; длительное хранение культур (6 и 12 месяцев) в замороженном состоянии при температуре (минус 20±2)° С.
В ходе экспериментов оценивали устойчивость бактерий к повреждающим воздействиям по сохранению ими способности к образованию колоний на плотной питательной среде (ППС) в чашках Петри, то есть определением биологической концентрации (БК) микробных культур. Показатель сохранения жизнеспособности выражали в доле бактерий, сохранивших способность к колонеобразованию, принимая значение БК исходной популяции бактерий за 100%. Учитывая то обстоятельство, что основными мишенями подобного рода повреждающих воздействий являются мембранные структуры бактериальных клеток, выполняющие не только энергетические, но и барьерные функции, в качестве другого показателя устойчивости популяции бактерий использовали степень нарушения барьерных функций клеток, определяемую по параметру СДНАД·Н [С.Г.Игнатов, В.А.Красильников, В.В.Перелыгин и др. Исследование мембранного аппарата Escherichia coli после низкотемпературного замораживания /Биохимия, 1980.- Т.46. - С.1996-2003].
Экзогенный никотинамидадениндинуклеотид восстановленный (НАД·Н) не проникает в интактные клетки бактерий (СДнАд·н=1,00), однако нарушение избирательной проницаемости барьерных структур и, прежде всего, цитоплазматической мембраны (ЦПМ) приводит к его проникновению на внутреннюю сторону ЦПМ, где интегрирован его специфический фермент НАД· Н - дегидрогеназа. Результатом этого взаимодействия является стимуляция дыхания поврежденных клеток при добавлении в среду инкубации экзогенного НАД-Н (СДНАД· Н>1,00), причем выраженность эффекта стимуляции тем выше, чем сильнее нарушение избирательной проницаемости клеточных мембран.
Конечная концентрация субстратов в пробах в серии опытов, результаты которых представлены в таблице 4, составляла: НАД· Н - 50 мМ; олигосахаридов - 10 мМ; глюкозы - 10 мкМ и 25 мкМ.
Как следует из данных таблицы 4, по мере усиления повреждающих воздействий в популяции снижается относительная доля жизнеспособных клеток, возрастает степень нарушения барьерных функций бактериальных мембран и снижается уровень энергозависимой регуляции дыхательной активности, определяемой по параметру ДК, вплоть до утраты дыхательного контроля (ДК=1,00). В то же время параметр RCI, определяемый по прототипу, принимает минимальное значение уже после относительно мягкого воздействия на популяцию бактерий "замораживанием-оттаиванием" и при усилении интенсивности повреждающих воздействий на популяцию эндогенное дыхание бактерий олигосахаридами не стимулируется (RCI=1,00).
В таблице 5 приводятся значения коэффициента корреляции (r) между параметрами дыхательного контроля, определяемыми по прототипу (RCI) и предлагаемому (ДК) способу, и биологическими показателями бактериальной популяции, представленными в таблице 4. Значения r рассчитаны для выборок, каждая из которых состоит из 36 независимых определений показателя (для 6 микробных культур в каждом из 6 опытов). Сильная коррелятивная связь (r>0,7) выявлена между показателем дыхательного контроля (ДК) и сохраняемостью жизнеспособных клеток, а также степенью нарушения барьерных функций клеточных мембран. Для показателя RCI соответствующие значения r меньше 0,5.
Свидетельством применимости параметра ДК для оценки качества лиофильно высушенных эталонных культур чумного микроба (штамм EV), используемых для приготовления на их основе чумных вакцин, являются результаты, представленные в таблице 6.
Разнокачественные эталонные культуры вакцин штамма EV серий 47, 71 и 79 использовались для приготовления посевных культур и далее - нативных культур при выращивании последних в аппаратах вместимостью 100 дм3 методом глубинного культивирования с принудительной аэрацией и дробным добавлением в ферментеры глюкозы как основного источника углерода и энергии. Согласно технологическим условиям показатели выхода биомассы и потребления за весь цикл культивирования глюкозы (27... 30 часов) являются наиболее значимыми параметрами качества получаемых нативных культур. Перед измерением параметров дыхательного контроля эталонные культуры регидратировали в физиологическом растворе.
Данные таблицы 6 свидетельствуют о том, что наиболее высокие значения биомассы и потребления глюкозы бактериями характерны для нативных культур, выращенных на основе эталонов серии 71 и, особенно, серии 79. Для этих вакцин значения ДК>1,00, причем максимальные значения ДК соответствуют максимальным значениям показателей качества нативных культур. Для всех исследованных эталонных культур вакцин значения дыхательного контроля по прототипу (RCI) не превышали минимального значения 1,00, что не позволяет дифференцировать культуры по этому параметру.
