Описание
Область изобретения
Изобретение связано со способом лечения или предотвращения заболевания, происходящего в результате высвобождения брадикинина у млекопитающего, в частности у млекопитающего, которое продуцирует гепарин-связывающий белок, при котором указанный гепарин-связывающий белок связывается с антагонистом гепарин-связывающего белка, включающий в себя введение указанному млекопитающему антагониста гепарин-связывающего белка в количестве, эффективном для снижения уровня высвобождения брадикинина у млекопитающего. Более того, изобретение связано со способами и наборами для определения того, продуцирует ли млекопитающее гепарин-связывающий белок, который связывается с антагонистом гепарин-связывающего белка, и способом обнаружения антагониста гепарин-связывающего белка.
Предшествующий уровень техники
Воспаление
Острый воспалительный ответ связан с несколькими явлениями, включая изменения в размерах и тонусе сосудов, так же, как и в увеличении проницаемости сосудов, что приводит к образованию обогащенного белком экссудата (Lewis in Mediators of Inflammation, Wright, Bristol, U.K., 1986).
Поскольку нейтрофилы (ПМЯ) получают хемотаксические сигналы, они располагаются по периферии сосудов и прилипают к сосудистым эндотелиальным клеткам посредством специфических адгезивных молекул, синтезируемых как на поверхности эндотелиальных клеток, так и на поверхности нейтрофилов. После прикрепления нейтрофилов к эндотелиальным клеткам в эндотелии образуются зазоры, индуцирующие проницаемость сосудов и создающие возможность для миграции нейтрофилов в интерстициальную область ткани.
Фазово-контактная система охватывает три ферментативных фактора, фактор XI (F XI), фактор XII (F XII) и прокалликреин (пре-калликреин) плазмы (РК), а также неэнзиматический кофактор, Н-кининоген (НК), который образует эквимолярные комплексы с F XI и РК, соответственно. Контактная фаза присутствует на моноцитах, фибробластах и нейтрофилах. Специфические связывающие сайты, экспонируемые эндотелиальными клетками и неклеточными отрицательно заряженными поверхностями, такими как каолин, в целом называемыми здесь "контактной фазой", создают возможность локального скопления критических компонентов. Превращение зимогена F XII в активный фермент, F ХIIа, активирует как таковую фазово-контактную систему (Colman et al., 1986, Grit. Rev. Oncol. Hematol. 5: 57-85). Реципрокная активация связанного с поверхностью F XII и РК, заякоренного на поверхности посредством НК, способствует выработке F XIIа и Pka, амплифицируя тем самым сигнал инициации. Затем фактор XIIа может активировать фактор XI, и инициируется внутренний путь свертывания. Известно, что РК также гидролизует НК с образованием сильного нанопептида, брадикинина (Kaplan and Silverberg, 1987, Blood 70:1-15). Кинины считаются первичными медиаторами процесса воспаления, которое вызывает боль, расширение сосудов и увеличение проницаемости сосудов в результате прямого действия на эндотелиальные клетки, вызывая их сокращение и допуская одновременно и миграцию нейтрофилов, и образование транссудата содержимого плазмы (Oyvin et al., 1970, Experentia 26:843-844). Может быть вовлечен также процесс локального продуцирования простагландинов и окиси азота, NO (Hall, 1992, Pharmacol. Ther. 56:131-190). Следовательно, активация фазово-контактной системы может приводить к целому ряду повреждающих эффектов, например, к воспалению, септическому шоку, респираторному дистресс-синдрому взрослых, диссеминированному внутрисосудистому свертыванию, послеоперационному кровотечению при сердечно-сосудистой хирургии.
Фазово-контактная система может быть активирована, в частности, когда нейтрофилы связываются с эндотелиальными клетками, присутствием эндотоксинов и бактериальной инфекции (обзор Colman et al., 1997, Blood 90:3819-3843). Например, было обнаружено, что при сепсисе происходит активация фактора XII и прекалликреина, при которой они расщепляются и в результате превращаются в ферменты, которые быстро реагируют с С1-ингибитором с образованием комплексов фактор XIIa-C1-ингибитор и калликреин-C1-ингибитор. Значительное увеличение образования комплекса калликреин-С1-ингибитор наблюдается при стимулированном экстракорпоральном (искусственном) кровообращении.
Обнаружено, что апротинин, ингибитор одновременно и плазмина, и калликреина плазмы, уменьшает кровопотери после сердечной хирургии и уменьшает повышенное послеоперационное время свертывания. В частности, было показано, что апротинин в модели стимулированного экстракорпорального кровообращения способствует снижению образования комплексов калликреин-C1-ингибитор и С1-С1-ингибитор (Wachtfogel et al., 1993, J. Thorac. Cardiovasc Surg. 106:1). При добавлении апротинина к искусственному кругу кровообращения, перфузируемому всем объемом крови, антикоагулируемой гепарином, апротинин фактически дополнял действие гепарина (Bannan et al., 1998, Brit. J. Haem. 101:455-461). Следовательно, апротинин оказывает дополнительное гемостатически благоприятное воздействие по сравнению с индивидуальным действием гепарина в указанной модели.
Апротинин также повышает жизнеспособность эндотелиальных клеток в условиях гипоксии при охлаждении, когда такие клетки хранят в растворах для сохранения органов, и повышает сохранность легких и миокарда в моделях, в которых используются органы целиком (Sunamori et al., 1991, Ann. Thor. Surg. 52:971-978 and Roberts et al., 1998, Ann. Thorac. Surg. 66:225-230). Более того, в то время как брадикинин, как было показано, способствует повышению сосудистой проницаемости, апротинин уменьшает эту проницаемость сосудов и количество нейтрофилов (O'Brien et al., 1997, Can. J. Physiol. Pharmacol. 75:741-749 and Dwenger et al., 1995, Eur. J. Clin. Chem. Clin. Biochem. 34:207-214).
Гепарин-связывающий белок
В настоящее время обнаружена ковалентная структура двух тесно связанных белков, выделенных из нейтрофильных лейкоцитов периферической крови человека и свиньи (cf. H. Flodgaard et al., 1991, Eur. J. Biochem. 197: 535-547; J. Pohl et al., 1990, FEBS Lett. 272: 200 ff.). Оба белка обладают большим сходством с эластазой нейтрофилов, но в результате селективных мутаций активного серина 195 и гистидина 57 (нумерация по химотрипсину (B.S. Hartley, "Homologies in Serine Proteinases", Phil. Trans. Roy. Soc. Series 257,1970, p. 77 ff.)) эти белки лишены протеазной активности. Эти белки были названы гепарин-связывающим белком человека (hHBP) и гепарин-связывающим белком свиньи (рНВР), соответственно, благодаря их высокой аффинности к гепарину.
Schafer et al. (Schafer et al., 1984, Infec. Immun. 53:651) назвали этот белок катионным противомикробным белком (САР37) благодаря его противомикробной активности. НВР крепко связывается с липидным А-компонентом LPS и эндотоксином (Kass=0,8×109 М-1). Было высказано предположение, что бактерицидный эффект НВР обусловлен связыванием с липидом A (Petersen et al., 1993, Eur. J. Biochem. B214: 271-279, Flodgaard et al., 1994, J. Cell. Biochem. Suppl. 18A: Abstr. E505; Pereira et al., 1993, Proc. Natl. Acad. Sci. USA 90: 4733-4737). Предполагаемый сайт связывания нативного НВР с липидом A/LPS локализован в виде незаряженного участка между основным и кислотным участками на НВР и содержит остатки 20-26 и 41-43. В сайте связывания липид A/LPS Phe25, Cys26, Cys42, и Phe43 образуют гидрофобный карман, пригодный для связывания либо цепей жирной кислоты, либо гликозаминильного сахарного кольца липида А. Рядом с этим карманом расположен ионный и гидрофильный карман (Asn20, Gln21, и Аrg23), который должен весьма хорошо подходить для связывания гликозаминил-связанной фосфатной группы липида A (Iversen et al., 1997, Nature Struct Biology 4: 265-268).
Более того, на животных моделях фекального перитонита было показано, что лечение НВР спасает мышей от летального повреждения (Mercer-Jones et al., 1996, In Surgical Forum, pp. 105-108 and Wickel et al., 1997, In 4m International Congress on the Immune Consequences of Trauma, Shock and Sepsis, Munich, Germany, pp.413-416). В результате было выдвинуто предположение, что гепарин-связывающий белок или его LPS-связывающие фрагменты могут быть использованы для лечения септического шока (WO 95/28949, US Pat. Nos. 5458874, 5607916 and 5650392).
Первоначально НВР изучали в связи с тем, что он обладает антибиотическими и LPS-связывающими свойствами (Gabay et al. 1989, Proc. Natl. Acad. Sci. USA. 86:5610-5614 and Pereira et al., 1993, Proc. Natl. Acad. Sci. USA 90: 4733-7). Однако накопившиеся данные в настоящее время подтверждают концепцию, что НВР, кроме его бактерицидной роли, вовлечен в процесс развития воспаления благодаря его воздействию на рекрутмент и на активацию моноцитов (Pereira et al., 1990, J. Clin. Invest. 85:1468-1476 and Rasmussen et al., 1996, FEBS Lett. 390:109-112), рекрутинг Т-клеток (Chertov et al., 1996, J. Biol. Chem. 271: 2935-2940).
Было обнаружено, что НВР индуцирует сокращение эндотелиальных клеток и фибробластов (Ostergaard and Flodgaard, 1992, J. Leuk.Biol. 51: 316-323). В этой связи в WO 93/05396 описывается способ скрининга ингибиторов НВР путем инкубации НВР или клеток, продуцирующих НВР, в присутствии вещества, у которого предполагалось наличие свойств ингибитора НВР, и в присутствии ткани или клеток, которые способны взаимодействовать с НВР; ослабленное взаимодействие (например, сокращение эндотелиальных клеток) указывает на то, что это вещество является ингибитором НВР.
В заявке WO 99/26647 описано применение гепарин-связывающего белка в модулировании или профилактике апоптоза клеток млекопитающих. В этой заявке описано также, что НВР спасает клетки инсулиномы крысы от IL-1-индуцированного апоптоза.
Было также обнаружено, что гепарин-связывающий белок человека, в отличие от гепарин-связывающего белка свиньи, связывается с апротинином (BPTI) (Petersen et al., Eur. J. Biochem. 214:271-279). В частности, BPTI обладает способностью связывать НВР с Kd=0,1×10-6 M (Petersen et al., 1993, Eur. J. Biochem. B214: 271-279). P1-специфичность НВР, как было показано, в первую очередь связана с Lys или Leu (Kiczak et al., 1999, Biol. Chem. 380: 101-105). Было выдвинуто предположение, что наиболее характерными остатками в структуре НВР, которые ответственны за связывание BPTI, являются Gly169, Gly175, Ser192, и Asp201, в соответствии с Asp189, Ser195, Gly216, и Asp226 в трипсине (Petersen et al., 1993, Eur. J. Biochem. B214: 271-279). Kiczak et al., 1999, Biol. Chem. 380:101-106, используя методы фагового распределения, сконструировали библиотеку мутантов боковой цепи Р1 апротинина. Этими исследователями обнаружено, что НВР обладает большой аффинностью в отношении Р1 Lys, так же, как и в отношении незаряженных аминокислот P1 - Leu, Thr, Met, Gin.
Краткое содержание изобретения
Случайно было обнаружено, что гепарин-связывающий белок (НВР) служит сигналирующим звеном в индуцированной нейтрофилами проницаемости сосудов и в активации фазово-контактной системы с одновременным образованием брадикинина и что это, в частности, влияет на опосредованное РК расщепление НК с получением брадикининовой последовательности. Кроме того, было показано, что антагонисты НВР уменьшают проницаемость эндотелиальных клеток. Как здесь показано, "антагонистом НВР" является вещество, которое связывается с гепарин-связывающим белком и ингибирует действие гепарин-связывающего белка.
Данное изобретение направлено на способ лечения или предотвращения заболеваний, происходящих в результате высвобождения брадикинина у млекопитающего, в частности, у больного человека, в частности, у млекопитающего, у которого продуцируется гепарин-связывающий белок, который связывается с антагонистом НВР, включающий в себя введение указанному млекопитающему, в случае необходимости, антагониста гепарин-связывающего белка в количестве, эффективном для модуляции или снижения уровня высвобождения брадикинина у указанного млекопитающего. Такие заболевания включают в себя, не ограничиваясь ими, синдром системной воспалительной реакции, ишемическую реперфузию, анафилаксию и отторжение аллотрансплантата. Такие заболевания могут также включать в себя респираторный дистресс-синдром взрослых как побочный эффект синдрома системной воспалительной реакции. Анафилаксия может возникать на фоне нежелательной активации ПМЯ в процессе искусственного кровообращения, легочной хирургии, травмы головы и общей травмы всего организма. Модуляция или снижение степени высвобождения брадикинина происходит в результате предотвращения контакта НВР с эндотелиальными клетками и/или с фазово-контактной системой. В особом аспекте антагонист НВР модулирует или снижает опосредованное калликреином расщепление Н-кининогена с получением брадикининовой последовательности. Далее, изобретение направлено на применение антагониста НВР для изготовления лекарственного средства, применимого для лечения синдрома системной воспалительной реакции, ишемической реперфузии, анафилаксии и отторжения аллотрансплантата у больного, имеющего НВР, который связывается с антагонистом НВР. Антагонисты НВР могут, кроме того, быть использованы для лечения или изготовления лекарственного средства, применимого для лечения осложнения синдрома системной воспалительной реакции, респираторного дистресс-синдрома взрослых.
В особом аспекте изобретение направлено на способ предотвращения заболеваний, происходящих в результате высвобождения брадикинина у млекопитающего, в частности, у больного человека, в частности, у млекопитающего, у которого продуцируется гепарин-связывающий белок, который связывается с антагонистом НВР, включающий в себя введение указанному млекопитающему, в случае необходимости, домена ингибитора сериновой протеиназы наподобие сериновой протеиназы Кунитца или его аналога или производного, которое связывается с НВР, в количестве, эффективном для модуляции или снижения высвобождения брадикинина у указанного млекопитающего.
В другом, особом, аспекте данное изобретение направлено на способ предотвращения заболеваний, происходящих в результате высвобождения брадикинина у млекопитающего, в частности, у больного человека, в частности, у млекопитающего, у которого продуцируется гепарин-связывающий белок, который связывается с моноклональным антителом, которое связывается по меньшей мере с одним эпитопом на НВР, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина, включающий в себя введение указанному млекопитающему, в случае необходимости, моноклонального антитела, которое связывается по меньшей мере с одним эпитопом на НВР, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина, в количестве, эффективном для модуляции или снижения высвобождения брадикинина у указанного млекопитающего.
Данное изобретение, далее, направлено на обнаружение антагониста НВР. В одном из аспектов указанный способ включает в себя (а) культивирование эндотелиальных клеток в присутствии НВР и в присутствии и в отсутствие вещества, которое предположительно является указанным антагонистом, и (b) обнаружение любого действия указанного вещества на проницаемость эндотелиальных клеток, причем сниженная проницаемость указанных эндотелиальных клеток по сравнению с проницаемостью указанных клеток, когда их инкубируют в присутствии НВР, но в отсутствие указанного вещества, указывает на то, что указанное вещество является антагонистом. В другом аспекте указанный способ включает в себя (а) инкубирование НВР в присутствии первого вещества, которое взаимодействует с НВР, и второго вещества, которое по предположению является антагонистом НВР, и (b) обнаружение любого действия указанного второго вещества, которое по предположению является антагонистом НВР, на взаимодействие НВР с указаным первым веществом.
В особом аспекте данное изобретение направлено на способы идентификации моноклонального антитела, которое связывается по меньшей мере с одним эпитопом на НВР, в частности, НВР человека, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина. В одном из аспектов указанный способ включает в себя (а) культивирование эндотелиальных клеток в присутствии НВР и в присутствии и в отсутствие моноклонального антитела, которое предположительно связывается по меньшей мере с одним эпитопом НВР, в частности, НВР человека, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина, и (b) обнаружение любого действия указанного вещества на проницаемость эндотелиальных клеток, причем сниженная проницаемость указанных эндотелиальных клеток по сравнению с проницаемостью указанных клеток, когда их инкубируют в присутствии НВР, но в отсутствие указанного антитела, указывает на то, что указанное антитело является моноклональным антителом, которое связывается, по меньшей мере, с одним эпитопом НВР, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина. В другом аспекте указанный способ включает в себя (а) инкубирование прекалликреин-Н-кининогенового комплекса в присутствии НВР и в присутствии и в отсутствие моноклонального антитела, которое предположительно связывается, по меньшей мере, с одним эпитопом на НВР, в частности, НВР человека, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина, и (b) обнаружение любого действия указанного антитела на высвобождение брадикинина, причем сниженное высвобождение брадикинина указывает на то, что указанное антитело связывается, по меньшей мере, с указанным эпитопом на НВР.
Данное изобретение направлено также на способы и наборы для определения того, продуцируют ли млекопитающие НВР, который связывается с антагонистом НВР. Метод включает в себя следующие стадии: (а) выделение НВР или клеток или тканей, продуцирующих НВР, из млекопитающего, в частности больного человека; (b) инкубирование указанного НВР или клеток или тканей, продуцирующих НВР, с веществом, тканью, клетками или их компонентами, которые взаимодействуют с НВР и указанным антагонистом НВР, и (с) обнаружение воздействия указанного гепарин-связывающего антагониста на взаимодействие НВР с указанным веществом, тканью, клетками или их компонентами, причем сниженное взаимодействие указывает на то, что указанное НВР связывается с указанным антагонистом НВР. Набор для тестов включает (а) антагонист НВР; (b) нативный НВР и (с) вещество, ткань, клетки или их компонент, которые взаимодействуют с НВР.
В особом аспекте данное изобретение направлено также на способы и наборы для определения того, продуцирует ли млекопитающее, в частности больной человек, НВР, который связывается с моноклональным антителом, которое связывается по меньшей мере с одним эпитопом на нативном НВР, в частности, НВР человека, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина, включающие в себя (а) выделение НВР или клеток или тканей, продуцирующих НВР, из указанного млекопитающего; (b) культивирование указанного НВР или клеток или тканей, продуцирующих НВР, с эндотелиальными клетками в присутствии или в отсутствие указанного антитела, и (с) обнаружение любого воздействия указанного антитела на проницаемость эндотелиальных клеток, причем сниженная проницаемость указывает на то, что НВР связывается с указанным антителом. В другом аспекте способ включает в себя (а) выделение НВР или клеток или тканей, продуцирующих НВР, из указанного млекопитающего; (b) инкубирование указанного НВР или клеток или тканей, продуцирующих НВР, с прекалликреин-Н-кининогеновым комплексом в присутствии или в отсутствие указанного антитела, и (с) обнаружение любого воздействия указанного НВР на высвобождение брадикинина из прекалликреин-Н-кининогенового комплекса, причем сниженное высвобождение брадикинина указывает на то, что указанный НВР связывается с указанным антителом. Набор согласно изобретению включает в себя (а) моноклональное антитело, которое связывается по меньшей мере с одним эпитопом на нативном НВР, в частности, с НВР человека, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина, (b) нативный НВР человека, и (с) прекалликреин-Н-кининогеновый комплекс, прикрепленный к твердому носителю.
Краткое описание чертежей
На фиг.1 показан эффект перекрестного связывания CD 18 со вторичным козьим антителом против мышиного F{аb}2.
На фиг.2 показан зависимый от дозы эффект НВР (25-75 мкг/мл) на проницаемость монослоев эндотелиальных клеток.
На фиг.3 показано ингибирование НВР-индуцированного увеличения проницаемости эндотелиальных клеток (ЭК) поликлональной антисывороткой против НВР.
На фиг.4 показано, как влияют антитела против НВР на функцию эндотелиального барьера под действием продуктов секреции полиморфно-ядерных лейкоцитов (ПМЯ).
На фиг.5 показано ингибирование вызванного брадикинином и НВР увеличения проницаемости ЭК под действием моноклонального антитела МВК3 против брадикинина. Брадикинин (100 нМ; фиг. 5а) или НВР (75 мкг/мл; фиг.5b) вводили во время, соответствующее нулю, в боковую пластинку монослоев эндотелиальных клеток, проинкубированных в присутствии mAb МВК3 (40 мкг/мл).
На фиг.6 показано ингибирование вызванного НВР увеличения проницаемости ЭК под действием моноклонального антитела РКН4 против калликреина плазмы. НВР (75 мкг/мл; заполненные кружочки) или брадикинин (100 нМ; полые кружочки).
На фиг.7 показано ингибирование вызванного НВР увеличения проницаемости ЭК под действием обработки пептидом НКН20.
На фиг.8 показано ингибирование вызванного НВР увеличения проницаемости ЭК при обработке пептидом SDD31.
На фиг.9 показано ингибирование вызванного НВР увеличения проницаемости ЭК при обработке апротинином.
Фиг.10: высвобождение Il-6 из моноцитов человека. Моноциты культивировались в 1 мл бессывороточной среды в течение 24 часов в присутствии LPS и/или нативного НВР /[R23S, F25E]HBP/ [G175Q] HBP в количествах, указанных на фигуре.
Фиг.11: Насыщение нативного НВР, [R23S,F25E]HBP и [G175Q]HBP (A) 3H-LPS и (В) конкуренцией за связывание 125I-BPTI фиксированными, увеличивающимися концентрациями немеченого BPTI. (С) Концентрации 125I-BPTI показаны при 0 нМ % немеченого BPTI. Кажущееся различие в связывании между НВР и [R23S,F25E]HBP обсуждается в разделе "Результаты". Стрелками показаны стандартные отклонения.
Подробное описание изобретения
Гепарин-связывающий белок
В данном контексте термин "гепарин-связывающий белок" ("НВР") означает белок, который (i) протеолитически неактивен; (ii) содержался в азурофильных гранулах полиморфно-ядерных лейкоцитов; и (iii) является хемоаттрактантом для моноцитов и/или активирует моноциты. НВР может, соответственно, являться белком млекопитающего, в частности человека. В частности, НВР представляет собой зрелый НВР человека, имеющий по меньшей мере примерно 80% идентичность с аминокислотной последовательностью, представленной в SEQ ID NO:1, более предпочтительно, по меньшей мере, примерно 90%, еще более предпочтительно, по меньшей мере, примерно 95%, и наиболее предпочтительно, по меньшей мере, примерно 97% (далее в описании "гомологичные полипептиды"), которые качественно сохраняют активность НВР.
SEQ ID NO:1:

Аминокислотные последовательности гомологичных полипептидов отличаются от аминокислотной последовательности, представленной в SEQ ID NO:1, включением или делецией одного или более аминокислотных остатков и/или заменой одного или более аминокислотных остатков различными аминокислотными остатками. Предпочтительно, чтобы изменения аминокислот имели второстепенную природу, т.е. чтобы имели место такие консервативные аминокислотные замены, которые не оказывают существенного влияния на пространственную упаковку и/или активность протеина; малые делеции, обычно порядка 30 аминокислот; малые амино- или карбокси-концевые привески, такие как амино-концевой метиониновый остаток; малый линкерный пептид, состоящий примерно из 20-25 остатков; или малый привесок, который облегчает очистку путем изменения нейтрального заряда или выполняющий другую функцию, такой как полигистидиновый тракт, антигенный эпитоп или связывающий домен.
Примеры консервативных замен заключены в пределах группы основных аминокислот (таких как аргинин, лизин и гистидин), кислых аминокислот (таких как глутаминовая кислота и аспарагиновая кислота), полярных аминокислот (таких как глутамин и аспарагин), гидрофобных аминокислот (таких как лейцин, изолейцин и валин), ароматических аминокислот (таких как фенилаланин, триптофан и тирозин) и малых аминокислот (таких как глицин, аланин, серин, треонин и метионин). Аминокислотные замены, которые обычно не изменяют специфическую активность, известны в данной области и описаны, например, у Н. Neurath and R.L. Hill, 1979, в The Proteins, Academic Press, New York. Наиболее часто происходят следующие замены: Ala/Ser, Val/Ile, Asp/Glu, Thr/Ser, Ala/Gly, Ala/Thr, Ser/Asn, Ala/Val, Ser/Gly, Tyr/Phe, Ala/Pro, Lys/Arg, Asp/Asn, Leu/Ile, Leu/Val, Ala/Glu, Asp/Gly, и наоборот.
Этот термин специально введен для того, чтобы включить пептидные фрагменты НВР, в частности, фрагменты, имеющие хемотаксический эффект, сходный с таковым самого НВР. Более того, НВР, используемые в способах согласно настоящему изобретению, связываются с антагонистами НВР, такими как апротинин и/или моноклональное антитело, вырабатываемое против нативного НВР человека. Нативный НВР человека в его зрелой форме имеет аминокислотную последовательность, представленную в SEQ ID NO:1, и кроме того, является (i) протеолитически неактивным; (ii) содержащимся в азурофильных гранулах полиморфно ядерных лейкоцитов; и (iii) является хемоаттрактантом для моноцитов и/или активирует моноциты.
НВР может быть выделен из тромбоцитов крови, полученных из крови человека с применением способов, описанных в патенте США №5814602. Более конкретно, белок получен путем фракционирования экстракта тромбоцитов крови. Для этой цели применяется колоночная хроматография с использованием гепарин-сефарозы. Такой метод хроматографии, включающий в себя элюирование с колонки в градиенте NaCI от 0,5 М вплоть до 3 М, через которую сначала был пропущен экстракт тромбоцитов крови, дает в результате такой элюции два пика. Первый пик в области 1,2 М NaCI можно было зарегистрировать при 280 нм в виде широкого протеинового пика, который представляет собой известный сам по себе тромбоцитарный фактор (PF<). В области 1,8 М NaCI количество протеина ниже предела регистрации для используемой системы, но фракции в этой области обладают ангиогенной активностью. Активные фракции дополнительно очищали, используя микропробу, с помощью ВЭЖХ с обратимой фазой на колонке С4, и при 214 нм регистрировали полностью очищенный протеиновый пик; этот протеин оказался идентичным присутствующим НВР, будь то НВР свиного или человеческого типов, в зависимости от типа используемых тромбоцитов.
Если НВР используется для обнаружения антагонистов НВР, предпочтительно, чтобы он был получен методом рекомбинантной ДНК, как указано ниже. Последовательность нуклеиновой кислоты, кодирующая НВР, может быть получена искусственным путем с помощью установленных стандартных методов, например, фосфоамидитного метода, описанного S.L Beaucage and M.H. Caruthers, 1981, Tetrahedron Letters 22:1859-1869, или метода, описанного Matthes et al., 1984, EMBO Journal 3: 801-805. Согласно фосфоамидитному методу, олигонуклеотиды были синтезированы, например, в автоматическом ДНК-синтезаторе, очищены, гибридизованы, лигированы и клонированы в соответствующие векторы.
Методы, используемые для выделения или клонирования последовательности нуклеиновой кислоты, кодирующей гепарин-связывающий белок, используемый в способе согласно настоящему изобретению, хорошо известны в данной области и включают в себя выделение из геномной ДНК, получение из кДНК или их комбинацию. Клонирование последовательностей нуклеиновой кислоты согласно настоящему изобретению из таких геномных ДНК может быть реализовано, например, с использованием хорошо известной полимеразно-цепьевой реакции (PCR) или скрининга библиотек экспрессии антител для регистрации клонированных фрагментов ДНК с частичными структурными свойствами (см., например, Innis et al., 1990, A Guide to Methods and Application, Academic Press, New York). Можно использовать и другие способы амплификации нуклеиновой кислоты, такие как лигазно-цепьевая реакция (LCR), лигированная активированная транскрипция (LAT) и амплификация на основе последовательности нуклеиновой кислоты (NASBA).
Затем последовательность нуклеиновой кислоты включается в рекомбинантный вектор экспрессии, который может представлять собой любой вектор, который будет удобен для проведения методов рекомбинантной ДНК. Выбор вектора часто зависит от клетки-хозяина, в которую он будет включен. Так, вектор может быть автономно реплицирующимся вектором, т.е. вектором, который существует в виде внехромосомного вещества, и репликация которого не зависит от хромосомной репликации, например, плазмида. Альтернативно, в качестве вектора может быть выбран такой вектор, который при введении его в клетку-хозяина интегрируется в хозяйский геном и реплицируется вместе с хромосомой(ами), в которую он интегрировался.
Последовательность нуклеиновой кислоты SEQ ID NO:2, кодирующая последовательность SEQ ID NO:1, может быть оперативно связана с нуклеиновой кислотой, кодирующей гетерологичную про- и/или сигнальную последовательность. Альтернативно, нуклеиновая кислота, кодирующая SEQ ID NOS: 4 (сигнальная последовательность + зрелый HBF) и 6 (сигнальная последовательность + про-последовательность + зрелый НВР), SEQ ID NOS: 3 и 5, соответственно, могут быть включены в рекомбинантный вектор.



Последовательность нуклеиновой кислоты, кодирующая НВР, может быть также оперативно связана с соответствующим терминатором, таким как терминатор гормона роста человека (Palmiter et al., op. cit.) Вектор может дополнительно содержать другие элементы. Последние включают в себя сигналы полиаденилирования (например, из SV 40 или из области 5 Elb аденовируса), транскрипционные энхансерные последовательности (например, энхансер SV 40) и трансляционные энхансерные последовательности (например, последовательности, кодирующие РНК аденовируса VA). Рекомбинантный экспрессирующий вектор может также содержать последовательность ДНК, позволяющую вектору реплицироваться в клетке соответствующего хозяина. Вектор может также включать в себя селективный маркер, например ген, продукт которого восполняет дефект в клетке-хозяине, такой как ген, кодирующий дигидрофолатредуктазу (DHFR), или же такой ген, который придает резистентность к лекарству, например, неомицину, генетицину, ампициллину или гигромицину.
В особом аспекте НВР может продуцироваться культивируемыми хозяйскими клетками, содержащими последовательность ДНК, кодирующую зрелый НВР, которому предшествует N-концевая добавка в соответствующей культуральной среде в условиях, допускащих экспрессию НВР, в таком случае НВР получают из культуральной среды в виде НВР, содержащего дополнительный вставочный материал на N-конце.
N-концевая добавка может представлять собой последовательность примерно от 5 до 25 аминокислотных остатков, в частности примерно от 8 до 15 аминокислотных остатков. Природа аминокислотных остатков в N-концевой последовательности не считается критической.
Для того чтобы облегчить получение зрелого НВР, обычно считается предпочтительным, чтобы последовательность ДНК, кодирующая N-концевую добавку, включала в себя последовательность ДНК, кодирующую сайт, подверженный протеолитическому расщеплению, локализованный между последовательностью ДНК, кодирующей N-концевую добавку, и последовательностью ДНК, кодирующей зрелый НВР. Примерами соответствующих сайтов протеолитического расщепления являются сайт расщепления энтерокиназой с аминокислотной последовательностью, сайт расщепления фактором Ха с аминокислотной последовательностью.
Альтернативно, хозяйские клетки, содержащие ДНК, кодирующую зрелый НВР, предваряемый сигнальной последовательностью, могут быть культивированы в соответствующей культуральной среде в условиях, допускающих экспрессию НВР, и результирующий НВР получают из культуральной среды в виде зрелого НВР.
Процедура, используемая для лигирования последовательностей нуклеиновых кислот, кодирующих НВР или НВР с N-концевой добавкой, Дпро-НВР (сигнал + зрелый НВР), промотора и терминатора, соответственно, и для включения их в соответствующие векторы, содержащие информацию, необходимую для репликации, хорошо известны специалистам в данной области (ср., например, Sambrook et al., цитир).