Продолжительность фазы активного дыхания и время аккумуляции глюкозы микробной популяцией, состоящей из
1млрд. живых бактерий ( I95)
I95)
1) Время аккумуляции глюкозы измеряли по ее убыли из среды инкубации с использованием анализатора глюкозы фирмы "Beck-mаn", США
2) 1,8 мкг соответствует 10 нмоль глюкозы в среде инкубации
3) 4,5 мкг соответствует 25 нмоль глюкозы в среде инкубации
Стимуляция дыхания популяции чумного микроба (штамм 1300) различными субстратами ( ±I95, n=7)
±I95, n=7)
Изменение параметров дыхания популяции бактерий Y.pestis (штамм 1300) под действием разобщителя 2,4-динитрофенола (2,4-ДНФ), неметаболизируемого α -метил-β -D-[Улюкопиранозида (α -МГ) и ингибитора АТФ-азы дициклогексил-карбодиимида (ДЦКД), ( ± I95)
± I95)
1) За 100% принимаются значения параметров дыхания популяции бактерий в контрольной пробе (без добавок 2,4-ДНФ, α -МГ, ДЦКД)
2) n - количество исследованных популяций бактерий

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОЦЕНКИ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ МИКРОБНОЙ ПОПУЛЯЦИИ | 2001 |
|
RU2195496C2 |
| СРЕДСТВО НА ОСНОВЕ ПРИРОДНЫХ ФОСФОЛИПИДОВ | 2008 |
|
RU2367443C1 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО АНТИГИПОКСИЧЕСКОГО ДЕЙСТВИЯ | 2001 |
|
RU2189813C1 |
| СПОСОБ МОНИТОРИНГА СТАДИИ КУЛЬТИВИРОВАНИЯ В ТЕХНОЛОГИИ ЧУМНОЙ ВАКЦИНЫ | 2017 |
|
RU2677954C1 |
| Способ оценки функционального состояния митохондрий | 1988 |
|
SU1668945A1 |
| Способ индивидуального подбора фармакологических препаратов при терапии психических заболеваний | 1986 |
|
SU1406485A1 |
| СРЕДСТВО, НОРМАЛИЗУЮЩЕЕ МИТОХОНДРИИ ПЕЧЕНИ | 2011 |
|
RU2470655C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ БАКТЕРИОФИКСИРУЮЩЕЙ АКТИВНОСТИ ЭРИТРОЦИТОВ | 2007 |
|
RU2360969C1 |
| Способ определения активности мышиного токсина чумного микроба | 1990 |
|
SU1775472A1 |
| Способ получения бактериальных суспензий | 1987 |
|
SU1463754A1 |
Изобретение относится к способам исследования бактериальной популяции методом полярографии. Дыхательный контроль популяции бактерий определяют по соотношению скоростей до и после торможения активного дыхания популяции бактерий, окисляющих Д(+) глюкозу в конечной концентрации 10-25 нмоль/см2 в полярографической ячейке. Скорость убыли кислорода из среды инкубации регистрируют с помощью платинового электрода открытого типа. Способ прост в применении, обладает универсальностью и повышает достоверность результатов исследования. 1 ил., 6 табл.
Способ определения дыхательного контроля у популяции бактерий, включающий предварительное инкубирование популяции бактерий в полярографической ячейке с платиновым электродом открытого типа до достижения стабильного уровня эндогенного дыхания клеток, последующее одномоментное введение в среду инкубации с популяцией бактерий углевода и количественный анализ динамики убыли кислорода из среды инкубации, отличающийся тем, что в качестве углевода используют Д(+) глюкозу в конечной концентрации 10-25 нмоль/см3, а количественный анализ динамики убыли кислорода проводят, определяя соотношение скоростей до и после торможения активного дыхания популяции бактерий, окисляющих глюкозу, и по полученному коэффициенту судят о величине дыхательного контроля у популяции бактерий.
| TOMOFUSA TSUCHIYA, BARRY P | |||
| POSEN Respiratory control in Escherichia coli, FEBS Letters, 198, v.120, N1, p.128-130 | |||
| АДО А.Д | |||
| Патологическая физиология Учебник для мед.вузов, М., 2000, Триада-Х, с.23-25 | |||
| BURSTEIN C | |||
| ET.AL | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Eur | |||
| J | |||
| Biochem, 1979, № 94, p.387-392. | |||