Хозяйская клетка(и), в которую вводится вектор экспрессии, может быть любой клеткой(ами), которая способна продуцировать НВР и предпочтительно является эукариотической клеткой(ами), такой как клетка(и) беспозвоночных (насекомых) или клетка(и) позвоночных, например, клетка(и) млекопитающих, в частности, клетка(и) насекомых и клетка(и) млекопитающих. В предпочтительном аспекте, клетка млекопитающего представляет собой клетку, которую можно культивировать в анаэробных условиях после трансфекции нуклеиновой кислотой, кодирующей НВР млекопитающего. В более предпочтительном аспекте, клетка млекопитающего представляет собой трансформированную аденовирусом клетку(и) или клетку(и), полученную из эмбриона. В данном контексте, клетка, полученная из эмбриональной клетки, представляет собой клетку(и), полученную из первичной культуры эмбриональных клеток, или клетку(и), полученную из клеточной линии, первоначально пересеянную из первичной культуры эмбриональных клеток. Примером такой трансформированной аденовирусом клетки или клетки, полученной из эмбриона, является клетка(и) эмбриональной почки человека (НЕК), в частности, клетка НЕК 293. Среди клеток насекомого такими клетками могут быть, например, клетки Lepidoptera или Drosophila.
Методы трансфекции клеток млекопитающих и экспрессии последовательностей ДНК, введенных в эти клетки, описаны, например, у Kaufman and Sharp, 1982, J. Mol. Biol. 159:601-621; Southern and Berg, 1982, J. Mol. Appl. Genet. 1:327-341; Loyter et al., 1982, Proc. Natl. Acad. Sci. USA 79:422-426; Wigler et al., 1978, Cell 14:725; Corsaro and Pearson, 1981, Somatic Cell Genetics 7:603, Graham and van der Eb, 1973, Virology 52:456; Fraley et al., 1980, JBC 225:10431; Capecchi, 1980, Cell 22:479; Wiberg et al., 1983, NAR 11:7287; and Neumann et al., 1982, EMBO J. 1:841-845. Клетки насекомого могут быть соответствующим образом трансфицированы бакуловирусным вектором, как описано в патенте США №4745051.
Используемая для культивирования клеток среда может быть любой из обычно используемых сред, подходящих для роста клеток млекопитающих, таких как среда с содержанием сыворотки или бессывороточная среда, содержащая соответствующие добавки, или среда, подходящая для роста клеток млекопитающих. Необходимые среды можно приобрести в торговых фирмах или же приготовить самим в соответствии с опубликованными прописями (например, в каталогах Американской Коллекции Типовых Культур). Затем клетки подвергаются скринингу на резистентность к антибиотикам. После этого отобранные клоны подвергают анализу на определение активности НВР с использованием известных в данной области методов, таких как определение хемотаксической активности и анализ высвобождения цитокинов из моноцитов (см., например, Rasmussen et al., 1996, FEBS Lett. 390:109-112).
Продуцируемый клетками НВР может затем быть получен из культуральной среды с помощью обычных методов, включая отделение хозяйских клеток от среды путем центрифугирования или фильтрации, осаждение белковых компонентов супернатанта или фильтрата с помощью соли, например, сульфата аммония, очистку различными хроматографическими методами, например, методом ионообменной хроматографии, аффинной хроматографии и т.п.
Если НВР имеет N-концевую добавку, после его получения из культуральной среды N-концевая добавка НВР может быть успешно отщеплена под действием подходящей для этого протеиназы; и в результате получается зрелый (и активный) НВР. Примеры соответствующих ферментов включают в себя, не ограничиваясь ими, энтерокиназу и фактор Ха.
АНТАГОНИСТЫ НВР
Антагонисты НВР могут быть либо поликлональными антителами против НВР, либо моноклональными антителами против НВР. Для получения поликлональных антител против НВР могут быть использованы следующие процедуры. Для получения антител можно иммунизировать животных разных типов, используя их в качестве хозяина, путем инъецирования им НВР, предпочтительно НВР человека, включая, но не ограничиваясь ими, кроликов, мышей, крыс, овец, коз и т.д. В одном из аспектов, НВР может быть конъюгирован с иммуногенным носителем, например, бычьим сывороточным альбумином (BSA) или гемоцианином моллюска keyhole limpet (KLH). Для усиления иммунного ответа могут быть использованы различные адъюванты, в зависимости от видовой принадлежности хозяина, включая, но не ограничиваясь ими, адъювант Фрейнда (полный и неполный), минеральные гели, такие как гидроокись алюминия, поверхностно-активные вещества, такие как лизолецитин, плюроновые полиспирты, полианионы, пептиды, масляные эмульсии, гемоцианины моллюска keyhole limpet, динитрофенол и потенциально полезные адъюванты человека, такие как БЦЖ (бацилла Calmette-Guerin) и Corynebacterium parvum.
Для получения моноклональных антител, которые связываются по меньшей мере с одним эпитопом на НВР, в частности, НВР человека, в котором указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина, может быть использован следующий метод. В предпочтительном аспекте, моноклональное антитело представляет собой моноклональное антитело человека. Может быть использована любая технология, которая обеспечивает продукцию молекул антитела в культуре в результате длительного культивирования клеточных линий. Такие методы включают в себя, не ограничиваясь ими, гибридомную технологию, первоначально развиваемую Kohler и Milstein (1975, Nature 256:495-497), а также триомную технологию, технику В-клеточных гибридом человека (Kozbor et al., 1983, Immunology Today 4:72) и технологию EBV-гибридом для продукции моноклональных антител (Cole et al., 1985, in Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96). В дополнительном аспекте данного изобретения, с помощью современных технологий можно добиться продукции моноклональных антител у трансгенных животных (PCT/US90/02545; Green, 1999, J. Immunol. Methods 231:11-23 and US. Pat. Nos. 5625126 and 5633425). Согласно изобретению человеческие антитела могут быть использованы и могут быть получены с помощью человеческих гибридом (Cote et al., 1983, Proc. Natl. Acad. Sci. USA 80:2026-2030) или же путем трансформации В-клеток человека вирусом EBV in vitro (Cole et al., 1985, in Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, pp. 77-96).
Согласно изобретению имеющиеся технологии для получения одноцепочечных антител (US Pat. No. 4946778) можно адаптировать таким образом, чтобы наладить получение НВР-специфических одноцепочечных антител. В дополнительном аспекте данного изобретения используются технологии, в которых описан способ конструирования Fab-экспрессирующих библиотек (Huse et al., 1989, Science 246:1275-1281), чтобы иметь возможность быстро и легко идентифицировать Fab-фрагменты моноклональных антител требуемой специфичности в отношении НВР.
Фрагменты антител, которые содержат идиотип молекулы антитела, можно получить с помощью известных технологий. Например, такие фрагменты включают в себя, не ограничиваясь ими, фрагмент F(ab').sub.2, который может быть получен путем расщепления молекулы антитела под действием пепсина; Fab'-фрагменты, которые могут быть получены в результате восстановления дисульфидных мостиков F(ab').sub.2-фрагмента, и Fab-фрагменты, которые могут быть получены в результате обработки молекулы антитела папаином и восстанавливающим агентом.
При получении антител скрининг на требуемое антитело может быть произведен с помощью известных в данной области технологий, таких как радиоиммунный анализ, ELISA (иммуноферментный анализ), "сэндвич"-иммунный анализ, иммунорадиометрический анализ, реакции диффузной преципитации в геле, иммунодиффузионный анализ, иммунный анализ in situ (например, с использованием коллоидного золота, ферментных или радиоизотопных меток), вестерн-блот-анализ, реакции преципитации, анализ агглютинации (например, анализ гель-агглютинации, анализ гемагглютинации), анализ связывания комплемента, иммунофлуоресцентный анализ, анализ на протеин А и анализ методом иммуноэлектрофореза и т.д. В одном из аспектов связывание антитела определяется путем регистрации метки на первичном антителе. В другом аспекте, первичное антитело определяется путем регистрации связывания вторичного антитела или реагента с первичным антителом. Еще в одном аспекте, вторичное антитело связано с меткой. В данной области существует много известных способов регистрации связывания с помощью иммунного анализа, и все они находятся в рамках объема данного изобретения.
Альтернативно, библиотека анти-НВР-антител может быть получена с помощью технологии фагового распределения (см., например, Koscielska et al., 1998, Acta Biochimica Polinica 45:705-720). В результате скрининга этой библиотеки можно отобрать антитела, которые наиболее прочно связываются с НВР, и использовать их в качестве антагонистов НВР.
Антагонистом НВР может также быть полипептид, содержащий домен ингибитора сериновой протеазы типа ингибитора Кунитца, который связывается с НВР. Пептид, содержащий домены ингибитора сериновой протеазы типа ингибитора Кунитца, обычно содержит 60 остатков и шесть специфически пространственно расположенных цистеинов, которые образуют дисульфидные связи. В особом аспекте данного изобретения, если НВР является человеческим НВР, ингибитором типа ингибитора Кунитца может быть апротинин, известный также как BPTI или его аналог, который связывается с НВР. В предпочтительном аспекте, аналог апротинина содержит мутации в положениях 12-19 и 34-39, особенно предпочтительными являются мутации в положениях 15-19. Другие аналоги, которые также могут быть использованы, описаны в патентах США №№5162498, 5316923, 5395922, 5514585, 5510249, 5591603, 5618915, 5621074, 5673090, 5618696 и 5576294. В наиболее специфическом аспекте, мутации представляют собой А16Н, К15А, I18M, I19S, R17T.
Крупные библиотеки (107-109) полипептидов, включая домены типа домена Кунитца, можно получить, используя различные подходы. Например, метод error-prone - ПЦР (Saiki et al., 1988, Science 293:487-491) может быть применен к каждому домену типа домена Кунитца. В результате такой процедуры будут случайным образом включаться ошибочные нуклеотиды в ген каждого соответствующего домена типа домена Кунитца в процессе реакции ПЦР в условиях, при которых Тaq-полимераза на стадии амплификации включает ошибочные нуклеотиды. Библиотека может быть распределена на фаговых частицах с целью выбора полипептидов, наиболее прочно связывающихся с НВР.
Альтернативно, при использовании вырожденных олигонуклеотидов мутагенезу можно подвергнуть специфические петли (Schier et al., 1996, Gene 169:147-153). Искусственные случайные олигонуклеотиды включаются в домен типа домена Кунитца в процессе реакции ПЦР, и в результате нарабатывается библиотека вариантов для отдельной петли.
В другом аспекте, библиотеки могут вырабатываться путем перетасовки ДНК (Stemmer et al., 1994, Nature 370:389-391). Этот метод может быть использован для рекомбинации между гомологичными доменами типа доменов Кунитца. Вкратце, ДНК, кодирующая каждый из вариантов, смешивается и фрагментируется до малых фрагментов. Затем эти фрагменты вновь объединяются в процессе регулярной реакции ПЦР. В процессе такой реакции малые фрагменты будут объединяться друг с другом, поскольку они комплементарны друг другу, и в результате происходит рекомбинация включенных генов.
МЕТОДЫ РЕГИСТРАЦИИ
Антагонисты гепарин-связывающего белка (НВР) могут быть зарегистрированы путем определения того, связывается ли с НВР вещество, которое по предположению является антагонистом НВР, в результате (а) инкубации НВР с первым веществом, которое взаимодействует с НВР, и со вторым веществом, которое, по предположению, является антагонистом НВР, и (b) обнаружения любого эффекта указанного второго вещества, которое по предположению является антагонистом взаимодействия с НВР, с указанным первым веществом, причем ослабленное взаимодействие НВР с первым веществом указывает на то, что второе вещество является антагонистом НВР.
НВР или первое вещество могут быть связаны с меткой. Метку предпочтительно выбирают из группы, состоящей из ферментов, окрашенных или флуоресцентных веществ, радиоактивных изотопов и комплексобразующих агентов.
Примерами ферментов, применимых в качестве метящих веществ, являются пероксидазы (такие как пероксидаза хрена), фосфатазы (такие как кислая или щелочная фосфатаза), В-галактозидаза, уреаза, глюкозооксидаза, карбоангидраза, ацетилхолинэстераза, глюкоамилаза, лизоцим, малатдегидрогеназа, глюкозо-6-фосфатдегидрогеназа, b-глюкозидаза, протеиназы, пируватдекарбоксилаза, естеразы, люцифераза и т.д.
Ферменты сами по себе не определяются, однако они должны быть объединены с субстратом, чтобы катализировать реакцию, конечный продукт которой в результате и определяется. Примеры субстратов, которые могут быть использованы в способе согласно изобретению, включают в себя гидропероксид/тетраметилбензидин или хлорнафтол или о-фенилендиамин или 3-(пара-гидроксифенил) пропионовую кислоту или люминол, индоксилфосфат, пара-нитрофенилфосфат, нитрофенилгалактозу, 4-метилумбеллиферил-0-галактопиранозид или люциферин.
Альтернативно, меченое вещество может включать в себя окрашенное или флуоресцентное вещество, включая частички золота, окрашенные или флуоресцентные частички латекса, частички красителя, флуоресцеин, фикоэритрин или фикоцианин.
Радиоактивные изотопы, которые могут быть использованы в целях данного изобретения, могут быть выбраны из 125I, 131I, 111In 3H, 32P, 14С и 35S. Радиоактивность, излучаемая этими изотопами, может быть измерена с помощью гемма-счетчика или же сцинтилляционного счетчика способом, известным в данной области.
Комплексобразующие агенты, которые могут быть использованы в целях данного изобретения, могут быть выбраны из биотина (который образует комплексы с авидином или стрептавидином), авидина (который образует комплекс с биотином), протеина А (который образует комплекс с иммуноглобулинами) и лектинов (которые образуют комплекс с рецепторами углеводов). Поскольку комплекс напрямую не определяется, необходимо пометить вещество, с которым комплекс-образующие агенты образуют комплекс. Мечение может проводиться любым из меченых веществ, указанных выше для мечения фермента.
В особом аспекте связывание антагониста НВР с НВР может быть зарегистрировано, например, с помощью технологии SPA, позаимствованной из Amersham Pharmacia Biotech. Вкратце, биотинилированные плазматические мембраны, полученные из эндотелиальных клеток пупочной вены (HUVEC), связаны со стрептавидин-конъюгированными гранулами PVT SPA. Такие гранулы можно получить в компании Amersham Pharmacia Biotech. Меченый кининоген человека (например, 3H, 125I) добавляют к мембранам до тех пор, пока связывающие сайты кининогена не окажутся насыщенными. Добавляют НВР в увеличивающихся концентрациях, чтобы переместить меченый кининоген, и измеряют уменьшение включенной метки. Получают стандартную кривую такого снижения. Тестируют антагонист НВР на его способность ингибировать НВР-опосредованное снижение радиоактивного кининогена на плазматических мембранах. В частности, НВР инкубируют в присутствии различных концентраций вплоть до молярных избытков антагонистов НВР, и определяют их способность ингибировать НВР-опосредованное снижение радиоактивного кининогена на плазматических мембранах.
Альтернативно, антагонисты НВР могут быть обнаружены путем культивирования эндотелиальных клеток в присутствии НВР и в присутствии или в отсутствие вещества, которое предположительно является антагонистом. В особом аспекте, антагонист представляет собой моноклональное антитело, которое связывается, по меньшей мере, с одним эпитопом НВР, предпочтительно НВР человека, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина.
В предпочтительном аспекте моноклональное антитело представляет собой моноклональное антитело человека. В одном из аспектов антитело и НВР преинкубируются друг с другом перед инкубацией с эндотелиальными клетками в течение 10 мин при 37°С. Влияние вещества на проницаемость эндотелиальных клеток определяют путем измерения трансэндотелиального электрического сопротивления или же путем определения клиренса альбумина, как описано в примерах, ниже. Снижение проницаемости будет указывать на то, что вещество действует как антагонист НВР.
В особом аспекте связывание моноклонального антитела по меньшей мере с одним эпитопом НВР, где указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина, может быть определено путем инкубирования антитела с НВР и прекалликреин-Н-кининогеновым комплексом. Как будет подробно описано в дальнейшем в примерах, НВР связывается с прекалликреин-Н-кининогеновым комплексом, таким образом стимулируя высвобождение брадикинина. Моноклональное антитело, которое связывается, по меньшей мере, с одним эпитопом НВР, должно уменьшать или предотвращать высвобождение брадикинина. Высвобождение брадикинина может быть измерено с помощью методов, известных в данной области, например, с помощью иммунного анализа.
В одном из аспектов, покрывающий буфер содержит 0,05% Tween, 15,9 мМ бикарбонат натрия, 35 мМ карбонат натрия, рН 9,6. Добавляется Н-кининоген и проводится инкубирование в течение ночи при 4°С. Покрывающий буфер удаляют, и планшет промывают Tris-буфером, рН 7,4, содержащим примерно 50 мкМ цинка. Затем добавляют прекалликреин, и смесь инкубируют в течение 1 часа при комнатной температуре. После этого добавляют НВР в Tris-буфере, и последовательно, примерно с 10-минутными интервалами приблизительно в течение 1 час, начиная примерно через 5 мин после добавления НВР, образцы извлекаются, и в них определяется высвобождение брадикинина. В одном из аспектов высвобождение брадикинина может определяться с помощью коммерчески доступного набора для метода ELISA (MARKIT-M Bradykin, Dainippon Pharmaceutical Co., Ltd.). В таком методе брадикинину в пробе и меченному пероксидазой брадикинину (ВК) дают возможность конкурентно реагировать с анти-ВК-антителом (кроличьим), связанным с антителом против кроличьего IgG (козьим), которым покрыты полосатые лунки микропланшета. Концентрация брадикинина определяется по ферментативной активности меченного пероксидазой брадикинина, связанного с антибрадикининовым антителом.
Альтернативно, прекалликреин и Н-кининоген могут быть добавлены в эквимолярных количествах к монослою эндотелиальных клеток и проинкубированы в течение 1 час при 37°С. После этого добавляют НВР, и последовательно, примерно с 10-минутными интервалами приблизительно в течение 1 час, начиная примерно через 5 мин после добавления НВР, образцы извлекаются, и в них определяется высвобождение брадикинина с использованием метода, описанного выше.
Еще в одном аспекте прекалликреин и Н-кининоген добавляют в эквимолярных количествах к раствору плазмы, содержащему ингибитор АСЕ (ангиотензинпревращающий фермент), и инкубируют в течение 1 час при 37°С. После этого добавляют НВР, и последовательно, примерно с 10-минутными интервалами приблизительно в течение 1 час, начиная примерно через 5 мин после добавления НВР, образцы извлекаются, и в них определяют высвобождение брадикинина с использованием метода, описанного выше.
МЕТОДЫ ЛЕЧЕНИЯ
Композиция согласно изобретению содержит часть водного раствора. Предпочтительно, чтобы этот раствор был физиологически приемлемым, так, чтобы при его добавлении для доставки пациенту требуемой конструкции этот раствор не вызывал какого-либо неблагоприятного влияния на баланс электролитов и волюмометрический баланс у пациента. Таким образом, водная среда для композиции может включать в себя, например, нормальный физиологический раствор (0,9% NaCI, 0,15 М), рН 7-7,4 или другие фармацевтически приемлемые соли. Полезные растворы можно приготовить любым из методов, хорошо известных в фармацевтической области, описываемых, например, в "Remington's Pharmaceutical Sciences", (Gennaro, A., ed.). Mack Pub., 1990.
Концентрация антагониста НВР может широко варьировать, т.е. примерно менее чем от 0,5%, например, от 1%, до 15-20% по массе. Одноразовая доза композиции обычно может содержать примерно от 10 мг до 1 г антагониста НВР. Терапевтически эффективные дозы можно определять, используя либо методы in vitro, либо методы in vivo.
Антагонист НВР назначается местно, подкожно или в виде внутривенной инъекции. Дозы прописываются лечащим врачом в соответствии с конкретным состоянием и с учетом особенностей конкретного пациента, подвергаемого лечению. Тщательно подбираются дозировки и частота введения и приводятся в соответствие с показателями, определяемыми лечащим врачом при осмотре. Предпочтительным путем введения может быть, например, инъекционный внутрибрюшинный. Внутривенные внутрибрюшинные инъекции антагониста НВР могут вводиться каждые 24 час в диапазоне от 0,1 до 100 мг, в частности от 0,5 до 50 мг, в частности от 1 до 25 мг на кг веса тела. Эту дозу можно вводить в 1-4 приема за 24 час или же вводить непрерывно через катетер. Альтернативно, введения могут производиться путем непрерывной внутривенной инфузии в процессе хирургического вмешательства, а затем в течение 1-4 дней после операции.
Антагонист НВР будет вводиться только тем пациентам, которые продуцируют НВР, который связывается с антагонистом НВР. Для определения того, продуцируется ли у данного пациента такой НВР, из тромбоцитов периферической крови больного нужно выделить НВР с помощью методов, описанных выше. Связывание НВР с антагонистом НВР (например, моноклональным антителом) или же влияние НВР на проницаемость эндотелиальных клеток может быть оценено с помощью описанных выше способов и сравнено с действием нативного НВР. Альтернативно, действие НВР на сократимость эндотелиальных клеток или фибробластов или на агрегацию моноцитов в присутствии или в отсутствие антагониста НВР можно также определить с помощью методов, описанных в WO 93/05396. Сниженная степень клеточного сокращения или агрегации указывает на взаимодействие между НВР и антагонистом НВР. Набор для тестирования, который может быть использован для определения того, продуцируется ли у пациента такой НВР, включает в себя (а) антагонист НВР; (b) НВР или клетки, продуцирующие НВР, и (с) вещество, ткань, клетки или их компоненты, которые взаимодействуют с НВР.
В особом аспекте НВР или клетки или ткани, продуцирующие НВР, выделяются из организма пациента. НВР или клетки или ткани, продуцирующие указанный НВР, инкубируются с эндотелиальными клетками, и действие на проницаемость определяется в присутствии или в отсутствие моноклонального антитела, которое связывается по меньшей мере с одним эпитопом НВР, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина. Этот эффект сравним с эффектом нативного НВР человека. В особом аспекте нативный НВР человека может быть рекомбинантным НВР, который связывается с таким моноклональным антителом. Альтернативно, НВР или клетки или ткани, продуцирующие НВР, инкубируются в присутствии или в отсутствие прекалликреин-Н-кининогенового комплекса и в присутствии моноклонального антитела. Затем определяют влияние моноклонального антитела на выделение брадикинина. Уменьшенное высвобождение брадикинина в присутствии антитела свидетельствует о продукции у пациента НВР, который связывается с антителом. В подобном случае тест-набор будет содержать (а) моноклональное антитело, которое связывается, по меньшей мере, с одним эпитопом на НВР, причем указанный эпитоп связывается с прекалликреин-Н-кининогеновым комплексом и активирует высвобождение брадикинина; (b) нативный человеческий НВР и (с) прекалликреин-Н-кининогеновый комплекс, связанный с твердым носителем.
ПРИМЕРЫ
Пример 1
Активированные нейтрофилы запускают изменения в функции эндотелиального барьера путем сигналирования через В2-интегрины
МАТЕРИАЛЫ И МЕТОДЫ
Реагенты
Среда 199, RPMI-1640 (содержащая L-глутамин), эмбриональная бычья сыворотка (FBS), трипсин-EDTA, забуференный фосфатом физиологический раствор (PBS) и сбалансированный раствором Хэнкса физиологический раствор (HBSS) были приобретены в Life Technologies (Gaithersburg, MD, USA). Желатин, коллагеназа, пенициллин, стрептомицин, каталаза, бычий сывороточный альбумин (BSA), N-формил-метионин-лейцил-фенилаланин (fMLP), синий краситель Эванса, гексадецил-триметиламмония бромид и тетраметилбензидин были приобретены в Sigma Chemical Со. (St. Louis/ МО/ USA). Н202 в Е. Merck (Darmstadt, Germany). Биоматрикс I был приобретен в Biomedical Technologies Inc. (Stoughton, MA, USA), a Dextran 70 (Macrodex) и Ficoll-Paque в Pharmacia Biotech AB (Uppsala, Sweden). Основная культуральная среда состояла из смеси 1:1 М-199 и RPMI-1640, обогащенной 20% инактивированной нагреванием FBS, пенициллином (100 ед/мл) и стрептомицином (100 мкг/мл). Моноклональное антитело IB4 против общей β-цепи (CD18) β2-интегринов было предоставлено Dr. Claes Lundberg, Pharmacia & Up John (Uppsala, Sweden) с любезного согласия Dr. Samuel D. Wright, Rockefeller University, mAb 60.1, узнающее αm-цепь интегрина (CD11b), было предоставлено Dr. Manuel Patarroyo, Karolinska Institutet, mAb DREG 200 (анти-L-селектин) подарок Dr. Eugene Butcher, Stanford University, mAb SK11 (анти-L-селектин) было получено в Becton Dickinson (San Jose, CA) и mAb G44-26 (aHTH-CD44) было получено в PharMingen (San Diego, CA). F(ab')2-фрагмент козьего антитела против мышиного IgG был получен из Jackson immuno Research Laboratories, Inc. (West Grove, PA), FITC-конъюгированный F(ab')2-фрагмент кроличьего антитела против мышиного IgG был получен из Dako A/S (Glostrup, Дания).
Эндотелиальные клетки
Эндотелиальные клетки пупочной вены человека (HUVEC) и эндотелиальные клетки аорты быка (ВА2С) выделяли и культивировали в соответствии с описанным выше (Gautam et al. 1998, Brit. J. Pharm. 125:1109-1114). HUVEC или ВАЕС в первом-пятом пассажах снимали путем кратковременной (2 мин) обработки трипсином-EDTA (0,25% трипсин/0,01 % EDTA) и переносили либо на неорганическую мембрану Anopore с размером пор 0,2 мкм, либо на поликарбонатные фильтры с размером пор 3,0 мкм (Tissue Culture inserts, 10 мм; NUNC, Roskilde, Дания). Для ускорения клеточной дифференцировки и усиления прикрепления эндотелиальных клеток (ЭК) фильтры предварительно обрабатывали 50 мкл Biomatrix I (167 мкг/мл) и сушили на воздухе. ЭК высевали при плотности 2×105 клеток на фильтр и инкубировали в культуральной среде при 37°С в увлажняемой атмосфере 5% СО2 в воздухе. ЭК культивировали до образования конфлуэнтных монослоев, что контролировалось ежедневным микроскопическим наблюдением и измерением электрического сопротивления монослоя (Gautam et al. 1998, Brit. J. Pharm. 125:1109-1114).
Измерение барьерной функции ЭК
Для установления индуцированных стимуляцией изменений проницаемости ЭК использовали два различных подхода. Для измерения трансэндотелиального электрического сопротивления (TEER) фильтр-вставку с ЭК переносили в камеру для измерения электрического сопротивления (подробнее см. Gautam et al. 1998, Brit. J. Pharm. 125:1109-1114). Камера (нижний/освещенный компартмент) с фильтром-вставкой (верхний/освещенный компартмент), заполненная, соответственно, 2 мл и 400 мкл культуральной среды, помещена в инкубатор для культивирования клеток. Трансэндотелиальное электрическое сопротивление монослоя ЭК измеряли при 37°С в каждом компартменте с помощью электродов, точно установленных по отношению друг к другу и к монослою. Электрическое сопротивление монослоев ЭК получали путем вычитания сопротивления соответствующего интактного фильтра, покрытого слоем Biomatrix, измеряемого перед засеванием его эндотелиальными клетками.
В независимых экспериментах использовали альбумин, конъюгированный с синим красителем Эванса (ЕВА), в качестве маркера макромолекулярной проницаемости монослоев ЭК. Перед стимуляцией ЭК культуральную среду в верхнем компартменте меняли на среду, содержащую ЕВА (культуральная среда, содержащая 4% BSA, смешанного с синим красителем Эванса в конечной концентрации 0,67 мг/мл). Концентрацию ЕВА в жидких образцах верхнего и нижнего компартментов определяли спектрофотометрически путем измерения поглощения при 620 нм (Titertek Multiskan MCC; Flow Laboratories, Solna, Sweden). Клиренс альбумина вычисляли в соответствии с соотношением: V1=А2×V2×1/А1, где V1 - объем клиренса (т.е. теоретический объем освещенной среды, очищенной от альбумина путем диффузии к аблюминальному компартменту), V2 - аблюминальный объем, а А1 и А2 - поглощение освещенной и неосвещенной среды, соответственно. Базовый клиренс в отсутствие какого-либо стимула, в среднем 0,08±0,03 мкл/мин, вычитали из объема клиренса, полученного в ответ на соответствующий специфический стимул.
Получение и количественное определение ПМЯ
ПМЯ человека выделяли, как было описано (Gautam et al., 1998, Brit. J. Pharm. 125:1109-1114), из обогащенной лейкоцитами плазмы, и ресуспендировали в культуральной среде при концентрации 2-5×107 клеток/мл. Степень чистоты ПМЯ составляет > 98%, и жизнеспособность их, определяемая по исключению трипанового синего, составляет >95%. Перед добавлением к монослоям ЭК, ПМЯ инкубировали в течение 30 мин в присутствии моноклональных антител против молекул клеточной поверхности для последующего перекрестного связывания со вторичным mAb (см. экспериментальный метод).
В некоторых экспериментах экспрессию CD11b/CD18 на клеточной поверхности ПМЯ перед использованием усиливали путем изменений температуры инкубации. ПМЯ инкубировали в присутствии анти-GD18-mАb IB4 при 4°С в течение 10 мин, затем при 37°С в течение 10 мин, после чего вновь при 4°С в течение еще 10 мин, после чего клетки дважды промывали. ПМЯ, которые не инкубировали в присутствии антитела, подвергали обработке наполнителем вместо антитела, в полном соответствии с этой же методикой.
Адгезия ПМЯ к ЭК и трансэндотелиальная миграция количественно оценивались путем анализа ПМЯ-специфического фермента миелопероксидазы (МРО). Вкратце, ПМЯ лизировали в 0.5% гексадецилтриметиламмония бромида и центрифугировали, затем в супернатанте спектрофотометрически определяли ферментативную активность как изменение в поглощении при 650 нм, которое возникает в окислительно-восстановительной реакции Н2O2-тетраметилбензидина, катализируемой под действием МРО (Suzuki et al., 1983, Anal. Biochem. 132:345-352). Активность МРО прикрепившихся ПМЯ и мигрировавших сквозь слой ЭК, соответственно, связана с таковой общего количества ПМЯ, добавленных к монослою ЭК.
Экспериментальные методы
Перед переносом фильтр вносили в камеру для измерения сопротивления (поддерживаемую при 37°С), среду в фильтре-вставке и в камере заменяли свежей культуральной средой (37°С), обогащенной 10 mM Hepes. В некоторых экспериментах монослой ЭК перед стимуляцией 10 мкг/мл гербимицина А инкубировали в течение 15 мин при 37°С и дважды промывали. ПМЯ (2×106), содержащие и не содержащие моноклональные антитела, связанные с молекулами клеточной поверхности, добавляли к верхнему компартменту (отношение ПМЯ:ЭК=10:1) и в течение 10 мин давали им возможность осесть на монослой ЭК. Активацию ПМЯ индуцировали либо с помощью fMLP (10-7 М), добавляемого к нижнему компартменту, либо путем перекрестного связывания связанного с молекулами клеточной поверхности антитела с F(ab')2-фрагментами вторичного козьего антитела против мышиного IgG. Трансэндотелиальное электрическое сопротивление измеряли перед и каждую минуту после начала стимуляции, до тех пор, пока не достигалась фаза выхода на плато величины изменения сопротивления, а затем с 5-минутными интервалами. В тех экспериментах, где изучали проницаемость для альбумина и миграцию ПМЯ сквозь монослой ЭК, среда во вставленном фильтре содержала ЕВА. После определения фильтр-вставку с 5-10-минутными интервалами переносили в новые лунки, содержащие свежую среду. Все стадии инкубации проводили при 37°С. В конце эксперимента нижние лунки центрифугировали при 425хg и при 4°С в течение 20 мин, и в супернатанте определяли содержание ЕВА. Количество ПМЯ, которые мигрировали сквозь монослой, определяли путем анализа активности МРО в осадке, остающемся в лунке. Среду в фильтре-вставке подвергали количественному определению в ней неприкрепившихся ПМЯ, и мембрану фильтра с ЭК извлекали для количественного определения прикрепившейся фракции ПМЯ.
В отдельных экспериментах отмытые ПМЯ, предварительно обработанные анти-СD18-mАb IB4 (6 мкг на 2×106 клеток) в течение 30 мин, инкубировали в культуральной среде в присутствии и в отсутствие козьих анти-мышиных F(ab)2 в течение 10 мин при 37°С. ПМЯ осаждали путем центрифугирования при 300хg в течение 15 мин при комнатной температуре, и бесклеточный супернатант, содержащий продукты секреции ПМЯ, декантировали для применения в экспериментах in vivo (см. ниже) или с монослоями ЭК. В последнем случае среду в фильтре со встроенными ЭК заменяли средой, содержащей продукты секреции ПМЯ, и, как описано выше, определяли ее действие на TEER и на проницаемость для альбумина. В некоторых случаях среду, содержащую продукты секреции ПМЯ, перед использованием подвергали тепловой инкубации (80°С в течение 15 мин).
Определение внутриклеточного [Са2+] и образования f-актина в ЭК
Для определения изменений в содержании внутриклеточного свободного Са2+ использовали Са2+-чувствительную флуоресцентную пробу флуо-3/ДМ (Molecular Probes Europe BV, Leiden, Нидерланды). Конфлуэнтные монослои ЭК на фильтрах инкубировали в течение 30 мин при 37°С в присутствии fluo-3/ДМ (3 мкМ в HBSS, содержащем 2% FCS и 10 mM HEPES), добавляемого как на апикальную, так и на базолатеральную поверхность. Монослои трижды промывали и инкубировали в присутствии свежего HBSS в темноте в течение 20 мин при комнатной температуре для полного гидролиза эфирного красителя. ПМЯ, обработанные анти-CD18- или анти-L-селектиновыми mAb, добавляли к ЭК и, как указывалось выше, индуцировали перекрестное связывание рецептора со вторичными mAb. В отдельных экспериментах для стимуляции ЭК в отсутствие ПМЯ использовали среду, содержащую продукты секреции ПМЯ. Изменения в содержании внутриклеточного [Са2+]i ЭК в присутствии указанных стимулов измеряли в режиме непрерывной регистрации интенсивности флуоресценции с помощью лазерной сканирующей системы конфокального изображения (Insight Plus; Meridian Instruments Inc., Okemon, Michigan).
Для анализа образования f-актина в ЭК в ответ на активацию ПМЯ конфлуэнтные монослои ЭК, выращенные на покрытых Biomatrix полосатых поверхностях, инкубировали в течение 15 мин при 37°С либо в присутствии ПМЯ, подвергнутых перекрестному связыванию с антителом CD18, либо в присутствии среды, содержащей продукты секреции ПМЯ, в полном соответствии с процедурами, описываемыми выше. Контролем служили ЭК, инкубируемые в присутствии ПМЯ, обработанных IB4. Монослои фиксировали в 3,7% формальдегиде в PBS в течение 10 мин при комнатной температуре, дважды промывали и обрабатывали 0,2% Triton Х-100 в PBS (1 мин при 4°С). Клетки дважды промывали и окрашивали для визуализации актиновых филаментов FITC-конъюгированным фаллоидином в течение 20 мин при 37°С. После трех дополнительных промывок эндотелиальные клетки просматривали с помощью лазерной сканирующей системы конфокального изображения (Insight Plus; Meridian Instruments Inc., Okemon, Michigan).
Эксперименты in vivo
С целью изучения влияния продуктов секреции ПМЯ на проницаемость микрососудов in vivo проводили исследование микроциркуляции с помощью прижизненной внутриклеточной микроскопии в защечном мешке хомячка. Как было подробно описано ранее (Raud & Lindbom, 1994, Immunology of the Microcirculation, Academic Press, 1994, chapter 7), левый защечный мешок анестезированных золотистых сирийских хомячков выворачивали и подготавливали для микроскопического наблюдения в условиях постоянной гиперфузии бикарбонатным буфером при поддержании физиологических уровней температуры, рН и давления. FITC-конъюгированный декстран (М.м. 150 000), инъецированный внутривенно (250 мг/кг веса тела), использовали для визуализации изменений сосудистой проницаемости в отношении макромолекул. Микрососудистую систему в защечном мешке хомячка наблюдали во флуоресцентном свете при низком разрешении в микроскопе Leitz Orthoplan, и видеоизображение записывали. После контрольных видеозаписей гиперфузию прекращали, и на защечный мешок накладывали 1 мл среды, содержащей продукты секреции из ПМЯ, связанных с CD18, при 37°С. В параллельных экспериментах использовали среду ПМЯ, обработанных анти-CDlS, не подвергнутую перекрестному связыванию.
РЕЗУЛЬТАТЫ
ПМЯ (2×105), предварительно обработанные анти-СD18-mАb (IB4), добавляли к фильтру со встроенными ЭК (светлая поверхность ЭК/верхний компартмент камеры для измерения сопротивления) и оставляли на 10 мин. Перекрестное связывание CD18 со вторичным козьим F(ab)2 антимышиного антитела вызывало значительное снижение TEER, которое проявлялось в течение 1 мин, достигая своего максимума (36+13 от контроля) через 15 мин и оставаясь на этом уровне в течение всего периода наблюдения (фиг.1). Ни при обработке полиморфно-ядерных лейкоцитов одним только IB4, ни при их обработке вторичным mAb в отсутствие IB4 никаких изменений в TEER не происходило.
В дополнительных экспериментах изучали влияние способности перекрестного связывания CD18 на усиление макромолекулярного потока и миграции ПМЯ сквозь монослой ЭК. ПМЯ, предварительно обработанные насыщающей концентрацией IB4, добавляли к фильтру со встроенными ЭК и оставляли на 10 мин в среде, содержащей ЕВА. Перекрестное связывание CD18 сходным образом индуцировалось козьим F(ab)2 антимышиного антитела, и фильтры-вставки, содержащие ПМЯ/ЕВА, с регулярными интервалами переносились в новые лунки, содержащие свежую среду. В отсутствие вторичного антитела имеет место минутный и постоянный клиренс альбумина, 0,08±0,03 мкл·мин-1 (среднее ± SD, n=13), который не меняется со временем и который в точности совпадает со скоростью спонтанного клиренса, обнаруживаемого в фильтрах-вставках, содержащих необработанные ПМЯ. И наоборот, активация ПМЯ под действием перекрестного связывания CD18 вызывала увеличение клиренса альбумина, которое со временем прогрессировало (фиг.1). В чистом виде увеличение клиренса альбумина возрастало от 0,5±0,1 мкл в течение первого пятиминутного периода и достигало 20,0±4,5 мкл (среднее ± SD, n=10) через 60 мин. В отличие от возрастающего клиренса альбумина, перекрестное связывание CD18 не вызывало регистрируемой адгезии ПМЯ на ЭК или трансэндотелиальной миграции (фиг.1). Анализ активности МРО в различных компартментах через 60 мин после перекрестного связывания CD18 показал, что 91+6% (среднее ± SD, n=9) добавленных ПМЯ обнаруживаются в среде, собранной из верхнего компартмента, тогда как никакой заметной активности МРО не обнаружено в ячейке ЭК/фильтр или в среде нижнего компартмента. Непосредственное микроскопическое наблюдение прозрачных фильтров, промытых в конце эксперимента, подтвердило отсутствие ПМЯ, прикрепившихся к монослою ЭК.
Монослои эндотелиальных клеток метили Са2+-чувствительным флуорофором fluo-3, и индуцированные стимулом изменения содержания [Са2+]i в ЭК наблюдали методом лазерной сканирующей микроскопии. Добавление к монослою ЭК полиморфно-ядерных лейкоцитов, обработанных анти-CD18, не вызывало изменений [Са2+]i в ЭК. Однако перекрестное связывание CD18 со вторичным mAb приводило к быстрому возрастанию содержания свободного [Ca2+]i в цитозоле ЭК, которое достигало своего пика примерно через 100 сек, а затем медленно восстанавливалось. При перекрестном связывании таким же образом антитела L-селектином не удавалось индуцировать каких-либо изменений [Са2+]i в ЭК, тогда как после стимуляции необработанных ПМЯ под действием fMLP наблюдали Са2+-ответ, сходный с таковым, обнаруживаемым при перекрестном связывании CD18 (ср. Gautam et al., 1998, Brit. J. Pharmacol. 125:1109-1114). Причинная связь между изменениями активности внутриклеточного Са2+ и функциональным ответом ЭК на активацию ПМЯ очевиден из экспериментов, в которых эндотелиальные клетки предварительно обрабатывали кальций-хелатирующим агентом ВАРТА AM. Такая обработка полностью ингибировала как повышение содержания внутриклеточного свободного Са2+ в ЭК, так и изменения TEER в ответ на перекрестное связывание CD18.
В дальнейших экспериментах монослои ЭК инкубировали с ПМЯ, а затем проводили окрашивание FITC-конъюгированным фаллоидином с целью выявления актиновых филаментов. С помощью конфокальной лазерной сканирующей микроскопии показали, что в эндотелиальных клетках, инкубируемых с IB4-обработанными ПМЯ в отсутствие вторичного антитела, появлялось несколько волокон напряжения и тонкая полоса актина вдоль маргинальной зоны клеток при непосредственном соприкосновении с соседней клеткой. С другой стороны, эндотелиальные клетки, инкубируемые с ПМЯ, подвергнутыми перекрестному связыванию с CD18, обнаруживали значительное увеличение содержания f-актина, а также количества и плотности расположения волокон напряжения.
Для исследования фактора(ов), секретируемого ПМЯ, который регулирует ответ, обязательно приводящий к увеличению проницаемости ЭК, ПМЯ (2×106), предварительно обработанные анти-СD18-mАb IB4 и промытые с целью удаления несвязавшегося антитела, инкубировали в течение 30 мин в присутствии козьего F(ab')2-фрагмента антимышиного антитела, а затем центрифугировали в мягком режиме. Добавление бесклеточного супернатанта к монослою ЭК вызывало снижение TEER и усиление клиренса альбумина, причем оба этих изменения по величине и временной зависимости совпадали с таковыми в эксперименте, при котором к ЭК добавляли ПМЯ после перекрестного связывания CD8. Супернатант также вызывал изменения [Са2+]i и содержание и распределение f-актина в ЭК. Более того, местное нанесение супернатанта, содержащего продукты секреции ПМЯ, на защечный мешок хомячка in vivo вызывало быструю утечку плазмы из посткапиллярных и малых вен. Следы утечки наблюдались уже через 1,5 мин после нанесения, и максимальный ответ наблюдался уже примерно через 5 мин. После отмывания ткани буфером утечка постепенно утихала. Такое действие на проницаемость микрососудов утрачивается после тепловой обработки секрета ПМЯ, что указывает на чувствительность фактора(ов), ответственного за увеличение их проницаемости, к тепловому воздействию. Наложение на защечный мешок хомячка супернатанта IB4-обработанных ПМЯ, не подвергаемых перекрестному сязыванию, не вызывало сколько-нибудь заметной утечки плазмы.
ОБСУЖДЕНИЕ
Адгезия полиморфно-ядерных лейкоцитов к эндотелиальной выстилке и рекрутмент внесосудистой ткани при остром воспалении напрямую зависит от функции β2-интегринов. Повышенная проницаемость сосудов для макромолекул вызывается в тесной связи с таким ответом клеток, ведущим к появлению экссудата плазмы и образованию отека. Результаты описанных выше экспериментов доказывают причинную связь между трансмембранным (снаружи внутрь) сигналированием β2-интегринов в активированных ПМЯ и способностью этих клеток индуцировать дисфункцию барьера ЭК. Показано, что индуцируемое антителом лигирование и кластеризация β2-интегринов запускает процесс увеличения проницаемости ЭК, сходный с таковым, наблюдаемым после стимуляции ПМЯ хемоаттрактантом. Подобный межклеточный диалог происходит при отсутствии физического контакта ПМЯ с ЭК, свидетельствуя о том, что явление адгезии ПМЯ и структурной связи как таковой не связано напрямую с передачей сигналов на ЭК, которыми опосредовано повышение проницаемости. Скорее связывание лиганда β2-интегрина инициирует через внутриклеточное сигналирование каскад секреторных явлений в ПМЯ и высвобождение химического посредника, который способствует ПМЯ-зависимому изменению проницаемости ЭК. Далее, показано, что изменение сосудистой проницаемости, вызываемое посредством этого пути, является следствием активных, зависимых от тирозинкиназы, ответов ЭК, участвующих в реорганизации цитоскелета и в обратимом формировании межклеточных гэпов, что указывает на нелитическую в этом отношении функцию ПМЯ-зависимого фактора(ов).
Для более подробного изучения механизмов, с помощью которых активированные ПМЯ могут в этом процессе осуществлять передачу сигналов эндотелиальным клеткам, взаимодействия ПМЯ/ЭК и зависимое от адгезии вовлечение (ангажирование) β2-интегринов стимулировали путем перекрестного связывания CD11/CD18. Путем перекрестного связывания β2-интегринов, в обход индуцированной хемоаттрактантом активации клеток, показали, что ангажирование и кластеризация как таковых рецепторов β2-интегринов путем внутриклеточной передачи сигналов запускает процесс увеличения проницаемости ЭК. Поскольку этот ответ возникал независимо от адгезии ПМЯ на ЭК, передача сигналов между ПМЯ и ЭК не может быть отнесена за счет взаимодействия CD11/CD18 с противорецепторами на поверхности ЭК, а скорее должна достигаться путем высвобождения молекул-посредников из ПМЯ. Такой механизм дополнительно подтверждается способностью индуцировать гиперпроницаемость ЭК бесклеточным супернатантом, полученным после перекрестного связывания CD18 на ПМЯ в суспензии.
Пример 2
Идентификация полученного из нейтрофилов НВР (НВР) в качестве сигнала, связанного с индуцированной нейтрофилами утечкой плазмы из сосудов
В примере 1 адгезии ПМЯ посредством р2-интегринов предшествовали ПМЯ-индуцированные эффекты воздействия на проницаемость эндотелиальных клеток, а также Са2+-зависимая трансмембранная передача сигнала путем активируемой в результате адгезии β2-интегрин-индуцируемой секреции растворимого фактора(ов), который напрямую осуществляет запуск изменений в барьерной функции ЭК.
Как описано здесь в примере 2, определили, что растворимым фактором является человеческий НВР. В примере 2 охарактеризовано также взаимодействие НВР с фазово-контактной системой.
МАТЕРИАЛЫ И МЕТОДЫ
Реагенты
Среда 199, RPMI-1640 (содержащая L-глутамин), эмбриональная бычья сыворотка (FBS), трипсин-EDTA, забуференный фосфатом физиологический раствор (PBS) и сбалансированный раствором Хэнкса физиологический раствор (HBSS) были приобретены в Life Technologies (Gaithersburg, MD, USA). Желатин, коллагеназа, пенициллин, стрептомицин, бычий сывороточный альбумин (BSA), синий краситель Эванса и FITC-конъюгированный фаллоидин были приобретены в Sigma Chemical Co. (St. Louis, МО, USA). Fluo-3/AM в Molecular Probes (Eugene, OR, USA). Биоматрикс I был приобретен в Biomedical Technologies Inc. (Stoughton, MA, USA), a Dextran 70 (Macrodex) и Ficoll-Paque в Pharmacia Biotech AB (Uppsala, Sweden). Основная культуральная среда состояла из смеси 1:1 М-199 и RPMI-1640, обогащенной 20% инактивированной нагреванием FBS, пенициллином (100 ед/мл) и стрептомицином (100 мкг/мл). Моноклональное антитело IB4 против общей (5-цепи (CD18) (В-интегринов было предоставлено Dr. Claes Lundberg/ Pharmacia & Up John (Uppsala, Sweden) с любезного согласия Dr. Samuel D. Wright, Rockefeller University. F(ab) 2-фрагмент козьего антитела против IgG мыши было получено в Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA).
Поликлональные НВР-антитела были получены в соответствии со следующей процедурой. Новозеландских белых кроликов подкожно инъецировали 50 мкг человеческого гепарин-связывающего белка (НВР), эмульгированного в полном адъюванте Фрейнда (FCA), затем тремя бустер-инъекциями, содержащими 50 мкг человеческого НВР, эмульгированного в полном адъюванте Фрейнда (FIA), раз в две недели. Через десять дней после последней инъекции животных умерщвляли кровопусканием, и сыворотку крови собирали.
Моноклональные НВР-антитела были получены в соответствии со следующей процедурой. Мышей RBF иммунизировали путем подкожного инъецирования 20 мкг очищенного человеческого НВР в FCA, затем двумя инъекциями, содержащими 20 мкг человеческого НВР в FIA. Мыши, у которых наблюдали сильную реакцию, получили внутривенную бустер-инъекцию, содержащую 20 мкг НВР, и через три дня их селезенки были извлечены. Клетки селезенки сливали с лисьей миеломной клеточной линией. Супернатанты подвергали скринингу на наличие продукции анти-НВР-антитела с помощью НВР-специфического метода ELISA. Положительные линии клеток подвергали скринингу на предмет связывания с внутриклеточной частью макрофагальной линии с помощью стандартного FACS-метода. Положительные линии клонировали и использовали в обоих анализах. Моноклональные антитела очищали с помощью аффинной хроматографии с использованием протеина А.
Эндотелиальные клетки
Эндотелиальные клетки пупочной вены человека (HUVEC) выделяли из свежей пуповины, как описано в публикации Jaffe et al., 1973, J. Clin. Invest. 52:2745-2756, а эндотелиальные клетки аорты быка (ВАЕС) выделяли в соответствии с методом Booyse et al., 1975, Thromb. Diath. Haemmorrh. 24:825-839, с некоторыми модификациями. Вкратце, просвет пупочной вены или аорты быка инкубировали в присутствии 0,25% раствора коллагеназы в течение 10 мин, и открепившиеся эндотелиальные клетки промывали раствором PBS. Собранные клетки осаждали и ресуспендировали в культуральной среде, высевали во флаконы для культивирования тканей и инкубировали в стандартных условиях. Среду меняли каждые 48 час, пока культуры достигали стадии конфлуэнтности (3-5 дней). В экспериментах использовали HUVEC или ВАЕС от первого до пятого пассажей. Идентификацию и определение чистоты ЭК проводили на основании морфологии, напоминающей булыжник, и по фактор VIII/vWf-антигену (Booyse et al., 1975, Thromb. Diath. Haemmorrh. 24:825-839). Окрашивание на фактор VIII/vWf-антиген носило в клетках гомогенный характер.
Посев на проницаемых мембранах
Культивируемые ЭК открепляли путем кратковременной (2 мин) обработки трипсин-EDTA (0,25% трипсин/0,01% EDTA) и помещали либо на неорганическую мембрану Anopore с размером пор 0,2 мкм, либо на поликарбонатные фильтры с размером пор 3,0 мкм (Tissue Culture inserts, 10 mm; NUNC, Roskilde, Дания). Для ускорения клеточной дифференцировки и усиления прикрепления ЭК фильтры обрабатывали 50 мкл Biomatrix I (167 мкг/мл) и оставляли высыхать на воздухе. ЭК высевали с плотностью 2×105 клеток на фильтр и инкубировали в культуральной среде при 37°С в увлажненной атмосфере 5% содержания СО2 в воздухе. ЭК выращивали до образования конфлуэнтных монослоев, что контролировалось ежедневным микроскопическим наблюдением и измерением электрического сопротивления монослоя.
Измерение трансэндотелиального электрического сопротивления (TEER)
Для измерения электрического сопротивления монослоя ЭК фильтр-вставку (со встроенными клетками) переносили в камеру для измерения сопротивления (ENDOHM-12; World Precision Instruments, Sarasota, FL, USA), модифицированную таким образом, чтобы в процессе повторных измерений электроды были точно установлены по отношению друг к другу и ко встроенному фильтру. Камера (нижний компартмент) и фильтр-вставка (верхний компартмент) заполнена, соответственно, 2 мл и 400 мкл культуральной среды, помещена в инкубатор для культивирования клеток. Все измерения проводили при 37°С с помощью оборудования, установленного в инкубаторе для культивирования клеток. Непосредственное определение электрического сопротивления проводилось с помощью омметра (EVOM; World Precision Instruments, Sarasota, FL, USA), соединенного с электродами камеры для измерения электрического сопротивления. Электрическое сопротивление каждого из индивидуальных монослоев ЭК получали путем вычитания сопротивления соответствующего интактного фильтра, покрытого слоем Biomatrix, измеряемого перед засеванием его эндотелиальными клетками.
Определение клиренса альбумина
Альбумин, конъюгированный с синим красителем Эванса (ЕВА), использовали в качестве маркера макромолекулярной проницаемости монослоев ЭК. Синий краситель Эванса в конечной концентрации 0,67 мг/мл смешивали с культуральной средой, содержащей 4% BSA. В экспериментах, в которых изучали проницаемость для альбумина, культуральную среду из верхней камеры заменяли средой, содержащей ЕВА. Концентрацию ЕВА в жидких образцах верхнего и нижнего компартментов определяли спектрофотометрически путем измерения поглощения при 620 нм (Titertek Multiskan MCC; Flow Laboratories, Solna, Sweden). Клиренс альбумина вычисляли в соответствии с соотношением: V1=А2×V2×1/А1, где V1 - объем клиренса (т.е. теоретический объем апикальной среды, очищенной от альбумина путем диффузии к базолатеральному компартменту), V2 - базолатеральный объем, а А1 и А2 поглощение апикальной и базолатеральной среды, соответственно. Базовый клиренс в отсутствие какого-либо стимула, в среднем 0,08±0,01 мкл/мин, вычитывали из объема клиренса, полученного в ответ на соответствующий специфический стимул.
Выделение и активация ПМЯ
Обогащенную лейкоцитами плазму, полученную из цельной крови человека путем седиментации в декстране, тщательно наслаивали на Ficoll-Paque и центрифугировали при 400хg в течение 30 мин. Содержащий ПМЯ осадок ресуспендировали и дважды промывали (400хg, 7 мин) ледяным PBS. ПМЯ ресуспендировали в культуральной среде при концентрации 2-5×106 клеток/мл. Степень чистоты ПМЯ составляет > 98%, и жизнеспособность их, определяемая по исключению трипанового синего, составляет >95%.
Перекрестное связывание антителом молекул CD18 на поверхности нейтрофилов индуцировали, как указано выше. Вкратце, очищенные ПМЯ (5×106 мл) инкубировали в присутствии анти-CD18-mAb IB4, 10 мкг/мл в HBSS, в течение 30 мин. Во время периода инкубации клетки подвергали температурным изменениям от 4 до 37°С, чтобы усилить экспрессию CD11b/CD18 на клеточной поверхности. Клетки дважды промывали и подвергали перекрестному связыванию CD18 путем добавления F(ab')2-фрагментов козьего антитела против мышиного IgG (разбавленного в соотношении 1:20). ПМЯ осаждали путем центрифугирования при 300хg в течение 15 мин при комнатной температуре, и бесклеточный супернатант собирали. В некоторых экспериментах перекрестное связывание антителами CD18 вызывали преинкубацией ПМЯ в присутствии mAb IB4, добавленного к монослоям ЭК (см. ниже).
Экспериментальные методы
Перед переносом фильтров-вставок в камеру для измерения сопротивления (поддерживаемую при 37°С), культуральную среду во вставке и в камере заменяли свежей культуральной средой (37°С), обогащенной 10 mM Hepes. HBP (25-100 мкг/мл) или продукт секреции ПМЯ добавляли либо к верхнему, либо к нижнему компартменту, и каждую минуту, до тех пор, пока не достигалась фаза выхода на плато, измеряли величину изменения трансэндотелиального электрического сопротивления, а затем с 5-минутными интервалами. В отдельных экспериментах ПМЯ (2×106), проинкубированные в присутствии mAb IB4 (10 мкг/мл), добавляли к верхнему компартменту (отношение ПМЯ:ЭК=10:1) и в течение 10 мин давали им возможность осесть. Активацию ПМЯ индуцировали путем перекрестного связывания антитела с CD18 на поверхности лейкоцитов путем добавления F(ab')2-фрагментов козьего антитела против мышиного IgG, разбавленного в соотношении 1:20. В тех экспериментах, где изучали проницаемость монослоя ЭК для альбумина, среда в фильтре-вставке содержала ЕВА. В процессе эксперимента фильтр-вставку переносили с 5-10-минутными интервалами в новые лунки, содержащие свежую среду. Все стадии инкубации проводили при 37°С. В конце эксперимента среду из нижних лунок отбирали, и в ней определяли содержание ЕВА.
Определение внутриклеточного Са2+ и распределение F-актина в ЭК
Для определения изменений в содержании внутриклеточного свободного Са2+ использовали Са2+-чувствительную флуоресцентную пробу флуо-3/АМ (Molecular Probes Europe BV, Leiden, Нидерланды). Конфлуэнтные монослои HUVEC или BAEC, культивированные на фильтрах, покрытых Biomatrix I, инкубировали в течение 30 мин при 37°С в присутствии раствора fluo-3/AМ (3 мкМ в HBSS, содержащем 2% PCS и 10 мкМ HEPES), добавляемого как на апикальную, так и на базолатеральную поверхность. Перед использованием монослои трижды промывали и инкубировали в свежем HBSS в темноте в течение 20 мин при комнатной температуре для полного гидролиза эфирного красителя до его кальций-чувствительной свободной кислой формы. ЭК стимулировали добавлением НВР к их апикальной поверхности. Изменения в содержании [Са2+]i в ЭК в ответ на стимуляцию НВР измеряли в режиме непрерывной регистрации интенсивности флуоресценции с помощью лазерной сканирующей системы конфокального изображения (Insight Plus; Meridian Instruments Inc., Okemon, Michigan).
Для анализа распределения F-актина в ЭК в ответ на активацию под действием НВР конфлуэнтные монослои ЭК, выращенные на поверхностях, покрытых полосами Biomatrix, инкубировали в течение 15 мин при 37°С либо в присутствии НВР, либо в присутствии одного лишь наполнителя. Монослои дважды промывали культуральной средой и фиксировали в 3,7% формальдегиде в PBS в течение 10 мин при комнатной температуре.
ЭК для пермеабилизации обрабатывали 0,2% Triton X-100 в PBS (1 мин при 4°С), дважды промывали и окрашивали для визуализации актиновых филаментов FITC-конъюгированным фаллоидином в течение 20 мин при 37°С. После еще трех дополнительных промывок PBS эндотелиальные клетки просматривали с помощью лазерного сканирующего конфокального микроскопа.
Вестерн-блот-анализ
Образец НВР или секрета ПМЯ добавляли к аликвоте ДСН-буфера и кипятили в течение 5 мин. Электрофорез проводили в градиентном (3-10%) ДСН-полиакриламидном геле (ДСН-ПААГ). Для анализа методом иммуноблоттинга белки электрофоретически разгоняли на нитроцеллюлозной мембране (Bio-Rad Laboratories, Richmond, CA). Дополнительные связывающие сайты блокировали путем инкубирования нитроцеллюлозной мембраны в молоке в течение 2 час. НВР определяли методом вестерн-блоттинга.
РЕЗУЛЬТАТЫ
Активированные ПМЯ высвобождают НВР
ПМЯ в суспензии активировали путем перекрестного связывания антигена с CD18. ПМЯ центрифугировали, и супернатант собирали и прогоняли на ДСН-ПААГ. Методом вестерн-блоттинга с использованием антител против НВР распознавали заметную полосу в секрете ПМЯ, соответствующую 27 кД. В контрольном IgG кролика никаких специфических полос обнаружено не было.
НВР увеличивает проницаемость монослоев ЭК
Добавление НВР (75 мкг/мл) к верхнему компартменту вызывало значительное снижение трансэндотелиального сопротивления, которое проявлялось в течение 1 мин, достигая своего максимума (30±3% от контроля) через 15 мин и оставаясь на этом уровне в течение всего периода наблюдения (фиг.2). В ответ на НВР происходило также увеличение проницаемости ЭК для макромолекул, что выражается в прогрессивном увеличении клиренса альбумина, когда чистый объем клиренса через 60-минутный период достигал 20,4±3,4 мкл (фиг.2). Ответы ЭК на стимуляцию под действием НВР носили строго дозозависимый характер, как это видно из менее выраженного уменьшения TEER при непрерывном воздействии 25 и 50 мкг/мл НВР (фиг.2). Однако скорость возникновения ответов одинакова для всех доз НВР.
Стимуляция ЭК под действием НВР из базолатеральной части монослоя (НВР добавляли к нижнему компартменту) вызывала изменения в проницаемости ЭК, подобные таковым, вызываемым при действии НВР из апикальной части, однако при более высоких концентрациях.
Нейтрализация НВР ослабляет индуцированное ПМЯ увеличение проницаемости ЭК
Очищенный IgG антисыворотки кролика против НВР использовали для нейтрализации активности белка. Добавление НВР-антител (50 мкг/мл) к монослою ЭК перед стимуляцией НВР (75 мкг/мл) значительно уменьшало вызванные изменения в TEER и в проницаемости для макромолекул. Изменение в TEER возникало после некоторого латентного периода и достигало 75±5% от контрольных значений (фиг.3), указывая примерно на 65% ингибирование НВР-индуцируемого ответа. Сходным образом, утечка альбумина ингибировалась примерно на 60% и достигала чистого объема клиренса порядка 8,64±1,74 мкл через 60 мин (фиг.3). Обработка неспецифическим IgG кролика (50 мкг/мл) не оказывало влияния на НВР-индуцированное снижение TEER, достигая 31%±5% от контрольных значений через 15 мин.
Проводили определение того, могут ли антитела против НВР ингибировать также и изменения в барьерной функции ЭК, запускаемые активированными ПМЯ. С этой целью ПМЯ в суспензии активировали путем перекрестного связывания антител с CD18 лейкоцитов. Супернатант, полученный после центрифугирования, содержащий продукты секреции ПМЯ (см. раздел Материалы и методы), наносили на монослой ЭК. В присутствии антител против НВР наблюдали существенную супрессию уменьшения TEER и увеличения проницаемости ЭК в отношении макромолекул, по сравнению с ответами на стимуляцию продуктами секреции ПМЯ в отсутствие анти-НВР (фиг.4). Почти идентичные эффекты возникали после того, как индуцировалось перекрестное связывание антител с CD18 после добавления ПМЯ к монослою ЭК, и, следовательно, продукты секреции ПМЯ непосредственно действуют на монослой ЭК.
Стимуляция НВР повышает уровень внутриклеточного свободного Ca2+ в ЭК
С целью определения того, отвечают ли ЭК подъемом уровня свободного Са2+ в цитозоле в ответ на стимуляцию НВР, использовали метод лазерной сканирующей конфокальной микроскопии. Добавление НВР (75 мкг/мл) к конфлуэнтным монослоям HUVEC или ВАЕС, нагруженным Са2+-чувствительным флуорофором fluo-3/АМ, вызывало быстрое возрастание интенсивности флуоресценции, которая достигала своего максимального значения через 1 мин, а затем снижалась до исходных уровней. Такая мобилизация внутриклеточного Са2+ строго указывает на активный ответ ЭК на стимуляцию под действием НВР.
Стимуляция НВР вызывает реорганизацию микрофиламентов ЭК
Конфлуэнтные монослои ЭК, инкубируемые в присутствии НВР (75 мкг/мл) или же в культуральной среде как таковой в течение 15 минут, окрашивали FITC-конъюгированным фаллоидином с целью визуализации микрофиламентов и наблюдали в лазерном сканирующем конфокальном микроскопе. По сравнению с необработанными ЭК, имевшими низкую плотность F-актиновых волокон напряжения и отличающуюся плотность периферических полос вдоль маргинальной зоны клеток, в НВР-стимулированных ЭК были обнаружены существенные изменения в содержании и распределении F-актиновых волокон напряжения. Повышенная плотность покрывающих клетку F-актиновых волокон напряжения и прерывистость периферических полос указывают на реорганизацию цитоскелета ЭК вследствие стимуляции под действием НВР.
НВР увеличивает проницаемость сосудов in vivo
Местное нанесение НВР (75 мкг/мл) на микрососуды защечного мешка хомячка через 2 мин приводило к заметной утечке плазмы как из посткапиллярных, так и из больших вен. Максимальный ответ утечки наблюдался через 7-10 мин после нанесения. Затем утечка постепенно утихала и полностью прекращалась в пределах 30-минутного периода.
Ингибирование брадикинин- и НВР-индуцированного увеличения проницаемости ЭК под действием моноклонального антитела МВКЗ против брадикинина
Брадикинин (100 нМ; фиг.5а) или НВР (75 мкг/мл; фиг.5b) вводили в нулевое время на светлую поверхность монослоев ЭК, преинкубированных в присутствии моноклонального антитела против брадикинина, МВКЗ (40 мкг/мл) (Haasemann et al., 1991, J. Iniiounol. 147:3882-3892). МВК3 полностью предотвращало повышение проницаемости ЭК, вызываемое как брадикинином, так и НВР. Специфичность действия mAb определяли на основании отсутствия ингибирования изменений проницаемости, вызываемых гистамином (гистамин вводили на 15 минуте после стимуляции брадикинином или НВР), N=6.
Ингибирование НВР-индуцированного увеличения проницаемости ЭК под действием моноклонального антитела РКН4 против калликреина плазмы
НВР (75 мкг/мл; заполненные символы) или брадикинин (100 нМ; полые символы) вводили в нулевое время на светлую поверхность монослоев ЭК, преинкубированных в присутствии моноклонального антитела mAb PKH4 (40 мкг/мл). Результаты показаны на фиг.6. PKH4 предотвращало повышение проницаемости ЭК, вызываемое НВР, но не брадикинином. Обработка PKH4 не действовала на изменение проницаемости, вызываемое гистамином (гистамин вводили на 15 минуте после стимуляции НВР), N=6.
Ингибирование НВР-индуцированного увеличения проницаемости ЭК при обработке пептидом НКН20
Монослои ЭК инкубировали в присутствии НКН20 (3 мкг/мл) в течение 30 мин, такое воздействие вызывало перераспределение связанного с поверхностью Н-кининогена. После отмывки, в нулевое время, на светлую поверхность монослоя ЭК вводили НВР (75 мкг/мл), а затем, на 15 минуте брадикинин (100 нМ). НК прикреплялся к поверхности ЭК с помощью домена 5 и 6, главным образом, с помощью домена 5. Минимальная аминокислотная последовательность в домене 5, требуемая для связывания НК человека с ЭК, представлена 20-аминокислотным пептидом (НКН 20), имеющим следующую последовательность: KKHGHGHHKKNKGKKNGKH (SEQ ID NO:7) (Н-кининоген человека 479-498). Этот пептид ингибирует прикрепление НК к ЭК (Hasan et al., 1995, J. Biol. Chem. 270:19256-19261 and Herwald et al., 1996, J. Biol. Chem. 271:13040-13047). Результаты представлены на фиг. 7. Обработка ЭК пептидом НКН20 предотвращала повышение проницаемости ЭК, вызываемое НВР, но не изменений проницаемости, вызываемых последующим введением брадикинина, N=6.
Ингибирование НВР-индуцированного увеличения проницаемости ЭК при обработке пептидом SDD31
Монослои ЭК инкубировали в присутствии SDD31 (3 мкг/мл) в течение 30 мин. SDD31, обозначаемый также НК31, представляет собой пептид, полученный из домена 6 человеческого Н-кининогена, НК), который блокирует связывание НК с калликреином плазмы и с фактором XII (Tait et al, 1986, J. Biol. Chem. 261:15396-15401 and Vogel et al., 1990, J. Biol. Chem. 265:12494-12502). Он имеет последовательность SDDDWIPDIQTDPNGLSFNPISDFPDTTSPK (SEQ ID NO:8) (Н-кининоген человека 565-595). После промывки, в нулевое время, к освещенной поверхности монослоя ЭК вводили НВР (75 мкг/мл), а затем на 15 минуте брадикинин (100 нМ). Результаты представлены на фиг.8. Обработка ЭК пептидом SDD31 в большой степени предотвращает индуцируемое НВР повышение проницаемости ЭК. Ответ ЭК на непосредственную стимуляцию брадикинином после обработки пептидом SDD31 сохранялся, N=6.
Ингибирование НВР-индуцированного увеличения проницаемости ЭК при обработке апротинином
НВР (75 мкг/мл) вводили в нулевое время на освещенную поверхность монослоев ЭК, проинкубированных в присутствии апротинина (100 мкг/мл; 30 мин). Результаты представлены на фиг. 9. Обработка апротинином в большой степени предотвращает индуцируемое НВР повышение проницаемости ЭК, но не вызывала изменений проницаемости, индуцированных последующим введением брадикинина (100 нМ), N=6.
Мутант НВР (G175N) в результате мутации не обладает способностью связывать апротинин, хотя его пространственная структура и характерное поверхностное распределение заряда не изменены. При тестировании в модели утечки этот мутант индуцировал высвобождение брадикинина и увеличение проницаемости ЭК. Однако добавление апротинина не вызывало ингибирующего эффекта. Подобный результат был получен при использовании НВР свиньи, имеющего высокую степень гомологии с человеческим НВР, но не способного связывать апротинин из-за наличия R на дне вырожденного каталитического кармана.
Пример 3
Два мутанта гепарин-связывающего белка
Для исследований структурно-функциональных отношений остатков в предполагаемом сайте связывания липид A/LPS и сайте связывания BPTI (ингибитор трипсина поджелудочной железы быка) были получены два мутанта НВР человека, [R23S,F25E]HBP и [G175Q]HBP. Мутация G175Q не изменяет структуру белка, однако лишает его способности связывать BPTI, и мутант опосредует лишь весьма ограниченную стимуляцию липополисахарид (LPS)-индуцируемого высвобождения цитокинов из моноцитов человека. Способность связывания липид A/LPS сравнима с таковой нативного НВР. Мутации R23S.F25E не влияют на связывание липид A/LPS и BPTI или липополисахарид (LPS)-индуцируемого высвобождения цитокинов из моноцитов человека.
МАТЕРИАЛЫ И МЕТОДЫ
Конструирование мутантных белков
ДНК-полимеразу Pwo приобретали в Boehringer Mannheim, ДНК-лигазы BamH1 и Т4 в New England Biolabs, клетки spodoptera frugiperda (sf9) и переносящую плазмиду pVL1393 в Invitrogen, TC100 и SF900-II продуцированы на фирме Novo Nordisk A/S, эмбриональная телячья сыворотка (FCS) на фирме GibcoBRL. Все остальные химические реагенты высокой степени чистоты являются широко доступными.
Мутант [R23S, F25E]HBP
Мутант [R23S, F25E]HBP сконструировали с помощью техники перекрывающихся удлиняющих сегментов ПЦР (Но, S.N., Hunt, H.D., Horton, R.M., Pullen, J.K., and Pease, L.R., (1989) Gene (Amst) 77: 51-59). Два частично перекрывающихся мутированных фрагмента получены с помощью ПЦР с использованием олигонуклеотидов 1+2 и 3+6, и полученные в результате фрагменты соединены с помощью олигонуклеотидов 1+6 (Таблица 1). Нативная кДНК НВР человека использована в качестве матрицы в реакции ПЦР. Полученная в результате кДНК кодировала пропептид, состоящий из 19 аминокислот, часть зрелого белка из 222 аминокислот и С-концевой вставочный материал из трех аминокислот. Сайты BamH1 включены с каждой стороны мутированной кДНК для клонирования. Мутированный фрагмент лигирован в трансфер-плазмиду pVL1393 (Invitrogen). ДНК для трансфекции получали с использованием наборов Qiagen midiprep. После очистки вставку секвенировали.

Мутант [G175Q]HBP
В целом, этот мутант сконструирован, как описано выше. Два частично перекрывающихся фрагмента получены методом ПЦР с использованием олигонуклеотидов 1+4 и 5+6, и полученные в результате фрагменты соединены с помощью олигонуклеотидов 1+6 (Таблица 1).
Экспрессия и очистка
Клетки насекомого sf9 поддерживали в среде ТС100, обогащенной 10% FCS. Перед инфицированием среду заменяли бессывороточной средой SF900-II. Экспрессию обоих мутантов в бессывороточной среде и очистку мутантов от культурального супернатанта производили описанным выше способом для нативного НВР человека (Rasmussen, Р.В., Bjorn, S., Hastrup, S., Nielsen, P.F., Norris, K., Thim, L., Wiberg, F.C., and Flodgaard, H., (1996) FEBS Lett. 390:109-112).
Анализ индуцированного LPS высвобождения IL-6 из моноцитов
Моноциты человека выделяли и обрабатывали в соответствии с методикой (Rasmussen, Р.В., Bjorn, S., Hastrup, S., Nielsen, P.F., Norris, К., Thim, L., Wiberg, F.C., and Flodgaard, H., (1996) FEBS Lett. 390:109-112, за исключением того, что из среды был исключен 1% не-незаменимых аминокислот.
Анализ связывания 3H-LPS с НВР
Аффинность к липополисахариду (3H-LPS) измеряли, используя технику сцинтилляционного анализа микрозазоров (scintillation proximity assay technology) (SPA), описанную в литературе (Linde, V., Bjorn, S., Kastrup, J.S., and Flodgaard, H., 2000, Biotechniques 28:218-222). Все образцы были представлены тремя параллельными пробами.
Анализ связывания 125I-BPTI с НВР
Аффинность в отношении BPTI определяли, используя технику SPA, в основном как описано для анализа LPS (Linde, V., Bjorn, S., Kastrup, J.S., and Fiodgaard, H., 2000, Biotechniques 28:218-222), с несколькими модификациями. Так, 3H-LPS был заменен на 125I-BPTI (полученный на фирме Novo Nordisk A/S), и буфер был заменен буфером PBS (20 мМ фосфат, рН 7,4, и 150 мМ NaCl), включающим в себя 0,05% BSA и 0,05% Triton X-100. Относительные аффинности НВР и мутантов определяли методом конкурентного анализа, используя фиксированные, возрастающие концентрации немеченого BPTI.
РЕЗУЛЬТАТЫ
Биологические свойства
Биологическую активность двух мутантов сравнивали с нативным НВР в отношении их способности усиливать, дозо-зависимым способом, индуцированное LPS высвобождение I1-6 из моноцитов человека in vitro. Аффинность в отношении LPS двух мутантов и НВР измеряли методом сцинтилляционного анализа микрозазоров (SPA) (Linde, V., Bjorn, S., Kastrup, J.S., and Flodgaard, H., 2000, Biotechniques 28:218-222). Оба мутанта связывали 3H-LPS насыщающим образом, со сходным кажущимся Ка (приблизительно 0,7 мкг/мл), как и в случае нативного НВР (фиг. 11а). Более того, для обоих мутантов и для нативного НВР измеряли афинность в отношении BPTI сходным методом сцинтилляционного анализа микрозазоров, где в качестве лиганда для НВР использовали 125I-BPTI. Мутант [R23S,F25E]НВР, так же, как и нативный НВР, связывал BPTI, тогда как никакого связывания мутанта [G175Q]HBP обнаружено не было (фиг. 11b). С помощью конкурентного анализа с использованием фиксированных, возрастающих концентраций немеченого BPTI показали, что 125I-BPTI связывается с НВР и [R23S, F25E]НВР со сходными кажущимися значениями IC50, примерно составляющими 7 нМ. Как показано на фиг.11с, [R23S, F25E]НВР обладает большей кажущейся способностью связывания по сравнению с НВР. Возможно, это различие объясняется способностью [R23S, F25E]НВР связывать различные формы использованных йодированных BPTI. Однако такая повышенная способность связывания не влияет на аффинность 125I-BPTI. Кроме того, в опытах конкурентного анализа связывания изучали способность 3H-LPS вытеснять 125I-BPTI, связанный с нативным НВР, и наоборот: конкуренцию за связывание 125I-BPTI определяли при добавлении вплоть до 5 мкг/мл немеченого LPS, а конкуренцию за связывание 3H-LPS добавлением вплоть до 50 мМ немеченого BPTI. В обеих ситуациях эффектов ингибирования не наблюдали, что указывает на то, что сайты связывания LPS и BPTI локализованы в различных частях молекулы НВР.
Данные анализа биологической активности in vitro показали, что [G175Q1HBP, в отличие от [R23S, F25E]HBP и нативного НВР, является посредником лишь весьма ограниченной стимуляции индуцированного липополисахаридом (LPS) высвобождения цитокинов из моноцитов человека (фиг. 10).
Объем описанного выше и заявленного изобретения не ограничивается перечисленными здесь специфическими аспектами, поскольку эти аспекты приведены лишь в качестве иллюстрации некоторых приложений данного изобретения. Любые эквивалентные применения включаются в объем данного изобретения. В действительности специалистам в данной области нетрудно будет внести некоторые модификации данного изобретения, помимо описанных и показанных здесь в предшествующем описании. Такие модификации также считаются включенными в объем прилагаемой формулы изобретения.
В описании приведен целый ряд различных ссылок на публикации, которые в полном объеме считаются включенными в данное изобретение.
Список последовательностей приведен в конце описания.






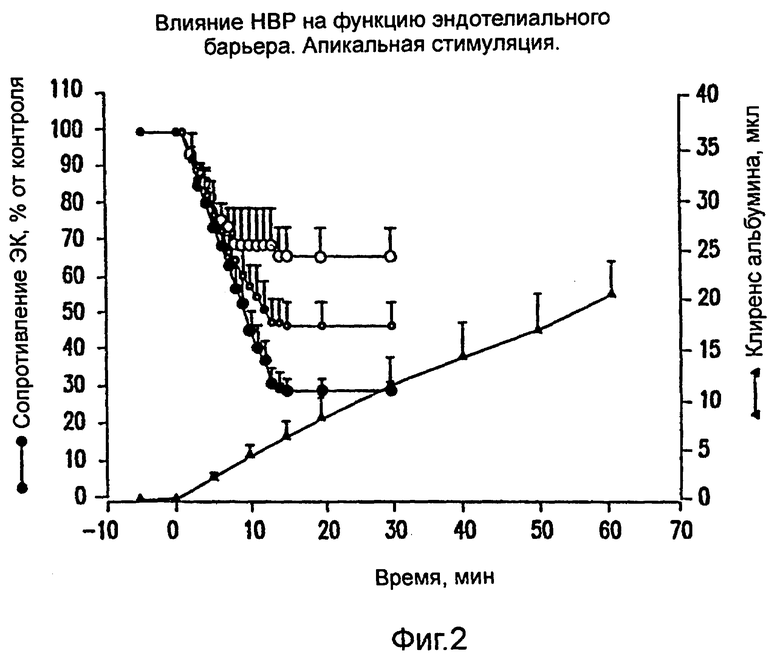

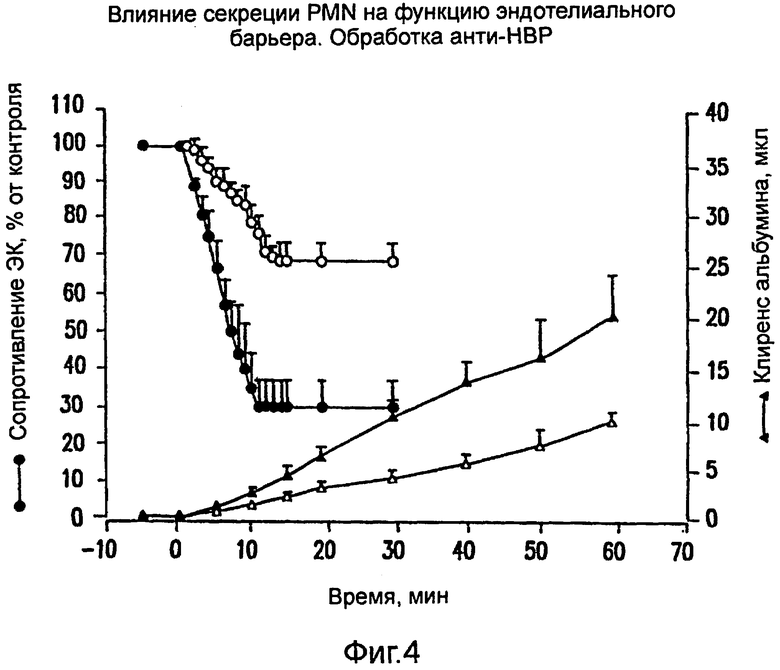
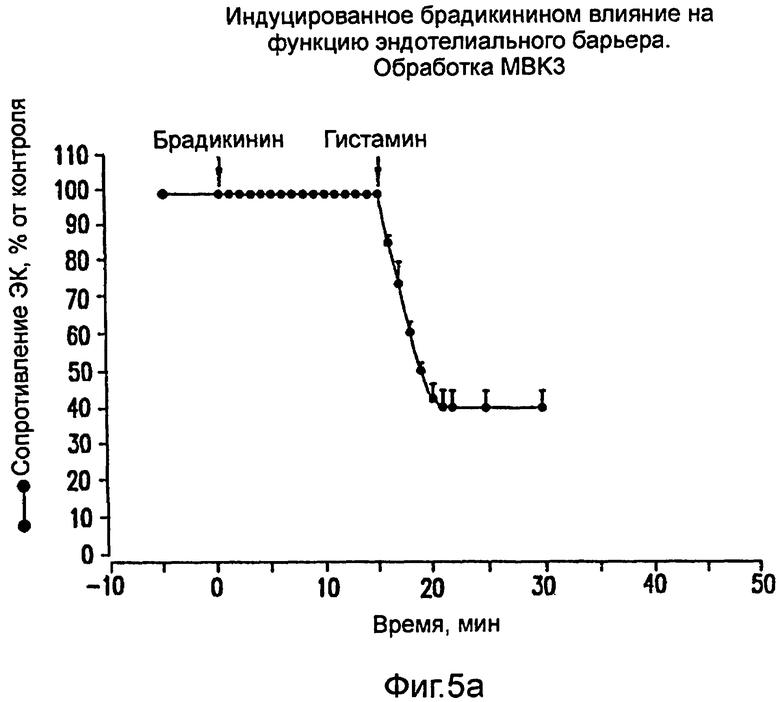
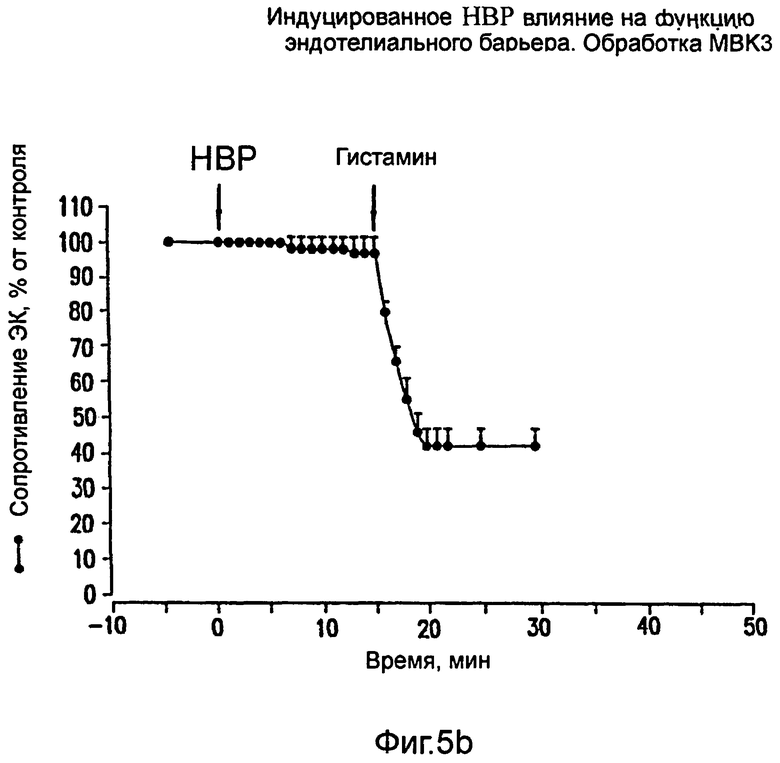
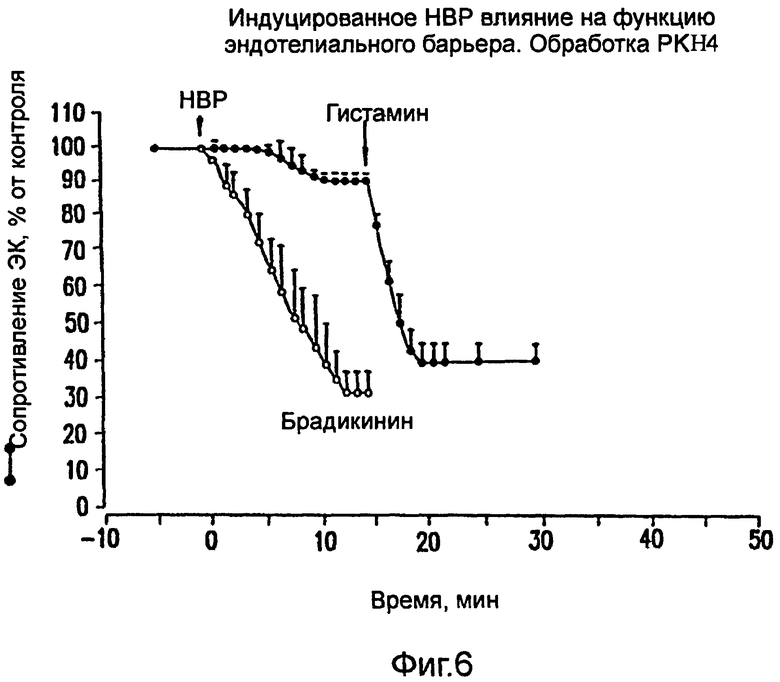
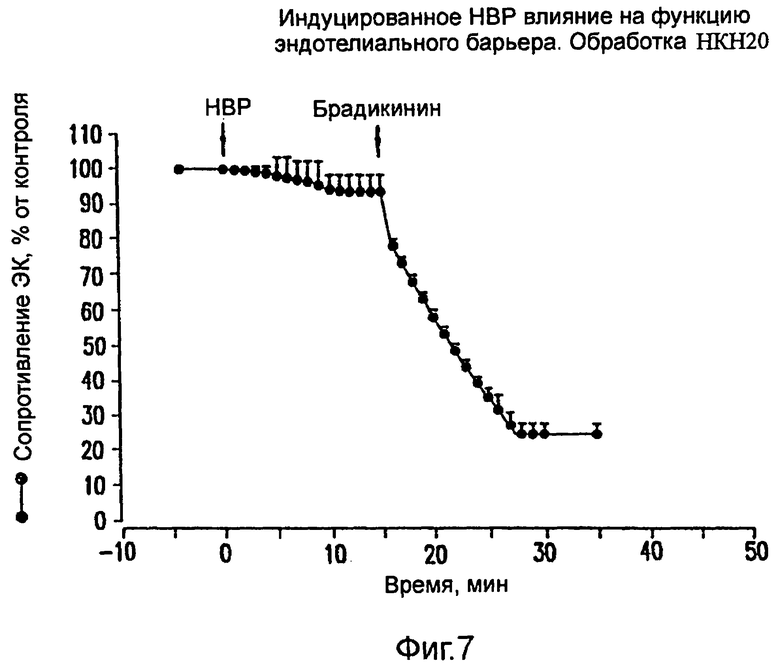

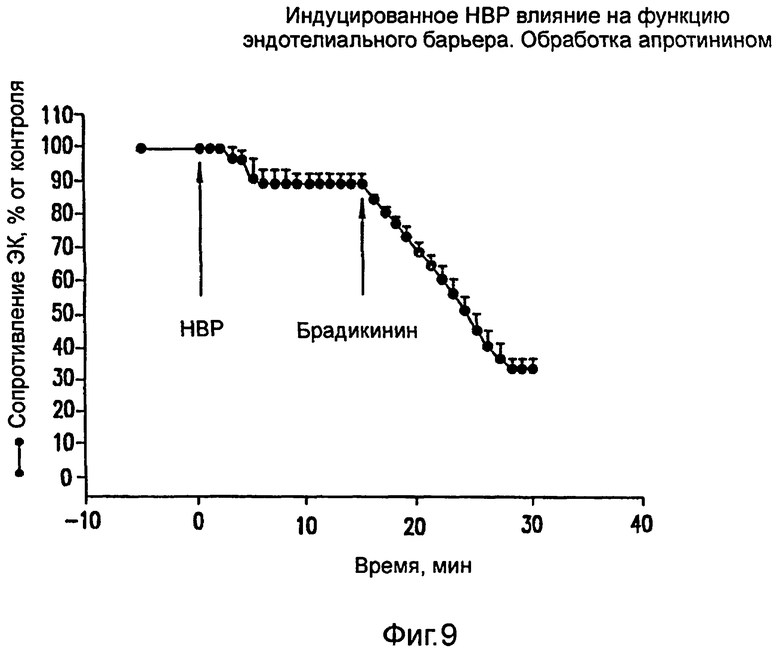
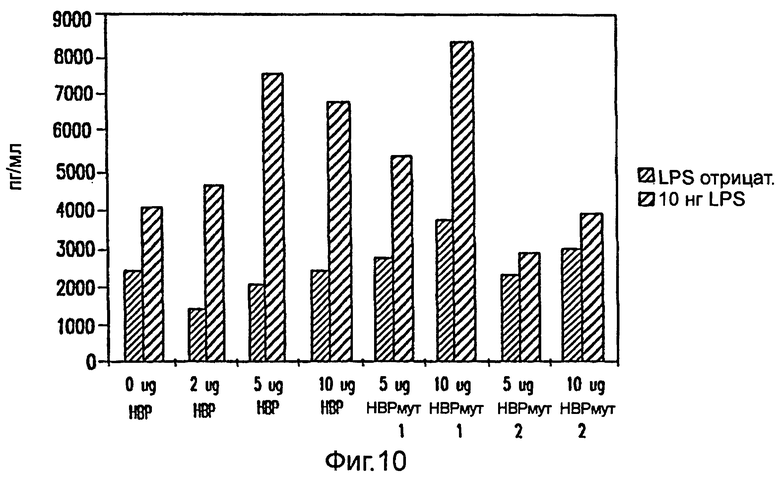

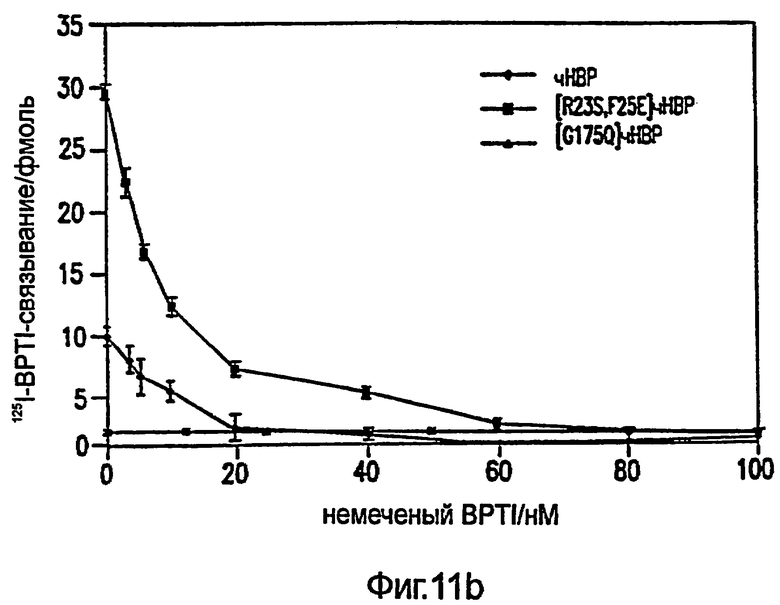
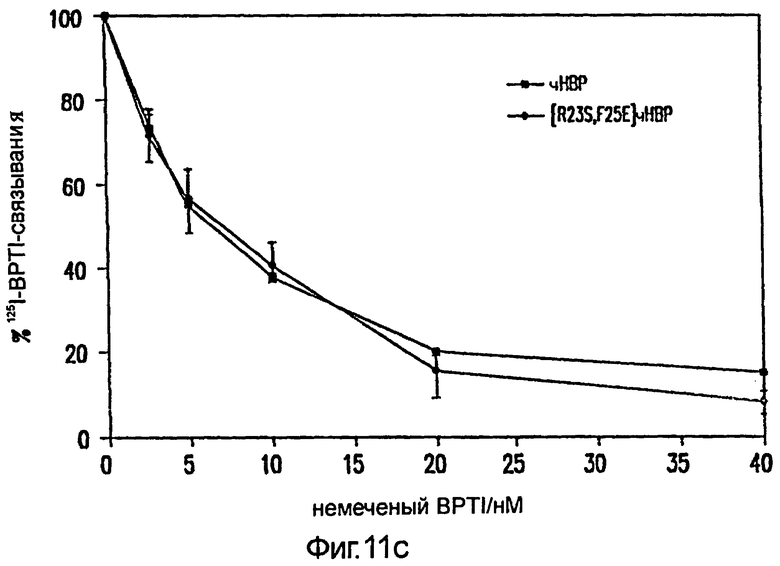
Изобретение относится к медицине, в частности к фармации и фармакологии, и касается создания лекарственного средства для лечения нарушений, связанных с высвобождением брадикинина. Для этого предложено использовать анти-НВР-антитела млекопитающего. Предлагаются способы идентификации моноклонального антитела, которое связывается по меньшей мере с одним эпитопом на нативном НВР (гепарин-связывающем белке), способы определения того, продуцирует ли млекопитающее НВР, который связывается с моноклональным антителом, а также наборы для этого. Изобретение обеспечивает возможность использования указанных антител в решении задач предупреждения и лечения нарушений, ассоциированных с высвобождением брадикинина. 6 с. и 19 з.п. ф-лы, 1 табл., 11 ил.
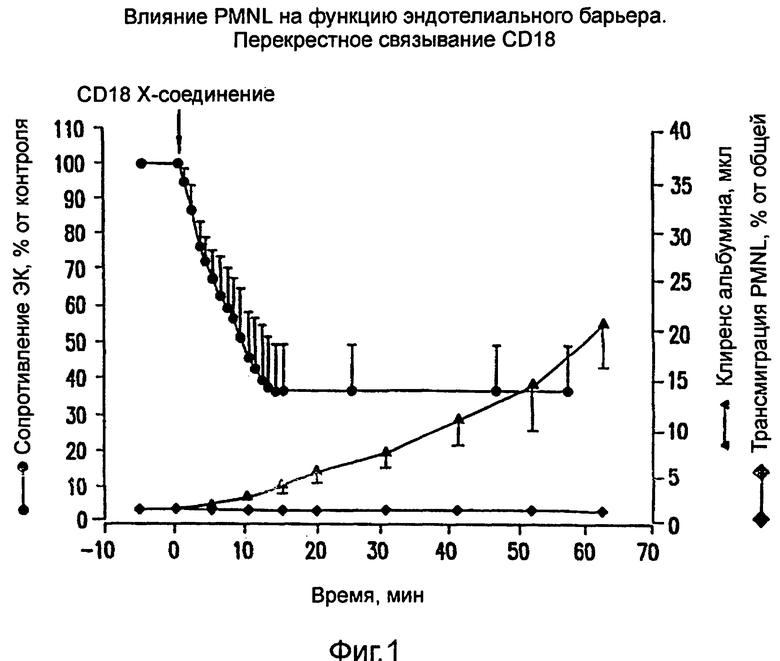
Приоритет по пунктам:
| WO 9305396, 18.03.1993 | |||
| OSTERGAARD et al | |||
| J | |||
| Lenc | |||
| Biol | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| PATTON W.A | |||
| et al | |||
| Biochem | |||
| J | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| Способ приготовления массы для карандашей | 1921 |
|
SU311A1 |
| RU 96105493 A, 10.06.1998 | |||
| RU 92016227 A, 10.03.1995 | |||
| ПЕПТИДЫ, СПОСОБ ИХ ПОЛУЧЕНИЯ, СРЕДСТВО, ОБЛАДАЮЩЕЕ БРАДИКИНИН-АНТАГОНИСТИЧЕСКОЙ АКТИВНОСТЬЮ | 1993 |
|
RU2109747C1 |
| БЕРТРАМ Г | |||
| КАТЦУНГ | |||
| Базисная и клиническая фармакология | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| - М., СПб., 1998, с.61-62. | |||

